Immunologv, 1971, 21, 33. Cytotoxic Action of Antiserum and Complement: Quantification with a Colony Inhibition Method A. R. BAKER,* T. BORSOS AND H. R. COLTEN Biology Branch, National Cancer Institute, National Institutes of Health, Bethesda, Maryland 20014, U.S.A. (Received 22nd July 1970) INTRODUCTION The availability of precise quantitative methods for the study of immune haemolysis has made it possible to study in detail the mechanism of action of antibody and complement on erythrocytes (Mayer, 1961; Borsos, Colten, Spalter, Rogentine and Rapp, 1968). Most of the methods for detecting complement mediated lysis of nucleated cells are not suitable to critically examine the mechanism of action of antiserum and complement on nucleated cells (Gorer and O'Gorman, 1956; Reif, 1962; Sanderson, 1964; Rapp, Colten and Borsos, 1966). These methods are limited in one or more of the following ways: lysis of cells can be quantified over a limited range only (20-80 per cent), the criteria for cell death are imprecise, baseline values of controls are variable and excessive, methods of detecting cell death are insensitive and radioactive markers introduced into cells may be cytotoxic. The availability of a rapidly growing cell line with high plating efficiency and with the ability to form discrete colonies has made it possible for us to adapt the colony inhibition technique (Hellstrom, Hellstrbm and Pierce, 1968) to quantify studies of complement mediated lysis of nucleated cells. With this technique cell death is defined as a loss of colony forming ability, lysis can be quantified over a wide range (10-99.99 %) baseline values of controls are small and constant, the detection of cell death is sensitive and requires the incorporation of no markers. Furthermore, since damage to a cell by antibody and complement leads to a loss of its colony forming ability the lysis of a single cell can be detected. The purpose of this report is to describe the application of the colony inhibition technique to the study of the mechanism of complement mediated injury to nucleated cells. MATERIALS AND METHODS Tissue culture cells Chinese hamster lung (CHL) cells (clonal line V-79-753-B3) (Elkind and Sutton, 1960) kindly provided by Drs M. M. Elkind and W. U. Shipley were used as the target cell in all experiments. The cells were grown in modified Eagle's medium. This medium was made by mixing 400 ml of Eagle's medium No. 2 with 7-6 ml of a 2 x concentrate of NCTC 109, 15X8 ml of 0*15 per cent trypsin, 2 ml of 3 per cent glutamine solution (all obtained from NIH Media Unit), and 75 ml of foetal bovine serum (Flow Laboratories, Rockville, Md). The cells were incubated at 370 in a humidified atmosphere of 5 per cent CO2: 95 per cent * Present address: Peter Bent Brigham Hospital, 721 Huntington Avenue, Boston, Mass, 02115. U.S.A. Please send reprint requests to the above address in care of Dr Harvey R. Colten. 33
Transcript
Immunologv, 1971, 21, 33.
Cytotoxic Action of Antiserum and Complement:Quantification with a Colony Inhibition Method
A. R. BAKER,* T. BORSOS AND H. R. COLTEN
Biology Branch, National Cancer Institute, National Institutes ofHealth,Bethesda, Maryland 20014, U.S.A.
(Received 22nd July 1970)
INTRODUCTION
The availability of precise quantitative methods for the study ofimmune haemolysis hasmade it possible to study in detail the mechanism of action of antibody and complementon erythrocytes (Mayer, 1961; Borsos, Colten, Spalter, Rogentine and Rapp, 1968).Most of the methods for detecting complement mediated lysis of nucleated cells are not
suitable to critically examine the mechanism of action of antiserum and complement onnucleated cells (Gorer and O'Gorman, 1956; Reif, 1962; Sanderson, 1964; Rapp, Coltenand Borsos, 1966). These methods are limited in one or more of the following ways: lysisof cells can be quantified over a limited range only (20-80 per cent), the criteria for celldeath are imprecise, baseline values of controls are variable and excessive, methods ofdetecting cell death are insensitive and radioactive markers introduced into cells may becytotoxic.The availability of a rapidly growing cell line with high plating efficiency and with the
ability to form discrete colonies has made it possible for us to adapt the colony inhibitiontechnique (Hellstrom, Hellstrbm and Pierce, 1968) to quantify studies of complementmediated lysis of nucleated cells. With this technique cell death is defined as a loss ofcolony forming ability, lysis can be quantified over a wide range (10-99.99 %) baselinevalues of controls are small and constant, the detection of cell death is sensitive andrequires the incorporation ofno markers. Furthermore, since damage to a cell by antibodyand complement leads to a loss of its colony forming ability the lysis of a single cell can bedetected. The purpose of this report is to describe the application of the colony inhibitiontechnique to the study of the mechanism ofcomplement mediated injury to nucleated cells.
MATERIALS AND METHODSTissue culture cellsChinese hamster lung (CHL) cells (clonal line V-79-753-B3) (Elkind and Sutton, 1960)
kindly provided by Drs M. M. Elkind and W. U. Shipley were used as the target cell in allexperiments. The cells were grown in modified Eagle's medium. This medium was made bymixing 400 ml of Eagle's medium No. 2 with 7-6 ml of a 2 x concentrate of NCTC 109,15X8 ml of 0*15 per cent trypsin, 2 ml of 3 per cent glutamine solution (all obtained fromNIH Media Unit), and 75 ml of foetal bovine serum (Flow Laboratories, Rockville, Md).The cells were incubated at 370 in a humidified atmosphere of5 per cent CO2: 95 per cent
* Present address: Peter Bent Brigham Hospital, 721 Huntington Avenue, Boston, Mass, 02115. U.S.A.Please send reprint requests to the above address in care of Dr Harvey R. Colten.
33
A. R. Baker, T. Borsos and H. R. Colten
air. Cells were plated in Falcon plastic tissue culture dishes (Becton Dickinson, LosAngeles, Calif.). Under these conditions 95 per cent ofthe plated CHL cells were capable offorming colonies. The cells had a doubling time of about 8 hours.
Preparation of anti-CHL antiseraCHL cells (7.5 x 107 cells) in Freund's complete adjuvant (Difco Laboratories, Detroit,
Mich.) were injected into the foot pads ofNew Zealand white rabbits. Three weeks later therabbits were injected intravenously with a washed CHL cell suspension (9.3 x 107 cells).The rabbits were bled 6 days thereafter, and the serum was stored at 40. Guinea-piganti-CHL antiserum was prepared by injecting 1 x 107 CHL cells in Freund's adjuvantinto the foot pads of guinea-pigs. The animals received weekly intradermal injections ofCHL cells for 5 weeks and were bled 6 days after the last injection.
All of the antisera were heated at 560 for 30 minutes immediately before use.Antisera were separated into 'IgM' and 'IgG' fractions by Sephadex G-200 chromato-
graphy as described (Rapp and Borsos, 1966). The 'IgG' and 'IgM' fractions were thenconcentrated to the volume of original serum by ultrafiltration in a Diaflo cell (AmiconCorp., Cambridge, Mass.) with a UM-10 membrane.
ComplementRabbit complement was obtained from New Zealand white rabbits. Freshly drawn
blood was allowed to clot at 00 for 8 hours, centrifuged in the cold, the serum decanted,and stored at-40'.
Pooled fresh rabbit, guinea-pig, human and rat serum were obtained from SuburbanSerum Laboratories (Silver Spring, Md).Mouse complement was obtained from male C57BL6/N mice. Freshly drawn blood was
allowed to clot at 00, centrifuged and the serum was stored at-400.Rabbit complement was absorbed three times at 00 with washed Syrian hamster red
blood cells. All sera were sterilized before storage by Micropore filtration (GelmanInstrument Co., Ann Arbor, Mich., pore size 0i20 y).
Colony inhibition assay procedureCytotoxic activity was quantified by determining the reduction in the colony forming
ability of a suspension of CHL cells. Reduction in colony forming ability was comparedin each experiment to controls which consisted of untreated cells, cells exposed to antiserumalone and cells exposed to complement alone.A single cell suspension ofCHL cells was prepared by exposing a 3-day-old culture to a
0 03 per cent trypsin solution (Grand Island Biological Company, Grand Island, NewYork) for 9 minutes at 370. The cells were washed in Eagle's medium with 15 per cent foetalbovine serum (Flow Laboratories, Rockville, Md) (EM-15) and the concentration of thecells was adjusted to 1-5 x 105 cells/ml. At this stage 100 per cent of the cells were viable asdetermined by trypan blue dye exclusion. The target cells in suspension were firstincubated with antiserum, then with complement. After an appropriate incubation periodserial dilutions of the cell suspensions were made and replicate samples were plated.Dilutions were made to yield about 200 surviving cells per plate. After incubating for 6days at 370 the plates were removed from the incubator, the colonies were stained with1 per cent methylene blue dye (Fisher Scientific Co., Fairview, New Jersey), and thenumber of colonies were counted (Fig. 1).
34
Cytotoxic Action of Antiserum and ComplementThe preparation of sheep erythrocytes (E) and Veronal buffered saline I= 0 15 (VBS)
was described (Mayer 1961). Veronal-buffered saline sucrose, I=0-065 (VBS-sucrose)was prepared as given in reference (Rapp and Borsos, 1963). The preparation of partiallypurified guinea-pig CT (Colten, Bond, Borsos and Rapp, 1969), C2 (the second com-ponent of complement) (Borsos, Rapp and Mayer, 1961) and C-EDTA (C mixed with0-01 M ethylendiamine tetra acetate-Na3 buffer, pH 7.6) (Rapp and Borsos, 1966) hasbeen described.The CT fixation and transfer test, a method permitting estimation ofcell bound antibody
on a molecular basis, has been described in detail (Borsos and Rapp, 1965). In brief, thetest consists of an initial stage in which CT is fixed to a particulate antigen-antibodycomplex, followed by a second stage in which the CT is transferred quantitatively to
FIG. 1. The gross appearance of CHL cell colonies stained with methylene blue. A: control. B: coloniesformed after cells were exposed to antiserum and complement.
appropriate indicator cells. The indicator cells, consisting of sheep erythrocytes (E)sensitized with antibody and the fourth component ofC (EAC4), were made as described(Borsos and Rapp, 1967).
RESULTSMORPHOLOGIC CHANGES INDUCED BY COMPLEMENT MEDIATED CELL LYSIS
We examined by phase contrast microscopy the appearance of CHL cells before andafter exposure to anti-CHL antiserum and complement. The cells were treated with anamount of antiserum and complement sufficient to kill more than 99 9 per cent of the cells.Fig. 2 shows that by 8-5 minutes after the addition of antiserum and complement an in-crease in the cytoplasmic granularity was noted and small blebs appeared on the surfaceof the cells. The cells were also smaller than control cells. By 20 minutes these changeswere more pronounced. These changes are similar to the changes observed in other studiesof the lysis of nucleated cells by antibody and complement (Goldberg and Green, 1959).The following experiments were designed to determine the optimal conditions for the
inhibition ofcolony formation by antibody and complement. Preliminary experiments hadindicated that the rate of sensitization of CHL cells by rabbit anti-CHL antiserum, under
35
A. R. Baker, T. Borsos and H. R. Colten
conditions oflimited antibody, was a function ofantiserum concentration, was slow and anend point was not reached until approximately 180 minutes.
KINETICS OF COMPLEMENT LYSIS OF SENSITIZED CHL CELLS
A suspension of CHL cells (I 5 x 105/ml) was sensitized for 2 hours at 370 with a1/10,000 dilution of rabbit anti-CHL cell antiserum. Rabbit complement (1/6 dilution of
Before addition of C 7 minutes after C added
1X Z. # <K,35* t... iis s@ .... .. ....4'S
8 5 minutes after C' added 20 minutes after Gadded
FIG. 2. Appearance ofsensitized CHL cells before and after the addition ofcomplement as observed withphase contrast microscopy.
rabbit serum) was then added to the sensitized cells. The cells were incubated at 370 andthe lytic reaction stopped at timed intervals by dilution. Up to 25 per cent lysis wasobserved in control preparations consisting of CHL cells plus complement alone withprolonged periods of incubation. The data were corrected for the extent of lysis in thecontrols. In other experiments the kinetics of complement lysis was investigated withcells sensitized with 1/16,000 and with 1/32,000 dilutions of antiserum.The results of these experiments summarized in Fig. 3 show that at 1/10,000 and
1/16,000 dilutions of antiserum an endpoint is reached 120 minutes after the addition of
36
Cytotoxic Action of Antiserum and Complementcomplement to the sensitized cells. At a 1/32,000 dilution an endpoint was not reached by240 minutes.
EFFECT OF DOSE AND SPECIES OF COMPLEMENT ON CELL LYSIS
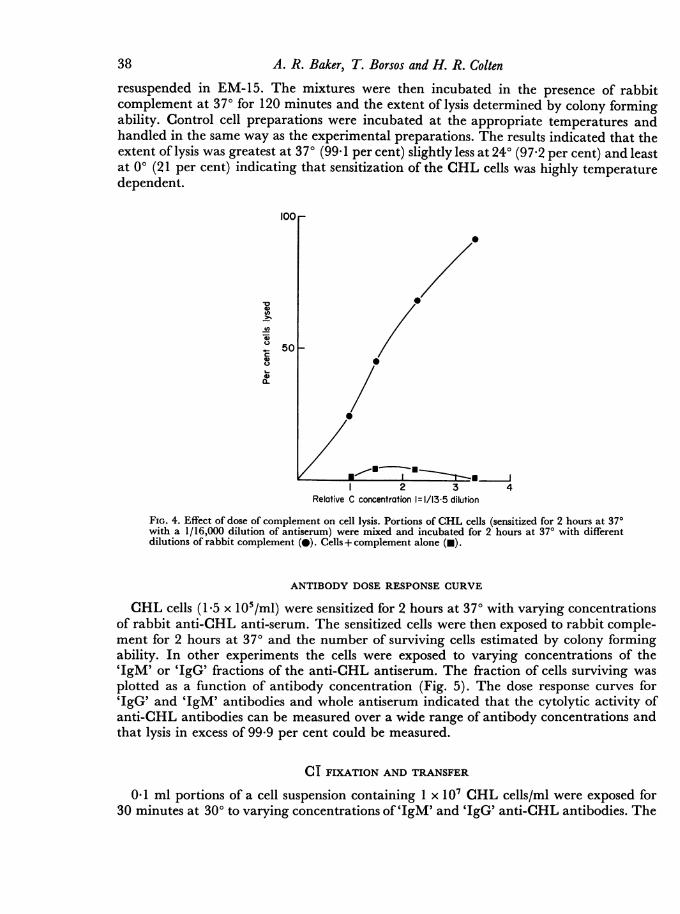
A CHL cell suspension (15 x 105/ml) was sensitized with an equal volume of a 1/16,000dilution of rabbit anti-CHL cell antiserum for 2 hours at 370. 1 0-ml aliquots of thesensitized cell suspension were then exposed to varying dilutions of fresh rabbit comple-ment for an additional 2 hours at 37°. The results of this experiment (Fig. 4) indicate thatthe extent of lysis is a function of complement concentration. Lysis of CHL cells withantibody alone or unsensitized CHL cells with complement was less than 5 per cent.Heating the complement source at 560 for 60 minutes completely abolished its lytic activity.
100_
21~~~~~~A
5 50
A
0V
60 120 180 240Timne (mirutes)
FIG. 3. Kinetics of complement lysis of sensitized CHL cells. Cells were sensitized for 2 hours at 370with a 1/10,000 (e), 1/16,000 (i), 1/32,000 (A) dilution of antiserum. 0 time: addition of rabbitcomplement.
In the following experiments the lytic activity of rabbit, human, rat, mouse andguinea-pig complement were compared. In the first experiment CHL cells (1-5 x 105/ml)were sensitized at 370 with either rabbit anti-CHL cell antiserum (1/10,000) or withguinea-pig anti-CHL antiserum (1/1000). The sensitized cell preparations were thenexposed for 1 hour at 370 to rabbit complement (1/6) or to guinea-pig complement (1/6).Antiserum, complement and untreated cell controls were included. The results indicatedthat regardless of the source of antibody, rabbit complement was more effective thanguinea-pig complement in the lysis of sensitized CHL cells. In other experiments, usingcells sensitized with rabbit anti-CHL antiserum we found that rabbit serum was a bettersource of complement than human, rat, or mouse serum.
EFFECT OF TEMPERATURE ON SENSITIZATION OF CHL CELLS
A CHL cell suspension (1-5 x 105 cells/ml) was divided into three portions. One 2-mlportion was incubated with an equal volume ofanti-CHL antiserum (1/4000) at 370; anotherat 240 and the third portion at 00. After 2 hours the cells were washed and each portion
37
A. R. Baker, T. Borsos and H. R. Colten
resuspended in EM-15. The mixtures were then incubated in the presence of rabbitcomplement at 370 for 120 minutes and the extent of lysis determined by colony formingability. Control cell preparations were incubated at the appropriate temperatures andhandled in the same way as the experimental preparations. The results indicated that theextent of lysis was greatest at 370 (99.1 per cent) slightly less at 240 (97-2 per cent) and leastat 00 (21 per cent) indicating that sensitization of the CHL cells was highly temperaturedependent.
100 _
_ 50 -
C
C) /
1 2 3 4Relative C concentration 1-1/13 5 dilution
FIG. 4. Effect of dose of complement on cell lysis. Portions of CHL cells (sensitized for 2 hours at 370with a 1/16,000 dilution of antiserum) were mixed and incubated for 2 hours at 370 with differentdilutions of rabbit complement (@). Cells+ complement alone (-).
ANTIBODY DOSE RESPONSE CURVE
CHL cells (1.5 x 105/ml) were sensitized for 2 hours at 370 with varying concentrationsof rabbit anti-CHL anti-serum. The sensitized cells were then exposed to rabbit comple-ment for 2 hours at 370 and the number of surviving cells estimated by colony formingability. In other experiments the cells were exposed to varying concentrations of the'IgM' or 'IgG' fractions of the anti-CHL antiserum. The fraction of cells surviving wasplotted as a function of antibody concentration (Fig. 5). The dose response curves for'IgG' and 'IgM' antibodies and whole antiserum indicated that the cytolytic activity ofanti-CHL antibodies can be measured over a wide range of antibody concentrations andthat lysis in excess of 99-9 per cent could be measured.
CT FIXATION AND TRANSFER
0 1 ml portions of a cell suspension containing 1 x 107 CHL cells/ml were exposed for30 minutes at 300 to varying concentrations of '1gM' and 'IgG' anti-CHL antibodies. The
38
Cytotoxic Action of Antiserum and Complementnumber of Cl fixing sites per cell was then determined with the Cl fixation and transfertest. To determine the slope of the dose response curves, the logarithm of the number ofCT fixing sites/cell was plotted as a function of the logarithm of relative antibody con-centration. It is seen from Fig. 6 that the slope of the 'IgM' dose response curve is close to1 and that of the 'IgG' is greater than 2.
.0
*0 IgG~~~~~~~~~g
0.01~~A
0 gI
8 i serum
U_~~~~
0 001
0 serum4812 1
Relative antibody concentratkin
Fic. 5. Lysis ofCHL cells as a function of antiserum and antibody concentration. (A) whole serum; (A)'IgM' fraction; (-) 'IgG' fraction. Relative antibody concentration of 1= 1/64,000 whole serum;1/6400 'IgM' fraction; 1/2700 'IgG' fraction.
CT FIXATION AND THE GENERATION OF CYTOLYTIC SITES
In the next experiment we investigated the quantitative relationship between CTfixation and the generation of cytolytic sites. Portions of a CHL cell suspension (1 x 1 06/ml)were sensitized with equal volumes ofdilutions ofthe 'IgM' fraction of the rabbit anti-CHLantiserum. Each of the sensitized cell preparations was then divided into two portions.One 2-ml portion was exposed to whole rabbit complement and the number of survivingcells estimated by colony forming ability. Another 2-ml portion was removed, centrifugedand CT added to the cell button. The number of CT fixing sites per CHL cell was thendetermined with the CT fixation and transfer test. The results of this experiment, shown in
39
A. R. Baker, T. Borsos and H. R. ColtenFig. 7, indicated that the concentration of antibody that produced one cytolytic site percell produced 1200 C 1 fixing sites/cell. The data also showed that the number oflytic sites percell was directly proportional to the relative antibody concentration. This observationmakes it unlikely that cell death was due to an accumulation of damaged sites/cell andmakes it likely that only one out of the 1200 CT fixing sites led to a cytolytic site.
U
800 _
IgG400 -
200
*9200 1gM
Relative antibody concentration
FIG. 6. Determination of the number ofCT fixing sites per cell generated as a function of 'IgM' (0) andIgG' (-) antibody concentration. The slope ofthe 'IgM' dose response curve is close to 1 and that ofthe'IgG' dose response curve is greater than 2. The precise slope of the 'IgG' dose response curve is some-what uncertain since the curve is based on only two experimental points.
EFFECT OF CELL DIVISION
Five ml of a CHL cell suspension (1.5 x 105/ml) were incubated with 5 ml of a 1/500dilution of anti-CHL antiserum for 2 hours at 370. The cell preparation was then dilutedin EM-15 to a final concentration ofapproximately 350 cells/ml. Samples were plated byadding 1 ml of the cell suspension to 10 ml of EM-15 in Petri dishes, and placed in theincubator for 6 hours to allow the cells to attach to the dishes. The cells were washed byremoving the medium from each of the dishes and by adding 10 ml of fresh growthmedium. The growth medium was removed from one set of dishes 8 hours after platingthe cells and 4-5 ml of a 1/18 dilution ofrabbit complement in EM-15 were added to oneset of dishes. Additional sets were exposed to complement at 24, 48 and 72 hours. In eachcase the cells were incubated with the complement for 2 hours at 37°. The complement wasthen removed and 10 ml of fresh growth medium was added. The number ofsurvivors wasdetermined after 6 days by counting the colonies formed. At each time interval controlswere handled in the same way as the experimental preparations.
40
41Cytotoxic Action of Antiserum and Complement
4 0 - 4000
3*0~~~~~~~~-300030 --
230 2000
- .0 0II
0-3
0-22 4 8 16
Relative IgM concentration
FIG. 7. Determination of the number of CT fixing (0) and cytolytic (0) sites per cell as a function of'IgM' antibody concentration.
TABLE 1
SUSCEPTIBILITY OF CHL CELLS TO COMPLEMENT MEDIATED LYSIS AT DIFFERENTTIMES AFTER SENSITIZATION
Colonies per plate
Time after Antiserumsensitization Control Antiserum Complement plus
Antiserum= 1/500, Complement= 1/18, No. of cells plated= 335.
A reduction in colony forming ability was noted only in the cell preparation exposed tocomplement 8 hours (i.e. one generation time) after sensitization. At 24, 48 and 72 hoursafter sensitization no reduction in colony forming ability was noted (Table 1).
DISCUSSIONThe data presented in this paper show that the colony inhibition technique is applicable
to the study of the mechanism of complement mediated cell injury. The results suggestthat there are several differences between cytolysis of red cells and nucleated cells. For
42 A. R. Baker, T. Borsos and H. R. Colten
example, we found that the rate of sensitization of CHL cells by the rabbit anti-CHLantiserum is slow; it takes 1-3 hours to reach an endpoint. In contrast, sensitization ofred cells reaches an endpoint a few minutes after the addition of antiserum (Borsos et al.,1968). Furthermore, sensitization of red cells proceeds equally well at 00 and 370, whereaswe find the extent of sensitization ofCHL cells by antibody is much less at 00 than at 370.The data do not permit one to decide whether these differences are due to differences
among the antibodies, to the nature and/or distribution of the number of antigenic sites onthe cell surfaces or to differences in the metabolic activity of the two cell types.Red cells once sensitized by haemolysin remain susceptible to the lytic action of com-
plement for many days. CHL cells sensitized with high concentrations of antibody appearto lose susceptibility to the lytic action of complement after three cell divisions. There areat least three possible explanations for this observation: (1) Antibody molecules areunequally distributed among the progeny of the sensitized cell. A consequence of unequaldistribution of the antibody could lead to an insufficiently sensitized daughter cell whichwould then form a colony. (2) Metabolic activity of the target cell may alter the cytolyticfunction of the antibody. (3) The antibody may dissociate from the cell surface.
It has been reported (Amos, Cohen and Klein, 1970) that sensitized mouse leukaemiacells also lose their susceptibility to lysis by complement after incubating at 370. This loss oflysability was not observed with sensitized cells incubated at 00 or at 370 with actinomycinD.
Studies ofimmune haemolysis showed that a single lytic site was necessary and sufficientto lyse a red cell. For red cells it has been found that one of every two CT fixing 'IgM'haemolysin molecules is capable of generating a haemolytic site (Rapp and Borsos, 1966).On the other hand, we have found that at least 1200 'IgM' anti-CHL antibody moleculeswere necessary to establish one cytolytic site. Previously we found that about 3000 CTfixing sites were necessary to generate one lytic site on the surface of a mouse leukaemiacell (Borsos et al., 1968). These findings should not be interpreted to mean that cumulativedamage is necessary for the lysis of these cell types for the slopes of the lytic dose responsecurves with 'IgM' anti-CHL and 'IgM' anti-mouse leukaemia cells were between 1 and1*5.The experiments in this paper also confirm the findings that rabbit complement is more
efficient in lysing nucleated cells than guinea-pig complement (Fass and Herberman,1969). In addition, rabbit complement was more efficient than mouse, rat or humancomplement.Our study also showed some of the difficulties inherent in studying the mechanism of
lysis of nucleated cells. For example the kinetics of sensitization and lysis are slow. Duringthis time the cells are metabolically active and are dividing and may be inactivating cellbound antibody. Thus, it is difficult to achieve optimal experimental conditions; for thisreason in some cases we had to select conditions that were practicable but not quite optimal.
REFERENCES
AMos, D. B., COHEN, I. and KLEIN, W.J.,JR. (1970).'Mechanisms of immunologic enhancement.' Trans-plant. Proc., 2, 68.
BoRsos, T., COLTEN, H. R., SPALTER,J. S., ROGENTINE,N. and RAPP, H. J. (1968). 'The C'la fixation andtransfer test: Examples of its applicability to thedetection and enumeration of antigens and anti-
bodies at cell surfaces.'_J. Immuol., 101, 392.BoRsos, T. and RAPP, H. J. (1965). 'Hemolysin
titration based on fixation of the activated firstcomponent of complement: Evidence that onemolecule of hemolysin suffices to sensitize an erythro-cyte.'_J. Immunol., 95, 559.
BoRsos, T. and RAPP, H.J. (1967). 'Immune hemolysis:
Cytotoxic Action qf Antiserum and Complement 43A simplified method for the preparation of EAc'4with guinea pig or with human complement.' J.Immunol., 99, 263.
BoRsos, T., RAPP, H.J. and MAYER, M. M. (1961).'Studies on the second component of complement. I.The reaction between EAc'1 and c'2: Evidence onthe single site mechanism of immune hemolysis anddetermination of c'2 on a molecular basis.' J.Immunol., 87, 310.
COLTEN, H. R., BOND, H. E., BORSOS, T. and RAPP, H.J. (1969). 'Purification of the first component ofcomplement by zonal ultracentrifugation.' J. Im-munol., 103, 862.
ELKIND, M. M. and SurrON, H. (1960). 'Radiationresponse of mammalian cells grown in culture. I.Repair of x-ray damage in surviving Chinesehamster lung cells.' Radiat. Res., 13, 556.
FASs, L. and HERBERMAN, R. B. (1969). 'A cytotoxicantiglobulin technique for assay of antibodies tohistocompatibility antigens.' J. Immunol., 102, 140.
GOLDBERG, B. and GREEN, H. (1959). 'The cytotoxicaction of immune gammaglobulin and complementon Krebs ascites tumor cells.' J. exp. Med., 109, 505.
GORER, P. A. and O'GORMAN, P. (1956). 'The cyto-toxic activity of isoantibodies in mice.' Transplant.Bull., 3, 142.
HELLSTROM, I., HELLSTROM, K. E. and PIERCE, G. E.(1968). 'In vitro studies of immune reactions againstautochthonous and syngeneic mouse tumors inducedby methylcholanthrene and plastic discs.' Int. J.Cancer, 3, 467.
MAYER, M. M. (1961) 'Complement and complementfixation.' Experimental Immunochemistry (Ed. by E. A.Kabat and M. M. Mayer), 2nd edn, p. 133. C.Thomas, Springfield, Illinois.
RAPP, H. J. and BoRsos, T. (1963). 'Effects of lowionic strength on immune hemolysis.' J. Immunol.,91, 826.
RAPP, H. J. and BoRsos, T. (1966). 'Forssman antigenand antibody: Preparation of water soluble antigenand measurement of antibody concentration byprecipitin analysis, by C'la fixation and by hemolyticactivity.'_7. Immunol., 96, 913.
RAPP, H. J., COLTEN, H. R. and BORSOS, T. (1966).'Immune cytolysis measurement by electrolyticconductance: Preliminary note.' J. nat. Cancer Inst.,36, 559.
REIF, A. E. (1962). 'Immune cytolysis of three mouseascites tumors.'J. Immunol., 89, 849.
SANDERSON, A. R. (1964). 'Cytotoxic reactions ofmouse isoantisera: Preliminary considerations.'Brit. J. exp. Path., 65, 398.