HAL Id: hal-00929220 https://hal.archives-ouvertes.fr/hal-00929220 Submitted on 1 Jan 1990 HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci- entific research documents, whether they are pub- lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers. L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés. Purification and characterization of a proline iminopeptidase from Propionibacterium shermanii 13673 G Panon To cite this version: G Panon. Purification and characterization of a proline iminopeptidase from Propionibacterium sher- manii 13673. Le Lait, INRA Editions, 1990, 70 (5_6), pp.439-452. hal-00929220
Transcript
HAL Id: hal-00929220https://hal.archives-ouvertes.fr/hal-00929220
Submitted on 1 Jan 1990
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Purification and characterization of a prolineiminopeptidase from Propionibacterium shermanii 13673
G Panon
To cite this version:G Panon. Purification and characterization of a proline iminopeptidase from Propionibacterium sher-manii 13673. Le Lait, INRA Editions, 1990, 70 (5_6), pp.439-452. �hal-00929220�
Purification and characterizationof a proline iminopeptidase
from Propionibacterium shermanii 13673
G Panon .:" '.
Laboratoire de Recherche de Technologie Laitière, INRA.65, rue de Seint-Brieuc, 35042 Rennes Cedex, France
(Received 18 July 1990; accepted 17 8eptember 1990)
Summary - A proline lminopepndase activity was round in P shermanii 13673. The enzyme activitywas present in the intracellular fraction. The peptidase was purifisd approximately 80 times by ion-exchange chromatography on Fractogel T8K DEAE 650 and gel filtration on 8ephacryl 8-100 HR.The purified enzyme appeared as a single band after sodium dodecyl sulfate-polyacrylamide gelelectrophoresis. It had a molecular weight of 61 000. Optima for activity of the.purified enzyme were40 oC and pH 8.0. The enzymatic activity was inhibited by phenylmethylsulfonylfluoride (1 mM). Di-valent ions Zn, Co, Cu and Fe alsohad an inhibitor}t effect. The peptidase hydrolyzed specificallyPro p-NA amino acid, dipeptides Pro-Met, Pro-Phe, Pro-Le, Pro-lieu, Pro-G1yand 13-casomorphindes-Tyr fragment 7.
Résumé - Purification et caractérisation d'une praline iminopeptidase à partir de la souchePropionibacterium shermanii 13673. Une activité de proline iminopeptidase a été mise en évi-dence dans la souche P shermanii 13673. L'activité enzymatique est présente dans la fraction intra-cellulaire. L'enzyme a été purifiée environ 80 fois par chromatographie d'échange d'ions sur fracto-gel TSK DEAE 650 et gel filtration sur Sephacryl S-100 HR. L'enzyme purifiée présente une seulebande après électrophorèse en gel de polyacrylamide et en présence de SDS. 'Elle a un poids mo-léculaire de 61 000. La température est de 40°C et le pH optimal de 8,0. L'activité enzymatique estinhibée par le phénylméthylfulfonyl f1uoride (1 mM). Les ions divalents Zn, Co, Cu et Fe ont aussi uneffet inhibiteur. La peptidase hydrolyse spécifiquement la Pro p-NA, les dipeptides Pro-Met, Pro-Phe, Pro-Leu, Pro-Ile, Pro-Gly et le fragment 7 des- Tyr de la {J-easomorphine. .
Proteolysis is probably the most importantbiochemical event during the ripening ofmost cheese varieties with a major impacton flavor and texture. The mechanisms offlavor compound formation in hard cheesesinvolve successive steps and severalagents: rennet, indigenous milk protein-ases, starter proteinases and peptidases,enzymes from secondary starters andfrom non starter microorganisms. Thermo-philic acid bacteria such as Streptoeoccusthermophilus, Laetobaeillus helvetieus andLaetobaeillus laetis are normally used asstarter bacteria in Swiss cheese industries,whereas propionic acid bacteria are addedas secondary starters. The initial lactic fer-mentation by the starters is followed byfermention of lactate and residual sugarsdue to presence of propionic acid bacteria.This is a vital stage in Swiss cheese ripen-ing.
Enzyme activities and proteolysis dur-ing cheese maturation have been re-viewed by Fox (1989) and by Kamaly andMarth (1989). Purification of peptidasesfrom starter bacteria has been reported byRabier and Desmazeaud (1973), Desmaz-eaud and Juge (1976), Eggimann andBachmann (1980), Desmazeaud (1982),El Soda and Demazeaud (1982), Ezzat etal (1982), Hickey et al (1983), Casey andMeyer (1985), Meyer and Jordi (1987) andKhalid and Marth (1990).
Proteolytic system from secondarystarters has also been studied by Langs-rud (1974). Four or 5 peptidases in a strainof Propionibaeterium shermanii have beendetected, one of which was a proline imino-peptidase. Langsrud et al (1977) foundthat propionibacteria released largeamounts of proline when grown in mediacontaining peptides. Langsrud et al(1978b) established that propionibacteriaalso produced intracellular peptidases
G Panon
which were released by autolysis in hardcheeses. Sahlstrom et al (1989) foundpeptidase activities in cell wall, membraneand intracellular fractions of P shermanii.Perez Chaia et al (1990), studying the ac-tivity of peptidases belonging to propioni-bacteria, found a greater affinity for prolinep-nitroanilide than for leucine p-nitroanilide.
ln this work we have purified (with goodreasons for presuming homogeneity) andcharacterized a proline iminopeptidasefrom P shermanii 13673. This is the first re-port describing the purification and charac-terization of a proline iminopeptidase froma cell extract of propionic acid bacteria.
MATE RIAL AND METHODS
Chemicals
Lysozyme, mutanolysin, DNase, RNase, ail pep-tide derivatives were obtained from SigmaChemical Co, St Louis, MO, USA. FractogelTSK DEAE 650 was purchased from E Merck,Darmstadt, Germany. Sephacryl S100 HR andmolecular weight markers were obtained fromPharmacia-LKB, Saint-Quentin en Yvelines,France.
Bacterial strains
Five strains from our collection were studied.The strain of P g/obosum 408 was supplied by astarter bacteria producer, Standa-Industrie,Caen, France. The strain of P freudenreichiiCNRZ 435 was supplied from the culture collec-tion of the Centre de Recherches (CNRZ), Jouy-en-Josas, France. P shermanii 13673 and the 2strains of P acidi propionici 4875 and P01 wereobtained from one private Emmental industry.
Ali strains were identified according to Ber-gey's Manua/ of Determinative Bacteri%gy(More and Holdeman, 1974) and the method ofBritz and Steyn (1980) and the method of Britz
Proline iminopeptidase from P shermanii 441
and Steyn (1980) and also by control of propion-ic acid production (HPLC method).
Growth conditions
The strains were maintained on yeast extractlactate (VEL) medium (Hettinga et al, 1968): so-dium lactate 1% (Merck), K2HP04 0.025% (Pro-labo), MgS04 0.005% (Prolabo), biotrypticase1% (BioMérieux), yeast extract 1% (Biokar); pHwas ajusted to 7.0 and medium autoclaved for15 min at 121°C. Strains were transferred 3times in 10-ml tubes. Cultures of 2 1 were grownin VEL medium with an inoculum of 1%. Growthwas maintained at 30°C for 48 h.
Preparation of cell extracts
At the end of the exponential growth period theceUswere harvested and centrifuged at 10 0009 for 15 min. The supernatants were stored at-20 "C, The cell pellets were washed twice in200 ml of 50 mM 13glycerophosphate, pH 7.0.
Washed cells were then treated by protopiastbuffer: 100 ml of 50 mMll Tris buffer, pH 7.0 con-taining 0.5 M saccharose, lysozyme at 5 mg/mland mutanolysin at 5 Ilg!ml (Kondo and McKay,1982). After incubation at 37°C for 2 h the proto-plasts were recovered by centrifugation at 5 5009 for 20 min. Protoplast supernatants werestored at -20 oC after filtration. CeIls were dis-rupted by osmotic shock in alkaline buffer (pH12). pH was immediately adjusted to 7.0 with 2M Tris buffer.
After DNase and RNase treatment (0.1 mg!ml) the crude extract was then centrifuged at24000 9 for 30 min at 4°C in order to eliminateintact ceIls and cell debris. The cell-free extractsdesignated as the cytoplasmic or intracellularfractions were stored at -20°C, before amine-peptidase activity investigations. The intracellu-lar fraction of P shermanii 13673 was used asstarting material for the purification of the pepti-dase.
Substrate specificity
Intracellular fractions were tested with L-Ieucinep-nitroanilide (Leu p-NA), L-Iysine p-nitroanilide
(Lys p-NA) , L-Proline p-nitroanilide (Pro p-NA)and L-glycine p-nitroanilide (Gly p-NA) sub-strates. Protein extracts from purification wereroutinely tested with Pro p-NA substrate.
Aminopeptidase activity was assayed by in-cubation of intracellular preparation With 0.75mM p-nitroanilide amino acids in 20 mM Tris-HCI buffer pH 7.0. Incubation was carried out at37°C for 1 h. The reaction was stopped by theaddition of 30% acetic acid. The supernatantwas separated by centrifugation at 10 000 9 for5 min. The release of p-nitroanilide was followedby measuring absorbance at 410 nm (A410) inan Uvikon Kontron 860 spectrophotometer. Oneunit of aminopeptidase activity was defined asthe amount of enzyme which produced a vari-ation of 0.01 unit of A410 per min at 37 oC. Thepurified enzyme was also tested against severaldi- and tripeptides. Purified enzyme was incuba-ted with 0.4 mM substrate in phosphate bufferpH 7.0. After incubation at 37°C for 1 h, free (X
amino groups were determined with 1 vol of nin-hydrin (20 9 of ninhydrin, 750 ml of methylcello-solve (ethylene glycol monomethyl ether), 250ml of 4 M acetate buffer (pH 5.51), 7 ml of titani·um chloride). The preparation was boiled at100°C for 5 min and 3 vol of 50% ethanol wereadded. Hydrolysis was followed at A44Qfor pro-line, and A570 for the other amino acids.
Substrate specificity of the peptidase wasalso tested by HPLC against bovine 13-casomorphin fragments: fragment 1-3, morpho-ceptin (13-casomorphin1-4 amide), fragment 1-5 and des-Tyr-fragment 7. The purified enzymewas incubated with 0.4 mM substrate in 20 mMTris buffer pH 7.0 for 3 h at 40°C. The reactionmixtures and substrates were injected onto aVydac column C 18 5 Il. Peptides were elutedby a gradient water/acetonitrile (1 vol/4 vol).
Purification
First ion exchange chromatography
Thè first ion exchange chromatography was per-formed with a Fractogel TSK DEAE 650 column.The column (2.6 cm x 40) was equilibrated with20 mM Tris-HCI buffer pH 7.0. One hundred mlof intracellular fraction of P shermanii 13673were dialyzed and centrifuged. This fraction(3 000 mg) was applied to the column. After
442
washing the column with the equilibration buffer,the enzyme was eluted with a linear gradient ofo to 0.6 M NaCI in the same buffer at a flow rateof 90 ml/h. Fractions of 11.2 ml each were col-lected and tested for protein concentration bymeasurinq absorbance at 280 nm (A280). Pepti-dase activity was tested with Pro p-NA sub-strate. Active fractions (Nos 36-45) were pooledand stored at -20 oC in the presence of stabiliz-ers aceordinq to the method of Neviani et al(1989).
Second ion exchange chromatography
The enzyme active fractions of the first per-formed chromatography were dialyzed against20 mM Tris-HCI pH 7.0 at 4 "O and applied tothe same Fractogel TSK OEAE column previ-ously equilibrated with 20 mM Tris-HCI bufferpH 7.0. The column was washed with 20 mMTris-HCI; the enzyme was eluted with a Iineargradient of NaCI 0-0.5 M in 20 mM Tris-HCI pH7.0 at a flow rate of 90 mlth, and 11.2-ml frac-tions collected. Active fractions (Nos 38-46)against Pro p-NA substrate were pooled and di-alyzed against 20 mM Tris-HCI buffer pH 7.0 at4°C. They were concentrated by PEG 3000 be-fore gel filtration.
Gel filtration
The concentrated enzyme (9 ml) was furtherpurified on a Sephacryl S-100 HR column (2.6cm x 100) previously equilibrated with 20 mMTris-HCI buffer pH 7.0 containing 1 M NaCI. Atlow rate of 120 mlth was applied and 8-ml frac-tions were collected from the column. The en-zyme was eluted from the column as a singlepeak. Active fractions (Nos 11-15) against Prop-NA substrate were pooled and dialysedagainst 20 mM Tris-HCI buffer pH 7.0 at 4 oC.Characterization of the enzyme was performedwith this preparation ..
Polyacrylamide gel electrophoresis
Each purification step was examined by sodiumdodecyl sultate--polyacrylarnide gel electrophor-esis (SDS-PAGE) with the Laemmli buffer sys-tem (Laemmli',1970). A 3% stacking gel and a
G Panon
10% running gel were prepared. The proteinsampi es were mixed 1/1 with sample buffer(10 mM Tris-HCI pH 6.8, 20% glycerol, 2%SOS, 5% 2-mercapto-ethanol and 0.001% bro-mophenol bJue), boiled for 5 min, and applied tothe gel. After a s-h run at 150 V, proteins werestained with Coomassie blue R. For destaining,the gels were put into a solution of 10% (v/v)acetic acid, 10% (vtv) methanol in water.
Determination of molecular weight
Molecular size of the active enzyme was esti-mated by gel filtration on Sephacryl S-100 HR incolumn 2.6 x 100. The column was first calibra-ted with bovine serum aJbumin (67 kDa), ovalbu-min (43 kOa) and lysozyme (14.3 kDa). Molecu-Jar size was also estimated by SOS-PAGE. Thelow molecuJar weight standards used were ob-tained from Pharmacia (Uppsala, Sweden):phosphorylase b (94 kDa), bovine serum albu-min (67 kOa), ovalbumin (43 kOa), bovine car-bonic anhydrase (30 kDa), soybean trypsin in-hibitor (20.1 kOa) and u-lactaloumin (14.4 kOa).The molecular weight of the aminopeptidasewas determined by a reference to a standardgraph relating log of molecular weight of eachstandard to its relative mobility.
Effect of reducing agents,meta/ che/atorsand inhibitors on enzyme activity
A diverse range of compounds known to inhibitvarious proteases were tested for their effectson the peptidase activity of P shermanii 13673.Reducing agents and thiol, metallo, serine pro-tease inhibitors were tested. The enzyme waspreincubated in 20 mM Tris-HCI pH 7.0 for 15min at 20 oC with the following agents: dithio-threitol, L-cysteine, 2-merceptoethanol, N-ethylmaleimide, iodoacetic acid, iodoacetamide,p-chloromercuric benzoic acid, ethylenediamine-tetraacetic acid (EOTA), 1-10 phenanthroline,bestatin, phosphoramidon, phenylmethylsulfonylf1uoride, pepstatin A and sodium citrate. Two fi-nal concentrations were used: 0.1 mM and1 mM (1 and 10 mM for EDTA). Pro p-NA wasadded as substrate and enzyme activity wasmeasured after incubation at 37 oC for 60 min at
A410•
Proline iminopeptidase from P shermanii 443
Effect of metal ions on enzyme activity
Effects of divalent ions were tested on enzymeactivity. The purified and dialyzed enzyme wasincubated in the presence or the absence of Cachloride, Co acetate, Cu sulfate, Zn, Mn sulfateand Mg chloride at a final concentration of 0.1mM. Reaction mixtures were preincubated for15 min at 20 oC in 20 mM Tris-HCI buffer (pH7.0) prior to the addition of Pro p-NA. The en-zyme activity was determined at 37°C for 60 minatA410•
Protein estimation
The BioRad method (BioRad Laboratories, Wat-ford, England) was used with bovine serum al-bumin as standard (Bradford, 1976). Elution pro-files of the proteins separated after FractogelTSK DEAE 650 and Sephacryl S-100 HR weredetermined at A2BO•
Effect of pH on enzyme activity
The effect of pH from 4.0 to 10.0 on peptidaseactivity was determined by using 20 mM gly-cine-NaCI or Tris-HCI buffer adjusted to the ap-propriate pH values. Enzyme activity was meas-ured at 37°C for 60 min with Pro p-NA assubstrate at A410.
Effect of temperatureon enzyme activity
The effect of temperature was measured in therange of 5-70°C. The 20 mM Tris-HCI buffer(pH 7.5) was equilibrated for 15 min at the testtemperature. Fifty lil of purified enzyme and 50lil of Pro p-NA as substrate were added and theenzyme activity was measured atter 60 min ofincubation at the test temperatures at A410•
ids were quantitatively assayed with an aminoacid analyzer Pharmacia-LKB alpha plus.
RESULTS
Peptidase purification
Enzymatic treatrnent of propionibacterialcells was preferred to physical treatmentbecause it does not generate high shearstresses and minimises product damage(Andrews and Asenjo, 1987). Peptidaseactivity was observed in the cell wall frac-tion (data not shown), probably due to par-tial cell Iysis during protoplast formation.
The major peptidase activities with leu-cine, lysine, proline and glycine p-nitroanilide substrates were present in thesoluble intracellular fraction of 5 strains ofpropionibacteria (table 1). For ail extracts
Table 1. Hydrolysis of p-nitroanilide (P-NA) sub-stituted amino acids (leucine, lysine, proline, gly-cine) by intracellular fraction of P globosum 408,P freudenreichii CNRZ 435, P shermanii 13673,P acidi propionici 4875, P acidi propionici P01.Results are expressed in units of activity per minat 37°C.Hydrolyse des acides aminés (leucine, lysine,proline, glycine) substitués (p-NA) par les frac-tions intracellulaires de P globosum 408, P freu-denreichii CNRZ 435, P shermanii 13673,P acidi propionici 4875, P acidi propionici P01.Les résultats sont exprimés en unités d'activitépar min à 37 oC.
SubstrateLeu Lys .Pro Glyp-NA p-NA p-NA p-NA
P globosum 408 75 0 180 75Amino acid analysis P freudenreichii Z 435 150 0 750 0
P shermanii 13673 180 150 2660 0
The purified enzyme was hydrolyzed with 6 NP acidi propionici 4875 100 0 500 0
HCI under vacuum at 110°C for 24 h. Amino ac-P acidi propionici P01 200 0 600 0
444
we observed a stronger activity with Pro p-NA than with Leu p-NA substrate.
After this result we decided ta studyP shermenli 13673 more particularly andta purify the specific aminopeptidase. Inthe first ion exchange chromatographyperformed on Fractogel T8K DEAE 650the praline activity was eluted from the col-umn at 0.4 M NaCI (fig 1). During the sec-ond ion exchange chromatography on thesame Fractogel T8K DEAE 650, the en-zyme was eluted at 0.4 M NaCI.
After gel filtration on 8ephacryl 8-1 00HR the enzyme was eluted from the col-umn as a single and symmetrical peak (fig2). Results of the purification are summa-rized in table II. Using this protocol the pra-line iminopeptidase was purified '" 80-foldwith a yield of 7% from the crude extract.
1.280 P1,8 280 EP
200 T11.2 D
180AS
0.8 E
100 AC'
0.4 T1
V1T0 Y
0 10 20 30 40 80 80FRACTION NUM8ER
lU/mil-- PEPTIDA8E ACTIVITY - NICI 0.2M ID O.SM
..... A28D (nml
Fig 1. First ion exchange chromatography. Puri-fication of aminopeptidase from intracellular frac-tion of P shermanii 13673. Elution pattern of thepeptidase from a Fractogel T8K DEAE 650 co-lumn. The peptidase activity Â. (A410) was as-sayed with Pro p-NA substrate.Première chromatographie d'échange d'ions.Purification de /'aminopeptidase à partir de lafraction intracellulaire de P shermanii 13673.Profil d'élution de la peptidase de la colonne deFractogel T5K DEAE 650. L'activité de la pepti-dase Â. (A41OJ a été testée avec le substrat Prop-NA.
G Panon
Peptidase characterization
Molecular weight determination
The enzyme preparation showed a singleband after 8D8-PAGE. The molecularweight of the enzyme was estimated at61 000 even in the presence of 2-mercaptoethanol (data not shawn) (fig 3).The peptidase was also found to have amolecular weight of 61 000 by gel filtration.
Effect of pH and temperatureon enzyme activity
The optimal pH for Pro p-NA hydrolysingactivity appeared ta be pH 8.0 (fig 4A). AtpH 5.0 and 9.0, hydrolysing activity wasalso detected. The optimal temperature for
0,2 POOL 800.... PEP
0,15T1DA
A S2 0.1 E80 A
CT
0,05 1V1TY
00 20 40 80 80
FRACTION NUM8ER lU/mil
-&- PEPTIDASE ACTIVITY .........A280
Fig 2. Gel filtration on Sephacryl 8-100 HA. Pu-rification of peptidase from P shermanii 13673after the second ion exchange chromatographyon a 8ephacryl 8-100 HR column. The pepti-dase activity Â. (A410) was assayed with Pro p-NA substrate.Gel filtration sur 5ephacryl 5-100 HR. Purifica-tion de la peptidase de P shermanii 13673aprèsla seconde chromatographie d'échange d'ionssur colonne de 5ephacryl 5-100 HR. L'activitéde la peptidase Â. (A41oJ a été testée avec lesubstrat Pro p-NA.
the substrate. This specifie hydrolysis wascontinuous with time (data not shown). Foridentification of released products we in-jected proline as a marker. Under our con-ditions; Pro was not detectable. It is Iikelythat the peak which appeared was thePhe-Pro-Gly-Pro-lle peptide. Morever, thiscould be confirmed by identification of thispeptide by amino acid analysis.
Proline iminopeptidase from P shermanii 445
Table Il. Purification of the peptidase from P shermanii 13673.Purification de la peptidase de P shermanii 13673
Purification Vol Total Total' Specifie Yield Purificationstep protein activity activity
, Determined with the substrate Pro p-NA; , Déterminé avec le substrat Pro p-NA
Pro p-NA hydrolysing activity was found tobe 40 oC (fig 48). Significant activity wasdetected at 35 and 45°C; at 20°C, 50% ofspecifie activity was detected, but at 55°Conly 9% of the optimal activity was noted.
Substrate specificity
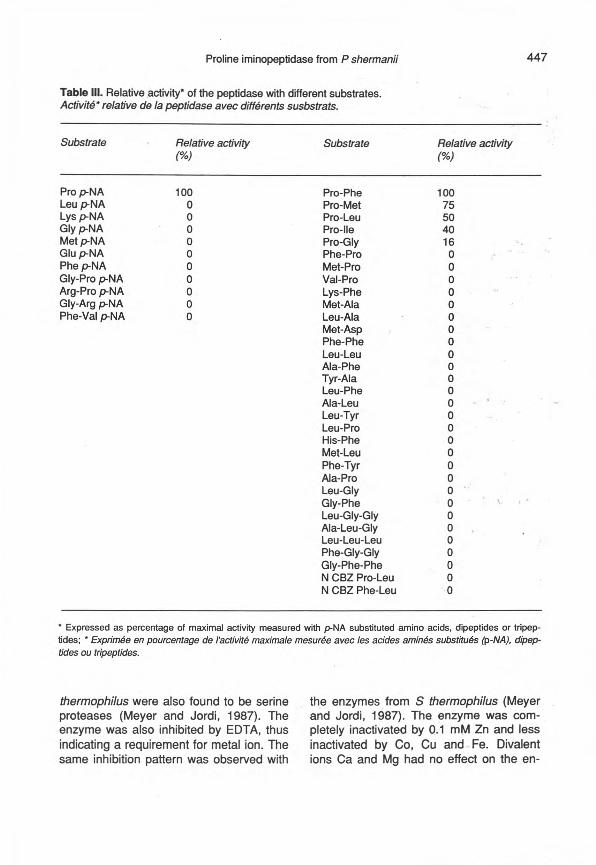
The purified enzyme was incubated with p-nitroanilide derivatives of amino acids ordipeptides, and with dipeptides and tri pep-tides (table III). Pro p-NA had the highestactivity among the p-NA substrates. Thepurified enzyme catalyzed hydrolysis of di-peptides only if they had a proline residuein the N-terminal position: Pro-Phe, Pro-Met, Pro-Leu, Pro-Ile, Pro-Gly. Carboben-zoxy peptides were not hydrolyzed by theenzyme. The purified peptidase had thesame high specificity for Pro p-NA as theintracellular fraction. The peptidase had noaction on B-cascmotphln fragments 1-3,1-4 and 1-5 (table IV). But the peptidasewas able to hydrolyze specifically the 13-casomorphin des-Tyr-fragment 7: Pro-Phe-Pro-G1y-Pro-lle. After 3 h of incubation at40°C the peptidase hydrolysed '" 50% of
Effect of chemical reagentsand inhibitors
Treatment of the peptidase with severalagents is summarized in table V. The rateof Pro p-NA hydrolysis in the absence ofany reducing agent, metal chelator or in-hibitor was taken as 100%. The enzymeactivity was reduced to 30% when incuba-ted in 1.0 mM phenylmethylsulfonyl fluo-ride (PMSF), a specifie inhibitor for serineprotease. The metal-complexing agentsEDTA (10 mM) and bestatin al50 de-creased enzyme activity, indicating a metalion requirement. Sulfhydryl inhibitors hadsorne effect on the proline iminopeptidaseactivity, whereas dithiothreitol and 2-
446
3
- 94 kDa
- 67 kDa
- 43 kOa
- 30 kDa
- 20.1 kDa
Fig 3. SD5-polyacrylamidegel electrophoresis.t, Low molecularweightstandards:fromtop tobottom, phosphorylaseb (94 kDa), bovineserum albumin(67 kDa), ovalbumin(43 kDa),bovinecarbonicanhydrase(30 kDa), soybeantrypsininhibitor(20.1kDa).2, Crudeintracellu-lar extractfrom P shermanii 13673.3, Purifiedpeptidase(1.6 J!gof protein)after gel filtrationon SephacrylS-100HA.Electrophorèse en gel de SD5-polyacrylamide.1, Standards de faible poids moléculaire : dehaut en bas, phosphorylase b (94 kDa), albu-mine bovine (67 kDa), ovalbumine (43 kDa), an-hydrase carbonique bovine (30 kDa), inhibiteurtrypsique de soja (20.1 kDa); 2, Extrait intracel-lulaire brut de P shermanii13673; 3, Peptidasepurifiée (1.6 Jlg de protéine) après gel filtrationsur Sephacryl S-100 HR.
mercaptoethanol had no effect Onenzymeactivity.
Several divalent metal ions were tested(fig 5). Addition of 0.1 mM Co acetate, Cusulfate, Fe sulfate inhibited the enzyme ac-tivity by 20, 53 and 77% respectively. Acomplete inhibition was observed with 0.1mM Zn. Addition of 0.1 mM Ca and Mgchloride had no inhibitory effect.
G Panon
Amino acid composition
The amino acid compositionis shown intable VI. The peptidase contained moder-ate quantities of hydrophobic amino acids,consistent with the intracellular origin ofthe protein.
DISCUSSION
ln the present study a proline iminopepti-dase was purified from the intracellularfraction of P shermanii 13673 in severalsteps. Purification, as estimated from thespecific activity, was approximately 80-foldwith a yield of 7%; but protein purificationof '" 1 OOO-foidwas achieved. Ouring thedifferent steps of purification, inactivationapparently occurred in spite of the pres-ence of stabilizers. The proline iminopepti-dase of P shermanii 13673 has a molecu-lar weight of 61 000 as estimated by SOS-PAGE and gel filtration. In comparison theaminopeptidase from S thermophilus has amolecular weight of 62 000, and its dipepti-dase 50 000 (Rabier and Oesmazeaud,1973), whereas the aminopeptidase from Llaetis has a molecular weight of 78 000 to81 000 (Eggimann and Bachmann, 1980).The prolyl-dipeptidyl aminopeptidase fromL helvetieus has a molecular weight of72 000 (Khalid and Marth, 1990) and theX-prolyl-dipeptidyl-aminopeptidase from Llaetis and from S thermophilus has a mo-lecular weight of '" 165 000 Da (Meyer andJordi, 1987). Incubation of the enzyme withdithiothreitol or 2-mercaptoethanol had noeffect on enzyme activity, suggesting thatan intact disulphide group was not essen-tial for the mechanism of action of this en-zyme. The enzyme was more markedly in-hibited by phenylmethylsulphonyl fluoride(1 mM). Theseresults suggest that the en-zyme is a serine protease. Bacterial dipep-tidyl aminopeptidases from L laetis, and S
Praline iminopeptidase from P shermanii
Table III. Relative ,activity' of the peptidase with different substrates.Activité" relative de la peptidase avec différents susbstrats.
447
Substrate Relative activity(%)
Pro p-NALeu p-NALys p-NAGly p-NAMetp-NAGlup-NAPhe p-NAGly-Pro p-NAArg-Pro p-NAGly-Arg p-NAPhe-Val p-NA
• Expressed as percentage of maximal activity measured with p-NA substituted amino acids, dipeptides or tripep-tides; • Exprimée en pourcentage de l'activité maximale mesurée avec les acides aminés substitués (P-NA), dipep-tides ou tripeptides.
thermophilus were also found to be serineproteases (Meyer and Jordi, 1987). Theenzyme was also inhibited by EDTA, thusindicating a requirement for metal ion. Thesame inhibition pattern was observed with
the enzymes. from S thermophilus (Meyerand Jordi, 1987). The enzyme was com-pletely inactivated by 0.1 mM Zn and lessinactivated by Co, Cu and- Fe. Divalentions Ca and Mg had no effect on the en-
448 GPanon
Table IV. Substrate specificity of the peptidase with B-casomorphin fragments. Hydrolysis ofB-casomorphin fragments was followed by HPLC. After incubation for 3 h at 40DC, the reactional mix-ture was injected cnte a Vydac C 18 5 ~ column and elution was pertormed with water/acetonitrilegradient.Specificité de substrats de la peptidase avec les fragments de B-casomorphine. L'hydrolyse des frag-ments de B-easomorphine a été suivie par HPLC. A près une incubation de 3 h à 40 DC, le mélangeréactionnel a été injecté sur une colonne Vydac C18 5Jl et l'élution faite par un gradient eau!acétonitrile.
zyme. The enzyme was most active at pH8.0. This result can be compared to thebroad pH optima of enzymes of S thermo-phi/us: pH 6.0-8.S (Rabier and Desmaz-eaud, 1973).
The optimum temperature was 40DC.The stability of the proline iminopeptidasewas high, SO% of the initial activity still re-
Relative activity C"l100 "
50
"pH
FIO.4A
maining at SODC. The considerable activityof this peptidase at 10DC (38% of relativeactivity), as weil as its good stability at pH6.0 indicate the potential role that it mayhave during cheese ripening in the libera-tion of amino acids. The quantity of intra-cellular enzyme is quite large. However,curd conditions are unlike the experimental
Relative activity C"l'00
50
'0
oL---,_~_~_~_~------I::::::;I<=-+_~o w ~ ~ ~ 60 BD 70 ~
Temperature C ClFIO.4B
Fig 4. Eftect of pH (A) and temperature (8) on peptidase activity. The peptidase activity (A410) wasassayed with Pro p-NA substrate.Role du pH (A) et de la température (B) sur l'activité de la peptidase. L'activité de la peptidase a ététestée avec le substrat Pro p-NA.
Praline iminopeptidase from P shermanii 449
Table V. Inhibition of peptidase activity.Inhibition de l'activité de la peptidase.
conditions: uncontrolled pH values, no Iy-sis during the exponential phase.
10080SO
70SO
8040302010o
none Co Cu Zn Mn F. C. MgDI\I\LENT IONS
~ CONCENTRATION 0.1 mM
After production of Swiss cheese, thepH is approximately 5.2 and rises to 5.6-5.8 during maturation. Also, during the rip-ening process the cheese is stored at dif-ferent temperatures between 10-24°C.During maturation the conditions are notoptimal for peptidase activity. But during ail
Fig 5. Effecl of divalent ions on peptidase activi-ty. The relative peptidase activity was measuredby the release of p-nitroanilide which was follo-wed spectrophotometrically.Rôle des ions divalents sur l'activité de la pepti-dase. L'activité relative de la peptidase a été me-surée par la libération de p-nitroanilide suivie auspectrophotomètre.
450 G Panon
Table VI. Amino acid composition of the proline iminopeptidase of P shermanii 13673.Composition en acides aminés de la proline iminopeptidase de P shermanii 13673.
the ripening process of ~ weeks, thetempe rature and pH values are favourablefor peptidase activity. The purified enzymecatalyzed hydrolysis of dipeptide and pep-tide derivatives containing proline residuesin the ultimate position. Pro-X peptideswere specifically hydrolyzed. The X-Propeptides were resistent to enzymatic hy-drolysis. The enzyme was identified as aproline iminopeptidase also with a proli-nase activity, according to the possible se-quences of enzyme activities proposed byBooth et al (1990). Several peptidaseshave been isolated and characterized inS eremoris (Geis et al, 1985; Exterkateand de Veer, 1987; Van Boven et al,1988), and L laetis (Tan and Konings,1990). None is capable of releasing N ter-minai prolyl residues. Beta-casomorphindes-Tyr was a good substrate for the pro-line iminopeptidase of P shermanii 13673.
The X-prolyl dipeptidyl peptidase puri-fied from L laetis (Zevaco et al, 1990) hy-drolyzed the B-casomorphin producing 4different peptides.
ln Swiss type cheese proteinases fromlactic acid bacteria provide peptides fromcaseins. These peptides will be furtherc1eaved by peptidases. Peptide hydrolasesfrom starters may include aminopeptidaseswith broad substrate specificities, dipepti-dases, and finally leucine aminopeptidaseor proline iminopeptidase in order to re-lease essential amino acids for cell nutri-tion. Degradation of casein by the action ofchymosin or other enzymes can result information and accumulation of bitter-tasting peptides in cheese (Vegarud andLangsrud, 1989).
Bitter-tasting peptides consistently havea high proportion of hydrophobie aminoacid residues. Bitterness will not develop if
Proline iminopeptidase from P shermanii 451
specifie peptidolytic enzymes with debitter-ing properties are present (Ardo et al,1989). Proline-containing peptides exhibit-ed bitterness (Ishibashi et al, 1988). Thisflavor defect may occur during acceleratedripening of cheese. Consequently, suffi-dent intracellular peptidases from starter,cells is a critical point in the release of ami-no acids and debittering of dairy products.The propionibacteria also produce intracel-lular peptidases which are released in thecheese by autolysis (Langsrud et al,1978b). Earlier observations by Langsrudet al (1977) suggested that proline-releasing peptidases predominate in Pshermanii. Proline has a sweet taste andits presence in particularly high concentra-tions in Emmental cheese led Langsrudand Reinbold (1973) to suggest that thisconstituted the basis of this aspect ofSwiss cheese flavor. In this paper we havedescribed the presence of a praline irnino-peptidase which enables P shermanii13673 to release free proline from interme-diary products of protein breakdown as ob-served by Langsrud et al (1978a).
Heat and freeze-shocking of prapioni-bacteria can be tested to accelerate ripen-ing of cheese, according to Frey et al(1986) and Bartels et al (1987a, b), cheesemanufacturers being interested in produc-ing mature-flavoured products in a shortperiod of time. More research is needed toobtain strains with Iytic properties as weilas a high content of stable intracellular en-zymes.
REFERENCESAndrews BA, Asenjo JA (1987) Enzymatic Iysis
and disruption of microbial ceUs. Tibteeh 5,273-277
Ardo Y, Larsson PO, Lindmark Mansson H, He-denberg A (1989) Studies of peptidolysis dur-ing early maturation and its influence on low-
fat cheese quality. Milehwissensehaft 44,485-490
Bartels HJ, Johnson ME, Oison NF (1987a) Ac-celerated ripening of Gouda cheese. 1. Effectof heat-shocked thermophilic lactobacilli andstreptococci on proteolysis and flavor devel-opment. Milehwissensehaft 42, 83-88
Bartels HJ, Johnson ME, Oison NF (1987b) Ac-celerated ripening of Gouda cheese. II. Effectof freeze-shocked Laetobaeillus helvetieus onproteolysis and f1avordevelopment. Milehwis-sensehaft 42, 139-144
Booth M, DonneUyWJ, Ni Fhaolain l, JenningsPV, O'Cuinn G (1990) Proline-specific pepti-dases of Streptocoeeus eremoris AM2.
, J Dairy Res 57,79-88Bradford MM (1976) A rapid and sensitive meth-
od for the quantitation of microgram quanti-ties of protein utilizing the principle of protein-dye binding. Anal Bioehem 72, 248-254
Casey MG, Meyer J (1985) Presence of X-prolyl-dipeptidyl-peptidase in lactic acid bac-teria. J Dairy Sei 68, 3212-3215
Desmazeaud MJ, Juge M (1976) Caractérisa-tion de l'activité protéolytique et fractionne-ment des dipeptidases et des aminopeptidas-es de Streptoeoeeus thermophilus. Lait 56,241-260
Eggimann B, Bachmann M (1980) Purificationand partial characterization of an aminopepti-dase from Laetobaeillus laetis. Appl EnvironMierobio/40, 876-882
El Soda M, Desmazeaud MJ (1982) Les pep-tide-hydrolases des lactobacilles du groupeThermobaeterium. 1. Mise en évidence deces activités chez Laetobaeillus helvetieus, Laeidophilus, L laetis et L bulgarieus. Gan JMierobio/28,1181-1188
Exterkate FA, de Veer GJCM (1987) Purificationand sorne properties of a membrane-boundaminopeptidase A from Streptocoeeus erem-oris. Syst Appl Mierobio/9, 183-191
Ezzat N, El Soda M, Desmazeaud MJ, Ismail A(1982) Peptide hydrolases from the Thermo-baeterium group of lactobacilli. II. Physiologi-cal factors and enzyme production. Milehwis-sensehaft 37, 666-668
452
Fox PF (1989) Proteolysis during cheese manu-facture and ripening. J Dairy Sei 72, 1379-1400
Frey JP, Marth EH, Johnson ME, Oison NF(1986) Heat-and freeze-shocking causechanges in peptidase and protease activityof Lactobacillus he/veticus. Milchwissens-chaft41 , 681-685
Geis A, Bockelman W, Teuber M (1985) Simul-taneous extraction and purification of a cellwall-associated peptidase and J}-caseinspe-cifie protease from Streptococcus cremorisAC1. App/ Microbio/ Biotechno/23, 79-84
Hettinga DH, Vedamuthu ER, Reinbold GW(1968) Pouch method for isolating and enu-merating propionibacteria. J Dairy Sci 51,1707-1709
Hickey MW, Hillier AJ, Jago GR (1983) Pepti-dase activities in lactobacilli. Aust J DairyTechno/38,118-123
Ishibashi N, Kubo T, Chino M, Fukui H, Shino-da l, Kikuchi E, Okai H, Fukui S (1988) Tasteof proline-eontaining peptides. Agric BiolChem 52, 95-98
Kamaly KM, Marth EH (1989) Enzyme activitiesof /actic streptococci and their role in matura-tion of cheese: a review. J Dairy Sci 72,1945-1966
Khalid NM, Marth EH (1990) Purification andpartial characterization of a prolyl-dipeptidylaminopeptidase from Lactobacillus he/veti-cus CNRZ 32. App/ Environ Microbio/ 56,381-388
Kondo JK, McKay LL (1982) Mutanolysin for im-proved Iysis and rapid protopiast formation indairy streptococci. J Dairy Sci 65, 1428-1431
Laemmli UK (1970) Cleavage of structural pro-teins during the assembly of the head of bac-teriophage T4. Nature (Lond) 227,680-685
Langsrud T (1974) Proline production by propi-onibacteria. Thesis, Iowa State University Li-brary, Ames, Iowa
Langsrud T, Reinbold CW (1973) Flavour devel-opment and microbiology of Swiss cheese -a review. J Milk Food Techno/36, 487-490
Langsrud T, Reinbold GW, Hammond EG(1977) Proline production by Propionibacteri-um shermanii P59. J Dairy Sci 60, 16-23
GPanon
Langsrud T, Reinbold GW, Hammond EG(1978a) Free proline production by strains ofpropionibacteria. J Dairy Sci 61, 303-308
Langsrud T, Reinbold GW, Hammond EG(1978b) Autolysis of propionibacteria. XX IntDairy Congr E, 595-596
Meyer J, Jordi R (1987) Purification and charac-terization of X-prolyl-dipeptidyl-aminopeptidase from Lactobacillus /actis andfrom Streptococcus thermophilus. J Dairy Sci70,738-745
More WEC, Holdeman LV (1974) Genus 1. Prop-ionibacterium. In: Bergey's Manua/ of Deter-minative Bacteri%gy (Buchanan RE, Gib-bons NE, eds) Williams and Wilkins Co,.Baltimore, 633-647
Neviani E, Boquien CY, Monnet V, Phan ThanhL, Grippon JC (1989) Purification and charac-terization of an aminopeptidase from Lacto-coccus /actis subsp cremoris AM2. App/ Envi-ron Microbio/55, 2308-2314
Perez Chaia A, Pesee de Ruiz Hoigado A, Oli-vier G (1990) Peptide hydrolases of propioni-bacteria: effect of pH and temperature.J Food Prot 53, 237-240
Rabier D, Desmazeaud MJ (1973) Inventairedes différentes activités intracellulaires deStreptococcus thermophilus. Purification etpropriétés d'une dipeptide-hydrolase et d'uneaminopeptidase. Biochimie 55, 389-404
Tan PST, Konings WN (1990) Purification andcharacterization of an aminopeptidase fromLactococcus /actis subsp cremoris Wg2. App/Environ Microbio/56, 526-532
Van Boven A, Tan PST, Konings WN (1988)Purification and characterization of a dipepti-dase from Streptococcus cremoris Wg2. App/Environ Microbio/54, 43-49
Vegarud GE, Langsrud T (1989) The level of bit-terness and solubility of hydrolysates pro-duced by controlled proteolysis of caseins.J Dairy Res 56, 375-379