Original article Reduction in dose of chemical fertilizers and growth enhancement of sesame (Sesamum indicum L.) with application of rhizospheric competent Pseudomonas aeruginosa LES4 Sandeep Kumar a , Piyush Pandey b , D.K. Maheshwari a, * a Department of Botany and Microbiology, Gurukul Kangri University, Haridwar (U.A.) 249404, India b S.B.S.P.G. Institute of Biomedical Sciences and Research, Balawala, Dehradun (U.A.) 248161, India a r t i c l e i n f o Article history: Received 9 September 2008 Received in revised form 4 April 2009 Accepted 7 April 2009 Available online 19 April 2009 Handling editor:Kristina Lindstro ¨m Keywords: PGPRChemical fertilizer Macrophomina phaseolina Fusarium oxysporumoilseed crop Sesame a b s t r a c t Pseudomonas aeruginosaLES4, an isolate of tomato rhizosphere was found to be positive for several plant growth-promo ting attributes like production of indole acetic acid, HCN and siderophore, solubilization ofinorganic phosphate along with urease, chitinase and b-1-3-glucanase activity. In addition, it showed strong antagonistic effect against Macrophomina phaseolina and Fusarium oxysporum. P. aeruginosaLES4 caus ed halo cell forma tion and othe r morp holog ical deformit ies in my celia ofM. phaseo lina and F. oxysporum. Root colonization was studied with Tn5 induced streptomycin resistant transconjugants ofspontaneous tetracycline-resistant LES4 (designated LES4 tetraþstrepþ ) after different durations. The strain was significantly rhizospheric competent, as 17.4% increase in its population was recorded in sesame rhizosphere. Seed bacterization with LES4 resulted in significant increase in vegetative growth param- eters and yield of sesame over non-bacterized seeds. However, application of LES4 with half dose offertilizers resulted in growth equivalent to full dose treatment, without compromising with the growth and yield of sesame. Moreover, the oil yield increased by 33.3%, while protein yield increased by 47.5% with trea tmen t of half dose of fertiliz er along with LES 4 bact erize d seeds, as comp ared to full dose offertilizers. 2009 Elsevier Masson SAS. All rights reserved. 1. Introduction Sesame (Sesamum indicumL.) is an important oilseed crop, next to soybean and groundnut but has low yield potential [35]. India rank first in the world in area (about 2.47 m ha annually, 40% of the world ) and pr oduction (0. 74 m ton es , 27%of theworld ). In int ensive cropping system, supplementing soil nutrients by use of chemical fertilizer is considered inevitable for obtaining optimum yield ofcrops, however, their utilization efficiency remains low, due to loss by vol ati zat ion , den itr ific ati on, lea chi ng and con versi on int o unavailable forms. Continuous use of chemical fertilizers subverts the soil ecol ogy , disru pt envir onment, degra de soil fertility and cons equ ently shows harmf ul effec ts on human health[1] and contaminates ground water[15]. Presence of chemicals in sesame had been major impediment in the promotion of sesame export. Export consignments of sesame are sometimes unsuitable in the inter national market due to the pres ence of pest icide residue, resulting in loss of revenues[6]. Role of plant growth-promoting rhizobacteria (PGPR) in plant growth promotion and biological control of soil borne pathogens has been intensively investigated[17,19]. Integration of PGPR with tra ditional ino rga nic fer tili zers in the field may pr ove to be effective means to increase the solubility of insoluble phosphorous ions and other minerals to plants with simultaneous reduction in diseases incidence. They take systemic and simultaneous account of envi- ronmenta l asp ect s, quali ty of the pr oduce and profitabilit y ofagriculture [22]. Alternatively, substitution of chemicals wit h bac teria l fer til izers and bio pes tic ide s, esp eci all y blendi ng ofchemical fertilizers with chemical adaptive PGPR is a promising approach to obtain sustainable fertility of the soil and plant growth [39]. There are seve ral PGPR curren tly commerc ialize d whose growth-promoting activity in crop plants have been demonstrated in seve ral way s inclu ding pro duct ion of iron- seq ueste ring side - rophores and antimicrobial compounds that hinder colonization ofhosts by phytopathogens [40] , induction of host systemic disease resistance, solubilization of precipitated mineral nutrients and for prod uction of plant growth hormones, ther eby enhancing the plants ability to take up nutri ents from soil and incre asing yield [8]. *Corresponding author. Tel.: þ91 1334 246 767 (O), 265 469 (R), þ91 983 730 8897 (M); fax:þ91 13 34 246 767 (O). E-ma il addre sses : [email protected](P . Pandey), maheshwaridk@ gmail.com (D.K. Maheshwari). Contents lists available at ScienceDirect European Journal of Soil Biology journal homepage: http://www.elsevier.com/locate/ejsobi 1164-5563/$ – see front matter 2009 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.ejsobi.2009.04.002 European Journal of Soil Biology 45 (2009) 334–340
Transcript
8/16/2019 Reduction in Dose of Chemical Fertilizers and Growth Enhancement of Sesame Sesamum Indicum L With Applicati…
Reduction in dose of chemical fertilizers and growth enhancement of sesame
(Sesamum indicum L.) with application of rhizospheric competent
Pseudomonas aeruginosa LES4
Sandeep Kumar a, Piyush Pandey b, D.K. Maheshwari a,*
a Department of Botany and Microbiology, Gurukul Kangri University, Haridwar (U.A.) 249404, Indiab S.B.S.P.G. Institute of Biomedical Sciences and Research, Balawala, Dehradun (U.A.) 248161, India
a r t i c l e i n f o
Article history:
Received 9 September 2008
Received in revised form
4 April 2009
Accepted 7 April 2009
Available online 19 April 2009
Handling editor: Kristina Lindstro m
Keywords:
PGPR
Chemical fertilizer
Macrophomina phaseolina
Fusarium oxysporum oilseed crop
Sesame
a b s t r a c t
Pseudomonas aeruginosa LES4, an isolate of tomato rhizosphere was found to be positive for several plant
growth-promoting attributes like production of indole acetic acid, HCN and siderophore, solubilization of
inorganic phosphate along with urease, chitinase and b-1-3-glucanase activity. In addition, it showed
strong antagonistic effect against Macrophomina phaseolina and Fusarium oxysporum. P. aeruginosa LES4
caused halo cell formation and other morphological deformities in mycelia of M. phaseolina and
F. oxysporum. Root colonization was studied with Tn5 induced streptomycin resistant transconjugants of
spontaneous tetracycline-resistant LES4 (designated LES4tetraþstrepþ) after different durations. The strain
was significantly rhizospheric competent, as 17.4% increase in its population was recorded in sesame
rhizosphere. Seed bacterization with LES4 resulted in significant increase in vegetative growth param-
eters and yield of sesame over non-bacterized seeds. However, application of LES4 with half dose of
fertilizers resulted in growth equivalent to full dose treatment, without compromising with the growth
and yield of sesame. Moreover, the oil yield increased by 33.3%, while protein yield increased by 47.5%
with treatment of half dose of fertilizer along with LES 4 bacterized seeds, as compared to full dose of
fertilizers. 2009 Elsevier Masson SAS. All rights reserved.
1. Introduction
Sesame (Sesamum indicum L.) is an important oilseed crop, next
to soybean and groundnut but has low yield potential [35]. India
rank first in the world in area (about 2.47 m ha annually, 40% of the
world) and production (0.74 m tones, 27%of theworld). In intensive
cropping system, supplementing soil nutrients by use of chemical
fertilizer is considered inevitable for obtaining optimum yield of
crops, however, their utilization efficiency remains low, due to loss
by volatization, denitrification, leaching and conversion into
unavailable forms. Continuous use of chemical fertilizers subvertsthe soil ecology, disrupt environment, degrade soil fertility and
consequently shows harmful effects on human health [1] and
contaminates ground water [15]. Presence of chemicals in sesame
had been major impediment in the promotion of sesame export.
Export consignments of sesame are sometimes unsuitable in the
international market due to the presence of pesticide residue,
resulting in loss of revenues [6].
Role of plant growth-promoting rhizobacteria (PGPR) in plant
growth promotion and biological control of soil borne pathogens
has been intensively investigated [17,19]. Integration of PGPR with
traditional inorganic fertilizers in the field may prove to be effective
means to increase the solubility of insoluble phosphorous ions and
other minerals to plants with simultaneous reduction in diseases
incidence. They take systemic and simultaneous account of envi-
ronmental aspects, quality of the produce and profitability of
agriculture [22]. Alternatively, substitution of chemicals withbacterial fertilizers and biopesticides, especially blending of
chemical fertilizers with chemical adaptive PGPR is a promising
approach to obtain sustainable fertility of the soil and plant growth
[39]. There are several PGPR currently commercialized whose
growth-promoting activity in crop plants have been demonstrated
in several ways including production of iron-sequestering side-
rophores and antimicrobial compounds that hinder colonization of
hosts by phytopathogens [40], induction of host systemic disease
resistance, solubilization of precipitated mineral nutrients and for
production of plant growth hormones, thereby enhancing the
plants ability to take up nutrients from soil and increasing yield [8].
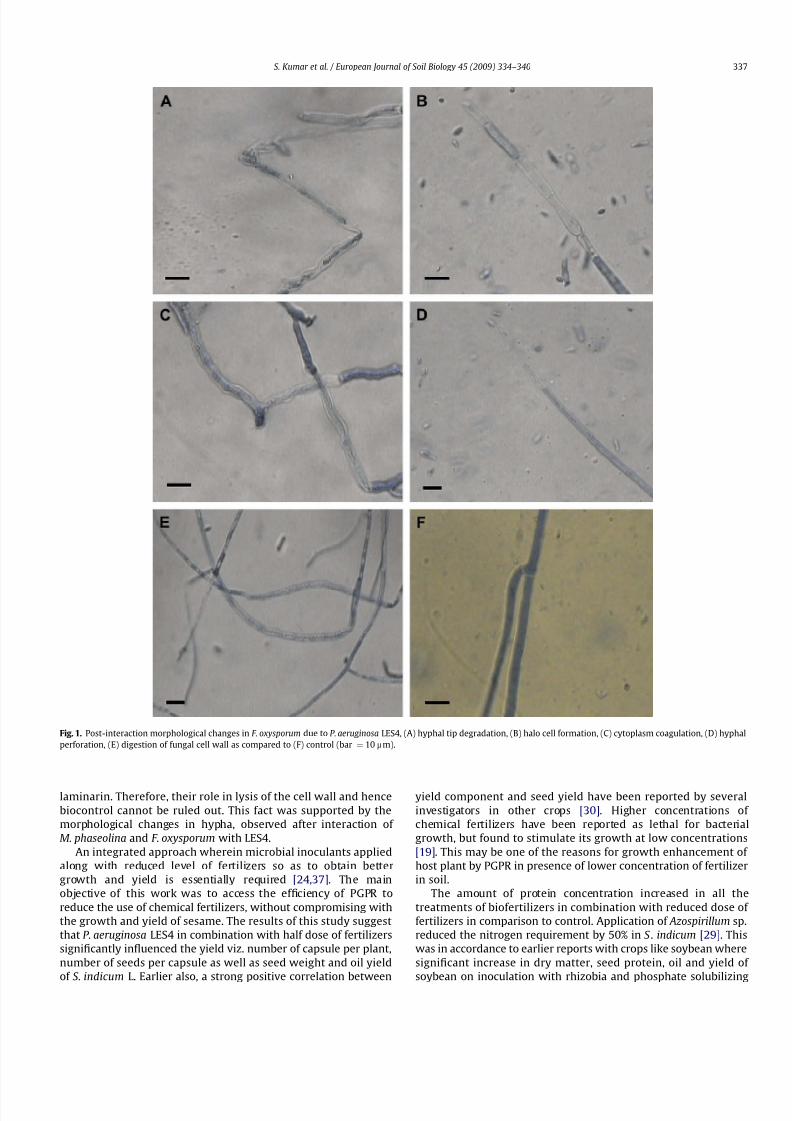
laminarin. Therefore, their role in lysis of the cell wall and hence
biocontrol cannot be ruled out. This fact was supported by the
morphological changes in hypha, observed after interaction of
M. phaseolina and F. oxysporum with LES4.
An integrated approach wherein microbial inoculants applied
along with reduced level of fertilizers so as to obtain better
growth and yield is essentially required [24,37]. The main
objective of this work was to access the efficiency of PGPR to
reduce the use of chemical fertilizers, without compromising with
the growth and yield of sesame. The results of this study suggest
that P. aeruginosa LES4 in combination with half dose of fertilizers
significantly influenced the yield viz. number of capsule per plant,
number of seeds per capsule as well as seed weight and oil yield
of S. indicum L. Earlier also, a strong positive correlation between
yield component and seed yield have been reported by several
investigators in other crops [30]. Higher concentrations of
chemical fertilizers have been reported as lethal for bacterial
growth, but found to stimulate its growth at low concentrations
[19]. This may be one of the reasons for growth enhancement of
host plant by PGPR in presence of lower concentration of fertilizer
in soil.
The amount of protein concentration increased in all the
treatments of biofertilizers in combination with reduced dose of
fertilizers in comparison to control. Application of Azospirillum sp.
reduced the nitrogen requirement by 50% in S . indicum [29]. This
was in accordance to earlier reports with crops like soybean where
significant increase in dry matter, seed protein, oil and yield of
soybean on inoculation with rhizobia and phosphate solubilizing
Fig. 1. Post-interaction morphological changes in F. oxysporum due to P. aeruginosa LES4, (A) hyphal tip degradation, (B) halo cell formation, (C) cytoplasm coagulation, (D) hyphal
perforation, (E) digestion of fungal cell wall as compared to (F) control (bar ¼ 10 mm).
S. Kumar et al. / European Journal of Soil Biology 45 (2009) 334–340 337
8/16/2019 Reduction in Dose of Chemical Fertilizers and Growth Enhancement of Sesame Sesamum Indicum L With Applicati…
bacteria in combination with chemical fertilizers was reported
[21,43]. Contrary to this, Poul and Savithri [28] reported that thegrowth and yield of sesame with application of half dose of urea in
combination with biofertilizers was less as compared to dose of
inorganic fertilizer. However, in present study, P. aeruginosa LES4
resulted equivalent yield as well as growth when applied with half
dose of fertilizers, as compared to application of full dose of
fertilizers, which may be because the difference in PGPR used.
Strong root colonization ability of LES4 lies in it being the
successful efficient colonizer in the rhizosphere of sesame. The use of
genetic markers such as intrinsic levels of resistance to various anti-
bioticsis oneof the simple and rapid methods for strain identification
in rhizosphere [25,27], and hence utilized in this study. Combined
resistance against two broad range antibiotics in introduced marker
strain LES4tetraþstrepþ allowed no background population of other
strains, which was further assured by checking the phenotypic and
physiological characteristics. The LES4tetraþstrepþ showed efficient
root colonization after different time intervals of observation (i.e. 30,
60, and 90 days) and enhanced plant growth and growth yield. The
population density increased up to 60 days of inoculation, which
became almost stable thereafter. It has been suggested that bacteria
that attain CFU of aboutz
10
3
per gram orhigher on rootmasscan beconsidered as good colonizers [31], and we obtained population of
LES4tetraþstrepþ of z107. The continued presence of P. aeruginosa
LES4tetraþstrepþ for 90 days in soil showed that it had reached
homeostasis after undergoing exchange with indigenous microflora
andis not affected by the active andpassive processes restricting soil
community. These results were similar to root colonization of
P. aeruginosa NBRI2650 [25], however they studied the colonization
up to60 days only.Earlier, P. aeruginosa hasbeen reported as potential
biocontrol agentand simultaneously enhancedplant growth and root
colonization of various vegetables and cereals without showing any
deleterious effects of plants [13,27,34].
On the basis of present findings, it may be concluded that
P. aeruginosa LES4 contains large number of plant growth-promoting
attributes, and its application contributed in enhancement of
sesame growth leading to better yield. In addition, its ability to
reduce the chemical requirement for obtaining optimum yield of
sesame appears to be of great ecological and economic importance,
with possibilities of an efficient bioinoculant.
Fig. 2. Post-interaction morphological changes in M. phaseolina due to P. aeruginosa LES4, (A) sclerotial cease in M. phaseolina, (B) halo cell formation, (C) mycelial deformities in
M. phaseolina as compare to (D) control (bar ¼ 10 mm).
Table 2
Root colonization of S. indicum L. by P. aeruginosa LES4 and standard strain MTCC-1934.
Interactive effect of Biofertilizers and Integrated Nutrient Management Protocol on the seed germination and growth and yield component of S. indicum L. cv. ST-1 after 120 DAS.
SEM 0.732 0.437 0.756 0.247 0.563 1.168 0.611 0.939 1.185
CD at 1% 3.161 1.886 3.264 1.068 2.432 5.044 2.640 4.054 5.119
CD at 5% 2.255 1.346 2.329 0.762 1.735 3.599 1.883 2.893 3.653
Values are mean of 10 randomly selected plants from each set; ns, not significant; *significant at 5% LSD; **significant at 1% level of LSD as compared to control; full dose of
chemical fertilizers N40þ40þ40, P30, K 30; half doses of chemical fertilizers N20þ20þ20, P15, K15.
Table 4
Plant growth-promoting and antifungal properties of isolated fluorescent Pseudomonas species.
Strains IAAA Phosphate solubilizationB SiderophoreC HCND ChitinaseE b-1,3-glucanaseF Antagonism against
M. phaseolina
Antagonism against
F. oxysporum
LES1 þ þ þ þ
LES2 þ þ þ LES3 þ þþ þ þ
LES4 þ þþþ þþþ þþ þ þ þþþ þþ
LES5 þ þ þ þ
LES6 þ þ þþ
LES7 þ þþ þ þ þ þ þþ þ
MTCC-1934 þ þ
MTCC-103 þ þ þ þ
MTCC-102 þ þ
Abbreviations: A, IAA negative; þ, IAA positive; B, phosphate solubilization negative; þ, phosphate solubilization positive; C, absence of halo formation; þ, small
halos < 0.5 cm wide surrounding colonies; þþ, medium halos > 0.5 cm wide surrounding colonies; þþþ, large halos > 1.0 cm wide surrounding colonies; D, HCN negative;
þ, HCN positive; E, chitinase negative; þ, chitinase positive small halos < 0.5 cm wide surrounding colonies; þþ, medium halos > 0.5 cm wide surrounding colonies; F, b-
1,3-glucanase negative, þ, b-1,3-glucanase positive;G, no inhibition; þ, 0–25% inhibition; þþ, 26–50% inhibition; þþþ, 60–85% inhibition against M. phaseolina; P. aeruginosa
MTCC-1934; P. fluorescens MTCC-103; P. putida MTCC-102.
Table 3
Morphological, physiological and biochemical characters of fluorescent Pseudomonas strains with standard strains.
The authors wish to thank the TMOP & M, CSIR, New Delhi for
financial assistance.
References
[1] S. Ayala, E.V.S.P. Rao, Perspective of soil fertility management with a focus onfertilizer use for crop productivity, Curr. Sci. 82 (2002) 797–807.
[2] M. Bano, J. Musarrat, Characterization of a new Pseudomonas aeruginosa strainNJ-15 as a potential biocontrol agent, Curr. Microbiol. 43 (2002) 182–186.
[3] S. Bhaita, R.C. Dubey, D.K. Maheshwari, Antagonistic effect of fluorescentpseudomonads against Macrophomina phaseolina that causes charcoal rot of groundnut, Ind. J. Exp. Biol. 41 (2003) 1442–1446.
[4] M.M. Bradford, A rapid and sensitive method for the quantitation of micro-organism quantities of protein utilizing the principle of protein dye binding,Anal. Biochem. 72 (1976) 248–254.
[5] K. Das, V. Katiyar, R. Goel, P solubilization potential of plant growth-promotingPseudomonas mutants at low temperature, Microbiol. Res. 158 (2003)359–362.
[6] S.S. Duhoon, A. Jyoteishi, M.R. Deshmukh, N.B. Singh, Optimization of Sesamum indicum Production through Bio-natural Inputs. In: 4th InternationalCrop Science Congress, The Regional Institute, Sydney, 2004.
[7] C. Dunne, J.J. Crowely, Y. Moenne-Loccoz, D.N. Dowling, F.J. de Brujin, F. O’Gara,Biological control of Pythium ultimum by Stenotrophomonas maltophislia W81
is mediated by an extracellular proteolytic activity, Microbiology 143 (1997)3921–3931.[8] B.R. Glick, The enhancement of plant growth by free-living bacteria, Can.
J. Microbiol. 47 (1995) 109–117.[9] A.K. Gomez, A.A. Gomez, Statistical Procedure for Agricultural Research, John
Wiley and Sons, New York, 1984.[10] A.O. Adesemoye, M. Obini, E.O. Ugoji, Comparison of plant growth-promotion
with Pseudomonas aeruginosa and Bacillus subtilis in three vegetables, Braz. J. Microbiol. 39 (2008) 423–426.
[11] M.Saraf Tank, Enhancement of plant growth and decontamination of nickel-spiked soil using PGPR, J. Basic Microbiol. 49 (2009) 195–204.
[12] C.P. Gupta, A. Sharma, R.C. Dubey, D.K. Maheshwari, Pseudomonas aeruginosaas a strong antagonist of Macrophomina phaseolina and Fusarium oxysporum,Cytobios 99 (1999) 183–189.
[13] M. Hofte, K.Y. Seong, E. Turkeyviteh, W. Verstraete, Pyoverdin production bythe plant growth beneficial Pseudomonas strains 7NSK2-ecological signifi-cance in soil, Plant Soil 130 (1991) 249–257.
[14] J.G. Holt, N.R. Krieg, P.H.A. Sneath, J.T. Staley, S.T. Williams (Eds.), Bergey’sManual of Determinative Bacteriology, Williams and Wilkins Press, Baltimore,
USA, 1994.[15] K.K. Joshi, V. Kumar, R.C. Dubey, D.K. Maheshwari, Effect of chemical fertilizer
adaptive variants, Pseudomonas aeruginosa GRC2 and Azotobacter chroococcumAC1 on Macrophomina phaseolina causing charcoal rot of Brassica juncea,Korean J. Environ. Agric. 25 (2006) 228–235.
[16] A.R.S. Kartha, A.S. Sethi, A cold percolation method for rapid gravimetricestimation of oil in small quantities of oilseeds, Ind. J. Agric. Sci. 27 (1957)211–217.
[18] A. Kumar, N. Vij, G.S. Randhawa, Isolation and symbiotic characterization of transposon Tn5-induced arginine auxotrophs of Sinorhizobium meliloti, Ind.
J. Exp. Biol. 41 (2003) 1198–1204.[19] B. Kumar, R.C. Dubey, D.K. Maheshwari, Biocontrol of Macrophomina
phaseolina: prospects and constraints, in: T. Satyanarayana, B.N. Johri (Eds.),Microbial Diversity: Current Perspectives and Potential Applications, IKInternational Pvt Ltd, New Delhi, 2005, pp. 471–492.
[20] V. Kumar, R.K. Behl, N. Nurula, Establishment of phosphate solubilizing strainsof Azotobacter chroococcum in rhizosphere and their effect on wheat cultivarsunder green house conditions, Microbiol. Res. 156 (2001) 87–93.
[21] P.W. Lanje, A.N. Buldeo, S.R. Zade, V.G. Gulhane, The effect of Rhizobium andphosphorous solubilizers on nodulation, dry matter, seed protein, oil and yieldof soybean, J. Oilseeds Res. 15 (2005) 132–135.
[22] Luc M. Maene, 10th World Food Prize Congress, Washington DC, 2000, pp.169–171.
[23] R.L. Miller, B.B. Higgins, Association of cyanide with infection of birds foottrefoil by Stemphylium loti, Phytopathology 60 (1970) 104–110.
[24] M.D. Mohiuddin, A.K. Das, D.C. Ghosh, Growth and productivity of wheat asinfluenced by integrated use of chemical fertilizer, biofertilizers and growthregulator, Ind. J. Plant Physiol. 5 (2000) 334–338.
[25] C.S. Nautiyal, J.K. Johri, H.B. Singh, Survival of rhizosphere-competentbiocontrol strain Pseudomonas fluorescens NBRI2650 in the soil and phyto-sphere, Can. J. Microbiol. 48 (2002) 588–601.
[26] C.S. Nautiyal, in: R.K. Upadhyay, K.G. Mukherji, B.P. Chamola (Eds.), BiocontrolPotential and its Exploitation in Sustainable Agriculture, Kluwer Academic/Plenum Publishers, New York, 2000, pp. 9–23.
[27] P. Pandey, S.C. Kang, C.P. Gupta, D.K. Maheshwari, Rhizosphere competentPseudomonas aeruginosa GRC1 produces characteristic siderophore andenhance growth of Indian mustard (Brassica campestris), Curr. Microbiol. 51(2005) 303–309.
[28] K.I. Poul, K.E. Savithri, Effect of biofertilizers vs. perfected chemical fertiliza-tion for sesame grown in summer rice fallow, J. Trop. Agric. 41 (2003) 47–49.
[29] B.N. Reddy, S.N. Sudhakarababu, Production potential and utilization andeconomics of fertilizer management in summer sunflower based crop, Ind.
J. Agric. Sci. 66 (1996) 16–19.
[30] P.M. Salimath, P.N. Bahl, Association analysis and plant ideotype in chick pea(Cicer arietinum L.), Exp. Gen. 2 (1986) 41–46.[31] F.M. Scher, J.W. Kloepper, C. Singleton, I. Zaleska, M. Laliberte, Colonization of
soybean roots by Pseudomonas and Serratia spp. relationship to bacterialmotility, chemotaxis and generation time, Phytopathology 78 (1988)1055–1059.
[32] B. Schwyn, J.B. Neilands, Universal assay for the detection and determinationof siderophores, Anal. Biochem. 160 (1987) 47–56.
[33] G. Selvaraj, V.N. Iyer, Suicide plasmid vehicles for insertion mutagenesis inRhizobium meliloti and related bacteria, J. Bacteriol. 156 (1983) 1292.
[34] R. Seong, M. Hofte, W. Verstraete, Acclimatization of plant growth promotingPseudomonas strain 7NSK2 in soil: effect on population dynamics and plantgrowth, Soil Biol. Biochem. 24 (1992) 750–759.
[35] R.D. Sharma, Judicious use of phosphatic fertilizers in crop production, Ind.Far. Dig. 27 (1994) 36–39.
[37] F. Stephan, E.V. Nybe, Organic manure and biofertilizers on nutrient
availability and yield in black pepper, J. Trop. Agric. 41 (2003) 52–55.[38] N.S. Subba-Rao, Advances in agricultural microbiology, in: N.S. Subba-Rao
(Ed.), Studies in the Agriculture and Food Sciences, Butterworth Scientific,London, 1982, pp. 295–303.
[39] M.A.T. Vargas, I.C. Mandes, M. Hungaria, Response of field-grown been (Pha-seolus vulgaris L.) to Rhizobium inoculation and nitrogen fertilization in twocarrados soils, Biol. Fert. Soil 33 (2000) 228–233.
[40] D.M. Weller, Biological control of soil borne plant pathogens in therhizosphere with bacteria, Ann. Phytopathol. 26 (1998) 379–407.
[41] D.M. Weller, R.J. Cook, Suppression of take all the wheat by seedtreatment with fluorescent pseudomonads, Phytopathology 23 (1983)23–54.
[42] J.M. Whipps, Microbial interactions and biocontrol in the rhizosphere, J. Exp.Bot. 52 (2001) 487–511.
[43] S.C. Wu, Z.H. Cao, Z.G. Li, K.C. Cheung, M.H. Wong, Effects of biofertilizercontaining N-fixer, P and K solubilizers and AM fungi on maize growth:a greenhouse trial, Geoderma 125 (2005) 155–166.
Table 6
Interactive effect of Biofertilizers and Integrated Nutrient Management Protocol on the yield and yield component of S. indicum L. after 120 DAS.
CD at 1% 1.459 6.539 6.183 0.365 0.500 0.685 4.729 5.654 5.626 4.773
CD at 5% 1.041 4.666 4.412 0.260 0.357 0.489 3.374 4.034 4.015 3.406
Data aremean of two years. Values of each yearare mean of 10randomlyselected plants from each set; ns,notsignificant; *significantat 5% LSD; **significantat 1% levelof LSD
as compared to control; full dose of chemical fertilizers N40þ40þ40, P30, K 30; half doses of chemical fertilizers N20þ20þ20, P15, K15.
S. Kumar et al. / European Journal of Soil Biology 45 (2009) 334–340340