64
Protein spectroscopy and dynamics Vibrational spectroscopy Time-resolved spectroscopy Hemoglobin Myoglobin Enzymes Protein Folding
| Date post: | 21-Dec-2015 |
| Category: |
Documents |
| View: | 237 times |
| Download: | 0 times |

Protein spectroscopy and dynamics
Vibrational spectroscopy
Time-resolved spectroscopy
Hemoglobin
Myoglobin
Enzymes
Protein Folding

Dynamics in Proteins• Dynamics consist of:
• Protein relaxation in response to - ligand/substrate binding - electron transfer
• Protein folding. - cyclic compared to -sheet peptides - unfolded - molten globule - folded
• Time-resolved vibrational spectroscopy is a tool for investigation of structural changes.

Vibrational Spectroscopy
Quantum theory
Normal modes
Infrared absorption
Raman scattering
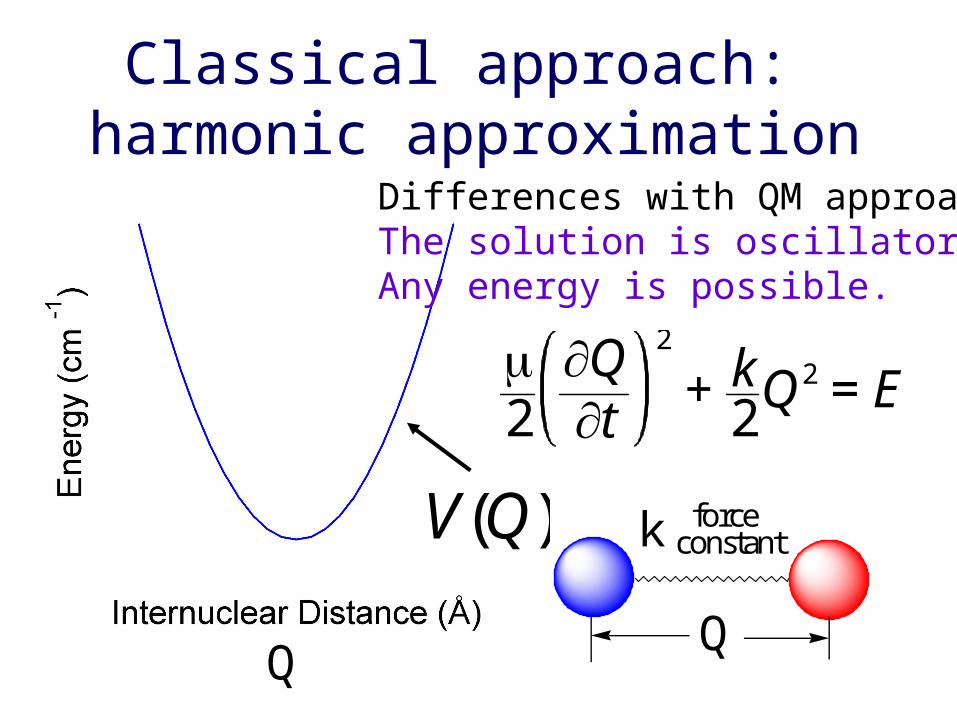
Classical approach: harmonic approximation
V(Q) k force constant
Q
Differences with QM approach:The solution is oscillatory.Any energy is possible.
2
Qt
2
+ k2Q2 = E
Q

Quantum theory of vibrationHarmonic approximation
– h
22Q2 + k
2Q2 = E
v is the quantum numberAllowed transitionsv’ v + 1, v’ v - 1
E = v + 12h
Q
Energy is quantized
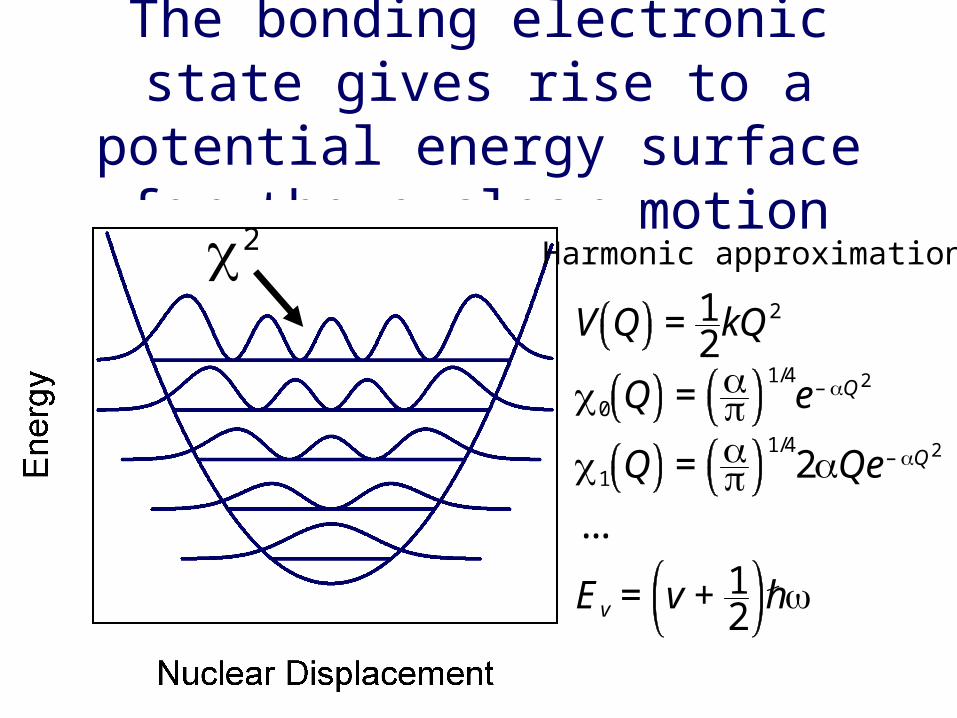
The bonding electronic state gives rise to a potential energy surface for
the nuclear motionHarmonic approximation
V Q = 12kQ2
0 Q =
1/4e– Q2
1 Q =
1/42Qe– Q2
...
Ev = v + 12h
2

There is a potential energy surface that corresponds to each electronic
state of the molecule
The shift in the nucleardisplacement arises fromthe fact that the bondlength increases in the* state compared to the state. We will show thatthe overlap of the vibra--tional wave functions is key to understanding theshape of absorption bands.

There are 3N-6 vibrational degrees of freedom in a molecule with N atoms
Three degrees of freedom are required for translation.Three degrees of freedom are required for rotation.For example, in H2O there are 9 total degrees of freedomand 3 vibrational degrees of freedom.In C6H6 there are 36 degrees of freedom and 30 vibrationaldegrees of freedom.Exception: In linear molecules there are only 2 rotationaldegrees of freedom and therefore the number of vibrationsis 3N - 5.

The vibrational degrees of freedom can be expressed as normal modes.
All normal modes have the same form for the harmonicoscillator wavefunction and differ only in the forceconstant k and mass . The total wavefunction is a product of normal modes. The total nuclear wavefunction for water is 123.The normal mode wavefunctions of water correspondto the symmetric stretch, bend, and asymmetric stretch.These are linear combinations of the stretching and bending internal coordinates of H2O.
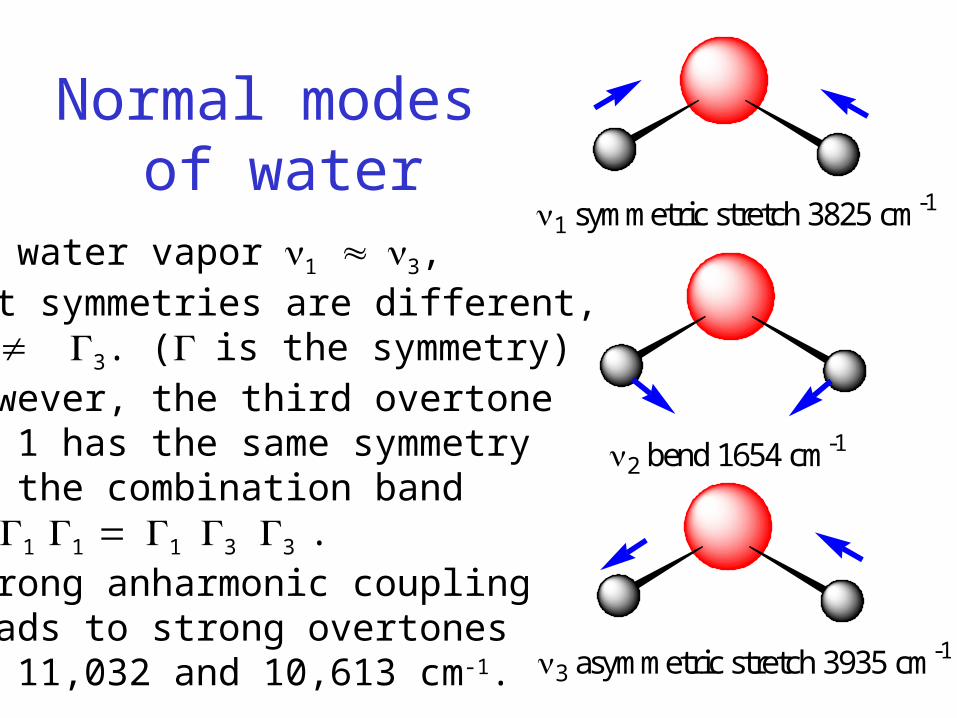
Normal modes of water
1 symmetric stretch 3825 cm-1
2 bend 1654 cm-1
3 asymmetric stretch 3935 cm-1
In water vapor 1 3, but symmetries are different, 1 3. (is the symmetry)However, the third overtoneof 1 has the same symmetryas the combination band1 1 1 13 3 .Strong anharmonic couplingleads to strong overtonesat 11,032 and 10,613 cm-1.
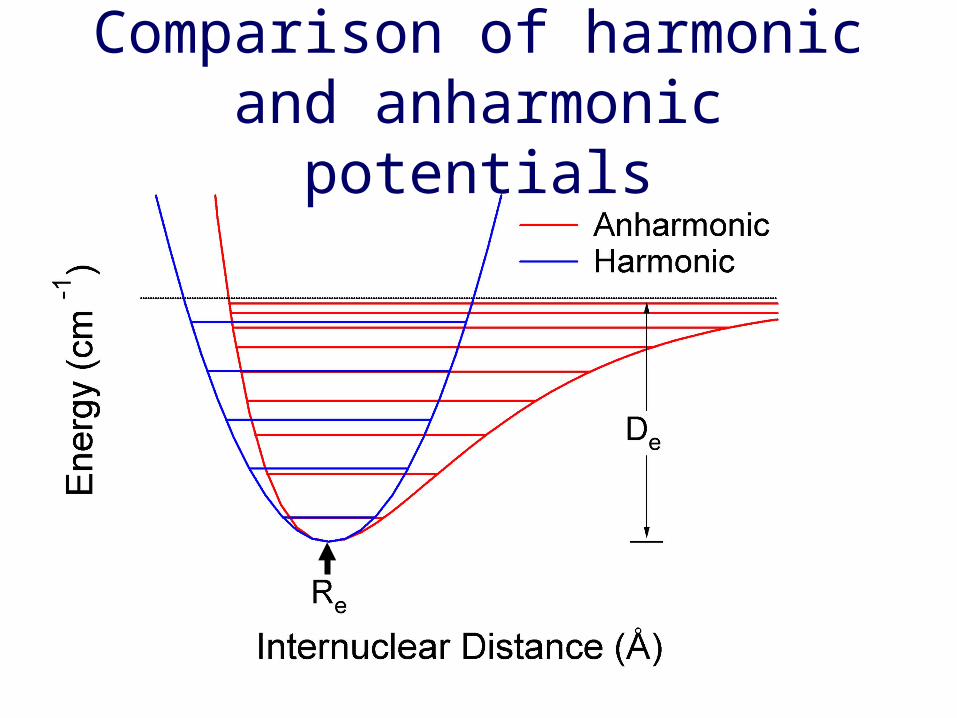
Comparison of harmonic and anharmonic potentials

Frequency shift due to molecular interactions
Hydrogen bonding lowers O-H force constantand H-O-H bending force constant.
The intermolecular hydrogen bondingstretching mode is difficult to observe.
vapor liquid1 3825 36572 1654 15953 3935 3756

VibrationalTransitions

Transition dipoles
In order for infrared light to be absorbed the polarization must be aligned with the directionof the transition moment. For a vibrational modethis is determined by the directional change inthe dipole moment. This is shown below forthe bending mode of H2O.
H
O
H H
O
H

Transition dipoles
The change in ground state dipole momentduring vibration interacts with light.
The first term is static and does not contributeto the transition. Calling the vibrational wave-functions i the transition moment is:
= g +g
Q Q +
10 =g
Q 1Q0dQ

Dipole derivatives
The vibrational wavefunctions i are Gaussians,thus the transition moment for transition fromvibrational state 0 to vibrational state 1 is:
The transition dipole moment is proportional tothe dipole derivative. This is true for any normal mode of vibration (i.e. harmonic).
10 =g
Q e– Q2/2Q2e– Q2/2dQ–
= 12
g
Q

Absorption of infrared radiation leads to vibrational transitions
v = 0

Absorption of infrared radiation leads to vibrational transitions
v = 0
v = 1

The selection rule for vibrational transitions is v = 1
v = 0
v = 1
v = 2

Analysis of isotope effectsVibrational spectra are analyzed within theharmonic approximation.
m1 m2k
xt2
+ kx = 0
Classical harmonic oscillator equation
=m1m2
m1 + m2
= kx = Acos t
Reduced mass

Raman spectroscopy
Goal: Study vibrational frequencies of the heme and theaxial ligands in order to obtain information
on the coupling of protein motion and electrostatics withthe heme iron
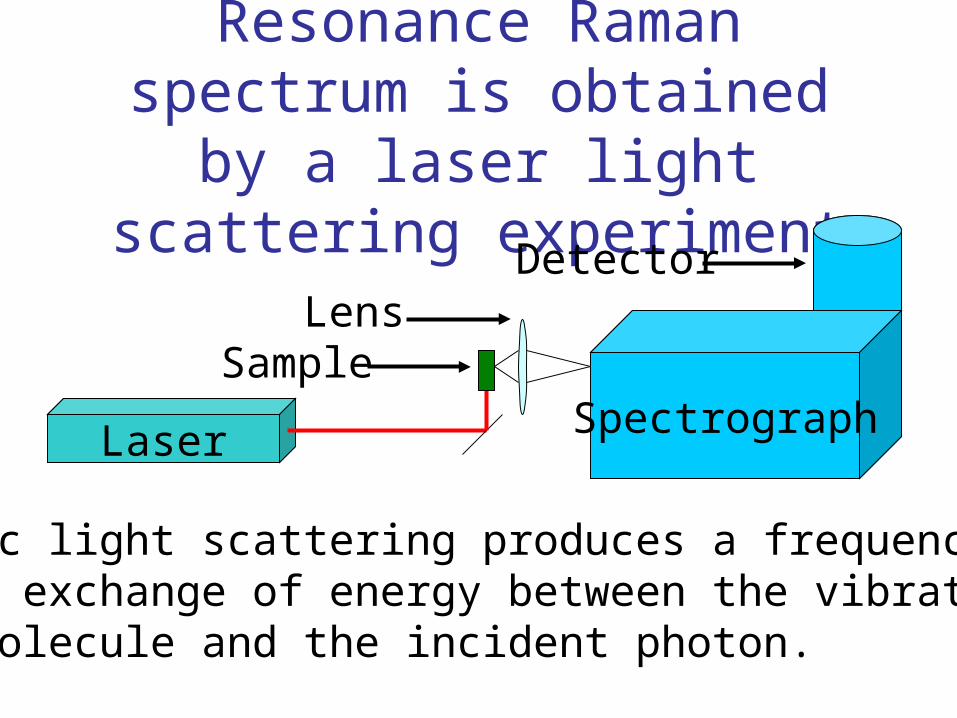
Resonance Raman spectrum is obtained by a laser light scattering experiment
Laser Spectrograph
Detector
Sample
Inelastic light scattering produces a frequency shift.There is exchange of energy between the vibrationsof the molecule and the incident photon.
Lens
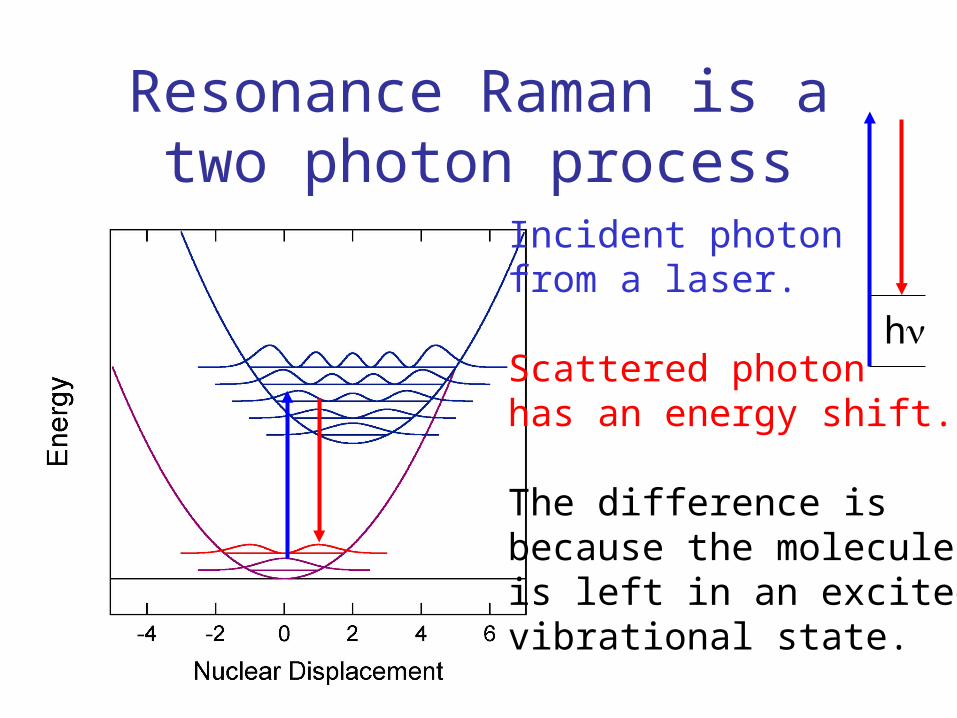
Resonance Raman is a two photon process
Incident photonfrom a laser.
Scattered photonhas an energy shift.
The difference isbecause the moleculeis left in an excitedvibrational state.
h

N N
NN
O O-O O-
Fe
The iron in heme is the binding site for oxygen and peroxide
Heme is iron protoporphyrin IX.
Functional aspects in Mb
O|||O

N N
NN
O O-O O-
Fe
The iron in heme is the binding site for oxygen and peroxide
Heme is iron protoporphyrin IX.
Functional aspects in Mb
1. Discrimination againstCO binding.
O|||C
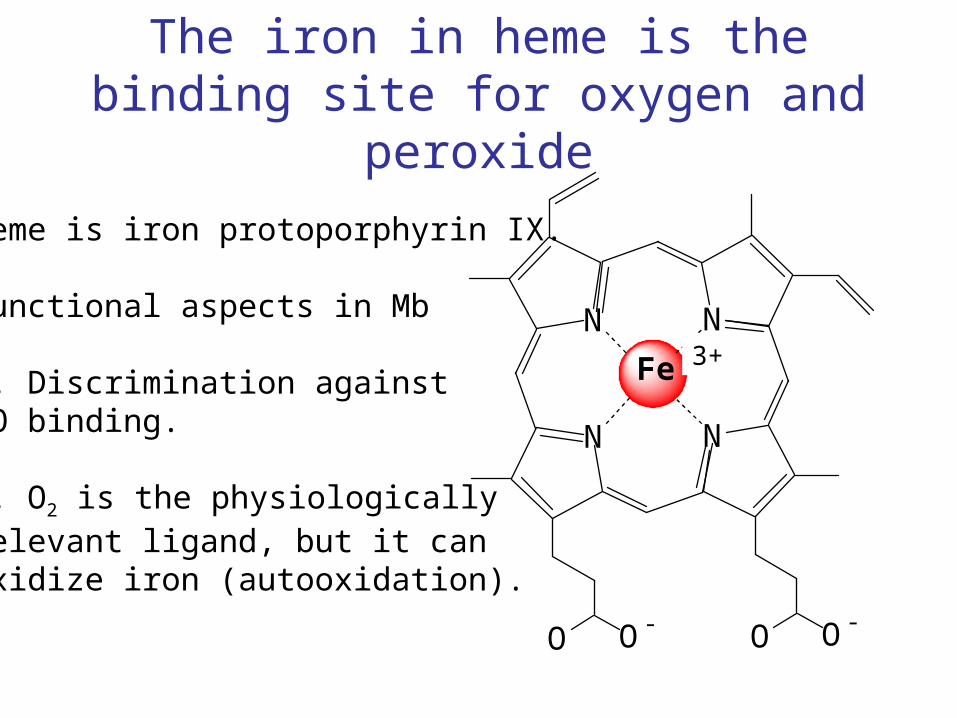
N N
NN
O O-O O-
Fe
The iron in heme is the binding site for oxygen and peroxide
Heme is iron protoporphyrin IX.
Functional aspects in Mb
1. Discrimination againstCO binding.
2. O2 is the physiologicallyrelevant ligand, but it canoxidize iron (autooxidation).
3+

Porphine orbitals
eg eg
a2u a1u

The four orbital model is used to represent the highest occupied and
lowest unoccupied MOs of porphyrins
eg
a1u a2u
The two highest occupiedorbitals (a1u,a2u) are nearly equal in energy. The eg orbitals are equal in energy.Transitions occur from:
a1u eg and a2u eg.
M1

The transitions from ground state orbitals
a1u and a2u to excited state * orbitals eg
can mix by configuration interaction
eg
a1u a2u
Two electronic transitionsare observed. One is verystrong (B or Soret) and the other is weak (Q).The transition moments are:MB = M1 + M2
MQ = M1 - M2 0
M1 M2
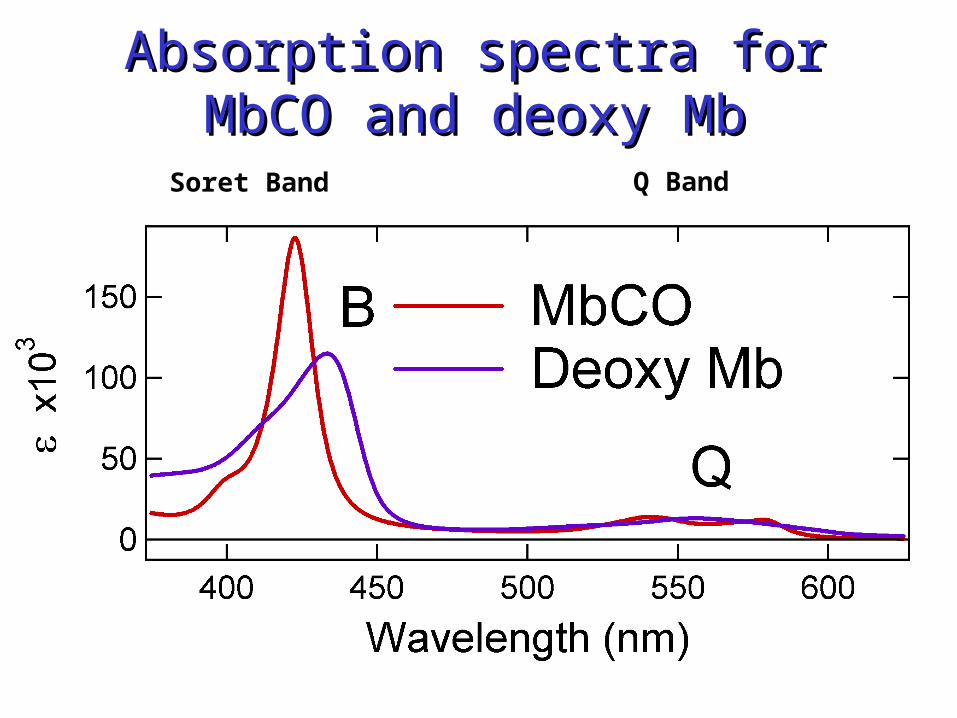
Absorption spectra for MbCO Absorption spectra for MbCO and deoxy Mband deoxy Mb
Soret Band Q Band

Resonance Raman spectrum Resonance Raman spectrum for excitation of heme Soret for excitation of heme Soret
bandband
Soret Band B Band Excitation Laser
Q Band
Raman spectrum

Soret (B) band Resonance Raman spectra of MbCO and Deoxy Mb
8

B band Resonance Raman spectra
of MbCO and Deoxy Mb

Hemoglobin
Time scale for the R-T switch
The trigger mechanism

The cooperative R - T switch
Hemoglobin is composed of two and two subunits whose structure s resemble myoglobin.
Eaton et al. Nature Struct. Biol. 1999, 6, 351
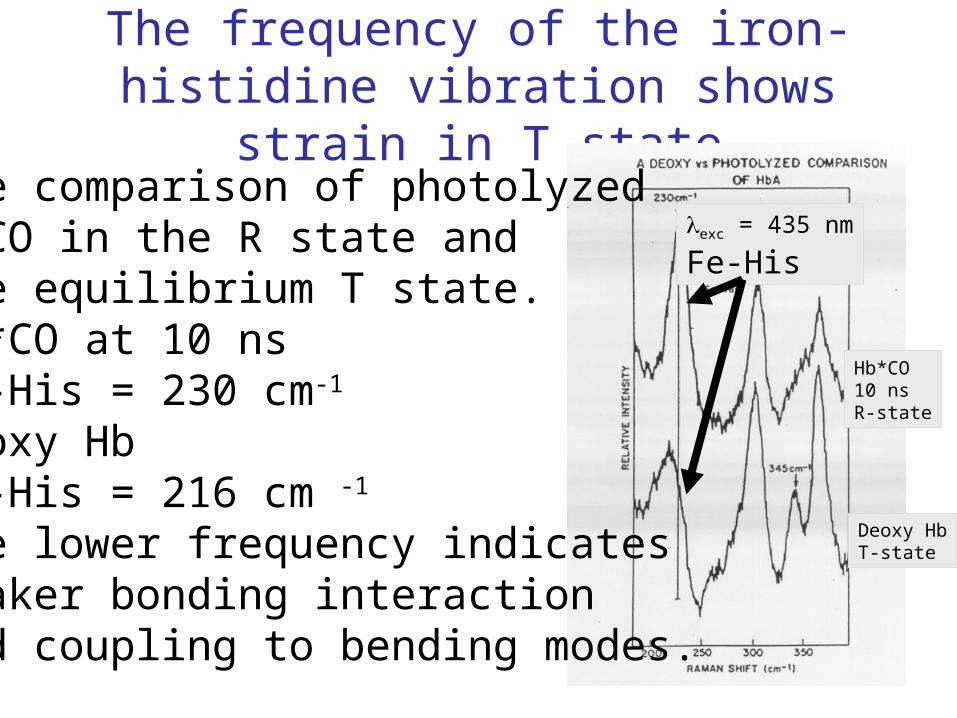
The frequency of the iron-histidine vibration shows strain in T state
The comparison of photolyzedHbCO in the R state andthe equilibrium T state.Hb*CO at 10 ns Fe-His = 230 cm-1
Deoxy HbFe-His = 216 cm -1
The lower frequency indicatesweaker bonding interactionand coupling to bending modes.
exc = 435 nm
Fe-His
Deoxy HbT-state
Hb*CO10 nsR-state
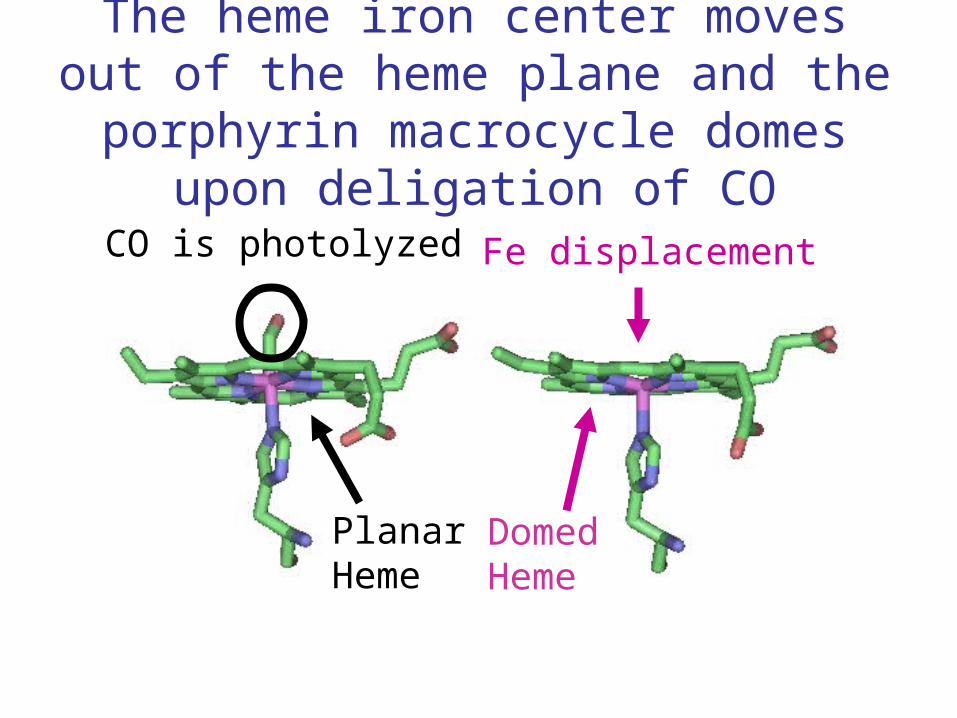
The heme iron center moves out of the heme plane and the porphyrin macrocycle
domes upon deligation of CO
CO is photolyzed Fe displacement
PlanarHeme
DomedHeme

The ligation of CO changes the spin state of the heme iron
dz2
dx2-y2
dxz,dyz
dxy
dz2
dx2-y2
dyz
dxz
dxy
Low spin Fe(II) High spin Fe(II)
S = 0 S = 2
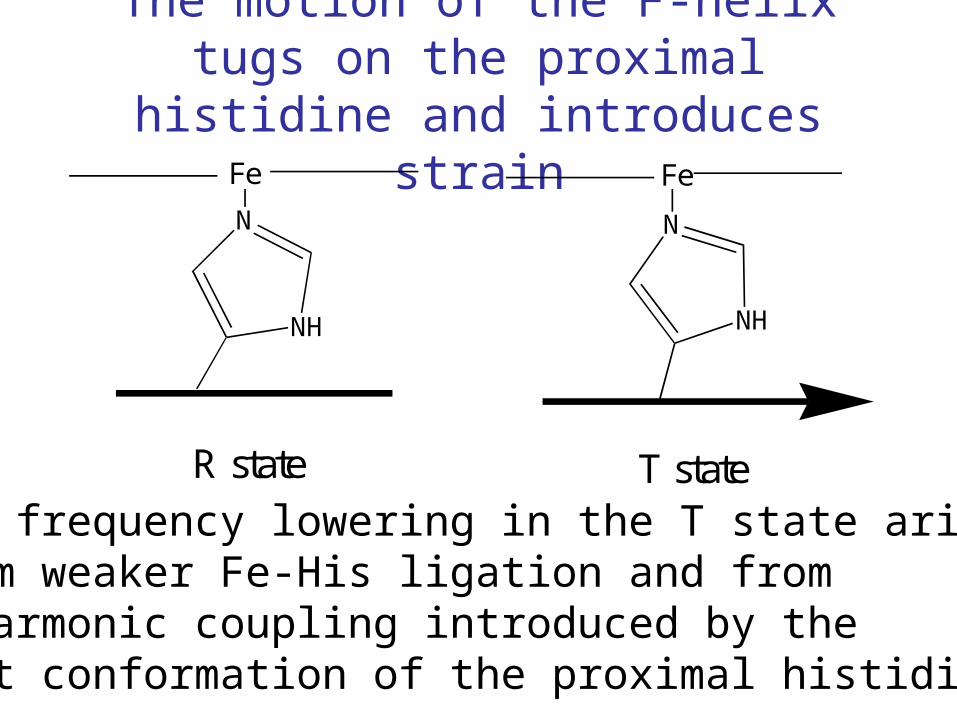
The motion of the F-helix tugs on the proximal histidine and introduces strain
The frequency lowering in the T state arisesfrom weaker Fe-His ligation and from anharmonic coupling introduced by thebent conformation of the proximal histidine.
Fe
N
NH
Fe
N
NH
R state T state
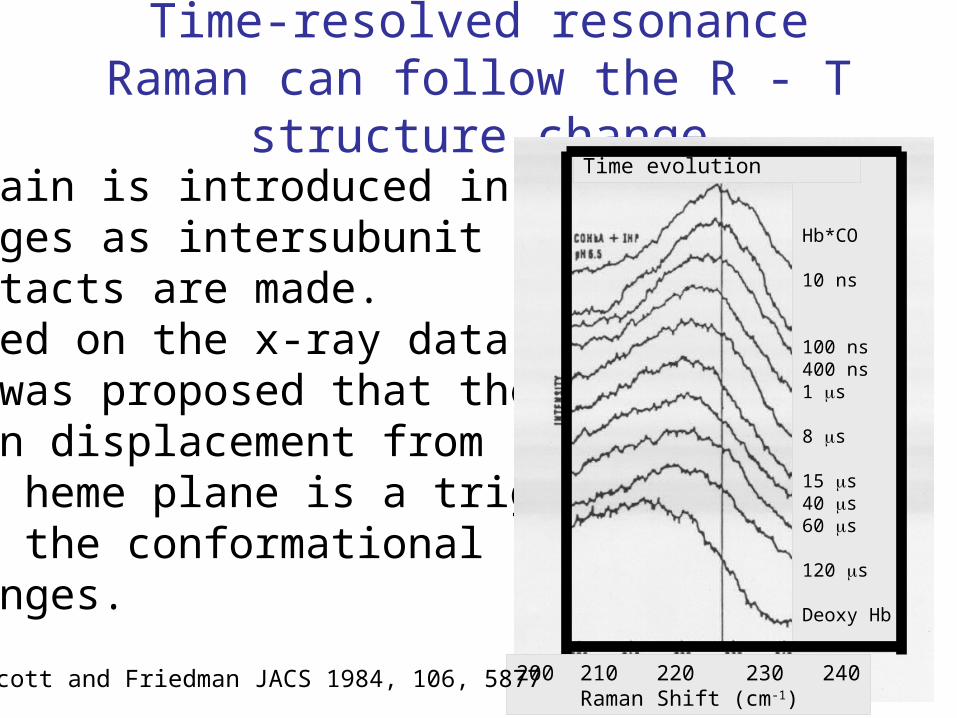
Time-resolved resonance Raman can follow the R - T structure change
Strain is introduced instages as intersubunitcontacts are made.Based on the x-ray datait was proposed that theiron displacement fromthe heme plane is a triggerfor the conformational changes.
Hb*CO
10 ns
100 ns400 ns1 s
8 s
15 s40 s60 s
120 s
Deoxy Hb
Time evolution
200 210 220 230 240 Raman Shift (cm-1)
Scott and Friedman JACS 1984, 106, 5877

Ultrafast resonance Raman spectroscopy shows that heme doming occurs in 1 ps
Equilibrium HbCO
Difference spectra obtainedby subtraction of the redspectrum from spectra obtained at the timedelays shown.
The evidence suggests that heme iron displacement is an ultrafast process that is independent of viscosity.
Franzen and Martin Nature Structural Biology 1994, 1, 230

Dehaloperoxidase: The First Enzymatically
Active Globin
NC State University

DHP oxidizes tribromophenol
DHP + TBP + H2O2
DHP + DBQ + H2O

Many Peroxidases belong to the Cytochrome c Peroxidase
family PDB: 1A2FCytochrome c Peroxidase (CCP)Class: All proteinsSuperfamily: Heme peroxidasesFamily: CCP-like Goodin and McCree
Scripps Institute
PDB: 2ATJHorseradish Peroxidase (HRP)Class: All proteinsSuperfamily: Heme peroxidasesFamily: CCP-like Hendrickson et al.
Biochemistry (1998)37, 8054

Dehaloperoxidase is a peroxidase that belongs to
the globin familyPDB: 1A6GMyoglobin (Mb)Class: All proteinsSuperfamily: Globin-likeFamily: Globins
Vojetchovsky,Berendzen,Schlichting
PDB: 1EW6Dehaloperoxidase (DHP)Class: All proteinsSuperfamily: Globin-likeFamily: Globins
Lebioda et al. J.Biol.Chem. 27518712 (2000)
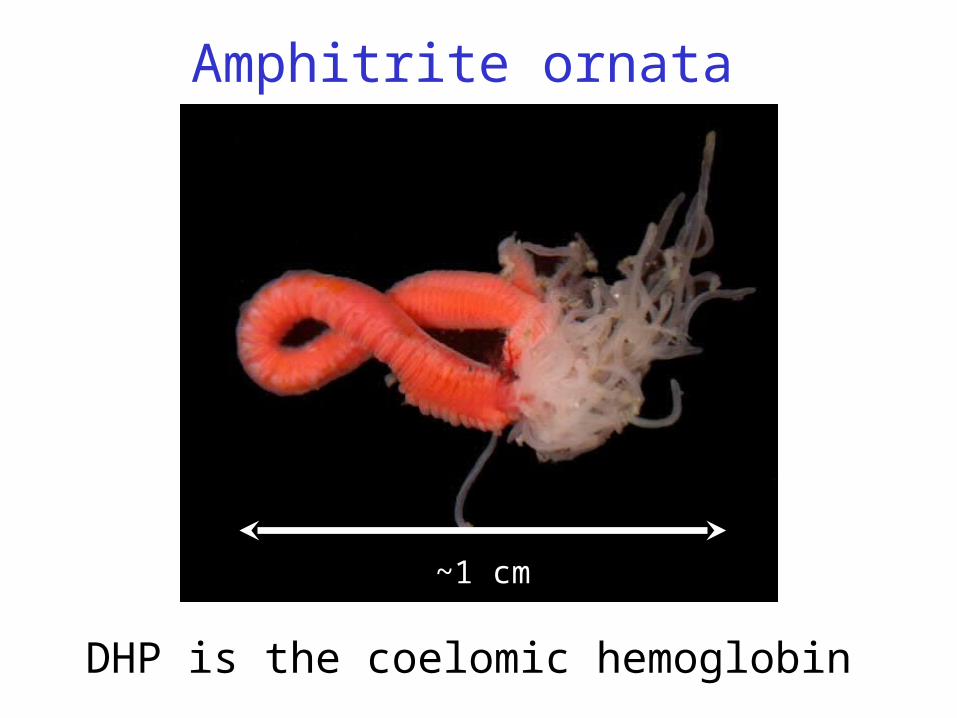
Amphitrite ornata
~1 cm
DHP is the coelomic hemoglobin

MbDHP
Comparison of DHP and Mb Structures
Superimpose hemes
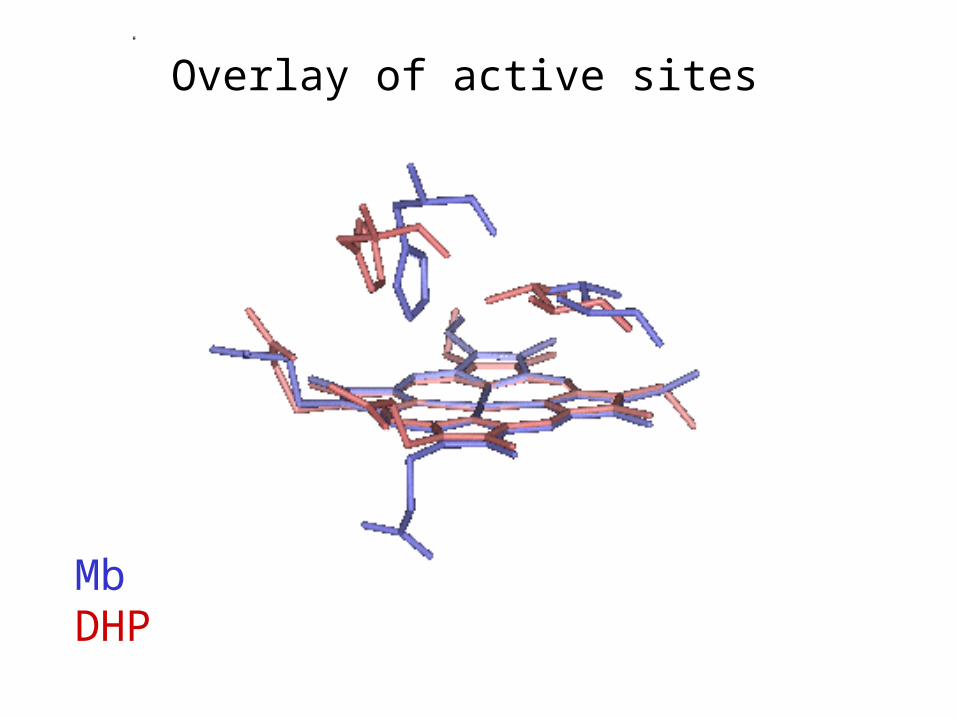
MbDHP
Overlay of active sites
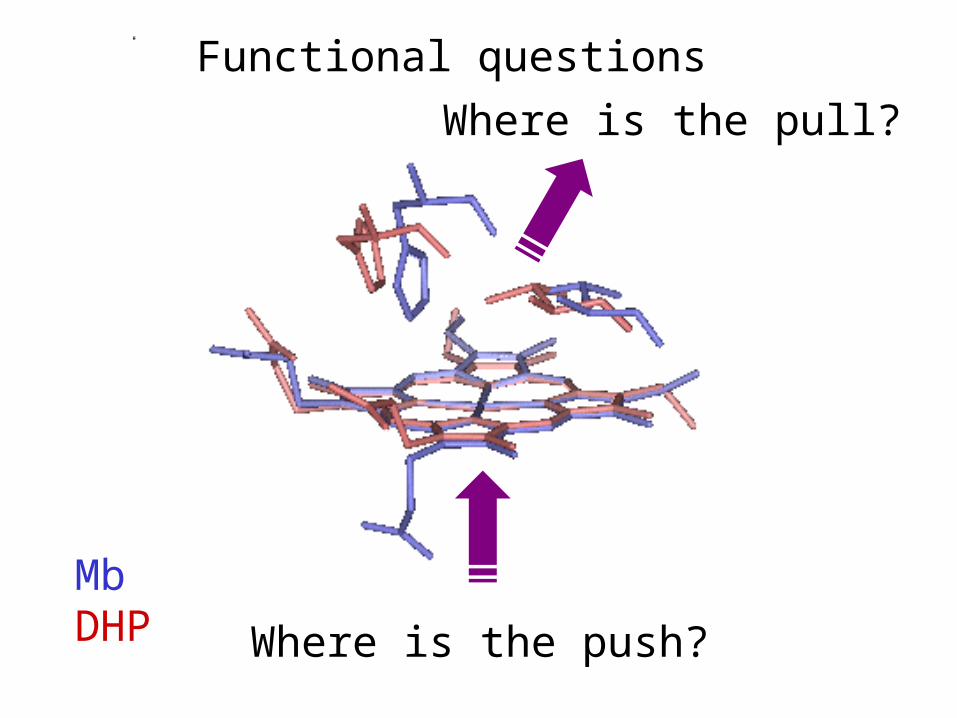
MbDHP
Functional questions
Where is the push?
Where is the pull?

Dehaloperoxidase looks like Mb,
but dehalogenates halophenols
OxoferrylCompound I
Ferric
Oxoferryl Compound II and phenol cation radical
P450
PeroxidaseHX + H2O
p-halophenol
Quinone
+
HX +
+.H2O
H2O2 H2O
HO XN
N
FeIV
N
N
N
N
O
N
N
FeIV
N
N
N
N
ON
N
N
N
FeIII
N
N
O O HO X
O O
HO X
X
X
X
X
X
X
Franzen et al., JACS (1998), 120, 4658-4661
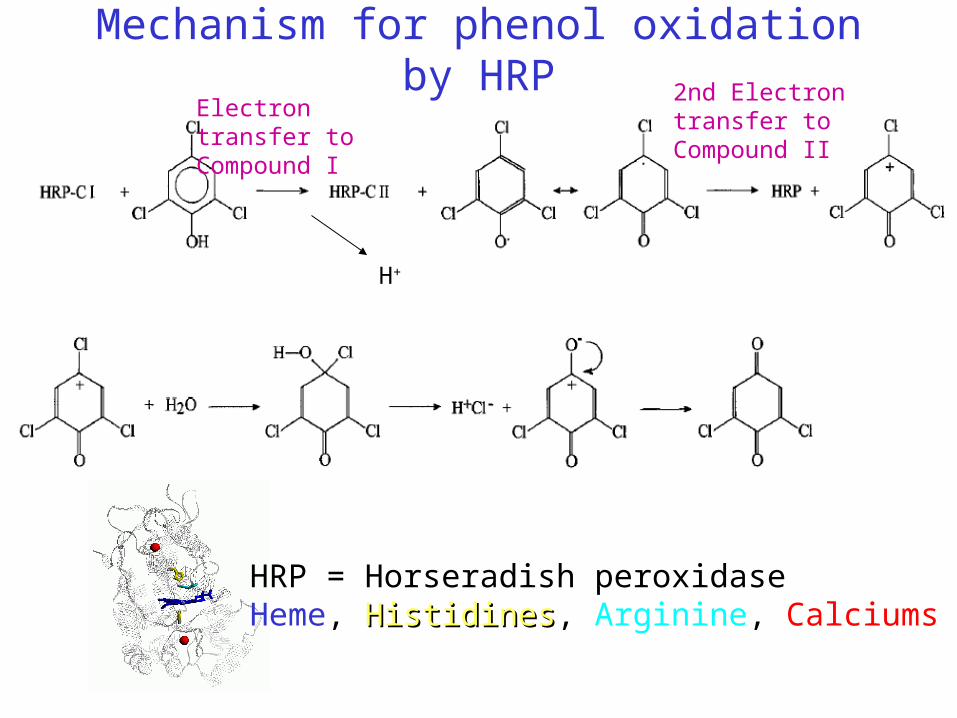
Mechanism for phenol oxidation by HRP
HRP = Horseradish peroxidaseHeme, HistidinesHistidines, Arginine, Calciums
Electron transfer to Compound I
2nd Electron transfer to Compound II
H+

X-ray structure of a substrate analog in the binding site of DHP
Lebioda et al., J.Biol.Chem. (2000) 275, 18712
4-iodophenolin internal siteUnprecedented in globins
N N
NN
O O-O O-
Fe
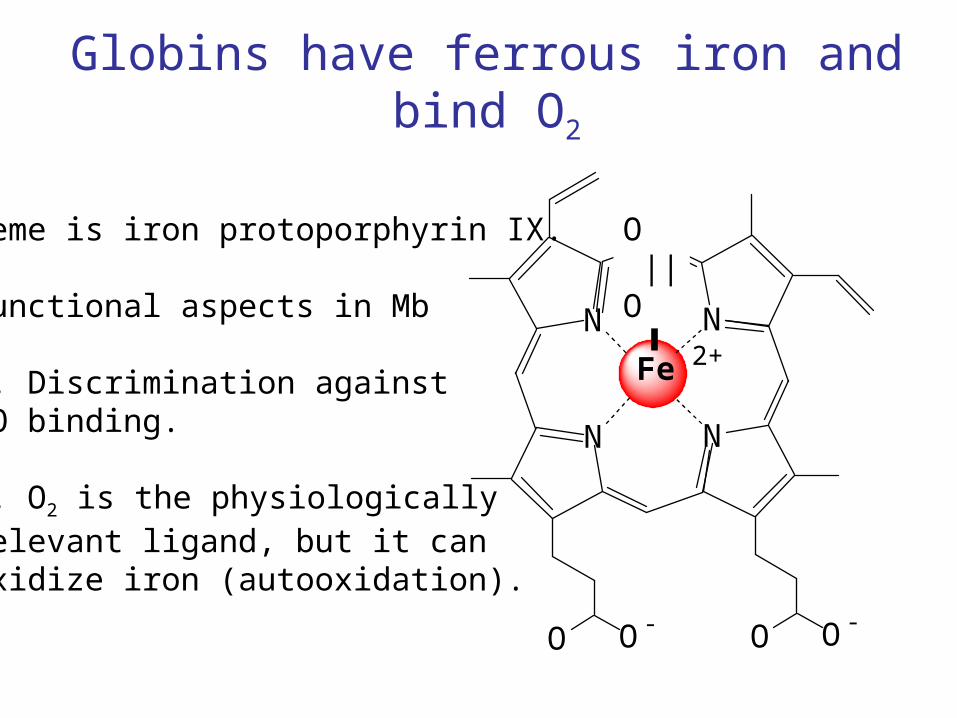
Globins have ferrous iron and bind O2
Heme is iron protoporphyrin IX.
Functional aspects in Mb
1. Discrimination againstCO binding.
2. O2 is the physiologicallyrelevant ligand, but it canoxidize iron (autooxidation).
O ||O
2+

N N
NN
O O-O O-
Fe
Peroxidases have ferric iron and bind H2O2
Heme is iron protoporphyrin IX.
Functional aspects in HRP
1. Activation involves formationof compounds I and II.
2. Edge electron transfer tosubstrate.
OH
/HO
3+
N N
NN
O O-O O-
Fe
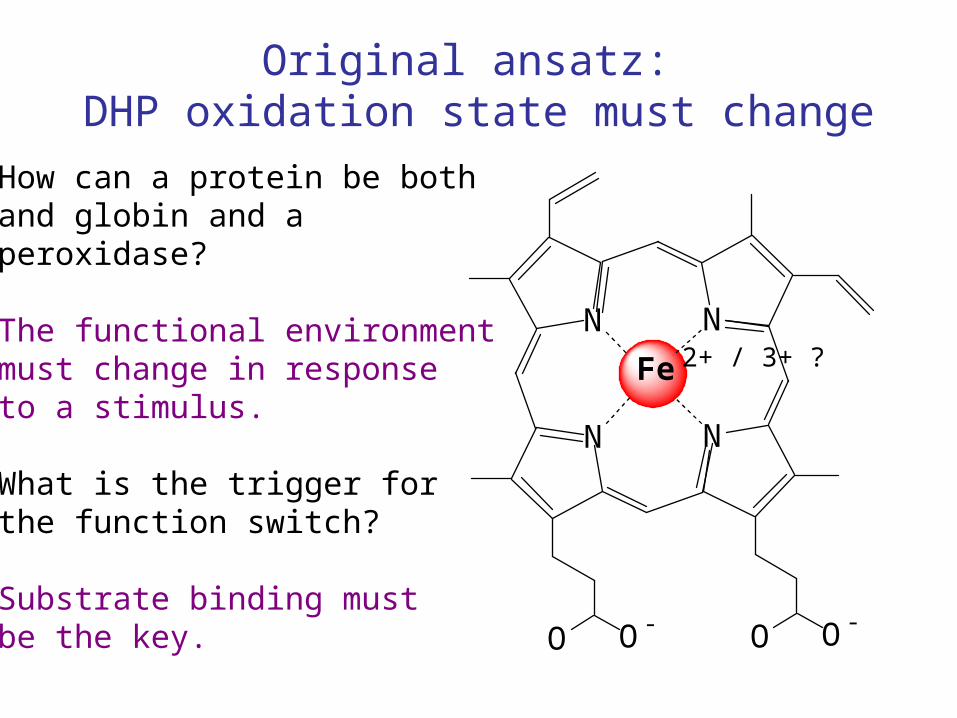
Original ansatz: DHP oxidation state must change
How can a protein be bothand globin and a peroxidase?
The functional environmentmust change in responseto a stimulus.
What is the trigger forthe function switch?
Substrate binding mustbe the key.
2+ / 3+ ?

Hardison, J. Exp. Biol. 1998, 102, 1099
Globins and Peroxidasesdiverged 1.8 billion years ago
Implicit meaning:Ancestral protein wasboth a hemoglobin anda peroxidase
Terrebellid polychaetesdo not figure in the scheme.
Convergent evolution?Divergent evolution?

Fe-histidine stretching mode of deoxy dehaloperoxidase
The frequency of the Fe-His mode is intermediate betweenthat of myoglobin (HHMb) and horseradish peroxidase (HRP).
Franzen et al., JACS (1998), 120, 4658-4661
Fe-His mode
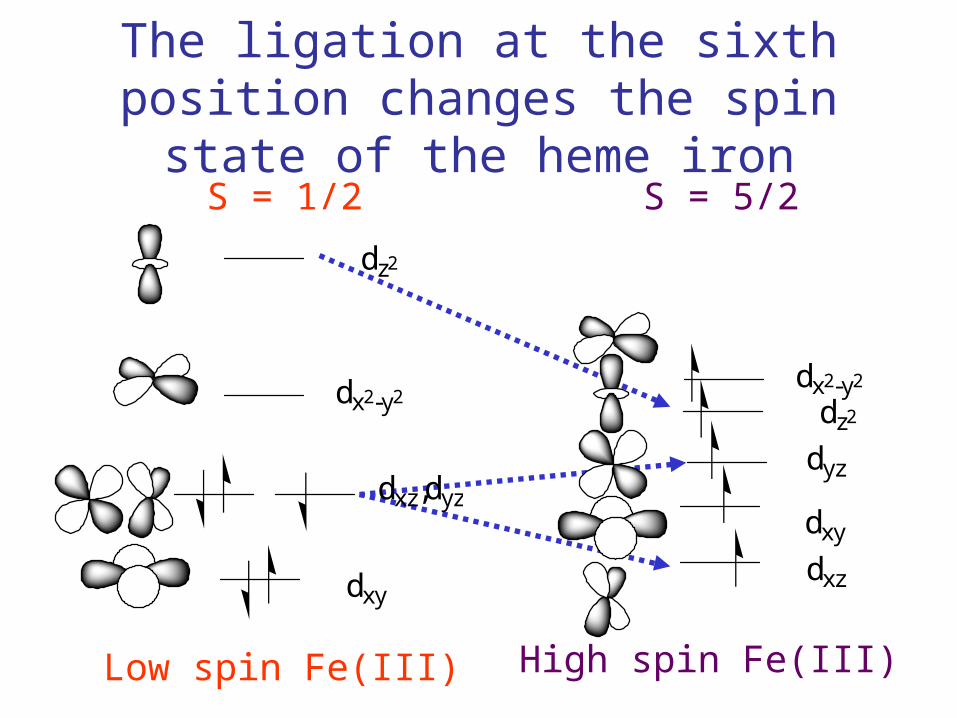
The ligation at the sixth position changes the spin state of the heme iron
Low spin Fe(III) High spin Fe(III)
S = 1/2 S = 5/2
dz2
dx2-y2
dxz,dyz
dxy
dz2
dx2-y2
dyz
dxz
dxy
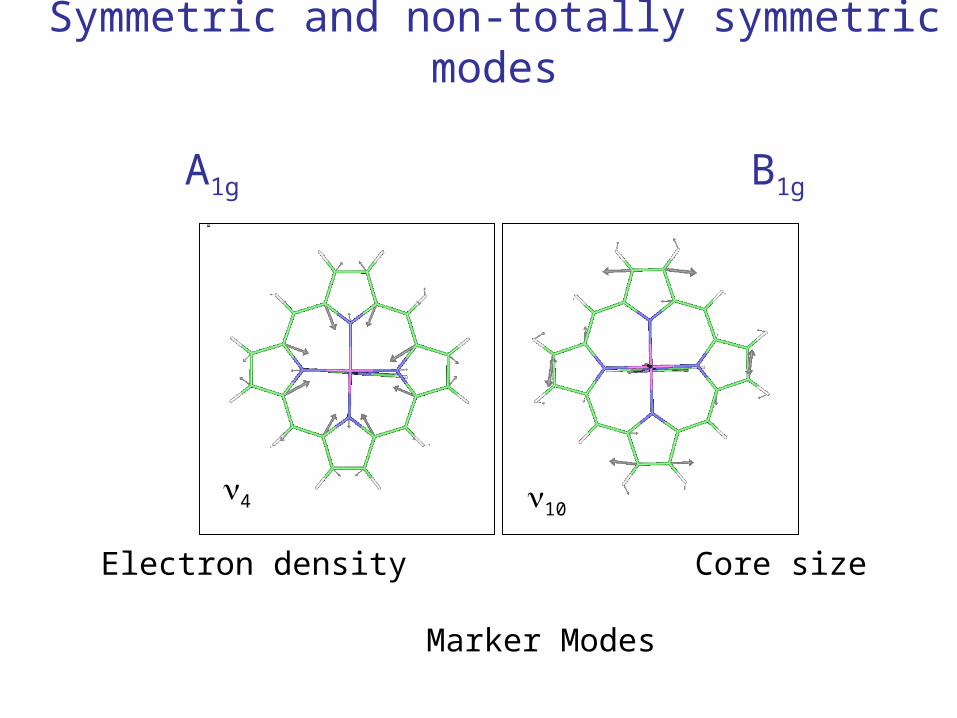
Symmetric and non-totally symmetric modes
A1g B1g
104
Electron density Core size Marker Modes

DHP core sizemarker modestudy
DHP is intermediate.It has more lowSpin character Than Mb, but less than HRP
Belyea, Franzen et al.Biochemistry, 2006
10234
high spin
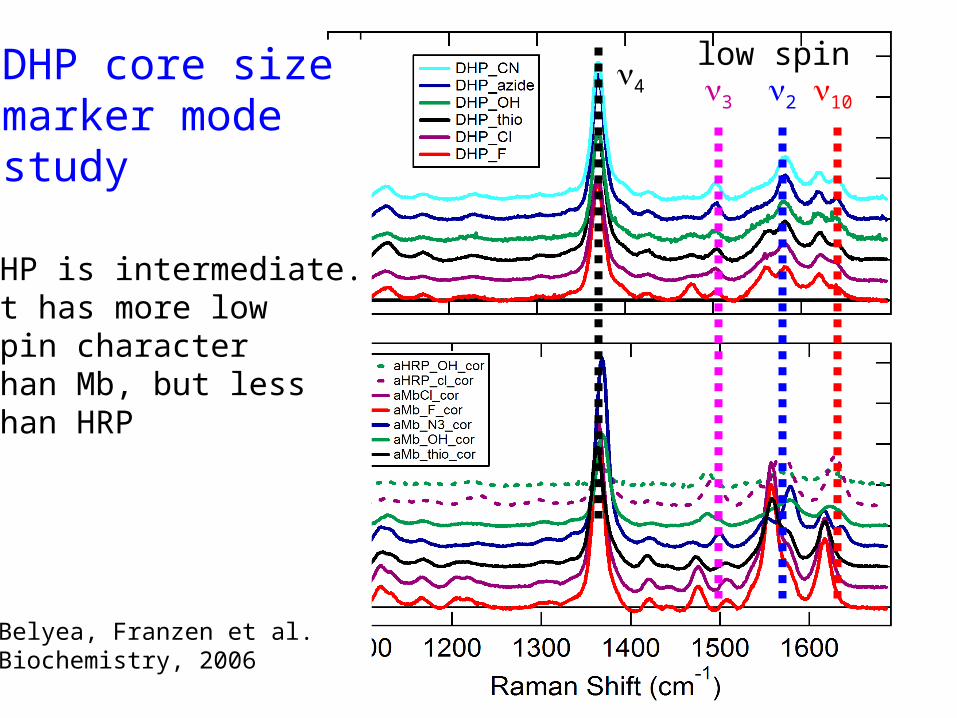
DHP core sizemarker modestudy
DHP is intermediate.It has more lowSpin character Than Mb, but less than HRP
Belyea, Franzen et al.Biochemistry, 2006
10234
low spin
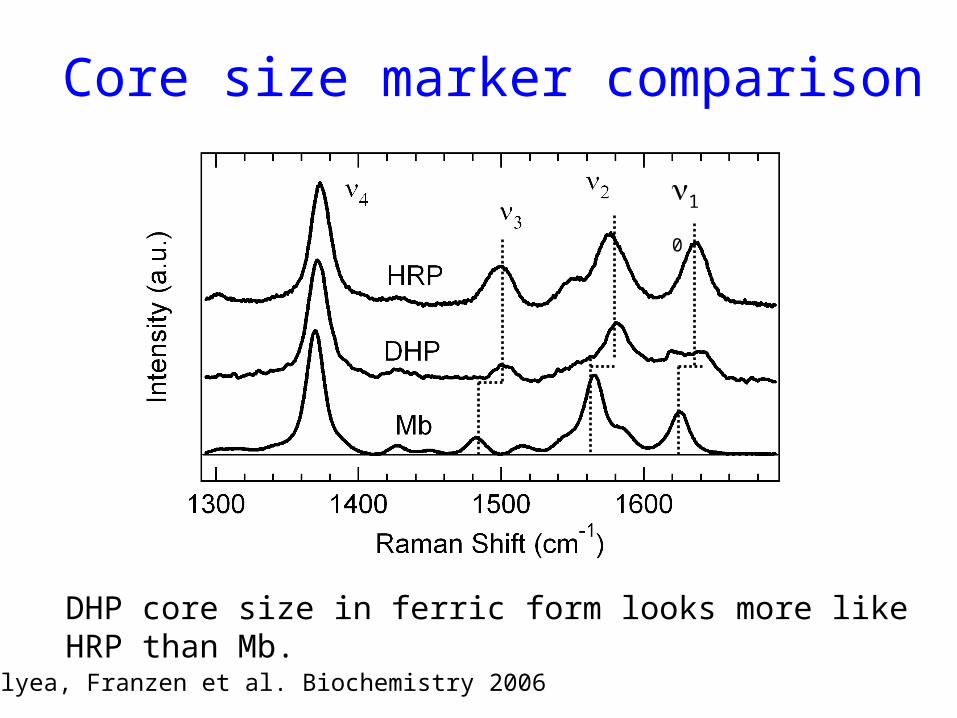
Core size marker comparison
DHP core size in ferric form looks more like HRP than Mb.
Belyea, Franzen et al. Biochemistry 2006
10
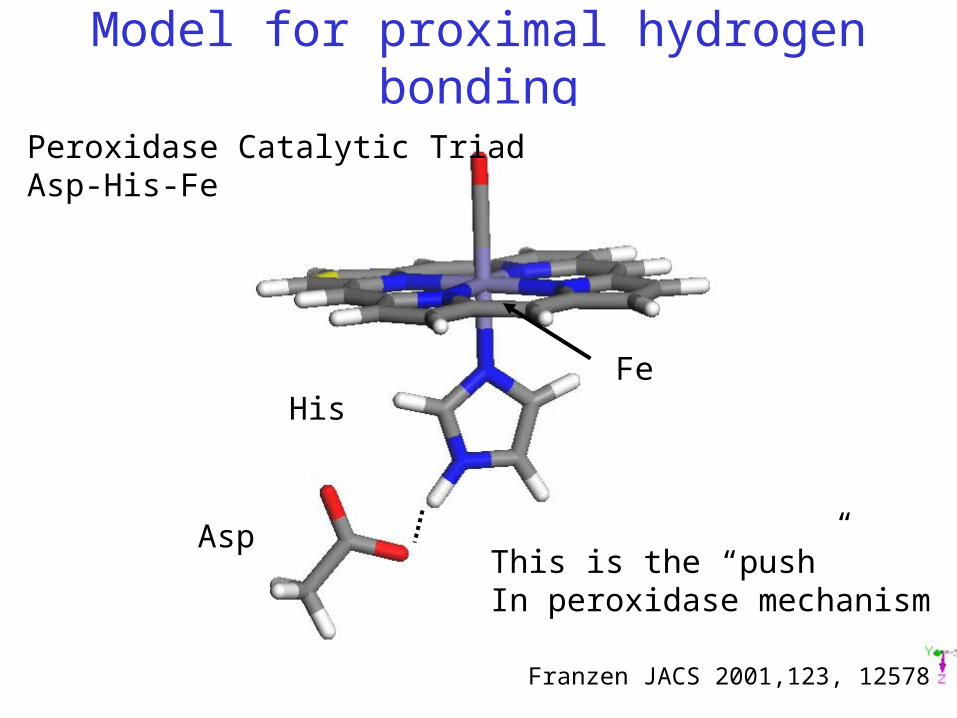
Model for proximal hydrogen bonding
Peroxidase Catalytic TriadAsp-His-Fe
Franzen JACS 2001,123, 12578
Asp
HisFe
This is the “push”In peroxidase mechanism

Model for proximal hydrogen bonding
Dehaloperoxidase Catalytic TriadC=O-His-Fe?
Franzen JACS 2001,123, 12578
Backbone C=O
HisFe
Hydrogen bond strengthis intermediate betweenMb and HRP/CcP.

















