ARTICLE Regional-scale patterns in community concordance: testing the roles of historical biogeography versus contemporary abiotic controls in determining stream community composition Margaret R. Neff and Donald A. Jackson Abstract: The determinants of a local-scale biological community can include both abiotic and biotic factors acting at a variety of spatial and temporal scales. Previous studies have shown that the Precambrian Shield, a broad-scale geological feature, has distinct abiotic conditions and aquatic communities in lotic systems as compared with systems off-Shield. The relative importance of historical biogeography and postglacial colonization in Ontario versus the role of contemporary abiotic conditions has been debated in determining fish community composition. In this study, we sampled fish and macroinvertebrate community data from south-central Ontario streams to examine the degree of community concordance between fishes and macroinvertebrates and to determine the main abiotic drivers for this biological distinction between Shield and off-Shield areas. This study shows that low-order lotic systems on the Shield differ both abiotically and biotically from adjacent off-Shield systems and that similar abiotic drivers are responsible for structuring both groups of taxa. However, water chemistry and land-use variables are the most important in structuring macroinver- tebrate communities, whereas physical habitat plays a larger role for fish communities. Résumé : Les déterminants d’une communauté biologique locale peuvent comprendre des facteurs tant abiotiques que bi- otiques qui opèrent a ` diverses échelles spatiales et temporelles. Des études antérieures ont démontré que les systèmes lotiques du Bouclier précambrien, une vaste entité géologique d’âge précambrien, sont caractérisés par des conditions abiotiques et des communautés aquatiques différentes de celles des systèmes situés a ` l’extérieur du Bouclier. Il existe un débat quant a ` l’importance relative de la biogéographie historique et de la colonisation postglaciaire en Ontario par rapport au rôle des conditions abiotiques contemporaines dans la détermination de la composition des communautés de poissons. Dans la présente étude, nous avons recueilli des données sur les communautés de poissons et de macroinvertébrés de cours d’eau du centre-sud de l’Ontario afin d’examiner le degré de concordance entre les communautés de poissons et celles de macroinvertébrés et de cerner les principaux facteurs abiotiques a ` l’origine de cette différence biologique entre les régions internes et externes au Bouclier. L’étude démontre que les systèmes lotiques d’ordre inférieur dans le Bouclier se distinguent sur le plan tant abiotique que biotique des systèmes avoisinants externes au Bouclier et que des facteurs abiotiques semblables déterminent la structure des deux groups de taxons. Cela dit, des variables associées a ` la chimie de l’eau et a ` l’utilisation du sol jouent un rôle particulièrement important dans la structuration des communautés de macroinvertébrés, alors que l’habitat physique joue un rôle plus important en ce qui concerne les communautés de poissons. [Traduit par la Rédaction] Introduction Regional-scale environmental factors often play a large role in determining local species assemblage patterns in aquatic systems (e.g., Heino et al. 2003; Wang et al. 2003; Mykrä et al. 2007). Al- though a number of abiotic and biotic variables can influence lotic community composition and structure, the relative impor- tance of certain variables can vary for different groups of taxa. Examining community patterns between different groups of taxa — i.e., community concordance — can provide a better un- derstanding of the effects of broad-scale environmental features on aquatic assemblages. Previous studies suggest that community concordance between different groups of aquatic taxa does exist, although the results vary depending on spatial and temporal scale, as well as the abiotic factors in question (Jackson and Harvey 1993; Kilgour and Barton 1999; Bowman et al. 2008). Ultimately, this type of analysis allows us to broaden the scope of interpreta- tion and generality of ecological understanding. The geological composition of a region is one broad-scale factor that can greatly influence the physical and chemical attributes of lakes, streams, and rivers and thus have an important impact on local species assemblages (e.g., Neff and Jackson 2011, 2012). In North America, the Precambrian Shield is a dominant surficial geological feature encompassing much of Canada and parts of the northern United States in the Midwest and New England. The Shield, as it is familiarly known, is characterized by ancient met- amorphic and igneous bedrock lying at or close to the surface, having been exposed during the glaciation in North America (Chapman and Putnam 1984). In areas near the Shield boundary, such as south-central Ontario, the Shield landscape stands in sharp contrast to the younger, primarily sedimentary bedrock of southern Ontario and most of the remaining area of North America. Differences in the weathering characteristics of these two geo- logical types create distinct physical and chemical attributes in aquatic systems; for example, slow weathering rates of Shield bedrock result in poor buffering capacity of Shield waters, characterized by low conductivities, a propensity for acidification, and calcium depletion (see Edwards et al. 2009; Jeziorski et al. 2008). Proximity of the bedrock to the surface may also affect the Received 16 November 2012. Accepted 26 May 2013. Paper handled by Associate Editor Cliff Kraft. M.R. Neff. Department of Ecology and Evolutionary Biology, University of Toronto, 25 Willcocks Street, Toronto, ON M5S 3B2, Canada. D.A. Jackson. Department of Ecology and Evolutionary Biology, University of Toronto, 25 Willcocks Street, Toronto, ON M5S 3B2, Canada; School of the Environment, University of Toronto, 33 Willcocks Street, Toronto, ON M5S 3E8, Canada. Corresponding author: Margaret R. Neff (e-mail: [email protected]and [email protected]). 1141 Can. J. Fish. Aquat. Sci. 70: 1141–1150 (2013) dx.doi.org/10.1139/cjfas-2012-0497 Published at www.nrcresearchpress.com/cjfas on 29 May 2013. Can. J. Fish. Aquat. Sci. Downloaded from www.nrcresearchpress.com by UNIVERSITY OF WYOMING on 09/22/13 For personal use only.
Transcript
ARTICLE
Regional-scale patterns in community concordance: testing the roles ofhistorical biogeography versus contemporary abiotic controls indetermining stream community compositionMargaret R. Neff and Donald A. Jackson
Abstract: The determinants of a local-scale biological community can include both abiotic and biotic factors acting at a variety ofspatial and temporal scales. Previous studies have shown that the Precambrian Shield, a broad-scale geological feature, has distinctabiotic conditionsandaquatic communities in lotic systemsas comparedwith systemsoff-Shield. The relative importanceofhistoricalbiogeography and postglacial colonization in Ontario versus the role of contemporary abiotic conditions has been debated indetermining fish community composition. In this study, we sampled fish andmacroinvertebrate community data from south-centralOntario streams to examine the degree of community concordance between fishes and macroinvertebrates and to determine themain abiotic drivers for this biological distinction between Shield and off-Shield areas. This study shows that low-order lotic systemson the Shield differ both abiotically and biotically fromadjacent off-Shield systems and that similar abiotic drivers are responsible forstructuring both groups of taxa. However, water chemistry and land-use variables are themost important in structuringmacroinver-tebrate communities, whereas physical habitat plays a larger role for fish communities.
Résumé : Les déterminants d’une communauté biologique locale peuvent comprendre des facteurs tant abiotiques que bi-otiques qui opèrent a diverses échelles spatiales et temporelles. Des études antérieures ont démontré que les systèmes lotiquesdu Bouclier précambrien, une vaste entité géologique d’âge précambrien, sont caractérisés par des conditions abiotiques et descommunautés aquatiques différentes de celles des systèmes situés a l’extérieur du Bouclier. Il existe un débat quant al’importance relative de la biogéographie historique et de la colonisation postglaciaire en Ontario par rapport au rôle desconditions abiotiques contemporaines dans la détermination de la composition des communautés de poissons. Dans la présenteétude, nous avons recueilli des données sur les communautés de poissons et de macroinvertébrés de cours d’eau du centre-sudde l’Ontario afin d’examiner le degré de concordance entre les communautés de poissons et celles de macroinvertébrés et decerner les principaux facteurs abiotiques a l’origine de cette différence biologique entre les régions internes et externes auBouclier. L’étude démontre que les systèmes lotiques d’ordre inférieur dans le Bouclier se distinguent sur le plan tant abiotiqueque biotique des systèmes avoisinants externes au Bouclier et que des facteurs abiotiques semblables déterminent la structuredes deux groups de taxons. Cela dit, des variables associées a la chimie de l’eau et a l’utilisation du sol jouent un rôleparticulièrement important dans la structuration des communautés de macroinvertébrés, alors que l’habitat physique joue unrôle plus important en ce qui concerne les communautés de poissons. [Traduit par la Rédaction]
IntroductionRegional-scale environmental factors often play a large role in
determining local species assemblage patterns in aquatic systems(e.g., Heino et al. 2003; Wang et al. 2003; Mykrä et al. 2007). Al-though a number of abiotic and biotic variables can influencelotic community composition and structure, the relative impor-tance of certain variables can vary for different groups of taxa.Examining community patterns between different groups oftaxa — i.e., community concordance — can provide a better un-derstanding of the effects of broad-scale environmental featureson aquatic assemblages. Previous studies suggest that communityconcordance between different groups of aquatic taxa does exist,although the results vary depending on spatial and temporalscale, as well as the abiotic factors in question (Jackson andHarvey1993; Kilgour and Barton 1999; Bowman et al. 2008). Ultimately,this type of analysis allows us to broaden the scope of interpreta-tion and generality of ecological understanding.
The geological composition of a region is one broad-scale factorthat can greatly influence the physical and chemical attributes of
lakes, streams, and rivers and thus have an important impact onlocal species assemblages (e.g., Neff and Jackson 2011, 2012). InNorth America, the Precambrian Shield is a dominant surficialgeological feature encompassingmuch of Canada and parts of thenorthern United States in the Midwest and New England. TheShield, as it is familiarly known, is characterized by ancient met-amorphic and igneous bedrock lying at or close to the surface,having been exposed during the glaciation in North America(Chapman and Putnam 1984). In areas near the Shield boundary,such as south-central Ontario, the Shield landscape stands insharp contrast to the younger, primarily sedimentary bedrock ofsouthern Ontario andmost of the remaining area of North America.Differences in the weathering characteristics of these two geo-logical types create distinct physical and chemical attributes inaquatic systems; for example, slow weathering rates of Shieldbedrock result in poor buffering capacity of Shield waters,characterized by low conductivities, a propensity for acidification,and calcium depletion (see Edwards et al. 2009; Jeziorski et al.2008). Proximity of the bedrock to the surface may also affect the
Received 16 November 2012. Accepted 26 May 2013.
Paper handled by Associate Editor Cliff Kraft.
M.R. Neff. Department of Ecology and Evolutionary Biology, University of Toronto, 25 Willcocks Street, Toronto, ON M5S 3B2, Canada.D.A. Jackson. Department of Ecology and Evolutionary Biology, University of Toronto, 25 Willcocks Street, Toronto, ON M5S 3B2, Canada; School of the Environment, University of Toronto,33 Willcocks Street, Toronto, ON M5S 3E8, Canada.
physical structure of aquatic systems, particularly streams andrivers, where erosional resistance of the underlying bedrock mayresult in poor pool development (Wohl and Legleiter 2003). Neffand Jackson (2011, 2012) showed that, at broad spatial scales, loticsystems on the Precambrian Shield not only exhibit distinct phys-ical and chemical characteristics, but also have distinct fish andmacroinvertebrate communities compared with streams of simi-lar size found in adjacent off-Shield sites in south-central Ontario.
There are other broad-scale environmental factors at play in deter-mining aquatic assemblages in this region, beyond the physical andchemical factors. For example, Jackson and Harvey (1989) showedthat lake fish communities from different regions of Ontario dem-onstrated strong relationships to the abiotic conditions aswell as theinfluence of historical effects. Mandrak (1995) noted the strong influ-ence of historical postglacial colonization routes into the province asbeing critical in determining these broad-scale patterns. This histor-ical influence assumes that species' current distributions are mostinfluenced by dispersal barriers arising following the last glaciation,when only cold-water species were able to colonize waters at theedge of the melting ice sheets (Hinch et al. 1991; Mandrak 1995). Bythe time these waters in the region warmed to a suitable degree forwarm-water fishes, aquatic pathways to upland areas of the provincehad been disconnected owing to isostatic rebound of the landscapeand subsequent formation ofmodern-daywatersheds.Many of theseboundaries to watersheds and species distributions coincide withthe Shield versus off-Shield boundary. Thus, both historical and cur-rent dispersal limitations have been considered to play significantroles in determiningwhich fish species are foundwhere throughoutthe province, and observations concerning the influence of the Pre-cambrian Shield on fish communities could be confounded withthese dispersal limitations.
Herewecapitalizedon inherentdifferences in lifehistorybetweentwo groups of aquatic taxa to test these competing hypotheses re-garding the role of (i) broad-scale environmental factors associatedwith Shield conditions (e.g., low pH, conductivity) in structuringaquatic communities versus (ii) historical factors influencingpostgla-cial dispersal and contemporary species distributions. A major eco-logical difference between fishes and benthic macroinvertebrateslies in their dispersal abilities. Manymacroinvertebrate taxa have anaerial lifehistory stage,wherebyadults emerge fromaquatic systemswith the ability to fly overland to new locations (Bilton et al. 2001;Bohonak and Jenkins 2003). In contrast, fish species are naturallylimited to aquatic pathways for their dispersal and can face barrierssuch as waterfalls and dams. Therefore, current fish distributionsmay be a reflection of historical limits to dispersal rather than localenvironmental conditions. However, if macroinvertebrate commu-nities — which should not show the same dispersal limitations thatrequire aquatic connections — also show distinct assemblages be-tween Shield and off-Shield systems, it supports the hypothesis thatthe abiotic conditions imposed in Shield lotic systems are more im-portant factors affecting the community composition of both fishand macroinvertebrate communities.
Based on previous studies examining the broad-scale communitycomposition of both fishes and macroinvertebrates in this region,which indicated that similar abiotioc variables were important instructuring both fish and macroinvertebrate communities in loticsystems (Neff and Jackson 2011, 2012), as well as other studies thatexamined community concordance in aquatic taxa (e.g., Jackson andHarvey 1993; Kilgour andBarton 1999; Paavola et al. 2006),wepredictthat fish and macroinvertebrate communities of Shield and off-Shield systemswill be influencedby similar abiotic in-streamfactors,and thus will show a strong degree of concordance in sites betweenthe two taxanomic groups. Significant community concordance be-tween these two groups of taxa will support the hypothesis that
broad-scale patterns of south-central Ontario fish communities are,at least in part, structured by abiotic conditions imposed by thePrecambrian Shield and are not merely an artifact of historical pro-cesses following deglaciation.
MethodsIn the summer of 2007 and 2008, first- to third-order stream sys-
tems located on- or off-Shield were sampled to assess fish and mac-roinvertebrate community composition, as well as physical andchemical attributes. Sampling was conducted within a limited geo-graphic range extending from �44.3°N to 45.9°N and �78.5°W to79.5°W in south-central Ontario (Fig. 1) to minimize the climaticdifferences among sites. To select sites, systemswithin this area thatmet the following criteria were considered for sampling: (i) betweenfirst and third stream order, inclusively, (ii) relatively unimpacted byanthropogenic influences, and (iii) on land locally accessible fromroads or pathways. Sites were classified as “Shield” or “off-Shield”using a GIS layer of bedrock geology (Ontario Geological Survey2003). Some sites classified as “Shield” using the GIS data had unusu-ally high conductance based on field sampling of water chemistryand (or)were very close to off-Shield geology,where therewas poten-tial for the true site location to be off-Shield, or on a combination ofShield and off-Shield geology. These siteswere tentatively deemed tobe “transition” sites and were examined more closely in the subse-quent analyses. In July–August 2007, 35 sites were sampled(34 Shield, 1 transition site), and in June–August 2008, 16 off-Shieldsites, 4 transition sites, and an additional 7 Shield sites were sam-pled, resulting in a total of 64 sites. Owing to absence of fishes orwater chemistry meter equipment malfunctions, 7 sites were re-moved from the dataset, resulting in 36 Shield, 5 transition, and16 off-Shield sites. A list of all sites and their associated location,classification, sampling year, and surficial bedrock geology can befound in the Supplementary Data (Table S11).
Abiotic samplingEach site was assessed for a set of physical and chemical param-
eters. Site length, which was first determined using a standard
1Supplementary data are available with the article through the journal Web site at http://nrcresearchpress.com/doi/suppl/10.1139/cjfas-2012-0497.
Fig. 1. Map of 2007–2008 sampling sites in south-central Ontario,including 36 Shield sites (black circles), 5 transition sites (greycircles), and 16 off-Shield sites (white circles). The dashed lineindicates the Shield boundary in this part of the province.
method for both Shield and off-Shield systems, was 10 times themean wetted width of the stream. As it was hypothesized thatShield systems, as a consequence of their hard, exposed bedrock,may not adhere to the same pattern in meander wavelengths asoff-Shield systems, this method allowed standardization acrosssites while still accounting for minor differences in stream size.Thus, this method incorporates the idea that as stream size in-creases, the effort required to adequately sample that systemshould increase proportionally and the use of a fixed site lengthcould result in under-representation of certain habitat features inlarger systems (Flotemersch et al. 2011). The relationships betweensite area (m2) and species or taxa richness are displayed in theSupplementary Data (Fig. S11) and did not differ from random foreither fish (linear regression, R2 = 0.007, df = 56, p = 0.57) ormacroinvertebrates (linear regression, R2 = 0.0008, df = 56, p = 0.83).
Dissolved oxygen (DO), pH, conductivity (Cond), and water tem-perature (T) were measured three times within the samplingreach and then averaged. Physical variables measured includedpercent canopy cover (Can), stream gradient (G), mean water ve-locity (V), and depth. Substrate composition (%) and abundance ofvegetation and woody debris for each site were determinedthrough a modified version of the methodology outlined in theOntario Stream Assessment Protocol (Stanfield 2005). Along eachtransect, substrate composition was assessed by taking a randomsample as well as an overall visual assessment at three, five, or sixpoints, depending on the length of the transect, as dictated by theOntario Stream Assessment Protocol (Stanfield 2005). The pres-ence of moss (M), woody debris (WD), algae (AL), and aquatic mac-rophytes (WV) was also assessed at each point along the transect.Presence or absence of these features at each point along thetransect was then used to calculate a proportional measure ofabundance; for example, if moss was found at five point measure-ments along a transect with six points, it received a value of 0.83for that transect.
As substrate composition recorded at each site represents com-positional variables that are not independent of each other andlead to biased assessment and interpretation using correlation-based methods (Jackson 1997), correspondence analysis (CA) wasused to summarize the variation across the different substratecategories into axes summarizing the dominant trends. The re-sulting site scores for the first three axes were used as new vari-ables, resulting in three substrate variables, S1, S2, and S3. Lastly,land-use data for the upstream catchment area for each site werecollected using the Aquatic Landscape Inventory Software (ALIS2008). Percent upstream catchment area of 28 land-use categorieswas assessed for each sampling site, and then CA was used tosummarize the variation across the difference categories in thesame manner as the substrate variables, resulting in two sum-mary land-use variables, ALIS1 and ALIS2. A summary on the in-terpretation of substrate and land-use variables is provided inTable 1. Mean values of all abiotic variables for Shield, off-Shield,
and transition sites are presented in the Supplementary Data (Ta-ble S21).
Variation in climatic attributes can impact biotic assemblagesin broad regional studies such as this one. Although this aspect ofthe abiotic environment was not explicitly assessed, care wastaken to minimize climatic variation within the study region, asthe off-Shield to Shield transition lies along a latitudinal gradient.An examination of mean yearly temperatures for the geographicrange of this study revealed some latitudinal variation inmean airtemperature (�1 °C; Environment Canada 2011). However, longi-tudinal variation, likely due to the influence of Lake Huron, wasalso observed, and thus any differences in climate among sam-pling sites is due tomultiple factors and is not restricted to the on-or off-Shield difference.
Biotic samplingFish community composition was sampled at each site using a
single pass with a backpack electrofisher. Block nets were set upat both ends of the site to prevent escapement during sampling.Fishes were identified on site and returned to the stream afterprocessing; reference samples were taken to confirm identifica-tions in the lab using taxonomic keys in Hubbs and Lagler (2004).Mean abundances of each species for Shield, transition, and off-Shield sites are presented in the Supplementary Data (Table S31).
Macroinvertebrate community composition was sampled ateach site with two kick-net samples from both a riffle and poolhabitat. If there was no distinct riffle or pool, one sample wastaken from a faster section of the site and the other from a slowersection. Sampling was done along a transect starting from thewater's edge and out into the centre of the stream for 1 min.Samples were preserved in the field in 75% ethanol and brought tothe lab for sorting and identification to family. Identifications tofamily were conducted in-lab using various taxonomic keys, in-cluding Clarke (1981), Peckarsky et al. (1990), Thorp and Covich(1991), and Merritt and Cummins (1996). Pool and riffle sampleswere pooled for each site prior to data analysis. Mean abundancesof each taxon for Shield, transition, and off-Shield sites are pre-sented in the Supplementary Data (Table S41).
Fish and macroinvertebrate sampling protocols were adaptedfrom the Ontario Benthos Biomonitoring Network (Jones et al.2004) and Aquatic Habitat Inventory Survey (AHIS 1987), both ofwhich were developed for sampling lotic systems in this region.
Statistical analysisPatterns in variation among sampling sites were first explored
using principal component analysis (PCA; abiotic data) and CA(biotic data). Abiotic data were standardized to amean of zero andunit variance, and biotic abundance data were log-transformedprior to analysis.
Canonical correspondence analysis (CCA) on log-transformedabundance data was used to examine general patterns in fishcommunity composition and in macroinvertebrate community
Table 1. Interpretation of compositional abiotic variables, describing how values for each new correspondenceanalysis (CA)-derived substrate (S1, S2, and S3) and land-use variables (ALIS1, ALIS2) relate to the original data.
Land use ALIS1 Positive Treed bog, unknown, settlement, water, recent cutoversNegative Cropland, pasture, conifer swamp
ALIS2 Positive Wetland, alvar, or mine tailingsNegative Dense deciduous, recent cutover, treed bog
Neff and Jackson 1143
Published by NRC Research Press
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F W
YO
MIN
G o
n 09
/22/
13Fo
r pe
rson
al u
se o
nly.
composition for Shield and off-Shield sites in relation to a singlepredictor variable indicating a site's location on- or off-Shield. Apermutation test (n = 1000) on the single constrained axis was usedto test whether Shield and off-Shield sites significantly differed intheir community composition. Fish and macroinvertebrate com-munity datasets were then individually constrained by abioticvariables using CCA to determine the most important factors ex-plaining the two community patterns in these sites. Partial CCAwas used to examine the relative influence of land-use, waterchemistry, and physical habitat variables on the overall explainedvariation.
To assess community concordance, Procrustes analysis wasused on the fish and macroinvertebrate CA results. This methodrotates and (or) reflects and translates an ordination result toproduce the maximum similarity with another ordination result(Jackson 1995; Peres-Neto and Jackson 2001). The overall measureof concordance is the m2 statistic, which provides an integratedleast-squares measurement of the deviations between the twoordinations. Significance of the m2 statistic was then assessed us-ing the permutation-based PROTEST function (n = 1000), and anal-ysis of the differences in the magnitude of Procrustes residualsbetween Shield and off-Shield sites was performed using Mann–Whitney's U test.
All analyses were performed using R (R Development CoreTeam 2010).
Results
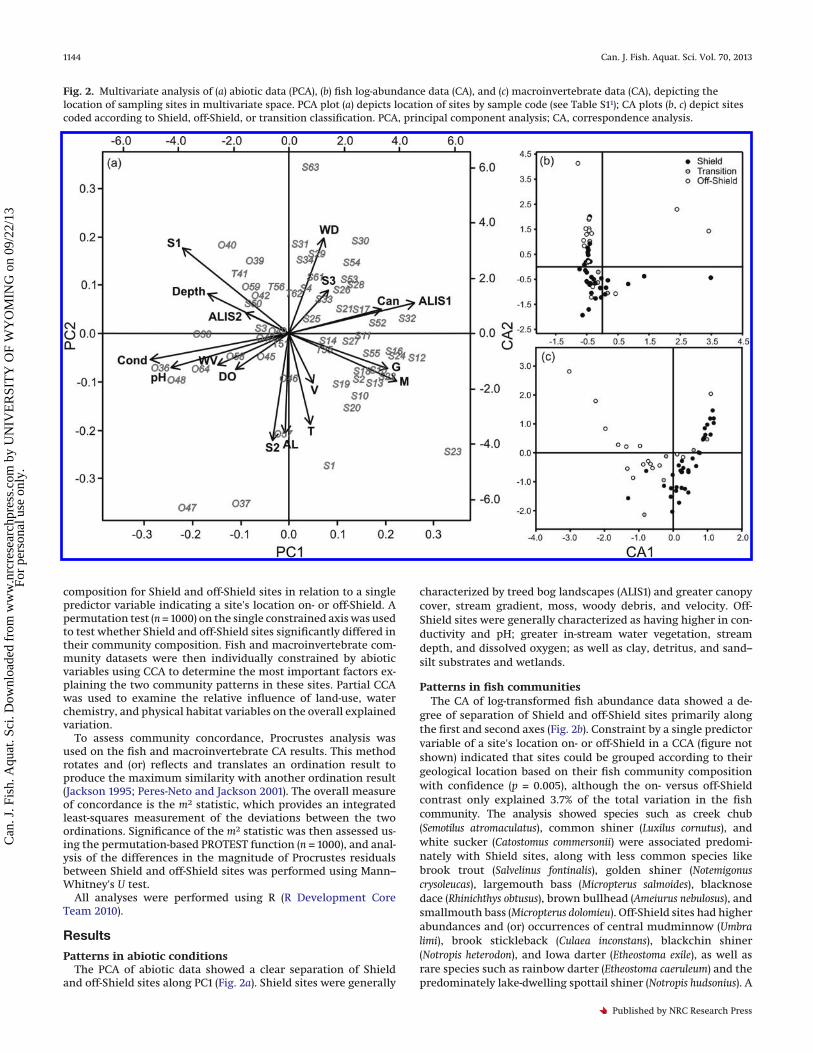
Patterns in abiotic conditionsThe PCA of abiotic data showed a clear separation of Shield
and off-Shield sites along PC1 (Fig. 2a). Shield sites were generally
characterized by treed bog landscapes (ALIS1) and greater canopycover, stream gradient, moss, woody debris, and velocity. Off-Shield sites were generally characterized as having higher in con-ductivity and pH; greater in-stream water vegetation, streamdepth, and dissolved oxygen; as well as clay, detritus, and sand–silt substrates and wetlands.
Patterns in fish communitiesThe CA of log-transformed fish abundance data showed a de-
gree of separation of Shield and off-Shield sites primarily alongthe first and second axes (Fig. 2b). Constraint by a single predictorvariable of a site's location on- or off-Shield in a CCA (figure notshown) indicated that sites could be grouped according to theirgeological location based on their fish community compositionwith confidence (p = 0.005), although the on- versus off-Shieldcontrast only explained 3.7% of the total variation in the fishcommunity. The analysis showed species such as creek chub(Semotilus atromaculatus), common shiner (Luxilus cornutus), andwhite sucker (Catostomus commersonii) were associated predomi-nately with Shield sites, along with less common species likebrook trout (Salvelinus fontinalis), golden shiner (Notemigonuscrysoleucas), largemouth bass (Micropterus salmoides), blacknosedace (Rhinichthys obtusus), brown bullhead (Ameiurus nebulosus), andsmallmouth bass (Micropterus dolomieu). Off-Shield sites had higherabundances and (or) occurrences of central mudminnow (Umbralimi), brook stickleback (Culaea inconstans), blackchin shiner(Notropis heterodon), and Iowa darter (Etheostoma exile), as well asrare species such as rainbow darter (Etheostoma caeruleum) and thepredominately lake-dwelling spottail shiner (Notropis hudsonius). A
Fig. 2. Multivariate analysis of (a) abiotic data (PCA), (b) fish log-abundance data (CA), and (c) macroinvertebrate data (CA), depicting thelocation of sampling sites in multivariate space. PCA plot (a) depicts location of sites by sample code (see Table S11); CA plots (b, c) depict sitescoded according to Shield, off-Shield, or transition classification. PCA, principal component analysis; CA, correspondence analysis.
rendering of the CA plot with species scores is available in theSupplementary Data (Fig. S21).
Patterns in macroinvertebrate communitiesThe CA of macroinvertebrate log-transformed abundance data
showed a clear separation of Shield and off-Shield sites using thefirst two axes (Fig. 2c). Constraint by a single predictor variable ofa site's location on- or off-Shield in a CCA indicated that sites couldbe grouped according to their geological location based on theirmacroinvertebrate community composition with confidence(p < 0.001), with the constrained axis explaining 8.1% of the totalvariation. The analysis showed a number of taxa that were locatednear the origin of the plot, such as Chironomidae, Simuliidaeand Tipulidae (Diptera), Hydropsychidae (Trichoptera), Elmidae(Coleoptera), Oligeochaeta, and Bivalvia, indicating that thesetaxa may be widely distributed and common in both groups ofsites, and this was confirmed through examination of the dataset.Shield sites were associated with taxa such as Aeshnidae, Cordule-gastridae, and Gomphidae (Odonata); Baetiscidae, Ephemerellidae,Heptegeniidae, and Leptophlebidae (Ephemeroptera); Glossoso-matidae, Leptoceridae, Philopotamidae, Odontoceridae, Polycen-tropidae, andRhyacophilidae (Trichoptera); Sialidae andCorydalidae(Megaloptera); and Leuctridae, Nemouridae, and Perlidae (Plecop-tera). Off-Shield sites were associated with increased prevalenceand abundance of Amphipoda, Asellidae (Isopoda), Baetidae andCaenidae (Ephemeroptera), Corixidae (Hemiptera), Haliplidae,Gyrinidae, Hydrophilidae and Dytiscidae (Coleoptera), Libelluli-dae and Lestidae (Odonata), Sciomyzidae and Tabanidae (Diptera),Hirudinea, and Lymnaeidae, Physidae, Planoboridae and Val-vatidae (Gastropoda). A rendering of the CA plot with taxa scoresis available in the Supplementary Data (Fig. S21).
Abiotic and community associationsShield and off-Shield fish communities were generally sepa-
rated using the first two CCA axes (Fig. 3a), where off-Shield com-munities tended to be associated with clay and detrital substrates(S1), higher conductivity, aquatic plants, pH, depth and, to a lesserdegree, wetlands and (or) alvar landscapes (ALIS2). Shield fishcommunities were primarily associated with increased density ofcanopy cover, treed bogs, unknown or unclassified land uses, set-tlement, water and (or) recent cutover land use (ALIS1), steepergradient, higher dissolved oxygen, in-stream moss, and instreamwoody debris.
Compared with the fish community analysis, sites were moreclearly separated in a plot of the first two CCA axes of macroin-vertebrate community composition constrained by abiotic vari-ables (Fig. 4a). Off-Shield macroinvertebrate communities wereprimarily associated with high conductivity, pH, small-sizedsubstrates (clay, detritus, sand–silt), bedrock and clay substrates,algae, wetlands and (or) alvars (ALIS2), and depth. Shield commu-nities were associated with land-use variable ALIS1 (greater treedbogs, settlements, water bodies, and (or) recent cutovers), woodydebris, higher gradient, in-stream moss, water velocity, andtemperature.
Partial CCA was used to assess the relative contribution of land-use, water chemistry, and physical habitat variables to the overallexplained variation in the constrained analyses. CCA summarized40.3% of the variation in fish communities, with 4% attributeduniquely to water chemistry variables (pH and conductivity), 3.2%to land-use variables (ALIS1 and ALIS2), 29.9% to the remainingphysical habitat variables, and 3.2% to shared variation. For mac-roinvertebrates, CCA summarized 43.3% of the total variation inmacroinvertebrate communities, with 3.5% attributed uniquely towater chemistry variables, 2.6% to land-use variables, 29.7% to theremaining physical habitat variables, and 7.5% to shared varia-tion. Plots of the various partial CCAs (i.e., water chemistryremoved, land-use removed, and physical habitat variation re-moved) revealed that the Shield – off-Shield separation of sites in
the CCA plot could be attributed to variation in all three types ofabiotic factors for fish communities (Figs. 3b, 3c and 3d). However,for macroinvertebrate communities, the distinction betweenShield and off-Shield points was removed when either land-use orwater chemistry variables were partialled out (Figs. 4b and 4c,respectively) and preserved when only physical variables are par-tialled out (Fig. 4d).
Community concordance and abiotic associationsProcrustes analysis between CA ordinations of fish and macro-
invertebrate abundances showed that the two ordinations aresignificantly associated (m2 = 0.771, p < 0.001), with the two com-munities showing similar patterns across the sampling locations.Analysis of residuals provided a comparison of sites based on theirfish communities relative to their invertebrate communities, andthose differing the most in the two communities had the largestresiduals, i.e., the greatest difference in site position between fishand macroinvertebrate CA results. The size of residuals for Shieldand off-Shield sites were significantly different, with off-Shieldsites having longer residuals and therefore more discordantfish and macroinvertebrate communities (Mann–Whitney U test,p = 0.02). This pattern appears to be strongly influenced by foursites — one Shield (S1) and three off-Shield (O37, O46, and O48) —which had very large residuals compared with all other sites. Mis-matches between the communities (i.e., residuals) were positivelycorrelated with pH (r = 0.39), conductivity (r = 0.26), and algae(r = 0.36), though negatively correlated with ALIS1 (r = 0.41), canopycover (r = 0.38), gradient (r = 0.29), and woody debris (r = 0.35).
Concordance between fish and macroinvertebrate communi-ties was also examined within Shield and off-Shield sites alone.Procrustes analysis between CA ordinations of fish and macroin-vertebrate abundances for Shield sites were significantly concor-dant (m2 = 0.47, p = 0.02), but not for off-Shield sites (m2 = 0.43,p = 0.1). However, the lack of significance for off-Shield sites islikely a reflection of lack of statistical power, as n = 16 for thisgroup of sites.
While not showing strong patterns with regard to Procrustesresiduals, therewere several sites that exhibited patterns in eitherabiotic or biotic conditions that were more intermediate betweena Shield and off-Shield site. One Shield site (S34) exhibited abioticconditions typical of other Shield sites and had a representativefish community, but had macroinvertebrate composition moretypical of off-Shield sites. Compared with other Shield sites, thissite was less associated with high dissolved oxygen and had agreater proportion of smaller substrates. Three off-Shield sites(O45, O49, and O57) exhibited typical abiotic characteristics andmacroinvertebrate communities compared with other off-Shieldsites (Fig. 2), but had fish communities similar to those of Shieldsites. These locations had greater canopy cover and were domi-nated by larger substrates comparedwith average values for otheroff-Shield sites.
For the sites that had been highlighted as potentially havinga transitional environment between the Shield and off-Shieldregions, all except one (T41) were abiotically similar to theShield sites, but with higher conductivity values (Fig. 2a). Therelationship between the abiotic conditions and the fish andmac-roinvertebrate communities in these sites further supports theabiotic–biotic patterns already noted. For example, T41 had off-Shield abiotic conditions with fish and macroinvertebrate com-munities at the interface of Shield and off-Shield compositionalpatterns, whereas T35 and T62 had Shield-like abiotic conditions(Fig. 2a), with Shield-like fish and macroinvertebrate communi-ties. The remaining two transition sites, T51 and T56, were alsogenerally associated with other Shield abiotic conditions (Fig. 2a),but had higher pH and conductivity. Likewise, the biotic commu-nity reflected both Shield and off-Shield attributes, with Shield-like fish communities but macroinvertebrate communities withhigher abundances of Amphipoda, Hirudinea, Oligochaeta, Plano-
Neff and Jackson 1145
Published by NRC Research Press
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F W
YO
MIN
G o
n 09
/22/
13Fo
r pe
rson
al u
se o
nly.
boridae and Viviparidae. However, low abundances of Shield-liketaxa were also present, including Coenagrionidae, Empididae, Gom-phidae, and Leuctridae.
There were also interesting patterns for three sites that arelocated on different tributaries of the same river system (S55, T56,and S63). Site S63, the most southern of these sites, is the closestsite to the Shield boundary and is abiotically Shield-like (Fig. 2a).However, it was associated with some off-Shield biota, such asIsopoda, Amphipoda, Hydracharina, and Gastropoda, as well asBrook Stickleback and Central Mudminnow. Compared with themean values for other Shield streams, it had a lower gradient anddissolved oxygen, but much higher values for substrate variablesthat include detritus (i.e., substrate variables S1 and S3). T56,which isnearby, had a similar fish and invertebrate community and was abi-otically Shield-like (Fig. 2a), but had higher conductivity than on
average. Site S55, which is the furthest from the Shield – off-Shieldinterface, exhibited Shield abiotic and biotic characteristics.
DiscussionExamination of community concordance between these two
groups of taxa has three main advantages for the purposes ofexamining the role of the Canadian Shield in structuring loticcommunities in south-central Ontario versus controls imposed byhistorical and contemporary colonization limitations. Firstly, anassessment can bemade as towhether fish andmacroinvertebratecomposition respond in the same way to the abiotic factors influ-enced by the Shield. In other words, for two sites with similar fishcommunities, do they also have similar macroinvertebrate com-munities? Secondly, if there is concordance between the two
Fig. 3. Full (a) and partial (b–d) canonical correspondence analyses (CCA) of fish community data, using abiotic variables standardized to amean of zero and unit variance. Abiotic variables include pH, conductivity (Cond), percent gradient (G), velocity (V), dissolved oxygen (DO),water temperature (T), moss (M), woody debris (WD), algae (AL), water vegetation (WV), depth, substrate (S1, S2, S3), and land use (ALIS1,ALIS2). Interpretation of compositional variables (i.e., S1, S2, S3, ALIS1, and ALIS2) can be found in Table 1. Full and partial CCA analyses wereused to determine which variables were important in distinguishing Shield and off-Shield fish communities, where (b) shows a partial CCAwith variation attributed to land-use variables removed, (c) partial CCA with water chemistry variables removed, and (d) partial CCA withremaining physical habitat variables removed.
groups of taxa, this analysis suggests that any potential limitationin dispersal of fishes is unlikely to be the main driver in distinc-tions between Shield and off-Shield systems, given that physicalbarriers (e.g., waterfalls, lack of water connections, etc.) will notimpede dispersal of invertebrates in a manner similar to the wayin which fish are impeded. Lastly, this analysis can also highlightthose sites where there might be a mismatch between fish andmacroinvertebrate communities, whether due to specific abioticconditions that are more influential to one group of taxa than theother ordue to anthropogenic impacts suchas species introductions.
Evidence from other studies has shown community concor-dance between different groups of aquatic taxa, although the re-sults vary depending on spatial and temporal scale, as well asthe abiotic factors in question (Heino 2002; Paavola et al. 2006;Bowman et al. 2008). In our study, fish and macroinvertebrate
communities were significantly concordant with each other,which is similar to what was found in Kilgour and Barton (1999),who examined communities in a more homogeneous landscape.Overall, the results from this study suggest that fish and macro-invertebrate communities are responding to similar abiotic driv-ers, further supporting the hypothesis that the Shield, via itsinfluence on various physical and chemical components of loticsystems, is an important broad-scale feature that affects lotic com-munity composition. Althoughwe did not set out to explicitly testthe relative influence of regional and local factors, this work pro-vides additional support to earlier studies showing the impor-tance of regional factors in structuring biological communities(e.g., Angermeier and Winston 1998; Heino et al. 2003; Mykrä et al.2007).
Fig. 4. Full (a) and partial (b–d) CCA of macroinvertebrate community data, using abiotic variables standardized to a mean of zero and unitvariance. Abiotic variables include pH, conductivity (Cond), percent gradient (G), velocity (V), dissolved oxygen (DO), water temperature (T),moss (M), woody debris (WD), algae (AL), water vegetation (WV), depth, substrate (S1, S2, S3), and land use (ALIS1, ALIS2). Full and partial CCAwere used to determine which variables were important in distinguishing Shield and off-Shield fish communities, where (b) shows a partialCCA with variation attributed to land-use variables removed, (c) partial CCA with water chemistry variables removed, and (d) partial CCA withremaining physical habitat variables removed.
While there was significant concordance between fish andmac-roinvertebrate communities, examination of the Procrustes resid-uals showed that as the abiotic environment moves to conditionsthat are more similar to those off-Shield, there was a greater mis-match between fish and macroinvertebrate communities. How-ever, this pattern appears to be dominated by four sites, all ofwhichhave unusual fish communities comparedwith those of othersites sampled in this study (e.g., relatively species-rich or -poor).These deviations may be explained by additional, unmeasured envi-ronmental factors, such as proximity to lake outflows.
Sites with abiotic or biotic conditions that blur the distinctionbetween Shield and off-Shield regions offer insight into the abi-otic factors influencing fish and macroinvertebrate compositionin these systems. Although fish andmacroinvertebrate communi-ties are, for the most part, responding to the same abiotic driversin distinguishing Shield and off-Shield systems, there is evidenceto suggest that the macroinvertebrate taxa most responsible forthe Shield – off-Shield distinction are those that appear limited tocertain chemical conditions. In contrast, fish communities appearto be responding more to differences in physical habitat ratherthan chemical conditions. For example, sites that resemble thechemical conditions of off-Shield sites but are physically Shield-like have macroinvertebrate communities similar to those ofother off-Shield sites, but have Shield-like fish communities.Jackson and Harvey (1993) reported similar results for lake com-munities in central Ontario, where significant concordance wasobserved between fish and invertebrate communities, but wherethe fish communities were structured by morphological charac-teristics of lakes and the invertebrate communities by chemicalcharacteristics.
This association of chemical variables and macroinvertebratecomposition was also observed when variation attributed to landuse andwater chemistry resulted in no distinction between Shieldand off-Shield sites on the macroinvertebrate CCA plot. This sug-gests that these two groups of variables are important drivers indistinguishing the macroinvertebrate communities betweenthese two geological regions. However, when only water chemis-try or land-use variables are removed, the distinction again breaksdown, further highlighting the importance of both sets of vari-ables and their covariation, rather than just land use or waterchemistry alone. When variation attributed to physical habitat isremoved and only water chemistry and land-use variation areincluded, the Shield – off-Shield distinction remains, thereby in-dicating that these latter variables are the more important typesof structuring factors.
Differences between Shield and off-Shield fish communities ap-pear to be more tightly linked to physical habitat preferences, asthere is a shift in community composition between Shield andoff-Shield sites, rather than a combination of Shield and off-Shieldspecies at sites with off-Shield chemistry and Shield physical hab-itat. For example, sites with a greater proportion of largersubstrates — even when also higher in conductivity and pH —were associated with species preferring larger substrates. Whenexamining the role of water chemistry, physical habitat, and landuse simultaneously, there is little change in the distinction ofShield and off-Shield sites when variation attributed to any groupof variables is removed, suggesting that unlike the factors affect-ing macroinvertebrate communities, all three types of factorsplay a comparable role in distinguishing Shield and off-Shield fishcommunities. It has been argued that while parameters such asconductivity and pH can impact stream fishes (e.g., Wood et al.1990; Matthews 1998; Dussault et al. 2008), the effects may beindirect. This is in contrast to findings for some macroinverte-brate taxa, where it has been shown that aspects of water chem-istry can limit distributions of Gastropoda and Amphipoda(McKillop and Harrison 1972; Glazier et al. 1992; Neff and Jackson2011).
Further, low conductivity and pH can negatively impact fishbehavior or interactions with other species, but may not directlylead to absences of species, except under extreme conditions (Neffand Jackson 2012). For example, species of darters were very rarein Shield systems, both in this study and in previous work encom-passing a broader geographic range (Neff and Jackson 2012), anddistribution maps indicate that several species are mostly limitedto off-Shield areas in Ontario (Mandrak and Crossman 1992a). Thiscould be due to historical factors, such as pathways of postglacialrecolonization (Mandrak and Crossman 1992b), but theremay alsobe other limiting factors such as unfavorable water chemistry(e.g., low ionic strength contributing to physiological stress),which exacerbates any effects of predation or competition withother Shield species. However, further study is necessary to ad-dress this physiological hypothesis.
Concordance between these two groups of taxa supports theidea that the distinctions between Shield and off-Shield fish com-munities are not primarily due to dispersal limitation. However,as significant community concordance alone does not completelydiscount the influence of dispersal limitation on Shield and off-Shield fish communities, concordance between fish and macroin-vertebrate communities within Shield and off-Shield sites wasalso assessed. For example, it is possible that across all sites, mac-roinvertebrate communities are responding to differences in abi-otic conditions between Shield and off-Shield areas, while fishcommunities are responding to dispersal limitations. This wouldstill result in community concordance, as community structure ofboth groups of taxa would vary similarly between Shield and off-Shield areas, but the factors influencing that structure would bedifferent. Significant community concordance within each groupwould indicate that both groups of taxa are responding to similarenvironmental factors, regardless of location relative to theShield. This was found to be the case for Shield sites, which hadsignificantly concordant fish and macroinvertebrate communi-ties and thus have similar responses across Shield sites. This out-come would not be expected if colonization or dispersalmechanisms were a dominant factor on the Shield, given thedifferent dispersal capabilities of the two groups of taxa. The lackof significant concordance for off-Shield sites is likely due to lowstatistical power.
Thus, the results are consistent with the hypothesis of differ-ences in abiotic factors imposed by the geological distinctionbetween Shield and off-Shield. Whereas previous studies haveshown that historical processes can play a role in determiningfish species distributions in the province of Ontario (Jackson andHarvey 1989; Hinch et al. 1991; Mandrak 1995), our results suggestthat, as a whole, the Precambrian Shield is another importantbroad-scale influence on the physical and chemical habitat of loticsystems and their biological communities. Although historicalfactors no doubt influence the present-day community composi-tion, their influence appears less dominant than that imposed bythe environmental conditions distinguishing Shield from off-Shield sites.
Here, we consider the geological composition to be an “um-brella” or over-riding abiotic factor directly influencing waterchemistry, land-use patterns, and physical habitat of lotic sys-tems. Other studies have sought to separate the relative effects ofgeology and land-use patterns on aquatic communities (e.g.,Kratzer et al. 2006; Dow et al. 2006) and generally concluded thatit is difficult at best, as geology often plays a large role in deter-mining land use (Allan et al. 1997). For example, the prominentbedrock outcroppings and nutrient-poor soil of the Shield effec-tively limits agricultural development, but encourages other prac-tices such as mining and logging. Land use was an importantaspect in the distinction of Shield and off-Shield systems, as wellas the composition of both fish and macroinvertebrate communi-ties. The goal of this study, however, was not to separate theeffects of land use from that of geology on aquatic communities,
1148 Can. J. Fish. Aquat. Sci. Vol. 70, 2013
Published by NRC Research Press
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F W
YO
MIN
G o
n 09
/22/
13Fo
r pe
rson
al u
se o
nly.
but to examine the Shield overall as a potential regional driver ofthe total species pool relative to that of the adjacent off-Shieldregion. In other words, the geological composition of the Shieldinfluences water chemistry, physical habitat composition inaquatic systems, and land-use patterns in the surrounding catch-ments that, in turn, create either favorable or unfavorable condi-tions for various species.
Our study lends further support to the importance of regionalfactors in structuring local fish and macroinvertebrate communi-ties (e.g., Heino et al. 2003; Wang et al. 2003; Astorga et al. 2011),where a broad-scale geological feature leads to a well-definedboundary delineating physical, chemical, and landscape charac-teristics in aquatic systems that are distinct from those of anadjacent geological region. However, it is important to note thatwhile the abiotic conditions created by the Shield lead to distinctbiological communities across the landscape of south-central On-tario, there is still potential for variation in abiotic conditionswithin these two groups of systems. This variation in local condi-tions can also have important implications for the types of biotafound at specific locations, and the range of conditions within aregion can have important implications for regional species rich-ness. For example, if a region is composed of a set of nearly uni-form habitats, that region is likely to have a smaller species poolthan an area where there is a range of abiotic conditions, allwithin the context of the broader-scale filters represented by theregion. Anecdotal information regarding the types of systemsfound on the Precambrian Shield suggests that there is variationin physical and chemical conditions among different lotic sys-tems; for example, some Shield streams are slow and boggy, withdark tea-colored water, while others are fast, clear, and run overexposed bedrock. Further study can contribute to more detailedunderstanding of the local abiotic variation in Shield lotic systemsin relation to biotic communities. However, it has been shownhere that, at a broad scale, the Shield influences regional abioticcharacteristics of lotic systems such that we see distinct fish andmacroinvertebrate communities from adjacent systems off-Shield, further solidifying the importance of broad-scale factors inshaping biological communities.
AcknowledgementsAlex Manning, Jesse Elders, Ariel Nesbitt, and Christopher
Luzczeck provided invaluable assistance in field collections andmacroinvertebrate sample processing and identifications. Thiswork was funded by the Natural Sciences and Engineering Re-search Council of Canada (NSERC) and the University of Toronto.
ReferencesAHIS. 1987. Manual of Instructions. Aquatic Habitat Inventory Survey, Ontario
ALIS. 2008. Aquatic Landscape Information System dataset. Land InformationOntario, managed by Ontario Ministry of Natural Resources, Peterborough,Ontario, Canada.
Allan, J.D., Erickson, D.L., and Fay, J. 1997. The influence of catchment land useon stream integrity across multiple spatial scales. Freshw. Biol. 37: 149–161.doi:10.1046/j.1365-2427.1997.d01-546.x.
Angermeier, P.L., and Winston, M.R. 1998. Local vs. regional influences on localdiversity in stream fish communities of Virginia. Ecology, 79: 911–927. doi:10.2307/176589.
Astorga, A., Heino, J., Luoto, M., and Muotka, T. 2011. Freshwater biodiversity atregional extent: determinants of macroinvertebrate taxonomic richness inheadwater streams. Ecography, 34: 705–713. doi:10.1111/j.1600-0587.2010.06427.x.
Bilton, D.T., Freeland, J.R., and Okamura, B. 2001. Dispersal in freshwater inver-tebrates. Annu. Rev. Ecol. Syst. 32: 159–181. doi:10.1146/annurev.ecolsys.32.081501.114016.
Bohonak, A.J., and Jenkins, D.G. 2003. Ecological and evolutionary significanceof dispersal by freshwater invertebrates. Ecol. Lett. 6: 783–796. doi:10.1046/j.1461-0248.2003.00486.x.
Bowman, M.F., Ingram, R., Reid, R.A., Somers, K.M., Yan, N.D., Paterson, A.M.,Morgan, G.E., and Gunn, J.M. 2008. Temporal and spatial concordance incommunity composition of phytoplankton, zooplankton, macroinverte-
brate, crayfish, and fish on the Precambrian Shield. Can. J. Fish. Aquat. Sci.65(5): 919–932. doi:10.1139/f08-034.
Chapman, L.J., and Putnam, D.F. 1984. The physiography of Southern Ontario.Special Vol. 2. Ontario Geological Survey, Sudbury, Ontario.
Clarke, A.H. 1981. The freshwater molluscs of Canada. National Museum ofNatural Sciences, National Museum of Canada, Ottawa, Canada.
Dow, C.L., Arscott, D.B., and Newbold, J.D. 2006. Relating major ions and nutri-ents to watershed conditions across a mixed-use, water-supply watershed.J. N. Am. Benthol. Soc. 25: 887–911. doi:10.1899/0887-3593(2006)025[0887:RMIANT]2.0.CO;2.
Dussault, E.B., Playle, R.C., Dixon, D.G., and McKinley, R.S. 2008. Effects of soft-water acclimation on the physiology, swimming performance, and cardiacparameters of the rainbow trout, Oncorhynchus mykiss. Fish Physiol. Biochem.34: 313–322. doi:10.1007/s10695-007-9190-1. PMID:18958588.
Edwards, B.A., Jackson, D.A., and Somers, K.M. 2009. Multispecies crayfish de-clines in lakes: implications for species distributions and richness. J. N. Am.Benthol. Soc. 28: 719–732. doi:10.1899/08-148.1.
Environment Canada. 2011. Canadian climate normals or averages, 1971–2000[online]. National Climate Data and Information Archive. Available fromhttp://www.climate.weatheroffice.gc.ca.
Flotemersch, J.E., Stribling, J.B., Hughes, R.M., Reynolds, L., Paul, M.J., andWolter, C. 2011. Site length for biological assessment of boatable rivers. RiverRes. Appl. 27: 520–535. doi:10.1002/rra.1367.
Glazier, D.S., Horne, M.T., and Lehman, M.E. 1992. Abundance, body composi-tion and reproductive output of Gammarus minus (Crustacea: Amphipoda) inten cold springs differing in pH and ionic content. Freshw. Biol. 28: 149–163.doi:10.1111/j.1365-2427.1992.tb00572.x.
Heino, J. 2002. Concordance of species richness patterns among multiple fresh-water taxa: a regional perspective. Biodivers. Conserv. 11: 137–147. doi:10.1023/A:1014075901605.
Heino, J., Muotka, T., and Paavola, R. 2003. Determinants of macroinvertebratediversity in headwater streams: regional and local influences. J. Anim. Ecol.72: 425–434. doi:10.1046/j.1365-2656.2003.00711.x.
Hinch, S.G., Collins, N.C., and Harvey, H.H. 1991. Relative abundance of littoralzone fishes: biotic interactions, abiotic factors, and postglacial colonization.Ecology, 72: 1314–1324. doi:10.2307/1941104.
Hubbs, C.L., and Lagler, K.F. 2004. Fishes of the Great Lakes Region. (Revised byG.R. Smith.) University of Michigan Press, Michigan, USA.
Jackson, D.A. 1995. PROTEST: a procrustean randomization test of communityenvironment concordance. Ecoscience, 2: 297–303.
Jackson, D.A. 1997. Compositional data in community ecology: The paradigm orperil of proportions? Ecology, 78: 929–940. doi:10.1890/0012-9658(1997)078[0929:CDICET]2.0.CO;2.
Jackson, D.A., and Harvey, H.H. 1989. Biogeographic associations in fish assem-blages: local vs. regional processes. Ecology, 70: 1472–1484. doi:10.2307/1938206.
Jackson, D.A., and Harvey, H.H. 1993. Fish and benthic invertebrates: commu-nity concordance and community–environment relationships. Can. J. Fish.Aquat. Sci. 50(12): 2641–2651. doi:10.1139/f93-287.
Jeziorski, A., Yan, N.D., Paterson, A.M., DeSellas, A.M., Turner, M.A., Jeffries, D.S.,Keller, B., Weeber, R.C., McNicol, D.K., Palmer, M.E., McIver, K., Arseneau, K.,Ginn, B.K., Cumming, B.F., and Smol, J.P. 2008. The widespread threat ofcalcium decline in fresh waters. Science, 322: 1374–1377. doi:10.1126/science.1164949. PMID:19039134.
Jones, C., Somers, K.M., Craig, B., and Reynoldson, T.B. 2004. Ontario BenthosBiomonitoring Network Protocol Manual v.1. Ontario Ministry of the Envi-ronment, Queen's Printer for Ontario, Toronto.
Kilgour, B.W., and Barton, D.R. 1999. Associations between stream fish andbenthos across environmental gradients in southern Ontario, Canada.Freshw. Biol. 41: 553–566. doi:10.1046/j.1365-2427.1999.00402.x.
Kratzer, E.B., Jackson, J.K., Arscott, D.B., Aufdenkampe, A.K., Dow, C.L.,Kaplan, L.A., Newbold, J.D., and Sweeney, B.W. 2006. Macroinvertebrate dis-tribution in relation to land use and water chemistry in New York Citydrinking-water-supply watersheds. J. N. Am. Benthol. Soc. 25: 954–976. doi:10.1899/0887-3593(2006)025[0954:MDIRTL]2.0.CO;2.
Mandrak, N.E. 1995. Biogeographic patterns of fish species richness in Ontariolakes in relation to historical and environmental factors. Can. J. Fish. Aquat.Sci. 52(7): 1462–1474. doi:10.1139/f95-141.
Mandrak, N.E., and Crossman, E.J. 1992a. A checklist of Ontario freshwaterfishes. Royal Ontario Museum, Toronto, Canada.
Mandrak, N.E., and Crossman, E.J. 1992b. Postglacial dispersal of freshwaterfishes into Ontario. Can. J. Zool. 70(11): 2247–2259. doi:10.1139/z92-302.
Matthews, W.J. 1998. Patterns in freshwater fish ecology. Chapman & Hall, NewYork, USA.
McKillop, W.B., and Harrison, A.D. 1972. Distribution of aquatic gastropodsacross an interface between the Canadian Shield and limestone formations.Can. J. Zool. 50(11): 1433–1445. doi:10.1139/z72-192.
Merritt, R.W., and Cummins, K.W. (Editors). 1996. An introduction to the aquaticinsects of North America. 3rd ed. Kendall Hunt Publishing Company, Iowa,USA.
Mykrä, H., Heino, J., and Muotka, T. 2007. Scale-related patterns in the spatialand environmental components of stream macroinvertebrate assemblage
variation. Global Ecol. Biogeogr. 16: 149–159. doi:10.1111/j.1466-8238.2006.00272.x.
Neff, M.R., and Jackson, D.A. 2011. Effects of broad-scale geological changes onpatterns in macroinvertebrate assemblages. J. N. Am. Benthol. Soc. 30: 459–473. doi:10.1899/10-052.1.
Neff, M.R., and Jackson, D.A. 2012. Geology as a structuring mechanism ofstream fish communities. Trans. Am. Fish. Soc. 141: 962–974. doi:10.1080/00028487.2012.676591.
Ontario Geological Survey. 2003. Surficial geology of southern Ontario – (includingToronto). Ontario Geological Survey, Miscellaneous Release Data 128.
Paavola, R., Muotka, T., Virtanen, R., Heino, J., Jackson, D.A., and Maki-Petäys, A.2006. Spatial scale affects community concordance among fishes, benthicmacroinvertebrates, and bryophytes in streams. Ecol. Appl. 16: 368–379. doi:10.1890/03-5410. PMID:16705986.
Peckarsky, B.L., Fraissinet, P., Pention, M.A., and Conklin, D.J., Jr. 1990. Freshwa-ter macroinvertebrates of northeastern North America. Cornell UniversityPress, Ithaca, New York.
Peres-Neto, P.R., and Jackson, D.A. 2001. How well do multivariate data sets
match? The advantages of a Procrustean superimposition approach over theMantel test. Oecologia, 129: 169–178. doi:10.1007/s004420100720.
R Development Core Team. 2010. R 2.10.1: a language and environment forstatistical computing. R Foundation for Statistical Computing, Vienna,Austria.
Stanfield, L. (Editor). 2005. Ontario Stream Assessment Protocol (OSAP), v.7.Thorp, J.H., and Covich, A.P. (Editors). 1991. Ecology and classification of North
American freshwater invertebrates. Academic Press Inc., San Diego, California.Wang, L., Lyons, J., Rasmussen, P., Seelbach, P., Simon, T., Wiley, M., Kanehl, P.,
Baker, E., Niemela, S., and Stewart, P.M. 2003.Watershed, reach, and riparianinfluences on stream fish assemblages in the Northern Lakes and ForestEcoregion, U.S.A. Can. J. Fish. Aquat. Sci. 60(5): 491–505. doi:10.1139/f03-043.
Wohl, E., and Legleiter, C.J. 2003. Controls on pool characteristics along aresistant-boundary channel. J. Geol. 111: 103–114. doi:10.1086/344667.
Wood, C.M., McDonald, D.G, Ingersoll, C.G., Mount, D.R., Johannsson, O.E.,Landsberger, S., and Bergman, H.L. 1990. Effects of water acidity, calcium,and aluminum on whole body ions of brook trout (Salvelinus fontinalis) con-tinuously exposed from fertilization to swim-up: a study by instrumentalneutron activation analysis. Can. J. Fish. Aquat. Sci. 47(8): 1593–1603. doi:10.1139/f90-181.