Protection of BALB/c Mice against Brucella abortus 544Challenge by Vaccination with Bacterioferritin or
P39 Recombinant Proteins with CpGOligodeoxynucleotides as Adjuvant
AYMAN AL-MARIRI,1 ANNE TIBOR,1 PASCAL MERTENS,1 XAVIER DE BOLLE,1
PATRICK MICHEL,2 JACQUES GODEFROID,2 KARL WALRAVENS,2
AND JEAN-JACQUES LETESSON1*
Unite de Recherche en Biologie Moleculaire, Laboratoire d’Immunologie et de Microbiologie,Facultes Universitaires Notre-Dame de la Paix, B-5000 Namur,1 and Centre d’Etude
et de Recherche Veterinaire et Agrochimique, B-1180 Brussels,2 Belgium
Received 28 February 2001/Returned for modification 11 April 2001/Accepted 7 May 2001
The P39 and the bacterioferrin (BFR) antigens of Brucella melitensis 16M were previously identified as Tdominant antigens able to induce both delayed-type hypersensivity in sensitized guinea pigs and in vitrogamma interferon (IFN-g) production by peripheral blood mononuclear cells from infected cattle. Here, weanalyzed the potential for these antigens to function as a subunitary vaccine against Brucella abortus infectionin BALB/c mice, and we characterized the humoral and cellular immune responses induced. Mice were injectedwith each of the recombinant proteins alone or adjuvanted with either CpG oligodeoxynucleotides (CpG ODN)or non-CpG ODN. Mice immunized with the recombinant antigens with CpG ODN were the only groupdemonstrating both significant IFN-g production and T-cell proliferation in response to either Brucella extractor to the respective antigen. The same conclusion holds true for the antibody response, which was onlydemonstrated in mice immunized with recombinant antigens mixed with CpG ODN. The antibody titers (bothimmunoglobulin G1 [IgG1] and IgG2a) induced by P39 immunization were higher than the titers induced byBFR (only IgG2a). Using a B. abortus 544 challenge, the level of protection was analyzed and compared to theprotection conferred by one immunization with the vaccine strain B19. Immunization with P39 and CpG ODNgave a level of protection comparable to the one conferred by B19 at 4 weeks postchallenge, and the mice werestill significantly protected at 8 weeks postchallenge, although to a lesser extent than the B19-vaccinated group.Intriguingly, no protection was detected after BFR vaccination. All other groups did not demonstrate anyprotection.
Brucella species are facultative intracellular gram-negativebacterial pathogens that infect both phagocytic and nonphago-cytic cells (42). Brucella abortus causes abortion and infertilityin cattle and also various chronic zoonotic infections in humans(8, 42). The intracellular localization of these bacteria impliesthat the immunity against Brucella requires a cell-mediatedimmune response, which makes the Th1 arm of the responsevery crucial for controlling the infection (44).
Brucella abortus strain B19 is one of the most commonlyused attenuated live vaccines against bovine brucellosis andinduces high level of protection in cattle (15). The presence ofsmooth lipopolysaccharide in the vaccine strain B19 may in-terfere with the discrimination between infected and vacci-nated individuals (32) and impair the test and slaughter strat-egy. Moreover, this strain can cause abortion whenadministered to pregnant cattle (9) and is still fully virulent forhumans (42). In order to avoid these drawbacks, alternativevaccination approaches are needed. Among these, subcellularvaccines able to induce protective Th1 cell-mediated immune
response are being developed. Recombinant antigens of Bru-cella spp. such as HtrA (40), GroEL (2, 30, 34), GroES (34),Cu,Zn superoxide dismutase (SOD) (47, 49), YajC (52), UvrA(34), and L7 and L12 (37) have been shown to induce humoraland cell-mediated immune responses in mice, but only L7/L12(36) and peptides comprising certain epitopes of Cu,Zn SOD(38, 47, 54) induced some level of protection in a mouse modelof infection. While the protection afforded could be improvedusing a multiple subunit vaccine, it remains also possible that amore powerful antigen or a better adjuvant or both may lead toprotection with a monovalent subunit vaccine.
Our laboratory has previously described bacterioferritin(BFR) (13) and P39 (a putative periplasmic binding protein)(10, 11) as T-cell immunodominant Brucella antigens (12),eliciting both a strong delayed-type hypersensitivity (DTH) inguinea pigs sensitized with brucellin and in vitro proliferationor gamma interferon (IFN-g) production by peripheral bloodmononuclear cells (PBMC) from infected cattle. The potentialof these antigens to induce a Th1-oriented immune responsemakes them attractive candidates as a subunitary vaccineagainst brucellosis.
While the type of antigen and microorganism administeredto a host (as well as the dose and route of immunization) areimportant factors that influence the type of immune response,
* Corresponding author. Mailing address. Unite de Recherche enBiologie Moleculaire (URBM), Laboratoire d’Immunologie et de Mi-crobiologie, Facultes Universitaires Notre-Dame de la Paix, Rue deBruxelles 61, B-5000 Namur, Belgium. Phone: 32-81-72-44-02. Fax:32-81-72-42-97. E-mail: [email protected].
it is also well established that the presence of certain cytokinesat the site of injection is crucial for orienting the emergingT-cell response (21, 33, 50). For this late concern, adjuvantsare of paramount importance. It has been shown that a syn-thetic phosphorothioate oligodeoxynucleotide containing anunmethylated, consensus immunostimulatory CpG motif (59-purine-purine-CpG-pyrimidine-pyrimidine-39oligodeoxynucleotide [CpG ODN]) can act as an adjuvantwhich favors cell-mediated immune mechanisms (19, 20, 25,26) with a Th1-like cytokine profile (7, 24). This suggests thatCpG ODN could act as an adjuvant for the clearance of intra-cellular pathogens (14). We decided to take this newly de-scribed adjuvant and test its potentiating effect with the Bru-cella T-cell antigens we have previously described.
In this study, we evaluated the potential of P39 and BFRwith CpG ODN as adjuvant in inducing a Th1 response and theefficiency of these vaccines to protect BALB/c mice against aninfectious B. abortus 544 challenge.
MATERIALS AND METHODS
Bacteria. The B. abortus virulent strain 544 was obtained from J.-M. Verger(Institut National de la Recherche Agronomique, Pathologie Infecteuse et Im-munologie, Nouzilly, France), and B. abortus vaccine strain B19 was obtainedfrom J. Goodfroid. They were grown in 2YT medium (10 g of yeast extract, 10g of tryptone, and 5 g of NaCl per liter) for 3 days at 37°C as described earlier(16). Escherichia coli BL21(DE3) was grown on Luria-Bertani medium thatcontained 100 mg of ampicillin per ml. This strain was used for production of therecombinant BFR protein using the pET-15b-bfr vector.
Oligodeoxynucleotides. Phosphorothioate-modified ODNs were synthesized atEurogentec. The ODNs used in these studies were the immunostimulatory CpG1826 (59-TCCATGACGTTCCTGACGTT-39) and non-CpG 1745 (59-TCCAATGAGCTTCCTGAGTCT-39), which are nonstimulatory and used as a control.(CpG motifs or reversed non-CpG motifs are underlined.) CpG 1826 has beenwell characterized for adjuvant activity with protein antigen (7).
Purification of BFR and P39 proteins. The bfr gene of Brucella melitensis 16Mwas subcloned into a pET-15b expression vector (Novagen, Madison, Wis.), andthe resulting plasmid pET-15b-bfr was introduced in E. coli BL21(DE3). After 2to 4 of induction with 1 mM IPTG (isopropyl-b-D-thiogalactopyranoside) (Pro-mega, Madison, Wis.), bacterial cells from a 100-ml culture were washed onceand then sonicated (seven times for 25 s each time on ice). The lysate wascentrifuged for 10 min at 9,000 3 g at 4°C. The pellet was kept frozen at 270°C.After it had thawed, the pellet was resuspended in lysis buffer (10 mM Tris-HCl[pH 8], 5 mM EDTA) containing 100 mg of lysozyme (Sigma, St. Louis, Mo.) perml. The resulting lysate was centrifuged at 9,000 3 g for 20 min at 4°C. Theinsoluble fraction of the His6-tagged BFR protein was solubilized in 50 ml of 6M guanidine-HCl (pH 6.5) buffer containing 0.05% Triton X-100, and the extractwas centrifuged at 9,000 3 g for 30 min at 4°C. The supernatant was purified onan Ni-affinity chromatography column (Pharmacia Biotech). The protein waseluted from the column in elution buffer (1 M imidazole, 0.5 M NaCl, 20 mMTris-HCl [pH 7.0]). The purified protein was divided into aliquots and stored at270°C until use. The purification of P39 was as described previously (28).
Expression and purification of the recombinant BFR was monitored by sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), followed byCoomassie brilliant blue staining or Western blot analyses.
SDS-PAGE and immunoblotting. SDS-PAGE and immunoblotting were per-formed as previously described (48).
Immunization of chicken. Sonicated E. coli BL21(DE3) lysate (150 mg ofproteins) was injected into the breast muscle for the induction of E. coli-specificantibodies in the egg yolk. Inoculations were repeated 3 and 6 weeks later. Theeggs were collected 7 days after the last injection. The egg yolk was diluted inH2O (10 times the yolk weight) and then frozen at 220°C to precipitate thelipids. The sample was thawed and centrifuged at 2,500 3 g at 4°C for 45 min.The supernatant was filtered on a 0.45-mm (pore-size) filter and mixed withammonium sulfate (25% saturation, final concentration). After a 20-min incu-bation at room temperature, the sample was centrifuged at 2,500 3 g at 4°C for30 min. The supernatant was mixed with ammonium sulfate (40% saturation,final concentration) and processed as before. Finally, the pellet was resuspendedin 1 ml of phosphate-buffered saline (PBS)-azide (0.1%).
Immunization of mice. Female BALB/c mice were obtained from IFFa Credo,Brussels, Belgium, at 4 weeks of age. Mice were separated into nine groups of 12mice. Groups 1, 2, and 3 received PBS, CpG ODN, and non-CpG ODN, respec-tively, and served as negative controls. Groups 4 and 5 were injected with thepurified P39 and BFR alone, respectively. Groups 6 and 7 were injected with therecombinant protein with CpG ODN adjuvant. Finally, groups 8 and 9 receivedthe recombinant antigens with the non-CpG ODN. Vaccines were prepared inPBS and contained combinations of the following: 20 mg of recombinant proteinand/or 20 mg of oligonucleotides when needed. Vaccines were given intramus-cularly (i.m.) into the left tibial anterior muscles in a total volume of 50 ml threetimes at 3-week intervals. Three weeks after the last injection, four mice ran-domly selected in each group were sacrificed by cervical dislocation. Their serawere harvested to determine the humoral immune response. Their spleens wereremoved aseptically to investigate the cellular immune response.
Isotype-specific immunoglobulin ELISA assays. Specific murine IgG1 andIgG2a isotypes were assayed by enzyme-linked immunosorbent assay (ELISA)using microplates (Nunc, Roskilde, Denmark) coated overnight at 4°C with anoptimal concentration of the different antigens in 50 ml of PBS–0.1% Thimerosal(Sigma). Plates were washed twice with PBS and blocked with PBS–2.5% caseinfor 2 h at room temperature (RT). After three washes in PBS, 100 ml of serialtwofold dilutions starting at 1/100 in dilution buffer (PBS–0.05% Tween 20–1.25% casein) were performed and then loaded in microwells and incubated atRT for 1 h. The sera from nonimmunized mice were used as negative controls.After five washes with washing buffer (PBS–0.05% Tween 20), biotinylated goatanti-mouse IgG1 or anti-mouse IgG2a antibodies (Amersham) were added at anoptimal dilution for 1 h at RT. Following five additional washes with PBS-Tween,the plates were incubated for 1 h with 50 ml of a 1:1,000 dilution of streptavidin-horseradish peroxidase (Amersham) at RT. Finally, the plates were washed fivetimes and developed for 10 min in the dark with TMB (3,39,5,59-tetramethylben-zidine) at 40 mg/ml in pH 4.0 citrate buffer containing 1.7 ml of hydrogenperoxide (KPL, Gaithersburg, Md.) per liter. The reaction was then stopped bythe addition of 2 N H2SO4 to each well. The absorbance was determined at 450nm (Bio Kinetics Reader EL-340). Titers were defined as the highest dilution ofmouse serum which gave an optical reading of three times the reading of thenegative control.
Lymphocyte proliferation assays. Spleens were homogenized with 2 ml oftissue culture medium (RPMI 1640–5% fetal bovine serum; Gibco-BRL), anderythrocytes were lysed with Gey’s solution. Splenocytes at 2 3 105 per well werestimulated with concanavalin A (ConA; 3 mg/ml), recombinant protein antigen(10 mg/ml), bacteria lysate (30 mg/ml), or no additive in culture medium for atotal volume of 0.2 ml per well. Cell proliferation was determined in triplicate,based on the uptake of [3H]thymidine ([methyl-3H]thymidine; CNA). After 2days of incubation at 37°C in 5% CO2, the plates were pulsed with [3H]thymidineat 0.5 mCi per well and processed, 18 h later with a cell harvester (Skatron, Inc.,Sterling, Va.) onto glass filter strips (Skatron, Inc.). Tritiated-thymidine incor-poration was counted by use of liquid scintillation spectroscopy with a Betaplatecounter (WALLAC Oy, Turku, Finland). The mean number of counts perminute and the standard error of the mean for each triplicate of cells weredetermined.
In vitro assay for cytokine production by spleen cells. Levels of IFN-g andinterleukin-5 (IL-5) in murine splenocyte culture supernatants were measuredafter 96 h of incubation with antigen or mitogen as described for the lymphocyteproliferation assay. IFN-g and IL-5 were assayed by specific ELISA kits (Pharm-ingen, San Diego, Calif.). Samples were tested in duplicate. The concentrationsof IFN-g or IL-5 were calculated from a standard curve for recombinant mouseIFN-g or IL-5 (Pharmingen) generated in each assay. Values of less than 40 and10 pg/ml were considered negative for IFN-g and IL-5, respectively.
Protection assay. Three weeks after the last injection, the remaining mice ofeach group were challenged by the intraperitoneal route (i.p.) with 5 3 104 CFUof B. abortus strain 544 in 100 ml of PBS. An additional group of eight micevaccinated i.p. with B19 (105 CFU) was challenged 4 weeks later in the same wayand served as a vaccinated control. Spleen colonization with the challenge strainwas determined at 4 and 8 weeks postinfection. Spleens were homogenized in 2ml of 0.1% Triton-PBS, and 100 ml of 10-fold serial dilutions were plated intriplicate onto 2YT agar for 3 to 4 days at 37°C with 10% CO2, and the CFU werecounted. The limit of detection of spleen counts is 20 CFU/spleen. For theB19-vaccinated group, dilutions were spread on 2YT agar alone or on 2YT plus0.1% erythritol for differentiation of B. abortus B19 strain from strain 544 (43).
Statistical analysis. Significances of differences were determined by use of theStudent’s t test or Fisher exact test as appropriate. A P value of ,0.05 wasconsidered significant.
VOL. 69, 2001 PROTECTION AGAINST BRUCELLA ABORTUS 4817
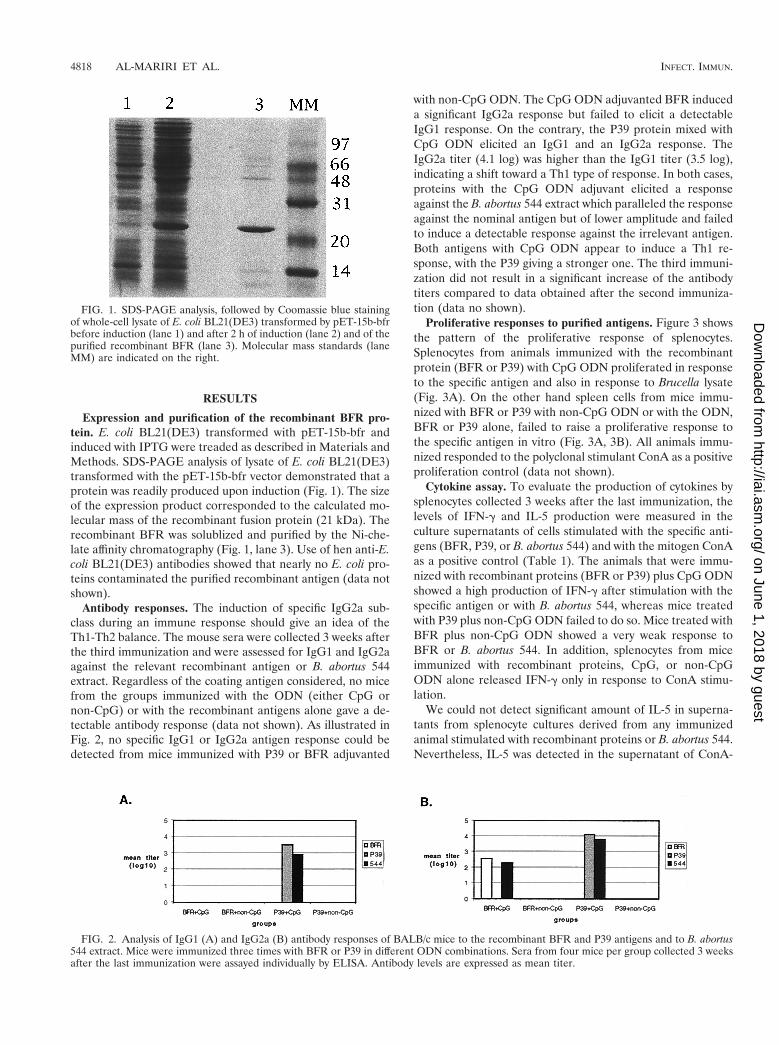
Expression and purification of the recombinant BFR pro-tein. E. coli BL21(DE3) transformed with pET-15b-bfr andinduced with IPTG were treaded as described in Materials andMethods. SDS-PAGE analysis of lysate of E. coli BL21(DE3)transformed with the pET-15b-bfr vector demonstrated that aprotein was readily produced upon induction (Fig. 1). The sizeof the expression product corresponded to the calculated mo-lecular mass of the recombinant fusion protein (21 kDa). Therecombinant BFR was solublized and purified by the Ni-che-late affinity chromatography (Fig. 1, lane 3). Use of hen anti-E.coli BL21(DE3) antibodies showed that nearly no E. coli pro-teins contaminated the purified recombinant antigen (data notshown).
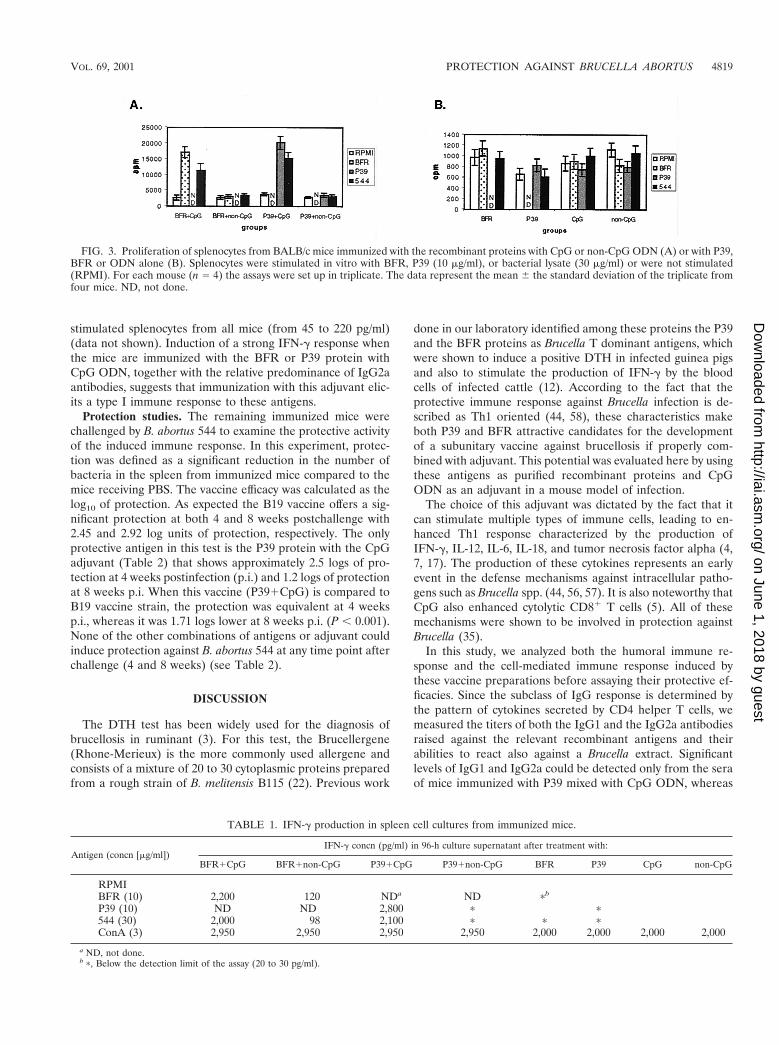
Antibody responses. The induction of specific IgG2a sub-class during an immune response should give an idea of theTh1-Th2 balance. The mouse sera were collected 3 weeks afterthe third immunization and were assessed for IgG1 and IgG2aagainst the relevant recombinant antigen or B. abortus 544extract. Regardless of the coating antigen considered, no micefrom the groups immunized with the ODN (either CpG ornon-CpG) or with the recombinant antigens alone gave a de-tectable antibody response (data not shown). As illustrated inFig. 2, no specific IgG1 or IgG2a antigen response could bedetected from mice immunized with P39 or BFR adjuvanted
with non-CpG ODN. The CpG ODN adjuvanted BFR induceda significant IgG2a response but failed to elicit a detectableIgG1 response. On the contrary, the P39 protein mixed withCpG ODN elicited an IgG1 and an IgG2a response. TheIgG2a titer (4.1 log) was higher than the IgG1 titer (3.5 log),indicating a shift toward a Th1 type of response. In both cases,proteins with the CpG ODN adjuvant elicited a responseagainst the B. abortus 544 extract which paralleled the responseagainst the nominal antigen but of lower amplitude and failedto induce a detectable response against the irrelevant antigen.Both antigens with CpG ODN appear to induce a Th1 re-sponse, with the P39 giving a stronger one. The third immuni-zation did not result in a significant increase of the antibodytiters compared to data obtained after the second immuniza-tion (data no shown).
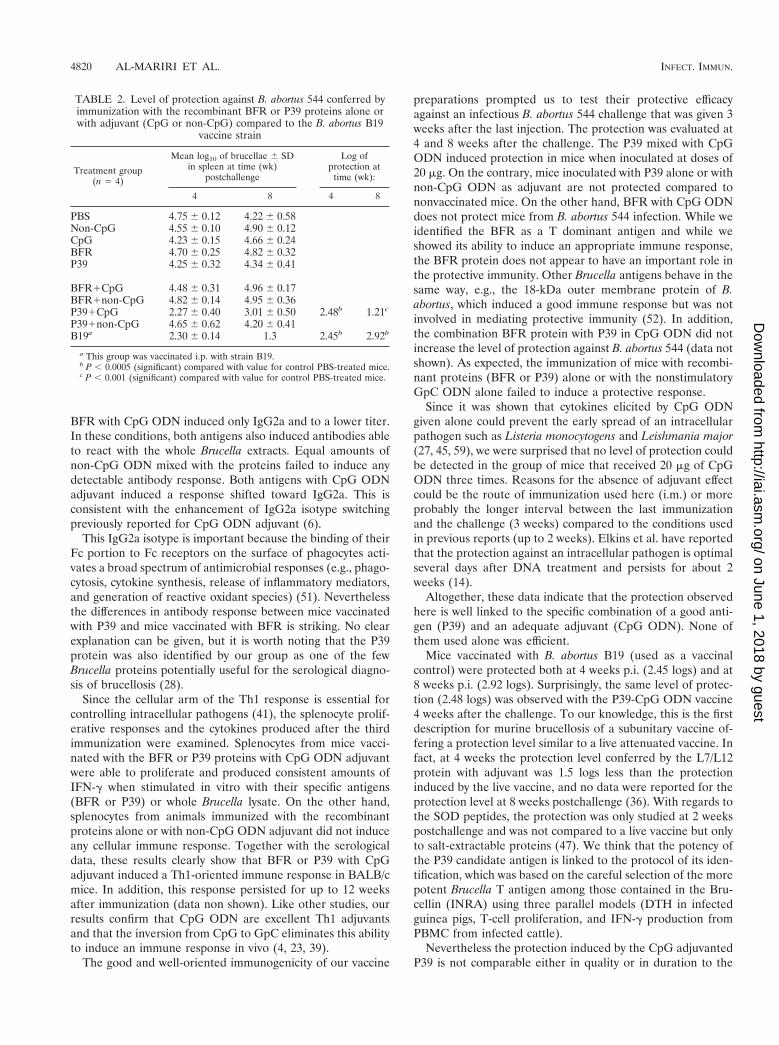
Proliferative responses to purified antigens. Figure 3 showsthe pattern of the proliferative response of splenocytes.Splenocytes from animals immunized with the recombinantprotein (BFR or P39) with CpG ODN proliferated in responseto the specific antigen and also in response to Brucella lysate(Fig. 3A). On the other hand spleen cells from mice immu-nized with BFR or P39 with non-CpG ODN or with the ODN,BFR or P39 alone, failed to raise a proliferative response tothe specific antigen in vitro (Fig. 3A, 3B). All animals immu-nized responded to the polyclonal stimulant ConA as a positiveproliferation control (data not shown).
Cytokine assay. To evaluate the production of cytokines bysplenocytes collected 3 weeks after the last immunization, thelevels of IFN-g and IL-5 production were measured in theculture supernatants of cells stimulated with the specific anti-gens (BFR, P39, or B. abortus 544) and with the mitogen ConAas a positive control (Table 1). The animals that were immu-nized with recombinant proteins (BFR or P39) plus CpG ODNshowed a high production of IFN-g after stimulation with thespecific antigen or with B. abortus 544, whereas mice treatedwith P39 plus non-CpG ODN failed to do so. Mice treated withBFR plus non-CpG ODN showed a very weak response toBFR or B. abortus 544. In addition, splenocytes from miceimmunized with recombinant proteins, CpG, or non-CpGODN alone released IFN-g only in response to ConA stimu-lation.
We could not detect significant amount of IL-5 in superna-tants from splenocyte cultures derived from any immunizedanimal stimulated with recombinant proteins or B. abortus 544.Nevertheless, IL-5 was detected in the supernatant of ConA-
FIG. 1. SDS-PAGE analysis, followed by Coomassie blue stainingof whole-cell lysate of E. coli BL21(DE3) transformed by pET-15b-bfrbefore induction (lane 1) and after 2 h of induction (lane 2) and of thepurified recombinant BFR (lane 3). Molecular mass standards (laneMM) are indicated on the right.
FIG. 2. Analysis of IgG1 (A) and IgG2a (B) antibody responses of BALB/c mice to the recombinant BFR and P39 antigens and to B. abortus544 extract. Mice were immunized three times with BFR or P39 in different ODN combinations. Sera from four mice per group collected 3 weeksafter the last immunization were assayed individually by ELISA. Antibody levels are expressed as mean titer.
stimulated splenocytes from all mice (from 45 to 220 pg/ml)(data not shown). Induction of a strong IFN-g response whenthe mice are immunized with the BFR or P39 protein withCpG ODN, together with the relative predominance of IgG2aantibodies, suggests that immunization with this adjuvant elic-its a type I immune response to these antigens.
Protection studies. The remaining immunized mice werechallenged by B. abortus 544 to examine the protective activityof the induced immune response. In this experiment, protec-tion was defined as a significant reduction in the number ofbacteria in the spleen from immunized mice compared to themice receiving PBS. The vaccine efficacy was calculated as thelog10 of protection. As expected the B19 vaccine offers a sig-nificant protection at both 4 and 8 weeks postchallenge with2.45 and 2.92 log units of protection, respectively. The onlyprotective antigen in this test is the P39 protein with the CpGadjuvant (Table 2) that shows approximately 2.5 logs of pro-tection at 4 weeks postinfection (p.i.) and 1.2 logs of protectionat 8 weeks p.i. When this vaccine (P391CpG) is compared toB19 vaccine strain, the protection was equivalent at 4 weeksp.i., whereas it was 1.71 logs lower at 8 weeks p.i. (P , 0.001).None of the other combinations of antigens or adjuvant couldinduce protection against B. abortus 544 at any time point afterchallenge (4 and 8 weeks) (see Table 2).
DISCUSSION
The DTH test has been widely used for the diagnosis ofbrucellosis in ruminant (3). For this test, the Brucellergene(Rhone-Merieux) is the more commonly used allergene andconsists of a mixture of 20 to 30 cytoplasmic proteins preparedfrom a rough strain of B. melitensis B115 (22). Previous work
done in our laboratory identified among these proteins the P39and the BFR proteins as Brucella T dominant antigens, whichwere shown to induce a positive DTH in infected guinea pigsand also to stimulate the production of IFN-g by the bloodcells of infected cattle (12). According to the fact that theprotective immune response against Brucella infection is de-scribed as Th1 oriented (44, 58), these characteristics makeboth P39 and BFR attractive candidates for the developmentof a subunitary vaccine against brucellosis if properly com-bined with adjuvant. This potential was evaluated here by usingthese antigens as purified recombinant proteins and CpGODN as an adjuvant in a mouse model of infection.
The choice of this adjuvant was dictated by the fact that itcan stimulate multiple types of immune cells, leading to en-hanced Th1 response characterized by the production ofIFN-g, IL-12, IL-6, IL-18, and tumor necrosis factor alpha (4,7, 17). The production of these cytokines represents an earlyevent in the defense mechanisms against intracellular patho-gens such as Brucella spp. (44, 56, 57). It is also noteworthy thatCpG also enhanced cytolytic CD81 T cells (5). All of thesemechanisms were shown to be involved in protection againstBrucella (35).
In this study, we analyzed both the humoral immune re-sponse and the cell-mediated immune response induced bythese vaccine preparations before assaying their protective ef-ficacies. Since the subclass of IgG response is determined bythe pattern of cytokines secreted by CD4 helper T cells, wemeasured the titers of both the IgG1 and the IgG2a antibodiesraised against the relevant recombinant antigens and theirabilities to react also against a Brucella extract. Significantlevels of IgG1 and IgG2a could be detected only from the seraof mice immunized with P39 mixed with CpG ODN, whereas
FIG. 3. Proliferation of splenocytes from BALB/c mice immunized with the recombinant proteins with CpG or non-CpG ODN (A) or with P39,BFR or ODN alone (B). Splenocytes were stimulated in vitro with BFR, P39 (10 mg/ml), or bacterial lysate (30 mg/ml) or were not stimulated(RPMI). For each mouse (n 5 4) the assays were set up in triplicate. The data represent the mean 6 the standard deviation of the triplicate fromfour mice. ND, not done.
TABLE 1. IFN-g production in spleen cell cultures from immunized mice.
Antigen (concn [mg/ml])IFN-g concn (pg/ml) in 96-h culture supernatant after treatment with:
BFR with CpG ODN induced only IgG2a and to a lower titer.In these conditions, both antigens also induced antibodies ableto react with the whole Brucella extracts. Equal amounts ofnon-CpG ODN mixed with the proteins failed to induce anydetectable antibody response. Both antigens with CpG ODNadjuvant induced a response shifted toward IgG2a. This isconsistent with the enhancement of IgG2a isotype switchingpreviously reported for CpG ODN adjuvant (6).
This IgG2a isotype is important because the binding of theirFc portion to Fc receptors on the surface of phagocytes acti-vates a broad spectrum of antimicrobial responses (e.g., phago-cytosis, cytokine synthesis, release of inflammatory mediators,and generation of reactive oxidant species) (51). Neverthelessthe differences in antibody response between mice vaccinatedwith P39 and mice vaccinated with BFR is striking. No clearexplanation can be given, but it is worth noting that the P39protein was also identified by our group as one of the fewBrucella proteins potentially useful for the serological diagno-sis of brucellosis (28).
Since the cellular arm of the Th1 response is essential forcontrolling intracellular pathogens (41), the splenocyte prolif-erative responses and the cytokines produced after the thirdimmunization were examined. Splenocytes from mice vacci-nated with the BFR or P39 proteins with CpG ODN adjuvantwere able to proliferate and produced consistent amounts ofIFN-g when stimulated in vitro with their specific antigens(BFR or P39) or whole Brucella lysate. On the other hand,splenocytes from animals immunized with the recombinantproteins alone or with non-CpG ODN adjuvant did not induceany cellular immune response. Together with the serologicaldata, these results clearly show that BFR or P39 with CpGadjuvant induced a Th1-oriented immune response in BALB/cmice. In addition, this response persisted for up to 12 weeksafter immunization (data non shown). Like other studies, ourresults confirm that CpG ODN are excellent Th1 adjuvantsand that the inversion from CpG to GpC eliminates this abilityto induce an immune response in vivo (4, 23, 39).
The good and well-oriented immunogenicity of our vaccine
preparations prompted us to test their protective efficacyagainst an infectious B. abortus 544 challenge that was given 3weeks after the last injection. The protection was evaluated at4 and 8 weeks after the challenge. The P39 mixed with CpGODN induced protection in mice when inoculated at doses of20 mg. On the contrary, mice inoculated with P39 alone or withnon-CpG ODN as adjuvant are not protected compared tononvaccinated mice. On the other hand, BFR with CpG ODNdoes not protect mice from B. abortus 544 infection. While weidentified the BFR as a T dominant antigen and while weshowed its ability to induce an appropriate immune response,the BFR protein does not appear to have an important role inthe protective immunity. Other Brucella antigens behave in thesame way, e.g., the 18-kDa outer membrane protein of B.abortus, which induced a good immune response but was notinvolved in mediating protective immunity (52). In addition,the combination BFR protein with P39 in CpG ODN did notincrease the level of protection against B. abortus 544 (data notshown). As expected, the immunization of mice with recombi-nant proteins (BFR or P39) alone or with the nonstimulatoryGpC ODN alone failed to induce a protective response.
Since it was shown that cytokines elicited by CpG ODNgiven alone could prevent the early spread of an intracellularpathogen such as Listeria monocytogens and Leishmania major(27, 45, 59), we were surprised that no level of protection couldbe detected in the group of mice that received 20 mg of CpGODN three times. Reasons for the absence of adjuvant effectcould be the route of immunization used here (i.m.) or moreprobably the longer interval between the last immunizationand the challenge (3 weeks) compared to the conditions usedin previous reports (up to 2 weeks). Elkins et al. have reportedthat the protection against an intracellular pathogen is optimalseveral days after DNA treatment and persists for about 2weeks (14).
Altogether, these data indicate that the protection observedhere is well linked to the specific combination of a good anti-gen (P39) and an adequate adjuvant (CpG ODN). None ofthem used alone was efficient.
Mice vaccinated with B. abortus B19 (used as a vaccinalcontrol) were protected both at 4 weeks p.i. (2.45 logs) and at8 weeks p.i. (2.92 logs). Surprisingly, the same level of protec-tion (2.48 logs) was observed with the P39-CpG ODN vaccine4 weeks after the challenge. To our knowledge, this is the firstdescription for murine brucellosis of a subunitary vaccine of-fering a protection level similar to a live attenuated vaccine. Infact, at 4 weeks the protection level conferred by the L7/L12protein with adjuvant was 1.5 logs less than the protectioninduced by the live vaccine, and no data were reported for theprotection level at 8 weeks postchallenge (36). With regards tothe SOD peptides, the protection was only studied at 2 weekspostchallenge and was not compared to a live vaccine but onlyto salt-extractable proteins (47). We think that the potency ofthe P39 candidate antigen is linked to the protocol of its iden-tification, which was based on the careful selection of the morepotent Brucella T antigen among those contained in the Bru-cellin (INRA) using three parallel models (DTH in infectedguinea pigs, T-cell proliferation, and IFN-g production fromPBMC from infected cattle).
Nevertheless the protection induced by the CpG adjuvantedP39 is not comparable either in quality or in duration to the
TABLE 2. Level of protection against B. abortus 544 conferred byimmunization with the recombinant BFR or P39 proteins alone orwith adjuvant (CpG or non-CpG) compared to the B. abortus B19
vaccine strain
Treatment group(n 5 4)
Mean log10 of brucellae 6 SDin spleen at time (wk)
a This group was vaccinated i.p. with strain B19.b P , 0.0005 (significant) compared with value for control PBS-treated mice.c P , 0.001 (significant) compared with value for control PBS-treated mice.
protection conferred by the B19 live vaccine, which was moreeffective at 8 weeks (2.92 logs) than the subunitary vaccine(1.21 logs).
The challenge by the infectious Brucella can only boost theanti-P39 humoral and cellular immunity, which appeared to besufficient to confer a short-term protection (4 weeks) but ismuch less able to generate a long-term protection. In contrast,the immunity induced by the live smooth Brucella B19 straininvolves not only protective cell-mediated immunity (1) againsta panel of Brucella T-cell epitopes but also humoral responsesagainst the LPS O chain and a variety of proteins. Theseantibodies were demonstrated as partially protective (18, 29,31, 55). This immunity linked to the use of a live vaccine is less“monotone” than the one induced by the P39-CpG ODN, andthis could be part of the explanation for the vanishing of theprotection at 8 weeks p.i.
Other factors may also be involved, such as differences inantigen presentation. It is well known that major histocompat-ibility complex type I-dependent CD81 cells are important foroptimal resistance to Brucella infection (44). These kinds ofeffector cells could well be more easily induced by a live atten-uated vaccine that is still able to replicate intracellularly thanby a recombinant protein even with CpG ODN as adjuvant.Finally, the antigen persistence is also quite different, since thedose used here in BALB/c mice the vaccinal strain B19 is stilldetectable in the spleen 6 weeks after the vaccination (46) andis thus more prone to offer a long-term protection than is theprotein with CpG adjuvant. These later points could be opti-mized by using other delivery systems for the P39 antigen.Naked DNA vaccine combines both the CpG adjuvant effectand an intracellular and persistent expression of the antigen.This is currently under investigation in our laboratory.
ACKNOWLEDGMENTS
We thank G. Houbeau for help with the mouse model.Ayman Al-Mariri holds a fellowship from the Atomic Energy Com-
mission of Syria (AECS). This work was supported by the Commissionof the European Communities, contract no. QLK2-CT-1999-00014.
REFERENCES
1. Araya, L. N., P. H. Elzer, G. E. Rowe, F. M. Enright, and A. J. Winter. 1989.Temporal development of protective cell-mediated and humoral immunity inBALB/c mice infected with Brucella abortus. J. Immunol. 143:3330–3337.
2. Baloglu, S., T. E. Toth, G. G. Schurig, N. Sriranganathan, and S. M. Boyle.2000. Humoral immune response of BALB/c mice to a vaccinia virus recom-binant expressing Brucella abortus GroEL does not correlate with protectionagainst a B. abortus challenge. Vet. Microbiol. 76:193–199.
3. Bercovich, Z., and E. A. ter Laak. 1990. An evaluation of the delayed-typehypersensitivity test for diagnosing brucellosis in individual cattle: a fieldstudy. Vet. Microbiol. 22:241–248.
4. Carson, D. A., and E. Raz. 1997. Oligonucleotide adjuvants for T helper 1(Th1)-specific vaccination. J. Exp. Med. 186:1621–1622.
5. Cho, H. J., K. Takabayashi, P. M. Cheng, M. D. Nguyen, M. Corr, S. Tuck,and E. Raz. 2000. Immunostimulatory DNA-based vaccines induce cytotoxiclymphocyte activity by a T-helper cell-independent mechanism. Nat. Bio-technol. 18:509–514.
6. Chu, R. S., T. McCool, N. S. Greenspan, J. R. Schreiber, and C. V. Harding.2000. CpG oligodeoxynucleotides act as adjuvants for pneumococcal po-lysaccharide-protein conjugate vaccines and enhance antipolysaccharide im-munoglobulin G2a (IgG2a) and IgG3 antibodies. Infect. Immun. 68:1450–1456.
7. Chu, R. S., O. S. Targoni, A. M. Krieg, P. V. Lehmann, and C. V. Harding.1997. CpG oligodeoxynucleotides act as adjuvants that switch on T helper 1(Th1) immunity. J. Exp. Med. 186:1623–1631.
8. Corbel, M. J. 1997. Recent advances in brucellosis. J. Med. Microbiol.46:101–103.
9. Corner, L. A., and G. G. Alton. 1981. Persistence of Brucella abortus strain 19infection in adult cattle vaccinated with reduced doses. Res. Vet. Sci. 31:342–344.
10. de Fays, K., A. Tibor, C. Lambert, C. Vinals, P. Denoel, X. De Bolle, J.Wouters, J. J. Letesson, and E. Depiereux. 1999. Structure and functionprediction of the Brucella abortus P39 protein by comparative modeling withmarginal sequence similarities. Protein Eng. 12:217–223.
11. Denoel, P. A., T. K. Vo, A. Tibor, V. E. Weynants, J. M. Trunde, G. Dubray,J. N. Limet, and J. J. Letesson. 1997. Characterization, occurrence, andmolecular cloning of a 39-kilodalton Brucella abortus cytoplasmic proteinimmunodominant in cattle. Infect. Immun. 65:495–502.
12. Denoel, P. A., T. K. Vo, V. E. Weynants, A. Tibor, D. Gilson, M. S. Zygmunt,J. N. Limet, and J. J. Letesson. 1997. Identification of the major T-cellantigens present in the Brucella melitensis B115 protein preparation, Brucel-lergene OCB. J. Med. Microbiol. 46:801–806.
13. Denoel, P. A., M. S. Zygmunt, V. Weynants, A. Tibor, B. Lichtfouse, P.Briffeuil, J. N. Limet, and J. J. Letesson. 1995. Cloning and sequencing of thebacterioferritin gene of Brucella melitensis 16M strain. FEBS Lett. 361:238–342.
14. Elkins, K. L., T. R. Rhinehart-Jones, S. Stibitz, J. S. Conover, and D. M.Klinman. 1999. Bacterial DNA containing CpG motifs stimulates lympho-cyte-dependent protection of mice against lethal infection with intracellularbacteria. J. Immunol. 162:2291–2298.
15. Fensterbank, R., and M. Plommet. 1979. Vaccination against bovine brucel-losis with a low dose of strain 19 administered by the conjunctival route. IV.Comparison between two methods of vaccination. Ann. Rech. Vet. 10:131–139.
16. Godfroid, F., A. Cloeckaert, B. Taminiau, I. Danese, A. Tibor, X. de Bolle, P.Mertens, and J. J. Letesson. 2000. Genetic organisation of the lipopolysac-charide O-antigen biosynthesis region of Brucella melitensis 16M (wbk). Res.Microbiol. 151:655–668.
17. Huang, L., A. M. Krieg, N. Eller, and D. E. Scott. 1999. Induction andregulation of Th1-inducing cytokines by bacterial DNA, lipopolysaccharide,and heat-inactivated bacteria. Infect. Immun. 67:6257–6263.
18. Jacques, I., A. Cloeckaert, J. N. Limet, and G. Dubray. 1992. Protectionconferred on mice by combinations of monoclonal antibodies directedagainst outer-membrane proteins or smooth lipopolysaccharide of Brucella.J. Med. Microbiol. 37:100–103.
19. Jakob, T., P. S. Walker, A. M. Krieg, M. C. Udey, and J. C. Vogel. 1998.Activation of cutaneous dendritic cells by CpG-containing oligodeoxynucle-otides: a role for dendritic cells in the augmentation of Th1 responses byimmunostimulatory DNA. J. Immunol. 161:3042–3049.
20. Jakob, T., P. S. Walker, A. M. Krieg, E. von Stebut, M. C. Udey, and J. C.Vogel. 1999. Bacterial DNA and CpG-containing oligodeoxynucleotides ac-tivate cutaneous dendritic cells and induce IL-12 production: implicationsfor the augmentation of Th1 responses. Int. Arch. Allergy Immunol. 118:457–461.
21. Janeway, C. A., S. Carding, B. Jones, J. Murray, P. Portoles, R. Rasmussen,J. Rojo, K. Saizawa, J. West, and K. Bottomly. 1988. CD41 T cells: specificityand function. Immunol. Rev. 101:39–80.
22. Jones, L. M., R. Diaz, and A. G. Taylor. 1973. Characterization of allergensprepared from smooth and rough strains of Brucella melitensis. Br. J. Exp.Pathol. 54:492–508.
23. Klinman, D. M., A. K. Yi, S. L. Beaucage, J. Conover, and A. M. Krieg. 1996.CpG motifs present in bacteria DNA rapidly induce lymphocytes to secreteinterleukin 6, interleukin 12, and interferon gamma. Proc. Natl. Acad. Sci.USA 93:2879–2883.
24. Kovarik, J., P. Bozzotti, L. Love-Homan, M. Pihlgren, H. L. Davis, P. H.Lambert, A. M. Krieg, and C. A. Siegrist. 1999. CpG oligodeoxynucleotidescan circumvent the Th2 polarization of neonatal responses to vaccines butmay fail to fully redirect Th2 responses established by neonatal priming.J. Immunol. 162:1611–1617.
25. Krieg, A. M. 1999. CpG DNA: a novel immunomodulator. Trends Microbiol.7:64–65.
26. Krieg, A. M. 2000. The role of CpG motifs in innate immunity. Curr. Opin.Immunol. 12:35–43.
27. Krieg, A. M., L. Love-Homan, A. K. Yi, and J. T. Harty. 1998. CpG DNAinduces sustained IL-12 expression in vivo and resistance to Listeria mono-cytogenes challenge. J. Immunol. 161:2428–2434.
28. Letesson, J. J., A. Tibor, G. van Eynde, V. Wansard, V. Weynants, P. Denoel,and E. Saman. 1997. Humoral immune responses of Brucella-infected cattle,sheep, and goats to eight purified recombinant Brucella proteins in an indi-rect enzyme-linked immunosorbent assay. Clin. Diagn. Lab. Immunol.4:556–564.
29. Limet, J. N., N. Bosseray, B. Garin-Bastuji, G. Dubray, and M. Plommet.1989. Humoral immunity in mice mediated by monoclonal antibodies againstthe A and M antigens of Brucella. J. Med. Microbiol. 30:37–43.
30. Lin, J., L. G. Adams, and T. A. Ficht. 1996. Immunological response to theBrucella abortus GroEL homolog. Infect. Immun. 64:4396–4400.
31. Montaraz, J. A., A. J. Winter, D. M. Hunter, B. A. Sowa, A. M. Wu, and L. G.Adams. 1986. Protection against Brucella abortus in mice with O-polysaccha-ride-specific monoclonal antibodies. Infect. Immun. 51:961–963.
32. Nielsen, K., J. W. Cherwonogrodzky, J. R. Duncan, and D. R. Bundle. 1989.Enzyme-linked immunosorbent assay for differentiation of the antibody re-
VOL. 69, 2001 PROTECTION AGAINST BRUCELLA ABORTUS 4821
sponse of cattle naturally infected with Brucella abortus or vaccinated withstrain 19. Am. J. Vet. Res. 50:5–9.
33. O’Garra, A., and K. Murphy. 1994. Role of cytokines in determining T-lymphocyte function. Curr. Opin. Immunol. 6:458–466.
34. Oliveira, S. C., J. S. Harms, M. Banai, and G. A. Splitter. 1996. RecombinantBrucella abortus proteins that induce proliferation and gamma-interferonsecretion by CD41 T cells from Brucella-vaccinated mice and delayed-typehypersensitivity in sensitized guinea pigs. Cell. Immunol. 172:262–268.
35. Oliveira, S. C., and G. A. Splitter. 1995. CD81 type 1 CD44hi CD45 RBlo Tlymphocytes control intracellular Brucella abortus infection as demonstratedin major histocompatibility complex class I- and class II-deficient mice. Eur.J. Immunol. 25:2551–2557.
36. Oliveira, S. C., and G. A. Splitter. 1996. Immunization of mice with recom-binant L7/L12 ribosomal protein confers protection against Brucella abortusinfection. Vaccine 14:959–962.
37. Oliveira, S. C., Y. Zhu, and G. A. Splitter. 1994. Recombinant L7/L12ribosomal protein and gamma-irradiated Brucella abortus induce a T-helper1 subset response from murine CD41 T cells. Immunology 83:659–664.
38. Onate, A. A., R. Vemulapalli, E. Andrews, G. G. Schurig, S. Boyle, and H.Folch. 1999. Vaccination with live Escherichia coli expressing Brucella abor-tus Cu/Zn superoxide dismutase protects mice against virulent B. abortus.Infect. Immun. 67:986–988.
39. Roman, M., E. Martin-Orozco, J. S. Goodman, M. D. Nguyen, Y. Sato, A.Ronaghy, R. S. Kornbluth, D. D. Richman, D. A. Carson, and E. Raz. 1997.Immunostimulatory DNA sequences function as T helper-1-promoting ad-juvants. Nat. Med. 3:849–854.
40. Roop, R. M., T. W. Fletcher, N. M. Sriranganathan, S. M. Boyle, and G. G.Schurig. 1994. Identification of an immunoreactive Brucella abortus HtrAstress response protein homolog. Infect. Immun. 62:1000–1007.
41. Seder, R. A., and A. V. Hill. 2000. Vaccines against intracellular infectionsrequiring cellular immunity. Nature 406:793–798.
42. Smith, L. D., and T. A. Ficht. 1990. Pathogenesis of Brucella. Crit. Rev.Microbiol. 17:209–230.
43. Sperry, J. F., and D. C. Robertson. 1975. Inhibition of growth by erythritolcatabolism in Brucella abortus. J. Bacteriol. 124:391–397.
44. Splitter, G., S. Oliveira, M. Carey, C. Miller, J. Ko, and J. Covert. 1996. Tlymphocyte mediated protection against facultative intracellular bacteria.Vet. Immunol. Immunopathol. 54:309–319.
45. Stacey, K. J., and J. M. Blackwell. 1999. Immunostimulatory DNA as anadjuvant in vaccination against Leishmania major. Infect. Immun. 67:3719–3726.
46. Stevens, M. G., S. C. Olsen, G. W. Pugh, Jr., and M. V. Palmer. 1994.Immune and pathologic responses in mice infected with Brucella abortus 19,RB51, or 2308. Infect. Immun. 62:3206–3212.
47. Tabatabai, L. B., and G. W. Pugh. 1994. Modulation of immune responses in
48. Tibor, A., V. Weynants, P. Denoel, B. Lichtfouse, X. De Bolle, E. Saman, J. N.Limet, and J. J. Letesson. 1994. Molecular cloning, nucleotide sequence, andoccurrence of a 16.5-kilodalton outer membrane protein of Brucella abortuswith similarity to PAL lipoproteins. Infect. Immun. 62:3633–3639.
49. Toth, T. E., J. A. Cobb, S. M. Boyle, R. M. Roop, and G. G. Schurig. 1995.Selective humoral immune response of BALB/c mice to Brucella abortusproteins expressed by vaccinia virus recombinants. Vet. Microbiol. 45:171–183.
50. Trinchieri, G. 1997. Cytokines acting on or secreted by macrophages duringintracellular infection (IL-10, IL-12, IFN-gamma). Curr. Opin. Immunol.9:17–23.
51. Unkeless, J. C., E. Scigliano, and V. H. Freedman. 1988. Structure andfunction of human and murine receptors for IgG. Annu. Rev. Immunol.6:251–281.
52. Vemulapalli, R., S. Cravero, C. L. Calvert, T. E. Toth, N. Sriranganathan,S. M. Boyle, O. L. Rossetti, and G. G. Schurig. 2000. Characterization ofspecific immune responses of mice inoculated with recombinant vacciniavirus expressing an 18-kilodalton outer membrane protein of Brucella abor-tus. Clin. Diagn. Lab. Immunol. 7:114–118.
53. Vemulapalli, R., A. J. Duncan, S. M. Boyle, N. Sriranganathan, T. E. Toth,and G. G. Schurig. 1998. Cloning and sequencing of yajC and secD homologsof Brucella abortus and demonstration of immune responses to YajC in micevaccinated with B. abortus RB51. Infect. Immun. 66:5684–5691.
54. Vemulapalli, R., Y. He, S. Cravero, N. Sriranganathan, S. M. Boyle, andG. G. Schurig. 2000. Overexpression of protective antigen as a novel ap-proach to enhance vaccine efficacy of Brucella abortus strain RB51. Infect.Immun. 68:3286–3289.
55. Vizcaino, N., and L. Fernandez-Lago. 1994. Protection and suppression ofthe humoral immune response in mice mediated by a monoclonal antibodyagainst the M epitope of Brucella. FEMS Immunol. Med. Microbiol. 8:133–139.
56. Zhan, Y., and C. Cheers. 1993. Endogenous gamma interferon mediatesresistance to Brucella abortus infection. Infect. Immun. 61:4899–4901.
57. Zhan, Y., and C. Cheers. 1995. Endogenous interleukin-12 is involved inresistance to Brucella abortus infection. Infect. Immun. 63:1387–1390.
58. Zhan, Y., Z. Liu, and C. Cheers. 1996. Tumor necrosis factor alpha andinterleukin-12 contribute to resistance to the intracellular bacterium Brucellaabortus by different mechanisms. Infect. Immun. 64:2782–2786.
59. Zimmermann, S., O. Egeter, S. Hausmann, G. B. Lipford, M. Rocken, H.Wagner, and K. Heeg. 1998. CpG oligodeoxynucleotides trigger protectiveand curative Th1 responses in lethal murine leishmaniasis. J. Immunol.160:3627–3630.