THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1985 by The American Society of Biological Chemists, Inc Vol. 260, No. 11, Issue of June 10. pp. 7029-7034,1985 Printed in V.S.A. Protease Nexin PROPERTIES AND A MODIFIED PURIFICATION PROCEDURE* (Received for publication, November 6, 1984) Randy W. Scott".', Barbara L. Bergman", Ani1 Bajpai", R. T. Hersh", Henry Rodriguez", Barry N. Jonesd, Carlos Barredad,Sherry Watts", and Joffre B. Bakera*' From the "Department of Biochemistry, University of Kansas, Lawrence, Kansas 66045, the 'Department of Protein Biochemistry, Genentech, Inc., South San Francisco, California 94080, and dUnigeneLaboratories, Fairfield, New Jersey 07006 The present paper describes chemical and functional properties of protease nexin, a serine protease inhibi- tor released from cultured human fibroblasts. It is shown that protease nexin is actually synthesized by fibroblasts and represents about 1%of their secreted protein. Analysis of the amino acid composition of pu- rified protease nexin indicates that it is evolutionarily related to antithrombin I11 and heparin cofactor 11. Protease nexin contains approximately 6% carbohy- drate, with 2.3% amino sugar, 1.1% neutral sugar, and 3.0% sialic acid. The M, calculated from equilibrium sedimentation analysis is 43,000. Protease nexin is a broad specificity inhibitor of trypsin-like serine pro- teases. It reacts rapidly with trypsin (k- = 4.2 f 0.4 x 10' M-' s-I), thrombin (k- = 6.0 f 1.3 X 10" M" s-'), urokinase (k- = 1.5 f 0.1 X lo6 M" s-'), and plasmin (k- = 1.3 f 0.1 X lo6 M" s-'), and slowly inhibits Factor Xa and the y subunit of nerve growth factor but does not inhibit chymotrypsin-like proteases or leukocyte elastase. In the presence of heparin, pro- tease nexin inhibits thrombin at a nearly diffusion- controlled rate. Two heparin affinity classes of pro- tease nexin can be detected. The present characteri- zation pertains to the fraction of protease nexin having the higher affinity for heparin. The low affinity ma- terial, which is the minor fraction, is lost during puri- fication. Protease nexin is a mammalian protein that forms 1:1 complexes with and inhibits several serine proteases including thrombin, plasmin, urokinase, and tissue-type plasminogen activator (1-4). Because protease nexin releases bound pro- teases at pH 12 or in 1 M hydroxylamine and does not complex serine proteases inhibited with diisopropylfluorophosphate (l), it appears that protease nexin forms carboxylic ester links with serine proteases at their active site serine residues, a reaction that is characteristic of inhibitors in the antithrom- bin III/al-antitrypsin family (5, 6). Protease nexin and anti- thrombin 111, although electrophoretically and immunologi- cally distinct have the additional similarity that heparin binds to both with high affinity and accelerates their binding of thrombin (1). Protease nexin is secreted by many different * This work was supported by National Institutes of Health Re- search Grant CA-29307. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. Section 1734 solelyto indicate this fact. bPresent address: Unigene Laboratories, 110 Little Falls Road, Fairfield, NJ 07006. Recipient of National Institutes of Health Research Career De- velopment Award CA-00886. anchorage-dependent cells including fibroblasts, heart muscle cells, and kidney epithelial cells (7). Fibroblasts and perhaps all protease nexin-secreting cells bear surface receptors that mediate the endocytosis and lysosomal degradation of pro- tease-protease nexin complexes (8).' In serum-free cultures of human fibroblasts, secreted pro- tease nexin suppresses cellular sensitivity to the mitogenic action of added thrombin (13,141, limits activation of secreted urokinase by exogenous proteases (4, 15), and inhibits acti- vated urokinase (3, 4). However, it is not certain that these are major physiological functions of this inhibitor. The present paper describes chemical, physical, and func- tional features of protease nexin. Since inhibitors often (al- though not always) react more rapidly with their physiological target proteases than with other proteases (16, 17) one aim of the present investigation was to identify serine proteases that react rapidly with protease nexin and might therefore be potential physiological targets of this inhibitor. EXPERIMENTAL PROCEDURES Materials-Human a-thrombin, originally purified in Dr. Roger Lundblad's laboratory (University of North Carolina), was a gift from L. L. Houston (University of Kansas). Human leukocyte elastase and human leukocyte cathepsin G were gifts from Dr. James Travis (University of Georgia). Mouse y-nerve growth factor, originally purified in Dr. Ralph Bradshaw's laboratory (University of California, Irvine), was a gift from Dr. Dan Raben (University of California, Irvine). Urokinase was from Abbott. Human factor Xa was obtained from Boehringer Mannheim. a-Chymotrypsin was from Worthington. Plasminogen was purified from outdated human serum by the pro- cedure of Deutsch and Mertz (18). Plasmin was prepared by the method of Dan0 and Reich (19). The following were obtained from Sigma: bovine trypsin, 4-meth- ylumbelliferyl-p-guanidinobenzoate, 4-methylumbelliferone, N-suc- cinyl-Ala-Ala-p-nitroanilide, D-Val-Leu-Lys-p-nitroanilide, N-ben- zoyl-Phe-Val-Arg-p-nitroanilide, p-nitrophenyl guanidinobenzoate, N-benzoyl-Arg-ethyl ester, and bovine serum albumin. Succinyl-Ala- Ala-Pro-Phe-p-nitroanilide was from Vega. The chromogenic sub- strates s-2444, S-2288, and S-2222 were from Helena Laboratories. Purification and Characterization-Protease nexin was purified from serum-free medium conditioned by human foreskin fibroblasts in microcarrier cultures, as previously described (2), with the follow- ing modifications. Sodium chloride was added to the harvest buffer to bring the final NaCl concentration in medium plus buffer to 0.28 M. Triton X-100 was omitted from the harvest buffer, andthe harvested medium was filtered through a 45-p Millipore filter before carrying out protein concentration by (Amicon) hollow filter ultra- ' Human fibroblasts secrete at least two other serine protease- binding factors, which have different protease-binding specificities, but also mediate the binding of their complexed proteases to the cells (9,lO). It has been proposed that these other factors be called protease nexin I1 and protease nexin 111, and that the urokinase and thrombin- binding protease nexin be called either protease nexin I or protease nexin (11, 12). The latter name is used here. 7029
Transcript
THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1985 by The American Society of Biological Chemists, Inc
Vol. 260, No. 11, Issue of June 10. pp. 7029-7034,1985 Printed in V.S.A.
Protease Nexin PROPERTIES AND A MODIFIED PURIFICATION PROCEDURE*
(Received for publication, November 6, 1984)
Randy W. Scott".', Barbara L. Bergman", Ani1 Bajpai", R. T. Hersh", Henry Rodriguez", Barry N. Jonesd, Carlos Barredad, Sherry Watts", and Joffre B. Bakera*' From the "Department of Biochemistry, University of Kansas, Lawrence, Kansas 66045, the 'Department of Protein Biochemistry, Genentech, Inc., South San Francisco, California 94080, and dUnigene Laboratories, Fairfield, New Jersey 07006
The present paper describes chemical and functional properties of protease nexin, a serine protease inhibi- tor released from cultured human fibroblasts. It is shown that protease nexin is actually synthesized by fibroblasts and represents about 1% of their secreted protein. Analysis of the amino acid composition of pu- rified protease nexin indicates that it is evolutionarily related to antithrombin I11 and heparin cofactor 11. Protease nexin contains approximately 6% carbohy- drate, with 2.3% amino sugar, 1.1% neutral sugar, and 3.0% sialic acid. The M, calculated from equilibrium sedimentation analysis is 43,000. Protease nexin is a broad specificity inhibitor of trypsin-like serine pro- teases. It reacts rapidly with trypsin (k- = 4.2 f 0.4 x 10' M-' s-I) , thrombin (k- = 6.0 f 1.3 X 10" M" s-'), urokinase (k- = 1.5 f 0.1 X lo6 M" s-'), and plasmin (k- = 1.3 f 0.1 X lo6 M" s-'), and slowly inhibits Factor Xa and the y subunit of nerve growth factor but does not inhibit chymotrypsin-like proteases or leukocyte elastase. In the presence of heparin, pro- tease nexin inhibits thrombin at a nearly diffusion- controlled rate. Two heparin affinity classes of pro- tease nexin can be detected. The present characteri- zation pertains to the fraction of protease nexin having the higher affinity for heparin. The low affinity ma- terial, which is the minor fraction, is lost during puri- fication.
Protease nexin is a mammalian protein that forms 1:1 complexes with and inhibits several serine proteases including thrombin, plasmin, urokinase, and tissue-type plasminogen activator (1-4). Because protease nexin releases bound pro- teases at pH 12 or in 1 M hydroxylamine and does not complex serine proteases inhibited with diisopropylfluorophosphate (l) , it appears that protease nexin forms carboxylic ester links with serine proteases at their active site serine residues, a reaction that is characteristic of inhibitors in the antithrom- bin III/al-antitrypsin family (5, 6). Protease nexin and anti- thrombin 111, although electrophoretically and immunologi- cally distinct have the additional similarity that heparin binds to both with high affinity and accelerates their binding of thrombin (1). Protease nexin is secreted by many different
* This work was supported by National Institutes of Health Re- search Grant CA-29307. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Recipient of National Institutes of Health Research Career De- velopment Award CA-00886.
anchorage-dependent cells including fibroblasts, heart muscle cells, and kidney epithelial cells (7). Fibroblasts and perhaps all protease nexin-secreting cells bear surface receptors that mediate the endocytosis and lysosomal degradation of pro- tease-protease nexin complexes (8).'
In serum-free cultures of human fibroblasts, secreted pro- tease nexin suppresses cellular sensitivity to the mitogenic action of added thrombin (13,141, limits activation of secreted urokinase by exogenous proteases (4, 15), and inhibits acti- vated urokinase (3, 4). However, it is not certain that these are major physiological functions of this inhibitor.
The present paper describes chemical, physical, and func- tional features of protease nexin. Since inhibitors often (al- though not always) react more rapidly with their physiological target proteases than with other proteases (16, 17) one aim of the present investigation was to identify serine proteases that react rapidly with protease nexin and might therefore be potential physiological targets of this inhibitor.
EXPERIMENTAL PROCEDURES
Materials-Human a-thrombin, originally purified in Dr. Roger Lundblad's laboratory (University of North Carolina), was a gift from L. L. Houston (University of Kansas). Human leukocyte elastase and human leukocyte cathepsin G were gifts from Dr. James Travis (University of Georgia). Mouse y-nerve growth factor, originally purified in Dr. Ralph Bradshaw's laboratory (University of California, Irvine), was a gift from Dr. Dan Raben (University of California, Irvine). Urokinase was from Abbott. Human factor Xa was obtained from Boehringer Mannheim. a-Chymotrypsin was from Worthington. Plasminogen was purified from outdated human serum by the pro- cedure of Deutsch and Mertz (18). Plasmin was prepared by the method of Dan0 and Reich (19).
The following were obtained from Sigma: bovine trypsin, 4-meth- ylumbelliferyl-p-guanidinobenzoate, 4-methylumbelliferone, N-suc- cinyl-Ala-Ala-p-nitroanilide, D-Val-Leu-Lys-p-nitroanilide, N-ben- zoyl-Phe-Val-Arg-p-nitroanilide, p-nitrophenyl guanidinobenzoate, N-benzoyl-Arg-ethyl ester, and bovine serum albumin. Succinyl-Ala- Ala-Pro-Phe-p-nitroanilide was from Vega. The chromogenic sub- strates s-2444, S-2288, and S-2222 were from Helena Laboratories.
Purification and Characterization-Protease nexin was purified from serum-free medium conditioned by human foreskin fibroblasts in microcarrier cultures, as previously described (2), with the follow- ing modifications. Sodium chloride was added to the harvest buffer to bring the final NaCl concentration in medium plus buffer to 0.28 M. Triton X-100 was omitted from the harvest buffer, and the harvested medium was filtered through a 45-p Millipore filter before carrying out protein concentration by (Amicon) hollow filter ultra-
' Human fibroblasts secrete at least two other serine protease- binding factors, which have different protease-binding specificities, but also mediate the binding of their complexed proteases to the cells (9,lO). It has been proposed that these other factors be called protease nexin I1 and protease nexin 111, and that the urokinase and thrombin- binding protease nexin be called either protease nexin I or protease nexin (11, 12). The latter name is used here.
7029
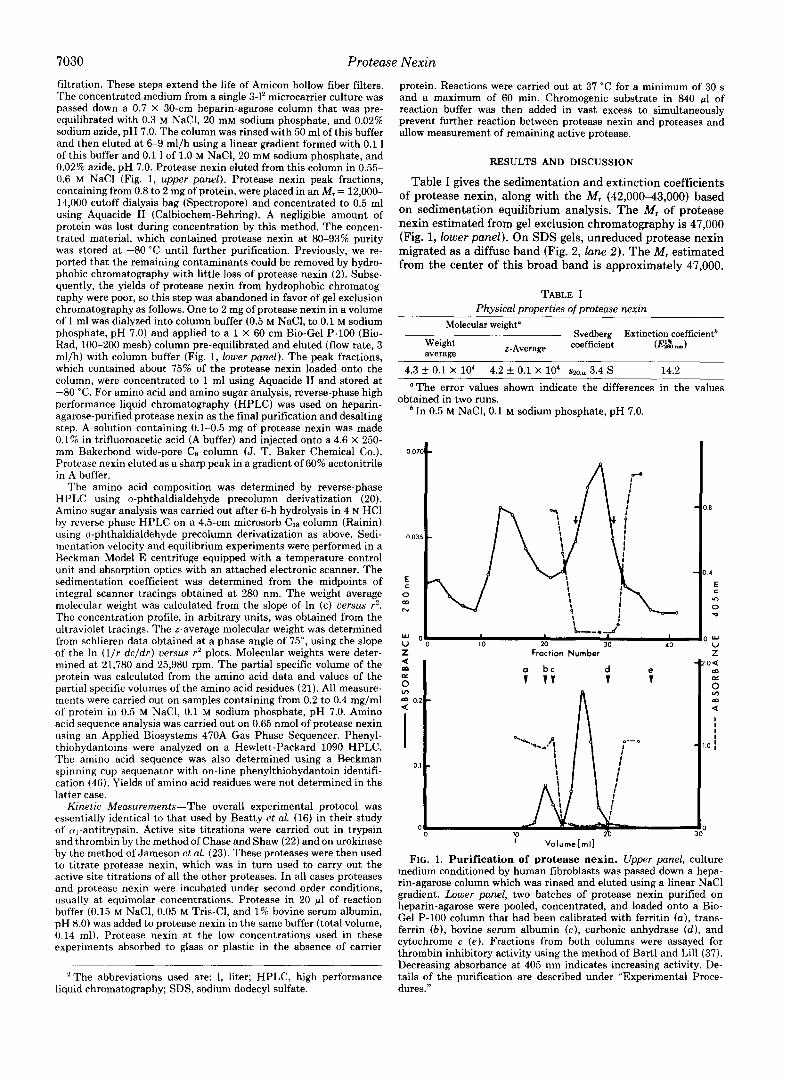
7030 Protease Nexin filtration. These steps extend the life of Amicon hollow fiber filters. The concentrated medium from a single 3-1' microcarrier culture was passed down a 0.7 X 30-cm heparin-agarose column that was pre- equilibrated with 0.3 M NaCl, 20 mM sodium phosphate, and 0.02% sodium azide, pH 7.0. The column was rinsed with 50 ml of this buffer and then eluted at 6-9 ml/h using a linear gradient formed with 0.1 1 of this buffer and 0.1 1 of 1.0 M NaC1, 20 mM sodium phosphate, and 0.02% azide, pH 7.0. Protease nexin eluted from this column in 0.55- 0.6 M NaCl (Fig. 1, upper panel). Protease nexin peak fractions, containing from 0.8 to 2 mg of protein, were placed in an M , = 12,000- 14,000 cutoff dialysis bag (Spectropore) and concentrated to 0.5 ml using Aquacide I1 (Calbiochem-Behring). A negligible amount of protein was lost during concentration by this method. The concen- trated material, which contained protease nexin at 80-93% purity was stored a t -80 "C until further purification. Previously, we re- ported that the remaining contaminants could be removed by hydro- phobic chromatography with little loss of protease nexin (2). Subse- quently, the yields of protease nexin from hydrophobic chromatog- raphy were poor, so this step was abandoned in favor of gel exclusion chromatography as follows. One to 2 mg of protease nexin in a volume of 1 ml was dialyzed into column buffer (0.5 M NaCl, to 0.1 M sodium phosphate, pH 7.0) and applied to a 1 X 60-cm Bio-Gel P-100 (Bio- Rad, 100-200 mesh) column pre-equilibrated and eluted (flow rate, 3 ml/h) with column buffer (Fig. 1, lower panel). The peak fractions, which contained about 75% of the protease nexin loaded onto the column, were concentrated to 1 ml using Aquacide I1 and stored a t -80 "C. For amino acid and amino sugar analysis, reverse-phase high performance liquid chromatography (HPLC) was used on heparin- agarose-purified protease nexin as the final purification and desalting step. A solution containing 0.1-0.5 mg of protease nexin was made 0.1% in trifluoroacetic acid (A buffer) and injected onto a 4.6 x 250- mm Bakerbond wide-pore C, column (J. T. Baker Chemical Co.). Protease nexin eluted as a sharp peak in a gradient of 60% acetonitrile in A buffer.
The amino acid composition was determined by reverse-phase HPLC using o-phthaldialdehyde precolumn derivatization (20). Amino sugar analysis was carried out after 6-h hydrolysis in 4 N HCl by reverse-phase HPLC on a 4.5-cm microsorb C l ~ column (Rainin) using o-phthaldialdehyde precolumn derivatization as above. Sedi- mentation velocity and equilibrium experiments were performed in a Beckman Model E centrifuge equipped with a temperature control unit and absorption optics with an attached electronic scanner. The sedimentation coefficient was determined from the midpoints of integral scanner tracings obtained a t 280 nm. The weight average molecular weight was calculated from the slope of In (c) uersus r2. The concentration profile, in arbitrary units, was obtained from the ultraviolet tracings. The z-average molecular weight was determined from schlieren data obtained at a phase angle of 75", using the slope of the In ( l / r d c l d r ) uersus r 2 plots. Molecular weights were deter- mined at 21,780 and 25,980 rpm. The partial specific volume of the protein was calculated from the amino acid data and values of the partial specific volumes of the amino acid residues (21). All measure- ments were carried out on samples containing from 0.2 to 0.4 mg/ml of protein in 0.5 M NaC1, 0.1 M sodium phosphate, pH 7.0. Amino acid sequence analysis was carried out on 0.65 nmol of protease nexin using an Applied Biosystems 470A Gas Phase Sequencer. Phenyl- thiohydantoins were analyzed on a Hewlett-Packard 1090 HPLC. The amino acid sequence was also determined using a Beckman spinning cup sequenator with on-line phenylthiohydantoin identifi- cation (46). Yields of amino acid residues were not determined in the latter case.
Kinetic Measurements-The overall experimental protocol was essentially identical to that used by Beatty et al. (16) in their study of nl-antitrypsin. Active site titrations were carried out in trypsin and thrombin by the method of Chase and Shaw (22) and on urokinase by the method of Jameson et al. (23). These proteases were then used to titrate protease nexin, which was in turn used to carry out the active site titrations of all the other proteases. In all cases proteases and protease nexin were incubated under second order conditions, usually a t equimolar concentrations. Protease in 20 pl of reaction buffer (0.15 M NaC1, 0.05 M Tris-C1, and 1% bovine serum albumin, pH 8.0) was added to protease nexin in the same buffer (total volume, 0.14 ml). Protease nexin at the low concentrations used in these experiments absorbed to glass or plastic in the absence of carrier
'The abbreviations used are: I, liter; HPLC, high performance liquid chromatography; SDS, sodium dodecyl sulfate.
protein. Reactions were carried out a t 37 "C for a minimum of 30 s and a maximum of 60 min. Chromogenic substrate in 840 pl of reaction buffer was then added in vast excess to simultaneously prevent further reaction between protease nexin and proteases and allow measurement of remaining active protease.
RESULTS AND DISCUSSION
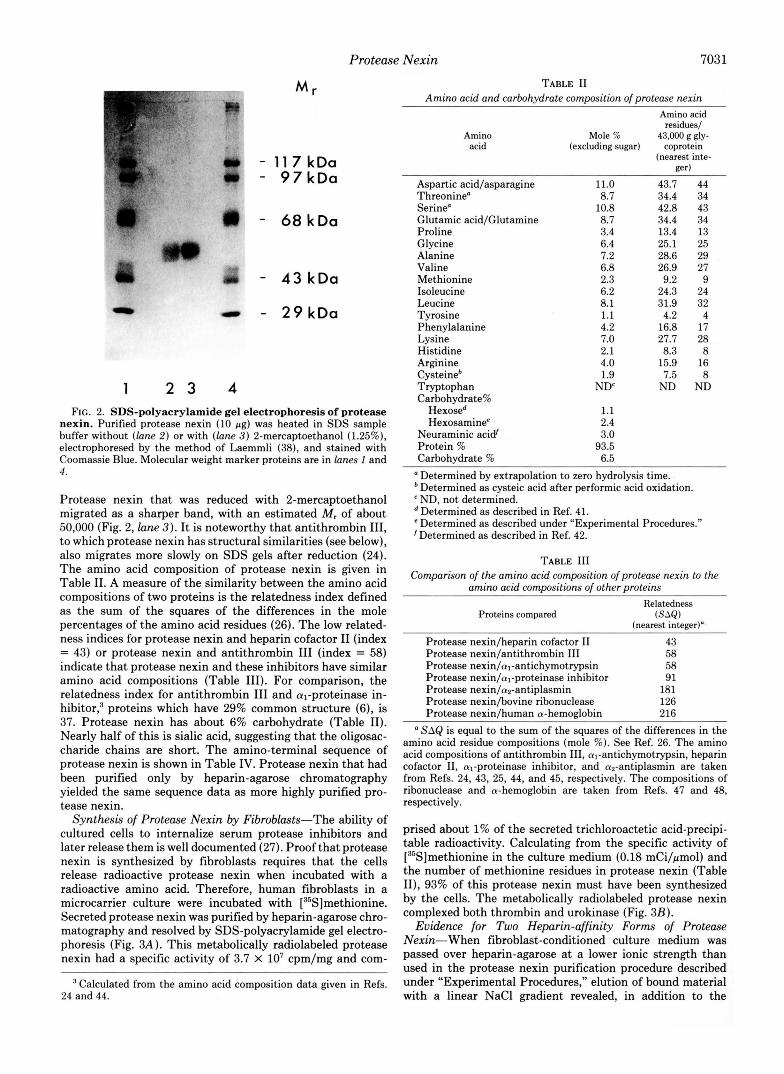
Table I gives the sedimentation and extinction coefficients of protease nexin, along with the M , (42,000-43,000) based on sedimentation equilibrium analysis. The M, of protease nexin estimated from gel exclusion chromatography is 47,000 (Fig. 1, lower panel). On SDS gels, unreduced protease nexin migrated as a diffuse band (Fig. 2, lane 2 ) . The M, estimated from the center of this broad band is approximately 47,000.
TABLE I Physical properties of protease nexin
Molecular weight"
Weight average
r-Average coefficient ( E z l nun) Svedberg Extinction coefficientb
4.3 k 0.1 X 104 4.2 k 0.1 x 10' Srn+ 3.4 s 14.2 "The error values shown indicate the differences in the values
obtained in two runs. In 0.5 M NaCl, 0.1 M sodium phosphate, pH 7.0.
0 0;
0.0:
E c 0
N W
U u
i b"+d
10 20 30 40 Fraction Number
I VoIurne[rn~l
FIG. 1. Purification of protease nexin. Upper panel, culture medium conditioned by human fibroblasts was passed down a hepa- rin-agarose column which was rinsed and eluted using a linear NaCl gradient. Lower panel, two batches of protease nexin purified on heparin-agarose were pooled, concentrated, and loaded onto a Bio- Gel P-100 column that had been calibrated with ferritin (a), trans- ferrin ( b ) , bovine serum albumin ( c ) , carbonic anhydrase (d ) , and cytochrome c (e). Fractions from both columns were assayed for thrombin inhibitory activity using the method of Bart1 and Lill (37). Decreasing absorbance at 405 nm indicates increasing activity. De- tails of the purification are described under "Experimental Proce- dures."
Protease Nexin 7031
- 11 7 kDa - 97kDa
- 68 kDa
- 43 kDa
- 29kDa
1 2 3 4 FIG. 2. SDS-polyacrylamide gel electrophoresis of protease
nexin. Purified protease nexin (10 pg) was heated in SDS sample buffer without (lane 2 ) or with (lane 3) 2-mercaptoethanol (1.25%), electrophoresed by the method of Laemmli (38), and stained with Coomassie Blue. Molecular weight marker proteins are in lunes 1 and 4.
Protease nexin that was reduced with 2-mercaptoethanol migrated as a sharper band, with an estimated M, of about 50,000 (Fig. 2, lane 3). It is noteworthy that antithrombin 111, to which protease nexin has structural similarities (see below), also migrates more slowly on SDS gels after reduction (24). The amino acid composition of protease nexin is given in Table 11. A measure of the similarity between the amino acid compositions of two proteins is the relatedness index defined as the sum of the squares of the differences in the mole percentages of the amino acid residues (26). The low related- ness indices for protease nexin and heparin cofactor I1 (index = 43) or protease nexin and antithrombin I11 (index = 58) indicate that protease nexin and these inhibitors have similar amino acid compositions (Table 111). For comparison, the relatedness index for antithrombin I11 and q-proteinase in- hibitor: proteins which have 29% common structure (6), is 37. Protease nexin has about 6% carbohydrate (Table 11). Nearly half of this is sialic acid, suggesting that the oligosac- charide chains are short. The amino-terminal sequence of protease nexin is shown in Table IV. Protease nexin that had been purified only by heparin-agarose chromatography yielded the same sequence data as more highly purified pro- tease nexin.
Synthesis of Protease Nexin by Fibroblasts-The ability of cultured cells to internalize serum protease inhibitors and later release them is well documented (27). Proof that protease nexin is synthesized by fibroblasts requires that the cells release radioactive protease nexin when incubated with a radioactive amino acid. Therefore, human fibroblasts in a microcarrier culture were incubated with [35S]methionine. Secreted protease nexin was purified by heparin-agarose chro- matography and resolved by SDS-polyacrylamide gel electro- phoresis (Fig. 3A). This metabolically radiolabeled protease nexin had a specific activity of 3.7 x lo7 cpm/mg and com-
Calculated from the amino acid composition data given in Refs. 24 and 44.
TABLE I1 Amino acid and carbohydrate composition of protease nexin
Neuraminic acid‘ 3.0 Protein % 93.5 Carbohydrate % 6.5 Determined by extrapolation to zero hydrolysis time.
*Determined as cysteic acid after performic acid oxidation. ND, not determined. Determined as described in Ref. 41. Determined as described under “Experimental Procedures.”
’Determined as described in Ref. 42.
TABLE I11 Comparison of the amino acid composition of protease nexin to the
amino acid compositions of other proteins Relatedness
a SAQ is equal to the sum of the squares of the differences in the amino acid residue compositions (mole %). See Ref. 26. The amino acid compositions of antithrombin 111, al-antichymotrypsin, heparin cofactor 11, al-proteinase inhibitor, and a*-antiplasmin are taken from Refs. 24, 43, 25, 44, and 45, respectively. The compositions of ribonuclease and a-hemoglobin are taken from Refs. 47 and 48, respectively.
prised about 1% of the secreted trichloroactetic acid-precipi- table radioactivity. Calculating from the specific activity of [35S]methionine in the culture medium (0.18 mCi/pmol) and the number of methionine residues in protease nexin (Table II), 93% of this protease nexin must have been synthesized by the cells. The metabolically radiolabeled protease nexin complexed both thrombin and urokinase (Fig. 3B).
Evidence for Two Heparin-affinity Forms of Protease Nexin-When fibroblast-conditioned culture medium was passed over heparin-agarose at a lower ionic strength than used in the protease nexin purification procedure described under “Experimental Procedures,” elution of bound material with a linear NaCl gradient revealed, in addition to the
7032 Protease Nexin TABLE IV
Amino-terminal sequence of protease nexin Yield is given in picomoles.
Yield 144 29 118 56 91 126 103 124 127 91 86 C y c l e 2 3 24 25 26 27 28
I l e - V a l - L y s - S e r - A r g - P r o Yield 101 105 81 15 60 108
Ser-His-Phe-Asn-Pro-Leu-Ser-Leu-Glu-Glu-Leu-
Gly-Ser-Asn-Thr-Gly-Ile-Gln-Val-Phe-Asn-Gln-
A
1
r
b
2
M r
- 9 7 kDa
- 6 8 kDa i - 4 3 kDa
- 2 9 kDa
B
1 2 3
FIG. 3. Metabolic labeling of protease nexin. Confluent hu- man fibroblasts in a 0.5-1 microcarrier culture were incubated for 3 days with 3.5 mCi of [“S]methionine in serum-free Dulbecco’s mod- ified Eagle’s medium containing methionine a t one-fifth the normal concentration. Medium was harvested after the third day, stored a t -20 “C, and the cells were incubated for another 3 days with medium containing methionine at the normal concentration. This medium was then harvested and pooled with the first harvest medium, and protease nexin was purified on heparin-Sepharose (2). A , samples were submitted to SDS-polyacrylamide gel electrophoresis and either fluorography (39) (gel I ) or staining with Coomassie Blue (gel 2) . B, the [”S]protease nexin (20 pg) was incubated with either 53-kDa urokinase (20 pg) or thrombin (20 pg) in 0.2 ml of phosphate-buffered saline for 0.5 h a t 37 “C. Fifty-pl samples were taken for SDS- polyacrylamide gel electrophoresis. Lane 1, [?S]protease nexin plus urokinase; lane 2, [S5S]protease nexin plus thrombin; and hne 3, [35S] protease nexin.
protease nexin that elutes a t 0.55 M NaCl, another peak of thrombin-inhibitory activity that eluted at 0.40 M NaCl (Fig. 4). As shown, this earlier-eluting peak of thrombin inhibitory activity coincided with a peak of protein that was electropho- retically identical to protease nexin, indicating that it also was protease nexin. The two protease nexin peaks were in- completely separated in most experiments, as shown in Fig. 4, but occasionally were completely separated (data not shown). Previously we showed that protease nexin that is eluted from heparin-agarose with a 1.0 M NaCl step is com- posed of about six charge forms that migrate between pH 7 and 8 on nonequilibrium pH gradient electrophoresis (2).
12 14 20
Fraction Number
FIG. 4. Resolution of two heparin-affinity classes of pro- tease nexin. Medium conditioned by human fibroblasts in a 3-1 microcarrier culture was mixed with 0.3 1 of harvest buffer containing 0.48 M NaCl and passed down a heparin-agarose column. The column was rinsed with 0.17 I of 0.2 M NaCI, 20 mM sodium phosphate, and 0.02% sodium azide (pH 7.0) and then eluted with a linear gradient formed with 0.1 1 of this buffer and 0.1 I of a buffer that was identical except that the NaCl concentration was 1.5 M. Fractions were assayed for thrombin-inhibitory activity (open circles; see Fig. 1 legend) and fraction proteins were resolved by SDS-polyacrylamide gel electro- phoresis and staining with Coomassie Blue. The arrow points to the protease nexin bands.
1 2 FIG. 5. Nonequilibrium pH gradient electrophoresis of two
heparin-affinity classes of protease nexin. Fractions containing protease nexin with low (lane 1 ) and high (lane 2 ) affinities for heparin were obtained from the column shown in Fig. 4 (pooled fractions 14 and 15, and pooled fractions 19 and 20, respectively) and electrophoresed by the method of O’Farrell et al. (40) on a vertical slab gel. The gel was stained with Coomassie Blue. The bottom of the gel is the basic end.
Samples from both of the protease nexin peaks shown in Fig. 4 were separately subjected to nonequilibrium pH gradient electrophoresis (Fig. 5). As shown, most of the same charge forms were present in both heparin affinity peaks, but the protease nexin fraction having the higher affinity for heparin was enriched in the more basic charge forms. This suggests that the affinity of protease nexin for heparin results a t least in part from an ion exchange-type of interaction and that charge heterogeneity may give rise to the two heparin-affinity classes of protease nexin.
The protease nexin purification procedure described under “Experimental Procedures” yields only the high heparin affin- ity protease nexin (Fig. 1). Nevertheless, this procedure is preferred for protease nexin purification because it results in greater purification than if medium is applied to heparin- agarose at reduced NaCl concentration. As seen in Fig. 4, and
Protease Nexin 7033
TABLE V Rates of protease inhibition by protease nexin
Human thrombin (3 nM) + heparin Tosyl-Gly-Pro-Arg-4-nitroanilide acetate (0.08 mM) 1.2 + 0.4 X 10'
Bovine trypsin (50 nM) N-Benzoyl-Ile-Glu-G1y-Arg-p-nitroanilide (0.16 mM) 4.2 & 0.4 X lo6 Human thrombin (5 nM) Tosyl-Gly-Pro-Arg-4-nitroanilide acetate (0.08 mM) 6.0 & 1.3 X lo5 Human urokinase (20 nM) <Glu-Gly-Arg-p-nitroanilide (0.4 mM) 1.5 & 0.1 X 10' Human plasmin (0.5 p M ) Benzoyl-Phe-Val-Arg-p-nitroanilide (0.2 mM) 1.3 & 0.1 X 10' Human tissue plasminogen activator
(12.5 units/ml)
Two chain (0.13 p M ) H-D-Ile-Pro-Arg-p-nitroanilide (1 mM) 3.0 & 0.2 X lok One chain (0.13 p M ) H-D-Ile-Pro-Arg-p-nitroanilide (1 mM) 1.5 & 0.4 X 1030
Human factor Xa (50 nM) N-Benzoyl-Ile-Glu-Gly-Arg-p-nitroanilide (0.16 mM) 7.3 & 0.5 X 10% Mouse y subunit nerve growth fac- H-D-Ile-Pro-Arg-p-nitroanilide (1 mM) 4.0 & 0.4 X lo3
Bovine chymotrypsin (20 nM) Succinyl-Ala-Ala-Pro-Phe-p-nitroanilide (0.2 mM) Not detected Human leukocyte elastase (0.1 pM) Methoxy-Succinyl-Ala-Ala-Pro-Val-p-nitroanilide (0.3 mM) Not detected Human cathepsin G (0.1 p ~ ) Succinyl-Ala-Ala-Pro-Phe-p-nitroanilide (0.2 mM) Not detected
tor (0.125 pM)
a Taken from Ref. 4. Calculated from inhibition after a 30-s incubation (see text).
in all other experiments, the high heparin affinity protease nexin was more abundant than the low heparin affinity pro- tease nexin.
Protease Specificity-Like the other inhibitors in the anti- thrombin I11 a,-proteinase inhibitor group, protease nexin complexes proteases essentially irreversibly. The sensitivity of proteases to protease nexin, therefore, can not be measured by KI. In many cases, formation of nondissociating protease- inhibitor complexes can be kinetically resolved into two steps: initial formation of a loose complex (PI*) that releases the unaltered protease (P) and inhibitor (I) at a finite rate and, secondly, conversion of the loose complex into a stable com- plex (P-I):
kl kl
k f P + I + PI* - P-I (1)
When the kinetics of inhibition of proteases by antithrombin I11 (28), a,-proteinase inhibitor (16), and protease nexin (4) have been examined using protease and inhibitor concentra- tions below the micromolar range, the reactions proceed ac- cording to simple second order kinetics, consistent with the following mechanism.
ka- P + I - P-I (2)
This apparent mechanism is not inconsistent with the two- step mechanism shown in Equation 1, for which, a t least in the case of antithrombin I11 (28, 29), there is strong evidence. The association constants (Equation 2 ) , obtained from reac- tions employing submicromolar reactant concentrations, have been used to compare the specificity of al-proteinase inhibitor for different proteases (16) and are here used for the same purpose with protease n e ~ i n . ~ From Equation 2 ,
d P / d t = -ka- (P)(I). (3)
At equimolar concentrations of reactants, Equation 3 yields
ka..,.t = 1/[Pl - 1/[Pol (4)
where [Po] and [PI represent the initial and final concentra- tions of protease, respectively. For all but one of the reactive
' The order of reactivity of protease nexin with proteases could differ at higher reactant concentrations. Protease nexin is not avail- able in sufficient quantities to carry out such experiments. The concentration of protease nexin in tissues is unknown. In culture we have never found it to exceed nanomolar levels (7) .
proteases tested, factor Xa, plots of 1/V versus t were linear, consistent with Equation 2.
Table V gives the rates of protease nexin-mediated inhibi- tion of 11 tested proteases. Three, cathepsin G, chymotrypsin, and leukocyte elastase, were completely insensitive to pro- tease nexin. These proteases all retained full activity, even in the presence of protease nexin at 100-fold molar excess (data not presented), suggesting that their insensitivity resulted from a failure of native protease nexin to bind them rather than from proteolytic inactivation of protease nexin. Cathep- sin G and chymotrypsin have similar substrate specificities. All of the sensitive proteases have trypsin-like substrate spec- ificities. However, their rates of inhibition varied over 1000- fold range. Protease nexin reacted at a slow but measurable rate with human factor Xa. The second order rate constant shown in Table V was calculated from the initial reaction rate. The plot of l / v versus t deviated downward from line- arity, suggesting that protease nexin and factor Xa formed a loose complex that decayed to a tight complex slowly or perhaps not at all. Thus, the second order rate constant was 7 X lo3 M" s-', if calculated from the 30-s inhibition data, and 2 X IO3 M-' s-', if calculated from the 5-min inhibition data. Protease nexin slowly inhibited the y subunit of mouse nerve growth factor, which supports earlier evidence that protease nexin competes with protease nexin I1 for the binding of nerve growth factor y subunit when this protease is added to fibroblast-conditioned medium (10).
Table V shows that protease nexin is a fast-acting inhibitor of trypsin, thrombin, urokinase, and plasmin. A comparison of the data in Table V to kinetic data in the literature suggests that protease nexin inhibits trypsin about as rapidly as does al-proteinase inhibitor (16) and inhibits thrombin signifi- cantly more rapidly than does antithrombin I11 (28, 29). Heparin accelerated the protease nexin-mediated inhibition of thrombin by more than lOO-fold, to a rate so rapid that it was difficult to measure and was probably near the diffusion- controlled limit.
Although protease nexin has thus far been studied and assayed only in cell culture systems, several comments about its possible physiological functions are worth making. The secretion of protease nexin by various connective tissue cells (7) suggests that protease nexin functions as an extravascular inhibitor. The high concentrations of al-proteinase inhibitor, antithrombin 111, ap-macroglobulin, and az-antiplasmin in plasma insure that protease nexin is not a significant plasma
7034 Protease Nexin
inhibitor of thrombin and plasmin and suggest, furthermore, that protease nexin may not be a significant inhibitor of these proteases even in interstitial fluid (although this will remain uncertain until the concentrations of protease nexin and plasma inhibitors in interstitial fluid are determined). Pro- tease nexin might be a significant source of anti-urokinase activity in connective tissue because, first, the present data indicate that it is one of the fastest acting urokinase inhibitors known and, second, plasma may provide little urokinase in- hibitory activity. The defined plasma protease inhibitors are very poor urokinase inhibitors (30-32). In a recent study Waller et al. (33) found that urokinase at 20 nM (1 pg/ml) retained over 80% of its activity when it was incubated in plasma for 2 h. The present finding that protease nexin inhibits trypsin more rapidly than it inhibits urokinase is not evidence against the possibility that urokinase is a major target of protease nexin. It is noteworthy that antithrombin 111, which is the major plasma inhibitor of thrombin, inhibits trypsin and several other proteases far more rapidly than it inhibits thrombin (29).
A number of urokinase-inhibitory proteins that are distinct from protease nexin have been discovered during the last several years (e.g. see Refs. 34-36). Data on their rates of association with urokinase are not yet available.
14. Low, D. A., Scott, R. W., Baker, J. B., and Cunningham, D. D.
15. Eaton, D. L., and Baker, J. B. (1983) J. Cell Biol. 97, 323-328 16. Beatty, K., Bieth, J., and Travis, J. (1980) J. Biol. Chem. 255,
17. Wiman, B., and Collen, D. (1978) Eur. J. Biochem. 8 4 , 573-578 18. Deutsch, D. G., and Mertz, E. T. (1970) Science (Wash. D. C.)
19. Dane, K., and Reich, E. (1979) Biochim. Biophys. Acta 566,138-
20. Jones, B. N., and Gilligan, J. P. (1983) J. Chromatogr. 266,471-
21. Zamyatnin, A. A. (1984) Ann. Rev. Biophys. Bioeng. 13,145-165 22. Chase, T., and Shaw, E. (1969) Biochemistry 8, 2212-2223 23. Jameson, G. W., Roberts, D. V., Adams, R. W., Kyle, W. S. A,,
and Elmore, D. T. (1973) Biochem. J . 131, 107-117 24. Kurachi, K., Schmer, G., Hermodson, M. A,, Teller, D. C., and
Davie, E. W. (1976) Biochemistry 15 , 368-373 25. Tollefsen, D. M., Majerus, D. W., and Blank, M. K. (1982) J.
Biol. Chem. 257,2162-2169 26. Marchalonis, J. J., and Weltman, J. K. (1971) Comp. Biochem.
Physiol. 38,609-625 27. Rohrlich, S. T., and Rifkin, D. B. (1981) J. Cell. Physiol. 109,l-
15 28. Olson, S. T., and Shore, J. D. (1982) J. Biol. Chem. 257, 14891-
14895 29. Feinman, R. D., Chang, T.-L., Wilson, S. M., and Li, E. H. H.
(1977) in Chemistry and Biology of Thrombin (Lundblad, R. L., Fenton. J. W.. 11, and Mann. K. G., eds) DD. 217-232. Ann
(1982) Nature 298, 476-478
3931-3934
170,1095-1096
151
482
centrifugations, and Dan Eaton for help with the sequencing. Acta 429.591-599
REFERENCES
1. Baker, J. B.. Low, D. A., Simmer, R. L. and Cunningham, D. D. (1980) cell 2 1 , 3 7 4 5
-
2. Scott, R. W., and Baker, J. B. (1983) J. Biol. Chem. 258,10439- 10444
Biol. Chem. 258,4397-4403 3. Scott, R. W., Eaton, D. L., Duran, N., and Baker, J. B. (1983) J.
4. Eaton, D. L., Scott, R. W., and Baker, J. B. (1984) J. Biol. Chem.
5. Owen, W. G. (1975) Biochem. Biophys. Acta 405 , 380-387 6. Carrell, R. W., Jeppsson, J.-O., Laurell, C.-B., Brennan, S. O.,
Owen, M. C., Vaughan, L., and Boswell, D. R. (1982) Nature
7. Eaton, D. L., and Baker, J. B. (1983) J. Cell. Physiol. 117 , 175- 182
8. Low, D. A., Baker, J. B., Koonce, W. C., and Cunningham, D. D. (1981) Proc. Natl. Acud. Sci. U. S. A. 78, 2340-2344
9. Knauer, D. J., and Cunningham, D. D. (1982) Proc. Natl. Acud. Sci. U. S. A. 7 9 , 2310-2314
10. Knauer, D. J., Scaparro, K. M., and Cunningham, D. D. (1982) J. Biol. Chem. 257,15098-15104
11. Knauer, D. J., Thompson, J. A., and Cunningham, D. D. (1983) J. Cell. Physiol. 1 1 7, 385-396
12. Baker, J. B., Knauer, D. J., and Cunningham, D. D. (1985) in The Receptors (Conn, P. M., ed) Vol. 3, Academic Press, New York, in press
13. Baker, J . B., Low, D. A., Eaton, D. L., and Cunningham, D. D. (1982) J. Cell. Physiol. 112 , 291-297
259,6241-6247
298,329-334
32. Vahtera, E.,’and Hamberg, U. (1978) Biochem. J . 171, 767-770 33. Waller, K. E., Schleuning, W.-D., and Reich, E. (1983) Biochem.
34. Loskutoff, D. J., van Mourik, J. A., Erickson, L. A., and Lawrence, D. (1983) Proc. Natl. Acud. Sci. U. 5‘. A. 80,2956-2960
35. Wiman, B., Chmielewska, J., and Rinby, M. (1984) J. Biol. Chem.
36. Holmberg, L., Lecander, I., Persson, B., and Astedt, B. (1978) Biochem. Biophys. Acta 544, 128-137
37. Bartl, K., and Lill, H. (1980) Thromb. Res. 18, 267-272 38. Laemmli, U. K. (1970) Nature 227 , 680-685 39. Bonner, W. M., and Laskey, R. A. (1974) Eur. J. Biochem. 46,
40. O’Farrell. D. Z.. Goodman. H. M.. and O’Farrell. P. H. (1977)
J. 2 1 5 , 123-131
259,3644-3647
83-88
Cell 12,113311141 . .
41. Dubois. M.. Gilles. K. A,. Hamilton. J. K.. Rebers. P. A.. and Smith, F.’(1956)Anal. Chem. 28, 350-356
42. Hammond, K. S., and Papermaster, D. S. (1976) Anal. Biochem. 74,292-297
43. Travis, J., Garner, D., and Bowen, J. (1978) Biochemistry 17,
44. Pannell, R., Johnson, D., and Travis, J . (1974) Biochemistry 13,
45. Wiman, B., and Collen, D. (1977) Eur. J. Biochem. 78 , 19-26 46. Rodrieuez. H.. Kohr. W. J.. and Harkins. R. N. (1984) Anal.
5647-5651
5439-5445.
Bio&ei. 140,538-547 ’
47. Smvth. D. G.. Stein. W. H.. and Moore. S. (1963) J. Biol. Chem. 238; 227-234 ‘
. . .
48. Hill, R. J., and Konigsberg, W. (1962) J. Biol. Chem. 237, 3151- 3156