Regulation of plasma testosterone, corticosterone, and metabolites in response to stress, reproductive stage, and social challenges in a desert male songbird Pierre Deviche a,⇑ , Benjamin Beouche-Helias b , Scott Davies a , Sisi Gao a , Samuel Lane a , Shelley Valle a a School of Life Sciences, Arizona State University, Tempe, AZ 85287-4501, USA b Universite de Poitiers, Faculte des Sciences Fondamentales et Appliquees, Poitiers F-86022, France article info Article history: Available online xxxx Keywords: Challenge hypothesis Glycemia Reproduction Simulated territorial intrusion Song playback Uric acid abstract In many male vertebrates, the secretion of reproductive (gonadal androgens) and adrenocortical (gluco- corticoids) hormones varies seasonally and in response to environmental stimuli, and these hormones exert numerous behavioral and metabolic effects. We performed two field studies on adult male Ruf- ous-winged Sparrows, Peucaea carpalis, a Sonoran Desert rain-dependent sedentary species, to (a) deter- mine seasonal changes in initial (baseline) and acute stress-induced plasma testosterone (T), corticosterone (CORT), and two metabolites (uric acid and glucose) and (b) compare the effects of two types of social challenge (song playback or simulated territorial intrusion consisting of song playback plus exposure to a live decoy bird) on plasma T, CORT, these metabolites, and territorial behavior. Initial plasma T was higher during the summer breeding period than during post-breeding molt. Acute stress resulting from capture and restraint for 30 min decreased plasma T in breeding condition birds but not in the fall, revealing that this decrease is seasonally regulated. Initial plasma CORT did not change season- ally, but plasma CORT increased in response to acute stress. This increase was likewise seasonally regu- lated, being relatively smaller during autumnal molt than in the summer. We found no evidence that acute stress levels of CORT are functionally related to stress-depressed plasma T and, therefore, that plasma T decreases during stress as a result of elevated plasma CORT. Thirty minutes of exposure to sim- ulated territorial intrusion resulted in different behavior than 30 min of exposure to song playback, with increased time spent near the decoy and decreased number of overhead flights. Neither type of social challenge influenced plasma T, thus offering no support for the hypothesis that plasma T either responds to or mediates the behavioral effects of social challenge. Exposure to both social challenges elevated plasma CORT, but simulated territorial intrusion was more effective in this respect than song playback. Plasma uric acid and glucose decreased during acute stress, but only plasma uric acid decreased during social challenge. Thus, an elevation in plasma CORT was consistently associated with a decrease in plasma uric acid, but not with a change in glycemia. These results enhance our understanding of the short-term relationships between T, CORT, and avian territorial behavior. They provide novel information on the endocrine effects of acute stress, in particular on plasma T, in free-ranging birds, and are among the first in these birds to link these effects to metabolic changes. Ó 2014 Elsevier Inc. All rights reserved. 1. Introduction In response to adverse conditions (i.e., stressors), vertebrates often activate their hypothalamic–pituitary–adrenal axis, resulting in elevated plasma glucocorticoids, and enter the ‘‘emergency life history stage’’ (Wingfield et al., 1998). The transition to this life history stage involves physiological and behavioral changes that promote survival and it is, therefore, thought to be adaptive (Sapol- sky et al., 2000; Wingfield and Kitaysky, 2002). These changes in- clude energy mobilization, increased cardiovascular tone, and sharpened cognition. Transition to the emergency life history stage can also affect reproductive processes, in particular the secretion of reproductive hormones, and studies suggest a role for glucocorti- coids such as cortisol and corticosterone (CORT) in this effect. The rapid (i.e., within minutes to hours) rise in plasma CORT in response to acute stress concurs with a rapid fall in plasma testosterone (T) in fish (Pickering et al., 1987), amphibians http://dx.doi.org/10.1016/j.ygcen.2014.01.010 0016-6480/Ó 2014 Elsevier Inc. All rights reserved. ⇑ Corresponding author. Fax: +1 480 965 6899. E-mail address: [email protected](P. Deviche). General and Comparative Endocrinology xxx (2014) xxx–xxx Contents lists available at ScienceDirect General and Comparative Endocrinology journal homepage: www.elsevier.com/locate/ygcen Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosterone, corticosterone, and metabolites in response to stress, reproductive stage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol. (2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010
Transcript
General and Comparative Endocrinology xxx (2014) xxx–xxx
Contents lists available at ScienceDirect
General and Comparative Endocrinology
journal homepage: www.elsevier .com/locate /ygcen
Regulation of plasma testosterone, corticosterone, and metabolitesin response to stress, reproductive stage, and social challenges in a desertmale songbird
http://dx.doi.org/10.1016/j.ygcen.2014.01.0100016-6480/� 2014 Elsevier Inc. All rights reserved.
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosterone, corticosterone, and metabolites in response to stress, reprostage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol. (2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010
Pierre Deviche a,⇑, Benjamin Beouche-Helias b, Scott Davies a, Sisi Gao a, Samuel Lane a, Shelley Valle a
a School of Life Sciences, Arizona State University, Tempe, AZ 85287-4501, USAb Universite de Poitiers, Faculte des Sciences Fondamentales et Appliquees, Poitiers F-86022, France
In many male vertebrates, the secretion of reproductive (gonadal androgens) and adrenocortical (gluco-corticoids) hormones varies seasonally and in response to environmental stimuli, and these hormonesexert numerous behavioral and metabolic effects. We performed two field studies on adult male Ruf-ous-winged Sparrows, Peucaea carpalis, a Sonoran Desert rain-dependent sedentary species, to (a) deter-mine seasonal changes in initial (baseline) and acute stress-induced plasma testosterone (T),corticosterone (CORT), and two metabolites (uric acid and glucose) and (b) compare the effects of twotypes of social challenge (song playback or simulated territorial intrusion consisting of song playback plusexposure to a live decoy bird) on plasma T, CORT, these metabolites, and territorial behavior. Initialplasma T was higher during the summer breeding period than during post-breeding molt. Acute stressresulting from capture and restraint for 30 min decreased plasma T in breeding condition birds but notin the fall, revealing that this decrease is seasonally regulated. Initial plasma CORT did not change season-ally, but plasma CORT increased in response to acute stress. This increase was likewise seasonally regu-lated, being relatively smaller during autumnal molt than in the summer. We found no evidence thatacute stress levels of CORT are functionally related to stress-depressed plasma T and, therefore, thatplasma T decreases during stress as a result of elevated plasma CORT. Thirty minutes of exposure to sim-ulated territorial intrusion resulted in different behavior than 30 min of exposure to song playback, withincreased time spent near the decoy and decreased number of overhead flights. Neither type of socialchallenge influenced plasma T, thus offering no support for the hypothesis that plasma T either respondsto or mediates the behavioral effects of social challenge. Exposure to both social challenges elevatedplasma CORT, but simulated territorial intrusion was more effective in this respect than song playback.Plasma uric acid and glucose decreased during acute stress, but only plasma uric acid decreased duringsocial challenge. Thus, an elevation in plasma CORT was consistently associated with a decrease inplasma uric acid, but not with a change in glycemia. These results enhance our understanding of theshort-term relationships between T, CORT, and avian territorial behavior. They provide novel informationon the endocrine effects of acute stress, in particular on plasma T, in free-ranging birds, and are amongthe first in these birds to link these effects to metabolic changes.
� 2014 Elsevier Inc. All rights reserved.
1. Introduction
In response to adverse conditions (i.e., stressors), vertebratesoften activate their hypothalamic–pituitary–adrenal axis, resultingin elevated plasma glucocorticoids, and enter the ‘‘emergency lifehistory stage’’ (Wingfield et al., 1998). The transition to this lifehistory stage involves physiological and behavioral changes that
promote survival and it is, therefore, thought to be adaptive (Sapol-sky et al., 2000; Wingfield and Kitaysky, 2002). These changes in-clude energy mobilization, increased cardiovascular tone, andsharpened cognition. Transition to the emergency life history stagecan also affect reproductive processes, in particular the secretion ofreproductive hormones, and studies suggest a role for glucocorti-coids such as cortisol and corticosterone (CORT) in this effect.The rapid (i.e., within minutes to hours) rise in plasma CORT inresponse to acute stress concurs with a rapid fall in plasmatestosterone (T) in fish (Pickering et al., 1987), amphibians
2 P. Deviche et al. / General and Comparative Endocrinology xxx (2014) xxx–xxx
(Woodley and Lacy, 2010), mammals (Dong et al., 2004), and birds(Lynn et al., 2010; Wingfield et al., 1982), including male Rufous-winged Sparrows, Peucaea carpalis (Deviche et al., 2012a), the spe-cies investigated in the present study. Mammalian studies reveal adirect action of glucocorticoids on testicular endocrine function,showing that glucocorticoids can inhibit plasma T production byacting on interstitial (Leydig) testicular cells through a non-geno-mic action (Dong et al., 2004; Hu et al., 2008; Martin and Tremblay2008). Avian testes contain glucocorticoid receptors and, similar tomammals, elevated plasma CORT may, therefore, acutely decreaseplasma T in birds through a testis-mediated mechanism (Kwoket al., 2007).
In most free-ranging male vertebrates, plasma T varies season-ally such that levels are highest when males are competing formates at the beginning of the breeding season (Landys et al.,2010; Van Duyse et al., 2003; Wingfield et al., 1987). Initial (oftenreferred to as baseline) and stress-induced plasma CORT can alsovary seasonally, but whether this is the case is species-specific.In some vertebrates, initial plasma CORT is elevated during ener-getically demanding times such as the early breeding season(Breuner and Orchinik, 2001; John-Alder et al., 2009; Place and Ke-nagy, 2000; Romero, 2002) or migration (Romero et al., 1997). Inother male vertebrates initial plasma CORT does, however, notchange seasonally (Landys et al., 2010). Likewise, stress-inducedplasma CORT fluctuates seasonally in some male vertebrates(Breuner and Orchinik, 2001; Klukowski, 2011; Romero, 2002;Romero et al., 1997) but not in others (Place and Kenagy, 2000).In free-ranging birds, the CORT response to acute stress is oftensmaller during than outside the molt period (Cyr et al., 2008; Re-mage-Healey and Romero 2000; Romero, 2002; Romero et al.,1998a).
Studies on species exhibiting seasonal variation in plasma T andin initial and stress-induced plasma CORT provide an opportunityto investigate functional relationships between these hormones.Here we addressed this question using free-ranging adult maleRufous-winged Sparrows sampled at different reproductive stages.This Sonoran Desert passerine breeds during the summer monsoon(Lowther et al., 1999). In males, plasma T is seasonally highest inJuly and August, when birds are in breeding condition, and lowerin September, after birds complete their breeding cycle (Devicheet al., 2006, 2012a). No study on Rufous-winged Sparrows hasdetermined seasonal changes in plasma CORT. The first mainobjective of the present study was to examine seasonal variationin initial and stress-induced plasma CORT and to relate this varia-tion to changes in initial and stress-induced plasma T in the samebirds. We predicted that if plasma CORT changes seasonally andthe changes are functionally related to plasma T, an inverse rela-tionship would exist between these hormones on a seasonal basisand in response to acute stress.
In addition to fluctuating during acute stress, plasma CORT andT in free-ranging territorial males can be socially modulated. Thismodulation is often investigated by presenting males with a socialchallenge consisting of exposure to conspecific song recordings(song playback, SPB) or to a conspecific decoy male and conspecificsong recordings (simulated territorial intrusion, STI). Exposure toSPB simulates an auditory social challenge whereas exposure toSTI simulates a combined visual and auditory social challenge.Exposure to SPB or STI often provokes an aggressive responseand in some cases, also elicits endocrine changes (Goymannet al., 2007; Landys et al., 2007). Several studies found that expo-sure to STI increases plasma CORT (Gill et al., 2008; Landys et al.,2007, 2010; Van Duyse et al., 2004). This exposure also stimulatesplasma T in some species (Wingfield, 1994), but either reduces orhas no effect on plasma T in others (Apfelbeck and Goymann2011; Hau and Beebe, 2011; Landys et al., 2007, 2010). Likewise,exposure to SPB either increases (Wikelski et al., 1999) or does
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosstage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol.
not influence plasma T, as was the case in the Rufous-wingedSparrow (Deviche et al., 2006) and in the congeneric Cassin’s Spar-row, Peucaea cassinii (Deviche et al., 2012b). Elevated plasma T inresponse to social challenges may function to increase the subse-quent persistence of aggressive responses (Challenge Hypothesis,Wingfield et al., 1987, 1990) and species-specific differences inendocrine responses to social challenges are postulated to be re-lated to life history characteristics such as varying mating systemsand brood numbers per season (Goymann et al., 2007; Hirschenha-user et al., 2003; Landys et al., 2007). However, reaching firm con-clusions on this subject has been complicated by methodologicaldifferences between studies. In particular, little research has beencarried out to compare the endocrine changes that are inducedby exposure to SPB vs. STI within the same study and in the samespecies. To address this issue, the second objective of the presentstudy was to compare the effects of auditory (SPB) vs. auditory plusvisual (STI) stimulation on the aggressive behavior and on plasma Tand CORT of male Rufous-Winged Sparrows in breeding condition.This comparison also offered the opportunity to further investigaterelationships between plasma T and CORT and, therefore, to com-pare this relationship to that observed in the above seasonal study.
The downstream metabolic effects of stress-related endocrinechanges are critical to determining whether these changes improvethe chances of survival and fitness, but these effects have not beenwell investigated in free-ranging birds. Glucose is a main substratefor energy production. In birds, CORT may, in some circumstances,regulate plasma glucose, but information on this subject is conflict-ing. Acute stress in captive European Starlings, Sturnus vulgaris, in-creased plasma glucose during the night but not during the dayand this effect is not observed in molting birds (Remage-Healeyand Romero, 2001, 2002). In this species, CORT administration tocaptive molting birds during the day elevated plasma glucose,but the significance of this finding is obscured by the fact thatthe treatment resulted in supra-physiological plasma CORT (Re-mage-Healey and Romero 2002). Other studies likewise found thatacute stress either increases (Corbel et al., 2010; Davies et al.,2013) or does not influence glycemia (Fokidis et al., 2011a; HouseSparrow, Passer domesticus: M. Romero, personal communication),and Corbel et al. (2010) obtained no evidence for an interaction be-tween CORT and the metabolic response (including plasma glu-cose) to stress in penguins. Uric acid is a product of proteindegradation (Costantini 2008) and a potent antioxidant that is ab-sorbed into tissues and oxidized to protect against tissue damage(Stinefelt et al., 2005). Stressful stimuli increase plasma uric acidin some species (Cohen et al., 2007; Tsahar et al., 2006), but hasthe opposite effect in others (Cohen et al., 2007; Davies et al.,2013). If CORT in free-ranging birds plays a major proximate rolein regulating metabolic responses to acute stress – including glyce-mia and plasma uric acid, we predicted changes in plasma CORT tobe consistently related to changes in the circulating levels of thesemetabolites across experimental conditions. To test this prediction,we measured plasma glucose and uric acid in free-ranging malesacross different reproductive stages, in response to social chal-lenge, and before and after acute stress.
2. Materials and methods
2.1. Study site, species, capture, and blood sample collection
We sampled adult male Rufous-winged Sparrows between June21st and September 26th 2012 in the Santa Rita ExperimentalRange, Pima County, Arizona, USA (altitude: 979 m; latitude:31�490N; longitude: 110�550W). This sedentary species is territorialand responds behaviorally to SPB year-round (Lowther et al., 1999;personal observations).
terone, corticosterone, and metabolites in response to stress, reproductive(2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010
P. Deviche et al. / General and Comparative Endocrinology xxx (2014) xxx–xxx 3
We caught sparrows using a mist net and conspecific SPB.Depending on the study, in addition to the SPB, some birds werealso exposed to conspecific STI (see below). Within two minutesof capture, we collected a first blood sample from each bird todetermine initial levels of plasma hormones or metabolites (seebelow). Blood samples were collected from the right jugular veinusing a heparinized syringe. Immediately after collection, theywere placed on ice until centrifuged later the same day. Plasmawas harvested using a Hamilton glass syringe and frozen until as-sayed for T, CORT, glucose, and uric acid (see below).
Prior to release at the capture site, we measured the body mass(±0.1 g), wing chord (±1 mm), cloacal protuberance width, which isa T-dependent secondary sexual characteristic (±0.1 mm), and fur-cular fat score (0–5; Rogers, 1991) of each male, and determinedwhether birds were molting their primary feathers. Each sparrowreceived an intramuscular (i.m.; pectoral muscle) injection of sal-ine solution (0.9% NaCl in water) to compensate for the volumeof blood removed and was marked with a uniquely numbered USGeological Survey aluminum tarsal band.
All procedures were approved by the Arizona State UniversityInstitutional Animal Care and Use Committee and were conductedunder appropriate scientific collecting permits issued by the Ari-zona Game and Fish Department and the US Fish and WildlifeService.
2.2. Seasonal study
The goal of this study was twofold. First, we investigatedwhether the acute stress-induced decrease in plasma T changeson a seasonal basis, and, second, how initial and stress-inducedplasma T relate to plasma CORT. Sparrows were sampled at fourtimes: June 21st (n = 9; pre-monsoon and prior to breeding), July16th–21st (n = 12; monsoon, during the breeding period), August12th (n = 10; monsoon), and September 26th (n = 10; end of themonsoon and the breeding season). We selected these periods be-cause plasma T naturally varies over this time frame (Deviche et al.,2006). During each month, we caught sparrows using SPB and col-lected an initial blood sample (110 ll) within two minutes of cap-ture. To elicit an acute stress response, we then confined birds toindividual breathable cloth bags. After 30 min of confinement, wecollected a second blood sample (110 ll) to determine stress-in-duced plasma hormones and metabolite concentrations.
2.3. Behavioral study
The first goal of this study was to compare the behavioral, endo-crine (plasma T and CORT), and metabolic (plasma glucose and uricacid) responses to SPB and STI. To this end, during July 16th–19thwe located males based either on their spontaneous singing behav-ior or on their response to brief (usually less than 1 min) SPB expo-sure. Once a focal male was located, we placed the speaker systemand a furled mist net at the estimated center of the area where abird had been observed. Next to the speaker, we also placed a cagethat was either empty (SPB group, n = 10) or contained a freshlycaught decoy male (STI group, n = 10). Decoy males were caughtusing SPB as described above and within 1 h of being used forSTI challenges. Challenged sparrows were alternatively assignedto the SPB or STI group and any given decoy male was used for asingle trial, and then released. Once the speaker broadcasting con-specific song was turned on, we did not collect behavioral mea-surements during the first 5 min of challenge because thepresence of the experimenter approaching and activating thespeaker may modify the focal bird’s behavior during this time. Inthe next 15 min, we recorded the number of songs, the numberof flights over the net, and the cumulative time spent on theground within <5 m of the cage. The period of behavioral
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosstage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol.
observations was followed with 5 min during which we did nottake any behavioral measurements. We then unfurled the mistnet and caught the focal male as quickly as possible (usually within5 min). Thus, challenged males were exposed to SPB or STI forapproximately 30 min before capture. Within two minutes of cap-ture, we collected a blood sample (200 ll) from each male as de-scribed above.
The second objective of the study was to compare the endocrineeffects of social challenges with those of capture and restraint forthe same duration. For this, data for SPB- and STI-exposed birdswere compared to those collected for birds in the Seasonal Studyand sampled immediately after capture (Initial) or after 30 minof restraint (stress-induced) during the same period (July 16th–21st).
2.4. Hormone and metabolite assays
Samples were assayed for each hormone or metabolite in dupli-cate and on the same day using the manufacturers’ recommendedprocedures, except that uric acid concentrations in samples col-lected during the Seasonal Study were measured on two separatedays. We quantified plasma T and CORT using commercial en-zyme-linked immunoassay kits (Enzo Life Sciences, Farmingdale,NY), both of which we have validated for use in the Rufous-wingedSparrow (Deviche et al., 2006, 2010). For the assays, we dilutedplasma 13� for T and 15� for CORT before dispensing into assayplate wells. The average inter- and intra-assay coefficients of vari-ation for T were 5.24% and 6.78%, respectively, and the assay sen-sitivity was 4.47 pg/ml. For the CORT assay, the average inter- andintra-assay coefficients of variation were 10.15% and 9.80%, respec-tively, and the assay sensitivity was 179 pg/ml. We measured plas-ma glucose and uric acid using colorimetric assay kits (glucose:Cayman Chemical Co., Ann Arbor, MI; uric acid: BioAssay Systems,Hayward, CA). To validate these assays for use in Rufous-wingedSparrows, we verified that a serially diluted plasma sample re-sulted in a dilution curve that was parallel to the standard curve.To measure plasma glucose, we diluted plasma by 5� using assaybuffer before dispensing into assay wells. The average inter- andintra-assay coefficients of variation for the glucose assay were0.6% and 1.2%, respectively, and the assay sensitivity was0.15 mg/dl. Uric acid was measured using 5 ll of undiluted plasmaper assay well. The average inter- and intra-assay coefficients ofvariation for the glucose assay were 1.7% and 2.7%, respectively,and the assay sensitivity was 0.025 mg/dl. Samples were randomlyassigned to assay plates. When two samples were collected fromthe same bird, they were assigned to the same plate.
2.5. Statistical analyses
We statistically analyzed data using SigmaPlot 12.5 (Systat Soft-ware, Inc., San Jose, CA) and GraphPad Prism 6 (GraphPad Software,Inc., San Diego, CA). We used Student’s t-tests to compare twogroups of interval-scale, normally distributed data. When pairs ofindependent data sets were not normally distributed or collectedon an ordinal scale, we compared them using a Mann–Whitneyrank sum test (Siegel, 1956). We used Fisher exact probability teststo compare the proportion of birds that were molting during differ-ent months (Seasonal Study). Comparisons of more than two setsof independent, interval-scale data were made using one-way or,when appropriate, two-way repeated measure analysis of variance(ANOVA) followed with Student–Newman–Keuls tests for pair-wise multiple comparisons. If ANOVA assumptions (normality(Shapiro–Wilk test) and/or homoscedasticity (equal variance test))were violated (p < 0.05), we either log10 transformed data toachieve normality and equal variance or ranked the data (Conoverand Iman, 1981) before ANOVA. In the case of one-way ANOVA,
terone, corticosterone, and metabolites in response to stress, reproductive(2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010
4 P. Deviche et al. / General and Comparative Endocrinology xxx (2014) xxx–xxx
when datum transformation did not result in normality and equalvariance, we compared groups using non-parametric Kruskal–Wal-lis one-way ANOVA on ranks (KW) followed with Dunn’s methodtests for pair-wise multiple comparisons. Data sets that we ana-lyzed using parametric statistical methods are presented asmeans ± standard errors (s.e.). Data sets that we analyzed usingnon-parametric tests are shown as medians ± 0.5 interquartileintervals (IQ). The statistical significance level of all tests was setat a = 0.05.
3. Results
3.1. Seasonal study
3.1.1. Capture times and external morphologyAll birds were caught between 6:15 AM and 13:20 PM. An AN-
OVA comparing average capture times found these times to differbetween the four experimental groups (F3,37 = 3.10, p = 0.039),but pair-wise multiple comparisons revealed no statistically signif-icant differences between months. Birds were caught, on average,after exposure to SPB for 3:30 min ± 5:48 min (median ± IQ); thisduration was similar whether birds were caught in June, July, Au-gust, or September (KW: H = 6.42, p > 0.09).
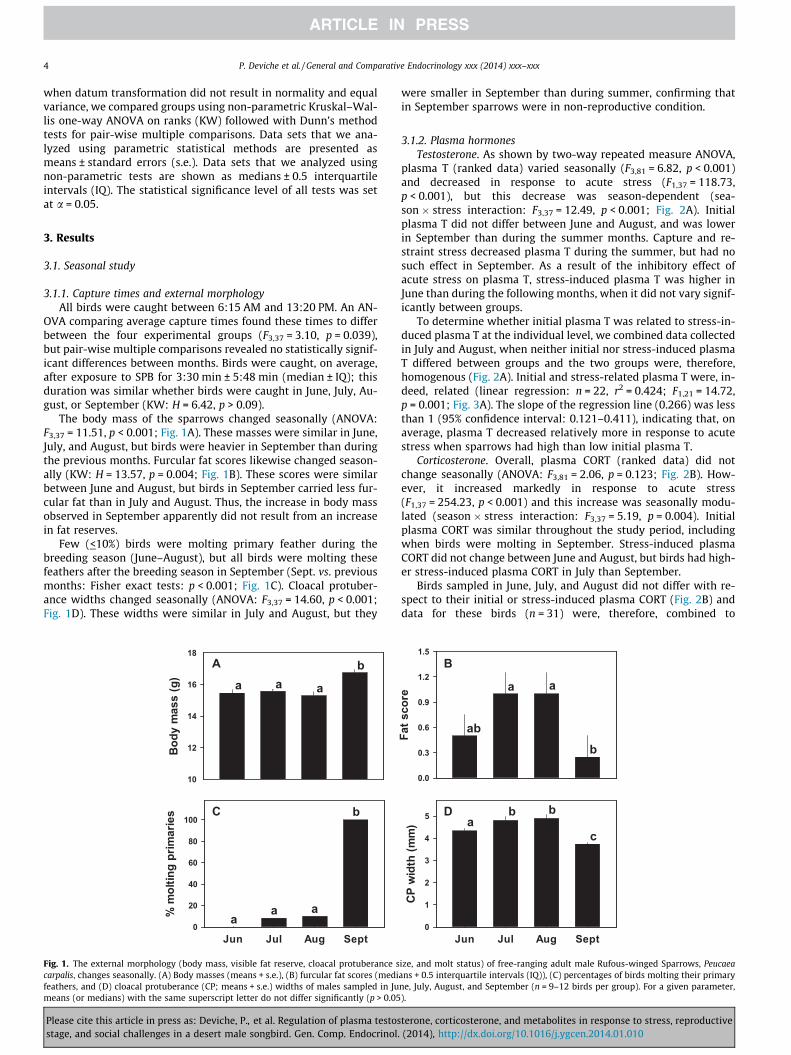
The body mass of the sparrows changed seasonally (ANOVA:F3,37 = 11.51, p < 0.001; Fig. 1A). These masses were similar in June,July, and August, but birds were heavier in September than duringthe previous months. Furcular fat scores likewise changed season-ally (KW: H = 13.57, p = 0.004; Fig. 1B). These scores were similarbetween June and August, but birds in September carried less fur-cular fat than in July and August. Thus, the increase in body massobserved in September apparently did not result from an increasein fat reserves.
Few (<10%) birds were molting primary feather during thebreeding season (June–August), but all birds were molting thesefeathers after the breeding season in September (Sept. vs. previousmonths: Fisher exact tests: p < 0.001; Fig. 1C). Cloacal protuber-ance widths changed seasonally (ANOVA: F3,37 = 14.60, p < 0.001;Fig. 1D). These widths were similar in July and August, but they
Body
mas
s (g
)
10
12
14
16
18
a a a
b
Jun Jul Aug Sept
% m
oltin
g pr
imar
ies
0
20
40
60
80
100
aa a
b
A
C
Fig. 1. The external morphology (body mass, visible fat reserve, cloacal protuberance scarpalis, changes seasonally. (A) Body masses (means + s.e.), (B) furcular fat scores (medifeathers, and (D) cloacal protuberance (CP; means + s.e.) widths of males sampled in Jumeans (or medians) with the same superscript letter do not differ significantly (p > 0.05
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosstage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol.
were smaller in September than during summer, confirming thatin September sparrows were in non-reproductive condition.
3.1.2. Plasma hormonesTestosterone. As shown by two-way repeated measure ANOVA,
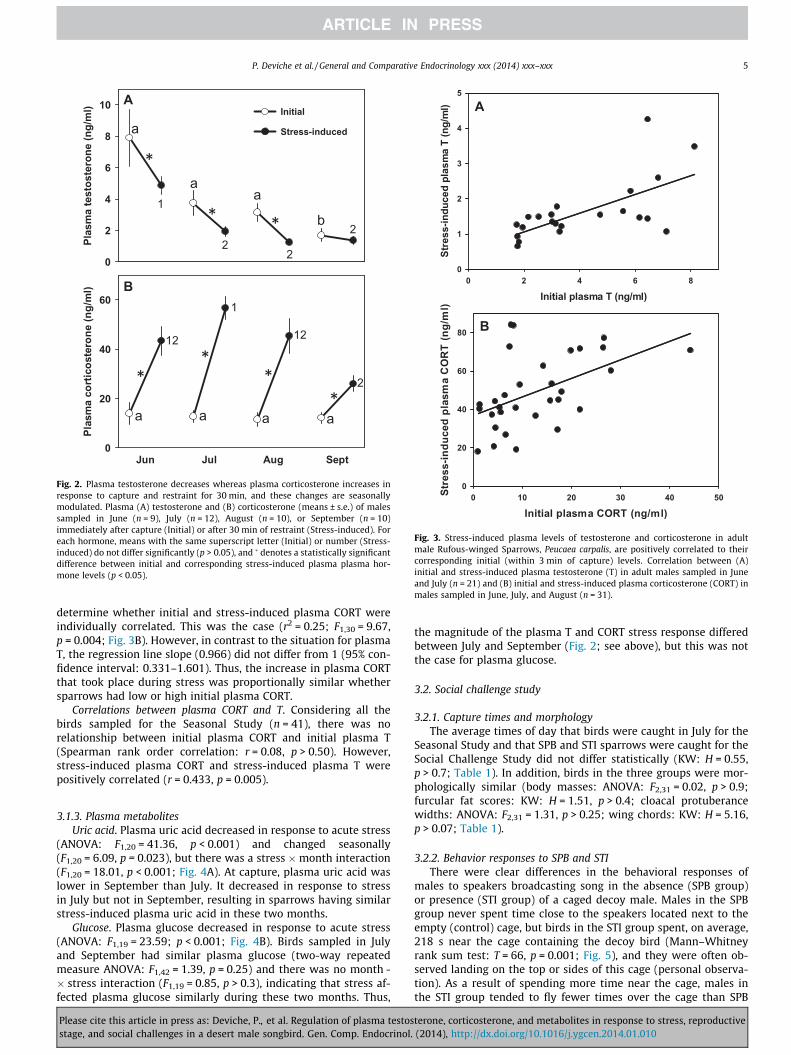
plasma T (ranked data) varied seasonally (F3,81 = 6.82, p < 0.001)and decreased in response to acute stress (F1,37 = 118.73,p < 0.001), but this decrease was season-dependent (sea-son � stress interaction: F3,37 = 12.49, p < 0.001; Fig. 2A). Initialplasma T did not differ between June and August, and was lowerin September than during the summer months. Capture and re-straint stress decreased plasma T during the summer, but had nosuch effect in September. As a result of the inhibitory effect ofacute stress on plasma T, stress-induced plasma T was higher inJune than during the following months, when it did not vary signif-icantly between groups.
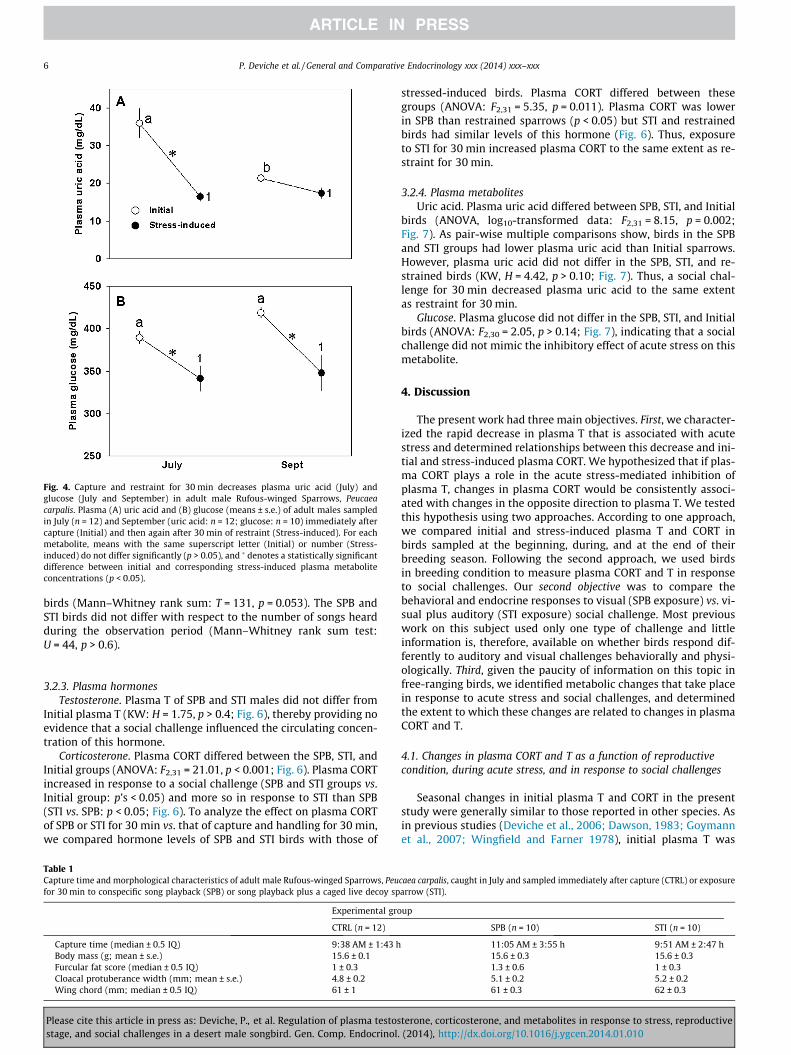
To determine whether initial plasma T was related to stress-in-duced plasma T at the individual level, we combined data collectedin July and August, when neither initial nor stress-induced plasmaT differed between groups and the two groups were, therefore,homogenous (Fig. 2A). Initial and stress-related plasma T were, in-deed, related (linear regression: n = 22, r2 = 0.424; F1,21 = 14.72,p = 0.001; Fig. 3A). The slope of the regression line (0.266) was lessthan 1 (95% confidence interval: 0.121–0.411), indicating that, onaverage, plasma T decreased relatively more in response to acutestress when sparrows had high than low initial plasma T.
Corticosterone. Overall, plasma CORT (ranked data) did notchange seasonally (ANOVA: F3,81 = 2.06, p = 0.123; Fig. 2B). How-ever, it increased markedly in response to acute stress(F1,37 = 254.23, p < 0.001) and this increase was seasonally modu-lated (season � stress interaction: F3,37 = 5.19, p = 0.004). Initialplasma CORT was similar throughout the study period, includingwhen birds were molting in September. Stress-induced plasmaCORT did not change between June and August, but birds had high-er stress-induced plasma CORT in July than September.
Birds sampled in June, July, and August did not differ with re-spect to their initial or stress-induced plasma CORT (Fig. 2B) anddata for these birds (n = 31) were, therefore, combined to
Jun Jul Aug Sept
CP w
idth
(mm
)
0
1
2
3
4
5
Fat s
core
0.0
0.3
0.6
0.9
1.2
1.5
ab b
c
abb
a a
B
D
ize, and molt status) of free-ranging adult male Rufous-winged Sparrows, Peucaeaans + 0.5 interquartile intervals (IQ)), (C) percentages of birds molting their primaryne, July, August, and September (n = 9–12 birds per group). For a given parameter,).
terone, corticosterone, and metabolites in response to stress, reproductive(2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010
Fig. 2. Plasma testosterone decreases whereas plasma corticosterone increases inresponse to capture and restraint for 30 min, and these changes are seasonallymodulated. Plasma (A) testosterone and (B) corticosterone (means ± s.e.) of malessampled in June (n = 9), July (n = 12), August (n = 10), or September (n = 10)immediately after capture (Initial) or after 30 min of restraint (Stress-induced). Foreach hormone, means with the same superscript letter (Initial) or number (Stress-induced) do not differ significantly (p > 0.05), and � denotes a statistically significantdifference between initial and corresponding stress-induced plasma plasma hor-mone levels (p < 0.05).
Initial plasma T (ng/ml)0 2 4 6 8
Stre
ss-in
duce
d pl
asm
a T
(ng/
ml)
0
1
2
3
4
5
A
Initial plasma CORT (ng/ml)0 10 20 30 40 50St
ress
-indu
ced
plas
ma
CO
RT
(ng/
ml)
0
20
40
60
80 B
Fig. 3. Stress-induced plasma levels of testosterone and corticosterone in adultmale Rufous-winged Sparrows, Peucaea carpalis, are positively correlated to theircorresponding initial (within 3 min of capture) levels. Correlation between (A)initial and stress-induced plasma testosterone (T) in adult males sampled in Juneand July (n = 21) and (B) initial and stress-induced plasma corticosterone (CORT) inmales sampled in June, July, and August (n = 31).
P. Deviche et al. / General and Comparative Endocrinology xxx (2014) xxx–xxx 5
determine whether initial and stress-induced plasma CORT wereindividually correlated. This was the case (r2 = 0.25; F1,30 = 9.67,p = 0.004; Fig. 3B). However, in contrast to the situation for plasmaT, the regression line slope (0.966) did not differ from 1 (95% con-fidence interval: 0.331–1.601). Thus, the increase in plasma CORTthat took place during stress was proportionally similar whethersparrows had low or high initial plasma CORT.
Correlations between plasma CORT and T. Considering all thebirds sampled for the Seasonal Study (n = 41), there was norelationship between initial plasma CORT and initial plasma T(Spearman rank order correlation: r = 0.08, p > 0.50). However,stress-induced plasma CORT and stress-induced plasma T werepositively correlated (r = 0.433, p = 0.005).
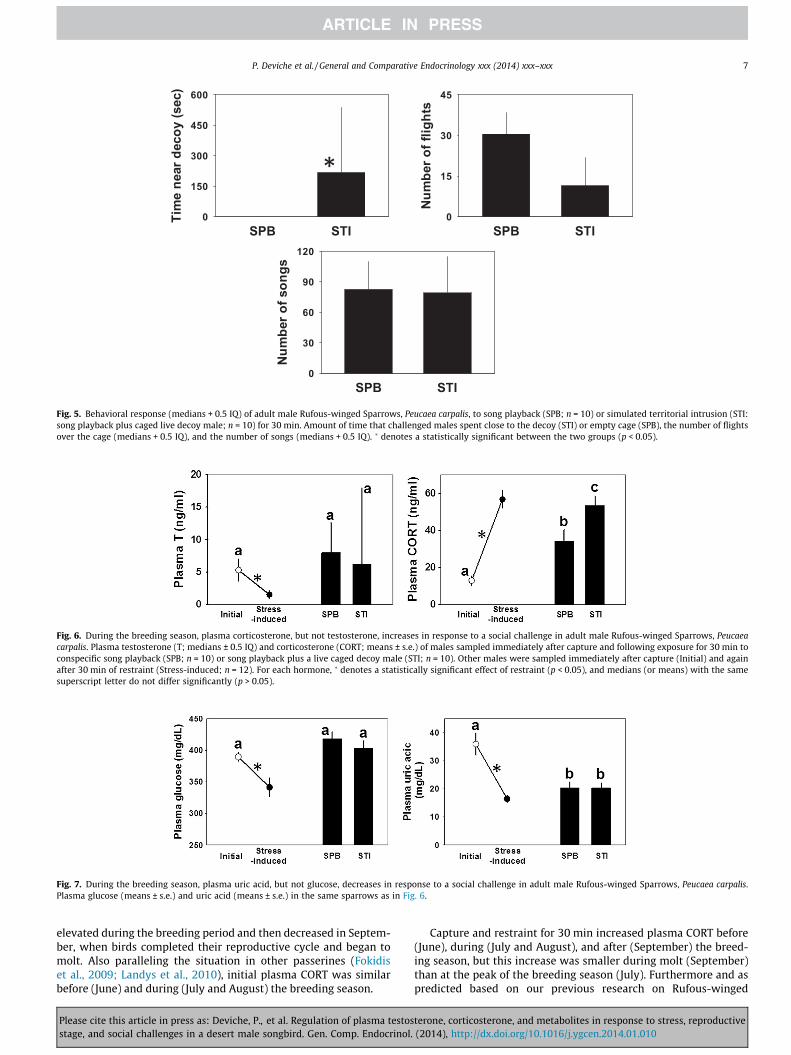
3.1.3. Plasma metabolitesUric acid. Plasma uric acid decreased in response to acute stress
(ANOVA: F1,20 = 41.36, p < 0.001) and changed seasonally(F1,20 = 6.09, p = 0.023), but there was a stress �month interaction(F1,20 = 18.01, p < 0.001; Fig. 4A). At capture, plasma uric acid waslower in September than July. It decreased in response to stressin July but not in September, resulting in sparrows having similarstress-induced plasma uric acid in these two months.
Glucose. Plasma glucose decreased in response to acute stress(ANOVA: F1,19 = 23.59; p < 0.001; Fig. 4B). Birds sampled in Julyand September had similar plasma glucose (two-way repeatedmeasure ANOVA: F1,42 = 1.39, p = 0.25) and there was no month -� stress interaction (F1,19 = 0.85, p > 0.3), indicating that stress af-fected plasma glucose similarly during these two months. Thus,
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosstage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol.
the magnitude of the plasma T and CORT stress response differedbetween July and September (Fig. 2; see above), but this was notthe case for plasma glucose.
3.2. Social challenge study
3.2.1. Capture times and morphologyThe average times of day that birds were caught in July for the
Seasonal Study and that SPB and STI sparrows were caught for theSocial Challenge Study did not differ statistically (KW: H = 0.55,p > 0.7; Table 1). In addition, birds in the three groups were mor-phologically similar (body masses: ANOVA: F2,31 = 0.02, p > 0.9;furcular fat scores: KW: H = 1.51, p > 0.4; cloacal protuberancewidths: ANOVA: F2,31 = 1.31, p > 0.25; wing chords: KW: H = 5.16,p > 0.07; Table 1).
3.2.2. Behavior responses to SPB and STIThere were clear differences in the behavioral responses of
males to speakers broadcasting song in the absence (SPB group)or presence (STI group) of a caged decoy male. Males in the SPBgroup never spent time close to the speakers located next to theempty (control) cage, but birds in the STI group spent, on average,218 s near the cage containing the decoy bird (Mann–Whitneyrank sum test: T = 66, p = 0.001; Fig. 5), and they were often ob-served landing on the top or sides of this cage (personal observa-tion). As a result of spending more time near the cage, males inthe STI group tended to fly fewer times over the cage than SPB
terone, corticosterone, and metabolites in response to stress, reproductive(2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010
Fig. 4. Capture and restraint for 30 min decreases plasma uric acid (July) andglucose (July and September) in adult male Rufous-winged Sparrows, Peucaeacarpalis. Plasma (A) uric acid and (B) glucose (means ± s.e.) of adult males sampledin July (n = 12) and September (uric acid: n = 12; glucose: n = 10) immediately aftercapture (Initial) and then again after 30 min of restraint (Stress-induced). For eachmetabolite, means with the same superscript letter (Initial) or number (Stress-induced) do not differ significantly (p > 0.05), and � denotes a statistically significantdifference between initial and corresponding stress-induced plasma metaboliteconcentrations (p < 0.05).
6 P. Deviche et al. / General and Comparative Endocrinology xxx (2014) xxx–xxx
birds (Mann–Whitney rank sum: T = 131, p = 0.053). The SPB andSTI birds did not differ with respect to the number of songs heardduring the observation period (Mann–Whitney rank sum test:U = 44, p > 0.6).
3.2.3. Plasma hormonesTestosterone. Plasma T of SPB and STI males did not differ from
Initial plasma T (KW: H = 1.75, p > 0.4; Fig. 6), thereby providing noevidence that a social challenge influenced the circulating concen-tration of this hormone.
Corticosterone. Plasma CORT differed between the SPB, STI, andInitial groups (ANOVA: F2,31 = 21.01, p < 0.001; Fig. 6). Plasma CORTincreased in response to a social challenge (SPB and STI groups vs.Initial group: p’s < 0.05) and more so in response to STI than SPB(STI vs. SPB: p < 0.05; Fig. 6). To analyze the effect on plasma CORTof SPB or STI for 30 min vs. that of capture and handling for 30 min,we compared hormone levels of SPB and STI birds with those of
Table 1Capture time and morphological characteristics of adult male Rufous-winged Sparrows, Peufor 30 min to conspecific song playback (SPB) or song playback plus a caged live decoy sp
Experimental gro
CTRL (n = 12)
Capture time (median ± 0.5 IQ) 9:38 AM ± 1:43 hBody mass (g; mean ± s.e.) 15.6 ± 0.1Furcular fat score (median ± 0.5 IQ) 1 ± 0.3Cloacal protuberance width (mm; mean ± s.e.) 4.8 ± 0.2Wing chord (mm; median ± 0.5 IQ) 61 ± 1
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosstage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol.
stressed-induced birds. Plasma CORT differed between thesegroups (ANOVA: F2,31 = 5.35, p = 0.011). Plasma CORT was lowerin SPB than restrained sparrows (p < 0.05) but STI and restrainedbirds had similar levels of this hormone (Fig. 6). Thus, exposureto STI for 30 min increased plasma CORT to the same extent as re-straint for 30 min.
3.2.4. Plasma metabolitesUric acid. Plasma uric acid differed between SPB, STI, and Initial
birds (ANOVA, log10-transformed data: F2,31 = 8.15, p = 0.002;Fig. 7). As pair-wise multiple comparisons show, birds in the SPBand STI groups had lower plasma uric acid than Initial sparrows.However, plasma uric acid did not differ in the SPB, STI, and re-strained birds (KW, H = 4.42, p > 0.10; Fig. 7). Thus, a social chal-lenge for 30 min decreased plasma uric acid to the same extentas restraint for 30 min.
Glucose. Plasma glucose did not differ in the SPB, STI, and Initialbirds (ANOVA: F2,30 = 2.05, p > 0.14; Fig. 7), indicating that a socialchallenge did not mimic the inhibitory effect of acute stress on thismetabolite.
4. Discussion
The present work had three main objectives. First, we character-ized the rapid decrease in plasma T that is associated with acutestress and determined relationships between this decrease and ini-tial and stress-induced plasma CORT. We hypothesized that if plas-ma CORT plays a role in the acute stress-mediated inhibition ofplasma T, changes in plasma CORT would be consistently associ-ated with changes in the opposite direction to plasma T. We testedthis hypothesis using two approaches. According to one approach,we compared initial and stress-induced plasma T and CORT inbirds sampled at the beginning, during, and at the end of theirbreeding season. Following the second approach, we used birdsin breeding condition to measure plasma CORT and T in responseto social challenges. Our second objective was to compare thebehavioral and endocrine responses to visual (SPB exposure) vs. vi-sual plus auditory (STI exposure) social challenge. Most previouswork on this subject used only one type of challenge and littleinformation is, therefore, available on whether birds respond dif-ferently to auditory and visual challenges behaviorally and physi-ologically. Third, given the paucity of information on this topic infree-ranging birds, we identified metabolic changes that take placein response to acute stress and social challenges, and determinedthe extent to which these changes are related to changes in plasmaCORT and T.
4.1. Changes in plasma CORT and T as a function of reproductivecondition, during acute stress, and in response to social challenges
Seasonal changes in initial plasma T and CORT in the presentstudy were generally similar to those reported in other species. Asin previous studies (Deviche et al., 2006; Dawson, 1983; Goymannet al., 2007; Wingfield and Farner 1978), initial plasma T was
caea carpalis, caught in July and sampled immediately after capture (CTRL) or exposurearrow (STI).
up
SPB (n = 10) STI (n = 10)
11:05 AM ± 3:55 h 9:51 AM ± 2:47 h15.6 ± 0.3 15.6 ± 0.31.3 ± 0.6 1 ± 0.35.1 ± 0.2 5.2 ± 0.261 ± 0.3 62 ± 0.3
terone, corticosterone, and metabolites in response to stress, reproductive(2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010
Fig. 5. Behavioral response (medians + 0.5 IQ) of adult male Rufous-winged Sparrows, Peucaea carpalis, to song playback (SPB; n = 10) or simulated territorial intrusion (STI:song playback plus caged live decoy male; n = 10) for 30 min. Amount of time that challenged males spent close to the decoy (STI) or empty cage (SPB), the number of flightsover the cage (medians + 0.5 IQ), and the number of songs (medians + 0.5 IQ). � denotes a statistically significant between the two groups (p < 0.05).
Fig. 6. During the breeding season, plasma corticosterone, but not testosterone, increases in response to a social challenge in adult male Rufous-winged Sparrows, Peucaeacarpalis. Plasma testosterone (T; medians ± 0.5 IQ) and corticosterone (CORT; means ± s.e.) of males sampled immediately after capture and following exposure for 30 min toconspecific song playback (SPB; n = 10) or song playback plus a live caged decoy male (STI; n = 10). Other males were sampled immediately after capture (Initial) and againafter 30 min of restraint (Stress-induced; n = 12). For each hormone, � denotes a statistically significant effect of restraint (p < 0.05), and medians (or means) with the samesuperscript letter do not differ significantly (p > 0.05).
Fig. 7. During the breeding season, plasma uric acid, but not glucose, decreases in response to a social challenge in adult male Rufous-winged Sparrows, Peucaea carpalis.Plasma glucose (means ± s.e.) and uric acid (means ± s.e.) in the same sparrows as in Fig. 6.
P. Deviche et al. / General and Comparative Endocrinology xxx (2014) xxx–xxx 7
elevated during the breeding period and then decreased in Septem-ber, when birds completed their reproductive cycle and began tomolt. Also paralleling the situation in other passerines (Fokidiset al., 2009; Landys et al., 2010), initial plasma CORT was similarbefore (June) and during (July and August) the breeding season.
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosstage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol.
Capture and restraint for 30 min increased plasma CORT before(June), during (July and August), and after (September) the breed-ing season, but this increase was smaller during molt (September)than at the peak of the breeding season (July). Furthermore and aspredicted based on our previous research on Rufous-winged
terone, corticosterone, and metabolites in response to stress, reproductive(2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010
8 P. Deviche et al. / General and Comparative Endocrinology xxx (2014) xxx–xxx
Sparrows and work on other songbirds (Deviche et al., 2010,2012a; Fokidis et al., 2011a,b), acute stress depressed plasma T (ex-cept in the fall; see below). Thus, acute stress influenced plasmaCORT (increase) and T (decrease) in opposite directions. Similarto previous findings (Deviche et al., 2012a; Goutte et al., 2010), ini-tial plasma CORT and T were not correlated at the individual level.However, stress-induced plasma levels of the two hormones werepositively correlated. This correlation suggests a functional rela-tionship between CORT and T during acute stress but analysis ofthe proportional changes in plasma CORT and T relative to theircorresponding initial levels indicates that, if present, this relation-ship is complex. Indeed, the increase in plasma CORT in response tostress, relative to initial plasma CORT, was proportionally similarregardless of initial plasma CORT. By contrast, plasma T decreasedrelatively more during stress when initially high than low. Thisfinding suggests that in stressful situations birds defend againstdecreasing plasma T below a minimum level that is needed formaintenance of T-dependent behavioral, morphological (secondarysexual characteristics), and/or physiological (negative feedback ongonadotropin secretion) functions. Further research is necessary toevaluate the validity of this interpretation but we note that it isconsistent with our observation that the relative magnitude ofthe stress-associated decrease in plasma T varies seasonally: stressexposure decreased plasma T when it was initially high (June–Au-gust) but not in September, when it was lower than during thesummer.
To further test whether there is a functional inverse relationshipbetween plasma CORT and T, we studied this relationship in birdsexposed to social challenges. Exposure to SPB or STI increased plas-ma CORT, and STI was more effective in this respect than SPB. Infact, plasma CORT was similar after 30 min of restraint and afterexposure to STI for the same duration, indicating that a social chal-lenge is physiologically stressful. By contrast, neither SPB nor STIaffected plasma T. Thus, the increase in plasma CORT during a so-cial challenge was not associated with a decrease in plasma T. Con-sidering two situations in which plasma CORT was elevated (acutestress and social challenges) the present findings do, therefore, notsupport the hypothesis that a short-term increase in plasma CORTcauses a decrease in plasma T.
In mammals, glucocorticoids influence T production via non-genomic inhibition of interstitial (Leydig) testicular cell function(Hardy et al., 2005; Hu et al., 2008). Avian testes contain glucocor-ticoid receptors (Kwok et al., 2007), but it is unknown in birdswhether glucocorticoids decrease plasma T through a similarmechanism as in mammals. Our findings that plasma CORT in-creases during acute stress and social challenges, whereas plasmaT decreases in response to acute stress but not social challenges,provides no support for a direct effect of glucocorticoids on testic-ular function. It also provides no support for the alternative mech-anism in which the stress-induced increase in CORT ishypothesized to cause displacement of T from plasma CORT-bind-ing globulin, thereby potentially accelerating the clearance of plas-ma T (Deviche et al., 2012a). Instead, our findings suggest that themechanism responsible for the rapid decrease in plasma T ob-served during acute stress is independent of glucocorticoids. Thisconclusion does not preclude direct inhibitory effects of acutestress on testicular endocrine function. Such effects may involvea role for gonadotropin-inhibitory hormone (GnIH; McGuire andBentley 2010) and/or follow activation of a sympathetic nervouspathway terminating in the gonads as demonstrated in rodents(Herman and Rivier 2006; James et al., 2008). This pathway is iden-tified in birds (Wakita et al., 1999), but its function in these verte-brates has not been evaluated.
Consistent with previous studies (Deviche et al., 2006, 2012b),we found that exposure to SPB does not affect plasma T. In addi-tion, in the male Rufous-winged Sparrow and other male
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosstage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol.
songbirds there is no relationship between plasma T and theduration of SPB to which birds are exposed immediately beforecapture (see below). Thus, in these species, auditory stimulationalone does not influence plasma T. However, we also found no ef-fect of STI, which combines auditory and visual stimulation, onplasma T and this result contrasts with those of the only otherstudy comparing plasma T in response to components of STI. Inthis study on Song Sparrows, plasma T increased in response toSTI but not in response either to visual or to auditory stimulationalone (Wingfield and Wada, 1989). Birds in the present studywere socially challenged during their breeding season. If at thistime their reproductive endocrine axis was secreting plasma Tat a seasonally maximum rate, a social challenge would not beexpected to further elevate the secretion of this hormone. Thiswas, however, probably not the case because administration ofgonadotropin-releasing hormone (GnRH) to Rufous-winged Spar-rows during the breeding season rapidly stimulates the release ofLH, and an injection of this hormone rapidly stimulates the re-lease of T (Deviche et al., 2010). Similar findings have been re-ported in other species (Hirschenhauser et al., 2000; Jaworet al., 2006; Lacombe et al., 1991). Consistent with the above con-clusion, studies conducted during stages of the breeding cyclewhen plasma T is not at its seasonal maximum likewise foundno plasma T response to STI (reviewed in Goymann, 2009).
The challenge hypothesis predicts that the T response to socialchallenge varies as a function of the breeding and mating systems(Wingfield et al., 1990). Specifically, males of multi-brooded spe-cies are predicted to respond to social challenges with increasedT whereas males of single-brooded species are predicted to be lessor not responsive to this challenge (Goymann et al., 2007; Landyset al., 2007). Rufous-winged Sparrows are multi-brooded (Austinand Ricklefs, 1977; Lowther et al., 1999) and were, therefore, antic-ipated to increase their plasma T when socially challenged duringthe breeding period. That this was not the case may reflect the factthat these desert-adapted birds are constrained in a similar way tomany single-brooded species due to their relatively short, mon-soon-dependent breeding season.
The behavioral response of Rufous-winged Sparrows to socialchallenges was context-dependent: males exposed to STI spentmore time near the decoy-containing cage that we placed withintheir territory and tended to fly fewer times over this cage thanmales exposed to SPB and an empty cage. These differences alsowere behavior-specific, as sparrows in the two experimentalgroups sang at similar rates during the experimental challenge.As no change in plasma T was associated with the behavioral ef-fects of STI, we conclude that plasma T does not determine theseeffects. Congruent with this conclusion, in the congeneric maleCassin’s Sparrow, exposure to SPB increased the expression ofmale-typical behavior without affecting plasma T (Deviche et al.,2012b). Furthermore, in a recent comparative study on severalsongbird species, urban birds exhibited more territorial behaviorthan their non-urban counterparts but this difference was notassociated with variation in plasma T (Fokidis et al., 2011a). In thiscomparative study as well as in Rufous-winged Sparrows (Devicheet al., 2006), there also was no relationship at the individual levelbetween the duration of exposure to SPB and plasma T. Taken col-lectively, these results indicate that short-term changes in theexpression of aggressive and territorial behavior in response to so-cial challenge are likely plasma T-independent. Additional supportfor this hypothesis comes from avian research showing that differ-ent intensities and frequencies of STIs cause different changes inbehavior that are not associated with a change in plasma T (Apfel-beck and Goymann, 2011), individual differences in behavior arenot correlated with individual plasma T levels (Silverin et al.,2004), and pharmacological inhibition of T does not producechanges in aggressive behavior in all species (Moore et al., 2004).
terone, corticosterone, and metabolites in response to stress, reproductive(2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010
P. Deviche et al. / General and Comparative Endocrinology xxx (2014) xxx–xxx 9
Plasma CORT increased in response to both STI and SPB, butmore so in response to STI. Other studies also found that plasmaCORT increases in response to social challenge (Charlier et al.,2009; Gill et al., 2008; Landys et al., 2007, 2010; Van Duyseet al., 2004). This increase may serve to coordinate responses to aphysical territorial intrusion (caged live decoy) through increasedmetabolism (see below), activity (Breuner et al., 1998), and aggres-sive behavior (Mikics et al., 2004). Evidence suggests that the neu-ral circuits that control aggression and the response to stressoverlap (Summers and Winberg, 2006). A functional link betweenplasma CORT and short-term changes in aggressive behavior re-quires further investigation because individual differences inexpression of this behavior are not necessarily associated withindividual variation in plasma CORT (Fokidis et al., 2011b). Thepresent results are nevertheless consistent with the idea that CORTplays a role in the regulation of short-term behavioral changesaccompanying social challenge.
4.2. Plasma glucose and uric acid
Little information is available regarding the effects of acutestress on plasma glucose in free-ranging birds and the results ofstudies on this subject are equivocal. Capture and restraint for60 min increased plasma glucose in the Abert’s Towhee (Davieset al., 2013) but this manipulation was ineffective in the Curve-billed Thrasher, Toxostoma curvirostre (Fokidis et al., 2011a,b) andHouse Sparrow (M. Romero, personal communication). Here wefound that acute stress decreased plasma glucose but this decreasewas not consistently related to changes in plasma CORT or T. In-deed, initial plasma glucose was similar during and outside thebreeding season, and plasma glucose decreased similarly in re-sponse to capture and restraint in breeding condition (July) andmolting (September) birds. However, plasma CORT increased moreduring stress in July than September and in addition, the socialchallenge-induced increase in plasma CORT was not accompaniedwith a change in glycemia. Finally, plasma T was higher in summer,when it decreased during stress, than in the fall, when not influ-enced by stress. These results provide no evidence for direct,short-term regulation of plasma glucose by either hormone. Inother vertebrates, the stimulating effect of acute stress on plasmaglucocorticoids often concurs with elevated glycemia (Carrhagerand Rees, 1993; Curi et al., 1990), but the contribution of glucocor-ticoids to this elevation remains unclear. For example, Corbel et al.(2010) found no association in penguins between plasma CORT andthe metabolic response (including plasma glucose) to stress. And incaptive European Starlings, CORT administration increased glyce-mia; however, this treatment resulted in supra-physiological plas-ma CORT and its effects are, therefore, difficult to interpret(Remage-Healey and Romero, 2002). The relationship betweenplasma T and glycemia is likewise unsettled. An injection of Tenanthate to laying hens increased their plasma glucose eighthours later (Mohammadi and Ansari-Pirsaraei, 2013). However,due to species, sex, body size, and methodological differences, thisfinding bears little relationship to the present study. It should alsobe pointed out that glycemia is regulated differently in birds andmammals (Braun and Sweazea, 2008) and, consequently, conclu-sions drawn from mammalian studies may not readily apply toavian models.
It cannot be excluded that the stress-associated decrease in gly-cemia in the Rufous-winged Sparrow was due to food deprivationduring the 30 min long period of restraint. In larger species (turkey,chicken, Yellow-legged Gull, Larus cachinnans, and raptors) onlyprolonged fasting (up to several days) normally decreases plasmaglucose (Alonso-Alvarez and Ferrer, 2001; Anthony et al., 1990;Belo et al., 1976; Minick, 1986). In a smaller species (House Spar-row), however, glycemia begins to decrease after a few hours of
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosstage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol.
fasting (Khalilieh et al., 2012). The body mass of Rufous-wingedSparrows (average: 15.6 g) is almost half that of House Sparrows(average: 28 g) and two or more orders of magnitude less thanthose of some of the above species. As a result, Rufous-wingedSparrows have a higher metabolic rate and energy expenditurethan these species (Kleiber’s law). Coupled with the fact that spar-rows carry relatively low fat reserves, this may result in these birdsrelying more on carbohydrates as a source of metabolic energyduring the initial stage of fasting than is the case in larger species.This could be the case particularly during the breeding season andduring molt because at these times, birds may benefit from usingcarbohydrates as a source of energy to protect against stress-re-lated proteolysis. Consistent with this proposition, plasma uric aciddid not increase in response to capture and restraint (the opposite,in fact, was observed), suggesting that this experimental protocoldid not cause substantial protein degradation. These hypothesesrequire further investigation. Indeed, we measured plasma glucosebefore and then only once (30 min) during stress and so do nothave information about the time course of stress-induced changesin glycemia after capture. In addition, glycemia at any given timereflects the balance between glucose secretion, utilization, and/orexcretion, which we did not determine. Finally, as plasma glucosein small free-ranging birds during stress or other experimental sit-uations is rarely measured, few studies are available for compari-son with the present work.
In the Rufous-winged Sparrow as in some other species (Cohenet al., 2007; Davies et al., 2013), plasma uric acid decreased in re-sponse to acute stress. However, this effect was reproductivestage-dependent: we observed this decrease in July, when birds re-sponded to acute stress by markedly elevating their plasma CORT,but not in September, when plasma CORT increased less duringstress than during summer. We also found that plasma uric aciddecreased during social challenge, when plasma CORT increased.Thus, in two independent experimental situations, an elevationin plasma CORT correlated with a decrease in plasma uric acid. Thiscorrelation suggests that elevated CORT may normally decreaseplasma uric acid, but further research is needed to test whetherthis is the case. Indeed, the relationship between plasma CORTand uric acid is not consistent across species (Cohen et al., 2008).Corticosterone administration to chickens increases their plasmauric acid (Lin et al., 2004) but Abert’s Towhees are impervious tothis treatment (Davies et al., 2013). Furthermore, acute stress in-creases, has no influence, or decreases plasma uric acid dependingon the species (Cohen et al., 2007). Uric acid is a potent antioxidant(Stinefelt et al., 2005) and as birds generally have higher glycemiathan mammals, in these vertebrates it may play a particularlyimportant protective function to ameliorate oxidative stress (Braunand Sweazea, 2008). Increased tissue uptake of uric acid duringstress could be especially beneficial if, as discussed above, acutestress were associated with increased uptake and utilization of glu-cose as a source of metabolic energy. However, this hypothesisdoes not account for the fact that plasma uric acid and glucose de-creased during stress in breeding birds, but only plasma glucosedecreased during molt. Further research, especially using smallspecies with a high metabolic rate and rapid metabolite turnoverrate, is warranted to assess these ideas and, in particular, to deter-mine functional relationships between glucocorticoids, glycemia,and uric acid as a function of the life history stage.
4.3. Plasma CORT and molt
Consistent with previous observations (Small et al., 2007), nei-ther the visible fat reserves nor the body mass of the study birdschanged between June and August, suggesting that males main-tained stable body condition during this period. Rufous-wingedSparrows breed, for the most part, at a time of the year when
terone, corticosterone, and metabolites in response to stress, reproductive(2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010
10 P. Deviche et al. / General and Comparative Endocrinology xxx (2014) xxx–xxx
energy-rich food resources (arthropods) become abundant. Inthese birds, enhanced use of readily available food resources dur-ing the breeding period conceivably compensates for heightenedenergy demand associated with reproduction, thereby alleviatingthe need to increase baseline plasma CORT to meet energy require-ments. Another life history stage of birds that is particularly en-ergy-demanding is the molt period, when large amounts ofproteins are synthesized and some of these proteins are incorpo-rated into new feathers (Cyr et al., 2008; Dlonik and Gavrilov,1979). Glucocorticoids promote proteolysis and baseline plasmaCORT is correspondingly often lower during molt than at othertimes (Cyr et al., 2008; Romero, 2002; Romero et al., 1998a). Wedid not observe this relationship in Rufous-winged Sparrows,which had similar initial plasma CORT prior to (June–August) andduring (September) molt. However, the magnitude of the CORT re-sponse to stress was lower during molt than at the peak of thebreeding season (July) and this result is consistent with previousresearch showing a suppression of the CORT stress response duringmolt (see below). An attenuated CORT response to stress duringmolt relative to the breeding period is commonly seen in birds(Cyr et al., 2008; Romero and Remage-Healey, 2000; Romeroet al., 1998b) and may protect against the proteolytic effect of thishormone. Supporting this proposition, CORT administration inhib-its feather replacement in the European Starling (Romero et al.,2005). However, whether CORT has the same effect in intact birdsremains conjectural (Strochlic and Romero, 2008). Thus, our resultsshowing a seasonal modulation of the stress response are consis-tent with others, but the significance of this modulation in rela-tionship to molt warrants further investigation.
5. Conclusions
Previous studies in male vertebrates demonstrated that T andCORT secretion depends on multiple factors that include the repro-ductive stage, whether an organism is or not exposed to stress, andsocial interactions between conspecifics. We measured the circu-lating concentrations of these hormones as a function of thesethree factors in free-ranging male birds and, confirming previousresearch, found that they can, indeed, modulate plasma T andCORT. Specifically, initial plasma T decreased between the breedingand non-breeding seasons but this was not the case for initial plas-ma CORT. Furthermore, plasma CORT and T rapidly increased anddecreased, respectively, in response to acute stress, but the magni-tude of these changes was smaller outside than during the breed-ing season, showing that the T and CORT responses to acute stressare seasonally modulated. When considered with the observationthat social challenges increased plasma CORT and this increasewas not associated with a change in plasma T, these results suggestthat rapid inhibitory effects of acute stress on plasma T are not di-rectly due to elevated plasma CORT, but rather result from a plas-ma glucocorticoid-independent mechanism. The nature of thismechanism and its role in the regulation of T secretion not onlyin challenged but potentially, also in unchallenged organisms, war-rant further investigation.
The present investigation has implications for our understand-ing of the endocrine mechanisms that control aggressive responsesto social challenge. We found STI to induce these responses moreeffectively than does SPB, indicating an important role in this re-spect of visual stimulation by the challenging individual. However,plasma T was not influenced by exposure to SPB or STI. Thus, theeffects of a social challenge on aggressive responses do not appearto depend on elevated plasma T, and the present study provides noevidence for rapid behavioral effects of the steroid. Whether plas-ma CORT, which increased during this challenge, plays a role in the
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosstage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol.
control of the behavioral response to social challenge is an impor-tant question that future research needs to address.
Finally, the present work extends previous research on the con-trol of glycemia and plasma uric acid in free-ranging birds, a topicthat has until now been the object of limited research and hasyielded conflicting results. Elevated plasma CORT, whether result-ing from social challenge or induced by acute stress, was not asso-ciated with elevated plasma glucose – in fact, the oppositesituation was observed in acutely stressed birds. This finding leadsto question the frequent assumption that elevated plasma CORTpromotes glycemia and it confirms the need to further investigatethe endocrine control of carbohydrate metabolism in free-rangingbirds, particularly in relationship to body size and reproductivecondition. Our observation that elevated plasma CORT during acutestress and social challenge parallels a decrease in plasma uric acidis consistent with some previous work on this subject in otheravian species. However, whether and how CORT normally regu-lates plasma uric acid is not known, and the physiological signifi-cance of rapid changes in plasma uric acid as observed duringstress and social challenge remains speculative. This topic likewisewarrants additional research.
Acknowledgments
The research was supported by National Science FoundationAward IOB 1026620 to PD. We are thankful to the managers ofthe Santa Rita Experimental Range for access to field sites.
References
Alonso-Alvarez, C., Ferrer, M., 2001. A biochemical study of fasting, subfeeding, andrecovery processes in yellow-legged gulls. Physiol. Biochem. Zool. 74, 703–713.
Anthony, N.B., Vasilatos-Younken, R., Bacon, W.L., Lilburn, M.S., 1990. Secretorypattern of growth hormone, insulin, and related metabolites in growing maleturkeys: effects of overnight fasting and refeeding. Poult. Sci. 69, 801–811.
Apfelbeck, B., Goymann, W., 2011. Ignoring the challenge? Male black redstarts(Phoenicurus ochruros) do not increase testosterone levels during territorialconflicts but they do so in response to gonadotropin-releasing hormone. Proc.Biol. Sci. 278, 3233–3242.
Austin, T., Ricklefs, E., 1977. Growth and development of the Rufous-wingedSparrow (Aimophila carpalis). Condor 79, 37–50.
Belo, P.S., Romsos, D.R., Leville, G.A., 1976. Blood metabolites and glucosemetabolism in the fed and fasted chicken. J. Nutr. 106, 1135–1143.
Breuner, C.W., Orchinik, M., 2001. Seasonal regulation of membrane andintracellular corticosteroid receptors in the house sparrow brain. J.Neuroendocrinol. 13, 412–420.
Charlier, T.D., Underhill, C., Hammond, G.L., Soma, K.K., 2009. Effects of aggressiveencounters on plasma corticosteroid-binding globulin and its ligands in white-crowned sparrows. Horm. Behav. 56, 339–347.
Cohen, A., Klasing, K., Ricklefs, R., 2007. Measuring circulating antioxidants in wildbirds. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 147, 110–121.
Cohen, A.A., Hau, M., Wikelski, M., 2008. Stress, metabolism, and antioxidants intwo wild passerine bird species. Physiol. Biochem. Zool. 81, 463–472.
Conover, W.J., Iman, R.L., 1981. Rank-transformations as a bridge betweenparametric and nonparametric statistics. Am. Stat. 35, 124–129.
Corbel, H., Geiger, S., Groscolas, R., 2010. Preparing to fledge: the adrenocortical andmetabolic responses to stress in king penguin chicks. Funct. Ecol. 24, 82–92.
Costantini, D., 2008. Oxidative stress in ecology and evolution: lessons from avianstudies. Ecol. Lett. 11, 1238–1251.
Curi, C.M., Ribeiro, E.B., Zaia, C.T., Dolnikoff, M.S., 1990. Glycemic response to stressstimulation by ether exposure in adrenalectomized rats. Pharmacol. Biochem.Behav. 37, 399–403.
Cyr, N.E., Wikelski, M., Romero, L.M., 2008. Increased energy expenditure butdecreased stress responsiveness during molt. Physiol. Biochem. Zool. 81, 452–462.
Davies, S., Rodriguez, N.S., Sweazea, K.L., Deviche, P., 2013. The effect of acute stressand long-term corticosteroid administration on plasma metabolites in an urbanand desert songbird. Physiol. Biochem. Zool. 86, 47–60.
terone, corticosterone, and metabolites in response to stress, reproductive(2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010
P. Deviche et al. / General and Comparative Endocrinology xxx (2014) xxx–xxx 11
Dawson, A., 1983. Plasma gonadal steroid levels in wild starlings (Sturnus vulgaris)during the annual cycle and in relation to the stages of breeding. Gen. Comp.Endocrinol. 49, 286–294.
Deviche, P., Small, T., Sharp, P., Tsutsui, K., 2006. Control of luteinizing hormone andtestosterone secretion in a flexibly breeding male passerine, the Rufous-wingedSparrow, Aimophila carpalis. Gen. Comp. Endocrinol. 149, 226–235.
Deviche, P.J., Hurley, L.L., Fokidis, H.B., Lerbour, B., Silverin, B., Silverin, B., Sabo, J.,Sharp, P.J., 2010. Acute stress rapidly decreases plasma testosterone in a free-ranging male songbird: potential site of action and mechanism. Gen. Comp.Endocrinol. 169, 82–90.
Deviche, P., Gao, S., Davies, S., Sharp, P.J., Dawson, A., 2012a. Rapid stress-inducedinhibition of plasma testosterone in free-ranging male rufous-winged sparrows,Peucaea carpalis: Characterization, time course, and recovery. Gen. Comp.Endocrinol. 117, 1–8.
Deviche, P., Sharp, P.J., Dawson, A., Sabo, J., Fokidis, B., Davies, S., Hurley, L., 2012b.Up to the challenge? Hormonal and behavioral responses of free-ranging maleCassin’s sparrows, Peucaea cassinii, to conspecific song playback. Horm. Behav.61, 741–749.
Dlonik, V.R., Gavrilov, V.M., 1979. Bioenergetics of molt in the Chaffinch (Fringillacoelebs). Auk 96, 253–264.
Dong, Q., Salva, A., Sottas, C.M., Niu, E., Holmes, M., Hardy, M.P., 2004. Rapidglucocorticoid mediation of suppressed testosterone biosynthesis in male micesubjected to immobilization stress. J. Androl. 25, 973–981.
Fokidis, H.B., Orchinik, M., Deviche, P., 2009. Corticosterone and corticosteroidbinding globulin in birds: relation to urbanization in a desert city. Gen. Comp.Endocrinol. 160, 259–270.
Fokidis, H.B., Hurley, L., Rogowski, C., Sweazea, K., Deviche, P., 2011a. Effects ofcaptivity and body condition on plasma corticosterone, locomotor behavior, andplasma metabolites in curve-billed thrashers. Physiol. Biochem. Zool. 84, 595–606.
Fokidis, H.B., Orchinik, M., Deviche, P., 2011b. Context-specific territorial behaviorin urban birds: no evidence for involvement of testosterone or corticosterone.Horm. Behav. 59, 133–143.
Gill, S.A., Costa, L.M., Hau, M., 2008. Males of a single-brooded tropical bird speciesdo not show increases in testosterone during social challenges. Horm. Behav.54, 115–124.
Goutte, A., Angelier, F., Chastel, C.C., Trouve, C., Moe, B., Bech, C., Gabrielsen, G.W.,Chastel, O., 2010. Stress and the timing of breeding: glucocorticoid-luteinizinghormones relationships in an arctic seabird. Gen. Comp. Endocrinol. 169, 108–116.
Goymann, W., 2009. Social modulation of androgens in male birds. Gen. Comp.Endocrinol. 163, 149–157.
Goymann, W., Landys, M.M., Wingfield, J.C., 2007. Distinguishing seasonal androgenresponses from male-male androgen responsiveness-revisiting the ChallengeHypothesis. Horm. Behav. 51, 463–476.
Hardy, M.P., Gao, H.B., Dong, Q., Ge, R., Wang, Q., Chai, W.R., Feng, X., Sottas, C., 2005.Stress hormone and male reproductive function. Cell Tissue Res. 322, 147–153.
Hau, M., Beebe, K., 2011. Plastic endocrine regulation of year-round territorialaggression in tropical male spotted antbirds. Gen. Comp. Endocrinol. 172, 305–313.
Herman, M., Rivier, C., 2006. Activation of a neural brain-testicular pathway rapidlylowers Leydig cell levels of the steroidogenic acute regulatory protein and theperipheral-type benzodiazepine receptor while increasing levels of neuronalnitric oxide synthase. Endocrinology 147, 624–633.
Hirschenhauser, K., Mostl, E., Peczely, P., Wallner, B., Dittami, J., Kotrschal, K., 2000.Seasonal relationships between plasma and fecal testosterone in response toGnRH in domestic ganders. Gen. Comp. Endocrinol. 118, 262–272.
Hirschenhauser, K., Winkler, H., Oliveira, R.F., 2003. Comparative analysis of maleandrogen responsiveness to social environment in birds: the effects of matingsystem and paternal incubation. Horm. Behav. 43, 508–519.
Hu, G.X., Lian, Q.Q., Lin, H., Latif, S.A., Morris, D.J., Hardy, M.P., Ge, R.S., 2008. Rapidmechanisms of glucocorticoid signaling in the Leydig cell. Steroids 73, 1018–1024.
James, P., Rivier, C., Lee, S., 2008. Presence of corticotrophin-releasing factor and/ortyrosine hydroxylase in cells of a neural brain-testicular pathway that arelabelled by a transganglionic tracer. J. Neuroendocrinol. 20, 173–181.
Jawor, J.M., McGlothlin, J.W., Casto, J.M., Greives, T.J., Snajdr, E.A., Bentley, G.E.,Ketterson, E.D., 2006. Seasonal and individual variation in response to GnRHchallenge in male dark-eyed juncos (Junco hyemalis). Gen. Comp. Endocrinol.149, 182–189.
John-Alder, H.B., Cox, R.M., Haenel, G.J., Smith, L.C., 2009. Hormones, performanceand fitness: Natural history and endocrine experiments on a lizard (Sceloporusundulatus). Integr. Comp. Biol. 49, 393–407.
Khalilieh, A., McCue, M.D., Pinshow, B., 2012. Physiological responses to fooddeprivation in the house sparrow, a species not adapted to prolonged fasting.Am. J. Physiol. Regul. Integr. Comp. Physiol. 303, R551–R561.
Klukowski, M., 2011. Effects of breeding season, testosterone and ACTH on thecorticosterone response of free-ranging male fence lizards (Sceloporusundulatus). Gen. Comp. Endocrinol. 173, 295–302.
Kwok, A.H., Wang, Y., Wang, C.Y., Leung, F.C., 2007. Cloning of chickenglucocorticoid receptor (GR) and characterization of its expression in pituitaryand extrapituitary tissues. Poult. Sci. 86, 423–430.
Lacombe, D., Cyr, A., Matton, P., 1991. Plasma LH and androgen levels in the red-winged blackbird (Agelaius phoeniceus) treated with a potent GnRH analogue.Comp. Biochem. Physiol. A Comp. Physiol. 99, 603–607.
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosstage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol.
Landys, M.M., Goymann, W., Raess, M., Slagsvold, T., 2007. Hormonal responses tomale–male social challenge in the blue tit Cyanistes caeruleus: single-broodedness as an explanatory variable. Physiol. Biochem. Zool. 80, 228–240.
Landys, M.M., Goymann, W., Schwabl, I., Trapschuh, M., Slagsvold, T., 2010. Impactof season and social challenge on testosterone and corticosterone levels in ayear-round territorial bird. Horm. Behav. 58, 317–325.
Lin, H., Decuypere, E., Buyse, J., 2004. Oxidative stress induced by corticosteroneadministration in broiler chickens (Gallus gallus domesticus) 1. Chronicexposure. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 139, 737–744.
Lowther, P.W., Groschupf, K.D., Russell, S.M., 1999. Rufous-winged Sparrow(Aimophila carpalis). In: Poole, A., Gill, F. (Eds.), The Birds of North America,No. 422. The Birds of North America Inc., Philadelphia, PA.
Martin, L.J., Tremblay, J.J., 2008. Glucocorticoids antagonize cAMP-induced Startranscription in Leydig cells through the orphan nuclear receptor NR4A1. J. Mol.Endocrinol. 41, 165–175.
McGuire, N.L., Bentley, G.E., 2010. A functional neuropeptide system in vertebrategonads: Gonadotropin-inhibitory hormone and its receptor in testes of field-caught house sparrow (Passer domesticus). Gen. Comp. Endocrinol. 166, 565–572.
Mikics, E., Kruk, M.R., Haller, J., 2004. Genomic and non-genomic effects ofglucocorticoids on aggressive behavior in male rats.Psychoneuroendocrinology 29, 618–635.
Minick, M.C., 1986. Correlation of simultaneously determined plasma insulin andglucose levels in fasting and mildly or severely stressed raptors. Comp.Biochem. Physiol. A Comp. Physiol. 83, 301–305.
Mohammadi, H., Ansari-Pirsaraei, Z., 2013. Changes in some blood parameters andproduction performance of old laying hens due to growth hormone andtestosterone injection. J. Anim. Physiol. Anim. Nutr. (Berl.), http://dx.doi.org/10.1111/jpn.12095.
Moore, I.T., Walker, B.G., Wingfield, J.C., 2004. The effects of combinedaromatase inhibitor and anti-androgen on male territorial aggression in atropical population of rufous-collared sparrows, Zonotrichia capensis. Gen.Comp. Endocrinol. 135, 223–229.
Pickering, A.D., Pottinger, T.G., Carragher, J., Sumpter, J.P., 1987. The effects of acuteand chronic stress on the levels of reproductive hormones in the plasma ofmature male brown trout, Salmo trutta L.. Gen. Comp. Endocrinol. 68, 249–259.
Place, N.J., Kenagy, G.J., 2000. Seasonal changes in plasma testosterone andglucocorticosteroids in free-living male yellow-pine chipmunks and theresponse to capture and handling. J. Comp. Physiol. B 170, 245–251.
Remage-Healey, L., Romero, L.M., 2000. Daily and seasonal variation in response tostress in captive starlings (Sturnus vulgaris): glucose. Gen. Comp. Endocrinol.119, 60–68.
Remage-Healey, L., Romero, L.M., 2001. Corticosterone and insulin interact toregulate glucose and triglyceride levels during stress in a bird. Am. J. Physiol.Regul. Integr. Comp. Physiol. 281, R994–R1003.
Remage-Healey, L., Romero, L.M., 2002. Corticosterone and insulin interact toregulate plasma glucose but not lipid concentrations in molting starlings. Gen.Comp. Endocrinol. 129, 88–94.
Rogers, C.M., 1991. An evaluation of the method of estimating body fat in birds byquantifying visible subcutaneous fat. J. Field Ornithol. 62, 349–356.
Romero, L.M., 2002. Seasonal changes in plasma glucocorticoid concentrations infree-living vertebrates. Gen. Comp. Endocrinol. 128, 1–24.
Romero, L.M., Remage-Healey, L., 2000. Daily and seasonal variation in response tostress in captive starlings (Sturnus vulgaris): corticosterone. Gen. Comp.Endocrinol. 119, 52–59.
Romero, L.M., Ramenofsky, M., Wingfield, J.C., 1997. Season and migrationalters the corticosterone response to capture and handling in an Arcticmigrant, the white-crowned sparrow (Zonotrichia leucophrys gambelii).Comp. Biochem. Physiol. C: Comp. Pharmacol. Toxicol. Endocrinol. 116C,171–177.
Romero, L.M., Soma, K.K., Wingfield, J.C., 1998a. Changes in pituitary and adrenalsensitivities allow the snow bunting (Plectrophenax nivalis), an Arctic-breedingsong bird, to modulate corticosterone release seasonally. J. Comp. Physiol. B168, 353–358.
Romero, L.M, Soma, K.K, Wingfield, J.C., 1998b. Hypothalamic–pituitary–adrenalaxis changes allow seasonal modulation of corticosterone in a bird. Am. J.Physiol.: Reg. Integr. Comp. Physiol. 274, R1338–R1344.
Romero, L.M., Strochlic, D., Wingfield, J.C., 2005. Corticosterone inhibits feathergrowth: potential mechanism explaining seasonal down regulation ofcorticosterone during molt. Comp. Biochem. Physiol. A Mol. Integr. Physiol.142, 65–73.
12 P. Deviche et al. / General and Comparative Endocrinology xxx (2014) xxx–xxx
Stinefelt, B., Leonard, S.S., Blemings, K.P., Shi, X., Klandorf, H., 2005. Free radicalscavenging, DNA protection, and inhibition of lipid peroxidation mediated byuric acid. Ann. Clin. Lab. Sci. 35, 37–45.
Strochlic, D.E., Romero, L.M., 2008. The effects of chronic psychological and physicalstress on feather replacement in European starlings (Sturnus vulgaris). Comp.Biochem. Physiol. A Mol. Integr. Physiol. 149, 68–79.
Summers, C.H., Winberg, S., 2006. Interactions between the neural regulation ofstress and aggression. J. Exp. Biol. 209, 4581–4589.
Tsahar, E., Arad, Z., Izhaki, I., Guglielmo, C.G., 2006. The relationship between uricacid and its oxidative product allantoin: a potential indicator for the evaluationof oxidative stress in birds. J. Comp. Physiol. B 176, 653–661.
Van Duyse, D.E., Pinxten, R., Eens, M., 2003. Seasonal fluctuations in plasmatestosterone levels and diurnal song activity in free-living male great tits. Gen.Comp. Endocrinol. 134, 1–9.
Van Duyse, E., Pinxten, R., Darras, V.M., Arckens, L., Eens, M., 2004. Opposite changesin plasma testosterone and corticosterone levels following a simulatedterritorial challenge in great tits. Behaviour 141, 451–467.
Wakita, T., Ohmori, Y., Watanabe, T., Fukuta, K., 1999. Location of sympatheticpostganglionic and sensory neurons innervating the testis in the male chicken.Anat. Histol. Embryol. 28, 299–302.
Wikelski, M., Hau, M., Wingfield, J., 1999. Social instability increases plasmatestosterone in a year-round territorial neotropical bird. Proc. R. Soc. London B22, 551–556.
Wingfield, J.C., 1994. Regulation of territorial behavior in the sedentary songsparrow, Melospiza melodia morphna. Horm. Behav. 28, 1–15.
Please cite this article in press as: Deviche, P., et al. Regulation of plasma testosstage, and social challenges in a desert male songbird. Gen. Comp. Endocrinol.
Wingfield, J.C., Farner, D.S., 1978. The endocrinology of a natural breedingpopulation of the White-crowned Sparrow (Zonotrichia leucophrys pugetensis).Physiol. Zool. 51, 188–205.
Wingfield, J.C., Kitaysky, A.S., 2002. Endocrine responses to unpredictableenvironmental events: stress or anti-stress hormones? Integr. Comp. Biol. 42,600–609.
Wingfield, J.C., Wada, M., 1989. Changes in plasma levels of testosterone duringmale-male interactions in the Song Sparrow, Melospiza melodia: time courseand specificity of response. J. Comp. Phyiol. A: Sensory, Neural Behav. Physiol.166, 189–194.
Wingfield, J.C., Smith, M.J., Farner, D.S., 1982. Endocrine responses of White-crowned Sparrows to environmental stress. Condor 84, 399–409.
Wingfield, J., Ball, G.F., Dufty, A.M., Hegner, R.E., Ramenofsky, M., 1987. Testosteroneand aggression in birds. Am. Sci. 75, 602–608.
Wingfield, J.C., Hegner, R.E., Dufty, A.M., Ball, G.F., 1990. The ‘‘ChallengeHypothesis’’: theoretical implications for patterns of testosterone secretion,mating systems, and breeding strategies. Am. Nat. 136, 829–846.
Wingfield, J., Maney, D., Breuner, C., Jacobs, J.D., Lynn, S., Ramenofsky, M.,Richardson, R.D., 1998. Ecological basis of hormone-behavior interactions:The ‘‘emergency life history stage’’. Integr. Comp. Biol. 38, 191–206.
Woodley, S.K., Lacy, E.L., 2010. An acute stressor alters steroid hormone levels andactivity but not sexual behavior in male and female Ocoee salamanders(Desmognathus ocoee). Horm. Behav. 58, 427–432.
terone, corticosterone, and metabolites in response to stress, reproductive(2014), http://dx.doi.org/10.1016/j.ygcen.2014.01.010