232 BIOCHIMICA ET BIOPHYSICA ACTA BBA 25345 PROLINE INCORPORATION INTO THE PROLINE AND HYDROXY- PROLINE OF EARTHWORM-CUTICLE COLLAGEN DAISABURO FUJIMOTO* AND ELIJAH ADAMS Department of Biochemistry, University of Maryland School of Medicine, Baltimore, Md. (U.S.A .) (Received January i2th, 1965) SUMMARY Studies in vivo of the incorporation of proline into earthworm cuticle have shown that, as with vertebrate collagens, proline and not free hydroxyproline is the source of both proline and hydroxyproline in cuticle collagen. Purified salt-soluble cuticle collagen preparations contain a component highly labeled in proline and re- sistant to collagenase (EC 3.4.4.i9), but sensitive to trypsin (EC 3.4.4.4). The labeled proline in crude collagen preparations, although much higher in specific activity than hydroxyproline, decays with about the same time-constant as the hydroxyproline; this and related observations suggest that injection-stimulated synthesis rather than normal turnover is being measured in these experiments. Digestion of cuticle collagen with collagenase releases most of the hydroxyproline in dialyzable form, implying extensive degradation to small fragments. INTRODUCTION The major fibrous protein of earthworm cuticle is classed as a collagen on the basis of wide-angle X-ray diffraction patterns 1, high levels of hydroxyproline2-5, characteristic optical-rotatory properties4, ~, and susceptibility to collagenase (EC 3.4.4.I9) B. It differs from most other collagens known, both vertebrate and inverte- brate, in lack of periodic striations on electron microscopy7, larger size of the tropo- collagen molecule4,5, composition of amino acids z-5 and larger quantity of associated carbohydrate 3-5. The most striking features of its amino acid composition are the high hydroxy- proline content (153-165 residues per thousand) and low proline content (7.7-13 resi- dues per thousand) 2-5. This imino acid distribution lends it unusual interest for certain aspects of collagen research. In particular, the stability of the cuticle collagenS, 6 and the marked difference between the composition of collagen from earthworm body and cuticle have been studied s. It is well-established that hydroxyproline of vertebrate collagen is derived from proline and not from free hydroxyproline (for a recent review see ROBERTSONg).Little information is yet available on the biosynthesis of invertebrate collagens, and it is of particular interest to examine this question in the case of earthworm-cuticle collagen because of its unusually high hydroxyproline content. * Permanent address: Tokyo Medical and Dental University, Tokyo, Japan. Biochim. Biophys. Acta, lO 7 (1965) 232-246
Transcript
232 BIOCHIMICA ET BIOPHYSICA ACTA
BBA 25345
PROLINE INCORPORATION INTO T H E PROLINE AND HYDROXY-
PROLINE OF EARTHWORM-CUTICLE COLLAGEN
DAISABURO FUJIMOTO* AND ELIJAH ADAMS
Department of Biochemistry, University of Maryland School of Medicine, Baltimore, Md. (U.S.A .)
(Received January i2th, 1965)
SUMMARY
Studies in vivo of the incorporation of proline into earthworm cuticle have shown that, as with vertebrate collagens, proline and not free hydroxyproline is the source of both proline and hydroxyproline in cuticle collagen. Purified salt-soluble cuticle collagen preparations contain a component highly labeled in proline and re- sistant to collagenase (EC 3.4.4.i9), but sensitive to trypsin (EC 3.4.4.4). The labeled proline in crude collagen preparations, although much higher in specific activity than hydroxyproline, decays with about the same time-constant as the hydroxyproline; this and related observations suggest that injection-stimulated synthesis rather than normal turnover is being measured in these experiments. Digestion of cuticle collagen with collagenase releases most of the hydroxyproline in dialyzable form, implying extensive degradation to small fragments.
INTRODUCTION
The major fibrous protein of earthworm cuticle is classed as a collagen on the basis of wide-angle X-ray diffraction patterns 1, high levels of hydroxyproline 2-5, characteristic optical-rotatory properties4, ~, and susceptibility to collagenase (EC 3.4.4.I9) B. It differs from most other collagens known, both vertebrate and inverte- brate, in lack of periodic striations on electron microscopy 7, larger size of the tropo- collagen molecule4, 5, composition of amino acids z-5 and larger quantity of associated carbohydrate 3-5.
The most striking features of its amino acid composition are the high hydroxy- proline content (153-165 residues per thousand) and low proline content (7.7-13 resi- dues per thousand) 2-5. This imino acid distribution lends it unusual interest for certain aspects of collagen research. In particular, the stability of the cuticle collagenS, 6 and the marked difference between the composition of collagen from earthworm body and cuticle have been studied s.
I t is well-established that hydroxyproline of vertebrate collagen is derived from proline and not from free hydroxyproline (for a recent review see ROBERTSONg). Little information is yet available on the biosynthesis of invertebrate collagens, and it is of particular interest to examine this question in the case of earthworm-cuticle collagen because of its unusually high hydroxyproline content.
* Permanent address: Tokyo Medical and Dental University, Tokyo, Japan.
Biochim. Biophys. Acta, lO 7 (1965) 232-246
INCORPORATION STUDIES WITH EARTHWORM-CUTICLE COLLAGEN 233
This paper reports observations of the incorporation in vivo of proline and hydroxyproline into earthworm-cuticle collagen. The generalizations that proline is the source of collagen hydroxyproline and that free hydroxyproline is not incorporated into collagen, based on many studies of vertebrate preparations, also hold for earth- worm cuticle. Certain differences from the results of vertebrate studies have appeared, however, notably evidence for a fraction extractable with collagen that is not sus- ceptible to collagenase, is highly labeled in proline, and that resembles cuticle collagen both in solubility properties and in certain metabolic features.
MATERIALS AND METHODS
Animals Earthworms, probably a species of Lumbricus, were purchased from Wholesale
Bait Co., Hamilton, Ohio. For long-term storage they were kept at 4 ° in a commercial soil preparation (Buss Manufacturing Co., Lanark, Ill.).
Chemicals and enzymes L-[14C~Proline (uniformly labeled) was purchased from New England Nuclear
Corp. Specific activity was adjusted to 33 t~C//~mole by diluting with non-labeled L-proline. DL-E2-14ClHydroxyproline (4-9/~C//~mole) was purchased from Nuclear Chicago Corp. L-[5-3H~Hydroxyproline (5o0/~C//~mole), obtained from New England Nuclear Corporation, was prepared by the reduction with 3H, gas of enzymatically made Al-pyrroline-3-hydroxy-5-carboxylate TM. Collagenase (EC 3.4.4.19) (Worthing- ton Catalog No. CLSP 63o3-I), crystalline trypsin (EC 3.4-4-4) and elastase (EC 3.4.4.7) were the products of Worthington Biochemical Corporation. Both racemates of syn- thetic 3-hydroxyprolineX!, as well as natural 3-hydroxyproline isolated from beef Achilles tendon 11, were gifts from Dr. M. A. LOGAN.
Administration of labeled compounds Worms were routinely injected using a 1.5-in, 27-gauge needle, passed through
the body wall in the anterior third and inserted posteriorly within the coelom. I-IO/~C of labeled amino acids in 0.2 ml of water were injected into each worm, usually during slow withdrawal of the needle to permit distribution of the injectate into a number of body segments. Injected worms were kept in moist soil-bedding at various tempera- tures.
Preparation of collagen At varying intervals after E14Clproline administration, worms were killed by
I5-min soaking in ether at room temperature 3. The cuticles were stripped from the body wall with a fine forceps and were routinely washed by rinsing repeatedly in distilled water at 4 °, usually about 6 washes of 500 ml each over a period of 6-24 h. Collagen was extracted from the washed cuticles either by trichloroacetic acid at 9 °o or 0.5 M NaC1. Extraction was carried out using IO % trichloroacetic acid, 0.5 ml per worm, at 9 °0 for 60 min (ref. 12). After centrifugation to remove undissolved material, the supernatant solution was dialyzed against several changes of water. The extract obtained contained 5-8/~moles of hydroxyproline per worm. NaCl-soluble collagen was prepared according to JOSSE AND HARRINGTON 5 by repeated extraction
Biochim. Biophys. Acta, Io7 (1965) 232-246
234 D. FUJIMOTO, E. ADAMS
with 0.5 M NaC1 and precipitation with 20 % saturated alkaline (NH4)2SO 4. Prepa- rations containing 0.2-0. 4/,mole of hydroxyproline per worm were obtained.
Preparation of labeled, non-collagen proteins Worms were injected with I/~C each of L-[14Clproline and kept for 24 h at I2 °.
Cuticles were removed and washed with two changes of IO ml of water. The washings were combined and centrifuged at 40000 rev./min for 60 rain. The supernatant solution from 5 worms, containing 5.7" lO4 counts/rain, was used for coprecipitation experiments with NaCl-soluble collagen. To another portion of the supernatant so- lution was added an equal volume of cold IO % trichloroacetic acid. The precipitate was collected and washed once with 5 % trichloroacetic acid, then dissolved by adding a small amount of 2 N NH4OH and warming at 5 o°. After trichloroacetic acid precipi- tation was repeated once more, the precipitate was taken up in o.i N NH,OH and dialyzed against several changes of water (I 1 each) for 48 h. The preparation from 15 worms contained about lO 5 counts/min and the specific activity of proline was measured as 4900 counts/min per/~mole. This preparation was used as "non-collagen" protein for testing collagenase specificity.
To obtain mucus, IO worms were immersed in IO ml of water and mucus secretion was stimulated by adding a piece of solid CO s. After IO min, the liquid was collected and centrifuged at IOOOO × g for 15 rain; the supernatant solution was dialyzed against cold water. This preparation, which contained about 3.5.1o 4 counts/rain, was used as "mucus" in coprecipitation experiments.
Determination of specific activity of proline and hydroxyproline Cuticles or extracted collagen preparations were hydrolyzed in tightly stoppered
bakelite screw-top tubes with 5 ml of 6 N HC1 at IiO ° for 20-24 h. After hydrolysis, HC1 was removed by evaporation in a boiling-water bath under an air stream, and the residue was taken up in I ml of water. NaNO~ (ioo mg in 0.5 ml of water) was added and the mixture was kept at room temperature for I h, after which 1.5 ml of concentrated HC1 were added and the mixture heated at ioo ° for i h. After removal of HC1 by evaporation under an air stream, the residue was taken up in 5 ml of water and applied to a Dowex-5o (H +) column (I cm x 7 cm). The column was washed with 5 volumes of water, after which amino acids were eluted with I N NH~OH. The eluate was concentrated and chromatographed by an ascending method on thick paper (Whatman No. 3MM or No. I7), with phenol-water (80:20, v/v) or tert.- butanol-formic acid-water (7o:15:15, v/v) as solvents. Regions corresponding to proline and hydroxyproline were cut out and eluted with water. Aliquots of paper eluates were counted in an end-window gas-flow counter giving an efficiency of 2o-3o %. Proline was determined by either Method A or B of PIEZ et al.13; hydroxy- proline was determined either by Method A of PIEZ et al. 13 or by a modification of the method of NEUMAN AND LOGAN 14.
Density-gradient centrifugation Sucrose gradient centrifugation was carried out as described by MARTIN .AND
AMES 15 in linear gradients from 5 % to 20 % in sucrose containing 0.5 M NaC1. Tubes were layered with 0. 3 ml of a collagen solution and centrifuged at 30000 rev./min for 18 h, at or below IO ° (chamber temperature). After centrifugation the tube was
Biochim. Biophys. Mcta, lO 7 (1965) 232-246
I N C O R P O R A T I O N S T U D I E S W I T H E A R T H W O R M - C U T I C L E C O L L A G E N 235
punctured at the bot tom and samples of 12 drops each were collected, dialyzed against water to remove sucrose, and counted for radioactivity.
Digestion by proteolytic enzymes NaCl-soluble collagen preparations were incubated with collagenase, trypsin or
elastase in buffers containing 0.05 M Tris (pH 7.5), and 0.5 mM CaC12. In the case of collagenase, the enzyme was usually about one-fifth the weight of the substrate and incubation was carried out at 37 ° for 18 h. In the case of the other proteases, amounts of enzyme about equal to substrate were used. In the latter cases, incubation was carried out at 12 ° to avoid denaturation of collagen and consequent susceptibility of collagen to general protease action. In each case one drop of toluene was added to prevent significant bacterial contamination. After incubation, digested fragments were separated from residual protein by dialysis with stirring against 250 ml of cold water for 24 h, or by addition of 2 volumes of cold ethanol followed by centrifugation.
Absence of non-specific protease activity in the collagenase preparation was confirmed by the release of only negligible counts (20 counts/rain) in dialyzable form from 14C-labeled "non-collagen" protein containing 20 ooo counts/min (4900 counts per min per /zmole of proline), following incubation with 0.2 mg of collagenase at 37 ° for 18 h. In contrast, 66 % of the counts of a collagen preparation were rendered ethanol-soluble after incubation with the same preparation of collagenase under similar conditions (Table I I I ) .
Isolation of hydroxyproline from cuticle-stripped worm bodies The method of extraction of worm bodies has been described separately s.
Hydroxyproline was isolated by the same procedure as described here for the cuticle. 3-Hydroxyproline (identified as trans-3-hydroxyproline, as described below) and 4-hydroxyproline were separated by paper electrophoresis at 30 V/cm for 2 h in a buffer of pH 2 containing 67 ml of acetic acid and 15.5 ml of formic acid per 1 (ref. 16).
R E S U L T S
Source of hydroxyproline in cuticles As shown in Table I, about o.I % of administered radioactivity was found in
the hydroxyproline of cuticle when L-[14C]proline was the source; labeled free
T A B L E I
INCORPORATION o F LABELED PROLINE OR HYDROXYPROLINE INTO CUTICLE HYDROXYPROLINE
I n j e c t e d w o r m s w e r e k e p t a t 25 ° f o r 24 h a f t e r w h i c h w h o l e w a s h e d c u t i c l e s w e r e h y d r o l y z e d a n d h y d r o x y p r o l i n e i s o l a t e d as d e s c r i b e d in t h e t e x t .
Injected compound #C given per worm Specific activity of cuticle hydroxy- proline (disinte- grations/rain per izmole)
Fraction of administered radio- activity in cuticle hydroxyproline (%)
L- [14C]Pro l ine I L- [14C]Pro l ine 7.5 L- [ 5 - a H ] H y d r o x y p r o l i n e IO DL- [2-14C] H y d r o x y p r o l i n e 1. 5
185 0 .07 3 5 2 0 o .16
2 6 o . o o o 6 o o
Biochim. Biophys. Acta, lO 7 (1965) 2 3 2 - 2 4 6
236 D. FUJIMOTO, E. ADAMS
hydroxyproline was not incorporated in significant amounts into the hydroxyproline of cuticles. These results, suggesting that free proline and not free hydroxyproline serves as the source of hydroxyproline in cuticle, are consistent with many similar observations with vertebrate collagen 9. It is of interest to compare the fraction of administered proline incorporated into the hydroxyproline of cuticle with data in vivo for other species. For example, MITOMA et al. 17 have reported incorporation data into the hydroxyproline of whole I2-day chick-embryo 24 h after injection of L-[14C~proline into the egg. Their data and ours (Table I), may be compared with respect to the resulting specific activity of hydroxyproline per unit of administered radioactivity per g of body weight. On this basis, chick-embryo collagen appears to be a more rapid incorporating system, by at least a factor of I0, than earthworm cuticle. On the basis of a similar calculation using data of ROBERTSON et al. ~s, incorporation of [14Clproline into the hydroxyproline of carageenin-granulomas in guinea pigs also appears to be considerably more efficient than into earthworm cuticle. The efficiency of incorporation into worm-body collagen hydroxyproline was only slightly greater than into cuticle (Table vii).
Relative specific activities of proline and hydroxyproline As noted in Table II, cuticle collagen extracted by hot trichloroacetic acid
showed a surprisingly high ratio of proline/hydroxyproline specific activity, varying between 29 and IOO. In the NaCl-soluble preparations, purified by the method of JOSSE AND HARRINGTON 5, there was also a marked deviation of this ratio from I, with values ranging between about 5 and 12. These data should be contrasted with proline/hydroxyproline specific-activity ratios measured in collagen extracted by comparable methods from vertebrate tissues. In the latter cases, ratios closer to I, varying only between about 0.6 and 1.6 have been reportedlS-~; these relatively small deviations from I have engendered discussion both as to their reality 2° and their interpretation18,19, ~1.
It is particularly notable that in contrast to the striking difference between
T A B L E I I
S P E C I F I C A C T I V I T Y OF P R O L I N E A N D H Y D R O X Y P R O L I N E I N C U T I C L E C O L L A G E N P R E P A R A T I O N S
W o r m s were k e p t for 24 h a f te r a d m i n i s t r a t i o n of L-[laC]proline. S torage t e m p e r a t u r e s were 25 ° (Expts . I, 5), 4 ° (Expt . 2), 12 ° (Expts . 3, 4, 6-8). Collagen p r epa ra t i ons used were ho t t r ichloro- acet ic acid-soluble (Expts . 1-4) and NaCl-soluble (Expts . 5-8).
Expt. I*C given Specific activity (counts/min per pmole) Molar ratio No. per worm hydroxyproline
i I 374 ° 36 lo4 23 2 I 66oo 227 29 No t measured 3 i 69oo 74 93 No t measu red 4 i 385o 5o 77 No t measured 5 i 254 o 356 7 .1 24 6 2. 5 65o0 56o 12 22 7 4 740o 9o6 8.2 2o 8 5 24000 512o 4.7 20
Biochim. Biophys. Acta, lo7 (1965) 232-246
INCORPORATION STUDIES WITH EARTHWORM-CUTICLE COLLAGEN 237
trichloroacetic acid-extracted collagen and NaCl-soluble collagen with respect to the specific-activity ratio, the molar ratios of these two amino acids agreed closely in both fractions and were similar to values reported by JOSSE AND HARRINGTON 5 for the NaCl-soluble fraction, which was carefully characterized by them and estimated to be rather pure by physical measurements.
These considerations suggested that an impurity accounted for the high specific activity of proline in both the trichloroacetic acid-soluble and NaCl-soluble collagen fractions of earthworm cuticle (since the specific-activity ratio was considerably reduced in the purer fraction), and that it must comprise only a small quantity of highly labeled proline (since the molar ratios were not significantly affected on purifi- cation, while the specific-activity ratio dropped sharply). From the extraction behavior of this "contaminant", it seemed possible that it might represent a proline-rich collagen precursor before hydroxylation. I t was therefore of interest to examine its susceptibility to collagenase and other proteinases.
T A B L E I I I
PROT]~INASE ACTION IN LIBERATING RADIOACTIVITY AND HYDROXYPROLINE FROM CUTICLE COLLAGEN
A single prepara t ion of NaCl-soluble cuticle collagen was isolated after adminis t ra t ion in vivo of I,-[I4C]proline. Aliquots containing o.9 #mole of hydroxyprol ine and 2ooo counts/rain were incu- bated for 18 h with 0. 5 mg of collagenase or i .o mg of t ryps in or i.o mg of elastase, in 1.2 ml of o.oi M Tris (pH 7-4), and o.I mM CaC1 v Collagenase incubat ion was at 37 °, the others at 12 °. After incubation, 2. 4 ml of cold absolute ethanol were added to precipitate the unhydrolyzed residue and large fragments. After centrifugation, radioactivi ty was measured in an al iquot of the supe rna tan t solution, while another aliquot was subjected to acid hydrolysis and subsequent determinat ion of hydroxyproline. See the tex t for other details of the digestion procedure. Radio- act ivi ty expected from hydroxyprol ine released was calculated from the hydroxyprol ine released on the basis t ha t the molar ratio of hydroxyprol ine/prol ine is 21 and the specific-activity ratio of prol ine/hydroxyprol ine is 8 (averages of values in Table I I for NaCl-soluble collagen). The hydroxyprol ine would contain 73 % of the total radioactivity.
Enzyme % of original remaining in ethanol supernatant
Hydroxyproline Radioactivity Radioactivity expected from hydroxyproline released
1Nlone 1.4 2 I Collagenase 92 66 67 Tryps in 14 26 I o Elastase 7.6 19 6
Treatment with collagenase and other proteinases The data of Table I I I show that after collagenase treatment most of the
hydroxyproline was no longer precipitable with 6 7 % ethanol and the radioactivity released was that expected from the hydroxyproline released. Trypsin and elastase digestion, in contrast, released relatively little hydroxyproline in proportion to counts released. These results are qualitatively in accord with the conclusion that the com- ponent highly labeled in proline has little or no susceptibility to collagenase but is hydrolyzed by trypsin and elastase. Similar observations were made when such proteolytic digestion experiments were followed by dialysis instead of ethanol treat- ment to separate the unliydrolyzed residue (Table IV).
Biochim. Biophys. Aeta, lO 7 (1965) 232-246
238 D. FUJIMOTO, E. ADAMS
T A B L E IV
S P ] ~ C l t t l C A C T I V I T Y O F P R O L I N ~ A N D H Y D R O X Y P R O L I N ] ~ I N P U R I I * I ] ~ D C O L L A G E N F R A C T I O N S
NaCl-soluble collagen was p repa red as described, and cen t r i fuged t h r o u g h a sucrose gradient . The sucrose -grad ien t peak compr ised ma te r i a l pooled f rom t u b e s shown in Reg ions I I I and IV (Fig. IA). The col lagenase digest was t he dia lyzable f rac t ion ob ta ined af te r d igest ion of NaC1- soluble collagen as descr ibed in t he t ex t ; t he co l lagenase- res i s tan t f rac t ion was t h a t r e ta ined in t he dialysis bag af te r digest ion.
Fraction Specific activity (eounts/min per/zmole)
Proline Hydroxyproline Proline Hydroxyproline
NaCl-soluble 7400 906 8.2
Sucrose-grad ien t peak 1690 780 2.2
Collagenase digest 116o 77 ° 1.5
Collagenase r e s i s t an t 17 ooo* - - - -
* The specif ic-act ivi ty va lue of 17000 is a m i n i m u m , since i t inc ludes prol ine der ived f rom col lagenase as well as f rom t he co l lagenase- res i s tan t c o m p o n e n t of NaCl-soluble collagen.
A z : IF : ~ F i I ~ " : I i i , i
200- , : ' g ~
m 100- I - -
z
"~. 2 4 6 8 I0 12 14 16 18 20 22 24
2oo- B
100-
(__3 2 4 6 8 I0 f2 14 16 18 20 22 24
Q < 200- ft.,-
iO0-
TUBE NUMBER
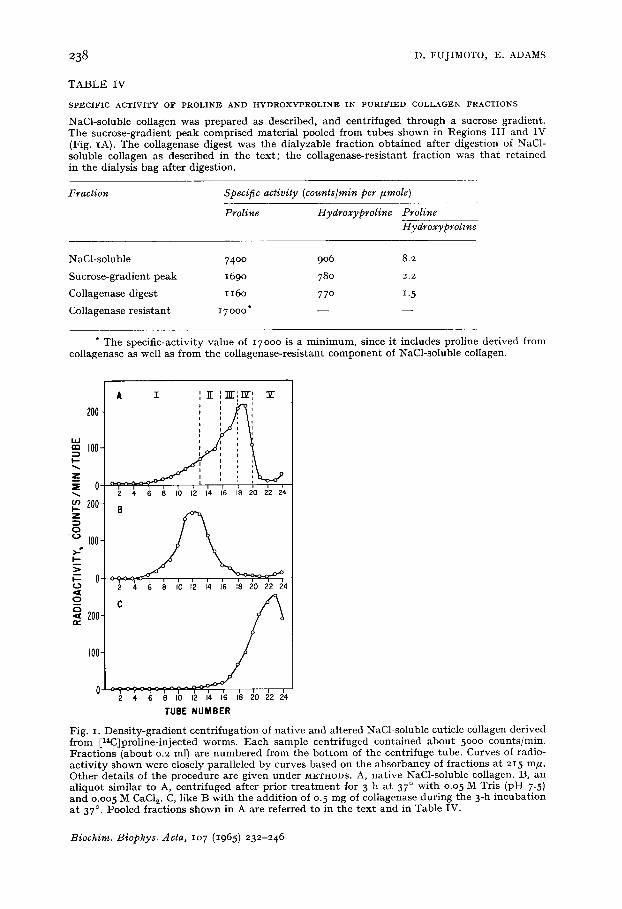
Fig. I. Dens i t y -g rad i en t cen t r i fuga t ion of na t i ve and al tered NaCl-soluble cuticle collagen der ived f rom [14C]proline-injected worms. E a c h sample cen t r i fuged con ta ined a b o u t 5000 counts / ra in . F rac t ions (about o.2 ml) are n u m b e r e d f rom t he b o t t o m of the cen t r i fuge tube. Curves of radio- ac t iv i ty s h o w n were closely paral le led by curves based on the a b s o r b a n c y of f rac t ions a t 2I 5 m# . O the r detai ls of t he p rocedure are g iven unde r M~THODS. A, na t ive NaCl-soluble collagen. B, an a l iquo t s imilar to A, cen t r i fuged af ter pr ior t r e a t m e n t for 3 h a t :37 ° wi th o.o 5 M Tris (pH 7.5) a n d o.oo 5 M CaC1 v C, like B wi th t he add i t ion of 0. 5 m g of col lagenase du r ing t he 3-h i ncuba t ion a t 37 °. Pooled f rac t ions shown in A are referred to in t he t ex t and in Table IV.
Biochim. Biophys. Acta, lO 7 (1965) 232-246
I N C O R P O R A T I O N S T U D I E S W I T H E A R T H W O R M - C U T I C L E C O L L A G E N 239
The small but significant release of hydroxyproline in dialyzable form by trypsin or elastase treatment may be due to partial denaturation of the collagen substrate during prolonged incubation.
Further purification of NaCl-soluble collagen On centrifugation through sucrose density gradients (3oooo rev./min for 18 h)
most of the radioactivity (about 7 ° % of the original counts were recovered) moved as a somewhat diffuse, asymmetric peak, shown in Fig. IA. That this peak was pre- dominantly native collagen was supported by the finding that its sedimentation velocity changed markedly after denaturation at 37 ° (Fig. IB) or after treatment with collagenase (Fig. IC).
The specific-activity ratio of proline to hydroxyproline in the peak region (Table IV) was only about one-fourth that of the original NaCl-soluble fraction, indi- cating removal of much of the contaminant. Separate fractions of the peak (Fractions I-V, Fig. I) each had approximately the same ratio of radioactivity of proline/ hydroxyproline. (Radioactivity ratios determined from chromatograms with carrier proline, rather than specific-activity ratios, were measured for these fractions because the low level of proline prevented reliable estimation.)
The contaminant separated from the main peak by sucrose-gradient centrifu- gation was not clearly identified elsewhere in the gradient. However, after removal of all gradient fractions, washings of the tubes contained material with a high ratio of radioactivity in proline to that in hydroxyproline (2.5), accounting for at least 5 % of the original total radioactivity applied to the gradient. Much of the contami- nant may have moved faster than the peak shown in Fig. IA and become adherent to the bottom of the tube.
The relative purity of the peak is suggested by the correspondence of its specific- activity ratio (proline/hydroxyproline) to that determined for dialyzable fragments released from collagenase digests of NaCl-soluble collagen (Table IV) when this procedure was carried out as described earlie#. It is noteworthy that residual proline remaining within the dialysis bag had a minimum specific activity many times that of the proline released by collagenase action (Table IV).
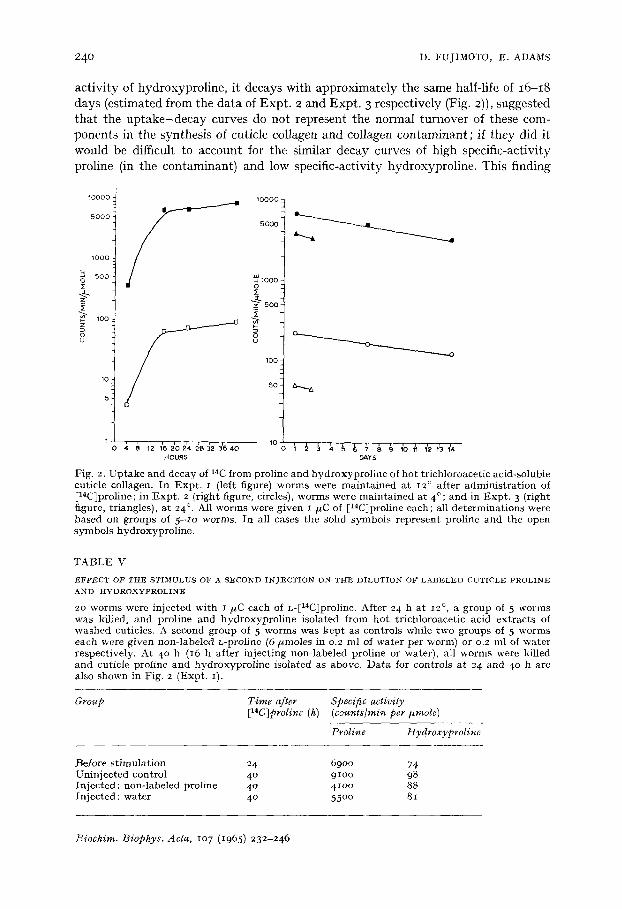
Kinetics of incorporation of [14Clproline Fig. 2 presents data on the appearance and decay of label in proline and
hydroxyproline derived from cuticle collagen after administering L@4C~proline to several groups of worms each kept at a different temperature during the experiment. Groups of worms used for different experiments showed considerable differences in the absolute value of imino acid specific activities attained, with no consistent relation to storage temperature. However, within a single group, uptake and decay curves showed consistent results and suggested (Expts. 2, 3) a slow exponential disappearance of label from both proline and hydroxyproline.
In all three experiments, it is striking that the specific activity of proline, although approx, i00 times greater than that of hydroxyproline (cf. Table II), decayed with the same time course, so that the ratio of specific activities is preserved at each point within the same experiment, both during the phase of uptake and of decay of label.
The unexpected finding that although proline attains IOO times the specific
Biochim. Biophys. Acta, IO 7 (1965) 2 3 2 - 2 4 6
240 D. FUJIMOTO, E. ADAMS
activity of hydroxyproline, it decays with approximately the same half-life of I6-I8 days (estimated from the data of Expt. 2 and Expt. 3 respectively (Fig. 2)), suggested that the uptake-decay curves do not represent the normal turnover of these com- ponents in the synthesis of cuticle collagen and collagen contaminant; if they did it would be difficult to account for the similar decay curves of high specific-activity proline (in the contaminant) and low specific-activity hydroxyproline. This finding
Fig, 2. U p t a k e and decay of 14C from prol ine and h y d r o x y p r o l i n e of ho t t r ich loroacet ic acid-soluble cut ic le collagen. In E x p t . I (left figure) worms were m a i n t a i n e d a t 12 ° af ter a d m i n i s t r a t i o n of [14C]proline; in E x p t . 2 ( r ight figure, circles), worms were m a i n t a i n e d a t 4°; and in E x p t . 3 (right figure, t r iangles) , a t 24 °. All worms were g iven ~/~C of [14C]proline each; al l de t e rmina t ions were based on groups of 5 ~ o worms. In al l cases the solid symbols represen t pro l ine and the open symbols hydroxypro l ine .
T A B L E V
~FFECT OF THE STIMULUS OF A SECOND INJECTION ON THE DILUTION O1~ LABELED CUTICLE PROLINE AND HyDROXYPROLINE
20 worms were in jec ted w i t h I /zC each of L-[14C~proline. Af ter 24 h a t I2 °, a g roup of 5 worms was killed, and prol ine and h y d r o x y p r o l i n e i so la ted f rom ho t t r i ch loroace t ic acid ex t r ac t s of washed cuticles. A second group of 5 worms was k e p t as controls while two groups of 5 worms each were g iven non- labeled L-proline (6/2moles in 0.2 ml of w a t e r per worm) or o.2 nil of wa te r respect ive ly . At 4 ° h (16 h af ter in jec t ing non- labeled prol ine or water) , al l worms were ki l led and cut ic le prol ine and h y d r o x y p r o l i n e isola ted as above. D a t a for controls a t 24 and 4 ° h are also shown in Fig. 2 (Expt . i).
Group Time after Specific activity [l*Clproline (h) (counts~rain per #mole)
Proline Hydroxyproline
Before s t imu la t i on 24 6900 74 Unin jec ted control 4 ° 91oo 98 In jec ted : non- labeled prol ine 4 ° 4 z ° ° 88 In j ec t ed : wa te r 4 ° 55oo 81
t~iochim. Biophys. Acta, lO 7 (1965) 232-246
INCORPORATION STUDIES WITH EARTHWORM-CUTICLE COLLAGEN 241
could be consistent, however, with the hypothesis that the manner of administering [14C}proline, i.e., by injection, of itself stimulated the rapid synthesis, for a short period of time, of the proline-rich contaminant, which normally has a low turnover rate, comparable to that of collagen.
An attempt to examine this possibility was made by observing the effects of injection alone (administration of non-labeled proline or of water) on the decay of label already present in cuticle proline and hydroxyproline. As shown in Table V, the injection of water or non-labeled proline 24 h after administering E14Clproline produced a sharp drop in the specific activity of [14Clproline, as measured 16 h after the second injection. The effect of these procedures on the subsequent specific activity of hydroxyproline, however, was too small to evaluate for significance. These results suggest that the act of injection of itself stimulated synthesis of the proline-containing contaminant, and offer a plausible explanation for the unexpected similarity of the decay rate shown for proline and hydroxyproline in cuticle. The interesting question of a possible parallel, or even linked, effect of injection on the stimulated synthesis of collagen could not be appraised in these experiments. If one makes the reasonable
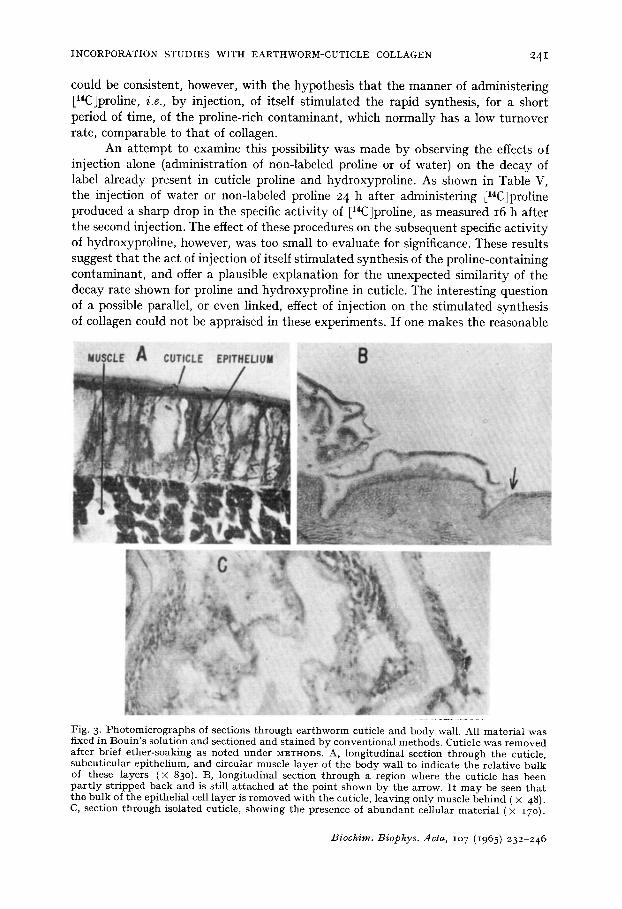
Fig. 3. P h o t o m i c r o g r a p h s of sect ions t h r o u g h e a r t h w o r m cuticle and body wall. All ma te r i a l was fixed in Bou in ' s solut ion and sect ioned and s ta ined by conven t iona l me thods . Cut ic le was r e m o v e d af ter brief e ther - soak ing as no ted unde r METHODS. A, longi tud ina l sect ion t h r o u g h t he cuticle, subcu t i cu la r ep i the l ium, and circular muscle layer of the body wall to indicate t he re la t ive bu lk of these layers (× 830). t3, long i tud ina l sect ion t h r o u g h a region where the cuticle ha s been p a r t l y s t r ipped back and is still a t t a c h e d a t the po in t shown by t he arrow. I t m a y be seen t h a t t he bu lk of t he epithelial cell l ayer is r emoved wi th t he cuticle, l eaving only musc le beh ind ( X 48). C, sect ion t h r o u g h isolated cuticle, showing t he presence of a b u n d a n t cellular ma te r i a l (X 17o ).
Biochim. Biophys. Acta, Io 7 (1965) 232-246
24 2 D. FUJIMOTO, E. ADAMS
postulation that the highly labeled proline component of cuticle is small relative to the collagen component, then stimulated synthesis of both components, clearly de- tectable in the contaminant, might well be imperceptible in the hydroxyproline of collagen.
Although it would be of interest to administer [14Clproline to worms in a manner that did not involve the stimulation of injection, preliminary trials of other routes of administration did not appear promising.
Possible source of contamination I t seemed relevant to consider the possible extent of contamination of cuticle
with underlying tissue when the former is removed as described above. Histological examination (Fig. 3) indicated that "cuticle" stripped from ether-treated worms contains a more-or-less intact adherent layer of underlying epithelium. There is thus an opportunity for gross contamination of cuticle fractions with non-collagenous material derived from the epithelium, even though much epithelial material may be removed by the subsequent washing procedure a.
I t should be noted in particular that the subcuticular epithelium would appear to be the source both of cuticle collagen and of the abundant mucus secretion of Lumbricus 22. Although little detailed chemical information concerning earthworm mucus is available, the solubility characteristics of other mucoproteins ~ would suggest that hot trichloroacetic acid could extract not only collagen but residual mucus or mucus precursors arising from contaminating epithelium. Experiments were done in which mucus or washings of cuticle from [14C]proline-injected worms were mixed with unlabeled NaCl-soluble collagen. After two or more successive precipitations of the latter with 20% saturated (NH4)2SO 4 only a small fraction (2- 3 %) of the initial counts were recovered with the precipitated collagen. Although the results of these artificial mixing experiments do not eliminate the possibility of absorption of a con- taminating soluble protein under conditions in vivo, they offered no support for the significant absorption to collagen of counts in unrelated proteins.
Extraction of various collagen fractions In vertebrate preparations, neutral-salt-soluble collagen has been shown to be
metabolically more active than acid-soluble or insoluble collagen 24. Similar relations were observed in Lumbricus cuticle. As shown in Table VI, when collagen was succes- sively extracted from cuticle with 0.5 M NaC1, 0.5 % acetic acid and IO % trichloro- acetic acid at 9 o°, the specific activity of proline and hydroxyproline decreased in the respective fractions. I t is of particular interest that the specific activity of proline in these fractions showed a progressive decrease quantitatively similar to that of hydroxyproline, indicating another point of similarity in the extraction behavior of the proline-labeled contaminant and that of collagen.
Incorporation of I14Clproline into 3- and 4-hydroxyproline of body collagen It was recently reported by the present authors s that collagen in the body of
Lumbricus shows a markedly different amino acid composition from cuticle collagen. Composite body collagen fractions contain less 4-hydroxyproline, more proline and much more 3-hydroxyproline than cuticle collagen. The 3-hydroxyproline component of worm body was found by high-voltage paper electrophoresis to migrate identically
Biochim. Biophys. Acta, lO7 (1965) 232-246
INCORPORATION STUDIES WITH EARTHWORM-CUTICLE COLLAGEN 243
TABLE VI
SPECIFIC ACTIVITY OF PROLINE AND HYDROXYPROLI.NE IN SUCCESSIV~ EXTRACTION FRACTIONS OF
CUTICLE COLLAGEN
Washed cuticles were extracted successively with o. 5 M NaC1 at 4 ° for 48 h, o. 5 % acetic acid at 4 ° for 48 h and IO ~o trichloroacetic acid at 90 ° for 60 min. No hydroxyprol ine was found in the residue after ho t trichloroacetic acid extraction.
Extraction solvent Specific activity (counts~rain per ~,mole)
Proline Hydroxyproline Proline Hydroxyproline
0. 5 M NaCI* 24000 512o 4.7 0.5 % acetic acid 7600 114o 6.7 Ho t trichloroacetic acid 4o60 318 13
* The 0. 5 M NaC1 extract was fur ther t reated with 2 successive (NH4)2SO 4 precipitat ions as in the NaCl-soluble prepara t ion noted under METHODS.
T A B L E V I I
INCORPORATION OF [14C]PROLINE INTO 4-HYDROXYPROLINE AND 3-HYDROXYPROLINE OF BODY AND
CUTICLE COLLAGEN
A single worm was given 7.5/ ,C of L-[14C]proline and kept at 12 ° for 2 4 h. The washed cuticle and cuticle-stripped body were extracted with hot trichloroacetic acid as described in the text . Proline and hydroxyprol ine were separated by paper ch romatography and the hydroxyprol ine fraction, after addition of I / , m o l e of carrier 3-hydroxyproline, was separated into 3- and 4- hydroxyprol ine by high-voltage paper electrophoresis.
Cuticle Body
4-Hydroxypro l ine , / ,moles recovered o. 173 o.694 4-Hydroxyproline, counts /min 165 849 4-Hydroxyprol ine, counts]min p e r / , m o l e 952 122o
3-Hydroxyproline, counts /min - - 4 ° 3-Hydroxyprol ine/4-hydroxyprol ine, counts /min - - 0.0 5 3-Hydroxyprol ine/4-hydroxyprol ine, /,moles* - - o. I I
* Determined f rom separate prepara t ions s.
with that synthetic racemate n which has recently been identified as the t rans form 25. As shown in Table VII, incorporation of Ei4Clproline into 4-hydroxyproline of
body collagen occurred a little more rapidly than in cuticle. An appreciable amount of radioactivity was found in the 3-hydroxyproline fraction, and the radioactivity ratio of 3-hydroxyproline/4-hydroxyproline, 0.05-0.06, roughly agreed with the molar ratio, which was measured as o . I i by automatic amino acid analysis s. The discrepancy can plausibly be explained by poor recovery of these amino acids during the isolation procedure and/or by the heterogeneity of body collagen with respect to the turnover rates of collagen fractions relative to their content of 3-hydroxyprollne. We interpret these results as consistent with the conclusion that 3-hydroxyproline of collagen is synthesised from free proline, probably in a manner similar to 4-hydroxyproline. (While this paper was in preparation a report appeared by KAPLAN e~ a/fi e concluding that proline is converted to 3-hydroxyproline of chick-embryo collagen, with the same specific activity as that of 4-hydroxyproline.)
Bioehim. Biophys. Acta, lO 7 (1965) 232-246
2 4 4 D. FUJIMOTO, E. ADAMS
DISCUSSION
The findings above indicate that despite its remarkably high content of hydroxy- proline and its other physical singularities, the collagen of earthworm cuticle re- sembles vertebrate collagen in that proline rather than free hydroxyproline is the source of collagen hydroxyproline.
An unexpected finding was the presence of a protein fraction which accompanies cuticle collagen through the salt extraction and precipitation procedure of JOSSE A.X~D HARRINGTON 5. This component was judged not to be collagen on the basis of its in- susceptibility to collagenase, its susceptibility to other proteinases at low temperature, and its selective content of highly labeled proline with little or no comparably labeled hydroxyproline. Further purification of collagen through sucrose gradients apparently removed much of the contaminant, the peak sucrose fractions yielding material with a proline/hydroxyproline specific-activity ratio of about 2, close to that obtained in the dialyzable fraction released after collagenase digestion.
The nature of the contaminant (if indeed it be a single component) and its possible biological relationship to collagen, are unsettled. I t would appear to be a protein and not a low-molecular-weight substance absorbed to collagen because it was retained by collagen preparations through extensive dialysis and repeated salt precipitation and was selectively released in dialyzable form after treatment with trypsin and with elastase, but not after treatment with collagenase.
The contaminating material could represent trivial contamination from a highly labeled protein fraction unrelated to collagen. In the latter case, repeated salt precipi- tation should markedly decrease the proline/hydroxyproline specific-activity ratio. When this was tested, the ratio of radioactivity in proline to that in hydroxyproline changed only slightly in NaCl-soluble collagen reprecipitated a third time with 20 o~/o saturated (NH4)2SO 4.
Alternatives of more biological interest, that the contaminating material is structurally and biologically closely related to collagen, receive some support. Thus the high specific-activity proline fraction appears soluble in hot trichloroacetic acid and also contaminates purified NaCl-soluble preparations, although not proportion- ally. It shows the same relationship as does hydroxyproline (and hence collagen) between specific activity and extraction procedure, and it decays at the same rate as does collagen hydroxyproline, even though present at much higher specific activity in both hot trichloroacetic acid-soluble and NaCl-soluble fractions.
The consideration that this material might be a non-hydroxylated collagen- precursor is greatly weakened by the finding that it appears resistant to collagenase, particularly since the specificity of collagenase appears to depend on primary structure in which proline and hydroxyproline are interchangeable without qualitative effect on susceptibility to enzymatic hydrolysis 27,2s. Other proteins plausibly associated with the collagen matrix, such as elastin or mucoprotein, might be considered.
The identity of the contaminant as elastin appears unlikely from the former's susceptibility to trypsin29; a mucoprotein cannot be dismissed particularly since such proteins may be soluble in hot trichloroacetic acid 23. Cuticle collagen preparations are rich in carbohydrate 3-5, some of which may well be associated with mucins, and the high specific activity of the contaminant in proline and apparent biosynthetic stimulation by trauma would fit a possible relationship to mucus synthesis.
Biochim. Biophys. Acta, lO 7 (i965) 232-246
INCORPORATION STUDIES WITH EARTHWORM-CUTICLE COLLAGEN 245
The small quantity of proline in the contaminant, however, is indicated by the marked disparity between varying specific-activity ratios and constant molar ratios of proline to hydroxyproline in the several collagen fractions studied. This in turn suggests that the contaminating protein may be present at only a few per cent of the collagen, assuming that the contaminant is a protein of average proline content. This consideration dictates caution in suggesting a resemblance to collagen based on solubility characteristics.
It is notable that many studies of vertebrate collagen labeled from proline have failed to detect a highly labeled proline contaminant, even though collagen prepa- rations were often quite crude. We have found only a few suggestions in the literature of a possible collagen-associated highly labeled proline component. Thus, trichloro- acetic acid extracts of carageenin-granulomas from ascorbate-deficient guinea pigs 21 showed high specific-activity ratios of proline to hydroxyproline. A striking obser- vation, which may be relevant to our own, comprises data presented in a recent paper by PETERKOFSKY AND UDENFRIEND 30. Here (Fig. 3, ref. 30) individual CM-cellulose fractions (derived from chick-embryo microsomes extracted with hot trichloroacetic acid), which resembled bovine Achilles-tendon gelatin fractions in chromatographic behavior, contained up to Io-fold greater labeling in proline than hydroxyproline; these findings indicate the presence of a component more highly labeled in proline than hydroxyproline, but one which resembles collagen enough to coincide with several chromatographically separated peaks of the gelatinized material. The data given do not permit an estimation of the specific-activity ratio of proline to hydroxy- proline.
In carrying out our proteolytic studies of the proline-rich contaminant, inci- dental information appeared concerning the susceptibility of earthworm-cuticle collagen to collagenase. In confirmation and extension of the finding of MASER AND RICE 6 that collagenase reduces the viscosity of cuticle collagen, we have found that more than 9 ° % of cuticle collagen hydroxyproline becomes dialyzable after collagen- * ase treatment.
Essentially complete degradation of cuticle collagen by collagenase to yield readily dialyzable fragments indicates hydrolysis at many points along the peptide chain. Assuming the generally accepted preference of collagenase for the sequence X-Pro-Y-Gly-Pro (where Pro can be either proline or hydroxyproline and hydrolysis occurs at the Y-Gly bond), one might postulate the frequent occurrence of triplets Gly-Hypro-X in earthworm-cuticle collagen, because of the predominance of hy- droxyproline as a pyrrolidine residue. This position for hydroxyproline has been con- sidered unlikely in vertebrate collagen, however, where digests show it to occupy the third position with considerable frequency in glycine triplets released by collagen- ase, i.e., Gly-X-Hypro (for a recent bibliographic summary, see ref. 5)- The possibility that earthworm-cuticle collagen contains significant numbers of the sequence Gly- Hypro-Hypro, has been questioned 5. It should be acknowledged that the action of collagenase is not strictly specific 28 for the sequence noted above, and that the ex- haustive digestions employed by us might reflect the results of a less prominent sequence-specificity of collagenase.
In any case, it is apparent that the unusual composition of earthworm-cuticle collagen raises further questions with regard to its collagenase susceptibility, just as it does for other structural features of collagen.
Biochim. Biophys. Acta, lO 7 (1965) 232-246
246 D. FUJIMOTO, E. ADAMS
ACKNOWLEDGEMENTS
This work was supported by U.S. Public Health Service Grant GM II lO 5. We wish to thank Dr. H. J. LINDER, Department of Zoology, University of Maryland, for advice and information concerning the biology and maintenance of earthworms, and Dr. H. I. FIRMINGER and Mr. H. BERGMANN of the Department of Pathology, University of Maryland School of Medicine, for help in the preparation and inter- pretation of histological material.
R E F E R E N C E S
I I{. M. RUDALL, Progr. Biophys. Biophys. Chem., I (195o) 39. 2 L. SINGLETON, Biochim. Biophys. Aeta, 24 (1957) 67. 3 M. R. WATSON, Bioehem. J., 68 (1958) 416. 4 M. D. MASER AND R. V. RICE, Biochim. Biophys. Aeta, 63 (1962) 255. 5 J- JossE AND W. F. HARRINGTON, J. Mol. Biol., 9 (1964) 269. 6 M. D. MASER AND R. V. RICE, Biochim. Biophys. Acta, 74 (1963) 283. 7 R. REED AND K. M. RUDALL, Bioehim. Biophys. Aeta, 2 (1948) 7. 8 D. FUJIMOTO AND E. ADAMS, Bioehem. Biophys. Res. Commun., 17 (1964) 437. 9 W. VAN B. ROBERTSON, Biophys. J. , 4 (1964) 93.
io E. ADAMS AND A. GOLDSTONR, J. Biol. Chem., 235 (196o) 3492. i i J. D. OGLE, R. B. ARLINGHAUS AND M. A. LOGAN, J. Biol. Chem., 237 (1962) 3667. 12 S. M. FITCH, M. R. HARKNESS ANn R. D. HARKNESS, Nature, 176 (1955) 163. 13 K. A. PIEZ, F. IEREVERRE AND H. L. WOLFF, J. Biol. Chem., 223 (1956) 687. 14 R. E. NEUMAN AND M. A. LOGAN, J. Biol. Chem., 184 (195 o) 299. 15 R. G. MARTIN AND B. N. AMES, J. Biol. Chem., 236 (1961) 1372. 16 J. C. SHEEHAN AND J. G. WHITNEY, J. Am. Chem. Soc., 84 (1962) 3980. 17 C. MITOMA, T. E. SMITH, F. FRIEDBERG AND C. R. RAYFORD, J. Biol. Chem., 234 (1959) 78. 18 W. VAN B. ROBERTSON, J. HIWETT AND C. HERMAN, J. Biol. Chem., 234 (1959) lO5. 19 N. M. GREEN AND D. A. LOWTHER, Biochem. J., 71 (1959) 55. 20 E. HAUSMANN AND W. F. NEUMAN, J. Biol. Chem., 236 (1961) 149. 21 N. STONE AND A. MEISTER, Nature, 194 (1962) 555. 22 M. S. LAVERACK, The Physiology of Earthworms, Pergamon Press, Oxford, 1963, p. 12. 23 R. G. SPIRO, New Engl. J. Med., 269 (I963J 566. 24 D. A. LOWTHER, lntern. Rev. Connective Tissue Res., I (1963) ioo. 25 F. IRREVERRE, K. MORITA, A.V. ROBERTSON AND B. WITKOP, J. Am. Chem. Soe., 85 (1963) 2824. 26 A. KAPLAN, B. WITKOP AND S. UDENFRIEND, J. Biol. Chem., 239 (19641 2559. 27 W. GRASSMANN, H. H6RMANN, A. NORDWlG AND E. WUNSCH, Z. Physiol. Chem., 316 (1959) 287. 28 Y. NAGAI, S. SAKAKIBARA, H. lXlODA AND S. AKABORI, Bioehim. Biophys. Acta, 37 (196o1 567- 29 I. MANDL, Advan. Enzymol., 23 (1961) 225. 30 B. PETERKOFSKY AND S. UDENFRIEND, J. Biol. Chem., 238 (1964) 3966.