33
Protein engineering: promising approach for the development of new antimicrobials Ismail Fliss & Riadh Hammami STELA conference, May13-14 2013
| Date post: | 14-Feb-2018 |
| Category: |
Documents |
| Upload: | phungkhanh |
| View: | 219 times |
| Download: | 3 times |

Protein engineering: promising approach for the development of new antimicrobials
Ismail Fliss & Riadh Hammami
STELA conference, May13-14 2013

STELA conference, May13-14 2013
Team at INAF: identification, caracterisation and study of the structure/fonction relationship of bactériocins produced by lactic acid and probiotic bacteria.
Team at CHUQ research centre, laboratory of medical chemistry: synthesis of bioactive peptides.
FQRNT-Team project

STELA conference, May13-14 2013
Large scale synthesis of new, more stable antimicrobial molecules with larger spectrum of antimicrobial activity from bacteriocin peptide sequences.
Microcine J25
General objective

STELA conference, May13-14 2013
Peptides or polypeptides naturally produced by several bacterial strains of food and environmental origin.
Antimicrobial activity (bactericidal or bacteriostatic) against microorganisms phylogenetically related to the producing strain.
Bacteriocins

STELA conference, May13-14 2013
Widespread phenomenon (Lactococci and Lactobacilli);
Broad spectrum of antimicrobial activity;
Acid and heat resistance;
150 Gram (+) and 25 Gram (-) bacteriocins: BACTIBASE
Bacteriocins

STELA conference, May13-14 2013
Nisin is the only bacteriocin approved in more than 50 countries. Carnobacterium maltaromaticum CB1: antimicrobial agent for preservation of meat products (Health Canada, food additive) C. divergens M35: bioconservation agent for smoked sea products (Health Canada, new foods section) Pediococcus acidilactici UL5 (pediocin producer): probiotic strain for veterinary use (Health Canada, Natural health products)
This image cannot currently be displayed.
Legal status of bacteriocins

STELA conference, May13-14 2013
Food : natural food additives (Nisin)
Pharmaceutical: skin infections, antibiotic resistant bacteria
Veterinary: Treatment and prevention of bovine mastitis (WIPE OUT® Immucell)
Applications

STELA conference, May13-14 2013
Advantages and limits
Limits Advantages
Food grade substances: completely
degraded in the digestive tract
High biological activity: 103 to 106
times higher than other antimicrobials,
including antibiotics;
High thermal stability: compatible with
various heat treatments
Narrow spectrum of inhibition activity
Instability in biological matrices
(proteases, extreme pH, salt).
Low production yields (10%): difficulty
for large-scale production, high costs,
etc.

STELA conference, May13-14 2013
Microbiological approach: selection of highly productive strains (L. lactis diacetylatis UL719, P. acidilactici UL5), optimization of fermentation processes (continuous fermentation with immobilized cells) and purification (immunoaffinity). Molecular biology approach: cloning and production of recombinant bacteriocins Protein engineering approach: peptide synthesis (knowledge of the structural characteristics and relationships with biological activity)
Improving yield

STELA conference, May13-14 2013
Microcin J25
Main Objective
Elucidate the relationship between the primary and secondary structures of bacteriocins and their biological function. The ultimate goal is to exploit the knowledge generated for the large-scale synthesis of new, more stable and biologically active antimicrobial molecules with a broad spectrum of inhibition activity.

STELA conference, May13-14 2013
Microcin J25

STELA conference, May13-14 2013
Microcin J25
21 AA Unique structure : Lasso 3 β-sheets
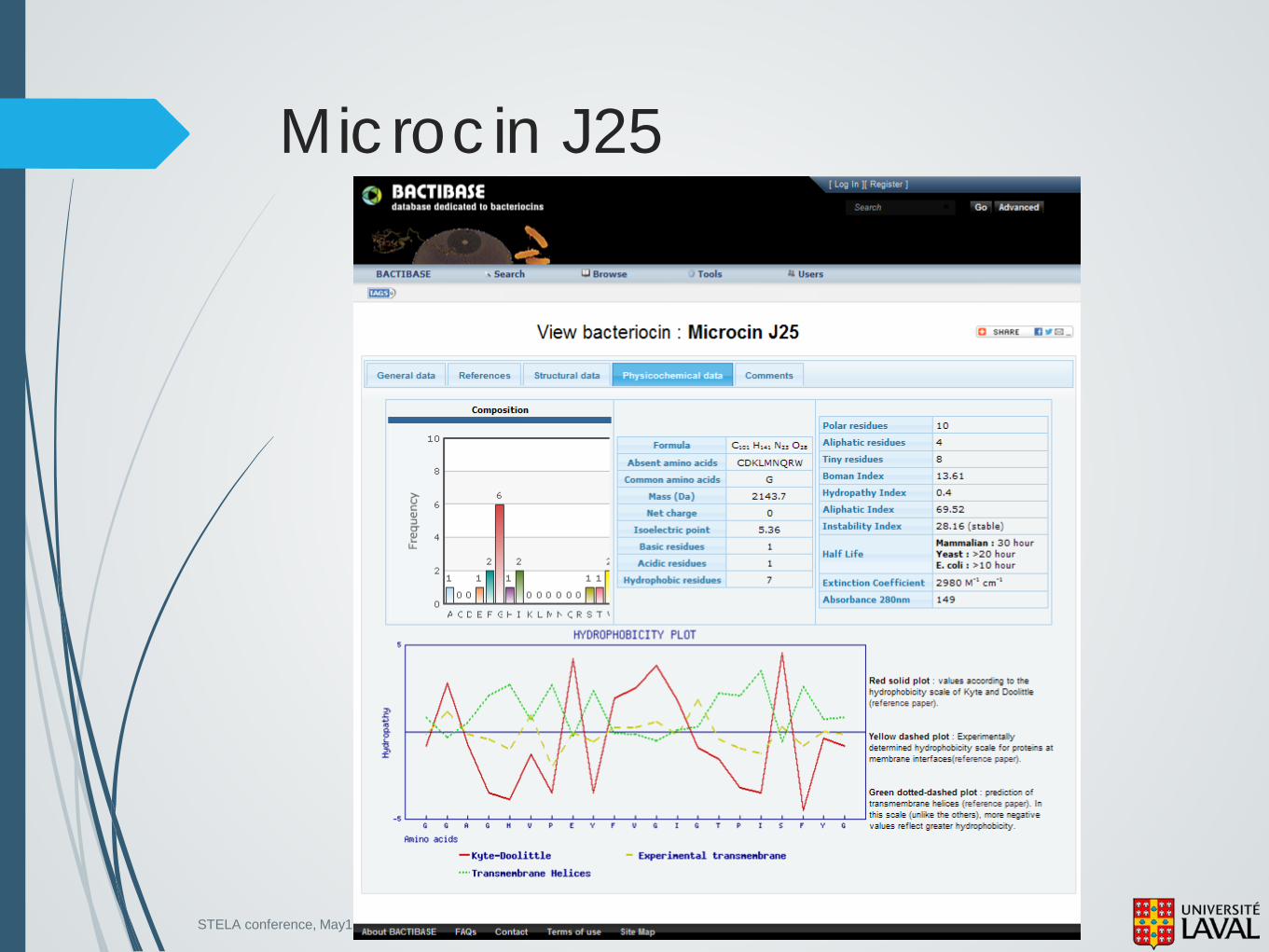
STELA conference, May13-14 2013
Microcin J25

STELA conference, May13-14 2013
Structure-function relationship of microcin J25
Several studies using molecular biology and chemical synthesis have attempted to produce microcin J25 or active derivatives but without success. Lack of knowledge about the relationship between the particular Lasso structure of microcin and its biological activity: - Role of the N-terminal 1-8 loop - The role of the C-terminal tail (with two segments: 9-17 and 18-21) - The presence of some amide bridges (cyclisation G1-E8) - The presence of some specific AA.

STELA conference, May13-14 2013
GGAGH VPEYF VGIGT PISFYG
Is overall conformation a prerequisite for Microcin biological activity?
J25
C1
C2
C3 GGAGH VPCYF VGIGT PICFYG
CGAGH VPCYF VGIGT PISFYG
CGAGH VPEYF VGIGT PICFYG C8 C18
C1 G1
C8
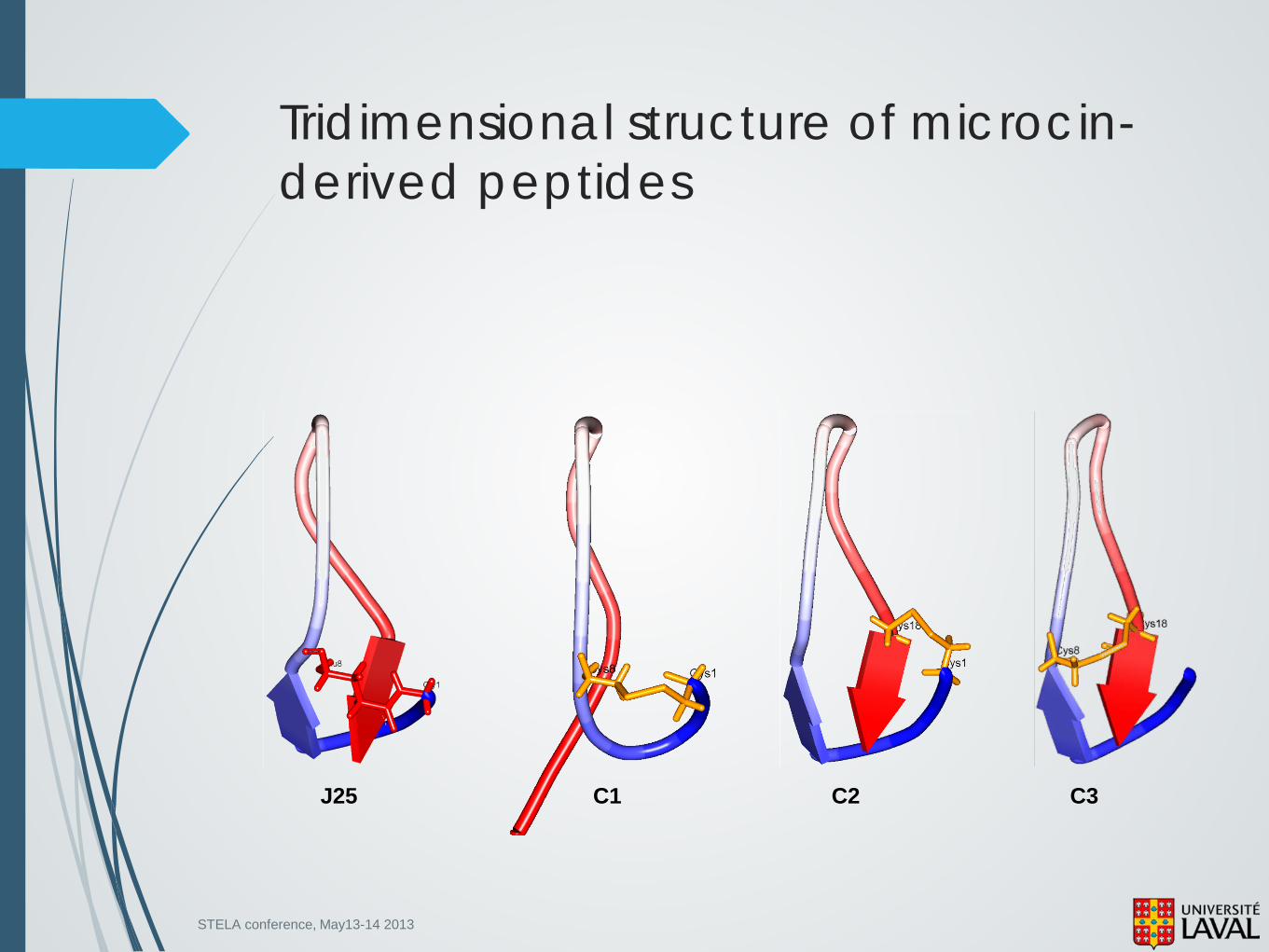
STELA conference, May13-14 2013
Tridimensional structure of microcin-derived peptides
J25 C1 C2 C3

STELA conference, May13-14 2013
Is the whole sequence of microcin required for its biological activity?
GGAGH VPEYF VGIGT PISFY G
--YF VGIGT PISFY G
-CYF VGIGT PICFY G
KCYF VGIGT PICFY G
J25
9-21
8-21
7-21

STELA conference, May13-14 2013
J25 7-21
Tridimensional structure of microcin-derived peptides

STELA conference, May13-14 2013
GGAGH VPEYF VGIGT PISFY G
GWKGK WKCYF VGIGT PICFY G
J25
WK_7-21
Does the overall net charge influence Microcin biological activity?
-1
+3

STELA conference, May13-14 2013
J25 WK-7-21
Tridimensional structure of microcin-derived peptides

STELA conference, May13-14 2013
Synthesized peptides
Peptide Sequence link Mass (Da) Net charge
MccJ25* GGAGHVPEYFVGIGTPISFYG 1-8 2 145.0 -1 C1 CGAGHVPCYFVGIGTPISFYG 1-8 2 145.0 0 C2 CGAGHVPEYFVGIGTPICFYG 1-18 2 145.0 -1 C3 GGAGHVPCYFVGIGTPICFYG 8-18 2 145.0 0
1-8L GGAGHVPE------------- - 723.0 -1 1-10C GGAGHVPEYF----------- 1-8 1033.0 -1
NC1 GGAGHVPCYF-GKG---CFYG 8-18 1717.8 +1 NC2 GGAGHVPCYF-KKK---CFYG 8-18 1861.0 +3 NC3 GGAGHVPCYF-WKW---CFYG 8-18 1976.8 +1
9-21L --------YFVGIGTPISFYG-NH2 - 1420.0 0 9-21C --------YFVGIGTPISFYG 9-18 1420.0 0 8-21C -------CYFVGIGTPICFYG 8-18 1540.0 0 7-21C ------KCYFVGIGTPICFYG 8-18 1668.0 +1
WK_7-21 GWKGKWKCYFVGIGTPICFYG 8-18 2410.0 +3

STELA conference, May13-14 2013
Minimal inhibitory concentration (MHI) of the peptides
Strains MIC (µM)
MccJ25* NC1 NC2 NC3 C1 C3 8-21C 7-21C WK_7_21
Salmonella enterica subsp. enterica ATCC 14028 6.5 - - - 125-250 - - - -
Salmonella enterica subsp. enterica ATCC 8387 0.1 - - 31.3 1.0 15.6-31.3 1.0 7.8-15.6 7.8
Salmonella enterica subsp. enterica ATCC 29628 6.5 - 250 - - - - - -
Salmonella enterica subsp. enterica ATCC 8400 0.8 - - - 62.5-125 - - 125-250 62.5
Salmonella enterica subsp. enterica ATCC 9607 1.6 - - - 250 - - - -
Salmonella enterica subsp. enterica ATCC 9700 0.4 - - - 250 - - - -
Escherichia coli ATCC 11229 0.2 - - - 62.5 - 125-250 - 250
Escherichia coli ATCC 25922 3.3 - - - 250.0 - - 250 - Escherichia coli ATCC 15144 - - - - - - - - -
Escherichia coli O157 H7 ATCC 35150 - - - - - - - - -
Escherichia coli MC4100 ATCC 35695 6.5 - - - 31.3-62.5 - - 125 -
Escherichia coli DH5a 6.5 - - - - - - - - Escherichia coli LR 05 - - - - >250 - - >250 >250
Listeria ivanovii HPB28 - - - - >250 - - >250 250
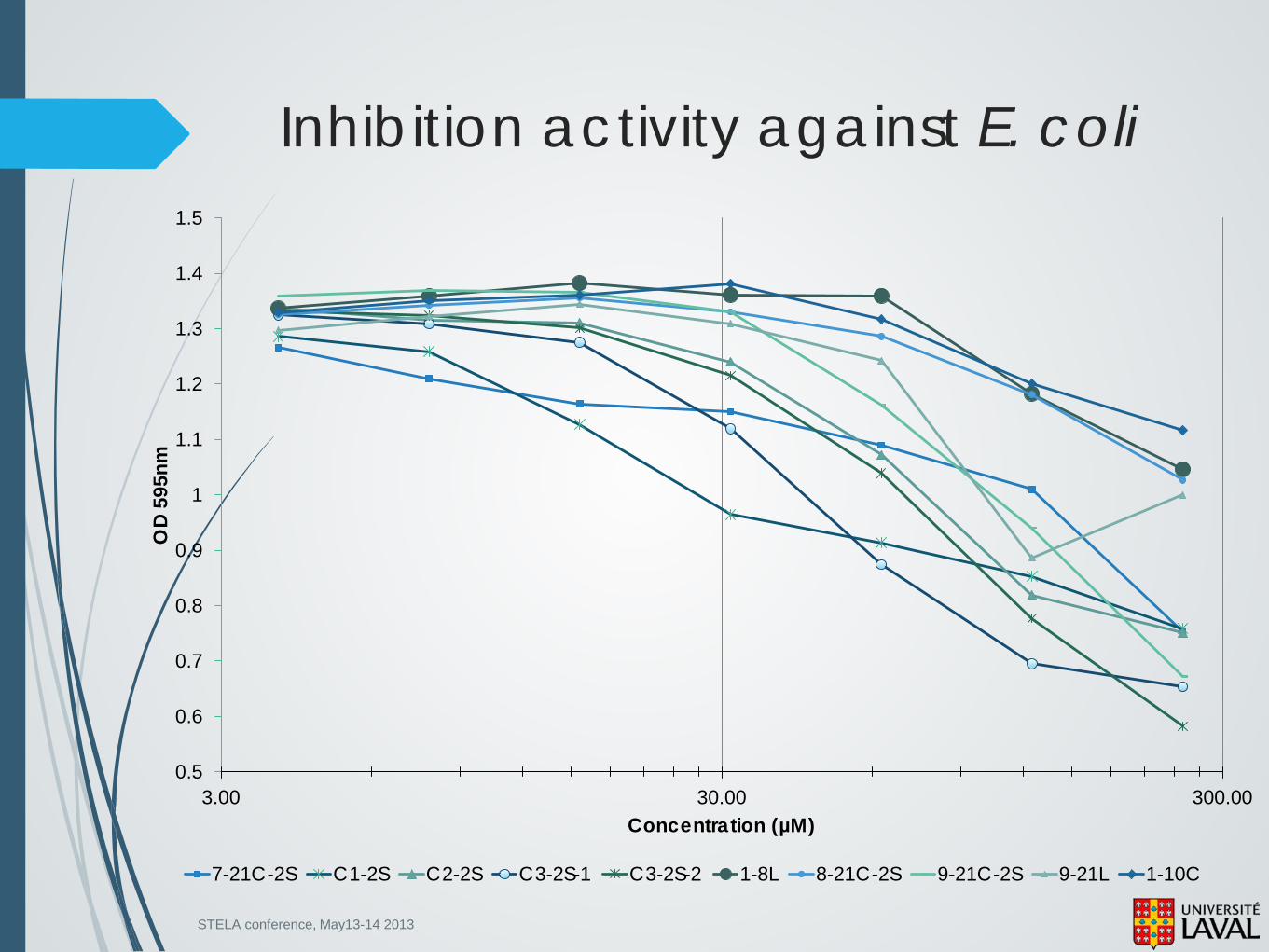
STELA conference, May13-14 2013
0.5
0.6
0.7
0.8
0.9
1
1.1
1.2
1.3
1.4
1.5
3.00 30.00 300.00
OD
595n
m
Concentration (µM)
7-21C-2S C1-2S C2-2S C3-2S-1 C3-2S-2 1-8L 8-21C-2S 9-21C-2S 9-21L 1-10C
Inhibition activity against E. coli

STELA conference, May13-14 2013
Inhibition activity against S. enterica ATCC 8387
0
20
40
60
80
100
MccJ25 C1 C3 NC3 8-21C 7-21C WK_7_21
Inhi
bitio
n (%
)
0.5 µM 7.8 µM

STELA conference, May13-14 2013
Growth inhibition of Listeria ivanovii by active peptides at 250µM
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0 4 8 12 16 20 24
Opt
ical
den
sity
(595
nm)
Time (Hours)
Control C1
7-21C WK_7-21

STELA conference, May13-14 2013
Electrostatic Potential (Face A)
Microcin J25 WK_7-21

STELA conference, May13-14 2013
Microcin J25 WK_7-21
Electrostatic Potential (Face B)

STELA conference, May13-14 2013
Secondary structure
Circular dichroïsm experiments :
The secondary structure of active peptides is very
similar to that of native microcin J25 (β sheets +
coils)

STELA conference, May13-14 2013
CONCLUSIONS
R1: yes, the lasso structure is important but not prerequisite for the biological activity.
R2: No, for example the ring plays an important role in the antimicrobial activity of microcin (C1) but its deletion doesn’t totally abolish the inhibitory activity (7-21)
R3: yes, the substitution of some amino acids to increase the overall charge seems to enhance the activity of microcin and extend its spectrum of action toward Gram+ (WK-7-21)

PERSPECTIVES
The new molecules can be synthesized on a large scale with high yields.
Opens the door to applications of these molecules in several areas (food, medical and veterinary) where the search for these new molecules is becoming more urgent.

Acknowledgements
François Bédard
Eric Biron
Pedro Alvares
Muriel Subirade
Fond québécois de recherche sur la nature et les technologies (FQRNT-Équipe)

Destruction of membrane potential
Nucleolytic and related activities
Inhibition of protein synthesis
Out
In
Leakage of cellular content
(ions, ATP)
insertion and pore formation
Nisin
Lipid II
Pediocin
Man-PTS
Electrostatic interactions
Translocation into the cytoplasm (e.g. colicins)
Bacteriocin
OR
Target recognition
Mem
bran
e
e.g. Sakacin
Receptor
or

+ + +
- - - -
-
Attraction électrostatique
Perturbation de la
membrane Insertion bioénergétique
et interaction
Peptides antimicrobiens: Mode d’action général
+ +
D’une manière générale le mode d’action se situe au niveau de la membrane :

















