Redox-coupled structural changes in nitrite reductase revealed by serial femtosecond and microfocus crystallography Received August 11, 2015; accepted November 19, 2015; published online January 14, 2016 Yohta Fukuda 1, *, Ka Man Tse 1,y , Mamoru Suzuki 2,3,y,z , Kay Diederichs 4,y , Kunio Hirata 3,y , Takanori Nakane 5 , Michihiro Sugahara 3 , Eriko Nango 3 , Kensuke Tono 6 , Yasumasa Joti 6 , Takashi Kameshima 6 , Changyong Song 3,7 , Takaki Hatsui 3 , Makina Yabashi 3 , Osamu Nureki 5 , Hiroyoshi Matsumura 1, *, Tsuyoshi Inoue 1,§ , So Iwata 3,8 and Eiichi Mizohata 1,ô 1 Department of Applied Chemistry, Graduate School of Engineering, Osaka University, 2-1 Yamadaoka, Suita, Osaka 565- 0871, Japan; 2 Institute for Protein Research, Osaka University, 3-2 Yamadaoka, Suita, Osaka 565-0871, Japan; 3 RIKEN SPring-8 Center, 1-1-1 Kouto, Sayo-cho, Sayo-gun, Hyogo 679-5148, Japan; 4 Department of Biology, University of Konstanz, D-78457 Konstanz, Germany; 5 Department of Biological Sciences, Graduate School of Science, The University of Tokyo, 7-3-1 Hongo, Bunkyo- ku, Tokyo 113-0033, Japan; 6 Japan Synchrotron Radiation Research Institute, 1-1-1 Kouto, Sayo-cho, Sayo-gun, Hyogo 679- 5198, Japan; 7 Department of Physics, Pohang University of Science and Technology, Pohang 790-784, Korea; and 8 Department of Cell Biology, Graduate School of Medicine, Kyoto University, Yoshidakonoe-cho, Sakyo-ku, Kyoto, 606-8501, Japan *Present addresses: Yohta Fukuda, Department of Biochemistry and Molecular Biophysics, Columbia University, 650 W 168 Street, NY 10032, USA; Hiroyoshi Matsumura, Department of Biotechnology, College of Life Sciences, Ritsumeikan University, 1-1-1 Noji-higashi, Kusatsu Shiga 525-8577, Japan. y These authors contributed equally to this work. z Mamoru Suzuki, Institute for Protein Research, Osaka University, 3-2 Yamadaoka, Suita, Osaka 565-0871, Japan. Tel: +81-6-6879-8637, Fax: +81-6-6879-4313, email: [email protected]§ Tsuyoshi Inoue, Department of Applied Chemistry, Graduate School of Engineering, Osaka University, 2-1 Yamadaoka, Suita, Osaka 565-0871, Japan. Tel: +81-6-6879-7408, Fax: +81-6-6879- 7409, email: [email protected]ô Eiichi Mizohata, Department of Applied Chemistry, Graduate School of Engineering, Osaka University, 2-1 Yamadaoka, Suita, Osaka 565-0871, Japan. Tel: +81-6-6879-7410, Fax: +81-6-6879- 7409, email: [email protected]Serial femtosecond crystallography (SFX) has enabled the damage-free structural determination of metalloen- zymes and filled the gaps of our knowledge between crystallographic and spectroscopic data. Crystallographers, however, scarcely know whether the rising technique provides truly new structural insights into mechanisms of metalloenzymes partly because of limited resolutions. Copper nitrite reductase (CuNiR), which converts nitrite to nitric oxide in denitrification, has been extensively studied by synchrotron radiation crystallography (SRX). Although catalytic Cu (Type 2 copper (T2Cu)) of CuNiR had been suspected to tolerate X-ray photoreduction, we here showed that T2Cu in the form free of nitrite is reduced and changes its coordination structure in SRX. Moreover, we deter- mined the completely oxidized CuNiR structure at 1.43 A ˚ resolution with SFX. Comparison between the high-resolution SFX and SRX data revealed the subtle structural change of a catalytic His residue by X-ray photoreduction. This finding, which SRX has failed to uncover, provides new insight into the reaction mechan- ism of CuNiR. Keywords: copper/electron transfer/enzyme/serial femtosecond crystallography/X-ray free-electron laser. Abbreviations: CuNiR, copper nitrite reductase; ET, electron transfer; GtNiR, Geobacillus thermodenitrifi- cans copper nitrite reductase; PCET, proton-coupled electron transfer; RT, room temperature; SFX, serial femtosecond crystallography; SRX, synchrotron ra- diation crystallography; T1Cu, Type 1 copper; T2Cu, Type 2 copper; XFEL, X-ray free-electron laser. Since the invention of the HaberBosch process, the amount of nitrogen oxides fixed in soils and waters has been increasing and the global nitrogen cycle has grad- ually changed (1, 2). In the cycle, nitrogen fixed in the form of ammonium salts are converted to nitrogen oxides and then reduced to a dinitrogen gas in a step- wise manner (NO 3 ! NO 2 ! NO ! N 2 O ! N 2 ) (3). This reduction process, denitrification, is the main path for fixed nitrogen to be removed and hence has major agronomic and environmental impacts. Chemical reactions in denitrification are performed by microorganisms and coupled with their anaerobic respiratory systems in which metalloenzymes are uti- lized (3, 4). Nitrite reduction to nitric oxide (NO 2 + e + 2H + ! NO + H 2 O) is an important step in denitrification where the ion is changed to the toxic and highly reactive gas. Two types of dissimilatory ni- trite reductase (NiR) have been identified to date (3, 4). One of them is cd 1 -type heme nitrite reductase (cd 1 NiR), which functions as a homodimer (5). The other one is copper nitrite reductase (CuNiR): a homo- trimeric copper-containing enzyme. CuNiR can also reduce dioxygen to hydrogen peroxide (6, 7) and cat- alyse the dismutation of superoxide (8). Each mono- mer of typical CuNiR contains two copper sites: Type 1 Cu (T1Cu) with a CysMetHis 2 ligand set and Type 2 Cu (T2Cu) with a His 3 ligand set (913). The T1Cu site accepts an electron from c-type cytochromes (14, 15) or blue copper proteins (6, 16, 17), when CuNiR and the donor protein form a transient elec- tron transfer (ET) complex. The received electrons are J. Biochem. 2016;112 doi:10.1093/jb/mvv133 ß The Authors 2016. Published by Oxford University Press on behalf of the Japanese Biochemical Society. This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons. org/licenses/by-nc/4.0/), which permits non-commercial re-use, distribution, and reproduction in any medium, provided the original work is properly cited. For commercial re-use, please contact [email protected]1 Journal of Biochemistry Advance Access published January 22, 2016 at University of Konstanz, Library on February 23, 2016 http://jb.oxfordjournals.org/ Downloaded from
Transcript
Redox-coupled structural changes in nitrite reductase revealedby serial femtosecond and microfocus crystallography
Received August 11 2015 accepted November 19 2015 published online January 14 2016
Yohta Fukuda1 Ka Man Tse1yMamoru Suzuki23yz Kay Diederichs4yKunio Hirata3y Takanori Nakane5Michihiro Sugahara3 Eriko Nango3Kensuke Tono6 Yasumasa Joti6Takashi Kameshima6 Changyong Song37Takaki Hatsui3 Makina Yabashi3Osamu Nureki5 Hiroyoshi Matsumura1Tsuyoshi Inoue1sect So Iwata38 andEiichi Mizohata1
1Department of Applied Chemistry Graduate School ofEngineering Osaka University 2-1 Yamadaoka Suita Osaka 565-0871 Japan 2Institute for Protein Research Osaka University 3-2Yamadaoka Suita Osaka 565-0871 Japan 3RIKEN SPring-8Center 1-1-1 Kouto Sayo-cho Sayo-gun Hyogo 679-5148 Japan4Department of Biology University of Konstanz D-78457Konstanz Germany 5Department of Biological Sciences GraduateSchool of Science The University of Tokyo 7-3-1 Hongo Bunkyo-ku Tokyo 113-0033 Japan 6Japan Synchrotron RadiationResearch Institute 1-1-1 Kouto Sayo-cho Sayo-gun Hyogo 679-5198 Japan 7Department of Physics Pohang University of Scienceand Technology Pohang 790-784 Korea and 8Department of CellBiology Graduate School of Medicine Kyoto UniversityYoshidakonoe-cho Sakyo-ku Kyoto 606-8501 Japan
Present addresses Yohta Fukuda Department of Biochemistryand Molecular Biophysics Columbia University 650 W 168 StreetNY 10032 USA Hiroyoshi Matsumura Department ofBiotechnology College of Life Sciences Ritsumeikan University1-1-1 Noji-higashi Kusatsu Shiga 525-8577 JapanyThese authors contributed equally to this workzMamoru Suzuki Institute for Protein Research Osaka University 3-2Yamadaoka Suita Osaka 565-0871 Japan Tel +81-6-6879-8637Fax +81-6-6879-4313 email mamorusuzukiproteinosaka-uacjpsectTsuyoshi Inoue Department of Applied Chemistry GraduateSchool of Engineering Osaka University 2-1 Yamadaoka SuitaOsaka 565-0871 Japan Tel +81-6-6879-7408 Fax +81-6-6879-7409 email inouetchemengosaka-uacjpEiichi Mizohata Department of Applied Chemistry GraduateSchool of Engineering Osaka University 2-1 Yamadaoka SuitaOsaka 565-0871 Japan Tel +81-6-6879-7410 Fax +81-6-6879-7409 email mizohatachemengosaka-uacjp
Serial femtosecond crystallography (SFX) has enabledthe damage-free structural determination of metalloen-zymes and filled the gaps of our knowledge betweencrystallographic and spectroscopic dataCrystallographers however scarcely know whether therising technique provides truly new structural insightsinto mechanisms of metalloenzymes partly because oflimited resolutions Copper nitrite reductase (CuNiR)which converts nitrite to nitric oxide in denitrificationhas been extensively studied by synchrotron radiationcrystallography (SRX) Although catalytic Cu (Type 2copper (T2Cu)) of CuNiR had been suspected to tolerateX-ray photoreduction we here showed that T2Cu in theform free of nitrite is reduced and changes its
coordination structure in SRX Moreover we deter-mined the completely oxidized CuNiR structure at143 A resolution with SFX Comparison between thehigh-resolution SFX and SRX data revealed the subtlestructural change of a catalytic His residue by X-rayphotoreduction This finding which SRX has failed touncover provides new insight into the reaction mechan-ism of CuNiR
Abbreviations CuNiR copper nitrite reductase ETelectron transfer GtNiR Geobacillus thermodenitrifi-cans copper nitrite reductase PCET proton-coupledelectron transfer RT room temperature SFX serialfemtosecond crystallography SRX synchrotron ra-diation crystallography T1Cu Type 1 copper T2CuType 2 copper XFEL X-ray free-electron laser
Since the invention of the HaberBosch process theamount of nitrogen oxides fixed in soils and waters hasbeen increasing and the global nitrogen cycle has grad-ually changed (1 2) In the cycle nitrogen fixed in theform of ammonium salts are converted to nitrogenoxides and then reduced to a dinitrogen gas in a step-wise manner (NO3
NO2
NO N2O N2)
(3) This reduction process denitrification is the mainpath for fixed nitrogen to be removed and hence hasmajor agronomic and environmental impactsChemical reactions in denitrification are performedby microorganisms and coupled with their anaerobicrespiratory systems in which metalloenzymes are uti-lized (3 4) Nitrite reduction to nitric oxide (NO2
+e + 2H+
NO + H2O) is an important step indenitrification where the ion is changed to the toxicand highly reactive gas Two types of dissimilatory ni-trite reductase (NiR) have been identified to date (3 4)One of them is cd1-type heme nitrite reductase(cd1NiR) which functions as a homodimer (5) Theother one is copper nitrite reductase (CuNiR) a homo-trimeric copper-containing enzyme CuNiR can alsoreduce dioxygen to hydrogen peroxide (6 7) and cat-alyse the dismutation of superoxide (8) Each mono-mer of typical CuNiR contains two copper sites Type1 Cu (T1Cu) with a CysMetHis2 ligand set andType 2 Cu (T2Cu) with a His3 ligand set (913) TheT1Cu site accepts an electron from c-type cytochromes(14 15) or blue copper proteins (6 16 17) whenCuNiR and the donor protein form a transient elec-tron transfer (ET) complex The received electrons are
J Biochem 2016112 doi101093jbmvv133
The Authors 2016 Published by Oxford University Press on behalf of the Japanese Biochemical SocietyThis is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (httpcreativecommonsorglicensesby-nc40) which permits non-commercial re-use distribution and reproduction in any medium provided the original work is properlycited For commercial re-use please contact journalspermissionsoupcom 1
Journal of Biochemistry Advance Access published January 22 2016 at U
niversity of Konstanz L
ibrary on February 23 2016httpjboxfordjournalsorg
Dow
nloaded from
transferred to the T2Cu site the catalytic centrethrough a CysHis pathway The AspHis pair(Aspcat and Hiscat) which is conserved above theT2Cu site and connected via a water molecule (brid-ging water) is essential to the enzymatic activity (1819) though the exact role has been ambiguous
In conventional synchrotron radiation crystallog-raphy (SRX) strong X-ray beams induce photoreduc-tion of metal centres and destroy their naturalstructures (2023) Spectroscopic analysis revealedthat T1Cu in CuNiR is rapidly reduced by synchrotronX-ray (24) T2Cu is more resistant than T1Cu to X-ray damage in the absence of NO2
When the sub-strate binds to T2Cu X-ray photoreduction of T1Cu isfollowed by ET from T1Cu to T2Cu which results inreduction of NO2
in crystallo (24) This gated ET isexplained by the concept of proton-coupled ET (2526) although the detailed mechanism remains to beelucidated Because the presence of NO2
acceleratesintramolecular ET also in solution (2527) it is obvi-ous that ET from T1Cu to T2Cu is gated to someextent However kinetic studies demonstrated therandom sequential mechanism of nitrite reductionie intramolecular ET can occur both with and with-out the binding of the substrate (28 29) Especiallyintramolecular ET before substrate binding is domin-ant at low pH (565) (28) Moreover we have recentlyshown that substrate-free T2Cu in a CuNiR crystalcrystallized at pH 45 may be reduced by synchrotronX-rays and that an unknown chemical reaction occurson T2Cu during data collection (30)
To investigate the nitrite reduction mechanism inCuNiR detailed structural comparison between itsoxidized and reduced state is necessary Here we clo-sely examined X-ray-induced structural changes andchemical reactions at the T2Cu site using a helicalscan method combined with microfocus X-ray beams(31) Furthermore we determined the first completelyoxidized CuNiR structure using serial femtosecondcrystallography (SFX) with X-ray free-electron laser(XFEL) (32) which has enabled damage-free struc-tural determination of metalloenzymes even at roomtemperature (RT) (3337) Because the Bragg spacingsof previously determined SFX structures of metalloen-zymes were longer than typical covalent bond lengthsfound in macromolecules (15 A) it has been difficultto obtain at the chemical level new structural insightsinto the reaction mechanisms of metalloenzymesCuNiR crystals used in this study have been knownto diffract X-rays well therefore we could determineits high-resolution SFX structure We here would liketo report our results in detail
Materials and Methods
Sample preparation of CuNiR from Geobacillusthermodenitrificans (GtNiR)Geobacillus thermodenitrificans copper nitrite reductase (GtNiR) wasexpressed and purified as described previously (30) We used chlor-ide-free buffers for all the steps of purification and crystallizationMicrocrystals for SFX were obtained by a rotational crystallizationtechnique using nanoseeds of the protein as follows Macrocrystalswere transferred to a 15ml tube (Eppendorf Hamburg Germany)containing 1ml of solution composed of 100mM sodium acetate
buffer (pH 45) 55 (wv) polyethylene glycol 4000 and 75mMCuSO4 After sonicating the crystals on ice with a UD-211 ultraso-nicator (Tomy Seiko Co Tokyo Japan) the solution was centri-fuged and the supernatant was collected as a nanoseed solution In a15ml centrifuge tube (AS ONE Co Osaka Japan) 4ml of the20mgml protein solution was mixed with 4ml of the precipitantsolution which was composed of 100mM sodium acetate buffer(pH 45) 15 (wv) polyethylene glycol 4000 75mM CuSO4 andthen 160ml of the nanoseed solution was added The centrifuge tubehad been rotated on a RT-50 culture rotator (TITEC SaitamaJapan) at 30 rpm for 1 week at RT The microcrystal solutionwas filtered through a 30 mm CellTrics filter (Chiyoda Sci CoTokyo Japan) and adjusted to a number density of 44108 crys-talsml by adding 4ml of the precipitant solution
Synchrotron data collectionCryogenic SRX datasets were collected using microfocus beamlineBL32XU at SPring-8 (38) A large single crystal of GtNiR(915620230mm3) was flash-cooled by immersion in liquid nitro-gen and mounted on a conventional goniometer with the longest axisroughly directed towards the horizontal rotation axis Along thelongest edge of the crystal 120 irradiation points were chosen Theregular intervals between the irradiation points were 76 mm which issufficient to separate the radiation damage at each irradiation pointThe beam at a wavelength of 07500 A was focused to 15 mm (heightH)10 mm (width W) with a photon flux of 81011 photonssUsing the helical scan method a total of seven datasets(SR1SR7) were repeatedly collected in order from the samepoints of the same crystal except for absorbed X-ray dose perframe The exposure time for each image was 1 s For datasetsSR1 SR3 SR5 and SR7 each image was collected with an absorbeddose of 0064MGyframe with a 923 attenuated beam while fordatasets SR2 SR4 and SR6 the dose corresponded to 8252MGyframe without attenuation The X-ray doses were calculated withRADDOSE (39) The parameters and statistics are summarized inTable I
RT data collection was performed at BL38B1 of SPring-8 (40) asdescribed previously (41) The dataset was collected from one pos-ition of a single crystal using an ADSC Quantum 315 charge-coupled device (CCD) detector (Area Detector Systems Co CAUSA) The beam size was 50 mm (H)88 mm (W) The oscillationangle and exposure time per image were set to 1 and 15 srespectively A total of 120 diffraction images were collected fromthe single crystal The parameters and statistics are summarized inTable II
Structure determination of the SRX structuresAll of the datasets were indexed and integrated using HKL2000 (42)The phases were determined by the molecular replacement methodusing MOLREP (43) with a GtNiR monomer (Protein Data Bank(PDB) code 4ZK8) as a search model Manual model building wasperformed using WinCoot 07 (44) The program REFMAC5 (45)from the CCP4 suite (ver 650) (46) was used for structural refine-ment Anisotropic displacement parameters were introduced afterwater molecules were built in the models The final models werechecked for stereochemical quality using MolProbity (47)
Single-shot XFEL data collectionThe experiment was performed at BL3 of SPring-8 AngstromCompact Free-Electron Laser (SACLA) in Hyogo Japan (48)Using 210 fs XFEL pulses we collected diffraction patterns fromgreenish-blue (aerobically oxidized) GtNiR microcrystals(Supplementary Fig S1a) The pulse duration shorter than 10 fs isquite important to obtain intact metalloprotein structures becauseultrabright XFEL beams damage electronic structures of heavyatoms in a few tens of femtoseconds (49) and can destroy the naturalstructures of metal centres (50) A liquid injector (nozzle aperturediameter 200mm) with a sample circulation system was used (51)Microcrystal sample (55ml) was placed in a reservoir The flow ratewas set to 53mlmin (70 cms) The injector was installed in a heliumambiance diffraction chamber enclosure Diverse ApplicationPlatform for Hard X-ray Diffraction in SACLA (DAPHNIS) (52)The liquidstream width was nearly the same as the aperture sizeThe sample chamber was maintained at a temperature of 300K witha humidity of 8599 The diffraction patterns were collected usinga short-working-distance octal multiport CCD detector (53) with
Y Fukuda et al
2
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
XFEL radiation The microcrystals were exposed to single X-raypulses at a photon energy of 110 keV The pulses consisted of51010 photonspulse were focused to 25 mm (H)20 mm (W) atthe interaction point using KirkpatrickBaez mirrors (54) Therepetition rate was 30Hz and the typical pulse energy at thesample was 90 mJpulse The parameters and statistics are summar-ized in Table III
Structural determination of the SFX structureA total of 180942 images were collected of which 139391 diffrac-tion images were identified and 37186 images were indexed andmerged using CrystFEL (55) The data was indexed as spacegroup R3 and processed as space group H3 Indexing ambiguity inSFX was solved by an algorithm that clusters snapshots (56) Thephase was determined by molecular replacement using MOLREPwith the GtNiR monomeric unit (PDB code 4ZK8) as a searchmodel Manual model building was performed using WinCoot 07The program REFMAC5 from the CCP4 suite (ver 650) was usedfor structure refinement The microcrystals diffracted X-rays beyond14 A resolution (Supplementary Fig S1b) but the statistics of high-resolution shells such as CC12 and Rsplit were poor (SupplementaryFig S2) We therefore performed paired refinement to determine thelsquonominal resolutionrsquo based on the idea that proper use of weakerand noisier but higher-resolution data can provide better models (5758) Actually inclusion of noisy high-resolution data enables thebetter analysis of SFX data (59) The result of paired refinementshowed that including higher-resolution data provided a bettermodel than rejecting it (Supplementary Fig S3a) The studentrsquos t-test P value calculated from CC12 in the highest resolution shell cutat 143 A resolution (00417 n=7859) was 0000214 which is lessthan the significance level =0001 (Supplementary Fig S3b)Conversely the P value calculated from CC12 in the highest reso-lution shell cut at 142 A resolution (00246 n=8164) was 00261which is greater than (Supplementary Fig S3b) Based on thesethings we chose to use data up to 143 A resolution and performedfurther refinement The final model was checked for stereochemicalquality using MolProbity
Results
Radiation damages to GtNiR in SRXThe structures for SR1 SR3 SR5 and SR7 data wererefined against data to 150 A resolution whereas thosefor SR2 SR4 and SR6 data were refined to 134 Aresolution (Table I) Root-mean-square deviations(RMSDs) for Ca atoms between seven structureswere at most 01 A
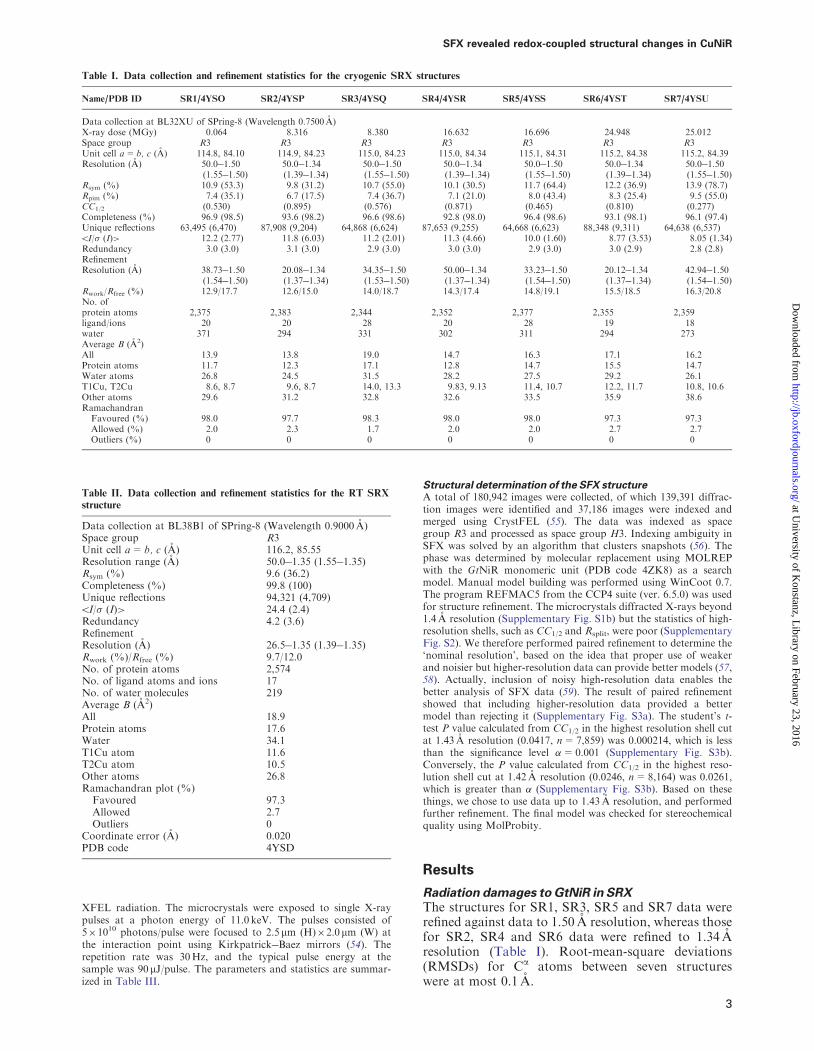
Table I Data collection and refinement statistics for the cryogenic SRX structures
NamePDB ID SR14YSO SR24YSP SR34YSQ SR44YSR SR54YSS SR64YST SR74YSU
Data collection at BL32XU of SPring-8 (Wavelength 07500 A)X-ray dose (MGy) 0064 8316 8380 16632 16696 24948 25012Space group R3 R3 R3 R3 R3 R3 R3Unit cell a= b c (A) 1148 8410 1149 8423 1150 8423 1150 8434 1151 8431 1152 8438 1152 8439Resolution (A) 500150
Table II Data collection and refinement statistics for the RT SRX
structure
Data collection at BL38B1 of SPring-8 (Wavelength 09000 A)Space group R3Unit cell a= b c (A) 1162 8555Resolution range (A) 500135 (155135)Rsym () 96 (362)Completeness () 998 (100)Unique reflections 94321 (4709)5I (I)gt 244 (24)Redundancy 42 (36)RefinementResolution (A) 265135 (139135)Rwork ()Rfree () 97120No of protein atoms 2574No of ligand atoms and ions 17No of water molecules 219Average B (A2)All 189Protein atoms 176Water 341T1Cu atom 116T2Cu atom 105Other atoms 268Ramachandran plot ()
Favoured 973Allowed 27Outliers 0
Coordinate error (A) 0020PDB code 4YSD
SFX revealed redox-coupled structural changes in CuNiR
3
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Changes in ligandT1Cu distance fell within therange of coordinate errors (Table IV) indicating thatthe T1Cu site did not show significant structuralchanges that can be detected at resolutions of our pre-sent structures Conversely the T2Cu site showed ob-vious structural changes induced by X-ray irradiationtwo water ligands (WatC and WatD) were present onT2Cu in the SR1 structure while the electron densityof WatD was not observed in other SR data (Fig 1a)Furthermore with increasing X-ray dose the T2Cuatom gradually sank towards a lsquoligand planersquocomposed of three N2 atoms of His ligands (Fig 1band c)
Completely oxidized GtNiR structure determined bySFXThe SFX structure was refined to 143 A resolution(Table III see Materials and Methods) The finalRwork and Rfree values were 137 and 149 respect-ively showing that the model has a good agreementwith the experimental data RMSDs of Ca atoms be-tween the SFX structure and cryogenic SRX structureswere 502 A Anomalous peaks of T1Cu and T2Cuwere clearly observed (Fig 2a) The ligandT1Cu dis-tances in the SFX structure were the same as those inthe SRX structures within the range of coordinateerrors (Table IV) T2Cu in the SFX structure wascoordinated by three histidine residues WatC andWatD (Fig 2b) The distance from T2Cu to theligand plane was longer than those of the SRX struc-tures (Fig 1c) although it is almost the same as that in
the SR1 structure within the range of coordinateerrors
An unidentified electron density was observed in thevicinity of T2Culigand water molecules in the SFXdata (Fig 2b) We tentatively assigned this to a 40occupancy sodium ion (Na+) from the crystallizationbuffer for the following reasons The peak assigned toNa+ was close to WatB WatC WatD and Asp98(Aspcat) as shown in Fig 2c The distances from theassigned Na+ to water molecules or Asp98 wereshorter than typical hydrogen bonds but longer thantypical covalent bonds in protein crystal structures Wecould rule out a disordered water model because allwater molecules in Fig 2c showed full occupancy Itis therefore reasonable to speculate that the atom atthe peak is a metal ion and forms coordination bondswith water and Aspcat The peak did not show ananomalous peak meaning that it is not Cu Na+ wasthe only metal ion other than Cu in the crystallizationsolution Furthermore Aspcat can easily form a coord-ination bond with a cation because it is deprotonatedin the resting state (60)
Presumably because of the high concentration ofCuSO4 in the crystallization buffer there existed ananomalous peak of 20 occupancy Cu bound toHis244 (Hiscat) in the SFX data (Fig 2aSupplementary Fig S5) Because the Cu atom wasvery close to bridging water (144 A SupplementaryFig S5) we regarded the Cu atom and bridgingwater were alternative structures The occupancy ofbridging water was 80 This anomalous peak of Cuwas also observed in a previously determined cryogenicSRX structure (PDB code 3X1E Supplementary FigS6) CuNiR has evolutionary and structural relation-ship with some multicopper oxidases (MCOs) (6164)and Hiscat in CuNiR is superimposed to one of the Hisligands to a trinuclear copper centre in the MCOs (6364) Therefore the binding of extra Cu to Hiscat is nota surprising phenomenon
RT SRX structure of GtNiRCryo-manipulations of protein crystals in SRX canchange the population of amino acid side chains (6566) Due to its thermostability GtNiR macrocrystalscan be used in SRX without cryo-cooling (41) there-fore we determined an RT SRX structure of GtNiR tojudge whether the observed structural differences be-tween cryogenic SRX and SFX structures were derivedfrom X-ray photoreduction and not from the differ-ence of measurement temperatures The RT SRXstructure was refined to 135 A resolution (Table II)The final Rwork and Rfree values were 97 and 120respectively RMSDs of Ca atoms between the RTSRX and cryogenic SRX structures were502 A TheRMSD of Ca atoms between the RT SRX and SFXstructure was 008 A
The T2Cu in the RT SRX structure was coordinatedby three His ligands and two water molecules(Supplementary Fig S7) The distance from T2Cu tothe ligand plane was shorter than that of the SR1structure and longer than those of other SRX struc-tures (Fig 1c) albeit within the range of errorsHowever it is obvious that the distance was shorter
Table III Data collection and refinement statistics for the SFX
structure
Data collectionBeamline SACLA BL3Wavelength (A) 1129Space group R3Unit cell a= b c (A) 1162 8555Resolution range (A) 348143 (147143)Rsplit () 1770 (1138)Completeness () 100 (9997)Unique reflections 79590 (7859)CC12 0970 (00417)5I (I)gt 338 (099)Redundancy 2453 (2070)RefinementResolution range (A) 3480143 (147143)Rwork ()Rfree () 137149No of protein atoms 2414No of heterogen atoms 9No of water molecules 134Average B (A2)All 240Protein atoms 233Water 366T1Cu 204T2Cu 179Other atoms 317Ramachandran plot ()
Favoured 964Allowed 36Outliers 0
PDB code 4YSA
Y Fukuda et al
4
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Fig 1 Structural changes in SRX (a) Changes in the hydration structures at the T2Cu site The 2FoFc maps contoured at 10 are shown ascyan meshes Carbon oxygen nitrogen and copper atoms are yellow red blue and brown respectively (b) The ligand plane composed of threeN2 atoms of His residues at the T2Cu site (yellow-dashed lines) Carbon nitrogen and copper atoms are coloured in bright red blue and brownrespectively (c) Distances from T2Cu to the ligand plane The error bars represent twice the values of the coordinate errors estimated by themaximum likelihood method (A colour version of this figure is available online at httpjboxfordjournalsorg)
Table IV Coordination geometries of the copper sites
SFX revealed redox-coupled structural changes in CuNiR
5
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
than that in the SFX structure The electron densitymap of the RT SRX data has a strong positive peakaround WatC with full occupancy (SupplementaryFig S4c) We also observed an unidentified positiveelectron density peak (Supplementary Fig S4c) at thecorresponding position of the putative Na+ site inthe SFX structure (Supplementary Fig S4b) thoughthe signal was too weak to place any atomic modelthere The observation that both RT SRX and SFXdata show this electron density means at least that itis not the result of photoreduction
The electron density map of the RT SRX datashowed a strong electron density peak near HiscatWhen water is assigned there the modelled moleculewas too close to Hiscat (520 A) Therefore it was likely
to be a Cu ion coordinated by Hiscat (SupplementaryFig S7) as was observed in the SFX structure and the3X1E structure
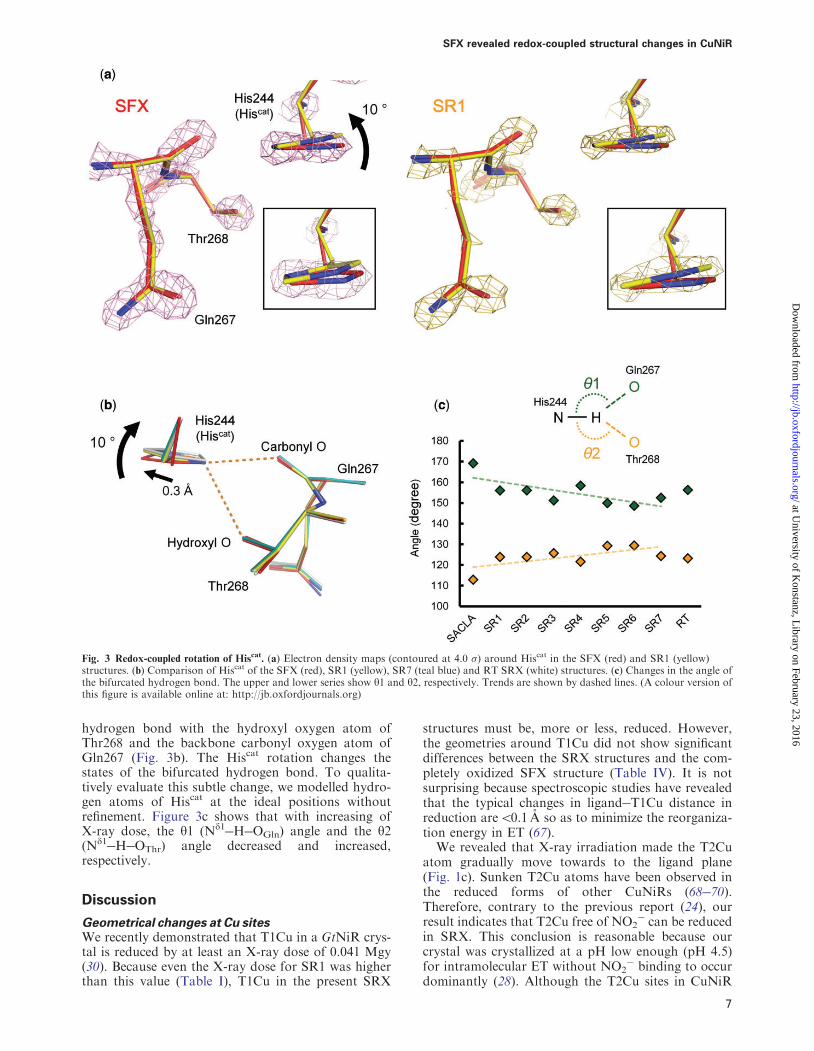
Redox-coupled conformational change in Hiscat
Compared with the SFX structure cryogenic and RTSRX structures revealed that the imidazole ring andthe Cb atom of Hiscat rotated 10 and moved 03 Arespectively (Fig 3a and b Supplementary Fig S8) Toconfirm that it is not the result of cutting the SFX dataat 143 A resolution we refined the SFX structureagainst the data to 165 A resolution where CC12
05 and Rsplit5 100 (Supplementary Fig S2) and theconformation of Hiscat did not change (SupplementaryFig S9) The Nd1 atom of Hiscat forms a bifurcated
Fig 2 SFX structure of GtNiR Carbon oxygen nitrogen sulfur and copper atoms are bright red red blue yellow and brown respectively (a)Copper binding sites in the SFX structure The anomalous Fourier maps are contoured at 40 (magenta) and 12 (dark blue) (b) Hydrationstructure of the T2Cu site in the SFX structure The 2FoFc maps contoured at 10 are shown as cyan meshes Na+ and water molecules areshown as a purple sphere (c) Penta-coordinated Na+ above the T2Cu site Distances are shown in A (A colour version of this figure is availableonline at httpjboxfordjournalsorg)
Y Fukuda et al
6
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
hydrogen bond with the hydroxyl oxygen atom ofThr268 and the backbone carbonyl oxygen atom ofGln267 (Fig 3b) The Hiscat rotation changes thestates of the bifurcated hydrogen bond To qualita-tively evaluate this subtle change we modelled hydro-gen atoms of Hiscat at the ideal positions withoutrefinement Figure 3c shows that with increasing ofX-ray dose the y1 (Nd1HOGln) angle and the y2(Nd1HOThr) angle decreased and increasedrespectively
Discussion
Geometrical changes at Cu sitesWe recently demonstrated that T1Cu in a GtNiR crys-tal is reduced by at least an X-ray dose of 0041 Mgy(30) Because even the X-ray dose for SR1 was higherthan this value (Table I) T1Cu in the present SRX
structures must be more or less reduced Howeverthe geometries around T1Cu did not show significantdifferences between the SRX structures and the com-pletely oxidized SFX structure (Table IV) It is notsurprising because spectroscopic studies have revealedthat the typical changes in ligandT1Cu distance inreduction are501 A so as to minimize the reorganiza-tion energy in ET (67)
We revealed that X-ray irradiation made the T2Cuatom gradually move towards to the ligand plane(Fig 1c) Sunken T2Cu atoms have been observed inthe reduced forms of other CuNiRs (6870)Therefore contrary to the previous report (24) ourresult indicates that T2Cu free of NO2
can be reducedin SRX This conclusion is reasonable because ourcrystal was crystallized at a pH low enough (pH 45)for intramolecular ET without NO2
binding to occurdominantly (28) Although the T2Cu sites in CuNiR
Fig 3 Redox-coupled rotation of Hiscat (a) Electron density maps (contoured at 40 ) around Hiscat in the SFX (red) and SR1 (yellow)
structures (b) Comparison of Hiscat of the SFX (red) SR1 (yellow) SR7 (teal blue) and RT SRX (white) structures (c) Changes in the angle ofthe bifurcated hydrogen bond The upper and lower series show y1 and y2 respectively Trends are shown by dashed lines (A colour version ofthis figure is available online at httpjboxfordjournalsorg)
SFX revealed redox-coupled structural changes in CuNiR
7
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
crystals lose ligand water when it is harshly reduced byartificial reductants (6870) it is just one of thereduced states of T2Cu called inactive reduced stateand a normal reduced state retains water ligand (29) aswas observed in our structures (Fig 1a)
Conformational change of Hiscat
The structural change of Hiscat cannot be caused bycoordination of Cu or the presence of Na+ above theT2Cu site because the former was found also in thecryogenic and RT SR structures (Supplementary FigS6 and S7) and the latter was found also in the RT SRstructure (Supplementary Fig S4c) The difference ofmeasurement temperature is not a major cause of thestructural change (Fig 3b) Thus the conformationaldifference in Hiscat between the SFX and SRX struc-tures is most likely to result from photoreduction ofthe Cu sites Indeed the observed conformationalchange was small and is probably due to the rigid cata-lytic site of thermostable GtNiR (71)
Interestingly there is a hydrogen bond betweenT2Cu ligand His100 and the side chain of Gln267
and His100 is located at the end of a sensor loop(Fig 4a) which is thought to transmit informationabout T1Cursquos redox state to T2Cu (8 72) Althoughthe ThrGln pair (or ThrGlu in some CuNiRs) com-posing the bifurcated hydrogen bond has been ignoredfor long they are conserved in CuNiRs except for afew CuNiRs which have Ser at the position of Thr(Fig 4b) The nitrite reduction activity of Ser-contain-ing CuNiR is lower than the activity of Thr-containingCuNiR (70 73) The crystal structures of Ser-contain-ing CuNiRs show that the Ser residue does not alwaysform a hydrogen bond with Hiscat (70 73) These factsimply the enzymatic importance of the hydrogen bondbetween Hiscat and Thr
The rotation of the imidazole ring changes the yangles of the bifurcated hydrogen bond (Fig 3c)Generally the y angle is used as an indicator of hydro-gen bond strength Strong moderate and weak hydro-gen bonds tend to show 170180 150180 90150
y angles respectively (74) Therefore the Hiscat rota-tion strengthens the hydrogen bond between Hiscat andThr and weakens the hydrogen bond between Hiscat
Fig 4 Conserved hydrogen bond network (a) Redox sensor loop in GtNiR The hydrogen bond network is shown by dotted lines (b) Amino acidsequences of CuNiRs Hiscat GlnGlu and ThrSer are indicated by filled circle triangle and diamond respectively Ax Ac Rs Rp Ph and Hdmeans Achromobacter xylosoxidans Achromobacter cycloclastes Rhodobacter sphaeroides Ralstonia pickettii Pseudoalteromonas haloplanktisand Hyphomicrobium denitrificans respectively (A colour version of this figure is available online at httpjboxfordjournalsorg)
Y Fukuda et al
8
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
and GlnGlu Because the hydroxyl oxygen atom is lessnegatively polarized than the carbonyl oxygen atomthis structural change may destabilize the positivecharge on Hiscat which may facilitate transfer of aproton to the bridging water between Aspcat andHiscat (Fig 4a) Hiscat has been suggested to directlyprovide a proton to NO2
(19 75) however our re-sults indicate another possibility Hiscat may functionas a switch for proton relay
Supplementary Data
Supplementary Data are available at JB Online
Acknowledgements
The SFX experiment was carried out at BL3 of the SPring-8Angstrom Compact Free-Electron Laser (SACLA) with the ap-proval of the Japan Synchrotron Radiation Research Institute(JASRI) (proposal no 2013B8045) We are grateful for supportfrom the SACLA High Performance Computing (HPC) systemand the Mini-K super computer system The microfocus SRX ex-periment at BL32XU of SPring-8 was supported by the Platform forDrug Discovery Informatics and Structural Life Science The au-thors thank MEP Murphy for discussion K Baba and N Mizunofor their help in the experiment at BL38B1 (proposal no2013A1592) E Yamashita and A Higashiura for their support atBL44XU of the SPring-8 and all staff at the SACLA for technicalassistance
FundingThis work was supported by the X-ray Free-Electron Laser PriorityStrategy Program of the Ministry of Education Culture SportsScience and Technology in Japan (MEXT) the Grant-in-Aid forScientific Research on Innovative Areas from MEXT the Grant-in-Aid for Japan Society for the Promotion of Science (JSPS)Research Fellows (Grant no 254626) and the JSPS KAKENHI(Grant no 15K18487)
Conflict of InterestNone declared
References
1 Gruber N and Galloway JN (2008) An Earth-systemperspective of the global nitrogen cycle Nature 451293296
2 Galloway JN Townsend AR Erisman JWBekunda M Cai Z Freney JR Martinelli LASeitzinger SP and Sutton MA (2008)Transformation of the nitrogen cycle recent trendsquestions and potential solutions Science 320 889892
3 Zumft WG (1997) Cell biology and molecular basis ofdenitrification Microbiol Mol Biol Rev 61 533616
4 Tavares P Pereira AS Moura JJ and Moura I(2006) Metalloenzymes of the denitrification pathwayJ Inorg Biochem 100 20872100
5 Fulop V Moir JWB Ferguson SJ and Hajdu J(1995) The anatomy of a bifunctional enzyme structuralbasis for reduction of oxygen to water and synthesis ofnitric oxide by cytochrome cd1 Cell 81 369377
6 Kakutani T Watanabe H Arima K and Beppu T(1981) A blue protein as inactivating factor for nitritereductase from Alcaligenes faecalis Strain S-6J Biochem 89 463472
7 MacPherson IS Rosell FI Scofield M MaukAG and Murphy ME (2010) Directed evolution of
copper nitrite reductase to a chromogenic reductantProtein Eng Des Sel 23 137145
8 Strange RW Murphy LM Dodd FE AbrahamZHL Eady RR Smith BE and Hasnain SS(1999) Structural and kinetic evidence for an orderedmechanism of copper nitrite reductase J Mol Biol287 10011009
9 Godden J Turley S Teller D Adman E Liu MPayne W and LeGall J (1991) The 23 angstrom X-raystructure of nitrite reductase from Achromobacter cyclo-clastes Science 253 438442
10 Kukimoto M Nishiyama M Murphy MEPTurley S Adman ET Horinouchi S and BeppuT (1994) X-ray structure and site-directed mutagenesisof a nitrite reductase from Alcaligenes Faecalis S-6 rolesof two copper atoms in nitrite reduction Biochemistry33 52465252
11 Dodd FE Beeumen JV Eady RR and HasnainSS (1998) X-ray structure of a blue-copper nitrite reduc-tase in two crystal forms The nature of the copper sitesmode of substrate binding and recognition by redox part-ner J Mol Biol 282 369382
12 Inoue T Gotowda M Deligeer Kataoka KYamaguchi K Suzuki S Watanabe H Gohow Mand Kai Y (1998) Type 1 Cu structure of blue nitritereductase from Alcaligenes xylosoxidans GIFU 1051 at205 A resolution comparison of blue and green nitritereductases J Biochem 124 876879
13 Merkle AC and Lehnert N (2012) Binding and acti-vation of nitrite and nitric oxide by copper nitrite reduc-tase and corresponding model complexes Dalton Trans41 33553368
14 Koteishi H Nojiri M Nakagami T Yamaguchi Kand Suzuki S (2009) Cytochrome c551 is a mediator ofelectron transfer between copper-containing nitrite re-ductase and azurin in a denitrifying bacteriumAchromobacter xylosoxidans Bull Chem Soc Jpn 8210031005
15 Nojiri M Koteishi H Nakagami T Kobayashi KInoue T Yamaguchi K and Suzuki S (2009)Structural basis of inter-protein electron transfer for ni-trite reduction in denitrification Nature 462 117120
16 Vlasie MD Fernandez-Busnadiego R Prudencio Mand Ubbink M (2008) Conformation of pseudoazurinin the 152 kDa electron transfer complex with nitrite re-ductase determined by paramagnetic NMR J Mol Biol375 14051415
17 Impagliazzo A Blok AJ Cliff MJ Ladbury JEand Ubbink M (2007) Redox-state-dependent complexformation between pseudoazurin and nitrite reductaseJ Am Chem Soc 129 226233
18 Boulanger MJ Kukimoto M Nishiyama MHorinouchi S and Murphy ME (2000) Catalyticroles for two water bridged residues (Asp-98 and His-255) in the active site of copper-containing nitrite reduc-tase J Biol Chem 275 2395723964
19 Kataoka K Furusawa H Takagi K Yamaguchi Kand Suzuki S (2000) Functional analysis of conservedaspartate and histidine residues located around the type 2copper site of copper-containing nitrite reductaseJ Biochem 127 345350
20 Yano J Kern J Irrgang KD Latimer MJBergmann U Glatzel P Pushkar Y Biesiadka JLoll B Sauer K Messinger J Zouni A andYachandra VK (2005) X-ray damage to the Mn4Cacomplex in single crystals of photosystem II a case
SFX revealed redox-coupled structural changes in CuNiR
9
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
study for metalloprotein crystallography Proc NatlAcad Sci USA 102 1204712052
21 Schlichting I Berendzen J Chu K Stock AMMaves SA Benson DE Sweet RM Ringe DPetsko GA and Sligar SG (2000) The catalytic path-way of cytochrome P450cam at atomic resolutionScience 287 16151622
22 Berglund GI Carlsson GH Smith AT Szoke HHenriksen A and Hajdu J (2002) The catalytic path-way of horseradish peroxidase at high resolution Nature417 463468
23 Beitlich T Kuhnel K Schulze-Briese C ShoemanRL and Schlichting I (2007) Cryoradiolytic reductionof crystalline heme proteins analysis by UV-Vis spec-troscopy and X-ray crystallography J Synch Rad 141123
24 Hough MA Antonyuk SV Strange RW EadyRR and Hasnain SS (2008) Crystallography withonline optical and X-ray absorption spectroscopies dem-onstrates an ordered mechanism in copper nitrite reduc-tase J Mol Biol 378 353361
25 Brenner S Heyes DJ Hay S Hough MA EadyRR Hasnain SS and Scrutton NS (2009)Demonstration of proton-coupled electron transfer inthe copper-containing nitrite reductases J Biol Chem284 2597325983
26 Leferink NG Han C Antonyuk SV Heyes DJRigby SE Hough MA Eady RR Scrutton NSand Hasnain SS (2011) Proton-coupled electron trans-fer in the catalytic cycle of Alcaligenes xylosoxidanscopper-dependent nitrite reductase Biochemistry 5041214131
27 Kobayashi K Tagawa S Deligeer and Suzuki S(1999) The pH-dependent changes of intramolecularelectron transfer on copper-containing nitrite reductaseJ Biochem 126 408412
28 Wijma HJ Jeuken LJ Verbeet MP ArmstrongFA and Canters GW (2006) A random-sequentialmechanism for nitrite binding and active site reductionin copper-containing nitrite reductase J Biol Chem281 1634016346
29 Wijma HJ Jeuken LJC Verbeet MP ArmstrongFA and Canters GW (2007) Protein film voltamme-try of copper-containing nitrite reductase reveals revers-ible inactivation J Am Chem Soc 129 85578565
30 Fukuda Y Tse KM Kado Y Mizohata EMatsumura H and Inoue T (2015) Insights into un-known foreign ligand in copper nitrite reductaseBiochem Biophys Res Commun 464 622628
31 Flot D Mairs T Giraud T Guijarro M LesourdM Rey V van Brussel D Morawe C Borel CHignette O Chavanne J Nurizzo D McSweeneyS and Mitchell E (2010) The ID23-2 structural biologymicrofocus beamline at the ESRF J Synch Rad 17107118
32 Chapman HN Fromme P Barty A White TAKirian RA Aquila A Hunter MS Schulz JDePonte DP Weierstall U Doak RB Maia FRMartin AV Schlichting I Lomb L Coppola NShoeman RL Epp SW Hartmann R Rolles DRudenko A Foucar L Kimmel N WeidenspointnerG Holl P Liang M Barthelmess M Caleman CBoutet S Bogan MJ Krzywinski J Bostedt CBajt S Gumprecht L Rudek B Erk B SchmidtC Homke A Reich C Pietschner D Struder LHauser G Gorke H Ullrich J Herrmann SSchaller G Schopper F Soltau H Kuhnel KU
Messerschmidt M Bozek JD Hau-Riege SPFrank M Hampton CY Sierra RG StarodubD Williams GJ Hajdu J Timneanu N SeibertMM Andreasson J Rocker A Jonsson OSvenda M Stern S Nass K Andritschke RSchroter CD Krasniqi F Bott M Schmidt KEWang X Grotjohann I Holton JM Barends TRNeutze R Marchesini S Fromme R Schorb SRupp D Adolph M Gorkhover T Andersson IHirsemann H Potdevin G Graafsma H NilssonB and Spence JC (2011) Femtosecond X-ray proteinnanocrystallography Nature 470 7377
33 Kern J Alonso-Mori R Hellmich J Tran RHattne J Laksmono H Glockner C Echols NSierra RG Sellberg J Lassalle-Kaiser B GildeaRJ Glatzel P Grosse-Kunstleve RW LatimerMJ McQueen TA DiFiore D Fry ARMesserschmidt M Miahnahri A Schafer DWSeibert MM Sokaras D Weng TC Zwart PHWhite WE Adams PD Bogan MJ Boutet SWilliams GJ Messinger J Sauter NK Zouni ABergmann U Yano J and Yachandra VK (2012)Room temperature femtosecond X-ray diffraction ofphotosystem II microcrystals Proc Natl Acad SciUSA 109 97219726
34 Johansson LC Arnlund D Katona G White TABarty A DePonte DP Shoeman RL WickstrandC Sharma A Williams GJ Aquila A Bogan MJCaleman C Davidsson J Doak RB Frank MFromme R Galli L Grotjohann I Hunter MSKassemeyer S Kirian RA Kupitz C Liang MLomb L Malmerberg E Martin AVMesserschmidt M Nass K Redecke L SeibertMM Sjohamn J Steinbrener J Stellato F WangD Wahlgren WY Weierstall U Westenhoff SZatsepin NA Boutet S Spence JC Schlichting IChapman HN Fromme P and Neutze R (2013)Structure of a photosynthetic reaction centre determinedby serial femtosecond crystallography Nat Commun 42911
35 Kupitz C Basu S Grotjohann I Fromme RZatsepin NA Rendek KN Hunter MSShoeman RL White TA Wang D James DYang JH Cobb DE Reeder B Sierra RG LiuH Barty A Aquila AL Deponte D Kirian RABari S Bergkamp JJ Beyerlein KR Bogan MJCaleman C Chao TC Conrad CE Davis KMFleckenstein H Galli L Hau-Riege SPKassemeyer S Laksmono H Liang M Lomb LMarchesini S Martin AV Messerschmidt MMilathianaki D Nass K Ros A Roy-ChowdhuryS Schmidt K Seibert M Steinbrener J Stellato FYan L Yoon C Moore TA Moore AL PushkarY Williams GJ Boutet S Doak RB WeierstallU Frank M Chapman HN Spence JC andFromme P (2014) Serial time-resolved crystallographyof photosystem II using a femtosecond X-ray laserNature 513 261265
36 Kern J Alonso-Mori R Tran R Hattne J GildeaRJ Echols N Glockner C Hellmich J LaksmonoH Sierra RG Lassalle-Kaiser B Koroidov SLampe A Han G Gul S DiFiore DMilathianaki D Fry AR Miahnahri A SchaferDW Messerschmidt M Seibert MM Koglin JESokaras D Weng T-C Sellberg J Latimer MJGrosse-Kunstleve RW Zwart PH White WEGlatzel P Adams PD Bogan MJ Williams GJBoutet S Messinger J Zouni A Sauter NK
Y Fukuda et al
10
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Yachandra VK Bergmann U and Yano J (2013)Simultaneous femtosecond X-ray spectroscopy and dif-fraction of photosystem II at room temperature Science340 491495
37 Kern J Tran R Alonso-Mori R Koroidov SEchols N Hattne J Ibrahim M Gul SLaksmono H Sierra RG Gildea RJ Han GHellmich J Lassalle-Kaiser B Chatterjee RBrewster AS Stan CA Glockner C Lampe ADiFiore D Milathianaki D Fry AR SeibertMM Koglin JE Gallo E Uhlig J Sokaras DWeng TC Zwart PH Skinner DE Bogan MJMesserschmidt M Glatzel P Williams GJ BoutetS Adams PD Zouni A Messinger J Sauter NKBergmann U Yano J and Yachandra VK (2014)Taking snapshots of photosynthetic water oxidationusing femtosecond X-ray diffraction and spectroscopyNat Commun 5 4371
38 Hirata K Kawano Y Ueno G Hashimoto KMurakami H Hasegawa K Hikima T KumasakaT and Yamamoto M (2013) Achievement of proteinmicro-crystallography at SPring-8 beamline BL32XUJ Phys Conf Ser 425 012002
39 Paithankar KS and Garman EF (2010) Know yourdose RADDOSE Acta Cryst D 66 381388
40 Tanida H Kikuchi A Miura K Takeshita KGoto S Shiro Y and Ishikawa T (2004) XAFS andprotein crystallography beamline BL38B1 at SPring-8AIP Conf Proc 705 486489
41 Fukuda Y and Inoue T (2015) High-temperatureand high-resolution crystallography of thermostablecopper nitrite reductase Chem Commun 5165326535
42 Otwinowski Z and Minor W (1997) Processing of X-ray diffraction data collected in oscillation modeMethods Enzymol 276 307326
43 Vagin A and Teplyakov A (2010) Molecular replace-ment with MOLREP Acta Cryst D 66 2225
44 Emsley P Lohkamp B Scott WG and Cowtan K(2010) Features and development of Coot Acta CrystD 66 486501
45 Murshudov GN Skubak P Lebedev AA PannuNS Steiner RA Nicholls RA Winn MD LongF and Vagin AA (2011) REFMAC5 for the refine-ment of macromolecular crystal structures Acta CrystD 67 355367
46 Winn MD Ballard CC Cowtan KD DodsonEJ Emsley P Evans PR Keegan RM KrissinelEB Leslie AG McCoy A McNicholas SJMurshudov GN Pannu NS Potterton EAPowell HR Read RJ Vagin A and Wilson KS(2011) Overview of the CCP4 suite and current develop-ments Acta Cryst D 67 235242
47 Chen VB Arendall WB III Headd JJ KeedyDA Immormino RM Kapral GJ Murray LWRichardson JS and Richardson DC (2010)MolProbity all-atom structure validation for macromol-ecular crystallography Acta Crystallogr D BiolCrystallogr 66 1221
48 Tono K Togashi T Inubushi Y Sato TKatayama T Ogawa K Ohashi H Kimura HTakahashi S Takeshita K Tomizawa H Goto SIshikawa T and Yabashi M (2013) Beamline experi-mental stations and photon beam diagnostics for thehard x-ray free electron laser of SACLA New J Phys15 083035
49 Son S-K Chapman HN and Santra R (2011)Multiwavelength anomalous diffraction at high X-rayintensity Phys Rev Lett 107 218102
50 Nass K Foucar L Barends TR Hartmann EBotha S Shoeman RL Doak RB Alonso-MoriR Aquila A Bajt S Barty A Bean R BeyerleinKR Bublitz M Drachmann N Gregersen JJonsson HO Kabsch W Kassemeyer S KoglinJE Krumrey M Mattle D Messerschmidt MNissen P Reinhard L Sitsel O Sokaras DWilliams GJ Hau-Riege S Timneanu N CalemanC Chapman HN Boutet S and Schlichting I (2015)Indications of radiation damage in ferredoxin microcrys-tals using high-intensity X-FEL beams J Synch Rad22 225238
51 Sugahara M Mizohata E Nango E Suzuki MTanaka T Masuda T Tanaka R Shimamura TTanaka Y Suno C Ihara K Pan D KakinouchiK Sugiyama S Murata M Inoue T Tono KSong C Park J Kameshima T Hatsui T Joti YYabashi M and Iwata S (2014) Grease matrix as aversatile carrier of proteins for serial crystallographyNat Methods 12 6163
52 Tono K Nango E Sugahara M Song C Park JTanaka T Tanaka R Joti Y Kameshima T OnoS Hatsui T Mizohata E Suzuki M Shimamura TTanaka Y Iwata S and Yabashi M (2015) Diverseapplication platform for hard X-ray diffraction inSACLA (DAPHNIS) application to serial protein crys-tallography using an X-ray free-electron laser J SynchRad 22 532537
53 Joti Y Kameshima T Yamaga M Sugimoto TOkada K Abe T Furukawa Y Ohata T TanakaR Hatsui T and Yabashi M (2015) Data acquisitionsystem for X-ray free-electron laser experiments atSACLA J Synch Rad 22 571576
54 Yumoto H Mimura H Koyama T Matsuyama STono K Togashi T Inubushi Y Sato T TanakaT Kimura T Yokoyama H Kim J Sano YHachisu Y Yabashi M Ohashi H Ohmori HIshikawa T and Yamauchi K (2012) Focusing of X-ray free-electron laser pulses with reflective optics NatPhotonics 7 4347
55 White TA Kirian RA Martin AV Aquila ANass K Barty A and Chapman HN (2012)CrystFEL a software suite for snapshot serial crystallog-raphy J Appl Cryst 45 335341
56 Brehm W and Diederichs K (2014) Breaking theindexing ambiguity in serial crystallography ActaCryst D 70 101109
57 Karplus PA and Diederichs K (2012) Linking crystal-lographic model and data quality Science 33610301033
58 Evans PR and Murshudov GN (2013) How good aremy data and what is the resolution Acta Cryst D 6912041214
59 Bublitz M Nass K Drachmann ND MarkvardsenAJ Gutmann MJ Barends TRM Mattle DShoeman RL Doak RB Boutet SMesserschmidt M Seibert MM Williams GJFoucar L Reinhard L Sitsel O Gregersen JLClausen JD Boesen T Gotfryd K Wang K-TOlesen C Moslashller JV Nissen P and Schlichting I(2015) Structural studies of P-type ATPaseligand com-plexes using an X-ray free-electron laser IUCrJ 2409420
SFX revealed redox-coupled structural changes in CuNiR
11
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046
Y Fukuda et al
12
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
transferred to the T2Cu site the catalytic centrethrough a CysHis pathway The AspHis pair(Aspcat and Hiscat) which is conserved above theT2Cu site and connected via a water molecule (brid-ging water) is essential to the enzymatic activity (1819) though the exact role has been ambiguous
In conventional synchrotron radiation crystallog-raphy (SRX) strong X-ray beams induce photoreduc-tion of metal centres and destroy their naturalstructures (2023) Spectroscopic analysis revealedthat T1Cu in CuNiR is rapidly reduced by synchrotronX-ray (24) T2Cu is more resistant than T1Cu to X-ray damage in the absence of NO2
When the sub-strate binds to T2Cu X-ray photoreduction of T1Cu isfollowed by ET from T1Cu to T2Cu which results inreduction of NO2
in crystallo (24) This gated ET isexplained by the concept of proton-coupled ET (2526) although the detailed mechanism remains to beelucidated Because the presence of NO2
acceleratesintramolecular ET also in solution (2527) it is obvi-ous that ET from T1Cu to T2Cu is gated to someextent However kinetic studies demonstrated therandom sequential mechanism of nitrite reductionie intramolecular ET can occur both with and with-out the binding of the substrate (28 29) Especiallyintramolecular ET before substrate binding is domin-ant at low pH (565) (28) Moreover we have recentlyshown that substrate-free T2Cu in a CuNiR crystalcrystallized at pH 45 may be reduced by synchrotronX-rays and that an unknown chemical reaction occurson T2Cu during data collection (30)
To investigate the nitrite reduction mechanism inCuNiR detailed structural comparison between itsoxidized and reduced state is necessary Here we clo-sely examined X-ray-induced structural changes andchemical reactions at the T2Cu site using a helicalscan method combined with microfocus X-ray beams(31) Furthermore we determined the first completelyoxidized CuNiR structure using serial femtosecondcrystallography (SFX) with X-ray free-electron laser(XFEL) (32) which has enabled damage-free struc-tural determination of metalloenzymes even at roomtemperature (RT) (3337) Because the Bragg spacingsof previously determined SFX structures of metalloen-zymes were longer than typical covalent bond lengthsfound in macromolecules (15 A) it has been difficultto obtain at the chemical level new structural insightsinto the reaction mechanisms of metalloenzymesCuNiR crystals used in this study have been knownto diffract X-rays well therefore we could determineits high-resolution SFX structure We here would liketo report our results in detail
Materials and Methods
Sample preparation of CuNiR from Geobacillusthermodenitrificans (GtNiR)Geobacillus thermodenitrificans copper nitrite reductase (GtNiR) wasexpressed and purified as described previously (30) We used chlor-ide-free buffers for all the steps of purification and crystallizationMicrocrystals for SFX were obtained by a rotational crystallizationtechnique using nanoseeds of the protein as follows Macrocrystalswere transferred to a 15ml tube (Eppendorf Hamburg Germany)containing 1ml of solution composed of 100mM sodium acetate
buffer (pH 45) 55 (wv) polyethylene glycol 4000 and 75mMCuSO4 After sonicating the crystals on ice with a UD-211 ultraso-nicator (Tomy Seiko Co Tokyo Japan) the solution was centri-fuged and the supernatant was collected as a nanoseed solution In a15ml centrifuge tube (AS ONE Co Osaka Japan) 4ml of the20mgml protein solution was mixed with 4ml of the precipitantsolution which was composed of 100mM sodium acetate buffer(pH 45) 15 (wv) polyethylene glycol 4000 75mM CuSO4 andthen 160ml of the nanoseed solution was added The centrifuge tubehad been rotated on a RT-50 culture rotator (TITEC SaitamaJapan) at 30 rpm for 1 week at RT The microcrystal solutionwas filtered through a 30 mm CellTrics filter (Chiyoda Sci CoTokyo Japan) and adjusted to a number density of 44108 crys-talsml by adding 4ml of the precipitant solution
Synchrotron data collectionCryogenic SRX datasets were collected using microfocus beamlineBL32XU at SPring-8 (38) A large single crystal of GtNiR(915620230mm3) was flash-cooled by immersion in liquid nitro-gen and mounted on a conventional goniometer with the longest axisroughly directed towards the horizontal rotation axis Along thelongest edge of the crystal 120 irradiation points were chosen Theregular intervals between the irradiation points were 76 mm which issufficient to separate the radiation damage at each irradiation pointThe beam at a wavelength of 07500 A was focused to 15 mm (heightH)10 mm (width W) with a photon flux of 81011 photonssUsing the helical scan method a total of seven datasets(SR1SR7) were repeatedly collected in order from the samepoints of the same crystal except for absorbed X-ray dose perframe The exposure time for each image was 1 s For datasetsSR1 SR3 SR5 and SR7 each image was collected with an absorbeddose of 0064MGyframe with a 923 attenuated beam while fordatasets SR2 SR4 and SR6 the dose corresponded to 8252MGyframe without attenuation The X-ray doses were calculated withRADDOSE (39) The parameters and statistics are summarized inTable I
RT data collection was performed at BL38B1 of SPring-8 (40) asdescribed previously (41) The dataset was collected from one pos-ition of a single crystal using an ADSC Quantum 315 charge-coupled device (CCD) detector (Area Detector Systems Co CAUSA) The beam size was 50 mm (H)88 mm (W) The oscillationangle and exposure time per image were set to 1 and 15 srespectively A total of 120 diffraction images were collected fromthe single crystal The parameters and statistics are summarized inTable II
Structure determination of the SRX structuresAll of the datasets were indexed and integrated using HKL2000 (42)The phases were determined by the molecular replacement methodusing MOLREP (43) with a GtNiR monomer (Protein Data Bank(PDB) code 4ZK8) as a search model Manual model building wasperformed using WinCoot 07 (44) The program REFMAC5 (45)from the CCP4 suite (ver 650) (46) was used for structural refine-ment Anisotropic displacement parameters were introduced afterwater molecules were built in the models The final models werechecked for stereochemical quality using MolProbity (47)
Single-shot XFEL data collectionThe experiment was performed at BL3 of SPring-8 AngstromCompact Free-Electron Laser (SACLA) in Hyogo Japan (48)Using 210 fs XFEL pulses we collected diffraction patterns fromgreenish-blue (aerobically oxidized) GtNiR microcrystals(Supplementary Fig S1a) The pulse duration shorter than 10 fs isquite important to obtain intact metalloprotein structures becauseultrabright XFEL beams damage electronic structures of heavyatoms in a few tens of femtoseconds (49) and can destroy the naturalstructures of metal centres (50) A liquid injector (nozzle aperturediameter 200mm) with a sample circulation system was used (51)Microcrystal sample (55ml) was placed in a reservoir The flow ratewas set to 53mlmin (70 cms) The injector was installed in a heliumambiance diffraction chamber enclosure Diverse ApplicationPlatform for Hard X-ray Diffraction in SACLA (DAPHNIS) (52)The liquidstream width was nearly the same as the aperture sizeThe sample chamber was maintained at a temperature of 300K witha humidity of 8599 The diffraction patterns were collected usinga short-working-distance octal multiport CCD detector (53) with
Y Fukuda et al
2
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
XFEL radiation The microcrystals were exposed to single X-raypulses at a photon energy of 110 keV The pulses consisted of51010 photonspulse were focused to 25 mm (H)20 mm (W) atthe interaction point using KirkpatrickBaez mirrors (54) Therepetition rate was 30Hz and the typical pulse energy at thesample was 90 mJpulse The parameters and statistics are summar-ized in Table III
Structural determination of the SFX structureA total of 180942 images were collected of which 139391 diffrac-tion images were identified and 37186 images were indexed andmerged using CrystFEL (55) The data was indexed as spacegroup R3 and processed as space group H3 Indexing ambiguity inSFX was solved by an algorithm that clusters snapshots (56) Thephase was determined by molecular replacement using MOLREPwith the GtNiR monomeric unit (PDB code 4ZK8) as a searchmodel Manual model building was performed using WinCoot 07The program REFMAC5 from the CCP4 suite (ver 650) was usedfor structure refinement The microcrystals diffracted X-rays beyond14 A resolution (Supplementary Fig S1b) but the statistics of high-resolution shells such as CC12 and Rsplit were poor (SupplementaryFig S2) We therefore performed paired refinement to determine thelsquonominal resolutionrsquo based on the idea that proper use of weakerand noisier but higher-resolution data can provide better models (5758) Actually inclusion of noisy high-resolution data enables thebetter analysis of SFX data (59) The result of paired refinementshowed that including higher-resolution data provided a bettermodel than rejecting it (Supplementary Fig S3a) The studentrsquos t-test P value calculated from CC12 in the highest resolution shell cutat 143 A resolution (00417 n=7859) was 0000214 which is lessthan the significance level =0001 (Supplementary Fig S3b)Conversely the P value calculated from CC12 in the highest reso-lution shell cut at 142 A resolution (00246 n=8164) was 00261which is greater than (Supplementary Fig S3b) Based on thesethings we chose to use data up to 143 A resolution and performedfurther refinement The final model was checked for stereochemicalquality using MolProbity
Results
Radiation damages to GtNiR in SRXThe structures for SR1 SR3 SR5 and SR7 data wererefined against data to 150 A resolution whereas thosefor SR2 SR4 and SR6 data were refined to 134 Aresolution (Table I) Root-mean-square deviations(RMSDs) for Ca atoms between seven structureswere at most 01 A
Table I Data collection and refinement statistics for the cryogenic SRX structures
NamePDB ID SR14YSO SR24YSP SR34YSQ SR44YSR SR54YSS SR64YST SR74YSU
Data collection at BL32XU of SPring-8 (Wavelength 07500 A)X-ray dose (MGy) 0064 8316 8380 16632 16696 24948 25012Space group R3 R3 R3 R3 R3 R3 R3Unit cell a= b c (A) 1148 8410 1149 8423 1150 8423 1150 8434 1151 8431 1152 8438 1152 8439Resolution (A) 500150
Table II Data collection and refinement statistics for the RT SRX
structure
Data collection at BL38B1 of SPring-8 (Wavelength 09000 A)Space group R3Unit cell a= b c (A) 1162 8555Resolution range (A) 500135 (155135)Rsym () 96 (362)Completeness () 998 (100)Unique reflections 94321 (4709)5I (I)gt 244 (24)Redundancy 42 (36)RefinementResolution (A) 265135 (139135)Rwork ()Rfree () 97120No of protein atoms 2574No of ligand atoms and ions 17No of water molecules 219Average B (A2)All 189Protein atoms 176Water 341T1Cu atom 116T2Cu atom 105Other atoms 268Ramachandran plot ()
Favoured 973Allowed 27Outliers 0
Coordinate error (A) 0020PDB code 4YSD
SFX revealed redox-coupled structural changes in CuNiR
3
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Changes in ligandT1Cu distance fell within therange of coordinate errors (Table IV) indicating thatthe T1Cu site did not show significant structuralchanges that can be detected at resolutions of our pre-sent structures Conversely the T2Cu site showed ob-vious structural changes induced by X-ray irradiationtwo water ligands (WatC and WatD) were present onT2Cu in the SR1 structure while the electron densityof WatD was not observed in other SR data (Fig 1a)Furthermore with increasing X-ray dose the T2Cuatom gradually sank towards a lsquoligand planersquocomposed of three N2 atoms of His ligands (Fig 1band c)
Completely oxidized GtNiR structure determined bySFXThe SFX structure was refined to 143 A resolution(Table III see Materials and Methods) The finalRwork and Rfree values were 137 and 149 respect-ively showing that the model has a good agreementwith the experimental data RMSDs of Ca atoms be-tween the SFX structure and cryogenic SRX structureswere 502 A Anomalous peaks of T1Cu and T2Cuwere clearly observed (Fig 2a) The ligandT1Cu dis-tances in the SFX structure were the same as those inthe SRX structures within the range of coordinateerrors (Table IV) T2Cu in the SFX structure wascoordinated by three histidine residues WatC andWatD (Fig 2b) The distance from T2Cu to theligand plane was longer than those of the SRX struc-tures (Fig 1c) although it is almost the same as that in
the SR1 structure within the range of coordinateerrors
An unidentified electron density was observed in thevicinity of T2Culigand water molecules in the SFXdata (Fig 2b) We tentatively assigned this to a 40occupancy sodium ion (Na+) from the crystallizationbuffer for the following reasons The peak assigned toNa+ was close to WatB WatC WatD and Asp98(Aspcat) as shown in Fig 2c The distances from theassigned Na+ to water molecules or Asp98 wereshorter than typical hydrogen bonds but longer thantypical covalent bonds in protein crystal structures Wecould rule out a disordered water model because allwater molecules in Fig 2c showed full occupancy Itis therefore reasonable to speculate that the atom atthe peak is a metal ion and forms coordination bondswith water and Aspcat The peak did not show ananomalous peak meaning that it is not Cu Na+ wasthe only metal ion other than Cu in the crystallizationsolution Furthermore Aspcat can easily form a coord-ination bond with a cation because it is deprotonatedin the resting state (60)
Presumably because of the high concentration ofCuSO4 in the crystallization buffer there existed ananomalous peak of 20 occupancy Cu bound toHis244 (Hiscat) in the SFX data (Fig 2aSupplementary Fig S5) Because the Cu atom wasvery close to bridging water (144 A SupplementaryFig S5) we regarded the Cu atom and bridgingwater were alternative structures The occupancy ofbridging water was 80 This anomalous peak of Cuwas also observed in a previously determined cryogenicSRX structure (PDB code 3X1E Supplementary FigS6) CuNiR has evolutionary and structural relation-ship with some multicopper oxidases (MCOs) (6164)and Hiscat in CuNiR is superimposed to one of the Hisligands to a trinuclear copper centre in the MCOs (6364) Therefore the binding of extra Cu to Hiscat is nota surprising phenomenon
RT SRX structure of GtNiRCryo-manipulations of protein crystals in SRX canchange the population of amino acid side chains (6566) Due to its thermostability GtNiR macrocrystalscan be used in SRX without cryo-cooling (41) there-fore we determined an RT SRX structure of GtNiR tojudge whether the observed structural differences be-tween cryogenic SRX and SFX structures were derivedfrom X-ray photoreduction and not from the differ-ence of measurement temperatures The RT SRXstructure was refined to 135 A resolution (Table II)The final Rwork and Rfree values were 97 and 120respectively RMSDs of Ca atoms between the RTSRX and cryogenic SRX structures were502 A TheRMSD of Ca atoms between the RT SRX and SFXstructure was 008 A
The T2Cu in the RT SRX structure was coordinatedby three His ligands and two water molecules(Supplementary Fig S7) The distance from T2Cu tothe ligand plane was shorter than that of the SR1structure and longer than those of other SRX struc-tures (Fig 1c) albeit within the range of errorsHowever it is obvious that the distance was shorter
Table III Data collection and refinement statistics for the SFX
structure
Data collectionBeamline SACLA BL3Wavelength (A) 1129Space group R3Unit cell a= b c (A) 1162 8555Resolution range (A) 348143 (147143)Rsplit () 1770 (1138)Completeness () 100 (9997)Unique reflections 79590 (7859)CC12 0970 (00417)5I (I)gt 338 (099)Redundancy 2453 (2070)RefinementResolution range (A) 3480143 (147143)Rwork ()Rfree () 137149No of protein atoms 2414No of heterogen atoms 9No of water molecules 134Average B (A2)All 240Protein atoms 233Water 366T1Cu 204T2Cu 179Other atoms 317Ramachandran plot ()
Favoured 964Allowed 36Outliers 0
PDB code 4YSA
Y Fukuda et al
4
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Fig 1 Structural changes in SRX (a) Changes in the hydration structures at the T2Cu site The 2FoFc maps contoured at 10 are shown ascyan meshes Carbon oxygen nitrogen and copper atoms are yellow red blue and brown respectively (b) The ligand plane composed of threeN2 atoms of His residues at the T2Cu site (yellow-dashed lines) Carbon nitrogen and copper atoms are coloured in bright red blue and brownrespectively (c) Distances from T2Cu to the ligand plane The error bars represent twice the values of the coordinate errors estimated by themaximum likelihood method (A colour version of this figure is available online at httpjboxfordjournalsorg)
Table IV Coordination geometries of the copper sites
SFX revealed redox-coupled structural changes in CuNiR
5
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
than that in the SFX structure The electron densitymap of the RT SRX data has a strong positive peakaround WatC with full occupancy (SupplementaryFig S4c) We also observed an unidentified positiveelectron density peak (Supplementary Fig S4c) at thecorresponding position of the putative Na+ site inthe SFX structure (Supplementary Fig S4b) thoughthe signal was too weak to place any atomic modelthere The observation that both RT SRX and SFXdata show this electron density means at least that itis not the result of photoreduction
The electron density map of the RT SRX datashowed a strong electron density peak near HiscatWhen water is assigned there the modelled moleculewas too close to Hiscat (520 A) Therefore it was likely
to be a Cu ion coordinated by Hiscat (SupplementaryFig S7) as was observed in the SFX structure and the3X1E structure
Redox-coupled conformational change in Hiscat
Compared with the SFX structure cryogenic and RTSRX structures revealed that the imidazole ring andthe Cb atom of Hiscat rotated 10 and moved 03 Arespectively (Fig 3a and b Supplementary Fig S8) Toconfirm that it is not the result of cutting the SFX dataat 143 A resolution we refined the SFX structureagainst the data to 165 A resolution where CC12
05 and Rsplit5 100 (Supplementary Fig S2) and theconformation of Hiscat did not change (SupplementaryFig S9) The Nd1 atom of Hiscat forms a bifurcated
Fig 2 SFX structure of GtNiR Carbon oxygen nitrogen sulfur and copper atoms are bright red red blue yellow and brown respectively (a)Copper binding sites in the SFX structure The anomalous Fourier maps are contoured at 40 (magenta) and 12 (dark blue) (b) Hydrationstructure of the T2Cu site in the SFX structure The 2FoFc maps contoured at 10 are shown as cyan meshes Na+ and water molecules areshown as a purple sphere (c) Penta-coordinated Na+ above the T2Cu site Distances are shown in A (A colour version of this figure is availableonline at httpjboxfordjournalsorg)
Y Fukuda et al
6
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
hydrogen bond with the hydroxyl oxygen atom ofThr268 and the backbone carbonyl oxygen atom ofGln267 (Fig 3b) The Hiscat rotation changes thestates of the bifurcated hydrogen bond To qualita-tively evaluate this subtle change we modelled hydro-gen atoms of Hiscat at the ideal positions withoutrefinement Figure 3c shows that with increasing ofX-ray dose the y1 (Nd1HOGln) angle and the y2(Nd1HOThr) angle decreased and increasedrespectively
Discussion
Geometrical changes at Cu sitesWe recently demonstrated that T1Cu in a GtNiR crys-tal is reduced by at least an X-ray dose of 0041 Mgy(30) Because even the X-ray dose for SR1 was higherthan this value (Table I) T1Cu in the present SRX
structures must be more or less reduced Howeverthe geometries around T1Cu did not show significantdifferences between the SRX structures and the com-pletely oxidized SFX structure (Table IV) It is notsurprising because spectroscopic studies have revealedthat the typical changes in ligandT1Cu distance inreduction are501 A so as to minimize the reorganiza-tion energy in ET (67)
We revealed that X-ray irradiation made the T2Cuatom gradually move towards to the ligand plane(Fig 1c) Sunken T2Cu atoms have been observed inthe reduced forms of other CuNiRs (6870)Therefore contrary to the previous report (24) ourresult indicates that T2Cu free of NO2
can be reducedin SRX This conclusion is reasonable because ourcrystal was crystallized at a pH low enough (pH 45)for intramolecular ET without NO2
binding to occurdominantly (28) Although the T2Cu sites in CuNiR
Fig 3 Redox-coupled rotation of Hiscat (a) Electron density maps (contoured at 40 ) around Hiscat in the SFX (red) and SR1 (yellow)
structures (b) Comparison of Hiscat of the SFX (red) SR1 (yellow) SR7 (teal blue) and RT SRX (white) structures (c) Changes in the angle ofthe bifurcated hydrogen bond The upper and lower series show y1 and y2 respectively Trends are shown by dashed lines (A colour version ofthis figure is available online at httpjboxfordjournalsorg)
SFX revealed redox-coupled structural changes in CuNiR
7
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
crystals lose ligand water when it is harshly reduced byartificial reductants (6870) it is just one of thereduced states of T2Cu called inactive reduced stateand a normal reduced state retains water ligand (29) aswas observed in our structures (Fig 1a)
Conformational change of Hiscat
The structural change of Hiscat cannot be caused bycoordination of Cu or the presence of Na+ above theT2Cu site because the former was found also in thecryogenic and RT SR structures (Supplementary FigS6 and S7) and the latter was found also in the RT SRstructure (Supplementary Fig S4c) The difference ofmeasurement temperature is not a major cause of thestructural change (Fig 3b) Thus the conformationaldifference in Hiscat between the SFX and SRX struc-tures is most likely to result from photoreduction ofthe Cu sites Indeed the observed conformationalchange was small and is probably due to the rigid cata-lytic site of thermostable GtNiR (71)
Interestingly there is a hydrogen bond betweenT2Cu ligand His100 and the side chain of Gln267
and His100 is located at the end of a sensor loop(Fig 4a) which is thought to transmit informationabout T1Cursquos redox state to T2Cu (8 72) Althoughthe ThrGln pair (or ThrGlu in some CuNiRs) com-posing the bifurcated hydrogen bond has been ignoredfor long they are conserved in CuNiRs except for afew CuNiRs which have Ser at the position of Thr(Fig 4b) The nitrite reduction activity of Ser-contain-ing CuNiR is lower than the activity of Thr-containingCuNiR (70 73) The crystal structures of Ser-contain-ing CuNiRs show that the Ser residue does not alwaysform a hydrogen bond with Hiscat (70 73) These factsimply the enzymatic importance of the hydrogen bondbetween Hiscat and Thr
The rotation of the imidazole ring changes the yangles of the bifurcated hydrogen bond (Fig 3c)Generally the y angle is used as an indicator of hydro-gen bond strength Strong moderate and weak hydro-gen bonds tend to show 170180 150180 90150
y angles respectively (74) Therefore the Hiscat rota-tion strengthens the hydrogen bond between Hiscat andThr and weakens the hydrogen bond between Hiscat
Fig 4 Conserved hydrogen bond network (a) Redox sensor loop in GtNiR The hydrogen bond network is shown by dotted lines (b) Amino acidsequences of CuNiRs Hiscat GlnGlu and ThrSer are indicated by filled circle triangle and diamond respectively Ax Ac Rs Rp Ph and Hdmeans Achromobacter xylosoxidans Achromobacter cycloclastes Rhodobacter sphaeroides Ralstonia pickettii Pseudoalteromonas haloplanktisand Hyphomicrobium denitrificans respectively (A colour version of this figure is available online at httpjboxfordjournalsorg)
Y Fukuda et al
8
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
and GlnGlu Because the hydroxyl oxygen atom is lessnegatively polarized than the carbonyl oxygen atomthis structural change may destabilize the positivecharge on Hiscat which may facilitate transfer of aproton to the bridging water between Aspcat andHiscat (Fig 4a) Hiscat has been suggested to directlyprovide a proton to NO2
(19 75) however our re-sults indicate another possibility Hiscat may functionas a switch for proton relay
Supplementary Data
Supplementary Data are available at JB Online
Acknowledgements
The SFX experiment was carried out at BL3 of the SPring-8Angstrom Compact Free-Electron Laser (SACLA) with the ap-proval of the Japan Synchrotron Radiation Research Institute(JASRI) (proposal no 2013B8045) We are grateful for supportfrom the SACLA High Performance Computing (HPC) systemand the Mini-K super computer system The microfocus SRX ex-periment at BL32XU of SPring-8 was supported by the Platform forDrug Discovery Informatics and Structural Life Science The au-thors thank MEP Murphy for discussion K Baba and N Mizunofor their help in the experiment at BL38B1 (proposal no2013A1592) E Yamashita and A Higashiura for their support atBL44XU of the SPring-8 and all staff at the SACLA for technicalassistance
FundingThis work was supported by the X-ray Free-Electron Laser PriorityStrategy Program of the Ministry of Education Culture SportsScience and Technology in Japan (MEXT) the Grant-in-Aid forScientific Research on Innovative Areas from MEXT the Grant-in-Aid for Japan Society for the Promotion of Science (JSPS)Research Fellows (Grant no 254626) and the JSPS KAKENHI(Grant no 15K18487)
Conflict of InterestNone declared
References
1 Gruber N and Galloway JN (2008) An Earth-systemperspective of the global nitrogen cycle Nature 451293296
2 Galloway JN Townsend AR Erisman JWBekunda M Cai Z Freney JR Martinelli LASeitzinger SP and Sutton MA (2008)Transformation of the nitrogen cycle recent trendsquestions and potential solutions Science 320 889892
3 Zumft WG (1997) Cell biology and molecular basis ofdenitrification Microbiol Mol Biol Rev 61 533616
4 Tavares P Pereira AS Moura JJ and Moura I(2006) Metalloenzymes of the denitrification pathwayJ Inorg Biochem 100 20872100
5 Fulop V Moir JWB Ferguson SJ and Hajdu J(1995) The anatomy of a bifunctional enzyme structuralbasis for reduction of oxygen to water and synthesis ofnitric oxide by cytochrome cd1 Cell 81 369377
6 Kakutani T Watanabe H Arima K and Beppu T(1981) A blue protein as inactivating factor for nitritereductase from Alcaligenes faecalis Strain S-6J Biochem 89 463472
7 MacPherson IS Rosell FI Scofield M MaukAG and Murphy ME (2010) Directed evolution of
copper nitrite reductase to a chromogenic reductantProtein Eng Des Sel 23 137145
8 Strange RW Murphy LM Dodd FE AbrahamZHL Eady RR Smith BE and Hasnain SS(1999) Structural and kinetic evidence for an orderedmechanism of copper nitrite reductase J Mol Biol287 10011009
9 Godden J Turley S Teller D Adman E Liu MPayne W and LeGall J (1991) The 23 angstrom X-raystructure of nitrite reductase from Achromobacter cyclo-clastes Science 253 438442
10 Kukimoto M Nishiyama M Murphy MEPTurley S Adman ET Horinouchi S and BeppuT (1994) X-ray structure and site-directed mutagenesisof a nitrite reductase from Alcaligenes Faecalis S-6 rolesof two copper atoms in nitrite reduction Biochemistry33 52465252
11 Dodd FE Beeumen JV Eady RR and HasnainSS (1998) X-ray structure of a blue-copper nitrite reduc-tase in two crystal forms The nature of the copper sitesmode of substrate binding and recognition by redox part-ner J Mol Biol 282 369382
12 Inoue T Gotowda M Deligeer Kataoka KYamaguchi K Suzuki S Watanabe H Gohow Mand Kai Y (1998) Type 1 Cu structure of blue nitritereductase from Alcaligenes xylosoxidans GIFU 1051 at205 A resolution comparison of blue and green nitritereductases J Biochem 124 876879
13 Merkle AC and Lehnert N (2012) Binding and acti-vation of nitrite and nitric oxide by copper nitrite reduc-tase and corresponding model complexes Dalton Trans41 33553368
14 Koteishi H Nojiri M Nakagami T Yamaguchi Kand Suzuki S (2009) Cytochrome c551 is a mediator ofelectron transfer between copper-containing nitrite re-ductase and azurin in a denitrifying bacteriumAchromobacter xylosoxidans Bull Chem Soc Jpn 8210031005
15 Nojiri M Koteishi H Nakagami T Kobayashi KInoue T Yamaguchi K and Suzuki S (2009)Structural basis of inter-protein electron transfer for ni-trite reduction in denitrification Nature 462 117120
16 Vlasie MD Fernandez-Busnadiego R Prudencio Mand Ubbink M (2008) Conformation of pseudoazurinin the 152 kDa electron transfer complex with nitrite re-ductase determined by paramagnetic NMR J Mol Biol375 14051415
17 Impagliazzo A Blok AJ Cliff MJ Ladbury JEand Ubbink M (2007) Redox-state-dependent complexformation between pseudoazurin and nitrite reductaseJ Am Chem Soc 129 226233
18 Boulanger MJ Kukimoto M Nishiyama MHorinouchi S and Murphy ME (2000) Catalyticroles for two water bridged residues (Asp-98 and His-255) in the active site of copper-containing nitrite reduc-tase J Biol Chem 275 2395723964
19 Kataoka K Furusawa H Takagi K Yamaguchi Kand Suzuki S (2000) Functional analysis of conservedaspartate and histidine residues located around the type 2copper site of copper-containing nitrite reductaseJ Biochem 127 345350
20 Yano J Kern J Irrgang KD Latimer MJBergmann U Glatzel P Pushkar Y Biesiadka JLoll B Sauer K Messinger J Zouni A andYachandra VK (2005) X-ray damage to the Mn4Cacomplex in single crystals of photosystem II a case
SFX revealed redox-coupled structural changes in CuNiR
9
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
study for metalloprotein crystallography Proc NatlAcad Sci USA 102 1204712052
21 Schlichting I Berendzen J Chu K Stock AMMaves SA Benson DE Sweet RM Ringe DPetsko GA and Sligar SG (2000) The catalytic path-way of cytochrome P450cam at atomic resolutionScience 287 16151622
22 Berglund GI Carlsson GH Smith AT Szoke HHenriksen A and Hajdu J (2002) The catalytic path-way of horseradish peroxidase at high resolution Nature417 463468
23 Beitlich T Kuhnel K Schulze-Briese C ShoemanRL and Schlichting I (2007) Cryoradiolytic reductionof crystalline heme proteins analysis by UV-Vis spec-troscopy and X-ray crystallography J Synch Rad 141123
24 Hough MA Antonyuk SV Strange RW EadyRR and Hasnain SS (2008) Crystallography withonline optical and X-ray absorption spectroscopies dem-onstrates an ordered mechanism in copper nitrite reduc-tase J Mol Biol 378 353361
25 Brenner S Heyes DJ Hay S Hough MA EadyRR Hasnain SS and Scrutton NS (2009)Demonstration of proton-coupled electron transfer inthe copper-containing nitrite reductases J Biol Chem284 2597325983
26 Leferink NG Han C Antonyuk SV Heyes DJRigby SE Hough MA Eady RR Scrutton NSand Hasnain SS (2011) Proton-coupled electron trans-fer in the catalytic cycle of Alcaligenes xylosoxidanscopper-dependent nitrite reductase Biochemistry 5041214131
27 Kobayashi K Tagawa S Deligeer and Suzuki S(1999) The pH-dependent changes of intramolecularelectron transfer on copper-containing nitrite reductaseJ Biochem 126 408412
28 Wijma HJ Jeuken LJ Verbeet MP ArmstrongFA and Canters GW (2006) A random-sequentialmechanism for nitrite binding and active site reductionin copper-containing nitrite reductase J Biol Chem281 1634016346
29 Wijma HJ Jeuken LJC Verbeet MP ArmstrongFA and Canters GW (2007) Protein film voltamme-try of copper-containing nitrite reductase reveals revers-ible inactivation J Am Chem Soc 129 85578565
30 Fukuda Y Tse KM Kado Y Mizohata EMatsumura H and Inoue T (2015) Insights into un-known foreign ligand in copper nitrite reductaseBiochem Biophys Res Commun 464 622628
31 Flot D Mairs T Giraud T Guijarro M LesourdM Rey V van Brussel D Morawe C Borel CHignette O Chavanne J Nurizzo D McSweeneyS and Mitchell E (2010) The ID23-2 structural biologymicrofocus beamline at the ESRF J Synch Rad 17107118
32 Chapman HN Fromme P Barty A White TAKirian RA Aquila A Hunter MS Schulz JDePonte DP Weierstall U Doak RB Maia FRMartin AV Schlichting I Lomb L Coppola NShoeman RL Epp SW Hartmann R Rolles DRudenko A Foucar L Kimmel N WeidenspointnerG Holl P Liang M Barthelmess M Caleman CBoutet S Bogan MJ Krzywinski J Bostedt CBajt S Gumprecht L Rudek B Erk B SchmidtC Homke A Reich C Pietschner D Struder LHauser G Gorke H Ullrich J Herrmann SSchaller G Schopper F Soltau H Kuhnel KU
Messerschmidt M Bozek JD Hau-Riege SPFrank M Hampton CY Sierra RG StarodubD Williams GJ Hajdu J Timneanu N SeibertMM Andreasson J Rocker A Jonsson OSvenda M Stern S Nass K Andritschke RSchroter CD Krasniqi F Bott M Schmidt KEWang X Grotjohann I Holton JM Barends TRNeutze R Marchesini S Fromme R Schorb SRupp D Adolph M Gorkhover T Andersson IHirsemann H Potdevin G Graafsma H NilssonB and Spence JC (2011) Femtosecond X-ray proteinnanocrystallography Nature 470 7377
33 Kern J Alonso-Mori R Hellmich J Tran RHattne J Laksmono H Glockner C Echols NSierra RG Sellberg J Lassalle-Kaiser B GildeaRJ Glatzel P Grosse-Kunstleve RW LatimerMJ McQueen TA DiFiore D Fry ARMesserschmidt M Miahnahri A Schafer DWSeibert MM Sokaras D Weng TC Zwart PHWhite WE Adams PD Bogan MJ Boutet SWilliams GJ Messinger J Sauter NK Zouni ABergmann U Yano J and Yachandra VK (2012)Room temperature femtosecond X-ray diffraction ofphotosystem II microcrystals Proc Natl Acad SciUSA 109 97219726
34 Johansson LC Arnlund D Katona G White TABarty A DePonte DP Shoeman RL WickstrandC Sharma A Williams GJ Aquila A Bogan MJCaleman C Davidsson J Doak RB Frank MFromme R Galli L Grotjohann I Hunter MSKassemeyer S Kirian RA Kupitz C Liang MLomb L Malmerberg E Martin AVMesserschmidt M Nass K Redecke L SeibertMM Sjohamn J Steinbrener J Stellato F WangD Wahlgren WY Weierstall U Westenhoff SZatsepin NA Boutet S Spence JC Schlichting IChapman HN Fromme P and Neutze R (2013)Structure of a photosynthetic reaction centre determinedby serial femtosecond crystallography Nat Commun 42911
35 Kupitz C Basu S Grotjohann I Fromme RZatsepin NA Rendek KN Hunter MSShoeman RL White TA Wang D James DYang JH Cobb DE Reeder B Sierra RG LiuH Barty A Aquila AL Deponte D Kirian RABari S Bergkamp JJ Beyerlein KR Bogan MJCaleman C Chao TC Conrad CE Davis KMFleckenstein H Galli L Hau-Riege SPKassemeyer S Laksmono H Liang M Lomb LMarchesini S Martin AV Messerschmidt MMilathianaki D Nass K Ros A Roy-ChowdhuryS Schmidt K Seibert M Steinbrener J Stellato FYan L Yoon C Moore TA Moore AL PushkarY Williams GJ Boutet S Doak RB WeierstallU Frank M Chapman HN Spence JC andFromme P (2014) Serial time-resolved crystallographyof photosystem II using a femtosecond X-ray laserNature 513 261265
36 Kern J Alonso-Mori R Tran R Hattne J GildeaRJ Echols N Glockner C Hellmich J LaksmonoH Sierra RG Lassalle-Kaiser B Koroidov SLampe A Han G Gul S DiFiore DMilathianaki D Fry AR Miahnahri A SchaferDW Messerschmidt M Seibert MM Koglin JESokaras D Weng T-C Sellberg J Latimer MJGrosse-Kunstleve RW Zwart PH White WEGlatzel P Adams PD Bogan MJ Williams GJBoutet S Messinger J Zouni A Sauter NK
Y Fukuda et al
10
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Yachandra VK Bergmann U and Yano J (2013)Simultaneous femtosecond X-ray spectroscopy and dif-fraction of photosystem II at room temperature Science340 491495
37 Kern J Tran R Alonso-Mori R Koroidov SEchols N Hattne J Ibrahim M Gul SLaksmono H Sierra RG Gildea RJ Han GHellmich J Lassalle-Kaiser B Chatterjee RBrewster AS Stan CA Glockner C Lampe ADiFiore D Milathianaki D Fry AR SeibertMM Koglin JE Gallo E Uhlig J Sokaras DWeng TC Zwart PH Skinner DE Bogan MJMesserschmidt M Glatzel P Williams GJ BoutetS Adams PD Zouni A Messinger J Sauter NKBergmann U Yano J and Yachandra VK (2014)Taking snapshots of photosynthetic water oxidationusing femtosecond X-ray diffraction and spectroscopyNat Commun 5 4371
38 Hirata K Kawano Y Ueno G Hashimoto KMurakami H Hasegawa K Hikima T KumasakaT and Yamamoto M (2013) Achievement of proteinmicro-crystallography at SPring-8 beamline BL32XUJ Phys Conf Ser 425 012002
39 Paithankar KS and Garman EF (2010) Know yourdose RADDOSE Acta Cryst D 66 381388
40 Tanida H Kikuchi A Miura K Takeshita KGoto S Shiro Y and Ishikawa T (2004) XAFS andprotein crystallography beamline BL38B1 at SPring-8AIP Conf Proc 705 486489
41 Fukuda Y and Inoue T (2015) High-temperatureand high-resolution crystallography of thermostablecopper nitrite reductase Chem Commun 5165326535
42 Otwinowski Z and Minor W (1997) Processing of X-ray diffraction data collected in oscillation modeMethods Enzymol 276 307326
43 Vagin A and Teplyakov A (2010) Molecular replace-ment with MOLREP Acta Cryst D 66 2225
44 Emsley P Lohkamp B Scott WG and Cowtan K(2010) Features and development of Coot Acta CrystD 66 486501
45 Murshudov GN Skubak P Lebedev AA PannuNS Steiner RA Nicholls RA Winn MD LongF and Vagin AA (2011) REFMAC5 for the refine-ment of macromolecular crystal structures Acta CrystD 67 355367
46 Winn MD Ballard CC Cowtan KD DodsonEJ Emsley P Evans PR Keegan RM KrissinelEB Leslie AG McCoy A McNicholas SJMurshudov GN Pannu NS Potterton EAPowell HR Read RJ Vagin A and Wilson KS(2011) Overview of the CCP4 suite and current develop-ments Acta Cryst D 67 235242
47 Chen VB Arendall WB III Headd JJ KeedyDA Immormino RM Kapral GJ Murray LWRichardson JS and Richardson DC (2010)MolProbity all-atom structure validation for macromol-ecular crystallography Acta Crystallogr D BiolCrystallogr 66 1221
48 Tono K Togashi T Inubushi Y Sato TKatayama T Ogawa K Ohashi H Kimura HTakahashi S Takeshita K Tomizawa H Goto SIshikawa T and Yabashi M (2013) Beamline experi-mental stations and photon beam diagnostics for thehard x-ray free electron laser of SACLA New J Phys15 083035
49 Son S-K Chapman HN and Santra R (2011)Multiwavelength anomalous diffraction at high X-rayintensity Phys Rev Lett 107 218102
50 Nass K Foucar L Barends TR Hartmann EBotha S Shoeman RL Doak RB Alonso-MoriR Aquila A Bajt S Barty A Bean R BeyerleinKR Bublitz M Drachmann N Gregersen JJonsson HO Kabsch W Kassemeyer S KoglinJE Krumrey M Mattle D Messerschmidt MNissen P Reinhard L Sitsel O Sokaras DWilliams GJ Hau-Riege S Timneanu N CalemanC Chapman HN Boutet S and Schlichting I (2015)Indications of radiation damage in ferredoxin microcrys-tals using high-intensity X-FEL beams J Synch Rad22 225238
51 Sugahara M Mizohata E Nango E Suzuki MTanaka T Masuda T Tanaka R Shimamura TTanaka Y Suno C Ihara K Pan D KakinouchiK Sugiyama S Murata M Inoue T Tono KSong C Park J Kameshima T Hatsui T Joti YYabashi M and Iwata S (2014) Grease matrix as aversatile carrier of proteins for serial crystallographyNat Methods 12 6163
52 Tono K Nango E Sugahara M Song C Park JTanaka T Tanaka R Joti Y Kameshima T OnoS Hatsui T Mizohata E Suzuki M Shimamura TTanaka Y Iwata S and Yabashi M (2015) Diverseapplication platform for hard X-ray diffraction inSACLA (DAPHNIS) application to serial protein crys-tallography using an X-ray free-electron laser J SynchRad 22 532537
53 Joti Y Kameshima T Yamaga M Sugimoto TOkada K Abe T Furukawa Y Ohata T TanakaR Hatsui T and Yabashi M (2015) Data acquisitionsystem for X-ray free-electron laser experiments atSACLA J Synch Rad 22 571576
54 Yumoto H Mimura H Koyama T Matsuyama STono K Togashi T Inubushi Y Sato T TanakaT Kimura T Yokoyama H Kim J Sano YHachisu Y Yabashi M Ohashi H Ohmori HIshikawa T and Yamauchi K (2012) Focusing of X-ray free-electron laser pulses with reflective optics NatPhotonics 7 4347
55 White TA Kirian RA Martin AV Aquila ANass K Barty A and Chapman HN (2012)CrystFEL a software suite for snapshot serial crystallog-raphy J Appl Cryst 45 335341
56 Brehm W and Diederichs K (2014) Breaking theindexing ambiguity in serial crystallography ActaCryst D 70 101109
57 Karplus PA and Diederichs K (2012) Linking crystal-lographic model and data quality Science 33610301033
58 Evans PR and Murshudov GN (2013) How good aremy data and what is the resolution Acta Cryst D 6912041214
59 Bublitz M Nass K Drachmann ND MarkvardsenAJ Gutmann MJ Barends TRM Mattle DShoeman RL Doak RB Boutet SMesserschmidt M Seibert MM Williams GJFoucar L Reinhard L Sitsel O Gregersen JLClausen JD Boesen T Gotfryd K Wang K-TOlesen C Moslashller JV Nissen P and Schlichting I(2015) Structural studies of P-type ATPaseligand com-plexes using an X-ray free-electron laser IUCrJ 2409420
SFX revealed redox-coupled structural changes in CuNiR
11
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046
Y Fukuda et al
12
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
XFEL radiation The microcrystals were exposed to single X-raypulses at a photon energy of 110 keV The pulses consisted of51010 photonspulse were focused to 25 mm (H)20 mm (W) atthe interaction point using KirkpatrickBaez mirrors (54) Therepetition rate was 30Hz and the typical pulse energy at thesample was 90 mJpulse The parameters and statistics are summar-ized in Table III
Structural determination of the SFX structureA total of 180942 images were collected of which 139391 diffrac-tion images were identified and 37186 images were indexed andmerged using CrystFEL (55) The data was indexed as spacegroup R3 and processed as space group H3 Indexing ambiguity inSFX was solved by an algorithm that clusters snapshots (56) Thephase was determined by molecular replacement using MOLREPwith the GtNiR monomeric unit (PDB code 4ZK8) as a searchmodel Manual model building was performed using WinCoot 07The program REFMAC5 from the CCP4 suite (ver 650) was usedfor structure refinement The microcrystals diffracted X-rays beyond14 A resolution (Supplementary Fig S1b) but the statistics of high-resolution shells such as CC12 and Rsplit were poor (SupplementaryFig S2) We therefore performed paired refinement to determine thelsquonominal resolutionrsquo based on the idea that proper use of weakerand noisier but higher-resolution data can provide better models (5758) Actually inclusion of noisy high-resolution data enables thebetter analysis of SFX data (59) The result of paired refinementshowed that including higher-resolution data provided a bettermodel than rejecting it (Supplementary Fig S3a) The studentrsquos t-test P value calculated from CC12 in the highest resolution shell cutat 143 A resolution (00417 n=7859) was 0000214 which is lessthan the significance level =0001 (Supplementary Fig S3b)Conversely the P value calculated from CC12 in the highest reso-lution shell cut at 142 A resolution (00246 n=8164) was 00261which is greater than (Supplementary Fig S3b) Based on thesethings we chose to use data up to 143 A resolution and performedfurther refinement The final model was checked for stereochemicalquality using MolProbity
Results
Radiation damages to GtNiR in SRXThe structures for SR1 SR3 SR5 and SR7 data wererefined against data to 150 A resolution whereas thosefor SR2 SR4 and SR6 data were refined to 134 Aresolution (Table I) Root-mean-square deviations(RMSDs) for Ca atoms between seven structureswere at most 01 A
Table I Data collection and refinement statistics for the cryogenic SRX structures
NamePDB ID SR14YSO SR24YSP SR34YSQ SR44YSR SR54YSS SR64YST SR74YSU
Data collection at BL32XU of SPring-8 (Wavelength 07500 A)X-ray dose (MGy) 0064 8316 8380 16632 16696 24948 25012Space group R3 R3 R3 R3 R3 R3 R3Unit cell a= b c (A) 1148 8410 1149 8423 1150 8423 1150 8434 1151 8431 1152 8438 1152 8439Resolution (A) 500150
Table II Data collection and refinement statistics for the RT SRX
structure
Data collection at BL38B1 of SPring-8 (Wavelength 09000 A)Space group R3Unit cell a= b c (A) 1162 8555Resolution range (A) 500135 (155135)Rsym () 96 (362)Completeness () 998 (100)Unique reflections 94321 (4709)5I (I)gt 244 (24)Redundancy 42 (36)RefinementResolution (A) 265135 (139135)Rwork ()Rfree () 97120No of protein atoms 2574No of ligand atoms and ions 17No of water molecules 219Average B (A2)All 189Protein atoms 176Water 341T1Cu atom 116T2Cu atom 105Other atoms 268Ramachandran plot ()
Favoured 973Allowed 27Outliers 0
Coordinate error (A) 0020PDB code 4YSD
SFX revealed redox-coupled structural changes in CuNiR
3
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Changes in ligandT1Cu distance fell within therange of coordinate errors (Table IV) indicating thatthe T1Cu site did not show significant structuralchanges that can be detected at resolutions of our pre-sent structures Conversely the T2Cu site showed ob-vious structural changes induced by X-ray irradiationtwo water ligands (WatC and WatD) were present onT2Cu in the SR1 structure while the electron densityof WatD was not observed in other SR data (Fig 1a)Furthermore with increasing X-ray dose the T2Cuatom gradually sank towards a lsquoligand planersquocomposed of three N2 atoms of His ligands (Fig 1band c)
Completely oxidized GtNiR structure determined bySFXThe SFX structure was refined to 143 A resolution(Table III see Materials and Methods) The finalRwork and Rfree values were 137 and 149 respect-ively showing that the model has a good agreementwith the experimental data RMSDs of Ca atoms be-tween the SFX structure and cryogenic SRX structureswere 502 A Anomalous peaks of T1Cu and T2Cuwere clearly observed (Fig 2a) The ligandT1Cu dis-tances in the SFX structure were the same as those inthe SRX structures within the range of coordinateerrors (Table IV) T2Cu in the SFX structure wascoordinated by three histidine residues WatC andWatD (Fig 2b) The distance from T2Cu to theligand plane was longer than those of the SRX struc-tures (Fig 1c) although it is almost the same as that in
the SR1 structure within the range of coordinateerrors
An unidentified electron density was observed in thevicinity of T2Culigand water molecules in the SFXdata (Fig 2b) We tentatively assigned this to a 40occupancy sodium ion (Na+) from the crystallizationbuffer for the following reasons The peak assigned toNa+ was close to WatB WatC WatD and Asp98(Aspcat) as shown in Fig 2c The distances from theassigned Na+ to water molecules or Asp98 wereshorter than typical hydrogen bonds but longer thantypical covalent bonds in protein crystal structures Wecould rule out a disordered water model because allwater molecules in Fig 2c showed full occupancy Itis therefore reasonable to speculate that the atom atthe peak is a metal ion and forms coordination bondswith water and Aspcat The peak did not show ananomalous peak meaning that it is not Cu Na+ wasthe only metal ion other than Cu in the crystallizationsolution Furthermore Aspcat can easily form a coord-ination bond with a cation because it is deprotonatedin the resting state (60)
Presumably because of the high concentration ofCuSO4 in the crystallization buffer there existed ananomalous peak of 20 occupancy Cu bound toHis244 (Hiscat) in the SFX data (Fig 2aSupplementary Fig S5) Because the Cu atom wasvery close to bridging water (144 A SupplementaryFig S5) we regarded the Cu atom and bridgingwater were alternative structures The occupancy ofbridging water was 80 This anomalous peak of Cuwas also observed in a previously determined cryogenicSRX structure (PDB code 3X1E Supplementary FigS6) CuNiR has evolutionary and structural relation-ship with some multicopper oxidases (MCOs) (6164)and Hiscat in CuNiR is superimposed to one of the Hisligands to a trinuclear copper centre in the MCOs (6364) Therefore the binding of extra Cu to Hiscat is nota surprising phenomenon
RT SRX structure of GtNiRCryo-manipulations of protein crystals in SRX canchange the population of amino acid side chains (6566) Due to its thermostability GtNiR macrocrystalscan be used in SRX without cryo-cooling (41) there-fore we determined an RT SRX structure of GtNiR tojudge whether the observed structural differences be-tween cryogenic SRX and SFX structures were derivedfrom X-ray photoreduction and not from the differ-ence of measurement temperatures The RT SRXstructure was refined to 135 A resolution (Table II)The final Rwork and Rfree values were 97 and 120respectively RMSDs of Ca atoms between the RTSRX and cryogenic SRX structures were502 A TheRMSD of Ca atoms between the RT SRX and SFXstructure was 008 A
The T2Cu in the RT SRX structure was coordinatedby three His ligands and two water molecules(Supplementary Fig S7) The distance from T2Cu tothe ligand plane was shorter than that of the SR1structure and longer than those of other SRX struc-tures (Fig 1c) albeit within the range of errorsHowever it is obvious that the distance was shorter
Table III Data collection and refinement statistics for the SFX
structure
Data collectionBeamline SACLA BL3Wavelength (A) 1129Space group R3Unit cell a= b c (A) 1162 8555Resolution range (A) 348143 (147143)Rsplit () 1770 (1138)Completeness () 100 (9997)Unique reflections 79590 (7859)CC12 0970 (00417)5I (I)gt 338 (099)Redundancy 2453 (2070)RefinementResolution range (A) 3480143 (147143)Rwork ()Rfree () 137149No of protein atoms 2414No of heterogen atoms 9No of water molecules 134Average B (A2)All 240Protein atoms 233Water 366T1Cu 204T2Cu 179Other atoms 317Ramachandran plot ()
Favoured 964Allowed 36Outliers 0
PDB code 4YSA
Y Fukuda et al
4
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Fig 1 Structural changes in SRX (a) Changes in the hydration structures at the T2Cu site The 2FoFc maps contoured at 10 are shown ascyan meshes Carbon oxygen nitrogen and copper atoms are yellow red blue and brown respectively (b) The ligand plane composed of threeN2 atoms of His residues at the T2Cu site (yellow-dashed lines) Carbon nitrogen and copper atoms are coloured in bright red blue and brownrespectively (c) Distances from T2Cu to the ligand plane The error bars represent twice the values of the coordinate errors estimated by themaximum likelihood method (A colour version of this figure is available online at httpjboxfordjournalsorg)
Table IV Coordination geometries of the copper sites
SFX revealed redox-coupled structural changes in CuNiR
5
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
than that in the SFX structure The electron densitymap of the RT SRX data has a strong positive peakaround WatC with full occupancy (SupplementaryFig S4c) We also observed an unidentified positiveelectron density peak (Supplementary Fig S4c) at thecorresponding position of the putative Na+ site inthe SFX structure (Supplementary Fig S4b) thoughthe signal was too weak to place any atomic modelthere The observation that both RT SRX and SFXdata show this electron density means at least that itis not the result of photoreduction
The electron density map of the RT SRX datashowed a strong electron density peak near HiscatWhen water is assigned there the modelled moleculewas too close to Hiscat (520 A) Therefore it was likely
to be a Cu ion coordinated by Hiscat (SupplementaryFig S7) as was observed in the SFX structure and the3X1E structure
Redox-coupled conformational change in Hiscat
Compared with the SFX structure cryogenic and RTSRX structures revealed that the imidazole ring andthe Cb atom of Hiscat rotated 10 and moved 03 Arespectively (Fig 3a and b Supplementary Fig S8) Toconfirm that it is not the result of cutting the SFX dataat 143 A resolution we refined the SFX structureagainst the data to 165 A resolution where CC12
05 and Rsplit5 100 (Supplementary Fig S2) and theconformation of Hiscat did not change (SupplementaryFig S9) The Nd1 atom of Hiscat forms a bifurcated
Fig 2 SFX structure of GtNiR Carbon oxygen nitrogen sulfur and copper atoms are bright red red blue yellow and brown respectively (a)Copper binding sites in the SFX structure The anomalous Fourier maps are contoured at 40 (magenta) and 12 (dark blue) (b) Hydrationstructure of the T2Cu site in the SFX structure The 2FoFc maps contoured at 10 are shown as cyan meshes Na+ and water molecules areshown as a purple sphere (c) Penta-coordinated Na+ above the T2Cu site Distances are shown in A (A colour version of this figure is availableonline at httpjboxfordjournalsorg)
Y Fukuda et al
6
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
hydrogen bond with the hydroxyl oxygen atom ofThr268 and the backbone carbonyl oxygen atom ofGln267 (Fig 3b) The Hiscat rotation changes thestates of the bifurcated hydrogen bond To qualita-tively evaluate this subtle change we modelled hydro-gen atoms of Hiscat at the ideal positions withoutrefinement Figure 3c shows that with increasing ofX-ray dose the y1 (Nd1HOGln) angle and the y2(Nd1HOThr) angle decreased and increasedrespectively
Discussion
Geometrical changes at Cu sitesWe recently demonstrated that T1Cu in a GtNiR crys-tal is reduced by at least an X-ray dose of 0041 Mgy(30) Because even the X-ray dose for SR1 was higherthan this value (Table I) T1Cu in the present SRX
structures must be more or less reduced Howeverthe geometries around T1Cu did not show significantdifferences between the SRX structures and the com-pletely oxidized SFX structure (Table IV) It is notsurprising because spectroscopic studies have revealedthat the typical changes in ligandT1Cu distance inreduction are501 A so as to minimize the reorganiza-tion energy in ET (67)
We revealed that X-ray irradiation made the T2Cuatom gradually move towards to the ligand plane(Fig 1c) Sunken T2Cu atoms have been observed inthe reduced forms of other CuNiRs (6870)Therefore contrary to the previous report (24) ourresult indicates that T2Cu free of NO2
can be reducedin SRX This conclusion is reasonable because ourcrystal was crystallized at a pH low enough (pH 45)for intramolecular ET without NO2
binding to occurdominantly (28) Although the T2Cu sites in CuNiR
Fig 3 Redox-coupled rotation of Hiscat (a) Electron density maps (contoured at 40 ) around Hiscat in the SFX (red) and SR1 (yellow)
structures (b) Comparison of Hiscat of the SFX (red) SR1 (yellow) SR7 (teal blue) and RT SRX (white) structures (c) Changes in the angle ofthe bifurcated hydrogen bond The upper and lower series show y1 and y2 respectively Trends are shown by dashed lines (A colour version ofthis figure is available online at httpjboxfordjournalsorg)
SFX revealed redox-coupled structural changes in CuNiR
7
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
crystals lose ligand water when it is harshly reduced byartificial reductants (6870) it is just one of thereduced states of T2Cu called inactive reduced stateand a normal reduced state retains water ligand (29) aswas observed in our structures (Fig 1a)
Conformational change of Hiscat
The structural change of Hiscat cannot be caused bycoordination of Cu or the presence of Na+ above theT2Cu site because the former was found also in thecryogenic and RT SR structures (Supplementary FigS6 and S7) and the latter was found also in the RT SRstructure (Supplementary Fig S4c) The difference ofmeasurement temperature is not a major cause of thestructural change (Fig 3b) Thus the conformationaldifference in Hiscat between the SFX and SRX struc-tures is most likely to result from photoreduction ofthe Cu sites Indeed the observed conformationalchange was small and is probably due to the rigid cata-lytic site of thermostable GtNiR (71)
Interestingly there is a hydrogen bond betweenT2Cu ligand His100 and the side chain of Gln267
and His100 is located at the end of a sensor loop(Fig 4a) which is thought to transmit informationabout T1Cursquos redox state to T2Cu (8 72) Althoughthe ThrGln pair (or ThrGlu in some CuNiRs) com-posing the bifurcated hydrogen bond has been ignoredfor long they are conserved in CuNiRs except for afew CuNiRs which have Ser at the position of Thr(Fig 4b) The nitrite reduction activity of Ser-contain-ing CuNiR is lower than the activity of Thr-containingCuNiR (70 73) The crystal structures of Ser-contain-ing CuNiRs show that the Ser residue does not alwaysform a hydrogen bond with Hiscat (70 73) These factsimply the enzymatic importance of the hydrogen bondbetween Hiscat and Thr
The rotation of the imidazole ring changes the yangles of the bifurcated hydrogen bond (Fig 3c)Generally the y angle is used as an indicator of hydro-gen bond strength Strong moderate and weak hydro-gen bonds tend to show 170180 150180 90150
y angles respectively (74) Therefore the Hiscat rota-tion strengthens the hydrogen bond between Hiscat andThr and weakens the hydrogen bond between Hiscat
Fig 4 Conserved hydrogen bond network (a) Redox sensor loop in GtNiR The hydrogen bond network is shown by dotted lines (b) Amino acidsequences of CuNiRs Hiscat GlnGlu and ThrSer are indicated by filled circle triangle and diamond respectively Ax Ac Rs Rp Ph and Hdmeans Achromobacter xylosoxidans Achromobacter cycloclastes Rhodobacter sphaeroides Ralstonia pickettii Pseudoalteromonas haloplanktisand Hyphomicrobium denitrificans respectively (A colour version of this figure is available online at httpjboxfordjournalsorg)
Y Fukuda et al
8
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
and GlnGlu Because the hydroxyl oxygen atom is lessnegatively polarized than the carbonyl oxygen atomthis structural change may destabilize the positivecharge on Hiscat which may facilitate transfer of aproton to the bridging water between Aspcat andHiscat (Fig 4a) Hiscat has been suggested to directlyprovide a proton to NO2
(19 75) however our re-sults indicate another possibility Hiscat may functionas a switch for proton relay
Supplementary Data
Supplementary Data are available at JB Online
Acknowledgements
The SFX experiment was carried out at BL3 of the SPring-8Angstrom Compact Free-Electron Laser (SACLA) with the ap-proval of the Japan Synchrotron Radiation Research Institute(JASRI) (proposal no 2013B8045) We are grateful for supportfrom the SACLA High Performance Computing (HPC) systemand the Mini-K super computer system The microfocus SRX ex-periment at BL32XU of SPring-8 was supported by the Platform forDrug Discovery Informatics and Structural Life Science The au-thors thank MEP Murphy for discussion K Baba and N Mizunofor their help in the experiment at BL38B1 (proposal no2013A1592) E Yamashita and A Higashiura for their support atBL44XU of the SPring-8 and all staff at the SACLA for technicalassistance
FundingThis work was supported by the X-ray Free-Electron Laser PriorityStrategy Program of the Ministry of Education Culture SportsScience and Technology in Japan (MEXT) the Grant-in-Aid forScientific Research on Innovative Areas from MEXT the Grant-in-Aid for Japan Society for the Promotion of Science (JSPS)Research Fellows (Grant no 254626) and the JSPS KAKENHI(Grant no 15K18487)
Conflict of InterestNone declared
References
1 Gruber N and Galloway JN (2008) An Earth-systemperspective of the global nitrogen cycle Nature 451293296
2 Galloway JN Townsend AR Erisman JWBekunda M Cai Z Freney JR Martinelli LASeitzinger SP and Sutton MA (2008)Transformation of the nitrogen cycle recent trendsquestions and potential solutions Science 320 889892
3 Zumft WG (1997) Cell biology and molecular basis ofdenitrification Microbiol Mol Biol Rev 61 533616
4 Tavares P Pereira AS Moura JJ and Moura I(2006) Metalloenzymes of the denitrification pathwayJ Inorg Biochem 100 20872100
5 Fulop V Moir JWB Ferguson SJ and Hajdu J(1995) The anatomy of a bifunctional enzyme structuralbasis for reduction of oxygen to water and synthesis ofnitric oxide by cytochrome cd1 Cell 81 369377
6 Kakutani T Watanabe H Arima K and Beppu T(1981) A blue protein as inactivating factor for nitritereductase from Alcaligenes faecalis Strain S-6J Biochem 89 463472
7 MacPherson IS Rosell FI Scofield M MaukAG and Murphy ME (2010) Directed evolution of
copper nitrite reductase to a chromogenic reductantProtein Eng Des Sel 23 137145
8 Strange RW Murphy LM Dodd FE AbrahamZHL Eady RR Smith BE and Hasnain SS(1999) Structural and kinetic evidence for an orderedmechanism of copper nitrite reductase J Mol Biol287 10011009
9 Godden J Turley S Teller D Adman E Liu MPayne W and LeGall J (1991) The 23 angstrom X-raystructure of nitrite reductase from Achromobacter cyclo-clastes Science 253 438442
10 Kukimoto M Nishiyama M Murphy MEPTurley S Adman ET Horinouchi S and BeppuT (1994) X-ray structure and site-directed mutagenesisof a nitrite reductase from Alcaligenes Faecalis S-6 rolesof two copper atoms in nitrite reduction Biochemistry33 52465252
11 Dodd FE Beeumen JV Eady RR and HasnainSS (1998) X-ray structure of a blue-copper nitrite reduc-tase in two crystal forms The nature of the copper sitesmode of substrate binding and recognition by redox part-ner J Mol Biol 282 369382
12 Inoue T Gotowda M Deligeer Kataoka KYamaguchi K Suzuki S Watanabe H Gohow Mand Kai Y (1998) Type 1 Cu structure of blue nitritereductase from Alcaligenes xylosoxidans GIFU 1051 at205 A resolution comparison of blue and green nitritereductases J Biochem 124 876879
13 Merkle AC and Lehnert N (2012) Binding and acti-vation of nitrite and nitric oxide by copper nitrite reduc-tase and corresponding model complexes Dalton Trans41 33553368
14 Koteishi H Nojiri M Nakagami T Yamaguchi Kand Suzuki S (2009) Cytochrome c551 is a mediator ofelectron transfer between copper-containing nitrite re-ductase and azurin in a denitrifying bacteriumAchromobacter xylosoxidans Bull Chem Soc Jpn 8210031005
15 Nojiri M Koteishi H Nakagami T Kobayashi KInoue T Yamaguchi K and Suzuki S (2009)Structural basis of inter-protein electron transfer for ni-trite reduction in denitrification Nature 462 117120
16 Vlasie MD Fernandez-Busnadiego R Prudencio Mand Ubbink M (2008) Conformation of pseudoazurinin the 152 kDa electron transfer complex with nitrite re-ductase determined by paramagnetic NMR J Mol Biol375 14051415
17 Impagliazzo A Blok AJ Cliff MJ Ladbury JEand Ubbink M (2007) Redox-state-dependent complexformation between pseudoazurin and nitrite reductaseJ Am Chem Soc 129 226233
18 Boulanger MJ Kukimoto M Nishiyama MHorinouchi S and Murphy ME (2000) Catalyticroles for two water bridged residues (Asp-98 and His-255) in the active site of copper-containing nitrite reduc-tase J Biol Chem 275 2395723964
19 Kataoka K Furusawa H Takagi K Yamaguchi Kand Suzuki S (2000) Functional analysis of conservedaspartate and histidine residues located around the type 2copper site of copper-containing nitrite reductaseJ Biochem 127 345350
20 Yano J Kern J Irrgang KD Latimer MJBergmann U Glatzel P Pushkar Y Biesiadka JLoll B Sauer K Messinger J Zouni A andYachandra VK (2005) X-ray damage to the Mn4Cacomplex in single crystals of photosystem II a case
SFX revealed redox-coupled structural changes in CuNiR
9
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
study for metalloprotein crystallography Proc NatlAcad Sci USA 102 1204712052
21 Schlichting I Berendzen J Chu K Stock AMMaves SA Benson DE Sweet RM Ringe DPetsko GA and Sligar SG (2000) The catalytic path-way of cytochrome P450cam at atomic resolutionScience 287 16151622
22 Berglund GI Carlsson GH Smith AT Szoke HHenriksen A and Hajdu J (2002) The catalytic path-way of horseradish peroxidase at high resolution Nature417 463468
23 Beitlich T Kuhnel K Schulze-Briese C ShoemanRL and Schlichting I (2007) Cryoradiolytic reductionof crystalline heme proteins analysis by UV-Vis spec-troscopy and X-ray crystallography J Synch Rad 141123
24 Hough MA Antonyuk SV Strange RW EadyRR and Hasnain SS (2008) Crystallography withonline optical and X-ray absorption spectroscopies dem-onstrates an ordered mechanism in copper nitrite reduc-tase J Mol Biol 378 353361
25 Brenner S Heyes DJ Hay S Hough MA EadyRR Hasnain SS and Scrutton NS (2009)Demonstration of proton-coupled electron transfer inthe copper-containing nitrite reductases J Biol Chem284 2597325983
26 Leferink NG Han C Antonyuk SV Heyes DJRigby SE Hough MA Eady RR Scrutton NSand Hasnain SS (2011) Proton-coupled electron trans-fer in the catalytic cycle of Alcaligenes xylosoxidanscopper-dependent nitrite reductase Biochemistry 5041214131
27 Kobayashi K Tagawa S Deligeer and Suzuki S(1999) The pH-dependent changes of intramolecularelectron transfer on copper-containing nitrite reductaseJ Biochem 126 408412
28 Wijma HJ Jeuken LJ Verbeet MP ArmstrongFA and Canters GW (2006) A random-sequentialmechanism for nitrite binding and active site reductionin copper-containing nitrite reductase J Biol Chem281 1634016346
29 Wijma HJ Jeuken LJC Verbeet MP ArmstrongFA and Canters GW (2007) Protein film voltamme-try of copper-containing nitrite reductase reveals revers-ible inactivation J Am Chem Soc 129 85578565
30 Fukuda Y Tse KM Kado Y Mizohata EMatsumura H and Inoue T (2015) Insights into un-known foreign ligand in copper nitrite reductaseBiochem Biophys Res Commun 464 622628
31 Flot D Mairs T Giraud T Guijarro M LesourdM Rey V van Brussel D Morawe C Borel CHignette O Chavanne J Nurizzo D McSweeneyS and Mitchell E (2010) The ID23-2 structural biologymicrofocus beamline at the ESRF J Synch Rad 17107118
32 Chapman HN Fromme P Barty A White TAKirian RA Aquila A Hunter MS Schulz JDePonte DP Weierstall U Doak RB Maia FRMartin AV Schlichting I Lomb L Coppola NShoeman RL Epp SW Hartmann R Rolles DRudenko A Foucar L Kimmel N WeidenspointnerG Holl P Liang M Barthelmess M Caleman CBoutet S Bogan MJ Krzywinski J Bostedt CBajt S Gumprecht L Rudek B Erk B SchmidtC Homke A Reich C Pietschner D Struder LHauser G Gorke H Ullrich J Herrmann SSchaller G Schopper F Soltau H Kuhnel KU
Messerschmidt M Bozek JD Hau-Riege SPFrank M Hampton CY Sierra RG StarodubD Williams GJ Hajdu J Timneanu N SeibertMM Andreasson J Rocker A Jonsson OSvenda M Stern S Nass K Andritschke RSchroter CD Krasniqi F Bott M Schmidt KEWang X Grotjohann I Holton JM Barends TRNeutze R Marchesini S Fromme R Schorb SRupp D Adolph M Gorkhover T Andersson IHirsemann H Potdevin G Graafsma H NilssonB and Spence JC (2011) Femtosecond X-ray proteinnanocrystallography Nature 470 7377
33 Kern J Alonso-Mori R Hellmich J Tran RHattne J Laksmono H Glockner C Echols NSierra RG Sellberg J Lassalle-Kaiser B GildeaRJ Glatzel P Grosse-Kunstleve RW LatimerMJ McQueen TA DiFiore D Fry ARMesserschmidt M Miahnahri A Schafer DWSeibert MM Sokaras D Weng TC Zwart PHWhite WE Adams PD Bogan MJ Boutet SWilliams GJ Messinger J Sauter NK Zouni ABergmann U Yano J and Yachandra VK (2012)Room temperature femtosecond X-ray diffraction ofphotosystem II microcrystals Proc Natl Acad SciUSA 109 97219726
34 Johansson LC Arnlund D Katona G White TABarty A DePonte DP Shoeman RL WickstrandC Sharma A Williams GJ Aquila A Bogan MJCaleman C Davidsson J Doak RB Frank MFromme R Galli L Grotjohann I Hunter MSKassemeyer S Kirian RA Kupitz C Liang MLomb L Malmerberg E Martin AVMesserschmidt M Nass K Redecke L SeibertMM Sjohamn J Steinbrener J Stellato F WangD Wahlgren WY Weierstall U Westenhoff SZatsepin NA Boutet S Spence JC Schlichting IChapman HN Fromme P and Neutze R (2013)Structure of a photosynthetic reaction centre determinedby serial femtosecond crystallography Nat Commun 42911
35 Kupitz C Basu S Grotjohann I Fromme RZatsepin NA Rendek KN Hunter MSShoeman RL White TA Wang D James DYang JH Cobb DE Reeder B Sierra RG LiuH Barty A Aquila AL Deponte D Kirian RABari S Bergkamp JJ Beyerlein KR Bogan MJCaleman C Chao TC Conrad CE Davis KMFleckenstein H Galli L Hau-Riege SPKassemeyer S Laksmono H Liang M Lomb LMarchesini S Martin AV Messerschmidt MMilathianaki D Nass K Ros A Roy-ChowdhuryS Schmidt K Seibert M Steinbrener J Stellato FYan L Yoon C Moore TA Moore AL PushkarY Williams GJ Boutet S Doak RB WeierstallU Frank M Chapman HN Spence JC andFromme P (2014) Serial time-resolved crystallographyof photosystem II using a femtosecond X-ray laserNature 513 261265
36 Kern J Alonso-Mori R Tran R Hattne J GildeaRJ Echols N Glockner C Hellmich J LaksmonoH Sierra RG Lassalle-Kaiser B Koroidov SLampe A Han G Gul S DiFiore DMilathianaki D Fry AR Miahnahri A SchaferDW Messerschmidt M Seibert MM Koglin JESokaras D Weng T-C Sellberg J Latimer MJGrosse-Kunstleve RW Zwart PH White WEGlatzel P Adams PD Bogan MJ Williams GJBoutet S Messinger J Zouni A Sauter NK
Y Fukuda et al
10
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Yachandra VK Bergmann U and Yano J (2013)Simultaneous femtosecond X-ray spectroscopy and dif-fraction of photosystem II at room temperature Science340 491495
37 Kern J Tran R Alonso-Mori R Koroidov SEchols N Hattne J Ibrahim M Gul SLaksmono H Sierra RG Gildea RJ Han GHellmich J Lassalle-Kaiser B Chatterjee RBrewster AS Stan CA Glockner C Lampe ADiFiore D Milathianaki D Fry AR SeibertMM Koglin JE Gallo E Uhlig J Sokaras DWeng TC Zwart PH Skinner DE Bogan MJMesserschmidt M Glatzel P Williams GJ BoutetS Adams PD Zouni A Messinger J Sauter NKBergmann U Yano J and Yachandra VK (2014)Taking snapshots of photosynthetic water oxidationusing femtosecond X-ray diffraction and spectroscopyNat Commun 5 4371
38 Hirata K Kawano Y Ueno G Hashimoto KMurakami H Hasegawa K Hikima T KumasakaT and Yamamoto M (2013) Achievement of proteinmicro-crystallography at SPring-8 beamline BL32XUJ Phys Conf Ser 425 012002
39 Paithankar KS and Garman EF (2010) Know yourdose RADDOSE Acta Cryst D 66 381388
40 Tanida H Kikuchi A Miura K Takeshita KGoto S Shiro Y and Ishikawa T (2004) XAFS andprotein crystallography beamline BL38B1 at SPring-8AIP Conf Proc 705 486489
41 Fukuda Y and Inoue T (2015) High-temperatureand high-resolution crystallography of thermostablecopper nitrite reductase Chem Commun 5165326535
42 Otwinowski Z and Minor W (1997) Processing of X-ray diffraction data collected in oscillation modeMethods Enzymol 276 307326
43 Vagin A and Teplyakov A (2010) Molecular replace-ment with MOLREP Acta Cryst D 66 2225
44 Emsley P Lohkamp B Scott WG and Cowtan K(2010) Features and development of Coot Acta CrystD 66 486501
45 Murshudov GN Skubak P Lebedev AA PannuNS Steiner RA Nicholls RA Winn MD LongF and Vagin AA (2011) REFMAC5 for the refine-ment of macromolecular crystal structures Acta CrystD 67 355367
46 Winn MD Ballard CC Cowtan KD DodsonEJ Emsley P Evans PR Keegan RM KrissinelEB Leslie AG McCoy A McNicholas SJMurshudov GN Pannu NS Potterton EAPowell HR Read RJ Vagin A and Wilson KS(2011) Overview of the CCP4 suite and current develop-ments Acta Cryst D 67 235242
47 Chen VB Arendall WB III Headd JJ KeedyDA Immormino RM Kapral GJ Murray LWRichardson JS and Richardson DC (2010)MolProbity all-atom structure validation for macromol-ecular crystallography Acta Crystallogr D BiolCrystallogr 66 1221
48 Tono K Togashi T Inubushi Y Sato TKatayama T Ogawa K Ohashi H Kimura HTakahashi S Takeshita K Tomizawa H Goto SIshikawa T and Yabashi M (2013) Beamline experi-mental stations and photon beam diagnostics for thehard x-ray free electron laser of SACLA New J Phys15 083035
49 Son S-K Chapman HN and Santra R (2011)Multiwavelength anomalous diffraction at high X-rayintensity Phys Rev Lett 107 218102
50 Nass K Foucar L Barends TR Hartmann EBotha S Shoeman RL Doak RB Alonso-MoriR Aquila A Bajt S Barty A Bean R BeyerleinKR Bublitz M Drachmann N Gregersen JJonsson HO Kabsch W Kassemeyer S KoglinJE Krumrey M Mattle D Messerschmidt MNissen P Reinhard L Sitsel O Sokaras DWilliams GJ Hau-Riege S Timneanu N CalemanC Chapman HN Boutet S and Schlichting I (2015)Indications of radiation damage in ferredoxin microcrys-tals using high-intensity X-FEL beams J Synch Rad22 225238
51 Sugahara M Mizohata E Nango E Suzuki MTanaka T Masuda T Tanaka R Shimamura TTanaka Y Suno C Ihara K Pan D KakinouchiK Sugiyama S Murata M Inoue T Tono KSong C Park J Kameshima T Hatsui T Joti YYabashi M and Iwata S (2014) Grease matrix as aversatile carrier of proteins for serial crystallographyNat Methods 12 6163
52 Tono K Nango E Sugahara M Song C Park JTanaka T Tanaka R Joti Y Kameshima T OnoS Hatsui T Mizohata E Suzuki M Shimamura TTanaka Y Iwata S and Yabashi M (2015) Diverseapplication platform for hard X-ray diffraction inSACLA (DAPHNIS) application to serial protein crys-tallography using an X-ray free-electron laser J SynchRad 22 532537
53 Joti Y Kameshima T Yamaga M Sugimoto TOkada K Abe T Furukawa Y Ohata T TanakaR Hatsui T and Yabashi M (2015) Data acquisitionsystem for X-ray free-electron laser experiments atSACLA J Synch Rad 22 571576
54 Yumoto H Mimura H Koyama T Matsuyama STono K Togashi T Inubushi Y Sato T TanakaT Kimura T Yokoyama H Kim J Sano YHachisu Y Yabashi M Ohashi H Ohmori HIshikawa T and Yamauchi K (2012) Focusing of X-ray free-electron laser pulses with reflective optics NatPhotonics 7 4347
55 White TA Kirian RA Martin AV Aquila ANass K Barty A and Chapman HN (2012)CrystFEL a software suite for snapshot serial crystallog-raphy J Appl Cryst 45 335341
56 Brehm W and Diederichs K (2014) Breaking theindexing ambiguity in serial crystallography ActaCryst D 70 101109
57 Karplus PA and Diederichs K (2012) Linking crystal-lographic model and data quality Science 33610301033
58 Evans PR and Murshudov GN (2013) How good aremy data and what is the resolution Acta Cryst D 6912041214
59 Bublitz M Nass K Drachmann ND MarkvardsenAJ Gutmann MJ Barends TRM Mattle DShoeman RL Doak RB Boutet SMesserschmidt M Seibert MM Williams GJFoucar L Reinhard L Sitsel O Gregersen JLClausen JD Boesen T Gotfryd K Wang K-TOlesen C Moslashller JV Nissen P and Schlichting I(2015) Structural studies of P-type ATPaseligand com-plexes using an X-ray free-electron laser IUCrJ 2409420
SFX revealed redox-coupled structural changes in CuNiR
11
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046
Y Fukuda et al
12
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Changes in ligandT1Cu distance fell within therange of coordinate errors (Table IV) indicating thatthe T1Cu site did not show significant structuralchanges that can be detected at resolutions of our pre-sent structures Conversely the T2Cu site showed ob-vious structural changes induced by X-ray irradiationtwo water ligands (WatC and WatD) were present onT2Cu in the SR1 structure while the electron densityof WatD was not observed in other SR data (Fig 1a)Furthermore with increasing X-ray dose the T2Cuatom gradually sank towards a lsquoligand planersquocomposed of three N2 atoms of His ligands (Fig 1band c)
Completely oxidized GtNiR structure determined bySFXThe SFX structure was refined to 143 A resolution(Table III see Materials and Methods) The finalRwork and Rfree values were 137 and 149 respect-ively showing that the model has a good agreementwith the experimental data RMSDs of Ca atoms be-tween the SFX structure and cryogenic SRX structureswere 502 A Anomalous peaks of T1Cu and T2Cuwere clearly observed (Fig 2a) The ligandT1Cu dis-tances in the SFX structure were the same as those inthe SRX structures within the range of coordinateerrors (Table IV) T2Cu in the SFX structure wascoordinated by three histidine residues WatC andWatD (Fig 2b) The distance from T2Cu to theligand plane was longer than those of the SRX struc-tures (Fig 1c) although it is almost the same as that in
the SR1 structure within the range of coordinateerrors
An unidentified electron density was observed in thevicinity of T2Culigand water molecules in the SFXdata (Fig 2b) We tentatively assigned this to a 40occupancy sodium ion (Na+) from the crystallizationbuffer for the following reasons The peak assigned toNa+ was close to WatB WatC WatD and Asp98(Aspcat) as shown in Fig 2c The distances from theassigned Na+ to water molecules or Asp98 wereshorter than typical hydrogen bonds but longer thantypical covalent bonds in protein crystal structures Wecould rule out a disordered water model because allwater molecules in Fig 2c showed full occupancy Itis therefore reasonable to speculate that the atom atthe peak is a metal ion and forms coordination bondswith water and Aspcat The peak did not show ananomalous peak meaning that it is not Cu Na+ wasthe only metal ion other than Cu in the crystallizationsolution Furthermore Aspcat can easily form a coord-ination bond with a cation because it is deprotonatedin the resting state (60)
Presumably because of the high concentration ofCuSO4 in the crystallization buffer there existed ananomalous peak of 20 occupancy Cu bound toHis244 (Hiscat) in the SFX data (Fig 2aSupplementary Fig S5) Because the Cu atom wasvery close to bridging water (144 A SupplementaryFig S5) we regarded the Cu atom and bridgingwater were alternative structures The occupancy ofbridging water was 80 This anomalous peak of Cuwas also observed in a previously determined cryogenicSRX structure (PDB code 3X1E Supplementary FigS6) CuNiR has evolutionary and structural relation-ship with some multicopper oxidases (MCOs) (6164)and Hiscat in CuNiR is superimposed to one of the Hisligands to a trinuclear copper centre in the MCOs (6364) Therefore the binding of extra Cu to Hiscat is nota surprising phenomenon
RT SRX structure of GtNiRCryo-manipulations of protein crystals in SRX canchange the population of amino acid side chains (6566) Due to its thermostability GtNiR macrocrystalscan be used in SRX without cryo-cooling (41) there-fore we determined an RT SRX structure of GtNiR tojudge whether the observed structural differences be-tween cryogenic SRX and SFX structures were derivedfrom X-ray photoreduction and not from the differ-ence of measurement temperatures The RT SRXstructure was refined to 135 A resolution (Table II)The final Rwork and Rfree values were 97 and 120respectively RMSDs of Ca atoms between the RTSRX and cryogenic SRX structures were502 A TheRMSD of Ca atoms between the RT SRX and SFXstructure was 008 A
The T2Cu in the RT SRX structure was coordinatedby three His ligands and two water molecules(Supplementary Fig S7) The distance from T2Cu tothe ligand plane was shorter than that of the SR1structure and longer than those of other SRX struc-tures (Fig 1c) albeit within the range of errorsHowever it is obvious that the distance was shorter
Table III Data collection and refinement statistics for the SFX
structure
Data collectionBeamline SACLA BL3Wavelength (A) 1129Space group R3Unit cell a= b c (A) 1162 8555Resolution range (A) 348143 (147143)Rsplit () 1770 (1138)Completeness () 100 (9997)Unique reflections 79590 (7859)CC12 0970 (00417)5I (I)gt 338 (099)Redundancy 2453 (2070)RefinementResolution range (A) 3480143 (147143)Rwork ()Rfree () 137149No of protein atoms 2414No of heterogen atoms 9No of water molecules 134Average B (A2)All 240Protein atoms 233Water 366T1Cu 204T2Cu 179Other atoms 317Ramachandran plot ()
Favoured 964Allowed 36Outliers 0
PDB code 4YSA
Y Fukuda et al
4
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Fig 1 Structural changes in SRX (a) Changes in the hydration structures at the T2Cu site The 2FoFc maps contoured at 10 are shown ascyan meshes Carbon oxygen nitrogen and copper atoms are yellow red blue and brown respectively (b) The ligand plane composed of threeN2 atoms of His residues at the T2Cu site (yellow-dashed lines) Carbon nitrogen and copper atoms are coloured in bright red blue and brownrespectively (c) Distances from T2Cu to the ligand plane The error bars represent twice the values of the coordinate errors estimated by themaximum likelihood method (A colour version of this figure is available online at httpjboxfordjournalsorg)
Table IV Coordination geometries of the copper sites
SFX revealed redox-coupled structural changes in CuNiR
5
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
than that in the SFX structure The electron densitymap of the RT SRX data has a strong positive peakaround WatC with full occupancy (SupplementaryFig S4c) We also observed an unidentified positiveelectron density peak (Supplementary Fig S4c) at thecorresponding position of the putative Na+ site inthe SFX structure (Supplementary Fig S4b) thoughthe signal was too weak to place any atomic modelthere The observation that both RT SRX and SFXdata show this electron density means at least that itis not the result of photoreduction
The electron density map of the RT SRX datashowed a strong electron density peak near HiscatWhen water is assigned there the modelled moleculewas too close to Hiscat (520 A) Therefore it was likely
to be a Cu ion coordinated by Hiscat (SupplementaryFig S7) as was observed in the SFX structure and the3X1E structure
Redox-coupled conformational change in Hiscat
Compared with the SFX structure cryogenic and RTSRX structures revealed that the imidazole ring andthe Cb atom of Hiscat rotated 10 and moved 03 Arespectively (Fig 3a and b Supplementary Fig S8) Toconfirm that it is not the result of cutting the SFX dataat 143 A resolution we refined the SFX structureagainst the data to 165 A resolution where CC12
05 and Rsplit5 100 (Supplementary Fig S2) and theconformation of Hiscat did not change (SupplementaryFig S9) The Nd1 atom of Hiscat forms a bifurcated
Fig 2 SFX structure of GtNiR Carbon oxygen nitrogen sulfur and copper atoms are bright red red blue yellow and brown respectively (a)Copper binding sites in the SFX structure The anomalous Fourier maps are contoured at 40 (magenta) and 12 (dark blue) (b) Hydrationstructure of the T2Cu site in the SFX structure The 2FoFc maps contoured at 10 are shown as cyan meshes Na+ and water molecules areshown as a purple sphere (c) Penta-coordinated Na+ above the T2Cu site Distances are shown in A (A colour version of this figure is availableonline at httpjboxfordjournalsorg)
Y Fukuda et al
6
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
hydrogen bond with the hydroxyl oxygen atom ofThr268 and the backbone carbonyl oxygen atom ofGln267 (Fig 3b) The Hiscat rotation changes thestates of the bifurcated hydrogen bond To qualita-tively evaluate this subtle change we modelled hydro-gen atoms of Hiscat at the ideal positions withoutrefinement Figure 3c shows that with increasing ofX-ray dose the y1 (Nd1HOGln) angle and the y2(Nd1HOThr) angle decreased and increasedrespectively
Discussion
Geometrical changes at Cu sitesWe recently demonstrated that T1Cu in a GtNiR crys-tal is reduced by at least an X-ray dose of 0041 Mgy(30) Because even the X-ray dose for SR1 was higherthan this value (Table I) T1Cu in the present SRX
structures must be more or less reduced Howeverthe geometries around T1Cu did not show significantdifferences between the SRX structures and the com-pletely oxidized SFX structure (Table IV) It is notsurprising because spectroscopic studies have revealedthat the typical changes in ligandT1Cu distance inreduction are501 A so as to minimize the reorganiza-tion energy in ET (67)
We revealed that X-ray irradiation made the T2Cuatom gradually move towards to the ligand plane(Fig 1c) Sunken T2Cu atoms have been observed inthe reduced forms of other CuNiRs (6870)Therefore contrary to the previous report (24) ourresult indicates that T2Cu free of NO2
can be reducedin SRX This conclusion is reasonable because ourcrystal was crystallized at a pH low enough (pH 45)for intramolecular ET without NO2
binding to occurdominantly (28) Although the T2Cu sites in CuNiR
Fig 3 Redox-coupled rotation of Hiscat (a) Electron density maps (contoured at 40 ) around Hiscat in the SFX (red) and SR1 (yellow)
structures (b) Comparison of Hiscat of the SFX (red) SR1 (yellow) SR7 (teal blue) and RT SRX (white) structures (c) Changes in the angle ofthe bifurcated hydrogen bond The upper and lower series show y1 and y2 respectively Trends are shown by dashed lines (A colour version ofthis figure is available online at httpjboxfordjournalsorg)
SFX revealed redox-coupled structural changes in CuNiR
7
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
crystals lose ligand water when it is harshly reduced byartificial reductants (6870) it is just one of thereduced states of T2Cu called inactive reduced stateand a normal reduced state retains water ligand (29) aswas observed in our structures (Fig 1a)
Conformational change of Hiscat
The structural change of Hiscat cannot be caused bycoordination of Cu or the presence of Na+ above theT2Cu site because the former was found also in thecryogenic and RT SR structures (Supplementary FigS6 and S7) and the latter was found also in the RT SRstructure (Supplementary Fig S4c) The difference ofmeasurement temperature is not a major cause of thestructural change (Fig 3b) Thus the conformationaldifference in Hiscat between the SFX and SRX struc-tures is most likely to result from photoreduction ofthe Cu sites Indeed the observed conformationalchange was small and is probably due to the rigid cata-lytic site of thermostable GtNiR (71)
Interestingly there is a hydrogen bond betweenT2Cu ligand His100 and the side chain of Gln267
and His100 is located at the end of a sensor loop(Fig 4a) which is thought to transmit informationabout T1Cursquos redox state to T2Cu (8 72) Althoughthe ThrGln pair (or ThrGlu in some CuNiRs) com-posing the bifurcated hydrogen bond has been ignoredfor long they are conserved in CuNiRs except for afew CuNiRs which have Ser at the position of Thr(Fig 4b) The nitrite reduction activity of Ser-contain-ing CuNiR is lower than the activity of Thr-containingCuNiR (70 73) The crystal structures of Ser-contain-ing CuNiRs show that the Ser residue does not alwaysform a hydrogen bond with Hiscat (70 73) These factsimply the enzymatic importance of the hydrogen bondbetween Hiscat and Thr
The rotation of the imidazole ring changes the yangles of the bifurcated hydrogen bond (Fig 3c)Generally the y angle is used as an indicator of hydro-gen bond strength Strong moderate and weak hydro-gen bonds tend to show 170180 150180 90150
y angles respectively (74) Therefore the Hiscat rota-tion strengthens the hydrogen bond between Hiscat andThr and weakens the hydrogen bond between Hiscat
Fig 4 Conserved hydrogen bond network (a) Redox sensor loop in GtNiR The hydrogen bond network is shown by dotted lines (b) Amino acidsequences of CuNiRs Hiscat GlnGlu and ThrSer are indicated by filled circle triangle and diamond respectively Ax Ac Rs Rp Ph and Hdmeans Achromobacter xylosoxidans Achromobacter cycloclastes Rhodobacter sphaeroides Ralstonia pickettii Pseudoalteromonas haloplanktisand Hyphomicrobium denitrificans respectively (A colour version of this figure is available online at httpjboxfordjournalsorg)
Y Fukuda et al
8
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
and GlnGlu Because the hydroxyl oxygen atom is lessnegatively polarized than the carbonyl oxygen atomthis structural change may destabilize the positivecharge on Hiscat which may facilitate transfer of aproton to the bridging water between Aspcat andHiscat (Fig 4a) Hiscat has been suggested to directlyprovide a proton to NO2
(19 75) however our re-sults indicate another possibility Hiscat may functionas a switch for proton relay
Supplementary Data
Supplementary Data are available at JB Online
Acknowledgements
The SFX experiment was carried out at BL3 of the SPring-8Angstrom Compact Free-Electron Laser (SACLA) with the ap-proval of the Japan Synchrotron Radiation Research Institute(JASRI) (proposal no 2013B8045) We are grateful for supportfrom the SACLA High Performance Computing (HPC) systemand the Mini-K super computer system The microfocus SRX ex-periment at BL32XU of SPring-8 was supported by the Platform forDrug Discovery Informatics and Structural Life Science The au-thors thank MEP Murphy for discussion K Baba and N Mizunofor their help in the experiment at BL38B1 (proposal no2013A1592) E Yamashita and A Higashiura for their support atBL44XU of the SPring-8 and all staff at the SACLA for technicalassistance
FundingThis work was supported by the X-ray Free-Electron Laser PriorityStrategy Program of the Ministry of Education Culture SportsScience and Technology in Japan (MEXT) the Grant-in-Aid forScientific Research on Innovative Areas from MEXT the Grant-in-Aid for Japan Society for the Promotion of Science (JSPS)Research Fellows (Grant no 254626) and the JSPS KAKENHI(Grant no 15K18487)
Conflict of InterestNone declared
References
1 Gruber N and Galloway JN (2008) An Earth-systemperspective of the global nitrogen cycle Nature 451293296
2 Galloway JN Townsend AR Erisman JWBekunda M Cai Z Freney JR Martinelli LASeitzinger SP and Sutton MA (2008)Transformation of the nitrogen cycle recent trendsquestions and potential solutions Science 320 889892
3 Zumft WG (1997) Cell biology and molecular basis ofdenitrification Microbiol Mol Biol Rev 61 533616
4 Tavares P Pereira AS Moura JJ and Moura I(2006) Metalloenzymes of the denitrification pathwayJ Inorg Biochem 100 20872100
5 Fulop V Moir JWB Ferguson SJ and Hajdu J(1995) The anatomy of a bifunctional enzyme structuralbasis for reduction of oxygen to water and synthesis ofnitric oxide by cytochrome cd1 Cell 81 369377
6 Kakutani T Watanabe H Arima K and Beppu T(1981) A blue protein as inactivating factor for nitritereductase from Alcaligenes faecalis Strain S-6J Biochem 89 463472
7 MacPherson IS Rosell FI Scofield M MaukAG and Murphy ME (2010) Directed evolution of
copper nitrite reductase to a chromogenic reductantProtein Eng Des Sel 23 137145
8 Strange RW Murphy LM Dodd FE AbrahamZHL Eady RR Smith BE and Hasnain SS(1999) Structural and kinetic evidence for an orderedmechanism of copper nitrite reductase J Mol Biol287 10011009
9 Godden J Turley S Teller D Adman E Liu MPayne W and LeGall J (1991) The 23 angstrom X-raystructure of nitrite reductase from Achromobacter cyclo-clastes Science 253 438442
10 Kukimoto M Nishiyama M Murphy MEPTurley S Adman ET Horinouchi S and BeppuT (1994) X-ray structure and site-directed mutagenesisof a nitrite reductase from Alcaligenes Faecalis S-6 rolesof two copper atoms in nitrite reduction Biochemistry33 52465252
11 Dodd FE Beeumen JV Eady RR and HasnainSS (1998) X-ray structure of a blue-copper nitrite reduc-tase in two crystal forms The nature of the copper sitesmode of substrate binding and recognition by redox part-ner J Mol Biol 282 369382
12 Inoue T Gotowda M Deligeer Kataoka KYamaguchi K Suzuki S Watanabe H Gohow Mand Kai Y (1998) Type 1 Cu structure of blue nitritereductase from Alcaligenes xylosoxidans GIFU 1051 at205 A resolution comparison of blue and green nitritereductases J Biochem 124 876879
13 Merkle AC and Lehnert N (2012) Binding and acti-vation of nitrite and nitric oxide by copper nitrite reduc-tase and corresponding model complexes Dalton Trans41 33553368
14 Koteishi H Nojiri M Nakagami T Yamaguchi Kand Suzuki S (2009) Cytochrome c551 is a mediator ofelectron transfer between copper-containing nitrite re-ductase and azurin in a denitrifying bacteriumAchromobacter xylosoxidans Bull Chem Soc Jpn 8210031005
15 Nojiri M Koteishi H Nakagami T Kobayashi KInoue T Yamaguchi K and Suzuki S (2009)Structural basis of inter-protein electron transfer for ni-trite reduction in denitrification Nature 462 117120
16 Vlasie MD Fernandez-Busnadiego R Prudencio Mand Ubbink M (2008) Conformation of pseudoazurinin the 152 kDa electron transfer complex with nitrite re-ductase determined by paramagnetic NMR J Mol Biol375 14051415
17 Impagliazzo A Blok AJ Cliff MJ Ladbury JEand Ubbink M (2007) Redox-state-dependent complexformation between pseudoazurin and nitrite reductaseJ Am Chem Soc 129 226233
18 Boulanger MJ Kukimoto M Nishiyama MHorinouchi S and Murphy ME (2000) Catalyticroles for two water bridged residues (Asp-98 and His-255) in the active site of copper-containing nitrite reduc-tase J Biol Chem 275 2395723964
19 Kataoka K Furusawa H Takagi K Yamaguchi Kand Suzuki S (2000) Functional analysis of conservedaspartate and histidine residues located around the type 2copper site of copper-containing nitrite reductaseJ Biochem 127 345350
20 Yano J Kern J Irrgang KD Latimer MJBergmann U Glatzel P Pushkar Y Biesiadka JLoll B Sauer K Messinger J Zouni A andYachandra VK (2005) X-ray damage to the Mn4Cacomplex in single crystals of photosystem II a case
SFX revealed redox-coupled structural changes in CuNiR
9
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
study for metalloprotein crystallography Proc NatlAcad Sci USA 102 1204712052
21 Schlichting I Berendzen J Chu K Stock AMMaves SA Benson DE Sweet RM Ringe DPetsko GA and Sligar SG (2000) The catalytic path-way of cytochrome P450cam at atomic resolutionScience 287 16151622
22 Berglund GI Carlsson GH Smith AT Szoke HHenriksen A and Hajdu J (2002) The catalytic path-way of horseradish peroxidase at high resolution Nature417 463468
23 Beitlich T Kuhnel K Schulze-Briese C ShoemanRL and Schlichting I (2007) Cryoradiolytic reductionof crystalline heme proteins analysis by UV-Vis spec-troscopy and X-ray crystallography J Synch Rad 141123
24 Hough MA Antonyuk SV Strange RW EadyRR and Hasnain SS (2008) Crystallography withonline optical and X-ray absorption spectroscopies dem-onstrates an ordered mechanism in copper nitrite reduc-tase J Mol Biol 378 353361
25 Brenner S Heyes DJ Hay S Hough MA EadyRR Hasnain SS and Scrutton NS (2009)Demonstration of proton-coupled electron transfer inthe copper-containing nitrite reductases J Biol Chem284 2597325983
26 Leferink NG Han C Antonyuk SV Heyes DJRigby SE Hough MA Eady RR Scrutton NSand Hasnain SS (2011) Proton-coupled electron trans-fer in the catalytic cycle of Alcaligenes xylosoxidanscopper-dependent nitrite reductase Biochemistry 5041214131
27 Kobayashi K Tagawa S Deligeer and Suzuki S(1999) The pH-dependent changes of intramolecularelectron transfer on copper-containing nitrite reductaseJ Biochem 126 408412
28 Wijma HJ Jeuken LJ Verbeet MP ArmstrongFA and Canters GW (2006) A random-sequentialmechanism for nitrite binding and active site reductionin copper-containing nitrite reductase J Biol Chem281 1634016346
29 Wijma HJ Jeuken LJC Verbeet MP ArmstrongFA and Canters GW (2007) Protein film voltamme-try of copper-containing nitrite reductase reveals revers-ible inactivation J Am Chem Soc 129 85578565
30 Fukuda Y Tse KM Kado Y Mizohata EMatsumura H and Inoue T (2015) Insights into un-known foreign ligand in copper nitrite reductaseBiochem Biophys Res Commun 464 622628
31 Flot D Mairs T Giraud T Guijarro M LesourdM Rey V van Brussel D Morawe C Borel CHignette O Chavanne J Nurizzo D McSweeneyS and Mitchell E (2010) The ID23-2 structural biologymicrofocus beamline at the ESRF J Synch Rad 17107118
32 Chapman HN Fromme P Barty A White TAKirian RA Aquila A Hunter MS Schulz JDePonte DP Weierstall U Doak RB Maia FRMartin AV Schlichting I Lomb L Coppola NShoeman RL Epp SW Hartmann R Rolles DRudenko A Foucar L Kimmel N WeidenspointnerG Holl P Liang M Barthelmess M Caleman CBoutet S Bogan MJ Krzywinski J Bostedt CBajt S Gumprecht L Rudek B Erk B SchmidtC Homke A Reich C Pietschner D Struder LHauser G Gorke H Ullrich J Herrmann SSchaller G Schopper F Soltau H Kuhnel KU
Messerschmidt M Bozek JD Hau-Riege SPFrank M Hampton CY Sierra RG StarodubD Williams GJ Hajdu J Timneanu N SeibertMM Andreasson J Rocker A Jonsson OSvenda M Stern S Nass K Andritschke RSchroter CD Krasniqi F Bott M Schmidt KEWang X Grotjohann I Holton JM Barends TRNeutze R Marchesini S Fromme R Schorb SRupp D Adolph M Gorkhover T Andersson IHirsemann H Potdevin G Graafsma H NilssonB and Spence JC (2011) Femtosecond X-ray proteinnanocrystallography Nature 470 7377
33 Kern J Alonso-Mori R Hellmich J Tran RHattne J Laksmono H Glockner C Echols NSierra RG Sellberg J Lassalle-Kaiser B GildeaRJ Glatzel P Grosse-Kunstleve RW LatimerMJ McQueen TA DiFiore D Fry ARMesserschmidt M Miahnahri A Schafer DWSeibert MM Sokaras D Weng TC Zwart PHWhite WE Adams PD Bogan MJ Boutet SWilliams GJ Messinger J Sauter NK Zouni ABergmann U Yano J and Yachandra VK (2012)Room temperature femtosecond X-ray diffraction ofphotosystem II microcrystals Proc Natl Acad SciUSA 109 97219726
34 Johansson LC Arnlund D Katona G White TABarty A DePonte DP Shoeman RL WickstrandC Sharma A Williams GJ Aquila A Bogan MJCaleman C Davidsson J Doak RB Frank MFromme R Galli L Grotjohann I Hunter MSKassemeyer S Kirian RA Kupitz C Liang MLomb L Malmerberg E Martin AVMesserschmidt M Nass K Redecke L SeibertMM Sjohamn J Steinbrener J Stellato F WangD Wahlgren WY Weierstall U Westenhoff SZatsepin NA Boutet S Spence JC Schlichting IChapman HN Fromme P and Neutze R (2013)Structure of a photosynthetic reaction centre determinedby serial femtosecond crystallography Nat Commun 42911
35 Kupitz C Basu S Grotjohann I Fromme RZatsepin NA Rendek KN Hunter MSShoeman RL White TA Wang D James DYang JH Cobb DE Reeder B Sierra RG LiuH Barty A Aquila AL Deponte D Kirian RABari S Bergkamp JJ Beyerlein KR Bogan MJCaleman C Chao TC Conrad CE Davis KMFleckenstein H Galli L Hau-Riege SPKassemeyer S Laksmono H Liang M Lomb LMarchesini S Martin AV Messerschmidt MMilathianaki D Nass K Ros A Roy-ChowdhuryS Schmidt K Seibert M Steinbrener J Stellato FYan L Yoon C Moore TA Moore AL PushkarY Williams GJ Boutet S Doak RB WeierstallU Frank M Chapman HN Spence JC andFromme P (2014) Serial time-resolved crystallographyof photosystem II using a femtosecond X-ray laserNature 513 261265
36 Kern J Alonso-Mori R Tran R Hattne J GildeaRJ Echols N Glockner C Hellmich J LaksmonoH Sierra RG Lassalle-Kaiser B Koroidov SLampe A Han G Gul S DiFiore DMilathianaki D Fry AR Miahnahri A SchaferDW Messerschmidt M Seibert MM Koglin JESokaras D Weng T-C Sellberg J Latimer MJGrosse-Kunstleve RW Zwart PH White WEGlatzel P Adams PD Bogan MJ Williams GJBoutet S Messinger J Zouni A Sauter NK
Y Fukuda et al
10
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Yachandra VK Bergmann U and Yano J (2013)Simultaneous femtosecond X-ray spectroscopy and dif-fraction of photosystem II at room temperature Science340 491495
37 Kern J Tran R Alonso-Mori R Koroidov SEchols N Hattne J Ibrahim M Gul SLaksmono H Sierra RG Gildea RJ Han GHellmich J Lassalle-Kaiser B Chatterjee RBrewster AS Stan CA Glockner C Lampe ADiFiore D Milathianaki D Fry AR SeibertMM Koglin JE Gallo E Uhlig J Sokaras DWeng TC Zwart PH Skinner DE Bogan MJMesserschmidt M Glatzel P Williams GJ BoutetS Adams PD Zouni A Messinger J Sauter NKBergmann U Yano J and Yachandra VK (2014)Taking snapshots of photosynthetic water oxidationusing femtosecond X-ray diffraction and spectroscopyNat Commun 5 4371
38 Hirata K Kawano Y Ueno G Hashimoto KMurakami H Hasegawa K Hikima T KumasakaT and Yamamoto M (2013) Achievement of proteinmicro-crystallography at SPring-8 beamline BL32XUJ Phys Conf Ser 425 012002
39 Paithankar KS and Garman EF (2010) Know yourdose RADDOSE Acta Cryst D 66 381388
40 Tanida H Kikuchi A Miura K Takeshita KGoto S Shiro Y and Ishikawa T (2004) XAFS andprotein crystallography beamline BL38B1 at SPring-8AIP Conf Proc 705 486489
41 Fukuda Y and Inoue T (2015) High-temperatureand high-resolution crystallography of thermostablecopper nitrite reductase Chem Commun 5165326535
42 Otwinowski Z and Minor W (1997) Processing of X-ray diffraction data collected in oscillation modeMethods Enzymol 276 307326
43 Vagin A and Teplyakov A (2010) Molecular replace-ment with MOLREP Acta Cryst D 66 2225
44 Emsley P Lohkamp B Scott WG and Cowtan K(2010) Features and development of Coot Acta CrystD 66 486501
45 Murshudov GN Skubak P Lebedev AA PannuNS Steiner RA Nicholls RA Winn MD LongF and Vagin AA (2011) REFMAC5 for the refine-ment of macromolecular crystal structures Acta CrystD 67 355367
46 Winn MD Ballard CC Cowtan KD DodsonEJ Emsley P Evans PR Keegan RM KrissinelEB Leslie AG McCoy A McNicholas SJMurshudov GN Pannu NS Potterton EAPowell HR Read RJ Vagin A and Wilson KS(2011) Overview of the CCP4 suite and current develop-ments Acta Cryst D 67 235242
47 Chen VB Arendall WB III Headd JJ KeedyDA Immormino RM Kapral GJ Murray LWRichardson JS and Richardson DC (2010)MolProbity all-atom structure validation for macromol-ecular crystallography Acta Crystallogr D BiolCrystallogr 66 1221
48 Tono K Togashi T Inubushi Y Sato TKatayama T Ogawa K Ohashi H Kimura HTakahashi S Takeshita K Tomizawa H Goto SIshikawa T and Yabashi M (2013) Beamline experi-mental stations and photon beam diagnostics for thehard x-ray free electron laser of SACLA New J Phys15 083035
49 Son S-K Chapman HN and Santra R (2011)Multiwavelength anomalous diffraction at high X-rayintensity Phys Rev Lett 107 218102
50 Nass K Foucar L Barends TR Hartmann EBotha S Shoeman RL Doak RB Alonso-MoriR Aquila A Bajt S Barty A Bean R BeyerleinKR Bublitz M Drachmann N Gregersen JJonsson HO Kabsch W Kassemeyer S KoglinJE Krumrey M Mattle D Messerschmidt MNissen P Reinhard L Sitsel O Sokaras DWilliams GJ Hau-Riege S Timneanu N CalemanC Chapman HN Boutet S and Schlichting I (2015)Indications of radiation damage in ferredoxin microcrys-tals using high-intensity X-FEL beams J Synch Rad22 225238
51 Sugahara M Mizohata E Nango E Suzuki MTanaka T Masuda T Tanaka R Shimamura TTanaka Y Suno C Ihara K Pan D KakinouchiK Sugiyama S Murata M Inoue T Tono KSong C Park J Kameshima T Hatsui T Joti YYabashi M and Iwata S (2014) Grease matrix as aversatile carrier of proteins for serial crystallographyNat Methods 12 6163
52 Tono K Nango E Sugahara M Song C Park JTanaka T Tanaka R Joti Y Kameshima T OnoS Hatsui T Mizohata E Suzuki M Shimamura TTanaka Y Iwata S and Yabashi M (2015) Diverseapplication platform for hard X-ray diffraction inSACLA (DAPHNIS) application to serial protein crys-tallography using an X-ray free-electron laser J SynchRad 22 532537
53 Joti Y Kameshima T Yamaga M Sugimoto TOkada K Abe T Furukawa Y Ohata T TanakaR Hatsui T and Yabashi M (2015) Data acquisitionsystem for X-ray free-electron laser experiments atSACLA J Synch Rad 22 571576
54 Yumoto H Mimura H Koyama T Matsuyama STono K Togashi T Inubushi Y Sato T TanakaT Kimura T Yokoyama H Kim J Sano YHachisu Y Yabashi M Ohashi H Ohmori HIshikawa T and Yamauchi K (2012) Focusing of X-ray free-electron laser pulses with reflective optics NatPhotonics 7 4347
55 White TA Kirian RA Martin AV Aquila ANass K Barty A and Chapman HN (2012)CrystFEL a software suite for snapshot serial crystallog-raphy J Appl Cryst 45 335341
56 Brehm W and Diederichs K (2014) Breaking theindexing ambiguity in serial crystallography ActaCryst D 70 101109
57 Karplus PA and Diederichs K (2012) Linking crystal-lographic model and data quality Science 33610301033
58 Evans PR and Murshudov GN (2013) How good aremy data and what is the resolution Acta Cryst D 6912041214
59 Bublitz M Nass K Drachmann ND MarkvardsenAJ Gutmann MJ Barends TRM Mattle DShoeman RL Doak RB Boutet SMesserschmidt M Seibert MM Williams GJFoucar L Reinhard L Sitsel O Gregersen JLClausen JD Boesen T Gotfryd K Wang K-TOlesen C Moslashller JV Nissen P and Schlichting I(2015) Structural studies of P-type ATPaseligand com-plexes using an X-ray free-electron laser IUCrJ 2409420
SFX revealed redox-coupled structural changes in CuNiR
11
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046
Y Fukuda et al
12
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Fig 1 Structural changes in SRX (a) Changes in the hydration structures at the T2Cu site The 2FoFc maps contoured at 10 are shown ascyan meshes Carbon oxygen nitrogen and copper atoms are yellow red blue and brown respectively (b) The ligand plane composed of threeN2 atoms of His residues at the T2Cu site (yellow-dashed lines) Carbon nitrogen and copper atoms are coloured in bright red blue and brownrespectively (c) Distances from T2Cu to the ligand plane The error bars represent twice the values of the coordinate errors estimated by themaximum likelihood method (A colour version of this figure is available online at httpjboxfordjournalsorg)
Table IV Coordination geometries of the copper sites
SFX revealed redox-coupled structural changes in CuNiR
5
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
than that in the SFX structure The electron densitymap of the RT SRX data has a strong positive peakaround WatC with full occupancy (SupplementaryFig S4c) We also observed an unidentified positiveelectron density peak (Supplementary Fig S4c) at thecorresponding position of the putative Na+ site inthe SFX structure (Supplementary Fig S4b) thoughthe signal was too weak to place any atomic modelthere The observation that both RT SRX and SFXdata show this electron density means at least that itis not the result of photoreduction
The electron density map of the RT SRX datashowed a strong electron density peak near HiscatWhen water is assigned there the modelled moleculewas too close to Hiscat (520 A) Therefore it was likely
to be a Cu ion coordinated by Hiscat (SupplementaryFig S7) as was observed in the SFX structure and the3X1E structure
Redox-coupled conformational change in Hiscat
Compared with the SFX structure cryogenic and RTSRX structures revealed that the imidazole ring andthe Cb atom of Hiscat rotated 10 and moved 03 Arespectively (Fig 3a and b Supplementary Fig S8) Toconfirm that it is not the result of cutting the SFX dataat 143 A resolution we refined the SFX structureagainst the data to 165 A resolution where CC12
05 and Rsplit5 100 (Supplementary Fig S2) and theconformation of Hiscat did not change (SupplementaryFig S9) The Nd1 atom of Hiscat forms a bifurcated
Fig 2 SFX structure of GtNiR Carbon oxygen nitrogen sulfur and copper atoms are bright red red blue yellow and brown respectively (a)Copper binding sites in the SFX structure The anomalous Fourier maps are contoured at 40 (magenta) and 12 (dark blue) (b) Hydrationstructure of the T2Cu site in the SFX structure The 2FoFc maps contoured at 10 are shown as cyan meshes Na+ and water molecules areshown as a purple sphere (c) Penta-coordinated Na+ above the T2Cu site Distances are shown in A (A colour version of this figure is availableonline at httpjboxfordjournalsorg)
Y Fukuda et al
6
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
hydrogen bond with the hydroxyl oxygen atom ofThr268 and the backbone carbonyl oxygen atom ofGln267 (Fig 3b) The Hiscat rotation changes thestates of the bifurcated hydrogen bond To qualita-tively evaluate this subtle change we modelled hydro-gen atoms of Hiscat at the ideal positions withoutrefinement Figure 3c shows that with increasing ofX-ray dose the y1 (Nd1HOGln) angle and the y2(Nd1HOThr) angle decreased and increasedrespectively
Discussion
Geometrical changes at Cu sitesWe recently demonstrated that T1Cu in a GtNiR crys-tal is reduced by at least an X-ray dose of 0041 Mgy(30) Because even the X-ray dose for SR1 was higherthan this value (Table I) T1Cu in the present SRX
structures must be more or less reduced Howeverthe geometries around T1Cu did not show significantdifferences between the SRX structures and the com-pletely oxidized SFX structure (Table IV) It is notsurprising because spectroscopic studies have revealedthat the typical changes in ligandT1Cu distance inreduction are501 A so as to minimize the reorganiza-tion energy in ET (67)
We revealed that X-ray irradiation made the T2Cuatom gradually move towards to the ligand plane(Fig 1c) Sunken T2Cu atoms have been observed inthe reduced forms of other CuNiRs (6870)Therefore contrary to the previous report (24) ourresult indicates that T2Cu free of NO2
can be reducedin SRX This conclusion is reasonable because ourcrystal was crystallized at a pH low enough (pH 45)for intramolecular ET without NO2
binding to occurdominantly (28) Although the T2Cu sites in CuNiR
Fig 3 Redox-coupled rotation of Hiscat (a) Electron density maps (contoured at 40 ) around Hiscat in the SFX (red) and SR1 (yellow)
structures (b) Comparison of Hiscat of the SFX (red) SR1 (yellow) SR7 (teal blue) and RT SRX (white) structures (c) Changes in the angle ofthe bifurcated hydrogen bond The upper and lower series show y1 and y2 respectively Trends are shown by dashed lines (A colour version ofthis figure is available online at httpjboxfordjournalsorg)
SFX revealed redox-coupled structural changes in CuNiR
7
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
crystals lose ligand water when it is harshly reduced byartificial reductants (6870) it is just one of thereduced states of T2Cu called inactive reduced stateand a normal reduced state retains water ligand (29) aswas observed in our structures (Fig 1a)
Conformational change of Hiscat
The structural change of Hiscat cannot be caused bycoordination of Cu or the presence of Na+ above theT2Cu site because the former was found also in thecryogenic and RT SR structures (Supplementary FigS6 and S7) and the latter was found also in the RT SRstructure (Supplementary Fig S4c) The difference ofmeasurement temperature is not a major cause of thestructural change (Fig 3b) Thus the conformationaldifference in Hiscat between the SFX and SRX struc-tures is most likely to result from photoreduction ofthe Cu sites Indeed the observed conformationalchange was small and is probably due to the rigid cata-lytic site of thermostable GtNiR (71)
Interestingly there is a hydrogen bond betweenT2Cu ligand His100 and the side chain of Gln267
and His100 is located at the end of a sensor loop(Fig 4a) which is thought to transmit informationabout T1Cursquos redox state to T2Cu (8 72) Althoughthe ThrGln pair (or ThrGlu in some CuNiRs) com-posing the bifurcated hydrogen bond has been ignoredfor long they are conserved in CuNiRs except for afew CuNiRs which have Ser at the position of Thr(Fig 4b) The nitrite reduction activity of Ser-contain-ing CuNiR is lower than the activity of Thr-containingCuNiR (70 73) The crystal structures of Ser-contain-ing CuNiRs show that the Ser residue does not alwaysform a hydrogen bond with Hiscat (70 73) These factsimply the enzymatic importance of the hydrogen bondbetween Hiscat and Thr
The rotation of the imidazole ring changes the yangles of the bifurcated hydrogen bond (Fig 3c)Generally the y angle is used as an indicator of hydro-gen bond strength Strong moderate and weak hydro-gen bonds tend to show 170180 150180 90150
y angles respectively (74) Therefore the Hiscat rota-tion strengthens the hydrogen bond between Hiscat andThr and weakens the hydrogen bond between Hiscat
Fig 4 Conserved hydrogen bond network (a) Redox sensor loop in GtNiR The hydrogen bond network is shown by dotted lines (b) Amino acidsequences of CuNiRs Hiscat GlnGlu and ThrSer are indicated by filled circle triangle and diamond respectively Ax Ac Rs Rp Ph and Hdmeans Achromobacter xylosoxidans Achromobacter cycloclastes Rhodobacter sphaeroides Ralstonia pickettii Pseudoalteromonas haloplanktisand Hyphomicrobium denitrificans respectively (A colour version of this figure is available online at httpjboxfordjournalsorg)
Y Fukuda et al
8
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
and GlnGlu Because the hydroxyl oxygen atom is lessnegatively polarized than the carbonyl oxygen atomthis structural change may destabilize the positivecharge on Hiscat which may facilitate transfer of aproton to the bridging water between Aspcat andHiscat (Fig 4a) Hiscat has been suggested to directlyprovide a proton to NO2
(19 75) however our re-sults indicate another possibility Hiscat may functionas a switch for proton relay
Supplementary Data
Supplementary Data are available at JB Online
Acknowledgements
The SFX experiment was carried out at BL3 of the SPring-8Angstrom Compact Free-Electron Laser (SACLA) with the ap-proval of the Japan Synchrotron Radiation Research Institute(JASRI) (proposal no 2013B8045) We are grateful for supportfrom the SACLA High Performance Computing (HPC) systemand the Mini-K super computer system The microfocus SRX ex-periment at BL32XU of SPring-8 was supported by the Platform forDrug Discovery Informatics and Structural Life Science The au-thors thank MEP Murphy for discussion K Baba and N Mizunofor their help in the experiment at BL38B1 (proposal no2013A1592) E Yamashita and A Higashiura for their support atBL44XU of the SPring-8 and all staff at the SACLA for technicalassistance
FundingThis work was supported by the X-ray Free-Electron Laser PriorityStrategy Program of the Ministry of Education Culture SportsScience and Technology in Japan (MEXT) the Grant-in-Aid forScientific Research on Innovative Areas from MEXT the Grant-in-Aid for Japan Society for the Promotion of Science (JSPS)Research Fellows (Grant no 254626) and the JSPS KAKENHI(Grant no 15K18487)
Conflict of InterestNone declared
References
1 Gruber N and Galloway JN (2008) An Earth-systemperspective of the global nitrogen cycle Nature 451293296
2 Galloway JN Townsend AR Erisman JWBekunda M Cai Z Freney JR Martinelli LASeitzinger SP and Sutton MA (2008)Transformation of the nitrogen cycle recent trendsquestions and potential solutions Science 320 889892
3 Zumft WG (1997) Cell biology and molecular basis ofdenitrification Microbiol Mol Biol Rev 61 533616
4 Tavares P Pereira AS Moura JJ and Moura I(2006) Metalloenzymes of the denitrification pathwayJ Inorg Biochem 100 20872100
5 Fulop V Moir JWB Ferguson SJ and Hajdu J(1995) The anatomy of a bifunctional enzyme structuralbasis for reduction of oxygen to water and synthesis ofnitric oxide by cytochrome cd1 Cell 81 369377
6 Kakutani T Watanabe H Arima K and Beppu T(1981) A blue protein as inactivating factor for nitritereductase from Alcaligenes faecalis Strain S-6J Biochem 89 463472
7 MacPherson IS Rosell FI Scofield M MaukAG and Murphy ME (2010) Directed evolution of
copper nitrite reductase to a chromogenic reductantProtein Eng Des Sel 23 137145
8 Strange RW Murphy LM Dodd FE AbrahamZHL Eady RR Smith BE and Hasnain SS(1999) Structural and kinetic evidence for an orderedmechanism of copper nitrite reductase J Mol Biol287 10011009
9 Godden J Turley S Teller D Adman E Liu MPayne W and LeGall J (1991) The 23 angstrom X-raystructure of nitrite reductase from Achromobacter cyclo-clastes Science 253 438442
10 Kukimoto M Nishiyama M Murphy MEPTurley S Adman ET Horinouchi S and BeppuT (1994) X-ray structure and site-directed mutagenesisof a nitrite reductase from Alcaligenes Faecalis S-6 rolesof two copper atoms in nitrite reduction Biochemistry33 52465252
11 Dodd FE Beeumen JV Eady RR and HasnainSS (1998) X-ray structure of a blue-copper nitrite reduc-tase in two crystal forms The nature of the copper sitesmode of substrate binding and recognition by redox part-ner J Mol Biol 282 369382
12 Inoue T Gotowda M Deligeer Kataoka KYamaguchi K Suzuki S Watanabe H Gohow Mand Kai Y (1998) Type 1 Cu structure of blue nitritereductase from Alcaligenes xylosoxidans GIFU 1051 at205 A resolution comparison of blue and green nitritereductases J Biochem 124 876879
13 Merkle AC and Lehnert N (2012) Binding and acti-vation of nitrite and nitric oxide by copper nitrite reduc-tase and corresponding model complexes Dalton Trans41 33553368
14 Koteishi H Nojiri M Nakagami T Yamaguchi Kand Suzuki S (2009) Cytochrome c551 is a mediator ofelectron transfer between copper-containing nitrite re-ductase and azurin in a denitrifying bacteriumAchromobacter xylosoxidans Bull Chem Soc Jpn 8210031005
15 Nojiri M Koteishi H Nakagami T Kobayashi KInoue T Yamaguchi K and Suzuki S (2009)Structural basis of inter-protein electron transfer for ni-trite reduction in denitrification Nature 462 117120
16 Vlasie MD Fernandez-Busnadiego R Prudencio Mand Ubbink M (2008) Conformation of pseudoazurinin the 152 kDa electron transfer complex with nitrite re-ductase determined by paramagnetic NMR J Mol Biol375 14051415
17 Impagliazzo A Blok AJ Cliff MJ Ladbury JEand Ubbink M (2007) Redox-state-dependent complexformation between pseudoazurin and nitrite reductaseJ Am Chem Soc 129 226233
18 Boulanger MJ Kukimoto M Nishiyama MHorinouchi S and Murphy ME (2000) Catalyticroles for two water bridged residues (Asp-98 and His-255) in the active site of copper-containing nitrite reduc-tase J Biol Chem 275 2395723964
19 Kataoka K Furusawa H Takagi K Yamaguchi Kand Suzuki S (2000) Functional analysis of conservedaspartate and histidine residues located around the type 2copper site of copper-containing nitrite reductaseJ Biochem 127 345350
20 Yano J Kern J Irrgang KD Latimer MJBergmann U Glatzel P Pushkar Y Biesiadka JLoll B Sauer K Messinger J Zouni A andYachandra VK (2005) X-ray damage to the Mn4Cacomplex in single crystals of photosystem II a case
SFX revealed redox-coupled structural changes in CuNiR
9
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
study for metalloprotein crystallography Proc NatlAcad Sci USA 102 1204712052
21 Schlichting I Berendzen J Chu K Stock AMMaves SA Benson DE Sweet RM Ringe DPetsko GA and Sligar SG (2000) The catalytic path-way of cytochrome P450cam at atomic resolutionScience 287 16151622
22 Berglund GI Carlsson GH Smith AT Szoke HHenriksen A and Hajdu J (2002) The catalytic path-way of horseradish peroxidase at high resolution Nature417 463468
23 Beitlich T Kuhnel K Schulze-Briese C ShoemanRL and Schlichting I (2007) Cryoradiolytic reductionof crystalline heme proteins analysis by UV-Vis spec-troscopy and X-ray crystallography J Synch Rad 141123
24 Hough MA Antonyuk SV Strange RW EadyRR and Hasnain SS (2008) Crystallography withonline optical and X-ray absorption spectroscopies dem-onstrates an ordered mechanism in copper nitrite reduc-tase J Mol Biol 378 353361
25 Brenner S Heyes DJ Hay S Hough MA EadyRR Hasnain SS and Scrutton NS (2009)Demonstration of proton-coupled electron transfer inthe copper-containing nitrite reductases J Biol Chem284 2597325983
26 Leferink NG Han C Antonyuk SV Heyes DJRigby SE Hough MA Eady RR Scrutton NSand Hasnain SS (2011) Proton-coupled electron trans-fer in the catalytic cycle of Alcaligenes xylosoxidanscopper-dependent nitrite reductase Biochemistry 5041214131
27 Kobayashi K Tagawa S Deligeer and Suzuki S(1999) The pH-dependent changes of intramolecularelectron transfer on copper-containing nitrite reductaseJ Biochem 126 408412
28 Wijma HJ Jeuken LJ Verbeet MP ArmstrongFA and Canters GW (2006) A random-sequentialmechanism for nitrite binding and active site reductionin copper-containing nitrite reductase J Biol Chem281 1634016346
29 Wijma HJ Jeuken LJC Verbeet MP ArmstrongFA and Canters GW (2007) Protein film voltamme-try of copper-containing nitrite reductase reveals revers-ible inactivation J Am Chem Soc 129 85578565
30 Fukuda Y Tse KM Kado Y Mizohata EMatsumura H and Inoue T (2015) Insights into un-known foreign ligand in copper nitrite reductaseBiochem Biophys Res Commun 464 622628
31 Flot D Mairs T Giraud T Guijarro M LesourdM Rey V van Brussel D Morawe C Borel CHignette O Chavanne J Nurizzo D McSweeneyS and Mitchell E (2010) The ID23-2 structural biologymicrofocus beamline at the ESRF J Synch Rad 17107118
32 Chapman HN Fromme P Barty A White TAKirian RA Aquila A Hunter MS Schulz JDePonte DP Weierstall U Doak RB Maia FRMartin AV Schlichting I Lomb L Coppola NShoeman RL Epp SW Hartmann R Rolles DRudenko A Foucar L Kimmel N WeidenspointnerG Holl P Liang M Barthelmess M Caleman CBoutet S Bogan MJ Krzywinski J Bostedt CBajt S Gumprecht L Rudek B Erk B SchmidtC Homke A Reich C Pietschner D Struder LHauser G Gorke H Ullrich J Herrmann SSchaller G Schopper F Soltau H Kuhnel KU
Messerschmidt M Bozek JD Hau-Riege SPFrank M Hampton CY Sierra RG StarodubD Williams GJ Hajdu J Timneanu N SeibertMM Andreasson J Rocker A Jonsson OSvenda M Stern S Nass K Andritschke RSchroter CD Krasniqi F Bott M Schmidt KEWang X Grotjohann I Holton JM Barends TRNeutze R Marchesini S Fromme R Schorb SRupp D Adolph M Gorkhover T Andersson IHirsemann H Potdevin G Graafsma H NilssonB and Spence JC (2011) Femtosecond X-ray proteinnanocrystallography Nature 470 7377
33 Kern J Alonso-Mori R Hellmich J Tran RHattne J Laksmono H Glockner C Echols NSierra RG Sellberg J Lassalle-Kaiser B GildeaRJ Glatzel P Grosse-Kunstleve RW LatimerMJ McQueen TA DiFiore D Fry ARMesserschmidt M Miahnahri A Schafer DWSeibert MM Sokaras D Weng TC Zwart PHWhite WE Adams PD Bogan MJ Boutet SWilliams GJ Messinger J Sauter NK Zouni ABergmann U Yano J and Yachandra VK (2012)Room temperature femtosecond X-ray diffraction ofphotosystem II microcrystals Proc Natl Acad SciUSA 109 97219726
34 Johansson LC Arnlund D Katona G White TABarty A DePonte DP Shoeman RL WickstrandC Sharma A Williams GJ Aquila A Bogan MJCaleman C Davidsson J Doak RB Frank MFromme R Galli L Grotjohann I Hunter MSKassemeyer S Kirian RA Kupitz C Liang MLomb L Malmerberg E Martin AVMesserschmidt M Nass K Redecke L SeibertMM Sjohamn J Steinbrener J Stellato F WangD Wahlgren WY Weierstall U Westenhoff SZatsepin NA Boutet S Spence JC Schlichting IChapman HN Fromme P and Neutze R (2013)Structure of a photosynthetic reaction centre determinedby serial femtosecond crystallography Nat Commun 42911
35 Kupitz C Basu S Grotjohann I Fromme RZatsepin NA Rendek KN Hunter MSShoeman RL White TA Wang D James DYang JH Cobb DE Reeder B Sierra RG LiuH Barty A Aquila AL Deponte D Kirian RABari S Bergkamp JJ Beyerlein KR Bogan MJCaleman C Chao TC Conrad CE Davis KMFleckenstein H Galli L Hau-Riege SPKassemeyer S Laksmono H Liang M Lomb LMarchesini S Martin AV Messerschmidt MMilathianaki D Nass K Ros A Roy-ChowdhuryS Schmidt K Seibert M Steinbrener J Stellato FYan L Yoon C Moore TA Moore AL PushkarY Williams GJ Boutet S Doak RB WeierstallU Frank M Chapman HN Spence JC andFromme P (2014) Serial time-resolved crystallographyof photosystem II using a femtosecond X-ray laserNature 513 261265
36 Kern J Alonso-Mori R Tran R Hattne J GildeaRJ Echols N Glockner C Hellmich J LaksmonoH Sierra RG Lassalle-Kaiser B Koroidov SLampe A Han G Gul S DiFiore DMilathianaki D Fry AR Miahnahri A SchaferDW Messerschmidt M Seibert MM Koglin JESokaras D Weng T-C Sellberg J Latimer MJGrosse-Kunstleve RW Zwart PH White WEGlatzel P Adams PD Bogan MJ Williams GJBoutet S Messinger J Zouni A Sauter NK
Y Fukuda et al
10
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Yachandra VK Bergmann U and Yano J (2013)Simultaneous femtosecond X-ray spectroscopy and dif-fraction of photosystem II at room temperature Science340 491495
37 Kern J Tran R Alonso-Mori R Koroidov SEchols N Hattne J Ibrahim M Gul SLaksmono H Sierra RG Gildea RJ Han GHellmich J Lassalle-Kaiser B Chatterjee RBrewster AS Stan CA Glockner C Lampe ADiFiore D Milathianaki D Fry AR SeibertMM Koglin JE Gallo E Uhlig J Sokaras DWeng TC Zwart PH Skinner DE Bogan MJMesserschmidt M Glatzel P Williams GJ BoutetS Adams PD Zouni A Messinger J Sauter NKBergmann U Yano J and Yachandra VK (2014)Taking snapshots of photosynthetic water oxidationusing femtosecond X-ray diffraction and spectroscopyNat Commun 5 4371
38 Hirata K Kawano Y Ueno G Hashimoto KMurakami H Hasegawa K Hikima T KumasakaT and Yamamoto M (2013) Achievement of proteinmicro-crystallography at SPring-8 beamline BL32XUJ Phys Conf Ser 425 012002
39 Paithankar KS and Garman EF (2010) Know yourdose RADDOSE Acta Cryst D 66 381388
40 Tanida H Kikuchi A Miura K Takeshita KGoto S Shiro Y and Ishikawa T (2004) XAFS andprotein crystallography beamline BL38B1 at SPring-8AIP Conf Proc 705 486489
41 Fukuda Y and Inoue T (2015) High-temperatureand high-resolution crystallography of thermostablecopper nitrite reductase Chem Commun 5165326535
42 Otwinowski Z and Minor W (1997) Processing of X-ray diffraction data collected in oscillation modeMethods Enzymol 276 307326
43 Vagin A and Teplyakov A (2010) Molecular replace-ment with MOLREP Acta Cryst D 66 2225
44 Emsley P Lohkamp B Scott WG and Cowtan K(2010) Features and development of Coot Acta CrystD 66 486501
45 Murshudov GN Skubak P Lebedev AA PannuNS Steiner RA Nicholls RA Winn MD LongF and Vagin AA (2011) REFMAC5 for the refine-ment of macromolecular crystal structures Acta CrystD 67 355367
46 Winn MD Ballard CC Cowtan KD DodsonEJ Emsley P Evans PR Keegan RM KrissinelEB Leslie AG McCoy A McNicholas SJMurshudov GN Pannu NS Potterton EAPowell HR Read RJ Vagin A and Wilson KS(2011) Overview of the CCP4 suite and current develop-ments Acta Cryst D 67 235242
47 Chen VB Arendall WB III Headd JJ KeedyDA Immormino RM Kapral GJ Murray LWRichardson JS and Richardson DC (2010)MolProbity all-atom structure validation for macromol-ecular crystallography Acta Crystallogr D BiolCrystallogr 66 1221
48 Tono K Togashi T Inubushi Y Sato TKatayama T Ogawa K Ohashi H Kimura HTakahashi S Takeshita K Tomizawa H Goto SIshikawa T and Yabashi M (2013) Beamline experi-mental stations and photon beam diagnostics for thehard x-ray free electron laser of SACLA New J Phys15 083035
49 Son S-K Chapman HN and Santra R (2011)Multiwavelength anomalous diffraction at high X-rayintensity Phys Rev Lett 107 218102
50 Nass K Foucar L Barends TR Hartmann EBotha S Shoeman RL Doak RB Alonso-MoriR Aquila A Bajt S Barty A Bean R BeyerleinKR Bublitz M Drachmann N Gregersen JJonsson HO Kabsch W Kassemeyer S KoglinJE Krumrey M Mattle D Messerschmidt MNissen P Reinhard L Sitsel O Sokaras DWilliams GJ Hau-Riege S Timneanu N CalemanC Chapman HN Boutet S and Schlichting I (2015)Indications of radiation damage in ferredoxin microcrys-tals using high-intensity X-FEL beams J Synch Rad22 225238
51 Sugahara M Mizohata E Nango E Suzuki MTanaka T Masuda T Tanaka R Shimamura TTanaka Y Suno C Ihara K Pan D KakinouchiK Sugiyama S Murata M Inoue T Tono KSong C Park J Kameshima T Hatsui T Joti YYabashi M and Iwata S (2014) Grease matrix as aversatile carrier of proteins for serial crystallographyNat Methods 12 6163
52 Tono K Nango E Sugahara M Song C Park JTanaka T Tanaka R Joti Y Kameshima T OnoS Hatsui T Mizohata E Suzuki M Shimamura TTanaka Y Iwata S and Yabashi M (2015) Diverseapplication platform for hard X-ray diffraction inSACLA (DAPHNIS) application to serial protein crys-tallography using an X-ray free-electron laser J SynchRad 22 532537
53 Joti Y Kameshima T Yamaga M Sugimoto TOkada K Abe T Furukawa Y Ohata T TanakaR Hatsui T and Yabashi M (2015) Data acquisitionsystem for X-ray free-electron laser experiments atSACLA J Synch Rad 22 571576
54 Yumoto H Mimura H Koyama T Matsuyama STono K Togashi T Inubushi Y Sato T TanakaT Kimura T Yokoyama H Kim J Sano YHachisu Y Yabashi M Ohashi H Ohmori HIshikawa T and Yamauchi K (2012) Focusing of X-ray free-electron laser pulses with reflective optics NatPhotonics 7 4347
55 White TA Kirian RA Martin AV Aquila ANass K Barty A and Chapman HN (2012)CrystFEL a software suite for snapshot serial crystallog-raphy J Appl Cryst 45 335341
56 Brehm W and Diederichs K (2014) Breaking theindexing ambiguity in serial crystallography ActaCryst D 70 101109
57 Karplus PA and Diederichs K (2012) Linking crystal-lographic model and data quality Science 33610301033
58 Evans PR and Murshudov GN (2013) How good aremy data and what is the resolution Acta Cryst D 6912041214
59 Bublitz M Nass K Drachmann ND MarkvardsenAJ Gutmann MJ Barends TRM Mattle DShoeman RL Doak RB Boutet SMesserschmidt M Seibert MM Williams GJFoucar L Reinhard L Sitsel O Gregersen JLClausen JD Boesen T Gotfryd K Wang K-TOlesen C Moslashller JV Nissen P and Schlichting I(2015) Structural studies of P-type ATPaseligand com-plexes using an X-ray free-electron laser IUCrJ 2409420
SFX revealed redox-coupled structural changes in CuNiR
11
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046
Y Fukuda et al
12
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
than that in the SFX structure The electron densitymap of the RT SRX data has a strong positive peakaround WatC with full occupancy (SupplementaryFig S4c) We also observed an unidentified positiveelectron density peak (Supplementary Fig S4c) at thecorresponding position of the putative Na+ site inthe SFX structure (Supplementary Fig S4b) thoughthe signal was too weak to place any atomic modelthere The observation that both RT SRX and SFXdata show this electron density means at least that itis not the result of photoreduction
The electron density map of the RT SRX datashowed a strong electron density peak near HiscatWhen water is assigned there the modelled moleculewas too close to Hiscat (520 A) Therefore it was likely
to be a Cu ion coordinated by Hiscat (SupplementaryFig S7) as was observed in the SFX structure and the3X1E structure
Redox-coupled conformational change in Hiscat
Compared with the SFX structure cryogenic and RTSRX structures revealed that the imidazole ring andthe Cb atom of Hiscat rotated 10 and moved 03 Arespectively (Fig 3a and b Supplementary Fig S8) Toconfirm that it is not the result of cutting the SFX dataat 143 A resolution we refined the SFX structureagainst the data to 165 A resolution where CC12
05 and Rsplit5 100 (Supplementary Fig S2) and theconformation of Hiscat did not change (SupplementaryFig S9) The Nd1 atom of Hiscat forms a bifurcated
Fig 2 SFX structure of GtNiR Carbon oxygen nitrogen sulfur and copper atoms are bright red red blue yellow and brown respectively (a)Copper binding sites in the SFX structure The anomalous Fourier maps are contoured at 40 (magenta) and 12 (dark blue) (b) Hydrationstructure of the T2Cu site in the SFX structure The 2FoFc maps contoured at 10 are shown as cyan meshes Na+ and water molecules areshown as a purple sphere (c) Penta-coordinated Na+ above the T2Cu site Distances are shown in A (A colour version of this figure is availableonline at httpjboxfordjournalsorg)
Y Fukuda et al
6
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
hydrogen bond with the hydroxyl oxygen atom ofThr268 and the backbone carbonyl oxygen atom ofGln267 (Fig 3b) The Hiscat rotation changes thestates of the bifurcated hydrogen bond To qualita-tively evaluate this subtle change we modelled hydro-gen atoms of Hiscat at the ideal positions withoutrefinement Figure 3c shows that with increasing ofX-ray dose the y1 (Nd1HOGln) angle and the y2(Nd1HOThr) angle decreased and increasedrespectively
Discussion
Geometrical changes at Cu sitesWe recently demonstrated that T1Cu in a GtNiR crys-tal is reduced by at least an X-ray dose of 0041 Mgy(30) Because even the X-ray dose for SR1 was higherthan this value (Table I) T1Cu in the present SRX
structures must be more or less reduced Howeverthe geometries around T1Cu did not show significantdifferences between the SRX structures and the com-pletely oxidized SFX structure (Table IV) It is notsurprising because spectroscopic studies have revealedthat the typical changes in ligandT1Cu distance inreduction are501 A so as to minimize the reorganiza-tion energy in ET (67)
We revealed that X-ray irradiation made the T2Cuatom gradually move towards to the ligand plane(Fig 1c) Sunken T2Cu atoms have been observed inthe reduced forms of other CuNiRs (6870)Therefore contrary to the previous report (24) ourresult indicates that T2Cu free of NO2
can be reducedin SRX This conclusion is reasonable because ourcrystal was crystallized at a pH low enough (pH 45)for intramolecular ET without NO2
binding to occurdominantly (28) Although the T2Cu sites in CuNiR
Fig 3 Redox-coupled rotation of Hiscat (a) Electron density maps (contoured at 40 ) around Hiscat in the SFX (red) and SR1 (yellow)
structures (b) Comparison of Hiscat of the SFX (red) SR1 (yellow) SR7 (teal blue) and RT SRX (white) structures (c) Changes in the angle ofthe bifurcated hydrogen bond The upper and lower series show y1 and y2 respectively Trends are shown by dashed lines (A colour version ofthis figure is available online at httpjboxfordjournalsorg)
SFX revealed redox-coupled structural changes in CuNiR
7
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
crystals lose ligand water when it is harshly reduced byartificial reductants (6870) it is just one of thereduced states of T2Cu called inactive reduced stateand a normal reduced state retains water ligand (29) aswas observed in our structures (Fig 1a)
Conformational change of Hiscat
The structural change of Hiscat cannot be caused bycoordination of Cu or the presence of Na+ above theT2Cu site because the former was found also in thecryogenic and RT SR structures (Supplementary FigS6 and S7) and the latter was found also in the RT SRstructure (Supplementary Fig S4c) The difference ofmeasurement temperature is not a major cause of thestructural change (Fig 3b) Thus the conformationaldifference in Hiscat between the SFX and SRX struc-tures is most likely to result from photoreduction ofthe Cu sites Indeed the observed conformationalchange was small and is probably due to the rigid cata-lytic site of thermostable GtNiR (71)
Interestingly there is a hydrogen bond betweenT2Cu ligand His100 and the side chain of Gln267
and His100 is located at the end of a sensor loop(Fig 4a) which is thought to transmit informationabout T1Cursquos redox state to T2Cu (8 72) Althoughthe ThrGln pair (or ThrGlu in some CuNiRs) com-posing the bifurcated hydrogen bond has been ignoredfor long they are conserved in CuNiRs except for afew CuNiRs which have Ser at the position of Thr(Fig 4b) The nitrite reduction activity of Ser-contain-ing CuNiR is lower than the activity of Thr-containingCuNiR (70 73) The crystal structures of Ser-contain-ing CuNiRs show that the Ser residue does not alwaysform a hydrogen bond with Hiscat (70 73) These factsimply the enzymatic importance of the hydrogen bondbetween Hiscat and Thr
The rotation of the imidazole ring changes the yangles of the bifurcated hydrogen bond (Fig 3c)Generally the y angle is used as an indicator of hydro-gen bond strength Strong moderate and weak hydro-gen bonds tend to show 170180 150180 90150
y angles respectively (74) Therefore the Hiscat rota-tion strengthens the hydrogen bond between Hiscat andThr and weakens the hydrogen bond between Hiscat
Fig 4 Conserved hydrogen bond network (a) Redox sensor loop in GtNiR The hydrogen bond network is shown by dotted lines (b) Amino acidsequences of CuNiRs Hiscat GlnGlu and ThrSer are indicated by filled circle triangle and diamond respectively Ax Ac Rs Rp Ph and Hdmeans Achromobacter xylosoxidans Achromobacter cycloclastes Rhodobacter sphaeroides Ralstonia pickettii Pseudoalteromonas haloplanktisand Hyphomicrobium denitrificans respectively (A colour version of this figure is available online at httpjboxfordjournalsorg)
Y Fukuda et al
8
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
and GlnGlu Because the hydroxyl oxygen atom is lessnegatively polarized than the carbonyl oxygen atomthis structural change may destabilize the positivecharge on Hiscat which may facilitate transfer of aproton to the bridging water between Aspcat andHiscat (Fig 4a) Hiscat has been suggested to directlyprovide a proton to NO2
(19 75) however our re-sults indicate another possibility Hiscat may functionas a switch for proton relay
Supplementary Data
Supplementary Data are available at JB Online
Acknowledgements
The SFX experiment was carried out at BL3 of the SPring-8Angstrom Compact Free-Electron Laser (SACLA) with the ap-proval of the Japan Synchrotron Radiation Research Institute(JASRI) (proposal no 2013B8045) We are grateful for supportfrom the SACLA High Performance Computing (HPC) systemand the Mini-K super computer system The microfocus SRX ex-periment at BL32XU of SPring-8 was supported by the Platform forDrug Discovery Informatics and Structural Life Science The au-thors thank MEP Murphy for discussion K Baba and N Mizunofor their help in the experiment at BL38B1 (proposal no2013A1592) E Yamashita and A Higashiura for their support atBL44XU of the SPring-8 and all staff at the SACLA for technicalassistance
FundingThis work was supported by the X-ray Free-Electron Laser PriorityStrategy Program of the Ministry of Education Culture SportsScience and Technology in Japan (MEXT) the Grant-in-Aid forScientific Research on Innovative Areas from MEXT the Grant-in-Aid for Japan Society for the Promotion of Science (JSPS)Research Fellows (Grant no 254626) and the JSPS KAKENHI(Grant no 15K18487)
Conflict of InterestNone declared
References
1 Gruber N and Galloway JN (2008) An Earth-systemperspective of the global nitrogen cycle Nature 451293296
2 Galloway JN Townsend AR Erisman JWBekunda M Cai Z Freney JR Martinelli LASeitzinger SP and Sutton MA (2008)Transformation of the nitrogen cycle recent trendsquestions and potential solutions Science 320 889892
3 Zumft WG (1997) Cell biology and molecular basis ofdenitrification Microbiol Mol Biol Rev 61 533616
4 Tavares P Pereira AS Moura JJ and Moura I(2006) Metalloenzymes of the denitrification pathwayJ Inorg Biochem 100 20872100
5 Fulop V Moir JWB Ferguson SJ and Hajdu J(1995) The anatomy of a bifunctional enzyme structuralbasis for reduction of oxygen to water and synthesis ofnitric oxide by cytochrome cd1 Cell 81 369377
6 Kakutani T Watanabe H Arima K and Beppu T(1981) A blue protein as inactivating factor for nitritereductase from Alcaligenes faecalis Strain S-6J Biochem 89 463472
7 MacPherson IS Rosell FI Scofield M MaukAG and Murphy ME (2010) Directed evolution of
copper nitrite reductase to a chromogenic reductantProtein Eng Des Sel 23 137145
8 Strange RW Murphy LM Dodd FE AbrahamZHL Eady RR Smith BE and Hasnain SS(1999) Structural and kinetic evidence for an orderedmechanism of copper nitrite reductase J Mol Biol287 10011009
9 Godden J Turley S Teller D Adman E Liu MPayne W and LeGall J (1991) The 23 angstrom X-raystructure of nitrite reductase from Achromobacter cyclo-clastes Science 253 438442
10 Kukimoto M Nishiyama M Murphy MEPTurley S Adman ET Horinouchi S and BeppuT (1994) X-ray structure and site-directed mutagenesisof a nitrite reductase from Alcaligenes Faecalis S-6 rolesof two copper atoms in nitrite reduction Biochemistry33 52465252
11 Dodd FE Beeumen JV Eady RR and HasnainSS (1998) X-ray structure of a blue-copper nitrite reduc-tase in two crystal forms The nature of the copper sitesmode of substrate binding and recognition by redox part-ner J Mol Biol 282 369382
12 Inoue T Gotowda M Deligeer Kataoka KYamaguchi K Suzuki S Watanabe H Gohow Mand Kai Y (1998) Type 1 Cu structure of blue nitritereductase from Alcaligenes xylosoxidans GIFU 1051 at205 A resolution comparison of blue and green nitritereductases J Biochem 124 876879
13 Merkle AC and Lehnert N (2012) Binding and acti-vation of nitrite and nitric oxide by copper nitrite reduc-tase and corresponding model complexes Dalton Trans41 33553368
14 Koteishi H Nojiri M Nakagami T Yamaguchi Kand Suzuki S (2009) Cytochrome c551 is a mediator ofelectron transfer between copper-containing nitrite re-ductase and azurin in a denitrifying bacteriumAchromobacter xylosoxidans Bull Chem Soc Jpn 8210031005
15 Nojiri M Koteishi H Nakagami T Kobayashi KInoue T Yamaguchi K and Suzuki S (2009)Structural basis of inter-protein electron transfer for ni-trite reduction in denitrification Nature 462 117120
16 Vlasie MD Fernandez-Busnadiego R Prudencio Mand Ubbink M (2008) Conformation of pseudoazurinin the 152 kDa electron transfer complex with nitrite re-ductase determined by paramagnetic NMR J Mol Biol375 14051415
17 Impagliazzo A Blok AJ Cliff MJ Ladbury JEand Ubbink M (2007) Redox-state-dependent complexformation between pseudoazurin and nitrite reductaseJ Am Chem Soc 129 226233
18 Boulanger MJ Kukimoto M Nishiyama MHorinouchi S and Murphy ME (2000) Catalyticroles for two water bridged residues (Asp-98 and His-255) in the active site of copper-containing nitrite reduc-tase J Biol Chem 275 2395723964
19 Kataoka K Furusawa H Takagi K Yamaguchi Kand Suzuki S (2000) Functional analysis of conservedaspartate and histidine residues located around the type 2copper site of copper-containing nitrite reductaseJ Biochem 127 345350
20 Yano J Kern J Irrgang KD Latimer MJBergmann U Glatzel P Pushkar Y Biesiadka JLoll B Sauer K Messinger J Zouni A andYachandra VK (2005) X-ray damage to the Mn4Cacomplex in single crystals of photosystem II a case
SFX revealed redox-coupled structural changes in CuNiR
9
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
study for metalloprotein crystallography Proc NatlAcad Sci USA 102 1204712052
21 Schlichting I Berendzen J Chu K Stock AMMaves SA Benson DE Sweet RM Ringe DPetsko GA and Sligar SG (2000) The catalytic path-way of cytochrome P450cam at atomic resolutionScience 287 16151622
22 Berglund GI Carlsson GH Smith AT Szoke HHenriksen A and Hajdu J (2002) The catalytic path-way of horseradish peroxidase at high resolution Nature417 463468
23 Beitlich T Kuhnel K Schulze-Briese C ShoemanRL and Schlichting I (2007) Cryoradiolytic reductionof crystalline heme proteins analysis by UV-Vis spec-troscopy and X-ray crystallography J Synch Rad 141123
24 Hough MA Antonyuk SV Strange RW EadyRR and Hasnain SS (2008) Crystallography withonline optical and X-ray absorption spectroscopies dem-onstrates an ordered mechanism in copper nitrite reduc-tase J Mol Biol 378 353361
25 Brenner S Heyes DJ Hay S Hough MA EadyRR Hasnain SS and Scrutton NS (2009)Demonstration of proton-coupled electron transfer inthe copper-containing nitrite reductases J Biol Chem284 2597325983
26 Leferink NG Han C Antonyuk SV Heyes DJRigby SE Hough MA Eady RR Scrutton NSand Hasnain SS (2011) Proton-coupled electron trans-fer in the catalytic cycle of Alcaligenes xylosoxidanscopper-dependent nitrite reductase Biochemistry 5041214131
27 Kobayashi K Tagawa S Deligeer and Suzuki S(1999) The pH-dependent changes of intramolecularelectron transfer on copper-containing nitrite reductaseJ Biochem 126 408412
28 Wijma HJ Jeuken LJ Verbeet MP ArmstrongFA and Canters GW (2006) A random-sequentialmechanism for nitrite binding and active site reductionin copper-containing nitrite reductase J Biol Chem281 1634016346
29 Wijma HJ Jeuken LJC Verbeet MP ArmstrongFA and Canters GW (2007) Protein film voltamme-try of copper-containing nitrite reductase reveals revers-ible inactivation J Am Chem Soc 129 85578565
30 Fukuda Y Tse KM Kado Y Mizohata EMatsumura H and Inoue T (2015) Insights into un-known foreign ligand in copper nitrite reductaseBiochem Biophys Res Commun 464 622628
31 Flot D Mairs T Giraud T Guijarro M LesourdM Rey V van Brussel D Morawe C Borel CHignette O Chavanne J Nurizzo D McSweeneyS and Mitchell E (2010) The ID23-2 structural biologymicrofocus beamline at the ESRF J Synch Rad 17107118
32 Chapman HN Fromme P Barty A White TAKirian RA Aquila A Hunter MS Schulz JDePonte DP Weierstall U Doak RB Maia FRMartin AV Schlichting I Lomb L Coppola NShoeman RL Epp SW Hartmann R Rolles DRudenko A Foucar L Kimmel N WeidenspointnerG Holl P Liang M Barthelmess M Caleman CBoutet S Bogan MJ Krzywinski J Bostedt CBajt S Gumprecht L Rudek B Erk B SchmidtC Homke A Reich C Pietschner D Struder LHauser G Gorke H Ullrich J Herrmann SSchaller G Schopper F Soltau H Kuhnel KU
Messerschmidt M Bozek JD Hau-Riege SPFrank M Hampton CY Sierra RG StarodubD Williams GJ Hajdu J Timneanu N SeibertMM Andreasson J Rocker A Jonsson OSvenda M Stern S Nass K Andritschke RSchroter CD Krasniqi F Bott M Schmidt KEWang X Grotjohann I Holton JM Barends TRNeutze R Marchesini S Fromme R Schorb SRupp D Adolph M Gorkhover T Andersson IHirsemann H Potdevin G Graafsma H NilssonB and Spence JC (2011) Femtosecond X-ray proteinnanocrystallography Nature 470 7377
33 Kern J Alonso-Mori R Hellmich J Tran RHattne J Laksmono H Glockner C Echols NSierra RG Sellberg J Lassalle-Kaiser B GildeaRJ Glatzel P Grosse-Kunstleve RW LatimerMJ McQueen TA DiFiore D Fry ARMesserschmidt M Miahnahri A Schafer DWSeibert MM Sokaras D Weng TC Zwart PHWhite WE Adams PD Bogan MJ Boutet SWilliams GJ Messinger J Sauter NK Zouni ABergmann U Yano J and Yachandra VK (2012)Room temperature femtosecond X-ray diffraction ofphotosystem II microcrystals Proc Natl Acad SciUSA 109 97219726
34 Johansson LC Arnlund D Katona G White TABarty A DePonte DP Shoeman RL WickstrandC Sharma A Williams GJ Aquila A Bogan MJCaleman C Davidsson J Doak RB Frank MFromme R Galli L Grotjohann I Hunter MSKassemeyer S Kirian RA Kupitz C Liang MLomb L Malmerberg E Martin AVMesserschmidt M Nass K Redecke L SeibertMM Sjohamn J Steinbrener J Stellato F WangD Wahlgren WY Weierstall U Westenhoff SZatsepin NA Boutet S Spence JC Schlichting IChapman HN Fromme P and Neutze R (2013)Structure of a photosynthetic reaction centre determinedby serial femtosecond crystallography Nat Commun 42911
35 Kupitz C Basu S Grotjohann I Fromme RZatsepin NA Rendek KN Hunter MSShoeman RL White TA Wang D James DYang JH Cobb DE Reeder B Sierra RG LiuH Barty A Aquila AL Deponte D Kirian RABari S Bergkamp JJ Beyerlein KR Bogan MJCaleman C Chao TC Conrad CE Davis KMFleckenstein H Galli L Hau-Riege SPKassemeyer S Laksmono H Liang M Lomb LMarchesini S Martin AV Messerschmidt MMilathianaki D Nass K Ros A Roy-ChowdhuryS Schmidt K Seibert M Steinbrener J Stellato FYan L Yoon C Moore TA Moore AL PushkarY Williams GJ Boutet S Doak RB WeierstallU Frank M Chapman HN Spence JC andFromme P (2014) Serial time-resolved crystallographyof photosystem II using a femtosecond X-ray laserNature 513 261265
36 Kern J Alonso-Mori R Tran R Hattne J GildeaRJ Echols N Glockner C Hellmich J LaksmonoH Sierra RG Lassalle-Kaiser B Koroidov SLampe A Han G Gul S DiFiore DMilathianaki D Fry AR Miahnahri A SchaferDW Messerschmidt M Seibert MM Koglin JESokaras D Weng T-C Sellberg J Latimer MJGrosse-Kunstleve RW Zwart PH White WEGlatzel P Adams PD Bogan MJ Williams GJBoutet S Messinger J Zouni A Sauter NK
Y Fukuda et al
10
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Yachandra VK Bergmann U and Yano J (2013)Simultaneous femtosecond X-ray spectroscopy and dif-fraction of photosystem II at room temperature Science340 491495
37 Kern J Tran R Alonso-Mori R Koroidov SEchols N Hattne J Ibrahim M Gul SLaksmono H Sierra RG Gildea RJ Han GHellmich J Lassalle-Kaiser B Chatterjee RBrewster AS Stan CA Glockner C Lampe ADiFiore D Milathianaki D Fry AR SeibertMM Koglin JE Gallo E Uhlig J Sokaras DWeng TC Zwart PH Skinner DE Bogan MJMesserschmidt M Glatzel P Williams GJ BoutetS Adams PD Zouni A Messinger J Sauter NKBergmann U Yano J and Yachandra VK (2014)Taking snapshots of photosynthetic water oxidationusing femtosecond X-ray diffraction and spectroscopyNat Commun 5 4371
38 Hirata K Kawano Y Ueno G Hashimoto KMurakami H Hasegawa K Hikima T KumasakaT and Yamamoto M (2013) Achievement of proteinmicro-crystallography at SPring-8 beamline BL32XUJ Phys Conf Ser 425 012002
39 Paithankar KS and Garman EF (2010) Know yourdose RADDOSE Acta Cryst D 66 381388
40 Tanida H Kikuchi A Miura K Takeshita KGoto S Shiro Y and Ishikawa T (2004) XAFS andprotein crystallography beamline BL38B1 at SPring-8AIP Conf Proc 705 486489
41 Fukuda Y and Inoue T (2015) High-temperatureand high-resolution crystallography of thermostablecopper nitrite reductase Chem Commun 5165326535
42 Otwinowski Z and Minor W (1997) Processing of X-ray diffraction data collected in oscillation modeMethods Enzymol 276 307326
43 Vagin A and Teplyakov A (2010) Molecular replace-ment with MOLREP Acta Cryst D 66 2225
44 Emsley P Lohkamp B Scott WG and Cowtan K(2010) Features and development of Coot Acta CrystD 66 486501
45 Murshudov GN Skubak P Lebedev AA PannuNS Steiner RA Nicholls RA Winn MD LongF and Vagin AA (2011) REFMAC5 for the refine-ment of macromolecular crystal structures Acta CrystD 67 355367
46 Winn MD Ballard CC Cowtan KD DodsonEJ Emsley P Evans PR Keegan RM KrissinelEB Leslie AG McCoy A McNicholas SJMurshudov GN Pannu NS Potterton EAPowell HR Read RJ Vagin A and Wilson KS(2011) Overview of the CCP4 suite and current develop-ments Acta Cryst D 67 235242
47 Chen VB Arendall WB III Headd JJ KeedyDA Immormino RM Kapral GJ Murray LWRichardson JS and Richardson DC (2010)MolProbity all-atom structure validation for macromol-ecular crystallography Acta Crystallogr D BiolCrystallogr 66 1221
48 Tono K Togashi T Inubushi Y Sato TKatayama T Ogawa K Ohashi H Kimura HTakahashi S Takeshita K Tomizawa H Goto SIshikawa T and Yabashi M (2013) Beamline experi-mental stations and photon beam diagnostics for thehard x-ray free electron laser of SACLA New J Phys15 083035
49 Son S-K Chapman HN and Santra R (2011)Multiwavelength anomalous diffraction at high X-rayintensity Phys Rev Lett 107 218102
50 Nass K Foucar L Barends TR Hartmann EBotha S Shoeman RL Doak RB Alonso-MoriR Aquila A Bajt S Barty A Bean R BeyerleinKR Bublitz M Drachmann N Gregersen JJonsson HO Kabsch W Kassemeyer S KoglinJE Krumrey M Mattle D Messerschmidt MNissen P Reinhard L Sitsel O Sokaras DWilliams GJ Hau-Riege S Timneanu N CalemanC Chapman HN Boutet S and Schlichting I (2015)Indications of radiation damage in ferredoxin microcrys-tals using high-intensity X-FEL beams J Synch Rad22 225238
51 Sugahara M Mizohata E Nango E Suzuki MTanaka T Masuda T Tanaka R Shimamura TTanaka Y Suno C Ihara K Pan D KakinouchiK Sugiyama S Murata M Inoue T Tono KSong C Park J Kameshima T Hatsui T Joti YYabashi M and Iwata S (2014) Grease matrix as aversatile carrier of proteins for serial crystallographyNat Methods 12 6163
52 Tono K Nango E Sugahara M Song C Park JTanaka T Tanaka R Joti Y Kameshima T OnoS Hatsui T Mizohata E Suzuki M Shimamura TTanaka Y Iwata S and Yabashi M (2015) Diverseapplication platform for hard X-ray diffraction inSACLA (DAPHNIS) application to serial protein crys-tallography using an X-ray free-electron laser J SynchRad 22 532537
53 Joti Y Kameshima T Yamaga M Sugimoto TOkada K Abe T Furukawa Y Ohata T TanakaR Hatsui T and Yabashi M (2015) Data acquisitionsystem for X-ray free-electron laser experiments atSACLA J Synch Rad 22 571576
54 Yumoto H Mimura H Koyama T Matsuyama STono K Togashi T Inubushi Y Sato T TanakaT Kimura T Yokoyama H Kim J Sano YHachisu Y Yabashi M Ohashi H Ohmori HIshikawa T and Yamauchi K (2012) Focusing of X-ray free-electron laser pulses with reflective optics NatPhotonics 7 4347
55 White TA Kirian RA Martin AV Aquila ANass K Barty A and Chapman HN (2012)CrystFEL a software suite for snapshot serial crystallog-raphy J Appl Cryst 45 335341
56 Brehm W and Diederichs K (2014) Breaking theindexing ambiguity in serial crystallography ActaCryst D 70 101109
57 Karplus PA and Diederichs K (2012) Linking crystal-lographic model and data quality Science 33610301033
58 Evans PR and Murshudov GN (2013) How good aremy data and what is the resolution Acta Cryst D 6912041214
59 Bublitz M Nass K Drachmann ND MarkvardsenAJ Gutmann MJ Barends TRM Mattle DShoeman RL Doak RB Boutet SMesserschmidt M Seibert MM Williams GJFoucar L Reinhard L Sitsel O Gregersen JLClausen JD Boesen T Gotfryd K Wang K-TOlesen C Moslashller JV Nissen P and Schlichting I(2015) Structural studies of P-type ATPaseligand com-plexes using an X-ray free-electron laser IUCrJ 2409420
SFX revealed redox-coupled structural changes in CuNiR
11
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046
Y Fukuda et al
12
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
hydrogen bond with the hydroxyl oxygen atom ofThr268 and the backbone carbonyl oxygen atom ofGln267 (Fig 3b) The Hiscat rotation changes thestates of the bifurcated hydrogen bond To qualita-tively evaluate this subtle change we modelled hydro-gen atoms of Hiscat at the ideal positions withoutrefinement Figure 3c shows that with increasing ofX-ray dose the y1 (Nd1HOGln) angle and the y2(Nd1HOThr) angle decreased and increasedrespectively
Discussion
Geometrical changes at Cu sitesWe recently demonstrated that T1Cu in a GtNiR crys-tal is reduced by at least an X-ray dose of 0041 Mgy(30) Because even the X-ray dose for SR1 was higherthan this value (Table I) T1Cu in the present SRX
structures must be more or less reduced Howeverthe geometries around T1Cu did not show significantdifferences between the SRX structures and the com-pletely oxidized SFX structure (Table IV) It is notsurprising because spectroscopic studies have revealedthat the typical changes in ligandT1Cu distance inreduction are501 A so as to minimize the reorganiza-tion energy in ET (67)
We revealed that X-ray irradiation made the T2Cuatom gradually move towards to the ligand plane(Fig 1c) Sunken T2Cu atoms have been observed inthe reduced forms of other CuNiRs (6870)Therefore contrary to the previous report (24) ourresult indicates that T2Cu free of NO2
can be reducedin SRX This conclusion is reasonable because ourcrystal was crystallized at a pH low enough (pH 45)for intramolecular ET without NO2
binding to occurdominantly (28) Although the T2Cu sites in CuNiR
Fig 3 Redox-coupled rotation of Hiscat (a) Electron density maps (contoured at 40 ) around Hiscat in the SFX (red) and SR1 (yellow)
structures (b) Comparison of Hiscat of the SFX (red) SR1 (yellow) SR7 (teal blue) and RT SRX (white) structures (c) Changes in the angle ofthe bifurcated hydrogen bond The upper and lower series show y1 and y2 respectively Trends are shown by dashed lines (A colour version ofthis figure is available online at httpjboxfordjournalsorg)
SFX revealed redox-coupled structural changes in CuNiR
7
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
crystals lose ligand water when it is harshly reduced byartificial reductants (6870) it is just one of thereduced states of T2Cu called inactive reduced stateand a normal reduced state retains water ligand (29) aswas observed in our structures (Fig 1a)
Conformational change of Hiscat
The structural change of Hiscat cannot be caused bycoordination of Cu or the presence of Na+ above theT2Cu site because the former was found also in thecryogenic and RT SR structures (Supplementary FigS6 and S7) and the latter was found also in the RT SRstructure (Supplementary Fig S4c) The difference ofmeasurement temperature is not a major cause of thestructural change (Fig 3b) Thus the conformationaldifference in Hiscat between the SFX and SRX struc-tures is most likely to result from photoreduction ofthe Cu sites Indeed the observed conformationalchange was small and is probably due to the rigid cata-lytic site of thermostable GtNiR (71)
Interestingly there is a hydrogen bond betweenT2Cu ligand His100 and the side chain of Gln267
and His100 is located at the end of a sensor loop(Fig 4a) which is thought to transmit informationabout T1Cursquos redox state to T2Cu (8 72) Althoughthe ThrGln pair (or ThrGlu in some CuNiRs) com-posing the bifurcated hydrogen bond has been ignoredfor long they are conserved in CuNiRs except for afew CuNiRs which have Ser at the position of Thr(Fig 4b) The nitrite reduction activity of Ser-contain-ing CuNiR is lower than the activity of Thr-containingCuNiR (70 73) The crystal structures of Ser-contain-ing CuNiRs show that the Ser residue does not alwaysform a hydrogen bond with Hiscat (70 73) These factsimply the enzymatic importance of the hydrogen bondbetween Hiscat and Thr
The rotation of the imidazole ring changes the yangles of the bifurcated hydrogen bond (Fig 3c)Generally the y angle is used as an indicator of hydro-gen bond strength Strong moderate and weak hydro-gen bonds tend to show 170180 150180 90150
y angles respectively (74) Therefore the Hiscat rota-tion strengthens the hydrogen bond between Hiscat andThr and weakens the hydrogen bond between Hiscat
Fig 4 Conserved hydrogen bond network (a) Redox sensor loop in GtNiR The hydrogen bond network is shown by dotted lines (b) Amino acidsequences of CuNiRs Hiscat GlnGlu and ThrSer are indicated by filled circle triangle and diamond respectively Ax Ac Rs Rp Ph and Hdmeans Achromobacter xylosoxidans Achromobacter cycloclastes Rhodobacter sphaeroides Ralstonia pickettii Pseudoalteromonas haloplanktisand Hyphomicrobium denitrificans respectively (A colour version of this figure is available online at httpjboxfordjournalsorg)
Y Fukuda et al
8
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
and GlnGlu Because the hydroxyl oxygen atom is lessnegatively polarized than the carbonyl oxygen atomthis structural change may destabilize the positivecharge on Hiscat which may facilitate transfer of aproton to the bridging water between Aspcat andHiscat (Fig 4a) Hiscat has been suggested to directlyprovide a proton to NO2
(19 75) however our re-sults indicate another possibility Hiscat may functionas a switch for proton relay
Supplementary Data
Supplementary Data are available at JB Online
Acknowledgements
The SFX experiment was carried out at BL3 of the SPring-8Angstrom Compact Free-Electron Laser (SACLA) with the ap-proval of the Japan Synchrotron Radiation Research Institute(JASRI) (proposal no 2013B8045) We are grateful for supportfrom the SACLA High Performance Computing (HPC) systemand the Mini-K super computer system The microfocus SRX ex-periment at BL32XU of SPring-8 was supported by the Platform forDrug Discovery Informatics and Structural Life Science The au-thors thank MEP Murphy for discussion K Baba and N Mizunofor their help in the experiment at BL38B1 (proposal no2013A1592) E Yamashita and A Higashiura for their support atBL44XU of the SPring-8 and all staff at the SACLA for technicalassistance
FundingThis work was supported by the X-ray Free-Electron Laser PriorityStrategy Program of the Ministry of Education Culture SportsScience and Technology in Japan (MEXT) the Grant-in-Aid forScientific Research on Innovative Areas from MEXT the Grant-in-Aid for Japan Society for the Promotion of Science (JSPS)Research Fellows (Grant no 254626) and the JSPS KAKENHI(Grant no 15K18487)
Conflict of InterestNone declared
References
1 Gruber N and Galloway JN (2008) An Earth-systemperspective of the global nitrogen cycle Nature 451293296
2 Galloway JN Townsend AR Erisman JWBekunda M Cai Z Freney JR Martinelli LASeitzinger SP and Sutton MA (2008)Transformation of the nitrogen cycle recent trendsquestions and potential solutions Science 320 889892
3 Zumft WG (1997) Cell biology and molecular basis ofdenitrification Microbiol Mol Biol Rev 61 533616
4 Tavares P Pereira AS Moura JJ and Moura I(2006) Metalloenzymes of the denitrification pathwayJ Inorg Biochem 100 20872100
5 Fulop V Moir JWB Ferguson SJ and Hajdu J(1995) The anatomy of a bifunctional enzyme structuralbasis for reduction of oxygen to water and synthesis ofnitric oxide by cytochrome cd1 Cell 81 369377
6 Kakutani T Watanabe H Arima K and Beppu T(1981) A blue protein as inactivating factor for nitritereductase from Alcaligenes faecalis Strain S-6J Biochem 89 463472
7 MacPherson IS Rosell FI Scofield M MaukAG and Murphy ME (2010) Directed evolution of
copper nitrite reductase to a chromogenic reductantProtein Eng Des Sel 23 137145
8 Strange RW Murphy LM Dodd FE AbrahamZHL Eady RR Smith BE and Hasnain SS(1999) Structural and kinetic evidence for an orderedmechanism of copper nitrite reductase J Mol Biol287 10011009
9 Godden J Turley S Teller D Adman E Liu MPayne W and LeGall J (1991) The 23 angstrom X-raystructure of nitrite reductase from Achromobacter cyclo-clastes Science 253 438442
10 Kukimoto M Nishiyama M Murphy MEPTurley S Adman ET Horinouchi S and BeppuT (1994) X-ray structure and site-directed mutagenesisof a nitrite reductase from Alcaligenes Faecalis S-6 rolesof two copper atoms in nitrite reduction Biochemistry33 52465252
11 Dodd FE Beeumen JV Eady RR and HasnainSS (1998) X-ray structure of a blue-copper nitrite reduc-tase in two crystal forms The nature of the copper sitesmode of substrate binding and recognition by redox part-ner J Mol Biol 282 369382
12 Inoue T Gotowda M Deligeer Kataoka KYamaguchi K Suzuki S Watanabe H Gohow Mand Kai Y (1998) Type 1 Cu structure of blue nitritereductase from Alcaligenes xylosoxidans GIFU 1051 at205 A resolution comparison of blue and green nitritereductases J Biochem 124 876879
13 Merkle AC and Lehnert N (2012) Binding and acti-vation of nitrite and nitric oxide by copper nitrite reduc-tase and corresponding model complexes Dalton Trans41 33553368
14 Koteishi H Nojiri M Nakagami T Yamaguchi Kand Suzuki S (2009) Cytochrome c551 is a mediator ofelectron transfer between copper-containing nitrite re-ductase and azurin in a denitrifying bacteriumAchromobacter xylosoxidans Bull Chem Soc Jpn 8210031005
15 Nojiri M Koteishi H Nakagami T Kobayashi KInoue T Yamaguchi K and Suzuki S (2009)Structural basis of inter-protein electron transfer for ni-trite reduction in denitrification Nature 462 117120
16 Vlasie MD Fernandez-Busnadiego R Prudencio Mand Ubbink M (2008) Conformation of pseudoazurinin the 152 kDa electron transfer complex with nitrite re-ductase determined by paramagnetic NMR J Mol Biol375 14051415
17 Impagliazzo A Blok AJ Cliff MJ Ladbury JEand Ubbink M (2007) Redox-state-dependent complexformation between pseudoazurin and nitrite reductaseJ Am Chem Soc 129 226233
18 Boulanger MJ Kukimoto M Nishiyama MHorinouchi S and Murphy ME (2000) Catalyticroles for two water bridged residues (Asp-98 and His-255) in the active site of copper-containing nitrite reduc-tase J Biol Chem 275 2395723964
19 Kataoka K Furusawa H Takagi K Yamaguchi Kand Suzuki S (2000) Functional analysis of conservedaspartate and histidine residues located around the type 2copper site of copper-containing nitrite reductaseJ Biochem 127 345350
20 Yano J Kern J Irrgang KD Latimer MJBergmann U Glatzel P Pushkar Y Biesiadka JLoll B Sauer K Messinger J Zouni A andYachandra VK (2005) X-ray damage to the Mn4Cacomplex in single crystals of photosystem II a case
SFX revealed redox-coupled structural changes in CuNiR
9
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
study for metalloprotein crystallography Proc NatlAcad Sci USA 102 1204712052
21 Schlichting I Berendzen J Chu K Stock AMMaves SA Benson DE Sweet RM Ringe DPetsko GA and Sligar SG (2000) The catalytic path-way of cytochrome P450cam at atomic resolutionScience 287 16151622
22 Berglund GI Carlsson GH Smith AT Szoke HHenriksen A and Hajdu J (2002) The catalytic path-way of horseradish peroxidase at high resolution Nature417 463468
23 Beitlich T Kuhnel K Schulze-Briese C ShoemanRL and Schlichting I (2007) Cryoradiolytic reductionof crystalline heme proteins analysis by UV-Vis spec-troscopy and X-ray crystallography J Synch Rad 141123
24 Hough MA Antonyuk SV Strange RW EadyRR and Hasnain SS (2008) Crystallography withonline optical and X-ray absorption spectroscopies dem-onstrates an ordered mechanism in copper nitrite reduc-tase J Mol Biol 378 353361
25 Brenner S Heyes DJ Hay S Hough MA EadyRR Hasnain SS and Scrutton NS (2009)Demonstration of proton-coupled electron transfer inthe copper-containing nitrite reductases J Biol Chem284 2597325983
26 Leferink NG Han C Antonyuk SV Heyes DJRigby SE Hough MA Eady RR Scrutton NSand Hasnain SS (2011) Proton-coupled electron trans-fer in the catalytic cycle of Alcaligenes xylosoxidanscopper-dependent nitrite reductase Biochemistry 5041214131
27 Kobayashi K Tagawa S Deligeer and Suzuki S(1999) The pH-dependent changes of intramolecularelectron transfer on copper-containing nitrite reductaseJ Biochem 126 408412
28 Wijma HJ Jeuken LJ Verbeet MP ArmstrongFA and Canters GW (2006) A random-sequentialmechanism for nitrite binding and active site reductionin copper-containing nitrite reductase J Biol Chem281 1634016346
29 Wijma HJ Jeuken LJC Verbeet MP ArmstrongFA and Canters GW (2007) Protein film voltamme-try of copper-containing nitrite reductase reveals revers-ible inactivation J Am Chem Soc 129 85578565
30 Fukuda Y Tse KM Kado Y Mizohata EMatsumura H and Inoue T (2015) Insights into un-known foreign ligand in copper nitrite reductaseBiochem Biophys Res Commun 464 622628
31 Flot D Mairs T Giraud T Guijarro M LesourdM Rey V van Brussel D Morawe C Borel CHignette O Chavanne J Nurizzo D McSweeneyS and Mitchell E (2010) The ID23-2 structural biologymicrofocus beamline at the ESRF J Synch Rad 17107118
32 Chapman HN Fromme P Barty A White TAKirian RA Aquila A Hunter MS Schulz JDePonte DP Weierstall U Doak RB Maia FRMartin AV Schlichting I Lomb L Coppola NShoeman RL Epp SW Hartmann R Rolles DRudenko A Foucar L Kimmel N WeidenspointnerG Holl P Liang M Barthelmess M Caleman CBoutet S Bogan MJ Krzywinski J Bostedt CBajt S Gumprecht L Rudek B Erk B SchmidtC Homke A Reich C Pietschner D Struder LHauser G Gorke H Ullrich J Herrmann SSchaller G Schopper F Soltau H Kuhnel KU
Messerschmidt M Bozek JD Hau-Riege SPFrank M Hampton CY Sierra RG StarodubD Williams GJ Hajdu J Timneanu N SeibertMM Andreasson J Rocker A Jonsson OSvenda M Stern S Nass K Andritschke RSchroter CD Krasniqi F Bott M Schmidt KEWang X Grotjohann I Holton JM Barends TRNeutze R Marchesini S Fromme R Schorb SRupp D Adolph M Gorkhover T Andersson IHirsemann H Potdevin G Graafsma H NilssonB and Spence JC (2011) Femtosecond X-ray proteinnanocrystallography Nature 470 7377
33 Kern J Alonso-Mori R Hellmich J Tran RHattne J Laksmono H Glockner C Echols NSierra RG Sellberg J Lassalle-Kaiser B GildeaRJ Glatzel P Grosse-Kunstleve RW LatimerMJ McQueen TA DiFiore D Fry ARMesserschmidt M Miahnahri A Schafer DWSeibert MM Sokaras D Weng TC Zwart PHWhite WE Adams PD Bogan MJ Boutet SWilliams GJ Messinger J Sauter NK Zouni ABergmann U Yano J and Yachandra VK (2012)Room temperature femtosecond X-ray diffraction ofphotosystem II microcrystals Proc Natl Acad SciUSA 109 97219726
34 Johansson LC Arnlund D Katona G White TABarty A DePonte DP Shoeman RL WickstrandC Sharma A Williams GJ Aquila A Bogan MJCaleman C Davidsson J Doak RB Frank MFromme R Galli L Grotjohann I Hunter MSKassemeyer S Kirian RA Kupitz C Liang MLomb L Malmerberg E Martin AVMesserschmidt M Nass K Redecke L SeibertMM Sjohamn J Steinbrener J Stellato F WangD Wahlgren WY Weierstall U Westenhoff SZatsepin NA Boutet S Spence JC Schlichting IChapman HN Fromme P and Neutze R (2013)Structure of a photosynthetic reaction centre determinedby serial femtosecond crystallography Nat Commun 42911
35 Kupitz C Basu S Grotjohann I Fromme RZatsepin NA Rendek KN Hunter MSShoeman RL White TA Wang D James DYang JH Cobb DE Reeder B Sierra RG LiuH Barty A Aquila AL Deponte D Kirian RABari S Bergkamp JJ Beyerlein KR Bogan MJCaleman C Chao TC Conrad CE Davis KMFleckenstein H Galli L Hau-Riege SPKassemeyer S Laksmono H Liang M Lomb LMarchesini S Martin AV Messerschmidt MMilathianaki D Nass K Ros A Roy-ChowdhuryS Schmidt K Seibert M Steinbrener J Stellato FYan L Yoon C Moore TA Moore AL PushkarY Williams GJ Boutet S Doak RB WeierstallU Frank M Chapman HN Spence JC andFromme P (2014) Serial time-resolved crystallographyof photosystem II using a femtosecond X-ray laserNature 513 261265
36 Kern J Alonso-Mori R Tran R Hattne J GildeaRJ Echols N Glockner C Hellmich J LaksmonoH Sierra RG Lassalle-Kaiser B Koroidov SLampe A Han G Gul S DiFiore DMilathianaki D Fry AR Miahnahri A SchaferDW Messerschmidt M Seibert MM Koglin JESokaras D Weng T-C Sellberg J Latimer MJGrosse-Kunstleve RW Zwart PH White WEGlatzel P Adams PD Bogan MJ Williams GJBoutet S Messinger J Zouni A Sauter NK
Y Fukuda et al
10
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Yachandra VK Bergmann U and Yano J (2013)Simultaneous femtosecond X-ray spectroscopy and dif-fraction of photosystem II at room temperature Science340 491495
37 Kern J Tran R Alonso-Mori R Koroidov SEchols N Hattne J Ibrahim M Gul SLaksmono H Sierra RG Gildea RJ Han GHellmich J Lassalle-Kaiser B Chatterjee RBrewster AS Stan CA Glockner C Lampe ADiFiore D Milathianaki D Fry AR SeibertMM Koglin JE Gallo E Uhlig J Sokaras DWeng TC Zwart PH Skinner DE Bogan MJMesserschmidt M Glatzel P Williams GJ BoutetS Adams PD Zouni A Messinger J Sauter NKBergmann U Yano J and Yachandra VK (2014)Taking snapshots of photosynthetic water oxidationusing femtosecond X-ray diffraction and spectroscopyNat Commun 5 4371
38 Hirata K Kawano Y Ueno G Hashimoto KMurakami H Hasegawa K Hikima T KumasakaT and Yamamoto M (2013) Achievement of proteinmicro-crystallography at SPring-8 beamline BL32XUJ Phys Conf Ser 425 012002
39 Paithankar KS and Garman EF (2010) Know yourdose RADDOSE Acta Cryst D 66 381388
40 Tanida H Kikuchi A Miura K Takeshita KGoto S Shiro Y and Ishikawa T (2004) XAFS andprotein crystallography beamline BL38B1 at SPring-8AIP Conf Proc 705 486489
41 Fukuda Y and Inoue T (2015) High-temperatureand high-resolution crystallography of thermostablecopper nitrite reductase Chem Commun 5165326535
42 Otwinowski Z and Minor W (1997) Processing of X-ray diffraction data collected in oscillation modeMethods Enzymol 276 307326
43 Vagin A and Teplyakov A (2010) Molecular replace-ment with MOLREP Acta Cryst D 66 2225
44 Emsley P Lohkamp B Scott WG and Cowtan K(2010) Features and development of Coot Acta CrystD 66 486501
45 Murshudov GN Skubak P Lebedev AA PannuNS Steiner RA Nicholls RA Winn MD LongF and Vagin AA (2011) REFMAC5 for the refine-ment of macromolecular crystal structures Acta CrystD 67 355367
46 Winn MD Ballard CC Cowtan KD DodsonEJ Emsley P Evans PR Keegan RM KrissinelEB Leslie AG McCoy A McNicholas SJMurshudov GN Pannu NS Potterton EAPowell HR Read RJ Vagin A and Wilson KS(2011) Overview of the CCP4 suite and current develop-ments Acta Cryst D 67 235242
47 Chen VB Arendall WB III Headd JJ KeedyDA Immormino RM Kapral GJ Murray LWRichardson JS and Richardson DC (2010)MolProbity all-atom structure validation for macromol-ecular crystallography Acta Crystallogr D BiolCrystallogr 66 1221
48 Tono K Togashi T Inubushi Y Sato TKatayama T Ogawa K Ohashi H Kimura HTakahashi S Takeshita K Tomizawa H Goto SIshikawa T and Yabashi M (2013) Beamline experi-mental stations and photon beam diagnostics for thehard x-ray free electron laser of SACLA New J Phys15 083035
49 Son S-K Chapman HN and Santra R (2011)Multiwavelength anomalous diffraction at high X-rayintensity Phys Rev Lett 107 218102
50 Nass K Foucar L Barends TR Hartmann EBotha S Shoeman RL Doak RB Alonso-MoriR Aquila A Bajt S Barty A Bean R BeyerleinKR Bublitz M Drachmann N Gregersen JJonsson HO Kabsch W Kassemeyer S KoglinJE Krumrey M Mattle D Messerschmidt MNissen P Reinhard L Sitsel O Sokaras DWilliams GJ Hau-Riege S Timneanu N CalemanC Chapman HN Boutet S and Schlichting I (2015)Indications of radiation damage in ferredoxin microcrys-tals using high-intensity X-FEL beams J Synch Rad22 225238
51 Sugahara M Mizohata E Nango E Suzuki MTanaka T Masuda T Tanaka R Shimamura TTanaka Y Suno C Ihara K Pan D KakinouchiK Sugiyama S Murata M Inoue T Tono KSong C Park J Kameshima T Hatsui T Joti YYabashi M and Iwata S (2014) Grease matrix as aversatile carrier of proteins for serial crystallographyNat Methods 12 6163
52 Tono K Nango E Sugahara M Song C Park JTanaka T Tanaka R Joti Y Kameshima T OnoS Hatsui T Mizohata E Suzuki M Shimamura TTanaka Y Iwata S and Yabashi M (2015) Diverseapplication platform for hard X-ray diffraction inSACLA (DAPHNIS) application to serial protein crys-tallography using an X-ray free-electron laser J SynchRad 22 532537
53 Joti Y Kameshima T Yamaga M Sugimoto TOkada K Abe T Furukawa Y Ohata T TanakaR Hatsui T and Yabashi M (2015) Data acquisitionsystem for X-ray free-electron laser experiments atSACLA J Synch Rad 22 571576
54 Yumoto H Mimura H Koyama T Matsuyama STono K Togashi T Inubushi Y Sato T TanakaT Kimura T Yokoyama H Kim J Sano YHachisu Y Yabashi M Ohashi H Ohmori HIshikawa T and Yamauchi K (2012) Focusing of X-ray free-electron laser pulses with reflective optics NatPhotonics 7 4347
55 White TA Kirian RA Martin AV Aquila ANass K Barty A and Chapman HN (2012)CrystFEL a software suite for snapshot serial crystallog-raphy J Appl Cryst 45 335341
56 Brehm W and Diederichs K (2014) Breaking theindexing ambiguity in serial crystallography ActaCryst D 70 101109
57 Karplus PA and Diederichs K (2012) Linking crystal-lographic model and data quality Science 33610301033
58 Evans PR and Murshudov GN (2013) How good aremy data and what is the resolution Acta Cryst D 6912041214
59 Bublitz M Nass K Drachmann ND MarkvardsenAJ Gutmann MJ Barends TRM Mattle DShoeman RL Doak RB Boutet SMesserschmidt M Seibert MM Williams GJFoucar L Reinhard L Sitsel O Gregersen JLClausen JD Boesen T Gotfryd K Wang K-TOlesen C Moslashller JV Nissen P and Schlichting I(2015) Structural studies of P-type ATPaseligand com-plexes using an X-ray free-electron laser IUCrJ 2409420
SFX revealed redox-coupled structural changes in CuNiR
11
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046
Y Fukuda et al
12
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
crystals lose ligand water when it is harshly reduced byartificial reductants (6870) it is just one of thereduced states of T2Cu called inactive reduced stateand a normal reduced state retains water ligand (29) aswas observed in our structures (Fig 1a)
Conformational change of Hiscat
The structural change of Hiscat cannot be caused bycoordination of Cu or the presence of Na+ above theT2Cu site because the former was found also in thecryogenic and RT SR structures (Supplementary FigS6 and S7) and the latter was found also in the RT SRstructure (Supplementary Fig S4c) The difference ofmeasurement temperature is not a major cause of thestructural change (Fig 3b) Thus the conformationaldifference in Hiscat between the SFX and SRX struc-tures is most likely to result from photoreduction ofthe Cu sites Indeed the observed conformationalchange was small and is probably due to the rigid cata-lytic site of thermostable GtNiR (71)
Interestingly there is a hydrogen bond betweenT2Cu ligand His100 and the side chain of Gln267
and His100 is located at the end of a sensor loop(Fig 4a) which is thought to transmit informationabout T1Cursquos redox state to T2Cu (8 72) Althoughthe ThrGln pair (or ThrGlu in some CuNiRs) com-posing the bifurcated hydrogen bond has been ignoredfor long they are conserved in CuNiRs except for afew CuNiRs which have Ser at the position of Thr(Fig 4b) The nitrite reduction activity of Ser-contain-ing CuNiR is lower than the activity of Thr-containingCuNiR (70 73) The crystal structures of Ser-contain-ing CuNiRs show that the Ser residue does not alwaysform a hydrogen bond with Hiscat (70 73) These factsimply the enzymatic importance of the hydrogen bondbetween Hiscat and Thr
The rotation of the imidazole ring changes the yangles of the bifurcated hydrogen bond (Fig 3c)Generally the y angle is used as an indicator of hydro-gen bond strength Strong moderate and weak hydro-gen bonds tend to show 170180 150180 90150
y angles respectively (74) Therefore the Hiscat rota-tion strengthens the hydrogen bond between Hiscat andThr and weakens the hydrogen bond between Hiscat
Fig 4 Conserved hydrogen bond network (a) Redox sensor loop in GtNiR The hydrogen bond network is shown by dotted lines (b) Amino acidsequences of CuNiRs Hiscat GlnGlu and ThrSer are indicated by filled circle triangle and diamond respectively Ax Ac Rs Rp Ph and Hdmeans Achromobacter xylosoxidans Achromobacter cycloclastes Rhodobacter sphaeroides Ralstonia pickettii Pseudoalteromonas haloplanktisand Hyphomicrobium denitrificans respectively (A colour version of this figure is available online at httpjboxfordjournalsorg)
Y Fukuda et al
8
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
and GlnGlu Because the hydroxyl oxygen atom is lessnegatively polarized than the carbonyl oxygen atomthis structural change may destabilize the positivecharge on Hiscat which may facilitate transfer of aproton to the bridging water between Aspcat andHiscat (Fig 4a) Hiscat has been suggested to directlyprovide a proton to NO2
(19 75) however our re-sults indicate another possibility Hiscat may functionas a switch for proton relay
Supplementary Data
Supplementary Data are available at JB Online
Acknowledgements
The SFX experiment was carried out at BL3 of the SPring-8Angstrom Compact Free-Electron Laser (SACLA) with the ap-proval of the Japan Synchrotron Radiation Research Institute(JASRI) (proposal no 2013B8045) We are grateful for supportfrom the SACLA High Performance Computing (HPC) systemand the Mini-K super computer system The microfocus SRX ex-periment at BL32XU of SPring-8 was supported by the Platform forDrug Discovery Informatics and Structural Life Science The au-thors thank MEP Murphy for discussion K Baba and N Mizunofor their help in the experiment at BL38B1 (proposal no2013A1592) E Yamashita and A Higashiura for their support atBL44XU of the SPring-8 and all staff at the SACLA for technicalassistance
FundingThis work was supported by the X-ray Free-Electron Laser PriorityStrategy Program of the Ministry of Education Culture SportsScience and Technology in Japan (MEXT) the Grant-in-Aid forScientific Research on Innovative Areas from MEXT the Grant-in-Aid for Japan Society for the Promotion of Science (JSPS)Research Fellows (Grant no 254626) and the JSPS KAKENHI(Grant no 15K18487)
Conflict of InterestNone declared
References
1 Gruber N and Galloway JN (2008) An Earth-systemperspective of the global nitrogen cycle Nature 451293296
2 Galloway JN Townsend AR Erisman JWBekunda M Cai Z Freney JR Martinelli LASeitzinger SP and Sutton MA (2008)Transformation of the nitrogen cycle recent trendsquestions and potential solutions Science 320 889892
3 Zumft WG (1997) Cell biology and molecular basis ofdenitrification Microbiol Mol Biol Rev 61 533616
4 Tavares P Pereira AS Moura JJ and Moura I(2006) Metalloenzymes of the denitrification pathwayJ Inorg Biochem 100 20872100
5 Fulop V Moir JWB Ferguson SJ and Hajdu J(1995) The anatomy of a bifunctional enzyme structuralbasis for reduction of oxygen to water and synthesis ofnitric oxide by cytochrome cd1 Cell 81 369377
6 Kakutani T Watanabe H Arima K and Beppu T(1981) A blue protein as inactivating factor for nitritereductase from Alcaligenes faecalis Strain S-6J Biochem 89 463472
7 MacPherson IS Rosell FI Scofield M MaukAG and Murphy ME (2010) Directed evolution of
copper nitrite reductase to a chromogenic reductantProtein Eng Des Sel 23 137145
8 Strange RW Murphy LM Dodd FE AbrahamZHL Eady RR Smith BE and Hasnain SS(1999) Structural and kinetic evidence for an orderedmechanism of copper nitrite reductase J Mol Biol287 10011009
9 Godden J Turley S Teller D Adman E Liu MPayne W and LeGall J (1991) The 23 angstrom X-raystructure of nitrite reductase from Achromobacter cyclo-clastes Science 253 438442
10 Kukimoto M Nishiyama M Murphy MEPTurley S Adman ET Horinouchi S and BeppuT (1994) X-ray structure and site-directed mutagenesisof a nitrite reductase from Alcaligenes Faecalis S-6 rolesof two copper atoms in nitrite reduction Biochemistry33 52465252
11 Dodd FE Beeumen JV Eady RR and HasnainSS (1998) X-ray structure of a blue-copper nitrite reduc-tase in two crystal forms The nature of the copper sitesmode of substrate binding and recognition by redox part-ner J Mol Biol 282 369382
12 Inoue T Gotowda M Deligeer Kataoka KYamaguchi K Suzuki S Watanabe H Gohow Mand Kai Y (1998) Type 1 Cu structure of blue nitritereductase from Alcaligenes xylosoxidans GIFU 1051 at205 A resolution comparison of blue and green nitritereductases J Biochem 124 876879
13 Merkle AC and Lehnert N (2012) Binding and acti-vation of nitrite and nitric oxide by copper nitrite reduc-tase and corresponding model complexes Dalton Trans41 33553368
14 Koteishi H Nojiri M Nakagami T Yamaguchi Kand Suzuki S (2009) Cytochrome c551 is a mediator ofelectron transfer between copper-containing nitrite re-ductase and azurin in a denitrifying bacteriumAchromobacter xylosoxidans Bull Chem Soc Jpn 8210031005
15 Nojiri M Koteishi H Nakagami T Kobayashi KInoue T Yamaguchi K and Suzuki S (2009)Structural basis of inter-protein electron transfer for ni-trite reduction in denitrification Nature 462 117120
16 Vlasie MD Fernandez-Busnadiego R Prudencio Mand Ubbink M (2008) Conformation of pseudoazurinin the 152 kDa electron transfer complex with nitrite re-ductase determined by paramagnetic NMR J Mol Biol375 14051415
17 Impagliazzo A Blok AJ Cliff MJ Ladbury JEand Ubbink M (2007) Redox-state-dependent complexformation between pseudoazurin and nitrite reductaseJ Am Chem Soc 129 226233
18 Boulanger MJ Kukimoto M Nishiyama MHorinouchi S and Murphy ME (2000) Catalyticroles for two water bridged residues (Asp-98 and His-255) in the active site of copper-containing nitrite reduc-tase J Biol Chem 275 2395723964
19 Kataoka K Furusawa H Takagi K Yamaguchi Kand Suzuki S (2000) Functional analysis of conservedaspartate and histidine residues located around the type 2copper site of copper-containing nitrite reductaseJ Biochem 127 345350
20 Yano J Kern J Irrgang KD Latimer MJBergmann U Glatzel P Pushkar Y Biesiadka JLoll B Sauer K Messinger J Zouni A andYachandra VK (2005) X-ray damage to the Mn4Cacomplex in single crystals of photosystem II a case
SFX revealed redox-coupled structural changes in CuNiR
9
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
study for metalloprotein crystallography Proc NatlAcad Sci USA 102 1204712052
21 Schlichting I Berendzen J Chu K Stock AMMaves SA Benson DE Sweet RM Ringe DPetsko GA and Sligar SG (2000) The catalytic path-way of cytochrome P450cam at atomic resolutionScience 287 16151622
22 Berglund GI Carlsson GH Smith AT Szoke HHenriksen A and Hajdu J (2002) The catalytic path-way of horseradish peroxidase at high resolution Nature417 463468
23 Beitlich T Kuhnel K Schulze-Briese C ShoemanRL and Schlichting I (2007) Cryoradiolytic reductionof crystalline heme proteins analysis by UV-Vis spec-troscopy and X-ray crystallography J Synch Rad 141123
24 Hough MA Antonyuk SV Strange RW EadyRR and Hasnain SS (2008) Crystallography withonline optical and X-ray absorption spectroscopies dem-onstrates an ordered mechanism in copper nitrite reduc-tase J Mol Biol 378 353361
25 Brenner S Heyes DJ Hay S Hough MA EadyRR Hasnain SS and Scrutton NS (2009)Demonstration of proton-coupled electron transfer inthe copper-containing nitrite reductases J Biol Chem284 2597325983
26 Leferink NG Han C Antonyuk SV Heyes DJRigby SE Hough MA Eady RR Scrutton NSand Hasnain SS (2011) Proton-coupled electron trans-fer in the catalytic cycle of Alcaligenes xylosoxidanscopper-dependent nitrite reductase Biochemistry 5041214131
27 Kobayashi K Tagawa S Deligeer and Suzuki S(1999) The pH-dependent changes of intramolecularelectron transfer on copper-containing nitrite reductaseJ Biochem 126 408412
28 Wijma HJ Jeuken LJ Verbeet MP ArmstrongFA and Canters GW (2006) A random-sequentialmechanism for nitrite binding and active site reductionin copper-containing nitrite reductase J Biol Chem281 1634016346
29 Wijma HJ Jeuken LJC Verbeet MP ArmstrongFA and Canters GW (2007) Protein film voltamme-try of copper-containing nitrite reductase reveals revers-ible inactivation J Am Chem Soc 129 85578565
30 Fukuda Y Tse KM Kado Y Mizohata EMatsumura H and Inoue T (2015) Insights into un-known foreign ligand in copper nitrite reductaseBiochem Biophys Res Commun 464 622628
31 Flot D Mairs T Giraud T Guijarro M LesourdM Rey V van Brussel D Morawe C Borel CHignette O Chavanne J Nurizzo D McSweeneyS and Mitchell E (2010) The ID23-2 structural biologymicrofocus beamline at the ESRF J Synch Rad 17107118
32 Chapman HN Fromme P Barty A White TAKirian RA Aquila A Hunter MS Schulz JDePonte DP Weierstall U Doak RB Maia FRMartin AV Schlichting I Lomb L Coppola NShoeman RL Epp SW Hartmann R Rolles DRudenko A Foucar L Kimmel N WeidenspointnerG Holl P Liang M Barthelmess M Caleman CBoutet S Bogan MJ Krzywinski J Bostedt CBajt S Gumprecht L Rudek B Erk B SchmidtC Homke A Reich C Pietschner D Struder LHauser G Gorke H Ullrich J Herrmann SSchaller G Schopper F Soltau H Kuhnel KU
Messerschmidt M Bozek JD Hau-Riege SPFrank M Hampton CY Sierra RG StarodubD Williams GJ Hajdu J Timneanu N SeibertMM Andreasson J Rocker A Jonsson OSvenda M Stern S Nass K Andritschke RSchroter CD Krasniqi F Bott M Schmidt KEWang X Grotjohann I Holton JM Barends TRNeutze R Marchesini S Fromme R Schorb SRupp D Adolph M Gorkhover T Andersson IHirsemann H Potdevin G Graafsma H NilssonB and Spence JC (2011) Femtosecond X-ray proteinnanocrystallography Nature 470 7377
33 Kern J Alonso-Mori R Hellmich J Tran RHattne J Laksmono H Glockner C Echols NSierra RG Sellberg J Lassalle-Kaiser B GildeaRJ Glatzel P Grosse-Kunstleve RW LatimerMJ McQueen TA DiFiore D Fry ARMesserschmidt M Miahnahri A Schafer DWSeibert MM Sokaras D Weng TC Zwart PHWhite WE Adams PD Bogan MJ Boutet SWilliams GJ Messinger J Sauter NK Zouni ABergmann U Yano J and Yachandra VK (2012)Room temperature femtosecond X-ray diffraction ofphotosystem II microcrystals Proc Natl Acad SciUSA 109 97219726
34 Johansson LC Arnlund D Katona G White TABarty A DePonte DP Shoeman RL WickstrandC Sharma A Williams GJ Aquila A Bogan MJCaleman C Davidsson J Doak RB Frank MFromme R Galli L Grotjohann I Hunter MSKassemeyer S Kirian RA Kupitz C Liang MLomb L Malmerberg E Martin AVMesserschmidt M Nass K Redecke L SeibertMM Sjohamn J Steinbrener J Stellato F WangD Wahlgren WY Weierstall U Westenhoff SZatsepin NA Boutet S Spence JC Schlichting IChapman HN Fromme P and Neutze R (2013)Structure of a photosynthetic reaction centre determinedby serial femtosecond crystallography Nat Commun 42911
35 Kupitz C Basu S Grotjohann I Fromme RZatsepin NA Rendek KN Hunter MSShoeman RL White TA Wang D James DYang JH Cobb DE Reeder B Sierra RG LiuH Barty A Aquila AL Deponte D Kirian RABari S Bergkamp JJ Beyerlein KR Bogan MJCaleman C Chao TC Conrad CE Davis KMFleckenstein H Galli L Hau-Riege SPKassemeyer S Laksmono H Liang M Lomb LMarchesini S Martin AV Messerschmidt MMilathianaki D Nass K Ros A Roy-ChowdhuryS Schmidt K Seibert M Steinbrener J Stellato FYan L Yoon C Moore TA Moore AL PushkarY Williams GJ Boutet S Doak RB WeierstallU Frank M Chapman HN Spence JC andFromme P (2014) Serial time-resolved crystallographyof photosystem II using a femtosecond X-ray laserNature 513 261265
36 Kern J Alonso-Mori R Tran R Hattne J GildeaRJ Echols N Glockner C Hellmich J LaksmonoH Sierra RG Lassalle-Kaiser B Koroidov SLampe A Han G Gul S DiFiore DMilathianaki D Fry AR Miahnahri A SchaferDW Messerschmidt M Seibert MM Koglin JESokaras D Weng T-C Sellberg J Latimer MJGrosse-Kunstleve RW Zwart PH White WEGlatzel P Adams PD Bogan MJ Williams GJBoutet S Messinger J Zouni A Sauter NK
Y Fukuda et al
10
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Yachandra VK Bergmann U and Yano J (2013)Simultaneous femtosecond X-ray spectroscopy and dif-fraction of photosystem II at room temperature Science340 491495
37 Kern J Tran R Alonso-Mori R Koroidov SEchols N Hattne J Ibrahim M Gul SLaksmono H Sierra RG Gildea RJ Han GHellmich J Lassalle-Kaiser B Chatterjee RBrewster AS Stan CA Glockner C Lampe ADiFiore D Milathianaki D Fry AR SeibertMM Koglin JE Gallo E Uhlig J Sokaras DWeng TC Zwart PH Skinner DE Bogan MJMesserschmidt M Glatzel P Williams GJ BoutetS Adams PD Zouni A Messinger J Sauter NKBergmann U Yano J and Yachandra VK (2014)Taking snapshots of photosynthetic water oxidationusing femtosecond X-ray diffraction and spectroscopyNat Commun 5 4371
38 Hirata K Kawano Y Ueno G Hashimoto KMurakami H Hasegawa K Hikima T KumasakaT and Yamamoto M (2013) Achievement of proteinmicro-crystallography at SPring-8 beamline BL32XUJ Phys Conf Ser 425 012002
39 Paithankar KS and Garman EF (2010) Know yourdose RADDOSE Acta Cryst D 66 381388
40 Tanida H Kikuchi A Miura K Takeshita KGoto S Shiro Y and Ishikawa T (2004) XAFS andprotein crystallography beamline BL38B1 at SPring-8AIP Conf Proc 705 486489
41 Fukuda Y and Inoue T (2015) High-temperatureand high-resolution crystallography of thermostablecopper nitrite reductase Chem Commun 5165326535
42 Otwinowski Z and Minor W (1997) Processing of X-ray diffraction data collected in oscillation modeMethods Enzymol 276 307326
43 Vagin A and Teplyakov A (2010) Molecular replace-ment with MOLREP Acta Cryst D 66 2225
44 Emsley P Lohkamp B Scott WG and Cowtan K(2010) Features and development of Coot Acta CrystD 66 486501
45 Murshudov GN Skubak P Lebedev AA PannuNS Steiner RA Nicholls RA Winn MD LongF and Vagin AA (2011) REFMAC5 for the refine-ment of macromolecular crystal structures Acta CrystD 67 355367
46 Winn MD Ballard CC Cowtan KD DodsonEJ Emsley P Evans PR Keegan RM KrissinelEB Leslie AG McCoy A McNicholas SJMurshudov GN Pannu NS Potterton EAPowell HR Read RJ Vagin A and Wilson KS(2011) Overview of the CCP4 suite and current develop-ments Acta Cryst D 67 235242
47 Chen VB Arendall WB III Headd JJ KeedyDA Immormino RM Kapral GJ Murray LWRichardson JS and Richardson DC (2010)MolProbity all-atom structure validation for macromol-ecular crystallography Acta Crystallogr D BiolCrystallogr 66 1221
48 Tono K Togashi T Inubushi Y Sato TKatayama T Ogawa K Ohashi H Kimura HTakahashi S Takeshita K Tomizawa H Goto SIshikawa T and Yabashi M (2013) Beamline experi-mental stations and photon beam diagnostics for thehard x-ray free electron laser of SACLA New J Phys15 083035
49 Son S-K Chapman HN and Santra R (2011)Multiwavelength anomalous diffraction at high X-rayintensity Phys Rev Lett 107 218102
50 Nass K Foucar L Barends TR Hartmann EBotha S Shoeman RL Doak RB Alonso-MoriR Aquila A Bajt S Barty A Bean R BeyerleinKR Bublitz M Drachmann N Gregersen JJonsson HO Kabsch W Kassemeyer S KoglinJE Krumrey M Mattle D Messerschmidt MNissen P Reinhard L Sitsel O Sokaras DWilliams GJ Hau-Riege S Timneanu N CalemanC Chapman HN Boutet S and Schlichting I (2015)Indications of radiation damage in ferredoxin microcrys-tals using high-intensity X-FEL beams J Synch Rad22 225238
51 Sugahara M Mizohata E Nango E Suzuki MTanaka T Masuda T Tanaka R Shimamura TTanaka Y Suno C Ihara K Pan D KakinouchiK Sugiyama S Murata M Inoue T Tono KSong C Park J Kameshima T Hatsui T Joti YYabashi M and Iwata S (2014) Grease matrix as aversatile carrier of proteins for serial crystallographyNat Methods 12 6163
52 Tono K Nango E Sugahara M Song C Park JTanaka T Tanaka R Joti Y Kameshima T OnoS Hatsui T Mizohata E Suzuki M Shimamura TTanaka Y Iwata S and Yabashi M (2015) Diverseapplication platform for hard X-ray diffraction inSACLA (DAPHNIS) application to serial protein crys-tallography using an X-ray free-electron laser J SynchRad 22 532537
53 Joti Y Kameshima T Yamaga M Sugimoto TOkada K Abe T Furukawa Y Ohata T TanakaR Hatsui T and Yabashi M (2015) Data acquisitionsystem for X-ray free-electron laser experiments atSACLA J Synch Rad 22 571576
54 Yumoto H Mimura H Koyama T Matsuyama STono K Togashi T Inubushi Y Sato T TanakaT Kimura T Yokoyama H Kim J Sano YHachisu Y Yabashi M Ohashi H Ohmori HIshikawa T and Yamauchi K (2012) Focusing of X-ray free-electron laser pulses with reflective optics NatPhotonics 7 4347
55 White TA Kirian RA Martin AV Aquila ANass K Barty A and Chapman HN (2012)CrystFEL a software suite for snapshot serial crystallog-raphy J Appl Cryst 45 335341
56 Brehm W and Diederichs K (2014) Breaking theindexing ambiguity in serial crystallography ActaCryst D 70 101109
57 Karplus PA and Diederichs K (2012) Linking crystal-lographic model and data quality Science 33610301033
58 Evans PR and Murshudov GN (2013) How good aremy data and what is the resolution Acta Cryst D 6912041214
59 Bublitz M Nass K Drachmann ND MarkvardsenAJ Gutmann MJ Barends TRM Mattle DShoeman RL Doak RB Boutet SMesserschmidt M Seibert MM Williams GJFoucar L Reinhard L Sitsel O Gregersen JLClausen JD Boesen T Gotfryd K Wang K-TOlesen C Moslashller JV Nissen P and Schlichting I(2015) Structural studies of P-type ATPaseligand com-plexes using an X-ray free-electron laser IUCrJ 2409420
SFX revealed redox-coupled structural changes in CuNiR
11
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046
Y Fukuda et al
12
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
and GlnGlu Because the hydroxyl oxygen atom is lessnegatively polarized than the carbonyl oxygen atomthis structural change may destabilize the positivecharge on Hiscat which may facilitate transfer of aproton to the bridging water between Aspcat andHiscat (Fig 4a) Hiscat has been suggested to directlyprovide a proton to NO2
(19 75) however our re-sults indicate another possibility Hiscat may functionas a switch for proton relay
Supplementary Data
Supplementary Data are available at JB Online
Acknowledgements
The SFX experiment was carried out at BL3 of the SPring-8Angstrom Compact Free-Electron Laser (SACLA) with the ap-proval of the Japan Synchrotron Radiation Research Institute(JASRI) (proposal no 2013B8045) We are grateful for supportfrom the SACLA High Performance Computing (HPC) systemand the Mini-K super computer system The microfocus SRX ex-periment at BL32XU of SPring-8 was supported by the Platform forDrug Discovery Informatics and Structural Life Science The au-thors thank MEP Murphy for discussion K Baba and N Mizunofor their help in the experiment at BL38B1 (proposal no2013A1592) E Yamashita and A Higashiura for their support atBL44XU of the SPring-8 and all staff at the SACLA for technicalassistance
FundingThis work was supported by the X-ray Free-Electron Laser PriorityStrategy Program of the Ministry of Education Culture SportsScience and Technology in Japan (MEXT) the Grant-in-Aid forScientific Research on Innovative Areas from MEXT the Grant-in-Aid for Japan Society for the Promotion of Science (JSPS)Research Fellows (Grant no 254626) and the JSPS KAKENHI(Grant no 15K18487)
Conflict of InterestNone declared
References
1 Gruber N and Galloway JN (2008) An Earth-systemperspective of the global nitrogen cycle Nature 451293296
2 Galloway JN Townsend AR Erisman JWBekunda M Cai Z Freney JR Martinelli LASeitzinger SP and Sutton MA (2008)Transformation of the nitrogen cycle recent trendsquestions and potential solutions Science 320 889892
3 Zumft WG (1997) Cell biology and molecular basis ofdenitrification Microbiol Mol Biol Rev 61 533616
4 Tavares P Pereira AS Moura JJ and Moura I(2006) Metalloenzymes of the denitrification pathwayJ Inorg Biochem 100 20872100
5 Fulop V Moir JWB Ferguson SJ and Hajdu J(1995) The anatomy of a bifunctional enzyme structuralbasis for reduction of oxygen to water and synthesis ofnitric oxide by cytochrome cd1 Cell 81 369377
6 Kakutani T Watanabe H Arima K and Beppu T(1981) A blue protein as inactivating factor for nitritereductase from Alcaligenes faecalis Strain S-6J Biochem 89 463472
7 MacPherson IS Rosell FI Scofield M MaukAG and Murphy ME (2010) Directed evolution of
copper nitrite reductase to a chromogenic reductantProtein Eng Des Sel 23 137145
8 Strange RW Murphy LM Dodd FE AbrahamZHL Eady RR Smith BE and Hasnain SS(1999) Structural and kinetic evidence for an orderedmechanism of copper nitrite reductase J Mol Biol287 10011009
9 Godden J Turley S Teller D Adman E Liu MPayne W and LeGall J (1991) The 23 angstrom X-raystructure of nitrite reductase from Achromobacter cyclo-clastes Science 253 438442
10 Kukimoto M Nishiyama M Murphy MEPTurley S Adman ET Horinouchi S and BeppuT (1994) X-ray structure and site-directed mutagenesisof a nitrite reductase from Alcaligenes Faecalis S-6 rolesof two copper atoms in nitrite reduction Biochemistry33 52465252
11 Dodd FE Beeumen JV Eady RR and HasnainSS (1998) X-ray structure of a blue-copper nitrite reduc-tase in two crystal forms The nature of the copper sitesmode of substrate binding and recognition by redox part-ner J Mol Biol 282 369382
12 Inoue T Gotowda M Deligeer Kataoka KYamaguchi K Suzuki S Watanabe H Gohow Mand Kai Y (1998) Type 1 Cu structure of blue nitritereductase from Alcaligenes xylosoxidans GIFU 1051 at205 A resolution comparison of blue and green nitritereductases J Biochem 124 876879
13 Merkle AC and Lehnert N (2012) Binding and acti-vation of nitrite and nitric oxide by copper nitrite reduc-tase and corresponding model complexes Dalton Trans41 33553368
14 Koteishi H Nojiri M Nakagami T Yamaguchi Kand Suzuki S (2009) Cytochrome c551 is a mediator ofelectron transfer between copper-containing nitrite re-ductase and azurin in a denitrifying bacteriumAchromobacter xylosoxidans Bull Chem Soc Jpn 8210031005
15 Nojiri M Koteishi H Nakagami T Kobayashi KInoue T Yamaguchi K and Suzuki S (2009)Structural basis of inter-protein electron transfer for ni-trite reduction in denitrification Nature 462 117120
16 Vlasie MD Fernandez-Busnadiego R Prudencio Mand Ubbink M (2008) Conformation of pseudoazurinin the 152 kDa electron transfer complex with nitrite re-ductase determined by paramagnetic NMR J Mol Biol375 14051415
17 Impagliazzo A Blok AJ Cliff MJ Ladbury JEand Ubbink M (2007) Redox-state-dependent complexformation between pseudoazurin and nitrite reductaseJ Am Chem Soc 129 226233
18 Boulanger MJ Kukimoto M Nishiyama MHorinouchi S and Murphy ME (2000) Catalyticroles for two water bridged residues (Asp-98 and His-255) in the active site of copper-containing nitrite reduc-tase J Biol Chem 275 2395723964
19 Kataoka K Furusawa H Takagi K Yamaguchi Kand Suzuki S (2000) Functional analysis of conservedaspartate and histidine residues located around the type 2copper site of copper-containing nitrite reductaseJ Biochem 127 345350
20 Yano J Kern J Irrgang KD Latimer MJBergmann U Glatzel P Pushkar Y Biesiadka JLoll B Sauer K Messinger J Zouni A andYachandra VK (2005) X-ray damage to the Mn4Cacomplex in single crystals of photosystem II a case
SFX revealed redox-coupled structural changes in CuNiR
9
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
study for metalloprotein crystallography Proc NatlAcad Sci USA 102 1204712052
21 Schlichting I Berendzen J Chu K Stock AMMaves SA Benson DE Sweet RM Ringe DPetsko GA and Sligar SG (2000) The catalytic path-way of cytochrome P450cam at atomic resolutionScience 287 16151622
22 Berglund GI Carlsson GH Smith AT Szoke HHenriksen A and Hajdu J (2002) The catalytic path-way of horseradish peroxidase at high resolution Nature417 463468
23 Beitlich T Kuhnel K Schulze-Briese C ShoemanRL and Schlichting I (2007) Cryoradiolytic reductionof crystalline heme proteins analysis by UV-Vis spec-troscopy and X-ray crystallography J Synch Rad 141123
24 Hough MA Antonyuk SV Strange RW EadyRR and Hasnain SS (2008) Crystallography withonline optical and X-ray absorption spectroscopies dem-onstrates an ordered mechanism in copper nitrite reduc-tase J Mol Biol 378 353361
25 Brenner S Heyes DJ Hay S Hough MA EadyRR Hasnain SS and Scrutton NS (2009)Demonstration of proton-coupled electron transfer inthe copper-containing nitrite reductases J Biol Chem284 2597325983
26 Leferink NG Han C Antonyuk SV Heyes DJRigby SE Hough MA Eady RR Scrutton NSand Hasnain SS (2011) Proton-coupled electron trans-fer in the catalytic cycle of Alcaligenes xylosoxidanscopper-dependent nitrite reductase Biochemistry 5041214131
27 Kobayashi K Tagawa S Deligeer and Suzuki S(1999) The pH-dependent changes of intramolecularelectron transfer on copper-containing nitrite reductaseJ Biochem 126 408412
28 Wijma HJ Jeuken LJ Verbeet MP ArmstrongFA and Canters GW (2006) A random-sequentialmechanism for nitrite binding and active site reductionin copper-containing nitrite reductase J Biol Chem281 1634016346
29 Wijma HJ Jeuken LJC Verbeet MP ArmstrongFA and Canters GW (2007) Protein film voltamme-try of copper-containing nitrite reductase reveals revers-ible inactivation J Am Chem Soc 129 85578565
30 Fukuda Y Tse KM Kado Y Mizohata EMatsumura H and Inoue T (2015) Insights into un-known foreign ligand in copper nitrite reductaseBiochem Biophys Res Commun 464 622628
31 Flot D Mairs T Giraud T Guijarro M LesourdM Rey V van Brussel D Morawe C Borel CHignette O Chavanne J Nurizzo D McSweeneyS and Mitchell E (2010) The ID23-2 structural biologymicrofocus beamline at the ESRF J Synch Rad 17107118
32 Chapman HN Fromme P Barty A White TAKirian RA Aquila A Hunter MS Schulz JDePonte DP Weierstall U Doak RB Maia FRMartin AV Schlichting I Lomb L Coppola NShoeman RL Epp SW Hartmann R Rolles DRudenko A Foucar L Kimmel N WeidenspointnerG Holl P Liang M Barthelmess M Caleman CBoutet S Bogan MJ Krzywinski J Bostedt CBajt S Gumprecht L Rudek B Erk B SchmidtC Homke A Reich C Pietschner D Struder LHauser G Gorke H Ullrich J Herrmann SSchaller G Schopper F Soltau H Kuhnel KU
Messerschmidt M Bozek JD Hau-Riege SPFrank M Hampton CY Sierra RG StarodubD Williams GJ Hajdu J Timneanu N SeibertMM Andreasson J Rocker A Jonsson OSvenda M Stern S Nass K Andritschke RSchroter CD Krasniqi F Bott M Schmidt KEWang X Grotjohann I Holton JM Barends TRNeutze R Marchesini S Fromme R Schorb SRupp D Adolph M Gorkhover T Andersson IHirsemann H Potdevin G Graafsma H NilssonB and Spence JC (2011) Femtosecond X-ray proteinnanocrystallography Nature 470 7377
33 Kern J Alonso-Mori R Hellmich J Tran RHattne J Laksmono H Glockner C Echols NSierra RG Sellberg J Lassalle-Kaiser B GildeaRJ Glatzel P Grosse-Kunstleve RW LatimerMJ McQueen TA DiFiore D Fry ARMesserschmidt M Miahnahri A Schafer DWSeibert MM Sokaras D Weng TC Zwart PHWhite WE Adams PD Bogan MJ Boutet SWilliams GJ Messinger J Sauter NK Zouni ABergmann U Yano J and Yachandra VK (2012)Room temperature femtosecond X-ray diffraction ofphotosystem II microcrystals Proc Natl Acad SciUSA 109 97219726
34 Johansson LC Arnlund D Katona G White TABarty A DePonte DP Shoeman RL WickstrandC Sharma A Williams GJ Aquila A Bogan MJCaleman C Davidsson J Doak RB Frank MFromme R Galli L Grotjohann I Hunter MSKassemeyer S Kirian RA Kupitz C Liang MLomb L Malmerberg E Martin AVMesserschmidt M Nass K Redecke L SeibertMM Sjohamn J Steinbrener J Stellato F WangD Wahlgren WY Weierstall U Westenhoff SZatsepin NA Boutet S Spence JC Schlichting IChapman HN Fromme P and Neutze R (2013)Structure of a photosynthetic reaction centre determinedby serial femtosecond crystallography Nat Commun 42911
35 Kupitz C Basu S Grotjohann I Fromme RZatsepin NA Rendek KN Hunter MSShoeman RL White TA Wang D James DYang JH Cobb DE Reeder B Sierra RG LiuH Barty A Aquila AL Deponte D Kirian RABari S Bergkamp JJ Beyerlein KR Bogan MJCaleman C Chao TC Conrad CE Davis KMFleckenstein H Galli L Hau-Riege SPKassemeyer S Laksmono H Liang M Lomb LMarchesini S Martin AV Messerschmidt MMilathianaki D Nass K Ros A Roy-ChowdhuryS Schmidt K Seibert M Steinbrener J Stellato FYan L Yoon C Moore TA Moore AL PushkarY Williams GJ Boutet S Doak RB WeierstallU Frank M Chapman HN Spence JC andFromme P (2014) Serial time-resolved crystallographyof photosystem II using a femtosecond X-ray laserNature 513 261265
36 Kern J Alonso-Mori R Tran R Hattne J GildeaRJ Echols N Glockner C Hellmich J LaksmonoH Sierra RG Lassalle-Kaiser B Koroidov SLampe A Han G Gul S DiFiore DMilathianaki D Fry AR Miahnahri A SchaferDW Messerschmidt M Seibert MM Koglin JESokaras D Weng T-C Sellberg J Latimer MJGrosse-Kunstleve RW Zwart PH White WEGlatzel P Adams PD Bogan MJ Williams GJBoutet S Messinger J Zouni A Sauter NK
Y Fukuda et al
10
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Yachandra VK Bergmann U and Yano J (2013)Simultaneous femtosecond X-ray spectroscopy and dif-fraction of photosystem II at room temperature Science340 491495
37 Kern J Tran R Alonso-Mori R Koroidov SEchols N Hattne J Ibrahim M Gul SLaksmono H Sierra RG Gildea RJ Han GHellmich J Lassalle-Kaiser B Chatterjee RBrewster AS Stan CA Glockner C Lampe ADiFiore D Milathianaki D Fry AR SeibertMM Koglin JE Gallo E Uhlig J Sokaras DWeng TC Zwart PH Skinner DE Bogan MJMesserschmidt M Glatzel P Williams GJ BoutetS Adams PD Zouni A Messinger J Sauter NKBergmann U Yano J and Yachandra VK (2014)Taking snapshots of photosynthetic water oxidationusing femtosecond X-ray diffraction and spectroscopyNat Commun 5 4371
38 Hirata K Kawano Y Ueno G Hashimoto KMurakami H Hasegawa K Hikima T KumasakaT and Yamamoto M (2013) Achievement of proteinmicro-crystallography at SPring-8 beamline BL32XUJ Phys Conf Ser 425 012002
39 Paithankar KS and Garman EF (2010) Know yourdose RADDOSE Acta Cryst D 66 381388
40 Tanida H Kikuchi A Miura K Takeshita KGoto S Shiro Y and Ishikawa T (2004) XAFS andprotein crystallography beamline BL38B1 at SPring-8AIP Conf Proc 705 486489
41 Fukuda Y and Inoue T (2015) High-temperatureand high-resolution crystallography of thermostablecopper nitrite reductase Chem Commun 5165326535
42 Otwinowski Z and Minor W (1997) Processing of X-ray diffraction data collected in oscillation modeMethods Enzymol 276 307326
43 Vagin A and Teplyakov A (2010) Molecular replace-ment with MOLREP Acta Cryst D 66 2225
44 Emsley P Lohkamp B Scott WG and Cowtan K(2010) Features and development of Coot Acta CrystD 66 486501
45 Murshudov GN Skubak P Lebedev AA PannuNS Steiner RA Nicholls RA Winn MD LongF and Vagin AA (2011) REFMAC5 for the refine-ment of macromolecular crystal structures Acta CrystD 67 355367
46 Winn MD Ballard CC Cowtan KD DodsonEJ Emsley P Evans PR Keegan RM KrissinelEB Leslie AG McCoy A McNicholas SJMurshudov GN Pannu NS Potterton EAPowell HR Read RJ Vagin A and Wilson KS(2011) Overview of the CCP4 suite and current develop-ments Acta Cryst D 67 235242
47 Chen VB Arendall WB III Headd JJ KeedyDA Immormino RM Kapral GJ Murray LWRichardson JS and Richardson DC (2010)MolProbity all-atom structure validation for macromol-ecular crystallography Acta Crystallogr D BiolCrystallogr 66 1221
48 Tono K Togashi T Inubushi Y Sato TKatayama T Ogawa K Ohashi H Kimura HTakahashi S Takeshita K Tomizawa H Goto SIshikawa T and Yabashi M (2013) Beamline experi-mental stations and photon beam diagnostics for thehard x-ray free electron laser of SACLA New J Phys15 083035
49 Son S-K Chapman HN and Santra R (2011)Multiwavelength anomalous diffraction at high X-rayintensity Phys Rev Lett 107 218102
50 Nass K Foucar L Barends TR Hartmann EBotha S Shoeman RL Doak RB Alonso-MoriR Aquila A Bajt S Barty A Bean R BeyerleinKR Bublitz M Drachmann N Gregersen JJonsson HO Kabsch W Kassemeyer S KoglinJE Krumrey M Mattle D Messerschmidt MNissen P Reinhard L Sitsel O Sokaras DWilliams GJ Hau-Riege S Timneanu N CalemanC Chapman HN Boutet S and Schlichting I (2015)Indications of radiation damage in ferredoxin microcrys-tals using high-intensity X-FEL beams J Synch Rad22 225238
51 Sugahara M Mizohata E Nango E Suzuki MTanaka T Masuda T Tanaka R Shimamura TTanaka Y Suno C Ihara K Pan D KakinouchiK Sugiyama S Murata M Inoue T Tono KSong C Park J Kameshima T Hatsui T Joti YYabashi M and Iwata S (2014) Grease matrix as aversatile carrier of proteins for serial crystallographyNat Methods 12 6163
52 Tono K Nango E Sugahara M Song C Park JTanaka T Tanaka R Joti Y Kameshima T OnoS Hatsui T Mizohata E Suzuki M Shimamura TTanaka Y Iwata S and Yabashi M (2015) Diverseapplication platform for hard X-ray diffraction inSACLA (DAPHNIS) application to serial protein crys-tallography using an X-ray free-electron laser J SynchRad 22 532537
53 Joti Y Kameshima T Yamaga M Sugimoto TOkada K Abe T Furukawa Y Ohata T TanakaR Hatsui T and Yabashi M (2015) Data acquisitionsystem for X-ray free-electron laser experiments atSACLA J Synch Rad 22 571576
54 Yumoto H Mimura H Koyama T Matsuyama STono K Togashi T Inubushi Y Sato T TanakaT Kimura T Yokoyama H Kim J Sano YHachisu Y Yabashi M Ohashi H Ohmori HIshikawa T and Yamauchi K (2012) Focusing of X-ray free-electron laser pulses with reflective optics NatPhotonics 7 4347
55 White TA Kirian RA Martin AV Aquila ANass K Barty A and Chapman HN (2012)CrystFEL a software suite for snapshot serial crystallog-raphy J Appl Cryst 45 335341
56 Brehm W and Diederichs K (2014) Breaking theindexing ambiguity in serial crystallography ActaCryst D 70 101109
57 Karplus PA and Diederichs K (2012) Linking crystal-lographic model and data quality Science 33610301033
58 Evans PR and Murshudov GN (2013) How good aremy data and what is the resolution Acta Cryst D 6912041214
59 Bublitz M Nass K Drachmann ND MarkvardsenAJ Gutmann MJ Barends TRM Mattle DShoeman RL Doak RB Boutet SMesserschmidt M Seibert MM Williams GJFoucar L Reinhard L Sitsel O Gregersen JLClausen JD Boesen T Gotfryd K Wang K-TOlesen C Moslashller JV Nissen P and Schlichting I(2015) Structural studies of P-type ATPaseligand com-plexes using an X-ray free-electron laser IUCrJ 2409420
SFX revealed redox-coupled structural changes in CuNiR
11
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046
Y Fukuda et al
12
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
study for metalloprotein crystallography Proc NatlAcad Sci USA 102 1204712052
21 Schlichting I Berendzen J Chu K Stock AMMaves SA Benson DE Sweet RM Ringe DPetsko GA and Sligar SG (2000) The catalytic path-way of cytochrome P450cam at atomic resolutionScience 287 16151622
22 Berglund GI Carlsson GH Smith AT Szoke HHenriksen A and Hajdu J (2002) The catalytic path-way of horseradish peroxidase at high resolution Nature417 463468
23 Beitlich T Kuhnel K Schulze-Briese C ShoemanRL and Schlichting I (2007) Cryoradiolytic reductionof crystalline heme proteins analysis by UV-Vis spec-troscopy and X-ray crystallography J Synch Rad 141123
24 Hough MA Antonyuk SV Strange RW EadyRR and Hasnain SS (2008) Crystallography withonline optical and X-ray absorption spectroscopies dem-onstrates an ordered mechanism in copper nitrite reduc-tase J Mol Biol 378 353361
25 Brenner S Heyes DJ Hay S Hough MA EadyRR Hasnain SS and Scrutton NS (2009)Demonstration of proton-coupled electron transfer inthe copper-containing nitrite reductases J Biol Chem284 2597325983
26 Leferink NG Han C Antonyuk SV Heyes DJRigby SE Hough MA Eady RR Scrutton NSand Hasnain SS (2011) Proton-coupled electron trans-fer in the catalytic cycle of Alcaligenes xylosoxidanscopper-dependent nitrite reductase Biochemistry 5041214131
27 Kobayashi K Tagawa S Deligeer and Suzuki S(1999) The pH-dependent changes of intramolecularelectron transfer on copper-containing nitrite reductaseJ Biochem 126 408412
28 Wijma HJ Jeuken LJ Verbeet MP ArmstrongFA and Canters GW (2006) A random-sequentialmechanism for nitrite binding and active site reductionin copper-containing nitrite reductase J Biol Chem281 1634016346
29 Wijma HJ Jeuken LJC Verbeet MP ArmstrongFA and Canters GW (2007) Protein film voltamme-try of copper-containing nitrite reductase reveals revers-ible inactivation J Am Chem Soc 129 85578565
30 Fukuda Y Tse KM Kado Y Mizohata EMatsumura H and Inoue T (2015) Insights into un-known foreign ligand in copper nitrite reductaseBiochem Biophys Res Commun 464 622628
31 Flot D Mairs T Giraud T Guijarro M LesourdM Rey V van Brussel D Morawe C Borel CHignette O Chavanne J Nurizzo D McSweeneyS and Mitchell E (2010) The ID23-2 structural biologymicrofocus beamline at the ESRF J Synch Rad 17107118
32 Chapman HN Fromme P Barty A White TAKirian RA Aquila A Hunter MS Schulz JDePonte DP Weierstall U Doak RB Maia FRMartin AV Schlichting I Lomb L Coppola NShoeman RL Epp SW Hartmann R Rolles DRudenko A Foucar L Kimmel N WeidenspointnerG Holl P Liang M Barthelmess M Caleman CBoutet S Bogan MJ Krzywinski J Bostedt CBajt S Gumprecht L Rudek B Erk B SchmidtC Homke A Reich C Pietschner D Struder LHauser G Gorke H Ullrich J Herrmann SSchaller G Schopper F Soltau H Kuhnel KU
Messerschmidt M Bozek JD Hau-Riege SPFrank M Hampton CY Sierra RG StarodubD Williams GJ Hajdu J Timneanu N SeibertMM Andreasson J Rocker A Jonsson OSvenda M Stern S Nass K Andritschke RSchroter CD Krasniqi F Bott M Schmidt KEWang X Grotjohann I Holton JM Barends TRNeutze R Marchesini S Fromme R Schorb SRupp D Adolph M Gorkhover T Andersson IHirsemann H Potdevin G Graafsma H NilssonB and Spence JC (2011) Femtosecond X-ray proteinnanocrystallography Nature 470 7377
33 Kern J Alonso-Mori R Hellmich J Tran RHattne J Laksmono H Glockner C Echols NSierra RG Sellberg J Lassalle-Kaiser B GildeaRJ Glatzel P Grosse-Kunstleve RW LatimerMJ McQueen TA DiFiore D Fry ARMesserschmidt M Miahnahri A Schafer DWSeibert MM Sokaras D Weng TC Zwart PHWhite WE Adams PD Bogan MJ Boutet SWilliams GJ Messinger J Sauter NK Zouni ABergmann U Yano J and Yachandra VK (2012)Room temperature femtosecond X-ray diffraction ofphotosystem II microcrystals Proc Natl Acad SciUSA 109 97219726
34 Johansson LC Arnlund D Katona G White TABarty A DePonte DP Shoeman RL WickstrandC Sharma A Williams GJ Aquila A Bogan MJCaleman C Davidsson J Doak RB Frank MFromme R Galli L Grotjohann I Hunter MSKassemeyer S Kirian RA Kupitz C Liang MLomb L Malmerberg E Martin AVMesserschmidt M Nass K Redecke L SeibertMM Sjohamn J Steinbrener J Stellato F WangD Wahlgren WY Weierstall U Westenhoff SZatsepin NA Boutet S Spence JC Schlichting IChapman HN Fromme P and Neutze R (2013)Structure of a photosynthetic reaction centre determinedby serial femtosecond crystallography Nat Commun 42911
35 Kupitz C Basu S Grotjohann I Fromme RZatsepin NA Rendek KN Hunter MSShoeman RL White TA Wang D James DYang JH Cobb DE Reeder B Sierra RG LiuH Barty A Aquila AL Deponte D Kirian RABari S Bergkamp JJ Beyerlein KR Bogan MJCaleman C Chao TC Conrad CE Davis KMFleckenstein H Galli L Hau-Riege SPKassemeyer S Laksmono H Liang M Lomb LMarchesini S Martin AV Messerschmidt MMilathianaki D Nass K Ros A Roy-ChowdhuryS Schmidt K Seibert M Steinbrener J Stellato FYan L Yoon C Moore TA Moore AL PushkarY Williams GJ Boutet S Doak RB WeierstallU Frank M Chapman HN Spence JC andFromme P (2014) Serial time-resolved crystallographyof photosystem II using a femtosecond X-ray laserNature 513 261265
36 Kern J Alonso-Mori R Tran R Hattne J GildeaRJ Echols N Glockner C Hellmich J LaksmonoH Sierra RG Lassalle-Kaiser B Koroidov SLampe A Han G Gul S DiFiore DMilathianaki D Fry AR Miahnahri A SchaferDW Messerschmidt M Seibert MM Koglin JESokaras D Weng T-C Sellberg J Latimer MJGrosse-Kunstleve RW Zwart PH White WEGlatzel P Adams PD Bogan MJ Williams GJBoutet S Messinger J Zouni A Sauter NK
Y Fukuda et al
10
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Yachandra VK Bergmann U and Yano J (2013)Simultaneous femtosecond X-ray spectroscopy and dif-fraction of photosystem II at room temperature Science340 491495
37 Kern J Tran R Alonso-Mori R Koroidov SEchols N Hattne J Ibrahim M Gul SLaksmono H Sierra RG Gildea RJ Han GHellmich J Lassalle-Kaiser B Chatterjee RBrewster AS Stan CA Glockner C Lampe ADiFiore D Milathianaki D Fry AR SeibertMM Koglin JE Gallo E Uhlig J Sokaras DWeng TC Zwart PH Skinner DE Bogan MJMesserschmidt M Glatzel P Williams GJ BoutetS Adams PD Zouni A Messinger J Sauter NKBergmann U Yano J and Yachandra VK (2014)Taking snapshots of photosynthetic water oxidationusing femtosecond X-ray diffraction and spectroscopyNat Commun 5 4371
38 Hirata K Kawano Y Ueno G Hashimoto KMurakami H Hasegawa K Hikima T KumasakaT and Yamamoto M (2013) Achievement of proteinmicro-crystallography at SPring-8 beamline BL32XUJ Phys Conf Ser 425 012002
39 Paithankar KS and Garman EF (2010) Know yourdose RADDOSE Acta Cryst D 66 381388
40 Tanida H Kikuchi A Miura K Takeshita KGoto S Shiro Y and Ishikawa T (2004) XAFS andprotein crystallography beamline BL38B1 at SPring-8AIP Conf Proc 705 486489
41 Fukuda Y and Inoue T (2015) High-temperatureand high-resolution crystallography of thermostablecopper nitrite reductase Chem Commun 5165326535
42 Otwinowski Z and Minor W (1997) Processing of X-ray diffraction data collected in oscillation modeMethods Enzymol 276 307326
43 Vagin A and Teplyakov A (2010) Molecular replace-ment with MOLREP Acta Cryst D 66 2225
44 Emsley P Lohkamp B Scott WG and Cowtan K(2010) Features and development of Coot Acta CrystD 66 486501
45 Murshudov GN Skubak P Lebedev AA PannuNS Steiner RA Nicholls RA Winn MD LongF and Vagin AA (2011) REFMAC5 for the refine-ment of macromolecular crystal structures Acta CrystD 67 355367
46 Winn MD Ballard CC Cowtan KD DodsonEJ Emsley P Evans PR Keegan RM KrissinelEB Leslie AG McCoy A McNicholas SJMurshudov GN Pannu NS Potterton EAPowell HR Read RJ Vagin A and Wilson KS(2011) Overview of the CCP4 suite and current develop-ments Acta Cryst D 67 235242
47 Chen VB Arendall WB III Headd JJ KeedyDA Immormino RM Kapral GJ Murray LWRichardson JS and Richardson DC (2010)MolProbity all-atom structure validation for macromol-ecular crystallography Acta Crystallogr D BiolCrystallogr 66 1221
48 Tono K Togashi T Inubushi Y Sato TKatayama T Ogawa K Ohashi H Kimura HTakahashi S Takeshita K Tomizawa H Goto SIshikawa T and Yabashi M (2013) Beamline experi-mental stations and photon beam diagnostics for thehard x-ray free electron laser of SACLA New J Phys15 083035
49 Son S-K Chapman HN and Santra R (2011)Multiwavelength anomalous diffraction at high X-rayintensity Phys Rev Lett 107 218102
50 Nass K Foucar L Barends TR Hartmann EBotha S Shoeman RL Doak RB Alonso-MoriR Aquila A Bajt S Barty A Bean R BeyerleinKR Bublitz M Drachmann N Gregersen JJonsson HO Kabsch W Kassemeyer S KoglinJE Krumrey M Mattle D Messerschmidt MNissen P Reinhard L Sitsel O Sokaras DWilliams GJ Hau-Riege S Timneanu N CalemanC Chapman HN Boutet S and Schlichting I (2015)Indications of radiation damage in ferredoxin microcrys-tals using high-intensity X-FEL beams J Synch Rad22 225238
51 Sugahara M Mizohata E Nango E Suzuki MTanaka T Masuda T Tanaka R Shimamura TTanaka Y Suno C Ihara K Pan D KakinouchiK Sugiyama S Murata M Inoue T Tono KSong C Park J Kameshima T Hatsui T Joti YYabashi M and Iwata S (2014) Grease matrix as aversatile carrier of proteins for serial crystallographyNat Methods 12 6163
52 Tono K Nango E Sugahara M Song C Park JTanaka T Tanaka R Joti Y Kameshima T OnoS Hatsui T Mizohata E Suzuki M Shimamura TTanaka Y Iwata S and Yabashi M (2015) Diverseapplication platform for hard X-ray diffraction inSACLA (DAPHNIS) application to serial protein crys-tallography using an X-ray free-electron laser J SynchRad 22 532537
53 Joti Y Kameshima T Yamaga M Sugimoto TOkada K Abe T Furukawa Y Ohata T TanakaR Hatsui T and Yabashi M (2015) Data acquisitionsystem for X-ray free-electron laser experiments atSACLA J Synch Rad 22 571576
54 Yumoto H Mimura H Koyama T Matsuyama STono K Togashi T Inubushi Y Sato T TanakaT Kimura T Yokoyama H Kim J Sano YHachisu Y Yabashi M Ohashi H Ohmori HIshikawa T and Yamauchi K (2012) Focusing of X-ray free-electron laser pulses with reflective optics NatPhotonics 7 4347
55 White TA Kirian RA Martin AV Aquila ANass K Barty A and Chapman HN (2012)CrystFEL a software suite for snapshot serial crystallog-raphy J Appl Cryst 45 335341
56 Brehm W and Diederichs K (2014) Breaking theindexing ambiguity in serial crystallography ActaCryst D 70 101109
57 Karplus PA and Diederichs K (2012) Linking crystal-lographic model and data quality Science 33610301033
58 Evans PR and Murshudov GN (2013) How good aremy data and what is the resolution Acta Cryst D 6912041214
59 Bublitz M Nass K Drachmann ND MarkvardsenAJ Gutmann MJ Barends TRM Mattle DShoeman RL Doak RB Boutet SMesserschmidt M Seibert MM Williams GJFoucar L Reinhard L Sitsel O Gregersen JLClausen JD Boesen T Gotfryd K Wang K-TOlesen C Moslashller JV Nissen P and Schlichting I(2015) Structural studies of P-type ATPaseligand com-plexes using an X-ray free-electron laser IUCrJ 2409420
SFX revealed redox-coupled structural changes in CuNiR
11
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046
Y Fukuda et al
12
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
Yachandra VK Bergmann U and Yano J (2013)Simultaneous femtosecond X-ray spectroscopy and dif-fraction of photosystem II at room temperature Science340 491495
37 Kern J Tran R Alonso-Mori R Koroidov SEchols N Hattne J Ibrahim M Gul SLaksmono H Sierra RG Gildea RJ Han GHellmich J Lassalle-Kaiser B Chatterjee RBrewster AS Stan CA Glockner C Lampe ADiFiore D Milathianaki D Fry AR SeibertMM Koglin JE Gallo E Uhlig J Sokaras DWeng TC Zwart PH Skinner DE Bogan MJMesserschmidt M Glatzel P Williams GJ BoutetS Adams PD Zouni A Messinger J Sauter NKBergmann U Yano J and Yachandra VK (2014)Taking snapshots of photosynthetic water oxidationusing femtosecond X-ray diffraction and spectroscopyNat Commun 5 4371
38 Hirata K Kawano Y Ueno G Hashimoto KMurakami H Hasegawa K Hikima T KumasakaT and Yamamoto M (2013) Achievement of proteinmicro-crystallography at SPring-8 beamline BL32XUJ Phys Conf Ser 425 012002
39 Paithankar KS and Garman EF (2010) Know yourdose RADDOSE Acta Cryst D 66 381388
40 Tanida H Kikuchi A Miura K Takeshita KGoto S Shiro Y and Ishikawa T (2004) XAFS andprotein crystallography beamline BL38B1 at SPring-8AIP Conf Proc 705 486489
41 Fukuda Y and Inoue T (2015) High-temperatureand high-resolution crystallography of thermostablecopper nitrite reductase Chem Commun 5165326535
42 Otwinowski Z and Minor W (1997) Processing of X-ray diffraction data collected in oscillation modeMethods Enzymol 276 307326
43 Vagin A and Teplyakov A (2010) Molecular replace-ment with MOLREP Acta Cryst D 66 2225
44 Emsley P Lohkamp B Scott WG and Cowtan K(2010) Features and development of Coot Acta CrystD 66 486501
45 Murshudov GN Skubak P Lebedev AA PannuNS Steiner RA Nicholls RA Winn MD LongF and Vagin AA (2011) REFMAC5 for the refine-ment of macromolecular crystal structures Acta CrystD 67 355367
46 Winn MD Ballard CC Cowtan KD DodsonEJ Emsley P Evans PR Keegan RM KrissinelEB Leslie AG McCoy A McNicholas SJMurshudov GN Pannu NS Potterton EAPowell HR Read RJ Vagin A and Wilson KS(2011) Overview of the CCP4 suite and current develop-ments Acta Cryst D 67 235242
47 Chen VB Arendall WB III Headd JJ KeedyDA Immormino RM Kapral GJ Murray LWRichardson JS and Richardson DC (2010)MolProbity all-atom structure validation for macromol-ecular crystallography Acta Crystallogr D BiolCrystallogr 66 1221
48 Tono K Togashi T Inubushi Y Sato TKatayama T Ogawa K Ohashi H Kimura HTakahashi S Takeshita K Tomizawa H Goto SIshikawa T and Yabashi M (2013) Beamline experi-mental stations and photon beam diagnostics for thehard x-ray free electron laser of SACLA New J Phys15 083035
49 Son S-K Chapman HN and Santra R (2011)Multiwavelength anomalous diffraction at high X-rayintensity Phys Rev Lett 107 218102
50 Nass K Foucar L Barends TR Hartmann EBotha S Shoeman RL Doak RB Alonso-MoriR Aquila A Bajt S Barty A Bean R BeyerleinKR Bublitz M Drachmann N Gregersen JJonsson HO Kabsch W Kassemeyer S KoglinJE Krumrey M Mattle D Messerschmidt MNissen P Reinhard L Sitsel O Sokaras DWilliams GJ Hau-Riege S Timneanu N CalemanC Chapman HN Boutet S and Schlichting I (2015)Indications of radiation damage in ferredoxin microcrys-tals using high-intensity X-FEL beams J Synch Rad22 225238
51 Sugahara M Mizohata E Nango E Suzuki MTanaka T Masuda T Tanaka R Shimamura TTanaka Y Suno C Ihara K Pan D KakinouchiK Sugiyama S Murata M Inoue T Tono KSong C Park J Kameshima T Hatsui T Joti YYabashi M and Iwata S (2014) Grease matrix as aversatile carrier of proteins for serial crystallographyNat Methods 12 6163
52 Tono K Nango E Sugahara M Song C Park JTanaka T Tanaka R Joti Y Kameshima T OnoS Hatsui T Mizohata E Suzuki M Shimamura TTanaka Y Iwata S and Yabashi M (2015) Diverseapplication platform for hard X-ray diffraction inSACLA (DAPHNIS) application to serial protein crys-tallography using an X-ray free-electron laser J SynchRad 22 532537
53 Joti Y Kameshima T Yamaga M Sugimoto TOkada K Abe T Furukawa Y Ohata T TanakaR Hatsui T and Yabashi M (2015) Data acquisitionsystem for X-ray free-electron laser experiments atSACLA J Synch Rad 22 571576
54 Yumoto H Mimura H Koyama T Matsuyama STono K Togashi T Inubushi Y Sato T TanakaT Kimura T Yokoyama H Kim J Sano YHachisu Y Yabashi M Ohashi H Ohmori HIshikawa T and Yamauchi K (2012) Focusing of X-ray free-electron laser pulses with reflective optics NatPhotonics 7 4347
55 White TA Kirian RA Martin AV Aquila ANass K Barty A and Chapman HN (2012)CrystFEL a software suite for snapshot serial crystallog-raphy J Appl Cryst 45 335341
56 Brehm W and Diederichs K (2014) Breaking theindexing ambiguity in serial crystallography ActaCryst D 70 101109
57 Karplus PA and Diederichs K (2012) Linking crystal-lographic model and data quality Science 33610301033
58 Evans PR and Murshudov GN (2013) How good aremy data and what is the resolution Acta Cryst D 6912041214
59 Bublitz M Nass K Drachmann ND MarkvardsenAJ Gutmann MJ Barends TRM Mattle DShoeman RL Doak RB Boutet SMesserschmidt M Seibert MM Williams GJFoucar L Reinhard L Sitsel O Gregersen JLClausen JD Boesen T Gotfryd K Wang K-TOlesen C Moslashller JV Nissen P and Schlichting I(2015) Structural studies of P-type ATPaseligand com-plexes using an X-ray free-electron laser IUCrJ 2409420
SFX revealed redox-coupled structural changes in CuNiR
11
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046
Y Fukuda et al
12
at University of K
onstanz Library on February 23 2016
httpjboxfordjournalsorgD
ownloaded from
60 Zhang H Boulanger MJ Mauk AG MurphyMEP (2000) Carbon monoxide binding to copper-con-taining nitrite reductase from Alcaligenes faecalisJ Phys Chem B 104 1073810742
61 Nakamura K and Go N (2005) Function and molecu-lar evolution of multicopper blue proteins Cell MolLife Sci 62 20502066
62 Lawton TJ Sayavedra-Soto LA Arp DJ andRosenzweig AC (2009) Crystal structure of a two-domain multicopper oxidase implications for the evolu-tion of multicopper blue proteins J Biol Chem 2841017410180
63 Komori H Miyazaki K and Higuchi Y (2009) X-raystructure of a two-domain type laccase a missing link inthe evolution of multi-copper proteins FEBS Lett 58311891195
64 MacPherson IS and Murphy ME (2007) Type-2copper-containing enzymes Cell Mol Life Sci 6428872899
65 Fraser JS Clarkson MW Degnan SC Erion RKern D and Alber T (2009) Hidden alternative struc-tures of proline isomerase essential for catalysis Nature462 669673
66 Fraser JS van den Bedem H Samelson AJ LangPT Holton JM Echols N and Alber T (2011)Accessing protein conformational ensembles usingroom-temperature X-ray crystallography Proc NatlAcad Sci USA 108 1624716252
67 Solomon EI Szilagyi RK DeBeer George S andBasumallick L (2004) Electronic structures of metalsites in proteins and models contributions to functionin blue copper proteins Chem Rev 104 419458
68 Murphy MEP Turley S and Adman ET (1997)Structure of nitrite bound to copper-containing nitrite
reductase from Alcaligenes faecalis J Biol Chem 2722845528460
69 Jacobson F Guo H Olesen K Okvist M NeutzeR and Sjolin L (2005) Structures of the oxidized andreduced forms of nitrite reductase from Rhodobactersphaeroides 243 at high pH changes in the interactionsof the type 2 copper Acta Cryst D 61 11901198
70 Lawton TJ Bowen KE Sayavedra-Soto LA ArpDJ and Rosenzweig AC (2013) Characterization of anitrite reductase involved in nitrifier denitrificationJ Biol Chem 288 2557525583
71 Fukuda Y Tse KM Lintuluoto M Fukunishi YMizohata E Matsumura H Takami H Nojiri Mand Inoue T (2014) Structural insights into the functionof a thermostable copper-containing nitrite reductaseJ Biochem 155 123135
72 Hough MA Ellis MJ Antonyuk S Strange RWSawers G Eady RR and Samar Hasnain S (2005)High resolution structural studies of mutants provide in-sights into catalysis and electron transfer processes incopper nitrite reductase J Mol Biol 350 300309
73 Boulanger MJ and Murphy MEP (2002) Crystalstructure of the soluble domain of the major anaerobic-ally induced outer membrane protein (AniA) frompathogenic Neisseria a new class of copper-containingnitrite reductases J Mol Biol 315 11111127
74 Desiraju GR and Steiner T (1999) The WeakHydrogen Bonds in Structural Chemistry and BiologyOxford University Press Oxford
75 Antonyuk SV Strange RW Sawers G Eady RRand Hasnain SS (2005) Atomic resolution structures ofresting-state substrate- and product-complexed Cu-ni-trite reductase provide insight into catalytic mechanismProc Natl Acad SciUSA 102 1204112046