1 Project no. GOCE-CT-2003-505539 Project acronym: ENSEMBLES Project title: ENSEMBLE-based Predictions of Climate Changes and their Impacts Instrument: Integrated Project Thematic Priority: Global Change and Ecosystems Deliverable 2B.35 Journal papers on the impacts application of methods for the construction of probabilistic regional climate projections Due date of deliverable: February 2009 Actual submission date: February 2010 Start date of project: 1 September 2004 Duration: 64 Months Organisation name of lead contractor for this deliverable: University of East Anglia Revision [Final] Project co-funded by the European Commission within the Sixth Framework Programme (2002-2006) Dissemination Level PU Public PU PP Restricted to other programme participants (including the Commission Services) RE Restricted to a group specified by the consortium (including the Commission Services) CO Confidential, only for members of the Consortium (including the Commission Services)
Transcript
1
Project no. GOCE-CT-2003-505539
Project acronym: ENSEMBLES
Project title: ENSEMBLE-based Predictions of Climate Changes and their Impacts
Instrument: Integrated Project Thematic Priority: Global Change and Ecosystems Deliverable 2B.35 Journal papers on the impacts application of methods for the
construction of probabilistic regional climate projections
Due date of deliverable: February 2009 Actual submission date: February 2010
Start date of project: 1 September 2004 Duration: 64 Months Organisation name of lead contractor for this deliverable: University of East Anglia
Revision [Final]
Project co-funded by the European Commission within the Sixth Framework Programme (2002-2006)
Dissemination Level PU Public PU PP Restricted to other programme participants (including the Commission Services) RE Restricted to a group specified by the consortium (including the Commission Services) CO Confidential, only for members of the Consortium (including the Commission Services)
2
Deliverable 2B.35 Journal papers on the impacts application of methods for the construction of probabilistic regional climate projections At the time of writing (February 2010), four papers are published, or in press, and another three are in preparation. They are referenced below and represent the output from three partners: ULUND, NIHWM and FUB. Another partner, PAS, is expected to publish results of relevance to this deliverable at a later date, and a summary of some of the work they have done using probabilistic projections may be found in the ENSEMBLES final science summary report, downloadable at: http://ensembles-eu.metoffice.com/docs/Ensembles_final_report_Nov09.pdf Papers marked with an asterisk (*) are reproduced in this document. Donat M.G., Leckebusch G.C., Wild S., Ulbrich U., 2010: Future changes of European winter storm losses and extreme wind speeds in multi-model GCM and RCM simulations. Special Issue of Natural Hazards and Earth System Sciences (NHESS): Applying ensemble climate change projections for assessing risks of impacts in Europe, in press. * Jönsson, A.M., Appelberg, G., Harding, S. and Bärring, L. 2009: Spatio-temporal impact of climate change on the activity and voltinism of the spruce bark beetle, Ips typographus Global Change Biology 15:486-499. * Mares C., Mares I., Mihailescu M., Hübener M., Cubasch U., Stanciu P., 2009: 21st century discharge estimation in the Danube lower basin with predictors simulated through EGMAM model. Revue Roumaine de Géophysique, tom 52, 2009. Rammig, A., Jönsson, A.M., Hickler, T., Smith, B., Bärring, L., Sykes, M.T. 2010: Impacts of changing frost regimes on Swedish forests: Incorporating cold hardiness in a regional ecosystem model. Ecological Modelling 303-313. * In preparation: Jönsson, A.M., Harding, S., Krokene, P., Lange, H., Lindelöw, Å., Økland, B. and Schroeder, L.M. Modelling the potential impact of global warming on Ips typographus voltinism and reproductive diapause. Jönsson, A.M., Bärring, L. Ensemble-based analysis of climate impact on Norway spruce bark beetle swarming activity in northern and central Europe. Jönsson, A.M., Bärring, L. Warming up for spring backlashes in Norway spruce forests.
Future changes of European winter storm losses and extreme wind speeds in multi-model GCM and RCM simulations
M.G. Donat, G.C. Leckebusch, S. Wild, U. Ulbrich
Institute for Meteorology, Freie Universität Berlin, Germany
Figure 1. Population density on a 0.25°x0.25° grid is used as a proxy for insured values in the
considered regions for which loss calculations were performed (unit: inhabitants per km²).
Figure 2. Daily maximum wind speed (WIMAX), 98th percentile in the GCM simulations
a) absolute values for 20C (unit: m/s)
Figure 2. Daily maximum wind speed (WIMAX), 98th percentile in the GCM simulations
b) ACC signals A1B-20C: magnitude of changes is displayed by black iso-lines (unit: m/s), coloured areas indicate statistical significance above 0.9 (Student-T-test).
Figure 3. Ensemble Mean of 98th percentile of WIMAX in the GCM simulations
a) absolute values for 20C (unit: m/s)
b) anomaly GCM ensemble (20C) relative to ERA40 (unit: m/s)
c) ACC signal A1B-20C: magnitude of changes is displayed by black iso-lines (unit:
m/s), coloured areas indicate statistical significance above 0.9 (Student-T-test)
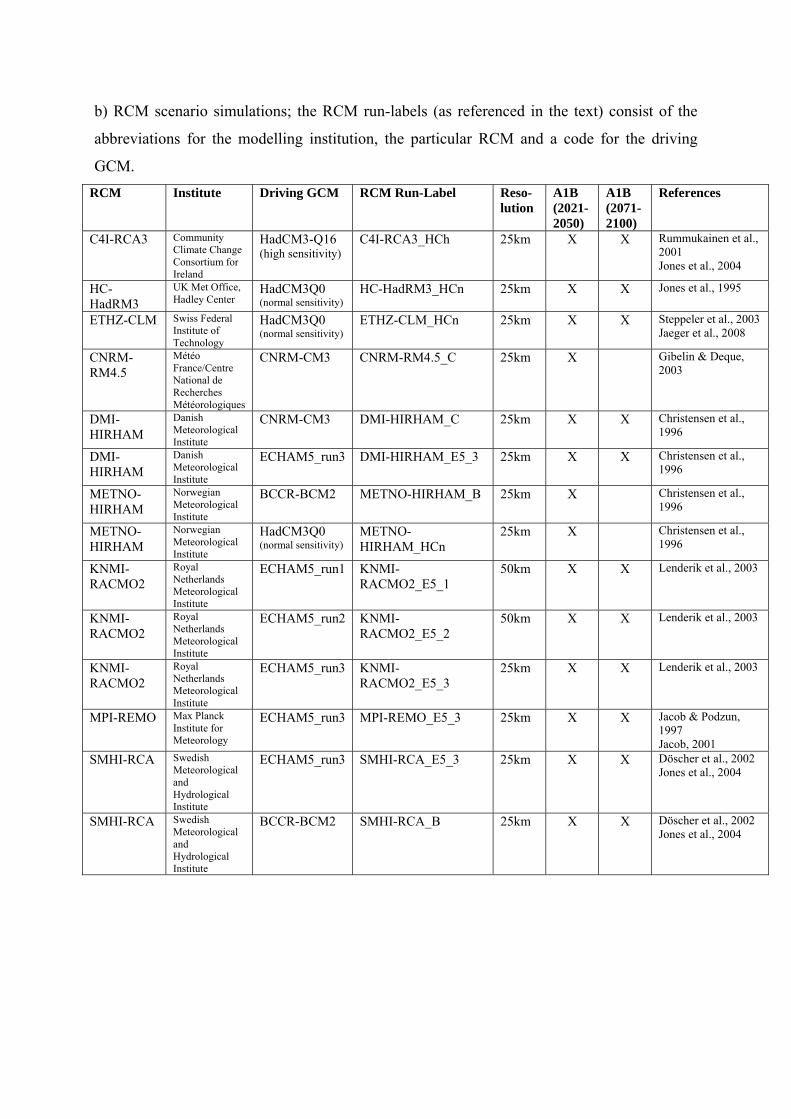
a) end of 21C (2071-2100), based on 9 GCM and 11 RCM simulations
b) middle of 21C (2021-2050), based on 9 GCM and 14 RCM simulations
Figure 4. Relative changes (unit: %) of mean annual storm loss potential based on the GCM
(upper row) and RCM (bottom row) simulations for the end (a) and middle (b) of the 21st
century compared to recent climate conditions (20C, 1961-2000). Values in parentheses are
inter-model standard deviations.
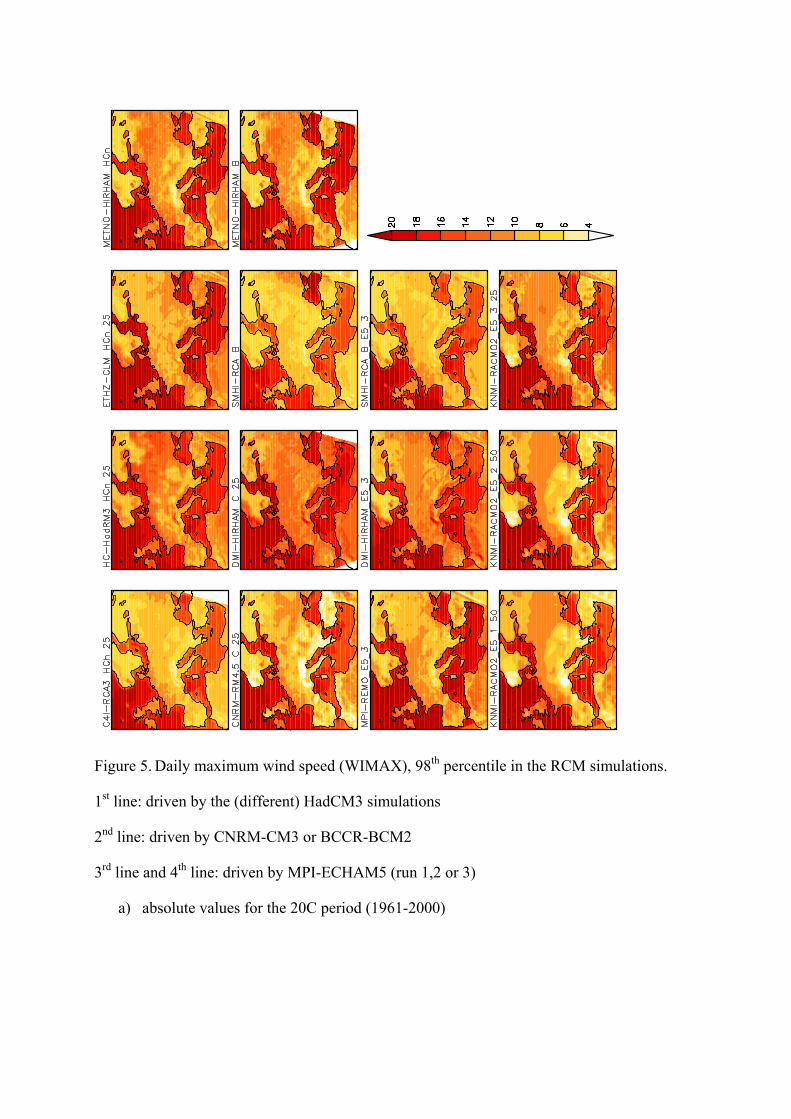
Figure 5. Daily maximum wind speed (WIMAX), 98th percentile in the RCM simulations.
1st line: driven by the (different) HadCM3 simulations
2nd line: driven by CNRM-CM3 or BCCR-BCM2
3rd line and 4th line: driven by MPI-ECHAM5 (run 1,2 or 3)
a) absolute values for the 20C period (1961-2000)
Figure 5. Daily maximum wind speed (WIMAX), 98th percentile in the RCM simulations.
1st line: driven by the (different) HadCM3 simulations, 2nd line: driven by CNRM-CM3 or
BCCR-BCM2, 3rd line and 4th line: driven by MPI-ECHAM5 (run 1,2 or 3)
b) ACC signals for 98th percentile of WIMAX in the RCM simulations, all results are for
the future period 2071-2100, except for METNO_HIRHAM_HCn*, METNO-
HIRHAM_B* and CNRM-RM4.5_C* (only integrated until 2050) signals for the
period 2021-2050 are presented. Magnitude of changes is displayed by black iso-lines
(unit: m/s), coloured areas indicate statistical significance above 0.9 (Student-T-test).
Figure 6. RCM-Ensemble Mean of ACC signal for 98th percentile of WIMAX in the future
scenario simulations. Magnitude of changes is displayed by black iso-lines (unit: m/s),
coloured areas indicate statistical significance above 0.9 (Student-T-test).
a) A1B (2021-2050) – 20C
b) A1B (2071-2100) – 20C
Figure 7. Probability of loss potential changes for the future climate period end of 21C (2071-
2100) compared to 20C (1961-2000) without adaptation of the loss threshold, based on all
possible model combinations of GCMs (green curve), RCMs (blue curve) and all available
GCM and RCM scenario (red curve) simulations. The red shaded areas mark the range where
90% of the change signals (between 5th and 95th percentile) based on all model combinations
are found.
Appendix A: Supplementary Material
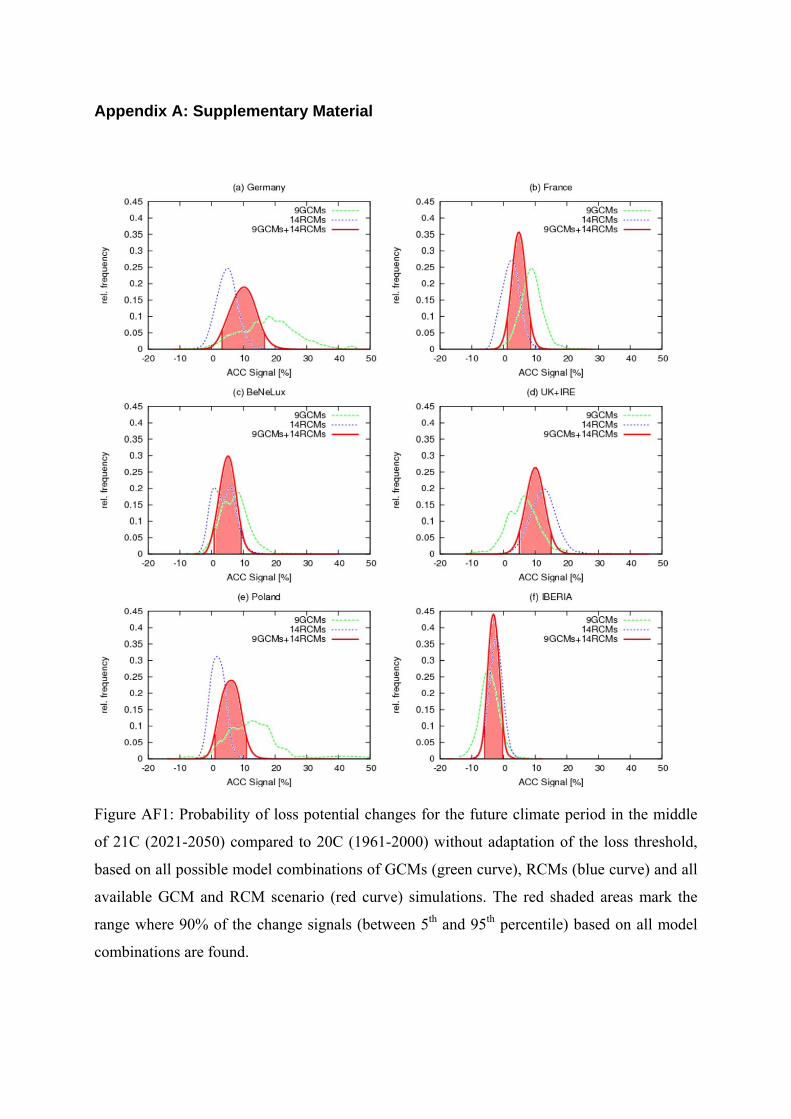
Figure AF1: Probability of loss potential changes for the future climate period in the middle
of 21C (2021-2050) compared to 20C (1961-2000) without adaptation of the loss threshold,
based on all possible model combinations of GCMs (green curve), RCMs (blue curve) and all
available GCM and RCM scenario (red curve) simulations. The red shaded areas mark the
range where 90% of the change signals (between 5th and 95th percentile) based on all model
combinations are found.
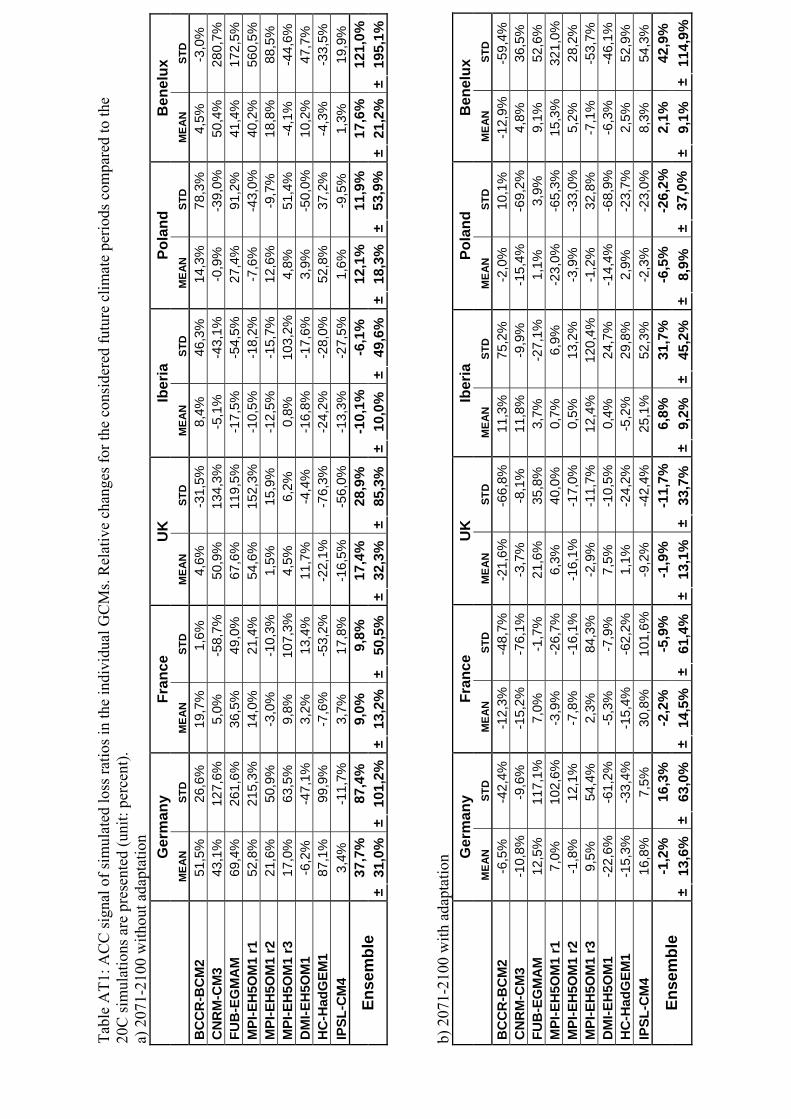
Tabl
e A
T1: A
CC
sign
al o
f sim
ulat
ed lo
ss ra
tios i
n th
e in
divi
dual
GC
Ms.
Rel
ativ
e ch
ange
s for
the
cons
ider
ed fu
ture
clim
ate
perio
ds c
ompa
red
to th
e 20
C si
mul
atio
ns a
re p
rese
nted
(uni
t: pe
rcen
t).
a) 2
071-
2100
with
out a
dapt
atio
n
Ger
man
y Fr
ance
U
K
Iber
ia
Pola
nd
Ben
elux
MEA
N
STD
M
EAN
ST
D
MEA
N
STD
M
EAN
ST
D
MEA
N
STD
M
EAN
ST
D
BC
CR
-BC
M2
51
,5%
26,6
%
19
,7%
1,6%
4,6%
-31,
5%
8,
4%
46
,3%
14,3
%
78
,3%
4,5%
-3,0
%
CN
RM
-CM
3
43,1
%
12
7,6%
5,0%
-58,
7%
50
,9%
134,
3%
-5,1
%
-4
3,1%
-0
,9%
-39,
0%
50,4
%
28
0,7%
FU
B-E
GM
AM
69,4
%
26
1,6%
36,5
%
49
,0%
67,6
%
11
9,5%
-1
7,5%
-5
4,5%
27
,4%
91,2
%
41
,4%
172,
5%
MPI
-EH
5OM
1 r1
52,8
%
21
5,3%
14,0
%
21
,4%
54,6
%
15
2,3%
-1
0,5%
-1
8,2%
-7
,6%
-43,
0%
40,2
%
56
0,5%
M
PI-E
H5O
M1
r2
21
,6%
50,9
%
-3
,0%
-10,
3%
1,
5%
15
,9%
-12,
5%
-15,
7%
12,6
%
-9
,7%
18,8
%
88
,5%
M
PI-E
H5O
M1
r3
17
,0%
63,5
%
9,
8%
10
7,3%
4,
5%
6,
2%
0,
8%
10
3,2%
4,
8%
51
,4%
-4,1
%
-4
4,6%
D
MI-E
H5O
M1
-6
,2%
-47,
1%
3,
2%
13
,4%
11,7
%
-4
,4%
-16,
8%
-17,
6%
3,9%
-50,
0%
10,2
%
47
,7%
H
C-H
adG
EM1
87
,1%
99,9
%
-7
,6%
-53,
2%
-2
2,1%
-7
6,3%
-24,
2%
-28,
0%
52,8
%
37
,2%
-4,3
%
-3
3,5%
IP
SL-C
M4
3,
4%
-1
1,7%
3,7%
17,8
%
-1
6,5%
-5
6,0%
-13,
3%
-27,
5%
1,6%
-9,5
%
1,
3%
19
,9%
37,7
%
87
,4%
9,0%
9,8%
17,4
%
28,9
%
-1
0,1%
-6
,1%
12,1
%
11,9
%
17
,6%
121,
0%
Ense
mbl
e ±
31,0
%
± 10
1,2%
±13
,2%
±50
,5%
±
32,3
%±
85,3
%
±10
,0%
±
49,6
%
±18
,3%
±53
,9%
±
21,2
%
± 19
5,1%
b)
207
1-21
00 w
ith a
dapt
atio
n
G
erm
any
Fran
ce
UK
Ib
eria
Po
land
B
enel
ux
M
EAN
ST
D
MEA
N
STD
M
EAN
ST
D
MEA
N
STD
M
EAN
ST
D
MEA
N
STD
B
CC
R-B
CM
2
-6,5
%
-4
2,4%
-12,
3%
-48,
7%
-2
1,6%
-6
6,8%
11,3
%
75,2
%
-2
,0%
10,1
%
-1
2,9%
-59,
4%
CN
RM
-CM
3
-10,
8%
-9
,6%
-15,
2%
-76,
1%
-3
,7%
-8,1
%
11
,8%
-9
,9%
-15,
4%
-69,
2%
4,
8%
36
,5%
FU
B-E
GM
AM
12,5
%
11
7,1%
7,
0%
-1
,7%
21,6
%
35
,8%
3,7%
-27,
1%
1,
1%
3,
9%
9,
1%
52
,6%
M
PI-E
H5O
M1
r1
7,
0%
10
2,6%
-3
,9%
-26,
7%
6,
3%
40
,0%
0,7%
6,9%
-23,
0%
-65,
3%
15
,3%
321,
0%
MPI
-EH
5OM
1 r2
-1,8
%
12
,1%
-7,8
%
-1
6,1%
-16,
1%
-17,
0%
0,
5%
13
,2%
-3,9
%
-3
3,0%
5,2%
28,2
%
MPI
-EH
5OM
1 r3
9,5%
54,4
%
2,
3%
84
,3%
-2,9
%
-1
1,7%
12,4
%
120,
4%
-1
,2%
32,8
%
-7
,1%
-53,
7%
DM
I-EH
5OM
1
-22,
6%
-6
1,2%
-5,3
%
-7
,9%
7,5%
-10,
5%
0,
4%
24
,7%
-14,
4%
-68,
9%
-6
,3%
-46,
1%
HC
-Had
GEM
1
-15,
3%
-3
3,4%
-15,
4%
-62,
2%
1,
1%
-2
4,2%
-5,2
%
29
,8%
2,9%
-23,
7%
2,
5%
52
,9%
IP
SL-C
M4
16
,8%
7,5%
30,8
%
10
1,6%
-9
,2%
-42,
4%
25
,1%
52
,3%
-2,3
%
-2
3,0%
8,3%
54,3
%
-1
,2%
16,3
%
-2
,2%
-5,9
%
-1
,9%
-11,
7%
6,8%
31,7
%
-6
,5%
-26,
2%
2,1%
42,9
%
Ense
mbl
e ±
13,6
%
± 63
,0%
±
14,5
%±
61,4
%
±13
,1%
±33
,7%
±
9,2%
±
45,2
%
±8,
9%
±37
,0%
±
9,1%
±
114,
9%
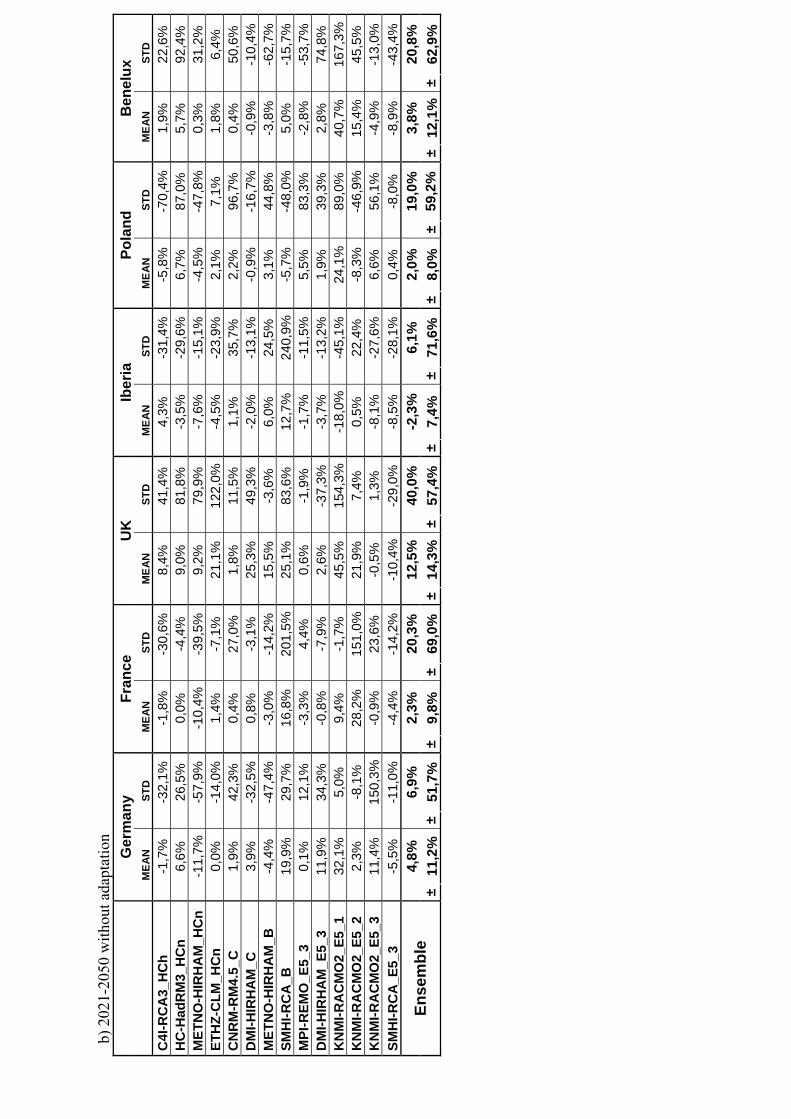
c) 2
021-
2050
with
out a
dapt
atio
n
Ger
man
y Fr
ance
U
K
Iber
ia
Pola
nd
Ben
elux
MEA
N
STD
M
EAN
ST
D
MEA
N
STD
M
EAN
ST
D
MEA
N
STD
M
EAN
ST
D
BC
CR
-BC
M2
14
,5%
-10,
1%
10
,8%
10
,0%
-0,8
%
-3
5,7%
11,4
%
69
,9%
-13,
4%
-68,
3%
-0
,9%
-9,9
%
CN
RM
-CM
3
19,8
%
55
,9%
3,1%
-51,
8%
8,
3%
17
,8%
1,0%
-5,4
%
-5
,9%
-55,
2%
16
,9%
120,
6%
FUB
-EG
MA
M
7,
1%
14
,1%
-3,4
%
-3
0,7%
19,6
%
11
9,5%
-6
,5%
-17,
9%
17,4
%
15
7,5%
5,
3%
26
,1%
M
PI-E
H5O
M1
r1
45
,5%
103,
4%
26,5
%
3,4%
33,7
%
77
,7%
-13,
5%
-29,
6%
17,3
%
-1
6,6%
32,4
%
25
4,6%
M
PI-E
H5O
M1
r2
4,
5%
7,
1%
14
,4%
11
9,7%
2,0%
31,6
%
-6
,9%
0,8%
10,1
%
13
,3%
9,7%
30,8
%
MPI
-EH
5OM
1 r3
-9,6
%
-1
6,3%
2,5%
71,9
%
1,
6%
7,
5%
-1
3,6%
-3
7,0%
11
,1%
22,3
%
-8
,7%
-55,
8%
DM
I-EH
5OM
1
16,8
%
-1
3,5%
6,6%
-7,6
%
1,
9%
-1
4,8%
3,7%
41,9
%
53
,3%
85,3
%
1,
4%
-9
,3%
H
C-H
adG
EM1
69
,2%
156,
8%
18,0
%
36,0
%
-1
2,7%
-6
1,6%
-11,
2%
-27,
6%
17,5
%
-2
3,9%
4,8%
80,8
%
IPSL
-CM
4
-6,1
%
-1
0,5%
0,7%
-5,0
%
3,
0%
3,
8%
-3
,5%
-16,
6%
2,2%
-15,
4%
3,
2%
62
,9%
17,9
%
31
,9%
8,8%
16,2
%
6,
3%
16
,2%
-4,3
%
-2
,4%
12,2
%
11,0
%
7,
1%
55
,6%
En
sem
ble
±25
,1%
±
61,4
%
±9,
5%
±52
,7%
±
13,3
%±
55,5
%
±8,
4%
±35
,7%
±
18,9
%±
71,1
%
±11
,8%
±
91,5
%
Tabl
e A
T2: A
CC
sign
al o
f los
ses c
alcu
late
d in
the
indi
vidu
al R
CM
scen
ario
sim
ulat
ions
. Cha
nge
sign
als A
1B-2
0C w
ithou
t ada
ptat
ion
of th
e lo
ss
thre
shol
d fo
r the
2 fu
ture
per
iods
in th
e m
iddl
e (2
021-
2050
) and
at t
he e
nd o
f 21C
(207
1-21
00).
a) 2
071-
2100
with
out a
dapt
atio
n
Ger
man
y Fr
ance
U
K
Iber
ia
Pola
nd
Ben
elux
MEA
N
STD
M
EAN
ST
D
MEA
N
STD
M
EAN
ST
D
MEA
N
STD
M
EAN
ST
D
C4I
-RC
A3_
HC
h
-2,8
%
3,
6%
-0
,4%
-19,
2%
7,
3%
50
,7%
-6,1
%
-5
9,2%
-8,6
%
-8
3,0%
3,7%
109,
4%
HC
-Had
RM
3_H
Cn
10
,5%
78,1
%
10
,5%
10
9,7%
12,3
%
99
,6%
0,8%
45,4
%
9,
4%
82
,9%
8,2%
281,
4%
ETH
Z-C
LM_H
Cn
19
,0%
65,6
%
9,
3%
16
,7%
27,1
%
20
2,3%
0,6%
32,3
%
8,
0%
10
1,5%
10,8
%
93
,6%
D
MI-H
IRH
AM
_C
2,
5%
-2
2,2%
-2,1
%
-2
1,5%
15,8
%
84
,9%
-7,8
%
-2
5,7%
10,4
%
67,2
%
-1
,7%
-23,
4%
SMH
I-RC
A_B
17,6
%
-6
,2%
15,9
%
99,1
%
23
,0%
57,3
%
7,
9%
12
5,7%
-1
,7%
-24,
0%
4,
4%
10
,6%
M
PI-R
EMO
_E5_
3
5,3%
55,7
%
0,
0%
67
,8%
0,9%
-24,
1%
-3
,5%
-34,
4%
2,
8%
13
,2%
1,8%
81,2
%
DM
I-HIR
HA
M_E
5_3
15
,2%
34,1
%
12
,1%
39
,0%
12,7
%
-3
1,8%
-3,5
%
-1
4,5%
19,6
%
351,
7%
9,
1%
72
,3%
K
NM
I-RA
CM
O2_
E5_1
54,8
%
11
4,0%
8,
7%
6,
2%
63
,6%
288,
0%
-1
1,8%
8,
3%
20
,4%
20
,0%
39,6
%
25
2,2%
K
NM
I-RA
CM
O2_
E5_2
14,3
%
26
,7%
14,8
%
37,8
%
35
,4%
10,2
%
-5
,1%
60,0
%
-3
,7%
-40,
3%
9,
9%
10
,2%
K
NM
I-RA
CM
O2_
E5_3
33,6
%
27
4,6%
-1
,7%
-13,
4%
8,
5%
36
,7%
-7,9
%
-0
,9%
14,1
%
128,
7%
4,
3%
35
,1%
SM
HI-R
CA
_E5_
3
-4,5
%
-9
,9%
-3,9
%
-3
8,4%
-2,6
%
-1
7,3%
-9,8
%
-3
0,5%
-4,3
%
-1
1,2%
-2,6
%
-1
4,2%
15,1
%
55
,8%
5,7%
25,8
%
18
,5%
68
,8%
-4,2
%
9,
7%
6,
1%
55
,2%
8,0%
82,6
%
Ense
mbl
e ±
17,1
%
± 83
,8%
±
7,4%
±
49,9
%
±18
,6%
±99
,1%
±
5,6%
±
53,0
%
±9,
9%
±11
7,6%
±11
,4%
±
101,
2%
b) 2
021-
2050
with
out a
dapt
atio
n
Ger
man
y Fr
ance
U
K
Iber
ia
Pola
nd
Ben
elux
MEA
N
STD
M
EAN
ST
D
MEA
N
STD
M
EAN
ST
D
MEA
N
STD
M
EAN
ST
D
C4I
-RC
A3_
HC
h
-1,7
%
-3
2,1%
-1,8
%
-3
0,6%
8,4%
41,4
%
4,
3%
-3
1,4%
-5,8
%
-7
0,4%
1,9%
22,6
%
HC
-Had
RM
3_H
Cn
6,
6%
26
,5%
0,0%
-4,4
%
9,
0%
81
,8%
-3,5
%
-2
9,6%
6,7%
87,0
%
5,
7%
92
,4%
M
ETN
O-H
IRH
AM
_HC
n
-11,
7%
-5
7,9%
-10,
4%
-39,
5%
9,
2%
79
,9%
-7,6
%
-1
5,1%
-4,5
%
-4
7,8%
0,3%
31,2
%
ETH
Z-C
LM_H
Cn
0,
0%
-1
4,0%
1,4%
-7,1
%
21
,1%
122,
0%
-4,5
%
-2
3,9%
2,1%
7,1%
1,8%
6,4%
C
NR
M-R
M4.
5_C
1,9%
42,3
%
0,
4%
27
,0%
1,8%
11,5
%
1,
1%
35
,7%
2,2%
96,7
%
0,
4%
50
,6%
D
MI-H
IRH
AM
_C
3,
9%
-3
2,5%
0,8%
-3,1
%
25
,3%
49,3
%
-2
,0%
-13,
1%
-0
,9%
-16,
7%
-0
,9%
-10,
4%
MET
NO
-HIR
HA
M_B
-4,4
%
-4
7,4%
-3,0
%
-1
4,2%
15,5
%
-3
,6%
6,0%
24,5
%
3,
1%
44
,8%
-3,8
%
-6
2,7%
SM
HI-R
CA
_B
19
,9%
29,7
%
16
,8%
201,
5%
25,1
%
83
,6%
12,7
%
24
0,9%
-5
,7%
-48,
0%
5,
0%
-1
5,7%
M
PI-R
EMO
_E5_
3
0,1%
12,1
%
-3
,3%
4,4%
0,6%
-1,9
%
-1
,7%
-11,
5%
5,
5%
83
,3%
-2,8
%
-5
3,7%
D
MI-H
IRH
AM
_E5_
3
11,9
%
34
,3%
-0,8
%
-7
,9%
2,6%
-37,
3%
-3
,7%
-13,
2%
1,
9%
39
,3%
2,8%
74,8
%
KN
MI-R
AC
MO
2_E5
_1
32
,1%
5,0%
9,4%
-1,7
%
45
,5%
154,
3%
-18,
0%
-45,
1%
24
,1%
89,0
%
40
,7%
167,
3%
KN
MI-R
AC
MO
2_E5
_2
2,
3%
-8
,1%
28,2
%
15
1,0%
21
,9%
7,4%
0,5%
22,4
%
-8
,3%
-46,
9%
15
,4%
45,5
%
KN
MI-R
AC
MO
2_E5
_3
11
,4%
150,
3%
-0,9
%
23
,6%
-0,5
%
1,
3%
-8
,1%
-27,
6%
6,
6%
56
,1%
-4,9
%
-1
3,0%
SM
HI-R
CA
_E5_
3
-5,5
%
-1
1,0%
-4,4
%
-1
4,2%
-10,
4%
-29,
0%
-8
,5%
-28,
1%
0,
4%
-8
,0%
-8,9
%
-4
3,4%
4,8%
6,9%
2,3%
20,3
%
12
,5%
40
,0%
-2,3
%
6,
1%
2,
0%
19
,0%
3,8%
20,8
%
Ense
mbl
e ±
11,2
%
± 51
,7%
±
9,8%
±
69,0
%
±14
,3%
±57
,4%
±
7,4%
±
71,6
%
±8,
0%
±59
,2%
±
12,1
%
± 62
,9%
Spatio-temporal impact of climate change on the activityand voltinism of the spruce bark beetle, Ips typographus
A N N A M A R I A J O N S S O N *, G U S T A F A P P E L B E R G *, S U S A N N E H A R D I N G w and L A R S
B A R R I N G *, z*Department of Physical Geography and Ecosystems Analysis, Geobiosphere Science Centre, Lund University, Solvegatan 12, SE-
223 62 Lund, Sweden, wDepartment of Agriculture and Ecology, Faculty of Life Sciences, University of Copenhagen, Thorvaldsensvej
40, DK-1871 Frederiksberg C, Denmark, zRossby Centre, Swedish Meteorological and Hydrological Institute, SE-601 76
Norrkoping, Sweden
Abstract
The spruce bark beetle Ips typographus is one of the major insect pests of mature Norway
spruce forests. In this study, a model describing the temperature-dependent thresholds
for swarming activity and temperature requirement for development from egg to adult
was driven by transient regional climate scenario data for Sweden, covering the period of
1961–2100 for three future climate change scenarios (SRES A2, A1B and B2). During the
20th century, the weather supported the production of one bark beetle generation per
year, except in the north-western mountainous parts of Sweden where the climate
conditions were too harsh. A warmer climate may sustain a viable population also in
the mountainous part; however, the distributional range of I. typographus may be
restricted by the migration speed of Norway spruce. Modelling suggests that an earlier
timing of spring swarming and fulfilled development of the first generation will
significantly increase the frequency of summer swarming. Model calculations suggest
that the spruce bark beetle will be able to initiate a second generation in South Sweden
during 50% of the years around the mid century. By the end of the century, when
temperatures during the bark beetle activity period are projected to have increased by
2.4–3.8 1C, a second generation will be initiated in South Sweden in 63–81% of the years.
The corresponding figures are 16–33% for Mid Sweden, and 1–6% for North Sweden.
During the next decades, one to two generations per year are predicted in response to
temperature, and the northern distribution limit for the second generation will vary. Our
study addresses questions applicable to sustainable forest management, suggesting that
adequate countermeasures require monitoring of regional differences in timing of
swarming and development of I. typographus, and planning of control operations during
summer periods with large populations of bark beetles.
Keywords: forest damage, impact modelling, Sweden, temperature
Received 31 March 2008; revised version received 12 July 2008 and accepted 18 July 2008
Introduction
A variety of biological effects of the recent climate
warming have been observed as phenological changes
of plant and animal species across Europe (Menzel et al.,
2006). Insects are highly sensitive to changes in climate.
Their metabolic rate is dictated by ambient temperature,
and their activity and development therefore respond
strongly to even minor changes in temperature. Hence,
phenology and developmental rate may change in
response to changes in temperature and, for multivol-
tine insect species, the number of generations per year
may be affected (Ayres & Lombardero, 2000; Volney &
Fleming, 2000). Also, mobility and geographical distri-
bution of insect species may change as a result of global
warming, some species increasing their geographical
range, others moving their limits of distribution
towards north or into higher elevations in mountainous
regions (Williams & Liebhold, 2002; Carroll et al., 2003).
Factors such as fragmentation of vegetation and migra-
tion speed of vegetation may prevent or delay theCorrespondence: Anna Maria Jonsson, tel. 1 46 46 222 94 10,
temporal changes in the life cycle and voltinism of
I. typographus in a gradually changing climate. The
frequency of summer swarming and of completed
development of a second generation can increase due
to earlier spring swarming and faster development of
the first generation. The northern limit for a second
generation will vary between years in response to
temperature conditions. Model calculations indicated
that I. typographus will shift from univoltine to primarily
bivoltine in South Sweden and a viable population may
be established in the north-western mountainous parts.
The expansion of the distributional range of I. typogra-
phus may, however, be restricted by the migration speed
of Norway spruce. Changes in the timing of phenolo-
gical events will gradually affect the recommendations
for timely bark beetle management operations. For
adequate timing of countermeasures during periods
with large populations of bark beetles, regional mon-
itoring of swarming, development and probability of
bivoltinism will be required.
Acknowledgements
This work was carried out within the EU/FP6 ENSEMBLESproject (GOCE-CT-2003-505539). Anonymous reviewers arethanked for valuable comments to an earlier version of themanuscript.
References
Ahti T, Hamet-Ahti L, Jalas J (1968) Vegetation zones and their
sections in northwestern Europe. Annales Botanici Fennici, 5,
169–211.
Anderbrant O (1989) Reemergence and second brood in the bark
Jonsson AM, Harding S, Barring L, Ravn HP (2007) Impact of the
climate change on the population dynamics of Ips typographus
in southern Sweden. Agricultural and Forest Meteorology, 146,
70–81.
Jungclaus JH, Keenlyside N, Botzet M et al. (2006) Ocean circula-
tion and tropical variability in the coupled model ECHAM5/
MPI-OM. Journal of Climate, 19, 3952–3972.
Kjellstrom E, Barring L, Gollvik S et al. (2005) A 140-year simula-
tion of European climate with the new version of the Rossby Centre
regional atmospheric climate model (RCA3). Reports on Meteor-
ology and Climatology, No. 108, December 2005, Swedish
Meteorological and Hydrological Institute, Norrkoping, Swe-
den, 54 pp.
Lange H, Økland B, Krokene P (2006) Thresholds in the life cycle
of the spruce bark beetle under climate change. Interjournal for
Complex Systems, 1648, 1–10.
Langstrom B (2007) Bilaga 5 Svarmningskontroll av barkborrar
pa Asa och Tonnersjohedens forsoksparker. In: Overvakning av
Insektsangrepp – Slutrapport fran Skogsstyrelsens Regeringsupp-
drag (ed. Svensson L), pp. 44–57, Skogsstyrelsen Meddelanden
1:2007. Skogsstyrelsen, Jonkoping, Sweden.
Logan JA, Regniere J, Powell JA (2003) Assessing the impact of
global warming on forest pest dynamics. Frontiers in Ecology
and Environment, 1, 130–137.
Loman JO (ed.) (2008) Statistical Yearbook of Forestry 2008. Official
Statistics of Sweden, Swedish Forest Agency, J o nk o ping.
Menzel A, Sparks TH, Estrella N et al. (2006) European pheno-
logical response to climate change matches the warming
pattern. Global Change Biology, 12, 1969–1976.
Nakicenovic N, Swart R (eds) (2000) Emission Scenarios, a
Special Report of Working Group III of the Intergovernmental Panel
on Climate Change. Cambridge University Press, Cambridge, UK.
Netherer S, Pennerstorfer J (2003) Parameters relevant for mod-
eling the potential development of Ips typographus (Coleoptera:
Scolytidae). Integrated Pest Management Reviews, 6, 177–184.
Økland B, Berryman A (2004) Ressource dynamics plays a key
role in regional fluctuations of the spruce bark beetles Ips
typographus. Agricultural and Forest Entomology, 6, 141–146.
Roeckner E, Bengtsson L, Feichter J, Lelieveld J, Rodhe H (1999)
Transient climate change simulations with a coupled atmo-
sphere-ocean GCM including the tropospheric sulfur cycle.
Journal of Climate, 12, 3004–3032.
Schelhaas MJ, Nabuurs GJ, Schuck A (2003) Natural distur-
bances in the European forests in the 19th and 20th centuries.
Global Change Biology, 9, 1620–1633.
Schopf A (1989) The effect of photoperiod on the induction of the
imaginal diapause of Ips typographus (L.) (Col., Scolytidae).
Journal of Applied Entomology, 107, 275–288.
Seidl R, Baier P, Rammer W, Schopf A, Lexer M (2007) Modelling
tree mortality by bark beetle infestation in Norway spruce
forests. Ecological Modelling, 206, 383–399.
Strand JF (2000) Some agrometeorological aspects of pest and
disease management for the 21st century. Agricultural and
Forest Meteorology, 103, 73–82.
Tragardh I, Butovitch V (1935) Redogorelse for barkborrekam-
panjen efter stormharjningarna 1931–1932. Meddelanden fran
Statens Skogsforsoksanstalt, 28, 1–268.
Uppala SM, Kallberg PW, Simmons AJ et al. (2005) The ERA-40
re-analysis. Quarterly Journal of the Royal Meteorological Society,
131, 2961–3012.
498 A . M . J O N S S O N et al.
r 2008 The AuthorsJournal compilation r 2008 Blackwell Publishing Ltd, Global Change Biology, 15, 486–499
van Ulden AP, van Oldenborgh GJ (2006) Large-scale atmo-
spheric circulation biases and changes in global climate model
simulations and their importance for climate change in
Central Europe. Atmospheric Chemistry and Physics, 6,
863–881.
Volney WJA, Fleming RA (2000) Climate change and impacts of
boreal forest insects. Agriculture, Ecosystems and Environment,
82, 283–294.
Wermelinger B (2004) Ecology and management of the spruce
bark beetle Ips typographus – a review of recent research. Forest
Ecology and Management, 202, 67–82.
Wermelinger B, Seifert M (1998) Analysis of the temperature
dependent development of the spruce bark beetle Ips typogra-
phus (L.) (Col., Scolytidae). Journal of Applied Entomology, 122,
185–191.
Wermelinger B, Seifert M (1999) Temperature-dependent repro-
duction of the spruce bark beetle Ips typographus, and analysis
of the potential population growth. Ecological Entomology, 24,
103–110.
Williams DW, Liebhold AM (2002) Climate change and the
outbreak ranges of two North American bark beetles. Agricul-
tural and Forest Entomology, 4, 87–99.
S PAT I O - T E M P O R A L I M PA C T O F C L I M A T E C H A N G E O N I P S T Y P O G R A P H U S 499
r 2008 The AuthorsJournal compilation r 2008 Blackwell Publishing Ltd, Global Change Biology, 15, 486–499
Ih
Aa
b
a
ARRAA
KNPLBPFHCC
1
tib7miwpc
S
0d
Ecological Modelling 221 (2010) 303–313
Contents lists available at ScienceDirect
Ecological Modelling
journa l homepage: www.e lsev ier .com/ locate /eco lmodel
mpacts of changing frost regimes on Swedish forests: Incorporating coldardiness in a regional ecosystem model
. Rammig a,∗, A.M. Jönsson a, T. Hickler a, B. Smith a, L. Bärring a,b, M.T. Sykes a
Geobiosphere Science Centre, Department of Physical Geography and Ecosystems Analysis, Lund University, Sölvegatan 12, SE-22362 Lund, SwedenSMHI Rossby Centre, Swedish Meteorological and Hydrological Institute, SE-60176 Norrköping, Sweden
r t i c l e i n f o
rticle history:eceived 27 October 2008eceived in revised form 5 May 2009ccepted 9 May 2009vailable online 21 June 2009
Understanding the effects of climate change on boreal forests which hold about 7% of the global terres-trial biomass carbon is a major issue. An important mechanism in boreal tree species is acclimatization toseasonal variations in temperature (cold hardiness) to withstand low temperatures during winter. Tem-perature drops below the hardiness level may cause frost damage. Increased climate variability underglobal and regional warming might lead to more severe frost damage events, with consequences fortree individuals, populations and ecosystems. We assessed the potential future impacts of changing frostregimes on Norway spruce (Picea abies L. Karst.) in Sweden. A cold hardiness and frost damage modelwere incorporated within a dynamic ecosystem model, LPJ-GUESS. The frost tolerance of Norway sprucewas calculated based on daily mean temperature fluctuations, corresponding to time and temperaturedependent chemical reactions and cellular adjustments. The severity of frost damage was calculated asa growth-reducing factor when the minimum temperature was below the frost tolerance. The hardinessmodel was linked to the ecosystem model by reducing needle biomass and thereby growth according tothe calculated severity of frost damage. A sensitivity analysis of the hardiness model revealed that theseverity of frost events was significantly altered by variations in the hardening rate and dehardening rateduring current climate conditions. The modelled occurrence and intensity of frost events was related to
observed crown defoliation, indicating that 6–12% of the needle loss could be attributed to frost dam-age. When driving the combined ecosystem-hardiness model with future climate from a regional climatemodel (RCM), the results suggest a decreasing number and strength of extreme frost events particularly innorthern Sweden and strongly increasing productivity for Norway spruce by the end of the 21st centuryas a result of longer growing seasons and increasing atmospheric CO2 concentrations. However, accord-ing to the model, frost damage might decrease the potential productivity by as much as 25% early in thecentury.
. Introduction
Boreal forests cover up to 14.7 Mio km2, which is about 11% ofhe earth’s land surface (Bonan and Shugart, 1989). They play anmportant role in global climate by storing about 42 Gt of Car-on in biomass and 200 Gt C in soil organic matter representing.5–10% of the global terrestrial amounts (Jarvis et al., 2001). Cli-ate change will affect these ecosystems not only through changes
n mean conditions – such as impacts of changing temperature,ater availability and atmospheric CO2 concentrations on plantroduction – but also via changes in the frequency and level oflimatic extremes (IPCC, 2007), such as changes in frost regimes,
prolonged water stress or drought, or damaging windstorms. Stud-ies with global models point to the likelihood of increased climaticvariability under future greenhouse forcing (IPCC, 2007). A hypoth-esis is that this could lead to an increased frequency of weatherevents beyond plant physiological tolerance thresholds, potentiallyresulting in stress, damage, mortality and changed biogeochemicalcycling in ecosystems.
Cannell and Smith (1986) presented the seemingly paradoxicalhypothesis that climatic warming in the boreal and temperate zoneswill cause increased frost damage due to dehardening or growthonset of trees during mild spells in winter and early spring, leadingto frost damage in subsequent cold periods. Increased frost dam-
age could reduce the positive effect of an extended growing periodand elevated atmospheric CO2 concentrations on forest growth(Woldendorp et al., 2008). In Norway spruce (Picea abies L. Karst.),frost is harmful when the ambient temperature falls below the“hardiness level” (Jönsson, 2005).
Hardiness is the ability of plant cells to tolerate cellular freezingnd occurs during the annual cycle of trees during acclimation toold temperatures. It is controlled by an additive effect of environ-ental factors, such as photoperiod and temperature (Chen and
i, 1978). During the active growth phase, the trees do not havehe potential for hardening. After growth cessation, the hardeningrocess is initiated (Kellomäki et al., 1995; Bigras and Colombo,000) and the trees use their energy resources to develop a freezeolerance (cf. Fuchigami et al., 1982; Harrison et al., 1978). Theehardening process starts with increasing temperatures (Bigrasnd Colombo, 2000).
Seedlings and newly developed shoots are particularly sensitiveCannell and Sheppard, 1982; Sakai and Larcher, 1987), but frostpisodes may also damage needles, bark and roots in mature trees.eversible damage of needles causes ion leakage due to disrup-ion of membrane transport functions and the recovery costs waternd energy (e.g. Sutinen et al., 2001; Linder and Flower-Ellis, 1992;urr et al., 2001). Damaged trees are prone to subsequent attacksy fungal pathogens, which may cause heavy crown defoliation orven kill the tree (Alden and Hermann, 1971; Schoeneweiss, 1975;arlman, 1986; Kowalski, 1991, 1996; Sieber et al., 1995). Jönssont al. (2004) showed that an earlier start of the vegetation periodaused by a warmer climate can increase the risk for spring frostamage on mature Norway spruce (P. abies L. Karst.) in southernweden. This hypothesis is also supported by electrolyte leakagexperiments (Repo et al., 1996) and model simulations for Scotsine (Pinus sylvestris L.) and other tree species in Finland (Kellomäkit al., 1995; Leinonen, 1996; Hänninen et al., 2001). However, Ögren2001) concluded from carbohydrate measurements after freezingreatments that the seasonal hardening and dehardening cyclesnd cold hardiness levels of Norway spruce, Scots pine and lodge-ole pine (Pinus contorta ssp. latifolia Loud.) would be unaffectedy global warming.
Most modelling studies of climate effects on tree frost hardinessave been very detailed in their description of the physiologicalechanisms of the investigated tree species over the annual cycle
e.g. Kellomäki et al., 1995; Leinonen, 1996). However, the modelsave to date only been applied at the site scale under simplifiedlimate change projections, such as a linear increase in mean tem-erature (Cannell and Smith, 1986; Kellomäki et al., 1995; Leinonen,996). Impacts of climate and atmospheric changes on ecosystemunctioning at regional scales may be investigated with the helpf process-based ecosystem models (e.g. Bergh et al., 2003; Kocat al., 2006; Morales et al., 2007). Current models include detailedormulations of physiological processes linking plant gas exchange,rowth, phenology, and neighbourhood interactions to gradualhanges in climate. Mechanisms of response to daily temperatureuctuations, including variations in frost hardiness, are, however,ot included in many current models, or are represented in sim-listic ways that are insufficient to capture the detailed ecosystemesponse. Many studies also use monthly mean values of climatearameters as input to the ecosystem model, so that thresholds foramage or reduced functioning, such as lethal temperatures, mayever be crossed (e.g. Hickler et al., 2004; Morales et al., 2007; Kocat al., 2006).
Thanks to developments in climate modelling, it is now possibleo obtain driving climate data for ecosystem model studies with aigh spatial resolution and realistic characterisation of diurnal vari-tion and extremes of temperature. Regional climate models (RCMs,.g. Christensen et al., 2007; Jacob et al., 2007) describe processeseading to seasonally and regionally non-uniform warming trends
n a more realistic manner compared to coarser-resolution global
odels (general circulation models, GCMs). For example, RCM sim-lations over northern Europe suggest that regional warming mayeduce the seasonal snow cover resulting in a positive albedo feed-ack that enhances the wintertime warming trend (Kjellström,
elling 221 (2010) 303–313
2004). The more limited spatial averaging that occurs in RCMs com-pared to GCMs may result in a better description of possible changesin extremes, such as low temperatures, under greenhouse forcingscenarios (Kjellström et al., 2005, 2007).
In this study, we aimed to address two hypotheses: (1) regionalclimate change will alter the risk for frost damage in mature Nor-way spruce, as a major dominant species of boreal forest in Sweden.(2) The productivity of spruce-dominated forests will be influencedby changes in the risk for frost damage under future climate con-ditions. In order to investigate these hypotheses, we incorporateda detailed physiological model of frost hardiness (Jönsson et al.,2004) and frost damage (Kellomäki et al., 1995; Leinonen, 1996) thataccounts for autumn, mid-winter and spring frost events within anexisting process-based ecosystem model, LPJ-GUESS (Smith et al.,2001). This is a step towards the development of tools for assess-ing the effects of changing frost regimes on forest productivity atthe regional scale. In this paper, we focus on model developmentand evaluation. The outcomes of our study provide first estimatesof changes in the risk of frost damage. We present model generatedhypotheses on the potential impacts of frost damage on the pro-ductivity of Norway spruce in different regions of Sweden underfuture climate conditions, based on RCM-generated high-resolutiontransient daily climate data.
2. Methods
An existing model to calculate the daily frost hardiness (Jönssonet al., 2004) and frost damage (Kellomäki et al., 1995; Leinonen,1996) accounting for autumn, mid-winter and spring frost eventsin Norway spruce was tested and incorporated within the process-based ecosystem model LPJ-GUESS (Smith et al., 2001; Koca et al.,2006). Model performance was evaluated by comparing simulationresults to observed data on needle loss and stand productivity.
2.1. Model descriptions
2.1.1. Ecosystem model (LPJ-GUESS)The generalized ecosystem model LPJ-GUESS (Smith et al., 2001)
combines the mechanistic representations of plant physiologicaland biogeochemical processes of the Lund-Potsdam-Jena DynamicGlobal Vegetation Model (LPJ-DGVM; Sitch et al., 2003) with adetailed description of vegetation structure and dynamics, simi-lar to forest gap models such as FORSKA2 (Prentice et al., 1993). Fortree individuals, the model simulates photosynthesis, respirationand allocation of annually accrued carbon (net primary production,NPP) to leaves, fine roots, sapwood, heartwood and reproductiveorgans, accounting for seasonal changes in needle phenology andadjustments in stomatal conductance. Neighbouring individualscompete for uptake of light and soil water. Population dynamics(establishment and mortality) are influenced by current resourcestatus, demography and the life history characteristics of each sim-ulated species or plant functional type (PFT). In this study, themodel was set up only to simulate growth and dynamics of even-aged stands of Norway spruce (parameters settings see Table 1, allother parameters were inherited from Smith et al. (2001), Sitchet al. (2003), Hickler et al. (2004) and Koca et al. (2006)). Inputdata to the model are daily or monthly mean values of climateparameters (temperature (◦C), precipitation (mm) and incomingshortwave radiation (Wm−2)), atmospheric CO2 concentrations anda soil texture class that governs soil hydrology and heat conduc-
tance. LPJ-GUESS has been applied to simulate north European orboreal vegetation in several previous studies (Badeck et al., 2001;Smith et al., 2001, 2008; Hickler et al., 2004; Koca et al., 2006;Morales et al., 2005, 2007; Zaehle et al., 2006) and has been evalu-ated, for example, using data on growth (Zaehle et al., 2006; Smith
A. Rammig et al. / Ecological Modelling 221 (2010) 303–313 305
Table 1Plant functional type and species parameter settings for simulations of Norwayspruce stands with LPJ-GUESS, all other parameters were inherited from Smith et al.(2001), Hickler et al. (2004) and Koca et al. (2006).
Parameter Unit Value Source
Tree specific parametersCarbon density of sapwood
and heartwood in treeskgC m−3 250 Jaakkola et al. (2006)
Fraction of roots in upper(0–50 cm) and lower(50–150 cm) soil layer
Dimensionless 0.9/0.1 Köstler et al. (1968)
Needleleaf specific parametersLeaf area to sapwood
cross-sectional area ratiom2 m−2 3500 Köstner et al. (2002)
Leaf longevity Years 4 Niinemets andLukjanova (2003)
Fine root turnover Year−1 0.7 Vogt et al. (1996), Li etal. (2003)
Shadetolerant specific parametersSapwood conversion rate Year−1 0.05 Bartelink (2000)
Norway spruce specific parametersExpected longevity under Years 500 Bugmann (1994)
ea
2
fbd“id
Table 2Parameters for the frost hardiness and frost damage model.
Parameters forstandard runs
Description Value and unit
Parameters for hardiness modelHmin Minimum hardiness levela −2 ◦CHmax Maximum hardiness levelc −30 ◦CSaut Start of autumn (start of
hardening)aJulian day 210
Sspr Start of spring (start ofdehardening) for SouthernSwedena
Julian day 1
H∗t Target hardiness levela F (daily mean temperature)
r∗h
Rate of hardeninga 0–1 ◦C/dayr∗
dhRate of dehardeninga 0–5 ◦C/day
Wd Winter dormancya From days 260 to 365
Parameters for calculation of the growth reducing factorb Slope parameterb 0.2 ◦C−1
LT50 “Lethal temperature”c 20 ◦C
where max(value 1, value 2) is a function that selects the maxi-
F(h
lifetime non-stressedconditions
t al., 2008) and carbon fluxes (Morales et al., 2005; Wramneby etl., 2008) for European forests.
.1.2. Hardiness modelCalculations of the hardiness level of Norway spruce were per-
ormed using an updated version of the hardiness model developedy Jönsson et al. (2004). In the hardiness model (Fig. 1B), the har-
◦
iness level Hday ( C) is adjusted on a daily basis, converging to atarget” hardiness level Ht (Fig. 1C, Table 2) according to a harden-ng or dehardening rate (rh or rdh; Fig. 1D and E, Table 2) based onaily mean temperature Tmean (◦C; Fig. 1B). Hday during the summer
ig. 1. (A) Distribution of the study sites (black dots) over Sweden showing forestry adm55.6–57.9◦N), “Central” (58.0–61.8◦N) and “North” (62.2–67.8◦N) regions. (B) Example ofardiness level. (C–E) Target hardiness (in ◦C), hardening and dehardening rates (in ◦C day
a Values from Jönsson et al. (2004).b Values from Kellomäki et al. (1995).c Values from Bigras and Colombo (2000).
is calculated as
Hday(day + 1) = Hmin
{if Hday(day) + rdh < Hmin
if aggd5 ≥ 120◦C and if day ≤ Saut
(1)
where day is the day of year, Hmin is the minimum hardiness, Saut isthe day of start of autumn (Table 2) and aggd5 are the accumulatedgrowing degree days (◦C), calculated as
aggd5 = aggd5 + max(0, Tmean − 5 ◦C) (2)
mum from two values. Bud burst in Central Sweden occurs after120 degree-days above 5 ◦C (Hannerz, 1994). At the first day of theyear, aggd5 is set to 0 ◦C. During springtime (if day < Saut and if Eq.(1) is not true), then both, hardening and dehardening is possible
inistrative regions; for some analyses the study sites were grouped into “South”a 1-year mean and minimum temperature curve for one site and the corresponding−1) in relation to the ambient mean temperature.
During winter (Wd, Table 2), only hardening is possible and Hdays calculated according to Eq. (4). The hardiness can not fall belowhe maximum hardiness (Hmax, Table 2) and thus
day(day + 1) = Hmax if Hday(day + 1) > Hmax (6)
The daily difference Dday (◦C) between the calculated hardinessevel Hday and the minimum temperature Tmin (◦C).
day = Hday − Tmin,day (7)
as taken to be the main factor determining the degree of frostamage to the trees (Fig. 1B). At a day with Dday > 0, the trees weressumed to experience a frost event. We counted the days whereday > 0 and defined this as the frequency of frost events FFE (num-er of days):
FEyear =365∑d=1
{1 if Dday > 00 if Dday ≤ 0
(8)
Additionally, the size of the frost event SFEday (◦C) was defineds
FEday = max(0, Dday) (9)
hich is the actual amount of the difference Dday if the minimumemperature falls below the hardiness level (Fig. 1B).
.1.3. Frost damage modelBased on the hardiness level and current temperature, a “growth
educing factor” (gfday) was calculated in LPJ-GUESS for each day.he growth reducing factor gives the relative effect of freeze dam-ge for each difference between the hardiness level and minimumemperature Dday (Eq. (1)) according to a logistic function:
fday = 11 + exp(b(Dday − LT50))
(10)
here b is the slope parameter, and Dday is the difference betweenhe hardiness level of the tree and the minimum temperature athe specified day. Thus, Dday was taken as a proxy for the length ofhe cold episode and thus determines the degree of the frost dam-ge. The parameter LT50 (◦C) is defined as the “lethal temperatureifference between the hardiness level and the minimum temper-ture” at which 50% of the trees are damaged, and determines thenflection-point of the curve, depending on the Dday. For the pur-oses of this study, we set LT50 to 20 ◦C, i.e. with Dday = 20 ◦C, 50%f the needles would be killed (Table 2). The slope parameter bas assumed to be 0.2 for the standard simulations. The growth
educing factor is dimensionless and ranges between 0.0 and 1.0,here values <0.1 signify strong damage and values >0.9 signify lit-
le damage (modified after Kellomäki et al., 1995). It was assumedhat frost damage primarily affects trees by causing needle necrosisnd a reduction in carbon assimilation in proportion to the needlesilled (Bigras and Colombo, 2000). The frost damage model thenelates needle loss to the growth reducing factor by using the min-mum daily value obtained for gfday during a simulation year, gfmin,o determine the annual amount of frost damage (modified after
einonen, 1996):
fmin = min(365d=1gfd) (11)
It was thus assumed that the maximum temperature differenceetween the daily minimum temperature and the daily hardiness
elling 221 (2010) 303–313
level on any particular day is an adequate predictor for the frostdamage for that year (Leinonen, 1996). Frost damage was imple-mented at the end of each simulation year based on the gfmin-value:the leaf carbon mass, sapwood mass and accrued NPP of eachaverage individual tree (representing average properties of the sim-ulated trees within a patch) were multiplied by gfmin. Killed needleswere transferred to the litter pool, and killed sapwood to heart-wood. It was assumed that roots were not affected by frost damage.
2.2. Data for model evaluation
Data on crown defoliation from 1999 to 2005 from 122 man-aged Norway spruce stands were obtained from the Swedish ForestAgency. These data were collected in accordance with ICP forestmonitoring (Eichhorn et al., 2006). The site locations range in lat-itude from 51.7 to 67.5◦N, spanning most forest areas in Sweden(Fig. 1A). In these data, crown defoliation is defined as the annualneedle loss, in 5% intervals, relative to a tree of the same type(species, age, size) with full foliage (Eichhorn et al., 2006), andranged between 0 and 35% for the observed forests. The observa-tions on crown defoliation were carried out annually by comparingthe selected tree to a “reference tree”, which is defined as “the besttree with full foliage that could grow at a particular site. . . [with]0% defoliation” (Eichhorn et al., 2006). The density of the observedstands was on average 740 trees ha−1 and average stand age 59years. Observed data of forest stand productivity in Sweden (mea-sured as mean annual stem volume increment, m3 ha−1 yr−1) wereobtained from the Swedish Forest Agency (2007) for the period2001–2005 for the 19 forestry administrative regions of Sweden.
2.3. Environmental driver data
Global atmospheric CO2 concentrations derived from ice-coremeasurements and atmospheric observations (Sitch et al., 2003)were used to drive the model for the historical period of the sim-ulations (see Section 4 below). For the period from 1999 to 2100,projections of CO2 concentrations were used (Joos et al., 2001).
Climate data for the ecosystem model runs were derived fromthree sources: (1) MESAN/ERA40, which is high-resolution (11 km)gridded meteorological data obtained from the mesoscale mete-orological analysis system MESAN (Häggmark et al., 2000) set upto use ERA-40 reanalysis data (Uppala et al., 2005) in combinationwith quality-controlled daily observations from weather stations(Jansson et al., 2007); (2) RCA3/ERA40 are output data from theSMHI Rossby Centre regional climate model RCA3 run for the cli-mate of Europe over the historical period 1961–2004 forced byERA-40 reanalysis data (Uppala et al., 2005) at the lateral bound-aries; and (3) RCA3/ECHAM4/A2 are output data from RCA3 froma simulation covering the period 1961–2100 (Kjellström et al.,2005) with lateral boundary conditions taken from an experimentwith the ECHAM4/OPYC3 general circulation model (Roeckner etal., 1999) forced by the SRES A2 emission scenario. The SRES A2assumes a “regional-economic” development of the future world(Nakicenovic et al., 2000). For both (2 and 3), RCA3 was configuredto have a spatial resolution of 0.5◦ (ca. 50 km) and it was operated ona time step of 30 min. The basic characteristics of RCA3 were eval-uated by Kjellström et al. (2005) and the new land surface schemeis presented by Samuelsson et al. (2006). For all three data sources,the data used were average daily values of temperature (◦C), precip-itation (mm) and total downward shortwave radiation at the landsurface (Wm−2) as well as daily minimum temperature (◦C), all on
a 0.5 × 0.5◦ regular grid covering Sweden.
Because the frost hardiness model is critically sensitive to theinput data for minimum daily temperature (Eq. (7)), it was impor-tant to correct for any bias in the mean and minimum temperaturedata. Three sources of bias were considered: bias that could arise
l Modelling 221 (2010) 303–313 307
fealdqambto‘Rptm(
2
pofaftaasf
2
icmwrutcp
2
upctRi
2
2
upv(eiHHi
Table 3Parameter variations in the hardiness and frost damage model for the sensitivityanalysis.
Simulation Parameter settings
Sensitivity analysis for the hardiness modelH1 StandardH2 Hardening rate +20%H3 Hardening rate −20%H4 Dehardening rate +20%H5 Dehardening rate −20%H6 “Target” hardiness level −50 ◦CH7 Start of autumn day 220 (∼August 8),
daylight ca. 15.2 (S)–16.5(N) hH8 Start of autumn day 230 (∼August 18),
daylight ca. 14.5 (S)–15.5(N) hH9 Start of autumn ca. day 237 (∼August 25;
S)–244 (∼September 1; N), daylight 14 h
Sensitivity analysis for the frost damage modelFNO No frost damageF1 Frost damage model, b = 0.2, LT50 = 20F2 Frost damage model, b = 0.1, LT50 = 20
A. Rammig et al. / Ecologica
rom RCA3, from ECHAM4/OPYC3, and from the up-scaling of mod-lled and observed data to the 0.5 × 0.5◦ grid. Dataset (1) wasssumed to be free from bias. Thus as a ‘Control Period I’, the over-apping period between the MESAN/ERA40 and the RCA3/ERA40atasets from 1998 to 2004 has been used. For this period we canuantify the bias in RCA3 versus observed climate conditions. Welso need to correct for the GCM bias, which is a combination ofodel bias (model imperfections) and natural variability induced
y the initial conditions and the fact that a climate scenario simula-ion is not constrained by observed weather conditions during thebserved reference period. For this purpose we use the overlapping
Control Period II’ from 1961 to 2004 between the RCA3/ERA40 andCA3/ECHAM4/A2 data has been used. To correct the mean tem-erature, a standardization method has been used. The minimumemperature has been corrected using the difference between the
ean and the minimum temperature, and an arithmetic methodSupplementary data).
.4. Modelling protocol
For our study, we applied the model to simulate even-aged,lanted Norway spruce stands at the 122 sites in Sweden for whichbserved frost damage data were available (Fig. 1). Climate dataor the grid cell encapsulating the observed site were used. Standge, density (trees ha−1) and disturbance history were prescribedor the simulation runs in order to get a realistic representation ofhe forest for the study period. For example, a stand with a recordedge of 75 years in 2005 was initialized (planted with spruce treest the density of the observed stand) in the simulation year corre-ponding to calendar year 1930. Tree regeneration was switched offor the remainder of the simulation.
.4.1. Model evaluation runsSimulations under current climate conditions were performed
n order to evaluate the performance of the model. The bias-orrected RCA3/ECHAM4/A2 climate data were used to drive theodel for the period 1930–1990. The simulations then continuedith input from the 8 years of MESAN-ERA40 data, which were
epeated twice. The first 8 years (1990–1998) were used as a “spinp” period in order to achieve an approximate steady state betweenhe simulated forest stand and the climate, the second 8 yearsorresponded to the actual period from 1998 to 2005 (evaluationeriod).
.4.2. Future scenario runsTo explore changes in frost damage and stand productivity
nder future climate conditions at each stand, simulations wereerformed with input from the bias-corrected RCA3/ECHAM4/A2limate data for the entire period 1961–2100. Climate data beforehis period were constructed by repeating the first 10 years of theCA3/ECHAM4/A2 climate dataset the number of times required to
nitialise each stand in the year it was actually planted.
.5. Analysis of model behaviour and performance
.5.1. Sensitivity of the simulated frost hardiness and frost damageTo test the sensitivity of the hardiness model, a range of sim-
lation experiments with different model configurations wereerformed for 1999–2005: simulation H1 corresponds to theersion with standard settings (Table 2). In simulations H2–H9Table 3), only one parameter at a time (OAT-experiments, Saltelli
t al., 2000) was varied. Hardening and dehardening rate was var-ed by +/−20% around the standard value (H2–H5). In simulation6, the maximum hardiness was set to −50 ◦C. In simulations7–H9, the start of autumn was retarded by 10–14 days. Accord-
ngly, changes in the main output variables of the hardiness model,
F3 Frost damage model, b = 0.3, LT50 = 20F4 Frost damage model, b = 0.2, LT50 = 10F5 Frost damage model, b = 0.2, LT50 = 30
namely derivates of FFE (frequency of frost events per year, whenminimum temperature drops below the hardiness level, Eq. (8)) andSFE (size of frost event, Eq. (9)) were analyzed over the simulationperiod (start = 1999, end = 2005, nyears = 6):
- The frequency of frost events over the simulation period.
SumFFE =end∑
y=start
FFEy (12)
- The average of the size of a maximum frost event.
AvSFEmax =∑end
y=startSFEmax,y
nyears(13)
where SFEmax is the largest value of SFE for any day in year y.The results of SumFFE and AvSFEmax were evaluated for the rangeof simulation experiments by ANOVA and pairwise comparisons(unpaired t-test; R statistical software).
The sensitivity of the frost damage model was also evaluatedby a range of simulation experiments, varying the parameters thatdetermine the growth reducing factor (gfday, Eq. (10)), and therebythe needle carbon mass, sapwood mass and accrued NPP of eachaverage individual tree: simulation F1 corresponds to the standardversion with b = 0.2 (Table 3, Kellomäki et al., 1995) and LT50 = 20 ◦C(Bigras and Colombo, 2000). The slope parameter b of the frost dam-age model (Eq. (10)) was varied between 0.1 and 0.3 (simulationsF2 and F3, Table 3). The parameter LT50, which denotes the pointwhere 50% of the needles are damaged, was varied between 10 and30 ◦C (simulation F4 and F5). To quantify the effect of the parametervariations, the “simulated crown defoliation” was used. To calculatesimulated crown defoliation we used the average simulated valuefor needle carbon biomass (kgC m−2) for the period 1999–2005. Theneedle carbon biomass of simulation FNO (no frost damage, Table 3)was set to 100%. Simulated crown defoliation was calculated forsimulations F1–F5 as the average needle carbon mass expressed asa percentage of the value from the FNO simulation.
2.5.2. Evaluation of the ecosystem model performanceIn order to evaluate the skill of the ecosystem model, we com-
pared simulated stem wood volume increments (m3 ha−1 yr−1)with average values for Swedish forestry administrative regions
308 A. Rammig et al. / Ecological Modelling 221 (2010) 303–313
F ., 200s reque( s the um uartil
baifww2at(1
uctoaiowddnsntdmduo
2c
o(a
ig. 2. Results of the sensitivity analysis of the frost hardiness model (Jönsson et alites. The simulation runs H1–H9 are described in Table 3. Given are the sum of the fAvSFEmax , Eq. (13)). The black line within the box denotes the median, the box give
inimum and maximum of the distribution within the range of 1.5 times the interq
y grouping the 122 sites into 21 forestry administrative regionsnd computing an average value for each region. Stem volumencrement was estimated from carbon mass increment obtainedrom LPJ-GUESS simulations. For conversion of carbon mass to stem
ood volume, it was assumed that 65% of the total carbon massould contribute to the actual stem wood volume (Koca et al.,
006). A wood density of 250 kgC m−3 was assumed (Jaakkola etl., 2006). Simulated and observed volume increments from 2001o 2005 were compared by linear regression, modelling efficiencyEF) and root mean square error (RMSE; after Mayer and Butler,993) among forestry regions.
Additionally, to estimate whether the ecosystem model sim-lates the appropriate magnitude of frost crown defoliation, weompared the simulated frost crown defoliation, as defined above,o the observed crown defoliation for each study site. Since thebserved crown defoliation may be related to several factors notccounted for in the frost damage model, such as tree age, acid-fication, and drought effects, we had to filter out the proportionf crown defoliation that is caused by extreme frost events. Thisas done by estimating the proportion of the total observed crownefoliation that can be explained by the average and the maximumifference between minimum daily temperatures and tree hardi-ess level during a frost event (AvSFEmax; see Eq. (13)) for eachite. This proportion was quantified as the coefficient of determi-ation for a simple linear regression model fitted to annual data onhe crown defoliation (dependent variable) and AvSFEmax (indepen-ent variable). Observed crown defoliation at each site was thenultiplied by this proportion to obtain an estimate of the crown
efoliation attributable to extreme frost events at that site. Sim-lated crown defoliation values were compared to these adjustedbservations to evaluate the model performance.
.6. Frost events and forest productivity under future climateonditions
Model output from the future scenario simulations was averagedver 30 year periods: 1976–2005 (current conditions), 1981–2010simulation period 1), 2011–2040 (period 2), 2041–2070 (period 3),nd 2071–2100 (period 4). Changes in frost regimes were charac-
4), driven by MESAN/ERA40 data for the period from 1999 to 2005 at all 122 studyncy of frost events (SumFFE, Eq. (12)) and the average size of a maximum frost eventpper and lower quartile, which contains 50% of the data and the whiskers give thee distance from the median. Outliers are marked with a circle.
terised by the average number of frost events per year
AvFFE = SumFFE
nyears(14)
(derived from Eq. (12)), and the average annual maximum size offrost events (AvSFEmax, Eq. (13)). Values of each of these indiceswere compared for simulation period 1–4. The frost damage model(Table 2) was characterised by comparing stand productivity forsimulation period 1–4 to productivity under current conditions.
3. Results
3.1. Evaluation of the hardiness model
The hardiness model was significantly sensitive to variationsin the hardening rate (Fig. 2, ANOVA: p < 0.001 for SumFFE andAvSFEmax). The frequency and the size of frost events (SumFFE andAvSFEmax, Fig. 2) were significantly lower with an increased hard-ening rate (simulation H2, Table 3, pairwise comparison: p < 0.001),and significantly higher with a decreased hardening rate (simula-tion H3) compared to the standard simulation runs (simulation H1,p < 0.001). The hardiness model showed only weak sensitivity for anincreased dehardening rate (H4; p = not significant). The hardinessmodel was not sensitive to changes in the “target” hardiness (H6)and in the start of autumn (H7–H9).
The average maximum frost event (AvSFEmax, Eq. (13)) explained12% of the variability in the observed average crown defoliationfrom 1999 to 2005 (R2 = 0.12, df = 120, F = 16.30, p < 0.001; otherexplanatory variables such as tree age, stand density, soil water con-tent and soil type were tested in multiple regression analysis butdid not significantly change the predictive power of the variableAvSFEmax).
3.2. Evaluation of the frost damage model
Observed crown defoliation for 1999–2005 was on average11.2% (Stdev: 5.9%). As described above, the simulated frost eventsexplained 12% of the variability in the observed crown defoliationover the period from 1999 to 2005. This result corresponded wellto Nihlgård (1990), who reported 5–15% of needle loss in Norway
A. Rammig et al. / Ecological Mod
Fig. 3. Evaluation of the frost damage model. Given is the average crown defoliation(%) for the time period 1999–2005. On the x-axis, “OBS” denotes 10% of the observedtptF
sacctiaald
Fa(
otal crown defoliation. Simulations F1–F5 show the results for variations in thearameters b and LT50 for the frost model (using gfmin). Simulation F1 is assumedo be the “standard” versions of the frost damage model. Details of the boxplots seeig. 2.
pruce forest stands in Scania during 1985 and 1987 due to frostnd subsequent fungal infections. Assuming that ∼10% of the totalrown defoliation resulted from frost damage, the “observed frostrown defoliation” could be calculated by taking a tenth of theotal observed crown defoliation which gives an estimated frost-
nduced crown defoliation of 1.1% (Fig. 3). This estimate was useds a benchmark for the simulated crown defoliation (which onlyccounts for frost damage). In general, the simulated crown defo-iation was very sensitive to changes in the parameters of the frostamage model. Particularly simulations F1 and F3 could reproduce
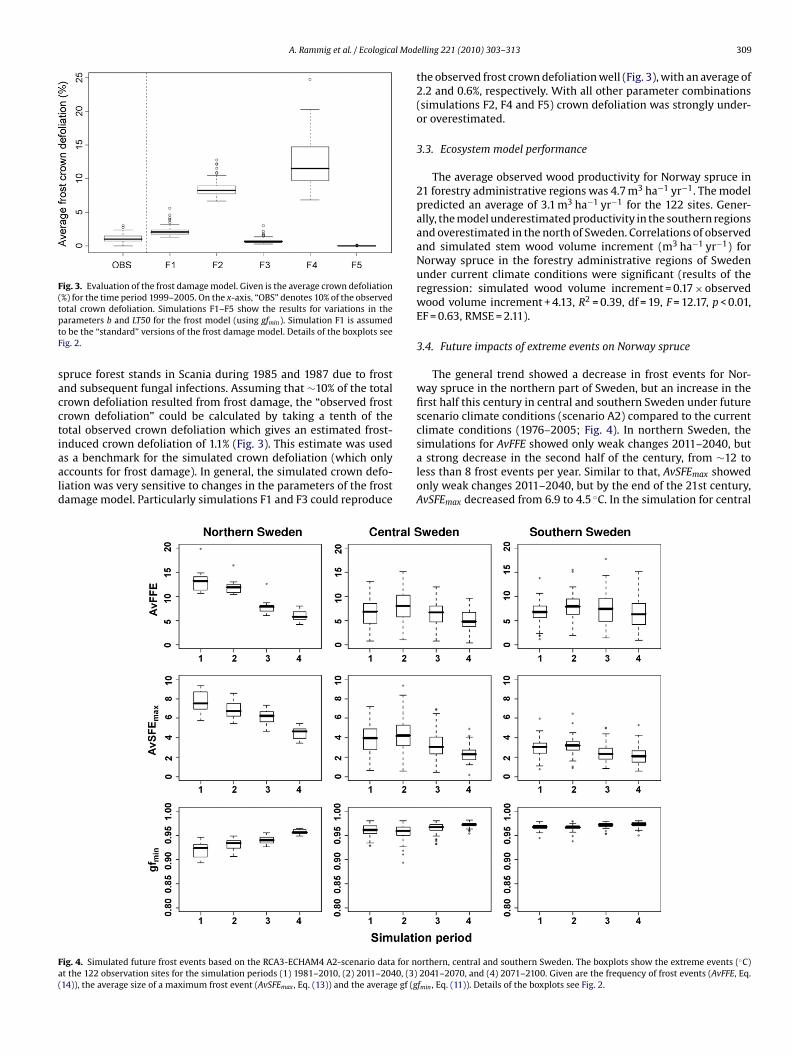
ig. 4. Simulated future frost events based on the RCA3-ECHAM4 A2-scenario data for nt the 122 observation sites for the simulation periods (1) 1981–2010, (2) 2011–2040, (3)14)), the average size of a maximum frost event (AvSFEmax , Eq. (13)) and the average gf (g
elling 221 (2010) 303–313 309
the observed frost crown defoliation well (Fig. 3), with an average of2.2 and 0.6%, respectively. With all other parameter combinations(simulations F2, F4 and F5) crown defoliation was strongly under-or overestimated.
3.3. Ecosystem model performance
The average observed wood productivity for Norway spruce in21 forestry administrative regions was 4.7 m3 ha−1 yr−1. The modelpredicted an average of 3.1 m3 ha−1 yr−1 for the 122 sites. Gener-ally, the model underestimated productivity in the southern regionsand overestimated in the north of Sweden. Correlations of observedand simulated stem wood volume increment (m3 ha−1 yr−1) forNorway spruce in the forestry administrative regions of Swedenunder current climate conditions were significant (results of theregression: simulated wood volume increment = 0.17 × observedwood volume increment + 4.13, R2 = 0.39, df = 19, F = 12.17, p < 0.01,EF = 0.63, RMSE = 2.11).
3.4. Future impacts of extreme events on Norway spruce
The general trend showed a decrease in frost events for Nor-way spruce in the northern part of Sweden, but an increase in thefirst half this century in central and southern Sweden under futurescenario climate conditions (scenario A2) compared to the currentclimate conditions (1976–2005; Fig. 4). In northern Sweden, the
simulations for AvFFE showed only weak changes 2011–2040, buta strong decrease in the second half of the century, from ∼12 toless than 8 frost events per year. Similar to that, AvSFEmax showedonly weak changes 2011–2040, but by the end of the 21st century,AvSFEmax decreased from 6.9 to 4.5 ◦C. In the simulation for central
orthern, central and southern Sweden. The boxplots show the extreme events (◦C)2041–2070, and (4) 2071–2100. Given are the frequency of frost events (AvFFE, Eq.fmin , Eq. (11)). Details of the boxplots see Fig. 2.
310 A. Rammig et al. / Ecological Modelling 221 (2010) 303–313
Table 4Evaluation of simulation results under future climate conditions (RCA3/ECHAM4/A2). The %-values in A) give the relative change of simulations where frost damage wasincluded in comparison to “No frost”-simulations.
(A) Average stem wood volume (m3 ha−1)a (B) Percentage of increase relative to 1976–2005 (%)
a Norway spruce stands were prescribed in the simulations to have the same stage was on average 69 years (Stdev: 12, min: 36, max: 95) and an average density o
nd southern Sweden, a different pattern was observed. In centralweden, AvFFE increased from 6 to 8 and AvSFEmax increased from.9 to 4.6 ◦C in the first half of the century. In southern Sweden, therequency of frost events (AvFFE) increased for a longer transienteriod until 2071. In the end of the 21st century, the frequency of
rost events (AvFFE) decreased to 5 in central and 6 frost events inouthern Sweden. The average maximum frost events (AvSFEmax)ecreased to 2.4 and 2.1 ◦C, respectively. The growth reducing fac-or (gfmin) varies according to AvSFEmax (Fig. 4). In northern Sweden,rost events have less impact on growth by the end of the century.n central Sweden, there is a transient period with increased frostamage during the first half of the century (Fig. 4).
.5. Forest productivity under future climate conditions
In general, all model versions projected an increase in Norwaypruce stand wood volume under the SRES-A2 future climate con-itions for all regions in Sweden (Table 4A). The model projectedn increase in mean annual stem wood volume in southern Swe-en from 219 to 265 m3 ha−1 for the period 1976–2100. In centralnd northern Sweden, stem wood volume was projected to increaserom 221 to 277 m3 ha−1, and from 137 to 170 m3 ha−1, respectively.
Simulated stem wood volume of Norway spruce was gener-lly lower if frost damage was included in the ecosystem modelTable 4A). In southern Sweden, when including frost damage, the
odel projected ∼5% less stem wood volume for 1976–2100 in com-arison to the FNO-simulations. For this period, stem volume wasecreased by ∼7% in central and 19% in northern Sweden.
The percentage of increase in stem wood volume under futurelimate conditions relative to 1976–2005 was positive for all modelersions (Table 4B). Under current conditions, frost damages areore important than under future climate conditions. A higher
umber of frost damages lowered the productivity in the F1-imulations under current conditions. Under future conditions,rost damage is occurring less frequently, leading to a higher pro-uctivity in the F1-simulations relative to 1976–2005 and relativeo the FNO-simulations. The increase of wood volume in the F1-imulations was projected to be 23% (south), 28% (central) and 46%north) for 2071–2100 relative to 1976–2005.
. Discussion
Seasonal variations in physiological processes in plants aretrongly driven by climate variables such as temperature and watervailability (Lambers et al., 2000). Climate change can thereforeffect such processes. The risk of future weather events exceeding
−12.0%
and density as the observed sites. At the end of each simulation period, the standrees ha−1 (Stdev: 269, min: 111, max: 1147).
the physiological tolerance of different species and thus causingdamage and reduced functioning can be explored by driving mod-els that encapsulate the relevant processes with data from climatescenarios. Ours is an example of such a study for the case of frostdamage in boreal forests. The hardiness model used (Jönsson et al.,2004) includes the gradual change in the environmental responseof frost hardiness. It considers the effect of both temperature andphotoperiod on the level of frost hardiness. In combination withthe frost damage model, frost damage in autumn, mid-winter andspring are considered.
We found a strong decrease of frost events in northern and onlylittle changes in central and southern Sweden under the futureclimate scenario investigated. Results from Jönsson et al. (2004)indicated latitudinal differences in the risk for frost, with a highnumber of spring frost events and almost no autumn frost events inthe South of Sweden for the period 1961–1990. In northern Sweden,their simulations showed opposite results: a low number of springfrost, but a high number of autumn frost events. Using data for theIPCC A2 and B2 future climate scenario for the period of 2070–2099from another combination of global and regional climate models,i.e. data from the Hadley Centre regional climate model HadRM3H,their model simulations showed a marked increase in the num-ber of spring frost events in the south and a decreased numberof autumn frost events in the north of Sweden. We did not dis-tinguish between spring and autumn frost events in the presentstudy; the results do not reflect an increase of spring frost eventsdue to a simultaneous decrease in autumn frost events (results notshown). Interestingly, our results suggest an increase in frost eventsin southern Sweden during the period 2011–2040, while simulatedfrost events strongly decreased in Sweden 2041–2100. This sug-gests that the increased risk for damage may not be permanentbut a temporary effect of increased risk for frost damage during atransient period caused by premature dehardening and therefore ahigher probability of temperature backlashes below freezing. Witha further increase in winter temperatures, the risk for frost dam-age could be reduced. The frequency and severity of harmful frostevents will change over time, but trajectories may vary depend-ing on climate scenarios. Data from another combination of globaland regional climate models, or from the same model forced by adifferent GHG emissions scenario, may result in a differently mod-elled frost damage. With a lower temperature increase, e.g. under
the SRES B1 and B2 scenarios (Nakicenovic et al., 2000), the risk forfrost damage could be elevated for a longer period of time. In thediscussion of options to reduce the risk of frost-related damages,such as by the adaptive selection of species and provenances, it isnecessary to consider the outcome of a range of contrasting climate
l Mod
cz(
fiepeffofrNiatess
waiH2d1eFaovstmafcBsit
(mwCiertacmelpoigmWpsr
A. Rammig et al. / Ecologica
hange scenarios in relation to species-specific ability for acclimati-ation, local adaptation and genetic adaptation by natural selectionHänninen et al., 2001; Jönsson et al., 2004).
To introduce the effect of frost events into the ecosystem model,rst, a “growth reducing factor” (gf) was calculated after Kellomäkit al. (1995) and Leinonen (1996). Their frost damage model wasarameterised for Scots pine (P. sylvestris), but resulted in appar-ntly realistic estimates for crown defoliation in our simulationsor Norway spruce. However, both the observed and simulatedrost crown defoliation are uncertain. We estimated that 6–12%f the total observed crown defoliation may be associated withrost events (Fig. 3). This is within the same range as valueseported from monitoring observations in southern Sweden (e.g.ihlgård, 1990), but crown defoliation of a stand is additionally
nfluenced by latitude, stand age, precipitation and nitrogen avail-bility (Rosengren-Brinck, 1998). However, the correlation betweenhe simulated frost damage and crown defoliation is unlikely to bexplained by covariance with these variables: frost events were notignificantly correlated with stand age and soil water (results nothown). Nitrogen dynamics were not explicitly addressed.
Sensitivity analysis showed that the simulated crown defoliationas very sensitive to changes in the parameters of the frost dam-
ge model. While much effort has been put into evaluating andmproving parameters for various models of frost hardiness (e.g.annerz and Westin, 2005; Hänninen, 2006; Hänninen and Kramer,007), parameters for estimating the actual frost damage on nee-les or whole forests have rarely been derived (but see Leinonen,996; Leinonen et al., 1997). Observations of damage to whole for-st stands have been made in the U.S.A. (Hagle et al., 2003) andinland (Jalkanen and Närhi, 1992; Jalkanen, 1996), where temper-ture inversions on mountain slopes damage large forest areas. Inur study, the simulated crown defoliation resembled the naturalariation and the amount of frost crown defoliation best when thetandard parameters were used for the calculation of growth reduc-ion (Table 3, simulation F1; Fig. 3). This suggests that simulated
aximum frost events (largest difference between hardiness levelnd daily minimum temperature) are best predictors for potentialrost damages. Such events are likely to be particularly sensitive tohanges in the seasonality and variability in daily temperatures (e.g.igras and Colombo, 2000; IPCC, 2007; Woldendorp et al., 2008),uggesting that impact modelling, such as projecting frost damagento the future, requires improved climate data at high spatial andemporal resolution.
Our study takes advantage of the high temporal resolution30 min; see Section 2) of data generated by the regional climate
odel RCA3. The input to the ecosystem model thus reflected dailyeather variations including the diurnal cycle in a realistic way.ompared with monthly mean data as used in many previous stud-
es of regional vegetation changes (e.g. Koca et al., 2006; Moralest al., 2007), this enables more realistic simulation of non-linearesponses of physiological processes to variable weather condi-ions, including extremes. However, in the context of many plantnd ecosystem processes, the spatial resolution of the RCM is ratheroarse. In reality, each simulated 50 km × 50 km gridcell containsany different vegetation stands and landscape units that influ-
nce local climate via variation in topography, hydrology, roughnessength and surface exchange. This heterogeneity is absent or highlyarameterised in the RCM which merely distinguishes the fractionsf ‘open land’ and ‘forest’ (Samuelsson et al., 2006). The variation
n local climate conditions this results in is aggregated to a sin-le value for each grid cell. The coarser the grid resolution, the
ore smoothed the simulated climate will be in terms of extremes.hile the spatial resolution of RCMs are steadily improved in
arallel with introduction of more sophisticated land-surfacechemes, the need for gridcell-to-point (site) downscaling willemain.
elling 221 (2010) 303–313 311
LPJ-GUESS has been shown to reproduce observed variations inecosystem productivity along broad climatic gradients (Morales etal., 2005; Zaehle et al., 2006) such as in Fennoscandia (Koca et al.,2006; Smith et al., 2008) and in response to elevated CO2 (Hickler etal., 2008). LPJ-GUESS is a generalised model which is not expected touniformly reproduce site-specific productivity because it does notinclude site-specific information such as detailed soil propertiesand scaling parameters for plant physiological processes as inputdata, as such data are not available for spatially extensive (grid-based) studies. One result of the generalised parameterisation wasthat the model underestimated productivity in the south of Swedenand overestimated it in the north. In addition, nutrient limitations,which are regarded as significant in the northern boreal forest, arenot included in the model. Bias in the simulated productivity gra-dient is, however, not critical for the effects of frost damage. Underfutures climate conditions, our simulation results suggest a strongincrease in the total productivity of Norway spruce stands (Table 4).However, including the physiological process of hardening and thesubsequent risk of frost damage by reducing the growth of the trees,the model projects lower productivity. Our simulation results sug-gest that frost events reduce stand productivity much more stronglyunder current climate conditions (i.e. 5% in the south and 25% inthe north of Sweden) than might be expected under an A2 futureclimate scenario (i.e. only 4% in the south and 12% in the north;Table 4). However, this reduction in productivity may be under-estimated, since the growth reducing effect of frost damage, wasonly calculated once per year in LPJ-GUESS, frost events were aggre-gated from among seasons into one value. In reality, there are oftenlarge differences in damage potential between spring and autumnfrost events. During spring, new buds and shoots are very sensi-tive to frost, whereas older shoots are more tolerant (Cannell andSheppard, 1982; Bigras and Colombo, 2000).
Our results point to the importance of including representationsof hardening/dehardening cycles and frost damage in ecosystemmodels to correctly capture transient changes in productivity andcarbon cycling of high-latitude forest ecosystems under globalchange.
Acknowledgements
The Project “Climate extremes & Ecosystem modelling” wasfunded by the Swedish Research Council. Part of this work was donewithin the EU/FP6 ENSEMBLES project (GOCE-CT-2003-505539).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ecolmodel.2009.05.014.
References
Alden, J., Hermann, R.K., 1971. Aspects of the cold-hardiness mechanism in plants.Botanical Review 37, 37–142.
Badeck, F.-W., Lischke, H., Bugmann, H., et al., 2001. Tree species composition in Euro-pean pristine forests. Comparison of stand data to model predictions. ClimaticChange 51, 307–347.
Bartelink, H.H., 2000. Effects of stand composition and thinning in mixed-speciesforests: a modeling approach applied to Douglas-fir and beech. Tree Physiology20, 399–406.
Bergh, J., Freeman, M., Sigurdsson, B., et al., 2003. Modelling the short-term effects ofclimate change on the productivity of selected tree species in Nordic countries.Forest Ecology and Management 183, 327–340.
ugmann, H., 1994. On the ecology of mountainous forests in a changing climate:a simulation study. Ph.D. Thesis. Swiss Federal Institute of Technology Zurich,Zurich, 258 pp.
annell, M.G.R., Sheppard, L.J., 1982. Seasonal changes in the frost hardiness of prove-nances of Picea sitchensis in Scotland. Forestry 55, 137–153.
annell, M.G.R., Smith, R.I., 1986. Climatic warming, spring budburst and forest dam-age on trees. Journal of Applied Ecology 23 (1), 177–191.
hen, H.-H., Li, P.H., 1978. Interactions of low temperature, water stress and shortdays in the induction of stem frost hardiness in red osier dogwood. Plant Physi-ology 62, 833–835.
hristensen, J.H., Hewitson, B., Busuioc, A., et al., 2007. Regional climate projections.In: Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M.,Miller, H.L. (Eds.), Climate Change 2007: The Physical Science Basis. Contributionof Working Group I to the Fourth Assessment Report of the IntergovernmentalPanel on Climate Change. Cambridge University Press, pp. 847–940.
ichhorn, J., Szepesi, A., Ferretti, M., Durrant, D., Roskams, P., 2006. Manual on meth-ods and criteria for harmonized sampling, assessment, monitoring and analysisof the effects of air pollution on forests, Part II: visual assessment of crowncondition. In: ICP Forests Manual. United Nations Economic Commission forEurope, Convention on Long-Range Transboundary Air Pollution, InternationalCo-operative Programme on Assessment and Monitoring of Air Pollution Effectson Forests, Hamburg, Germany.
uchigami, L.H., Weiser, C.J., Kobayashi, K., Timmis, R., Gusta, L.V., 1982. A degreegrowth stage (◦GS) model and cold acclimation in temperate woody plants. In:Li, P.H., Sakai, A. (Eds.), Plant Cold Hardiness and Freezing Stress. Mechanismsand Crop Implications. Academic Press, New York, pp. 93–116.
agle, S.K., Gibson, K.E., Tunnock, S., 2003. A Field Guide to Diseases and Insect Pestsof Northern and Central Rocky Mountain Conifers. United States Department ofAgriculture, Forest Service, Missoula, Montana.
annerz, M., 1994. Predicting the risk of frost occurrence after bud burst of Norwayspruce in Sweden. Silva Fennica 28, 243–249.
annerz, M., Westin, J., 2005. Autumn frost hardiness in Norway spruce plus treeprogeny and trees of the local and transferred provenances in central Sweden.Tree Physiology 25, 1181–1186.
änninen, H., 2006. Climate warming and the risk of frost damage to boreal for-est trees: identification of critical ecophysiological traits. Tree Physiology 26,889–898.
änninen, H., Beuker, E., Johnsen, O., Leinonen, I., Murray, M., Sheppard, L., Skroppa,T., 2001. Impacts of climate change on cold hardiness of conifers. In: Bigras,F.J., Colombo, S.J. (Eds.), Conifer Cold Hardiness. Kluwer Academic, Boston, pp.305–333.
änninen, H., Kramer, K., 2007. A framework for modelling the annual cycle of treesin boreal and temperate regions. Silva Fennica 41 (1), 167–205.
arrison, L.C., Weiser, C.J., Burke, M.J., 1978. Environmental and seasonal factorsaffecting the frost-induced stage of cold acclimation in Cornus stolonifera Michx.Plant Physiology 62, 894–898.
ickler, T., Smith, B., Sykes, M.T., Davis, M.B., Sugita, S., Walker, K., 2004. Using ageneralized vegetation model to simulate vegetation dynamics in northeasternUSA. Ecology 85 (2), 519–530.
ickler, T., Smith, B., Prentice, I.C., Mjöfors, K., Miller, P., Arneth, A., Sykes, M., 2008.CO2 fertilization in temperate FACE experiments not representative of borealand tropical forests. Global Change Biology 14, 1531–1542.
PCC, 2007. Climate Change 2007: The Physical Science Basis. Contribution of Work-ing Group I to the Fourth Assessment Report of the Intergovernmental Panel onClimate Change. Cambridge University Press, Cambridge, United Kingdom andNew York, NY, USA, 996 pp.
aakkola, T., Mäkinen, H., Saranpää, P., 2006. Wood density of Norway spruce:responses to timing and intensity of first commercial thinning and fertilisation.Forest Ecology & Management 237, 513–521.
acob, D., Barring, L., Christensen, O.B., Christensen, J.H., de Castro, M., Deque, M.,Giorgi, F., Hagemann, S., Lenderink, G., Rockel, B., Sanchez, E., Schar, C., Senevi-ratne, S.I., Somot, S., van Ulden, A., van den Hurk, B., 2007. An inter-comparison ofregional climate models for Europe: model performance in present-day climate.Climatic Change 81, 31–52.
alkanen, R., 1996. Repeated red belts at Lusmavaara, western Lapland. Pallas-Symposium 1996. Finnish Forest Research Institute, Research Papers 623, 91–95.
alkanen, R., Närhi P., 1992. Red Belt Phenomenon on the Slopes of the Levi fell inKittilä, Western Lapland. The Finnish Forest Research Institute, Research Papers,vol. 451, pp. 55–60.
ansson, A., Persson, C., Strandberg, G., 2007. 2D meso-scale re-analysis of precipita-tion, temperature and wind over Europe–ERAMESAN, Time period 1980–2004.SMHI Reports Meteorology and Climatology, No. 112, 28 pp.
arvis, P., Saugier, B., Schulze, E.-D., 2001. Productivity of boreal forests. In: Roy, J.,Saugier, B., Mooney, H.A. (Eds.), Terrestrial Global Productivity. Academic Press,San Diego, London, pp. 211–244.
önsson, A.M., Linderson, M.-L., Stjernquist, I., Schlyter, P., Bärring, L., 2004. Climatechange and the effect of temperature backlashes causing frost damage in Piceaabies. Global and Planetary Change 44, 195–207.
önsson, A.M., 2005. Tracing the impact of adverse environmental conditions on thefrost sensitivity in trees. Journal of Sustainable Forestry 21, 65–75.
oos, F., Prentice, I.C., Sitch, S., Meyer, R., Hooss, G., Plattner, G.K., Gerber, S., Hassel-mann, K., 2001. Global warming feedbacks on terrestrial carbon uptake under theIntergovernmental Panel on Climate Change (IPCC) emission scenarios. GlobalBiogeochemical Cycles 15, 891–907.
elling 221 (2010) 303–313
Karlman, M., 1986. Damage to Pinus contorta in northern Sweden with specialemphasis on pathogens. Stud. For. Sue, No. 176. Faculty of Forestry, SwedishUniv. of Agricultural Sciences, Uppsala, 42 pp.
Kellomäki, S., Hänninen, H., Kolström, M., 1995. Computations on frost damage toScots pine under climatic warming in boreal conditions. Ecological Applications5 (1), 42–52.
Kjellström, E., 2004. Recent and future signatures of climate change in Europe. Ambio33, 193–198.
Kjellström, E., Bärring, L., Gollvik, S., Hansson, U., Jones, C., Samuelsson, P., Rum-mukainen, M., Ullerstig, A., Willén, U., Wyser, K., 2005. A 140-year simulationof European climate with the new version of the Rossby Centre regional atmo-spheric climate model (RCA3). SMHI Reports Meteorology and Climatology 108,1–54.
Kjellström, E., Bärring, L., Jacob, D., Jones, R., Lenderink, G., Schär, C., 2007. Modellingdaily temperature extremes: recent climate and future changes over Europe.Climatic Change 81 (Suppl. 1), 249–265.
Koca, D., Smith, B., Sykes, M., 2006. Modelling regional climate change effects onpotential natural ecosystems in Sweden. Climatic Change 78, 381–406.
Köstler, J.N., Brückner, E., Biebelriether, H., 1968. Die Wurzeln der Waldbäume. PaulParey, Hamburg, 284 pp.
Köstner, B., Falge, E., Tenhunen, J.D., 2002. Age-related effects on leaf area/sapwoodarea relationships, canopy transpiration and carbon gain of Norway sprucestands (Picea abies) in the Fichtelgebirge, Germany. Tree Physiology 22, 567–574.
Kowalski, T., 1991. Oak decline: I. Fungi associated with various disease symptomson overground portions of middle-aged and old oak (Quercus robur L.). EuropeanJournal of Forest Pathology, 21.
Kowalski, T., 1996. Oak decline: II. Fungi associated with various types of lesions onstems and branches of young oaks (Quercus robur L.). Österr. Z. Pilzk. 5, 51–63.
Lambers, H., Chapin, F.S., Pons, T.L., 2000. Plant Physiological Ecology. Springer Ver-lag, New York, Berlin, Heidelberg, 540 pp.
Leinonen, I., 1996. A simulation model for the annual frost hardiness and freezedamage of Scots pine. Annals of Botany 78, 687–693.
Leinonen, I., Repo, T., Hänninen, H., 1997. Changing environmental effects onfrost hardiness of Scots pine during dehardening. Annals of Botany 79, 133–138.
Li, Z., Kurz, W.A., Apps, M.J., Beukema, S.J., 2003. Belowground biomass dynamics inthe Carbon Budget Model of the Canadian Forest Sector: recent improvementsand implications for the estimation of NPP and NEP. Canadian Journal of ForestResearch 33, 106–126.
Linder, S., Flower-Ellis, J.G.K., 1992. Environmental and physiological constraints toforest yield. In: Teller, A., Mathy, P., Jeffers, J.N.R. (Eds.), Responses of ForestEcosystems to Environmental Changes. Elsevier Applied Science, London, pp.149–164.
process-based ecosystem model predictions of carbon and water fluxes in majorEuropean forest biomes. Global Change Biology 11, 2211–2233.
Morales, P., Hickler, T., Rowell, D., Smith, B., Sykes, M., 2007. Changes in Europeanecosystem productivity and carbon balance driven by regional climate modeloutput. Global Change Biology 13, 108–122.
Nakicenovic, N., Alcamo, J., Davis, G., et al., 2000. IPCC Special Report on EmissionScenarios. Cambridge University Press, Cambridge, 612 pp.
Nihlgård, B., 1990. Dynamik i barrkemi och barrstrukturer på Skånska gran-och tall-provytor åren 1985–1987. Skånes samrådesgrupp mot skogsskador Rapport, Nr.8, 91 pp.
Niinemets, Ü., Lukjanova, A., 2003. Total foliar area and average leaf age may bemore strongly associated with branching frequency than with leaf longevity intemperate conifers. New Phytologist 158, 75–89.
Ögren, E., 2001. Effects of climatic warming on cold hardiness of some northernwoody plants assessed from simulation experiments. Physiologia Plantarum 112,71–77.
Prentice, I.C., Sykes, M.T., Cramer, W., 1993. A simulation model for the transienteffects of climate change on forest landscapes. Ecological Modelling 65, 51–70.
Repo, T., Hänninen, H., Kellomäki, S., 1996. The effects of long-term elevation ofair temperature and CO2 on the frost hardiness of Scots pine. Plant, Cell andEnvironment 19, 209–216.
Roeckner, E., Bengtsson, L., Feichter, J., Lelieveld, J., Rodhe, H., 1999. Transient cli-mate change simulations with a coupled atmosphere-ocean GCM including thetropospheric sulfur cycle. Journal of Climate 12, 3004–3032.
Rosengren-Brinck, U., 1998. Barrförlust och luftföroreningar: Samband mellan kro-nutglesning och miljöfaktorer I barrskog. Naturvårdsverket Rapport, 4890 pp.
Sakai, A., Larcher, W., 1987. Frost Survival of Plants. Springer Verlag, Berlin, Germany,321 pp.
Saltelli, A., Chan, K., Scott, E.M., 2000. Sensitivity Analysis. John Wiley & Sons, Ltd.,Chichester, New York, Weinheim, 475 pp.
Samuelsson, P., Gollvik, S., Ullerstig, A., 2006. The land-surface scheme of the RossbyCentre regional atmospheric climate model (RCA3). SMHI Rapport Meteorologi,Nr. 122, 25 pp.
Schoeneweiss, D.F., 1975. Predisposition, stress and plant disease. In: Baker-Kenneth,F. (Ed.), Annual Review of Phytopathology. Illus Annual Reviews, Palo Alto, CA,
USA, pp. 193–211.
Sieber, T.N., Kowalski, T., Holdenrieder, O., 1995. Fungal assemblages in stem andtwig lesions of Quercus robur in Switzerland. Mycological Research 99, 534–538.
Sitch, S., Smith, B., Prentice, I.C., Arneth, A., Bondeau, A., Cramer, W., Kaplans, J.O.,Levis, S., Lucht, W., Sykes, M.T., Thonicke, K., Venevsky, S., 2003. Evaluation of
l Mod
S
S
S
S
U
and ecosystem functioning in European forest biomes. Ecological Modelling 216,
A. Rammig et al. / Ecologica
ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJdynamic global vegetation model. Global Change Biology 9, 161–185.
mith, B., Knorr, W., Widlowski, J.L., Pinty, B., Gobron, N., 2008. Combining remotesensing data with process modelling to monitor boreal conifer forest carbonbalances. Forest Ecology & Management 255, 3985–3994.
mith, B., Prentice, I.C., Sykes, M., 2001. Representation of vegetation dynamics inthe modelling of terrestrial ecosystems: comparing two contrasting approacheswithin European climate space. Global Ecology and Biogeography 10, 621–637.
utinen, M.-L., Arora, R., Wisniewski, M., Ashworth, E., Strimbeck, R., Palta, J.P.,2001. Mechanisms of frost survival and freeze-damage in nature. In: Bigras,
wedish Forest Agency, 2007. Statistical Yearbook of Forestry 2007. Skogsstyrelsen,Jönköping, Sweden, 345 pp.
ppala, S.M., Kållberg, P.W., Simmons, A.J., et al., 2005. The ERA-40 re-analysis. Quar-terly Journal of the Royal Meteorological Society 131, 2961–3012.
Woldendorp, G., Hill, M.J., Doran, R., Ball, M.C., 2008. Frost in a future climate: mod-elling interactive effects of warmer temperatures and rising atmospheric [CO2]on the incidence and severity of frost damage in a temperate evergreen (Euca-lyptus pauciflora). Global Change Biology 14, 294–308.
Wramneby, A., Smith, B., Zaehle, S., Sykes, M.T., 2008. Parameter uncertainties inthe modelling of vegetation dynamics—effects on tree community structure
277–290.Zaehle, S., Sitch, S., Prentice, I.C., Liski, J., Cramer, W., Erhard, M., Hickler, T.,
Smith, B., 2006. The importance of representing age-related decline in forestNPP for modeling regional carbon balances. Ecological Applications 16, 1555–1574.