Protection against Mycobacterium tuberculosis challenge in mice by DNA vaccine Ag85A-ESAT-6-IL-21 priming and BCG boosting J. Dou 1, *, Y. Wang 1, *, F. Yu 1, †, H. Yang‡, J. Wang§, X. He†, W. Xu‡, J. Chen† & K. Hu† Summary Tuberculosis (TB) is one of most important chronic infectious diseases caused by Mycobacterium tubercu- losis and remains a major global health problem. In the study, we developed the DNA vaccine encoding fusion protein of antigen 85 A and 6 kDa early secretory antigen target of M. tuberculosis as well as the cytokine IL-21 to investigate its immune protective efficacy against M. tuberculosis challenge in mice after the DNA vaccine priming and Bacille Calmette-Gue ´rin (BCG) boosting. Compared with the different control groups, the intranasal DNA vaccine priming twice and BCG boosting once markedly increased the cytotoxic- ities of natural killer cells and splenocytes and enhanced the interferon-c level in the splenocyte supernatant as well as sIgA level in bronchoalveolar lavage in the vaccinated mice. Importantly, this heterologous prime– boost strategy significantly decreased the bacterial load in the mouse lungs in contrast to that of intranasal or subcutaneous BCG immunization alone. These findings provide further approaches for mucosal-targeted prime– boost vaccination to fight against TB. Introduction Mycobacterium tuberculosis has a penetrance of its host population. About one-third of the human popu- lation would have a positive skin test for the infection; thus, it is thought to harbour the M. tuberculosis (Russell et al., 2010). The latest estimate of the global tuberculosis (TB) mortality among human immuno- deficiency virus (HIV)-negative people fell from 30 to 20 per 100 000 population (36%) between 1990 and 2009 and could be halved by 2015. The overall decline (when including HIV-positive people, who comprise 12% of all TB cases) was 19%. Neverthe- less, a rapid spread of multi-drug resistant bacteria causing about 0.5 million TB cases per year has wors- ened the problem during recent years. Twenty-two ‘high-burden’ countries account for more than 80% of the active TB cases in the worldwide (Svenson et al., 2010; Glaziou et al., 2011). The conventional anti-TB vaccine, Mycobacterium bovis bacille Calmette-Gue ´rin (BCG), has still widely used to prevent TB and is given to millions of infants worldwide as unique vaccine since 1921. However, the protective efficacy of BCG has shown variable effects against adult pulmonary TB in different trials even though it continues to be used to prevent menin- gitis and phthisis miliaris in children in many coun- tries. In addition, the patients with HIV may be induced to be disseminated infection with M. tubercu- losis owing to BCG vaccination (Espinal et al., 2001; Kaufmann & McMichael, 2005; Claire et al., 2007). The current drug therapies are fraught with problems, predominantly because of the emergence of multi-drug resistant M. tuberculosis strains and the increase in patients with TB who are co-infected with HIV (Dou et al., 2010; Russell et al., 2010). There is no effec- tive vaccine against M. tuberculosis infection in view of the weakness of BCG; therefore, the development for a new TB vaccine candidate may consider it. Studies have revealed that the TB DNA vaccine is a very powerful and easy method for the induction of a strong immune responses in mice (Huygen, 2006; Dou et al., 2008); however, the clinical results of using the DNA vaccine have been disappointing with regard to the magnitude of the induced immune responses, and there is no one DNA vaccine candidate against TB to replace the current ‘gold standard’ vaccine BCG (von Reyn & Vuola, 2002; Huygen, 2006; Radosevic et al., 2007). Therefore, understanding of protective immu- nity against M. tuberculosis infection is very impor- tant for the current TB vaccine development. IL-21 plays a role in a wide range of relevant biological activities, including the activation and proliferation of * Department of Pathogenic Biology and Immunology, Medical School, Southeast University, Nanjing, 210009 China, † Medical School, Southeast University, Nanjing, China, ‡ Centers for Disease Control and Prevention of Jiangsu Province and § Zhongda Hospital, Medical School, Southeast University, Nanjing, China Received 31 August 2011; revised 18 October 2011; accepted 3 November 2011 Correspondence: Professor Jun Dou, Department of Pathogenic biology and Immunology, Medical School, Southeast University, # 87 Ding Jiaqiao Rd. Nanjing 210009, China. Tel: +86 25 83272454; Fax: +86 25 83272295; E-mail: [email protected]1 Co-author, equally contributed. ª 2011 Blackwell Publishing Ltd International Journal of Immunogenetics, 2012, 39, 183–190 183 doi: 10.1111/j.1744-313X.2011.01066.x
Transcript
doi: 10.1111/j.1744-313X.2011.01066.x
Protection against Mycobacterium tuberculosis challenge in mice by
DNA vaccine Ag85A-ESAT-6-IL-21 priming and BCG boosting
J. Dou1,*, Y. Wang1,*, F. Yu1,†, H. Yang‡, J. Wang§, X. He†, W. Xu‡, J. Chen† & K. Hu†
Summary
Tuberculosis (TB) is one of most important chronicinfectious diseases caused by Mycobacterium tubercu-losis and remains a major global health problem. In thestudy, we developed the DNA vaccine encoding fusionprotein of antigen 85 A and 6 kDa early secretoryantigen target of M. tuberculosis as well as the cytokineIL-21 to investigate its immune protective efficacyagainst M. tuberculosis challenge in mice after theDNA vaccine priming and Bacille Calmette-Guerin(BCG) boosting. Compared with the different controlgroups, the intranasal DNA vaccine priming twice andBCG boosting once markedly increased the cytotoxic-ities of natural killer cells and splenocytes and enhancedthe interferon-c level in the splenocyte supernatant aswell as sIgA level in bronchoalveolar lavage in thevaccinated mice. Importantly, this heterologous prime–boost strategy significantly decreased the bacterial loadin the mouse lungs in contrast to that of intranasal orsubcutaneous BCG immunization alone. These findingsprovide further approaches for mucosal-targeted prime–boost vaccination to fight against TB.
Introduction
Mycobacterium tuberculosis has a penetrance of itshost population. About one-third of the human popu-lation would have a positive skin test for the infection;thus, it is thought to harbour the M. tuberculosis(Russell et al., 2010). The latest estimate of the globaltuberculosis (TB) mortality among human immuno-
* Department of Pathogenic Biology and Immunology, Medical
School, Southeast University, Nanjing, 210009 China, † Medical
School, Southeast University, Nanjing, China, ‡ Centers for Disease
Control and Prevention of Jiangsu Province and § Zhongda Hospital,
Medical School, Southeast University, Nanjing, China
Received 31 August 2011; revised 18 October 2011; accepted 3
November 2011
Correspondence: Professor Jun Dou, Department of Pathogenic
biology and Immunology, Medical School, Southeast University, # 87
International Journal of Immunogenetics, 2012, 39, 183–190
deficiency virus (HIV)-negative people fell from 30 to20 per 100 000 population (36%) between 1990 and2009 and could be halved by 2015. The overalldecline (when including HIV-positive people, whocomprise 12% of all TB cases) was 19%. Neverthe-less, a rapid spread of multi-drug resistant bacteriacausing about 0.5 million TB cases per year has wors-ened the problem during recent years. Twenty-two‘high-burden’ countries account for more than 80% ofthe active TB cases in the worldwide (Svenson et al.,2010; Glaziou et al., 2011).
The conventional anti-TB vaccine, Mycobacteriumbovis bacille Calmette-Guerin (BCG), has still widelyused to prevent TB and is given to millions of infantsworldwide as unique vaccine since 1921. However,the protective efficacy of BCG has shown variableeffects against adult pulmonary TB in different trialseven though it continues to be used to prevent menin-gitis and phthisis miliaris in children in many coun-tries. In addition, the patients with HIV may beinduced to be disseminated infection with M. tubercu-losis owing to BCG vaccination (Espinal et al., 2001;Kaufmann & McMichael, 2005; Claire et al., 2007).The current drug therapies are fraught with problems,predominantly because of the emergence of multi-drugresistant M. tuberculosis strains and the increase inpatients with TB who are co-infected with HIV (Douet al., 2010; Russell et al., 2010). There is no effec-tive vaccine against M. tuberculosis infection in viewof the weakness of BCG; therefore, the developmentfor a new TB vaccine candidate may consider it.
Studies have revealed that the TB DNA vaccine is avery powerful and easy method for the induction of astrong immune responses in mice (Huygen, 2006; Douet al., 2008); however, the clinical results of using theDNA vaccine have been disappointing with regard tothe magnitude of the induced immune responses, andthere is no one DNA vaccine candidate against TB toreplace the current ‘gold standard’ vaccine BCG (vonReyn & Vuola, 2002; Huygen, 2006; Radosevic et al.,2007). Therefore, understanding of protective immu-nity against M. tuberculosis infection is very impor-tant for the current TB vaccine development. IL-21plays a role in a wide range of relevant biologicalactivities, including the activation and proliferation of
183
184 J. Dou et al.
CD8 T cells and natural killer (NK) cells, and servesas a costimulator for enhancing effector responses inthe control of a chronic viral infection including hepa-titis C virus and human immunodeficiency virus infec-tions (Yi et al., 2009) as well as the tumour vaccineapproaches. IL-21 is also as an immune molecularadjuvant for the development of DNA vaccine con-taining antigen 85 A (Ag85A) in mouse model (Douet al., 2008; Kim-Schulze et al., 2009). In our previ-ous studies, DNA vaccine expressing fusion protein ofAg85A and 6 kDa early secretory antigen target(ESAT-6) of M. tuberculosis as well as mouse IL-21could elicit mice to generate effective immuneresponses that included the activities of cytotoxic Tlymphocytes (CTL) and NK cells, the splenocyte pro-liferative responses to purified protein derivative aswell as the levels of antibody and serum cytokine (Jin-long et al., 2009). However, the protective efficacyagainst M. tuberculosis challenge in mice remains tobe confirmed.
Because TB is primarily a respiratory airway infec-tious disease, vaccination at the mucosal site has beenbelieved to be superior to vaccination at other sites foreliciting protective immune responses againstM. tuberculosis infection (Davis, 2001; Jian et al.,2009). Because current promising vaccines against TBare based on heterologous prime–boost combination,in the present study, we adopted the strategy of anintranasal (i.n.) DNA vaccine Ag85A-ESAT-6-IL-21priming and i.n. BCG boosting to investigate the pro-tective efficacy against M. tuberculosis challenge inmouse model. Our data demonstrated that the DNAvaccine priming–BCG boosting strategy significantlyreduced M. tuberculosis load in lungs compared withthat of the i.n. or subcutaneous (s.c.) BCG vaccina-tion. Therefore, these findings support the strategy ofi.n. DNA vaccine priming twice and i.n. BCG boostingonce under development for potential human applica-tions.
Material and methods
Mice, cell lines and DNA vaccine construct
Male C57BL ⁄ 6 mice between 4 and 6 weeks of agewere ordered from the Animal Center of Yang ZhouUniversity of China. The mice were raised at theExperimental Animal Center, Medical School, South-east University, Nanjing, China. All animal experi-ments were conducted following the guidelines of theAnimal Research Ethics Board of Southeast University.B16F10 murine melanoma cell line is syngeneic inC57BL ⁄ 6 mice and was a gift from Professor Pingsh-eng Chen, Southeast University Medical School.YAC-1 cell line (Moloney leukaemia-induced T-celllymphoma of A ⁄ Sn mouse origin) was purchased fromCellular Institute of China in Shanghai, and all cellsare cultured at 37�C in 5% CO2 atmosphere in RPMI1640 supplemented with 10% foetal bovine serum
containing 100 units mL)1 penicillin G sodium &100 lg mL)1 streptomycin sulphate. DNA vaccineconstruct Ag85A-ESAT-6-IL-21 was constructed byour laboratory (Dou et al., 2008; Jinlong et al.,2009).
Transfection and Western blot
The B16F10 cells grown to 70% confluent were trans-fected with the DNA vaccine construct pIRES-Ag85A-ESAT-6 or the pIRES-Ag85A-ESAT-6-IL-21 and themock plasmid as control using Lipofectamine� 2000reagent (Invitrogen, CA, USA), followed by selectionwith 800 lg mL)1 of G418 (Clontech, Palo Alto, CA,USA). G418-resistant B16F10 clones were screened forthe expression of different target proteins by Westernblot. The goat anti-Ag85A and goat anti-mouse IL-21antibody (I-18; Santa Cruz Biotechnology Company,CA, USA) were used in Western blot. The protocolwas based on the published reports (Jun et al., 2005,2006a,b).
Mycobacterium tuberculosis and vaccine BCG
H37Rv was used as the virulent M. tuberculosis strainfor challenge studies. The M. tuberculosis cultureswere grown at 37�C in suspension in Sauton mediumenriched with 0.5% sodium pyruvate and 0.5% glu-cose. Mycobacterium bovis BCG (Connaught Strain;products ID: 2009050321) was a gift from the Centersfor Disease Control and Prevention in Jiangsu Provinceof China.
Mouse immunization and Mycobacterium tuberculosis
challenge
C57BL ⁄ 6 mice were randomly divided into sevengroups, (1) the saline group (i.n. vaccinated with100 lL saline thrice), (2) the mock plasmid group(i.n.vaccinated with 100 lg mock plasmid thrice), (3) themock plasmid + BCG group (i.n. vaccinated with100 lg mock plasmid twice + i.n. 1 · 106 colonyforming unit (CFU) of BCG once), (4) the pIRES-Ag85A-ESAT-6 group + BCG group (i.n. vaccinatedwith 100 lg DNA vaccine pIRES-Ag85A-ESAT-6 twi-ce + i.n. 1 · 106 CFU of BCG once), (5) the pIRES-Ag85A-ESAT-6-IL-21 group (i.n. vaccinated with100 lg DNA vaccine pIRES-Ag85A-ESAT-6-IL-21twice + i.n. 1 · 106 CFU of BCG once), (6) the i.n.BCG group and (7) the s.c. BCG group (i.n. or s.c.vaccinated with 1 · 106 CFU of BCG thrice). Eachimmunization was at 2-week interval. Eighteen micewere used in groups 3–6. Two weeks after the finalimmunization, six mice were killed for detectingimmune efficiency and the remnant mice were used forM. tuberculosis challenge experiments. Twelve micewere only used in group 1, group 2 and group 7 forM. tuberculosis challenge experiments. BCG was pre-pared and s.c. or i.n. immunization was based on the
ª 2011 Blackwell Publishing Ltd
International Journal of Immunogenetics, 2012, 39, 183–190
Ag85A-ESAT-6-IL-21 vaccine against TB 185
published report (Wang et al., 2004). Each mouse waschallenged by the tail vein with 1 · 104 CFU of viru-lent M. tuberculosis H37Rv strain, diluted in 200 lL.Thirty days after challenge, six mice were killed fordetecting the bacterial burdens in the mouse lungs.After 60 days, six mice were weighed for mouse weightand then survival was continuously observed for 90days (Jeon et al., 2008; Sadagopal et al., 2009).
Cytotoxic assays
The cytotoxic activity of NK and splenocytes wasdetected using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide colorimetry (MTT)method (Mosmann, 1983; Duo et al., 2008). Briefly,for NK activity assay, 5 · 104 YAC-1 cells (targetcells) were seeded with 2.5 · 106 splenocytes (effectorcells) in 96-well plates at 50:1 ratios of effector to tar-get. For splenocyte cytotoxicities, 2.5 · 106 mL)1
splenocytes as effector cells were incubated for 24 hwith 5 · 104 B16F10 cells transfected with the pIRES-Ag85A-ESAT-6 or mock plasmid as target cells at50:1 ratios of effector cells to target cells in total vol-ume of 200 lL. After the plate was incubated at 37�Cin 5% CO2 for 20 h and supernatant was discarded,each well was added with 200 lL MTT for additional4 h incubation. Supernatants were discarded, and theplates were measured with the plate reader (Bio-Rad,Hercules, CA, USA) at optical densities (OD)490 after20 lL dimethylsulfoxide was added. All cytotoxicactivity assays were performed in triplicate.
Cytokine measurement
IFN-c lever in the splenocyte supernatants was mea-sured using a commercially available double antibodysandwich enzyme-linked immunosorbent assay(ELISA) kit according to the manufacturer’s protocol(eBioscience company, eBioscience, Vienna, Austria).Briefly, 2 · 106 splenocytes were obtained from themice immunized with the different vaccines describedabove and cultured in 24-well plates with mitomycinC inactivated 1 · 104 B16F10 cells transfected withthe pIRES-Ag85A-ESAT-6. After 2 days, the superna-tant was collected and the dilutions of the test sampleswere always tested (in triplicate) starting at a dilutionof 1:2. Samples and standards were run in triplicate,and the sensitivity of the assays was £5 pg mL)1
(Banerjee et al., 2004; Jun et al., 2007).
sIgA measurement in BAL
The secretory IgA (sIgA) level in bronchoalveolarlavage (BAL) was detected (Wang et al., 1999; Bastianet al., 2000; Davis et al., 2001). Briefly, lungs wereremoved along with the heart and a portion of the tra-chea. A segment of polyethylene tube was attached to a23-gauge needle with syringe. The tube was insertedinto the trachea portion, and lungs were lavaged thrice
ª 2011 Blackwell Publishing Ltd
International Journal of Immunogenetics, 2012, 39, 183–190
with 200 lL PBS and approximately 0.6 mL of BALfluid was acquired. BAL supernatant was removed andstored at )20�C for sIgA measurement with ELISAaccording to the manufacturer’s protocol (eBioscience).Both Ag85A and ESAT-6 were used as coated antigens.
Mycobacterial colony enumeration assay and
histopathology
Thirty days after M. tuberculosis challenge, lungswere removed and the right lung was placed in4.0 mL of PBS with 0.05% Tween-80 and homoge-nized with a tissue homogenizer until no tissue chunksremained. Two hundered microlitre serially dilutedhomogenates was plated onto Middlebrook 7H10 agarplates containing 10% oleic acid-albumin-dextrose-catalase enrichment and 0.5% glycerol. Plates weresemisealed in a plastic bag to prevent overdrying andincubated at 37�C for 18–24 days. Colonies were enu-merated with a dissection microscope (Davis et al.,2001). The left lung was fixed in 10% formalin andthen embedded in paraffin. Serial 5 lm tissue sectionswere cut and mounted on SuperFrost Plus glass slides(Brain Research Laboratories, Waban, Newton, MA,USA), fixed in methanol and stained in haematoxylinand eosin. The number of foci of caseation and necro-sis, and the number of calcified lesions were eachsemiquantitatively scored as absent, minimal, slight,moderate, strong, noted as 0, 1, 2, 3, 4 and 5, respec-tively. In this score, the frequency as well as the sever-ity of the lesions was incorporated. Granulomaformation was scored by estimating the occupied areaof the lung section (Dormans et al., 2004).
Statistical analysis
Statistical analysis was performed using the Student’st-test for the difference between the experimentalgroup and control group. Bonferroni correction wasused where multiple comparisons were made. P-values<0.05 were taken as statistically significant.
Results
Identification of target antigen expressions of DNA
vaccines in the B16F10 cells
We constructed the DNA vaccine pIRES-Ag85A-ESAT-6-IL-21 and successfully identified it by theanalyses of an endonuclease digestion and DNAsequence (data not shown). Subsequently, we devel-oped the B16F10 cells that were transfected with thepIRES-Ag85A-ESAT-6-IL-21 or the pIRES-Ag85A-ESAT-6 or the mock plasmid and further identified theexpressions of Ag85A-ESAT-6 fusion protein and IL-21 by Western blot. Figure 1 indicates that the targetantigen expressions of Ag85A-ESAT-6 and IL-21 inthe transfected B16F10 cells were confirmed by theanalysis of protein expressions. The results suggested
1 2 3
Ag85A-ESAT-6 (38 KD)
3
Ag85A (32 KD)
IL-21 (17 KD)
GAPDH (36 KD)
Figure 1. Target antigen expressions of Ag85A-ESAT-6 and IL-21
detected by Western blot. Lanes 1–3 represent the protein expres-
sions of Ag85A-ESAT-6 and IL-21 in the B16F10 cells transfected
with the mock plasmid or DNA vaccine pIRES-Ag85A-ESAT-6 or
pIRES-Ag85A-ESAT-6-IL-21 in order. The results suggested that the
Ag85A-ESAT-6 and IL-21 were correctly expressed in the stable
transfected B16F10 cells.
186 J. Dou et al.
that the DNA vaccine construct for pIRES-Ag85A-ESAT-6-IL-21 was developed appropriately for furtherinvestigating its immune protection from M. tubercu-losis infection in the mouse model.
Cytotoxicities of NK cells and splenocytes
There were four independent experiments for detectingcytotoxicities of NK cells and splenocytes in vitro asshown in Fig. 2(a,b). In Fig. 2(a), the NK cytotoxicactivity in pIRES-Ag85A-ESAT-6-IL-21 group washighest (72.37 ± 5.11%) among four groups and BCGi.n. vaccination group ranked second (58.90 ± 4.78%),followed by pIRES-Ag85A-ESAT-6 group (51.52 ±4.96%). The lowest NK activity was the mock plasmidgroup (33.92 ± 3.09%). The difference in NK activitywas a statistically significant between the first twogroups (P < 0.05), between the pIRES-Ag85A-ESAT-6-IL-21 group and the pIRES-Ag85A-ESAT-6 group(P < 0.05), and between the pIRES-Ag85A-ESAT-6-IL-21 group and the mock plasmid group (P < 0.03). The
**
607080 *
*
2030405060
1 2 3 40
1020
Cyt
otox
icity
of N
K c
ells
(%)
Groups
(a) (
Figure 2. Cytotoxic activity of natural killer (NK) cells and splenocytes det
group vaccinated thrice, pILES-Ag85A-ESAT-6 priming twice and Bacille Ca
ing twice and BCG boosting once, and Bacille Calmette-Guerin vaccinated
splenocyte cytotoxicity. All cytotoxic assays were performed in triplicate. A
(P = 0.014–0.028); *P < 0.05 (P = 0.032–0.043).
splenocyte cytotoxicity was markedly increased inpIRES-Ag85A-EAT-6-IL-21 group versus the mockplasmid group, whose difference was statistically signif-icant (P < 0.03). The splenocyte cytotoxicities in theBCG group and the pIRES-Ag85A-ESAT-6 group werealso increased, and the differences were statistically sig-nificant compared with that of the mock plasmid group(P < 0.05), respectively, as shown in Fig. 2(b).
ELISA for IFN-c in splenocyte supernatant
In the current study, we also detected type 1 cytokineIFN-c owing to its important function in host defenceagainst M. tuberculosis infection. The results showedthat the IFN-c level in splenocyte supernatant in themice immunized with the pIRES-Ag85A-ESAT-6-IL-21group was significantly increased compared with thelevel in the pIRES-Ag85A-ESAT-6 group (P < 0.05) orin the mock plasmid group (P < 0.03), and that theIFN-c level in mice immunized with the BCG groupwas also significantly increased compared to the miceimmunized with the pIRES-Ag85A-ESAT-6 group(P < 0.05) or the mock plasmid group (P < 0.05), asshown in Fig. 3. However, the increased IFN-c levelin the pIRES-Ag85A-ESAT-6-IL-21 group was not sta-tistically significant compared with that of BCGgroup.
sIgA in BAL detected by ELISA
Figure 4 gives that a sIgA levels in BAL were remark-ably elevated (1287.13 ± 115.26) in mice immunizedwith the pIRES-Ag85A-ESAT-6-IL-21 group (P < 0.05) orthe pIRES-Ag85A-ESAT-6 group (1201.35 ± 102.14,P < 0.05) as well as the BCG group (1164.08 ± 97.12,P < 0.05) in contrast to the level in the mice immu-nized with the mock plasmid (622.13 ± 121.74). ThesIgA levels, however, were no statistically significantdifference between the pIRES-Ag85A-ESAT-6-IL-21group and the BCG group (P > 0.05). A representative
**
708090
*
*
30405060
1 2 3 40
1020
Cyt
otox
icity
of s
plen
ocyt
es (%
)
Groups
b)
ected by the MTT method. Groups 1–4 represent the mock plasmid
set of data for six mice is shown in Fig. 4. Theresults (Figs 3 & 4) indicate that the DNA vaccinepIRES-Ag85A-ESAT-6-IL-21 elicits a high level ofIFN-c and a specific sIgA response to fusion protein ofAg85A and ESAT-6 in the immunized mice.
Immune protective efficacy Against Mycobacterium
Tuberculosis challenge
In order to acquire a robust immunogenicity of vaccineand effective protection from M. tuberculosis challengein mice, we used the strategy of the mucosal-targeted
ª 2011 Blackwell Publishing Ltd
International Journal of Immunogenetics, 2012, 39, 183–190
prime–boost vaccination approaches in the study. Micewere vaccinated with the different vaccines with the dif-ferent schedules that are shown in Fig. 5. We foundthat a significant protective efficacy against TB wasachieved in the mice using the various prime–booststrategies compared with that of saline i.n. group andthe mock plasmid group. There were significant differ-ences of CFU in the lung and the mouse weight betweentwo groups after immunized, infected mice as shown inFig. 5(a,b). For examples, the i.n. pAEI · 2 + i.n.BCG · 1 immunized, infected mice presented a reduc-tion of 2.8, log10 in the lung, a significantly greaterdegree in relation to the mock plasmid · 2 + i.n.BCG · 1 immunized, infected group. The other immu-nized, infected mice also presented a significant reduc-tion, as follows: the i.n. BCG · 3 group (1.73, log10),the s.c. BCG · 3 group (1.5, log10), the i.n.pAE · 2 + i.n. BCG · 1 group (1.15, log10). The Fig. 5suggested that the i.n. DNA vaccine pIRES-Ag85A-ESAT-6-IL-21 priming and i.n. BCG boosting strategywere efficient preventive strategies against TB and had astrong ability to reduce the bacterial burdens in themouse lungs after i.v. challenge of M. tuberculosis andlengthened the mouse survival were also (data notshown).
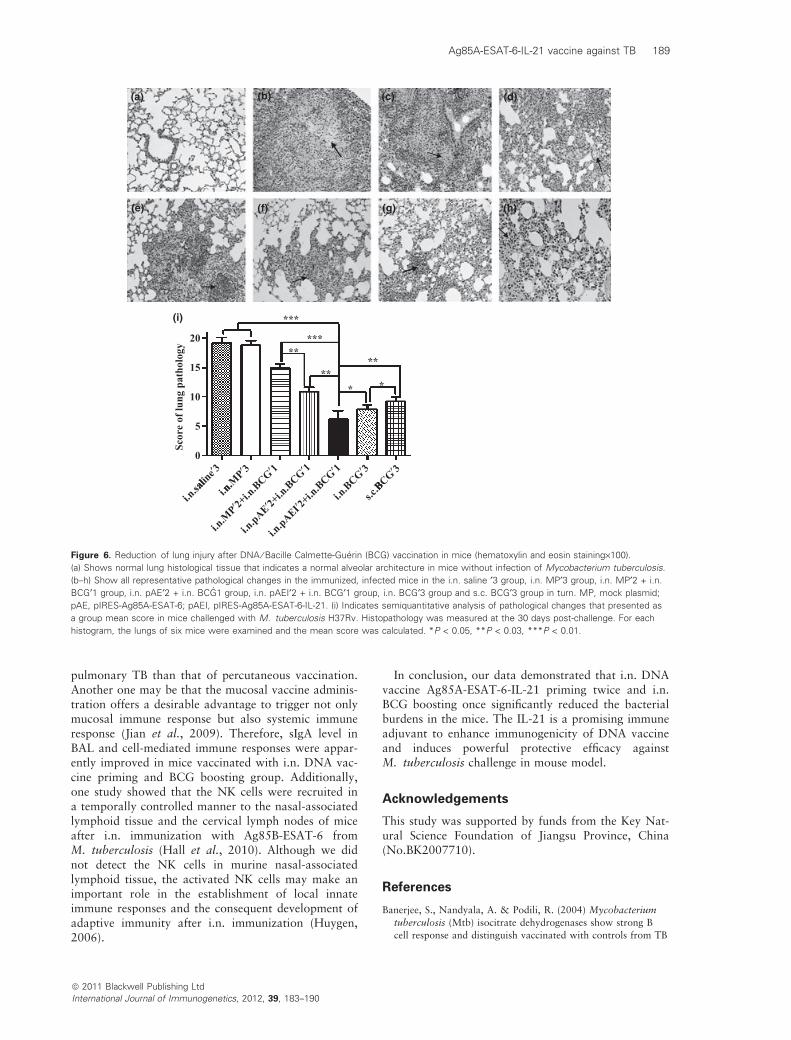
The lung injury was effectively lessened in miceimmunized with DNA vaccine pIRES-Ag85A-ESAT-6-IL-21 priming twice and BCG boosting once as shownin the Fig. 6(f), in where no obvious inflammationresponses were observed and the well-contained smallgranulomas (the arrows) exhibited generally infiltratednumerous lymphocytes and neutrophils. The patholog-ical changes of i.n. BCG · 3 group and s.c. BCG · 3group were shown in Fig. 6(g,h). Consistent with thehistological data, the pathological change of Fig. 6(f)is similar to the pathological changes of Fig. 6(g,h).Figure 6(d) shows a slight inflammation responses,which were generally infiltrated by numerous lympho-cytes and some neutrophils and a little caseationnecrotic (arrows) with moderate lymphocytic infiltra-tion in mice immunized with the mock plasmid twiceand BCG once group. The pathological change wasmore serious in mice immunized with saline threetimes (Fig. 6b) or the mock plasmid three times(Fig. 6c) than that of Fig. 6(d). Much more infiltratedinflammation cells, lymphocytes and TB lesions thatwere more likely to be necrotic (arrows) were found inFig. 6(b,c). Figure 6(i) showed that the semiquantita-tive analysis of histopathological changes as a groupmean score in lung challenged with M. tuberculosisH37Rv. We find that the score of pathology injury isthe lowest in DNA vaccine pIRES-Ag85A-ESAT-6-IL-21 priming twice–BCG boosting once group amongseven groups as shown in the Fig. 6(i).
Discussion
Intranasal or bronchial epithelial cells are known toplay an integral role in the airway defence mechanism
8 08.5
****
***
24.525.0 pre-infect.
Post-infect.***
******
6.06.57.07.58.0
***
CFU
(n p
er lu
ng)
22.022.523.023.524.0 *
*
Wei
ght (
g)
alinex
3
n.MPx3
BCGx1
BCGx1
BCGx1
BCGx3
BCGx34.55.05.56.0
Log 1
0
alinex
3M
Px3
BCGx1
BCGx1
BCGx1
BCGx3
BCGx320.020.521.021.5
i.n.sa
li.n
.
i.n.M
Px2+i.n
.BC
i.n.pAEx2
+i.n.BC
i.n.pAEIx2
+i.n.BC
i.n.B
s.c.B
i.n.sa
li.n
.M
i.n.M
Px2+i.n
.BC
i.n.pAEx2
+i.n.BC
i.n.pAEIx2
+i.n.BC
i.n.BC
s.c.BC
(a) (b)
Figure 5. DNA vaccine Prime-BCG boost strategy induced protective efficacy against Mycobacterium tuberculosis challenge. (a) Shows the
CFU in the mouse lung lobe 60 days after immunization ⁄ 30 days after challenge. (b) Indicates that the comparison of weight in pre-infection
and 30 days post- infection in the mice. The marks (*, ** and ***) show statistically significant difference between two groups as judged by
Student’s t-test and an ANOVA followed by multiple comparison tests were made. *P < 0.05; P < 0.03; ***P < 0.01. CFU, colony-forming unit;
21; BCG, Bacille Calmette-Guerin; ·1, ·2 and ·3: immunization once, immunization twice and immunization three times.
188 J. Dou et al.
and may make a crucial role in mucosal immunity.Thus, mucosal vaccination has received increasing atten-tion owing to its potency in inducing mucosa-associatedprotection from mucosal infectious diseases, such aspulmonary TB infection (Takizawa, 2005; Jian et al.,2009). Intranasal application of the BCG was highlyprotective efficacy against challenge infection withM. tuberculosis. The heterologous prime–boost immu-nization strategy can evoke powerful immuneresponses and may be of value in developing animproved TB vaccine (Davis, 2001; Jian et al., 2009).Dietrich et al. (2006) examined the i.n. administrationof a vaccine consisting of the mucosal adjuvant heat-labile enterotoxin mutant (LTK63) and the subunitfusion protein Ag85B-ESAT-6 against M. tuberculosis,and data showed that the vaccination withLTK63 ⁄ Ag85B-ESAT-6 gave a strong and sustainedTh1 response, which led to long-lasting protectionagainst TB. Based on the study and other reports (Hallet al., 2010; Jian et al., 2009), we investigated theprotective immunity elicited by the vaccine alone or incombination with BCG boosting strategy in all of theexperiment animals in order to improve the immuno-genicity of DNA vaccine and to enhance protectiveefficacy against M. tuberculosis infection. Our dataindicated that, compared with the i.n. BCG vaccina-tion alone, the i.n. DNA vaccine priming twice andi.n. BCG boosting once markedly increased the cyto-toxicities of NK cells and splenocytes as well as IFN-clevel, whose immune response was also stronger thanour previous results, especially in the IFN-c level (Jin-long et al., 2009). More importantly, the DNA prim-ing–BCG boosting strategy significantly reduced thepulmonary bacterial burdens and effectively lessenedthe pulmonary injury in mice after challenge of
M. tuberculosis, which was reflected in decreasing thenumber of CFU in lung, increasing mouse weight(Fig. 5), extending mouse survival until 90 days with-out death. In contrast, four of the six mice in thesaline group, three of the six mice in the mock plasmidgroup, three of the six mice in the mock plas-mid + BCG group and four of the six mice in thepIRES-Ag85A-ESAT-6 group + BCG group, two of thesix mice in the s.c. BCG group were all dead, but sixmice life were also survival in the i.n. BCG group. Inaddition, the effect of the DNA priming plus BCGboosting vaccination strategy resulted in a reducedtotal histopathology score, particularly in the pIRES-Ag85A-ESAT-6-IL-21 group (Fig. 6i). The resultsdemonstrated that the strategy of i.n. DNA vaccinepriming twice and i.n. BCG boosting once was betterthan single i.n. or s.c. BCG vaccination three times.
In the study, we used the IL-21 as an immune adju-vant and aimed to augment the immunogenicity ofDNA vaccine expressing fusion protein of Ag85A-ESAT-6 and to increase the protection againstM. tuberculosis infection in mice. Figures 2 & 3 showthat the cytotoxicity of NK cells and the IFN-c levelwere markedly increased in the pIRES-Ag85A-ESAT-6-IL-21 group compared with those of other groups.One of the mechanisms, we propose, is that the IL-21stimulated the production of IFN-c in the immunizedmice, and IFN-c and IL-21 enhanced the Ag85A-ESAT-6 presentation to lymphocytes by antigen-pre-senting cells that elicited T lymphocyte responses toAg85A-ESAT-6 and generated more IFN-c by NK cells(Dietrich et al., 2006; Dou et al., 2008). Based on thedecreased bacterial burdens in the mouse lungs, wefound that the mucosal vaccination directed to therespiratory system provided a better protection from
ª 2011 Blackwell Publishing Ltd
International Journal of Immunogenetics, 2012, 39, 183–190
(a) (b) (c) (d)
(e)
(i)
(f) (g) (h)
aline′3
n.MP′3
BCG′3
BCG′3
i.n.sa
li.n
.
i.n.M
P′2+i.n
.BCG′1
i.n.pAE′2+
i.n.BCG′1
i.n.pAEI′2
+i.n.BCG′1
i.n.
s.c.B
20
***
*****
****
* *
15
10
0
5
Scor
e of
lung
pat
holo
gy
Figure 6. Reduction of lung injury after DNA ⁄ Bacille Calmette-Guerin (BCG) vaccination in mice (hematoxylin and eosin staining·100).
(a) Shows normal lung histological tissue that indicates a normal alveolar architecture in mice without infection of Mycobacterium tuberculosis.
(b–h) Show all representative pathological changes in the immunized, infected mice in the i.n. saline ¢3 group, i.n. MP¢3 group, i.n. MP¢2 + i.n.
BCG¢1 group, i.n. pAE¢2 + i.n. BCG1 group, i.n. pAEI¢2 + i.n. BCG¢1 group, i.n. BCG¢3 group and s.c. BCG¢3 group in turn. MP, mock plasmid;
pAE, pIRES-Ag85A-ESAT-6; pAEI, pIRES-Ag85A-ESAT-6-IL-21. (i) Indicates semiquantitative analysis of pathological changes that presented as
a group mean score in mice challenged with M. tuberculosis H37Rv. Histopathology was measured at the 30 days post-challenge. For each
histogram, the lungs of six mice were examined and the mean score was calculated. *P < 0.05, **P < 0.03, ***P < 0.01.
Ag85A-ESAT-6-IL-21 vaccine against TB 189
pulmonary TB than that of percutaneous vaccination.Another one may be that the mucosal vaccine adminis-tration offers a desirable advantage to trigger not onlymucosal immune response but also systemic immuneresponse (Jian et al., 2009). Therefore, sIgA level inBAL and cell-mediated immune responses were appar-ently improved in mice vaccinated with i.n. DNA vac-cine priming and BCG boosting group. Additionally,one study showed that the NK cells were recruited ina temporally controlled manner to the nasal-associatedlymphoid tissue and the cervical lymph nodes of miceafter i.n. immunization with Ag85B-ESAT-6 fromM. tuberculosis (Hall et al., 2010). Although we didnot detect the NK cells in murine nasal-associatedlymphoid tissue, the activated NK cells may make animportant role in the establishment of local innateimmune responses and the consequent development ofadaptive immunity after i.n. immunization (Huygen,2006).
ª 2011 Blackwell Publishing Ltd
International Journal of Immunogenetics, 2012, 39, 183–190
In conclusion, our data demonstrated that i.n. DNAvaccine Ag85A-ESAT-6-IL-21 priming twice and i.n.BCG boosting once significantly reduced the bacterialburdens in the mice. The IL-21 is a promising immuneadjuvant to enhance immunogenicity of DNA vaccineand induces powerful protective efficacy againstM. tuberculosis challenge in mouse model.
Acknowledgements
This study was supported by funds from the Key Nat-ural Science Foundation of Jiangsu Province, China(No.BK2007710).
References
Banerjee, S., Nandyala, A. & Podili, R. (2004) Mycobacterium
tuberculosis (Mtb) isocitrate dehydrogenases show strong B
cell response and distinguish vaccinated with controls from TB
190 J. Dou et al.
patients. Proceedings of the National Academy of Sciences of
the United States of America, 101, 12652.
Bastian, A., Tunkel, C., Lins, M. Bottcher, H., Hirt, S.W.,
Cremer, J. & Bewig, B. (2000) Immunoglobulin A and secre-
tory immunoglobulin A in the bronchoalveolar lavage from
patients after lung transplantation. Clinical Transplantation, 6,
580.
Claire, S.A., Jes, D. & Else, M.A. (2007) The combined CTA1-
DD ⁄ ISCOMs vector is an effective intranasal adjuvant for
boosting prior Mycobacterium bovis BCG immunity to Myco-
bacterium tuberculosis. Infection and Immunity, 75, 408.
Davis, S.S. (2001) Nasal vaccines. Advanced Drug Delivery
Reviews, 51, 21.
Dietrich, J., Andersen, C., Rappuoli, R., Doherty, T.M., Jensen,
C.G. & Andersen, P. (2006) Mucosal administration of
Ag85B-ESAT-6 protects against infection with Mycobacterium
tuberculosis and boosts prior Bacillus Calmette-Gue¢rin immu-
nity. Journal of Immunology, 177, 6353.
Dormans, J., Burger, M. & Aguilar, D. (2004) Correlation of
virulence, lung pathology, bacterial load and delayed type
hypersensitivity responses after infection with different
Mycobacterium tuberculosis genotypes in a BALB ⁄ c mouse
model. Clinical and Experimental Immunology, 137, 468.