Page 1
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions,research libraries, and research funders in the common goal of maximizing access to critical research.
An In-silico Genomic Survey to Annotate Genes Coding for Early Development-Relevant Signaling Molecules in the Pearl Oyster, Pinctada fucataAuthor(s): Davin H. E. Setiamarga , Keisuke Shimizu , Junpei Kuroda , Kengo Inamura , Kei Sato ,Yukinobu Isowa , Makiko Ishikawa , Reo Maeda , Tomoyuki Nakano , Tomoko Yamakawa , RyoHatori , Akira Ishio , Kayo Kaneko , Kenjiroo Matsumoto , Isao Sarashina , Shinnosuke Teruya , RanZhao , Nori Satoh , Takenori Sasaki , Kenji Matsuno and Kazuyoshi EndoSource: Zoological Science, 30(10):877-888. 2013.Published By: Zoological Society of JapanDOI: http://dx.doi.org/10.2108/zsj.30.877URL: http://www.bioone.org/doi/full/10.2108/zsj.30.877
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological,and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and bookspublished by nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercialinquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
Page 2
2013 Zoological Society of JapanZOOLOGICAL SCIENCE 30: 877–888 (2013)
An In-silico Genomic Survey to Annotate Genes Coding for Early
Development-relevant Signaling Molecules in the
Pearl Oyster, Pinctada fucata
Davin H. E. Setiamarga1*†, Keisuke Shimizu1†, Junpei Kuroda2‡, Kengo Inamura1,
Kei Sato1,3, Yukinobu Isowa1, Makiko Ishikawa1, Reo Maeda2, Tomoyuki Nakano4,
Tomoko Yamakawa2‡, Ryo Hatori2, Akira Ishio2, Kayo Kaneko2, Kenjiroo Matsumoto2,
Isao Sarashina1,3, Shinnosuke Teruya1,3, Ran Zhao1, Nori Satoh5,
Takenori Sasaki1,3, Kenji Matsuno2‡, and Kazuyoshi Endo1
1Department of Earth and Planetary Science, Graduate School of Science, The University of Tokyo,
7-3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, Japan2Department of Biological Science and Technology, Tokyo University of Science, 2641 Yamazaki,
Noda, Chiba Prefecture 278-8510, Japan3The University Museum, The University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, Japan
4Seto Marine Biological Laboratory, Kyoto University, 459 Shirahama, Nishimuro,
Wakayama Prefecture 649-2211, Japan5Marine Genomics Unit, Okinawa Institute of Science and Technology Graduate University,
1919-1 Tancha, Onna, Okinawa 904-0495, Japan
The pearl oyster Pinctada fucata has great potential as a model system for lophotrochozoan devel-
opmental biology research. Pinctada fucata is an important commercial resource, and a significant
body of primary research on this species has emphasized its basic aquaculture biology such as
larval biology and growth, aquaculture, pearl formation and quality improvement, shell formation,
and biomineralization. Recently, a draft genome sequence of this species was published, and many
experimental resources are currently being developed, such as bioinformatics tools, embryo and
larva manipulation methods, gene knockdown technique, etc. In this paper, we report the results
from our genomic survey pertaining to gene families that encode developmental signaling ligands
(Fgf, Hedgehog, PDGF/VEGF, TGFββ, and Wnt families). We found most of the representative genes
of major signaling pathways involved in axial patterning, as well as copies of the signaling molecule
paralogs. Phylogenetic character mapping was used to infer a possible evolutionary scenario of the
signaling molecules in the protostomes, and to reconstruct possible copy numbers of signaling
molecule-coding genes for the ancestral protostome. Our reconstruction suggests that P. fucataretains the ancestral protostome gene complement, providing further justifications for the use of
this taxon as a model organism for developmental genomics research.
Key words: pearl oyster, signaling molecules, ancestral protostome, lophotrochozoans, early development
INTRODUCTION
Current progress in molecular developmental biology
has brought a deep understanding of the molecular under-
pinnings of animal morphogenesis. Although such research
has been performed on only a handful of ecdysozoans (the
fruit fly Drosophila melanogaster and the nematode
Caenorhabditis elegans) and vertebrates (the mouse Mus
musculus, the African clawed frog Xenopus tropicalis, the
chicken Gallus gallus, and the zebrafish Danio rerio), metic-
ulous molecular, genetic, and genomic dissections of the
development of such model systems have provided signifi-
cant information on the role of gene regulations and signal-
ing networks during animal embryonic development. We
now know that in metazoans, seven of about 20 signaling
pathways controlling cellular interactions and differentiations
are involved in the morphogenetic processes during embry-
onic development (Barolo and Posakony, 2002; Pires-da
Silva and Sommer, 2003; Gazave et al., 2009).
Although extrapolative and limited, information from
these model organisms has provided some insight into the
genetic mechanisms of metazoan body plan evolution.
* Corresponding author. Tel. : +81-3-5841-4553;
Fax : +81-3-5841-4555;
E-mail: [email protected] † These authors contributed equally.‡ Present address: Department of Biological Science, Osaka Uni-
versity, 1-1 Machikaneyama, Toyonaka, Osaka 560-0043, Japan
Supplemental material for this article is available online.
doi:10.2108/zsj.30.877
Page 3
D. H. E. Setiamarga et al.878
Recent progress in the field of evolutionary developmental
biology (evo-devo), which compares different modes of ani-
mal development and their evolution has revealed that a
basic core of signaling pathways is involved in the morpho-
genesis of all metazoans. Research utilizing non-model
experimental animals such as the acorn worm, amphioxus,
annelids, sea anemones, etc., have confirmed the presence
of at least some of the representative gene members of the
developmental signaling pathways in such animals (e.g.,
Kusserow et al., 2005; Saina et al., 2009; Dray et al., 2010;
Pani et al., 2012; Onai et al., 2012). Until relatively recently,
despite the current popularity of invertebrate evo-devo stud-
ies, developmental genomics research on lophotrochozoans
has remained arguably lacking. This may be due in part to
difficulties in obtaining sufficient numbers of fresh embryos,
lack of basic descriptive information and experimental tools,
and the paucity of genomic resources.
However, intensive studies on development and breed-
ing of some lophotrochozoan species has partially solved
the problem of obtaining fresh samples, while recent devel-
opments in sequencing technology have allowed for the
development of genomics tools. For example, one recent
report on spiralian genome sequences (Simakov et al.,
2013) shed further insights into the phylogeny and evolution
of metazoan at the genomic level. Some lophotrochozoans,
such as the pearl oyster, are cultured species, and therefore
have the potential to be developed as experimental model
systems. Recently, draft genome sequences of the Japanese
pearl oyster Pinctada fucata (Takeuchi et al., 2012) and the
oyster Crassostrea gigas (Zhang et al., 2012) have been
determined. The availability of these genome sequences
helps further development of these aquaculture lophotro-
chozoan species for developmental genomics and genome
evolution studies. In this report, we focus on the Japanese
pearl oyster P. fucata, as a part of the effort to annotate its
recently determined draft genome.
As a commercial species, the Japanese pearl oyster
Pinctada fucata has been cultured for more than 100 years
in Japan (Masaoka and Kobayashi, 2007). Its fertilized eggs
can be obtained constantly for ca. five months, and its con-
trolled fertilization and embryology have been intensely
studied, albeit from the aquaculture perspective (e.g.,
Fujimura et al., 1995). Abundant research on shell formation
and pearl development (e.g., Takeuchi et al., 2008; Fang et
al., 2011; McGinty et al., 2012) has been conducted, provid-
ing much insight into biomineralization and development. P.
fucata. EST libraries from an adult (Kinoshita et al., 2011)
and various developmental stages (Takeuchi et al., 2012)
have also been reported. The availability of aquaculture
information, experimental methods, recently available
genomic resources, and most importantly, copious fresh
embryos make this species ideal for development as a
model system for the investigation of early lophotrochozoan
development.
As one of the initial steps of providing basic information
needed to establish the Japanese pearl oyster as a model
system, we report the result of our genomic survey for early
development-relevant genes in the recently reported full
nuclear genome of the pearl oyster (Takeuchi et al., 2012).
We then focus on the signaling molecule gene families
involved in axial patterning (the FGF, Hedgehog, TGFβ,
VEGF, and Wnt families). Signaling molecules play impor-
tant roles in many morphogenetic events during various
stages of development, such as axis formation, muscle dif-
ferentiation, and nervous system development. These mole-
cules work by diffusing out from a signaling center, producing
a concentration gradient. Also known as morphogens, they
bind to receptors located on receptive surrounding cells,
inducing specific responses that depend upon the concen-
tration of the signaling molecules reaching them. Our results
reported herein indicate that most members of these signal-
ing molecule families are present in protostomes. We then
discuss the implications of our findings for interpretation of
the evolution of protostome signaling molecule genes
involved in their early development.
MATERIALS AND METHODS
Gene model searches and confirmations
We employed two different methods to identify Pinctada fucata
gene homologs. In the first, we obtained amino acid sequences of
the genes of interest from other organisms in GenBank. Retrieved
sequences were used as TBLASTN and BLASTP queries to search
the P. fucata gene models version 1.1 and the genome assembly
version 1.0. This analysis was conducted using the available
Genome Browser (http://marinegenomics.oist.jp/pinctada_fucata).
For the second method, we used the “Pfam domain search” function
available on the P. fucata Genome Browser.
Amino acid sequences of the obtained gene models from both
methods were then subjected to TBLASTN and BLASTP against
the NCBI non-redundant (nr) database for identification confirma-
tion. We also conducted TBLASTN and BLASTP searches against
the P. fucata transcriptome EST database, which is available at the
Genome Browser, to obtain additional confirmation of the gene
models. The EST sequence data were obtained from transcriptome
sequencing of several embryos and adults using Roche 454 Next
Generation Sequencer (Takeuchi et al., 2012).
To illuminate the evolution of the signaling molecule genes in
the protostomes, we browsed the available annotated draft
genomes of two lophotrochozoans (the gastropod Lottia gigantea
(Lg) and the polychaete Capitella teleta (Ct); Simakov et al., 2013),
for orthologous gene sequences.
Phylogenetic analyses of signaling molecule-coding genes
For phylogenetic analyses of signaling molecule genes with
multiple paralogous copies, we used sequences of the gene models
obtained from the P. fucata genome. We used EST sequences
when they showed a longer conserved domain sequence of the
gene than that predicted by gene models. We then used conserved
domain sequences from human (Hs) and fruit fly (Dm) obtained
from GenBank. We also obtained homologous sequences from the
draft genome of the two lophotrochozoans (the polychaete and the
limpet), by doing TBLASTN and BLASTP searches of genome
sequences using various sequence queries from mouse, fruit fly,
and molluscan sequences available in GenBank. We predicted their
conserved protein domains using SMART, and included the domain
sequences in our phylogenetic analyses.
Sequence alignments were conducted using the online version
of PROMALS3D, since this program allows users to input structural
constraints for known domains (Pei et al., 2008). Accordingly, we
first obtained “core” domain alignments from PROSITE (http://
prosite.expasy.org/) and used them as alignment constraints in
PROMALS3D. The obtained alignment was then edited manually by
using Mesquite v2.75 (Maddison and Maddison, 2011) or MEGA v5.0
(Tamura et al., 2011). Afterwards, we used MEGA v5.0 to search
for the best amino acid substitution model of the edited alignments.
In most occasions, the top four models suggested were WAG+G,
Page 4
Signaling Molecule Genes in Pearl Oyster 879
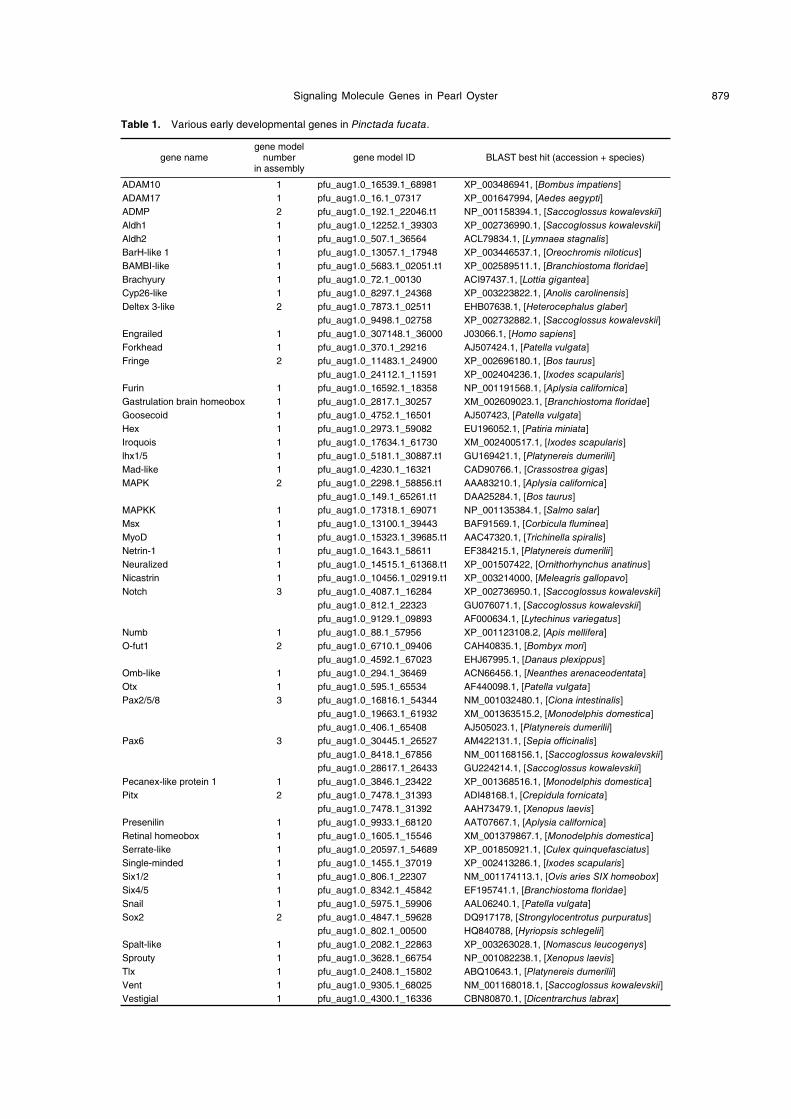
Table 1. Various early developmental genes in Pinctada fucata.
gene namegene model
numberin assembly
gene model ID BLAST best hit (accession + species)
ADAM10 1 pfu_aug1.0_16539.1_68981 XP_003486941, [Bombus impatiens]
ADAM17 1 pfu_aug1.0_16.1_07317 XP_001647994, [Aedes aegypti]
ADMP 2 pfu_aug1.0_192.1_22046.t1 NP_001158394.1, [Saccoglossus kowalevskii]
Aldh1 1 pfu_aug1.0_12252.1_39303 XP_002736990.1, [Saccoglossus kowalevskii]
Aldh2 1 pfu_aug1.0_507.1_36564 ACL79834.1, [Lymnaea stagnalis]
BarH-like 1 1 pfu_aug1.0_13057.1_17948 XP_003446537.1, [Oreochromis niloticus]
BAMBI-like 1 pfu_aug1.0_5683.1_02051.t1 XP_002589511.1, [Branchiostoma floridae]
Brachyury 1 pfu_aug1.0_72.1_00130 ACI97437.1, [Lottia gigantea]
Cyp26-like 1 pfu_aug1.0_8297.1_24368 XP_003223822.1, [Anolis carolinensis]
Deltex 3-like 2 pfu_aug1.0_7873.1_02511 EHB07638.1, [Heterocephalus glaber]
pfu_aug1.0_9498.1_02758 XP_002732882.1, [Saccoglossus kowalevskii]
Engrailed 1 pfu_aug1.0_307148.1_36000 J03066.1, [Homo sapiens]
Forkhead 1 pfu_aug1.0_370.1_29216 AJ507424.1, [Patella vulgata]
Fringe 2 pfu_aug1.0_11483.1_24900 XP_002696180.1, [Bos taurus]
pfu_aug1.0_24112.1_11591 XP_002404236.1, [Ixodes scapularis]
Furin 1 pfu_aug1.0_16592.1_18358 NP_001191568.1, [Aplysia californica]
Gastrulation brain homeobox 1 pfu_aug1.0_2817.1_30257 XM_002609023.1, [Branchiostoma floridae]
Goosecoid 1 pfu_aug1.0_4752.1_16501 AJ507423, [Patella vulgata]
Hex 1 pfu_aug1.0_2973.1_59082 EU196052.1, [Patiria miniata]
Iroquois 1 pfu_aug1.0_17634.1_61730 XM_002400517.1, [Ixodes scapularis]
lhx1/5 1 pfu_aug1.0_5181.1_30887.t1 GU169421.1, [Platynereis dumerilii]
Mad-like 1 pfu_aug1.0_4230.1_16321 CAD90766.1, [Crassostrea gigas]
MAPK 2 pfu_aug1.0_2298.1_58856.t1 AAA83210.1, [Aplysia californica]
pfu_aug1.0_149.1_65261.t1 DAA25284.1, [Bos taurus]
MAPKK 1 pfu_aug1.0_17318.1_69071 NP_001135384.1, [Salmo salar]
Msx 1 pfu_aug1.0_13100.1_39443 BAF91569.1, [Corbicula fluminea]
MyoD 1 pfu_aug1.0_15323.1_39685.t1 AAC47320.1, [Trichinella spiralis]
Netrin-1 1 pfu_aug1.0_1643.1_58611 EF384215.1, [Platynereis dumerilii]
Neuralized 1 pfu_aug1.0_14515.1_61368.t1 XP_001507422, [Ornithorhynchus anatinus]
Nicastrin 1 pfu_aug1.0_10456.1_02919.t1 XP_003214000, [Meleagris gallopavo]
Notch 3 pfu_aug1.0_4087.1_16284 XP_002736950.1, [Saccoglossus kowalevskii]
pfu_aug1.0_812.1_22323 GU076071.1, [Saccoglossus kowalevskii]
pfu_aug1.0_9129.1_09893 AF000634.1, [Lytechinus variegatus]
Numb 1 pfu_aug1.0_88.1_57956 XP_001123108.2, [Apis mellifera]
O-fut1 2 pfu_aug1.0_6710.1_09406 CAH40835.1, [Bombyx mori]
pfu_aug1.0_4592.1_67023 EHJ67995.1, [Danaus plexippus]
Omb-like 1 pfu_aug1.0_294.1_36469 ACN66456.1, [Neanthes arenaceodentata]
Otx 1 pfu_aug1.0_595.1_65534 AF440098.1, [Patella vulgata]
Pax2/5/8 3 pfu_aug1.0_16816.1_54344 NM_001032480.1, [Ciona intestinalis]
pfu_aug1.0_19663.1_61932 XM_001363515.2, [Monodelphis domestica]
pfu_aug1.0_406.1_65408 AJ505023.1, [Platynereis dumerilii]
Pax6 3 pfu_aug1.0_30445.1_26527 AM422131.1, [Sepia officinalis]
pfu_aug1.0_8418.1_67856 NM_001168156.1, [Saccoglossus kowalevskii]
pfu_aug1.0_28617.1_26433 GU224214.1, [Saccoglossus kowalevskii]
Pecanex-like protein 1 1 pfu_aug1.0_3846.1_23422 XP_001368516.1, [Monodelphis domestica]
Pitx 2 pfu_aug1.0_7478.1_31393 ADI48168.1, [Crepidula fornicata]
pfu_aug1.0_7478.1_31392 AAH73479.1, [Xenopus laevis]
Presenilin 1 pfu_aug1.0_9933.1_68120 AAT07667.1, [Aplysia californica]
Retinal homeobox 1 pfu_aug1.0_1605.1_15546 XM_001379867.1, [Monodelphis domestica]
Serrate-like 1 pfu_aug1.0_20597.1_54689 XP_001850921.1, [Culex quinquefasciatus]
Single-minded 1 pfu_aug1.0_1455.1_37019 XP_002413286.1, [Ixodes scapularis]
Six1/2 1 pfu_aug1.0_806.1_22307 NM_001174113.1, [Ovis aries SIX homeobox]
Six4/5 1 pfu_aug1.0_8342.1_45842 EF195741.1, [Branchiostoma floridae]
Snail 1 pfu_aug1.0_5975.1_59906 AAL06240.1, [Patella vulgata]
Sox2 2 pfu_aug1.0_4847.1_59628 DQ917178, [Strongylocentrotus purpuratus]
pfu_aug1.0_802.1_00500 HQ840788, [Hyriopsis schlegelii]
Spalt-like 1 pfu_aug1.0_2082.1_22863 XP_003263028.1, [Nomascus leucogenys]
Sprouty 1 pfu_aug1.0_3628.1_66754 NP_001082238.1, [Xenopus laevis]
Tlx 1 pfu_aug1.0_2408.1_15802 ABQ10643.1, [Platynereis dumerilii]
Vent 1 pfu_aug1.0_9305.1_68025 NM_001168018.1, [Saccoglossus kowalevskii]
Vestigial 1 pfu_aug1.0_4300.1_16336 CBN80870.1, [Dicentrarchus labrax]
Page 5
D. H. E. Setiamarga et al.880
WAG+G+I, JTT+G, and JTT+I+G. Whenever possible, we used the
best model suggested for subsequent phylogenetic analyses. Max-
imum Likelihood (ML) phylogenetic analyses were conducted using
the online version of RAxML (RAxML Blackbox; Stamatakis et al.,
2008; http://phylobench.vital-it.ch/raxml-bb/), with 100 bootstrap
replications.
Protein domain re-prediction using SMART for signaling mole-
cule genes
Although the Genome Browser provided a domain prediction
for all of its gene models, for further confirmation, we re-predicted
domain structures using the online version of the protein domain
annotation software, SMART (Letunic et al., 2012; Schultz et al.,
1998; http://smart.embl-heidelberg.de/). We based our diagrams of
signaling molecule domain structures on SMART results. In our fig-
ures, we provided diagrams of only our gene models, not of EST
sequences.
RESULTS AND DISCUSSION
Early development-relevant genes in the Pinctada fucata genome
We found and annotated 80 gene models, related to
various early developmental events, in the genome of
Pinctada fucata (Tables 1 and 2). Although we did not
exhaustively search for all currently known early develop-
ment-relevant genes, we did identify representatives of
major signaling pathways involved in early development. As
with all genes identified through wholesale in-silico gene
model predictions and annotations of draft genome
sequences, further experimental analyses through sequence
cloning and functional analyses will be necessary to confirm
their existence and functions. Further quality improvement of
the draft genome and the transcriptome data will also help
future verifications. Nonetheless, our results presented here
provide a starting point for such analyses, and at least at the
current stage, allow preliminary insights into the genomic
reality of development-relevant genes in P. fucata.
Signaling molecule gene families in the Pinctada fucata genome
We obtained gene models for most of the signaling mol-
ecule genes involved in axial patterning, confirming their
presence in the P. fucata genome (Table 2). We next
checked the EST data bank (obtained by transcriptome
Table 2. List of annotated gene models for early development-relevant signaling molecules and their receptors in Pinctada fucata.
Family Category gene namegene model
numberin assembly
gene model ID BLAST best hit (accession + species)
FGF Ligand FGF8/17/18 1 pfu_aug1.0_622.1_36612 NP_001164697.1, [Saccoglossus kowalevskii]
FGF19/21/23 1 pfu_aug1.0_1195.1_58407 XP_003226252.1, [Anolis carolinensis]
Receptor FGFR 1 pfu_aug1.0_1303.1_07891 BAI67805.1, [Idiosepius paradoxus]
Headghog Ligand Hedgehog 1 pfu_aug1.0_424.1_50858 EU394706.1, [Lottia gigantea]
Receptor Patched 1 pfu_aug1.0_5322.1_16629.t1 EFX90375.1, [Daphnia pulex]
PDGF/VEGF Ligand PVF1-like 1 pfu_aug1.0_938.1_58309 XP_002411998.1, [Ixodes scapularis]
Receptor PDGF- and VEGF-receptor 1 pfu_aug1.0_3981.1_16235.t1 BAI67804.1, [Idiosepius paradoxus]
TGFβ Ligand Dpp-BMP2/4 1 pfu_aug1.0_790.1_51081.t1 BAD16731.1, [Pinctada fucata]
BMP3 2 pfu_aug1.0_43625.1_63151.t1 ACF93445.1, [Branchiostoma japonicum]
pfu_aug1.0_102538.1_06302 XP_001494823.2, [Equus caballus]
Gbb-BMP5-8 1 pfu_aug1.0_23580.1_26106 XP_002407531.1, [Ixodes scapularis]
BMP9/10 1 pfu_aug1.0_2637.1_30198 CAD67715.1, [Crassostrea gigas]
Nodal 1 pfu_aug1.0_447.1_29262 ACB42422.1, [Biomphalaria glabrata]
Myostatin 1 pfu_aug1.0_156.1_22035 ABJ09581.2, [Chlamys farreri]
Marverick-like 1 pfu_aug1.0_6051.1_16824 CAD67714.1, [Crassostrea gigas]
Receptor BMPR1 1 pfu_aug1.0_257.1_50753 CAE11917.1, [Crassostrea gigas]
BMPR2 2 pfu_aug1.0_14312.1_32434 XP_001184902.1, [Strongylocentrotus purpuratus]
pfu_aug1.0_27640.1_04591 XP_001184902.1, [Strongylocentrotus purpuratus]
ACVR1 2 pfu_aug1.0_40446.1_70 ADD80738.1, [Pinctada fucata]
pfu_aug1.0_38196.1_0503 ADD80738.1, [Pinctada fucata]
ACVR1B 1 pfu_aug1.0_11.1_50544 CAD20573.1, [Crassostrea gigas]
ACVR2 1 pfu_aug1.0_3667.1_16148 CAR92545.1, [Crassostrea gigas]
Wnt Ligand WntA 1 pfu_aug1.0_964.1_00569 CAD37169.2, [Platynereis dumerilii]
Wnt1 1 pfu_aug1.0_11928.1_60961 CAD37164.2, [Platynereis dumerilii]
Wnt2 1 pfu_aug1.0_9858.1_38969 CAD37171.1, [Patella vulgata]
Wnt4 1 pfu_aug1.0_8897.1_53252 CAD37166.2, [Platynereis dumerilii]
Wnt6 1 pfu_aug1.0_39331.1_62990 ACE79727.1, [Branchiostoma lanceolatum]
Wnt7 1 pfu_aug1.0_76.1_29065 ABD16199.1, [Euprymna scolopes]
Wnt9 2 pfu_aug1.0_189.1_36368 XP_003206944.1, [Meleagris gallopavo]
pfu_aug1.0_189.1_36369 XP_003205766.1, [Meleagris gallopavo]
Wnt10 1 pfu_aug1.0_16143.1_54282 CAD37172.1, [Patella vulgata]
Wnt11 1 pfu_aug1.0_48418.1_70635 ADK38674.2, [Platynereis dumerilii]
Wnt16 1 pfu_aug1.0_5965.1_31091 ADR81924.2, [Platynereis dumerilii]
Receptor Frz-1/2/7 1 pfu_aug1.0_87126.1_42186.t1 NP_001079207.1, [Xenopus laevis]
Frz-4 1 pfu_aug1.0_3031.1_44550.t1 XP_002730495.1, [Saccoglossus kowalevskii]
Frz-5/8 1 pfu_aug1.0_16140.1_03676.t1 ADZ61652.1, [Ptychodera flava]
Page 6
Signaling Molecule Genes in Pearl Oyster 881
sequencing; Takeuchi et al., 2012), which is available on the
genome browser homepage, to see if the gene models for
the signaling molecules are expressed. We found ESTs for
some, but not all, gene models (Table 3). One possible
explanation for this is that developmental genes are
expressed at relatively low levels compared to housekeep-
ing and metabolic genes, and therefore the transcriptome
sequencing was unable to detect them.
1. FGF
A Pfam domain search for the FGF domain containing
genes on the P. fucata Genome Browser suggested five
prospective gene models. Subsequent confirmations identi-
fied the presence of only two fgf homologs, and the other
three were false hits. TBLASTN, BLASTP, and phylogenetic
analyses confirmed the identities of the two gene models as
fgf8 and fgf19 homologs. We named the gene models
accordingly (Pifuc-fgf8L and Pifuc-fgf19L – the suffix “L”
denotes “Like”) (Table 2; Fig. 1). SMART predicted the pres-
ence of a signal peptide motif and an FGF domain on Pifuc-
Fgf8L, indicating that this is a full-length gene. Pifuc-Fgf19L
lacked the signal peptide domain, suggesting that this gene
model is most likely partial, lacking the 5′ end (Fig. 1A).
However, we detected an EST sequence of this gene in the
transcriptome data, giving further support to the presence of
Pifuc-fgf19L in P. fucata. We also identified a single FGF
receptor (Pifuc-fgfr) gene model (Table 2).
Interestingly, dual copies of the fgf gene are found in
other protostomes too, with one copy being a supported
homolog of fgf8, while the homology of the other copy is
uncertain (e.g., Stathopoulos et al., 2004; Huang and Stern,
2005; Bertrand et al., 2011). Two of the three fgf genes in
the fruit fly, pyramus and thisbe, are fgf8 orthologs, and
they are most likely products of a lineage-specific local
duplication (Stathopoulos et al., 2004). Kadam et al. (2009)
showed that at least in the fruit fly, the two types of FGF
ligands (Pyramus+Thisbe vs. Branchless) are functionally
non-interchangeable. Although the phylogenetic interrela-
tionships among the non-fgf8 genes are unclear, it is possi-
bly a homolog to “the rest” of the fgfs (the non-fgf8s). These
observations suggest the possibility that the two copies of P.
fucata fgf gene models are not unique to this species, but
synapomorphic in the protostomes (Fig. 6). Future studies
involving gene structure and synteny analyses among vari-
ous metazoans and functional analyses may help to reveal
such interrelationships among fgf genes.
2. Hedgehog
We identified one hedgehog gene model (Pifuc-hh) in
the P. fucata genome. Homology of the gene model was
confirmed by TBLASTN, BLASTP and phylogenetic analy-
ses (Fig. 2A, B). The phylogenetic tree shows the affinity of
Pifuc-hh to its Hs, Dm, Lg, and Ct homologs. Protein domain
structure prediction using SMART indicated the presence of
a signal peptide, a Hedge domain, and a Hint (“Hog”)
domain, indicating that the gene model for Pifuc-hh is full-
length (Fig. 2A). We also identified one Hedgehog receptor
gene model (Pifuc-hhR, Table 2). The single gene copy of
Pifuc-hh is consistent with current insights about the copy
numbers of hh in protostomes (Fig. 6).
The presence of a complete hedgehog with both the
Hedge and Hint domains is unsurprising, as it is consistent
with findings from L. gigantea and C. teleta genomes. In
those two lophotrochozoans, we found single copies of
hedgehog with both the Hedge and Hint domains present,
as predicted by SMART. Previously, Matus et al. (2008)
suggested that lophotrochozoan Hedgehog might have lost
its Hedge domain. This was concluded because a genomic
search for hedgehog homologs in C. teleta found only Hint
domain-containing copies. However, the absence of Hedge
domain in C. teleta might have been caused by the quality
of the available draft genome data at that time. Meanwhile
in Ecdysozoa, although the fruit fly has a complete hedgehog
gene containing both the Hedge and the Hint domains, the
nematode C. elegans has apparently lost its Hedge domain
and has duplicated its remaining Hint containing hedgehog
homologs (Aspöck et al., 1999; Hao et al., 2006; Matus et
al., 2008). However, since other nematodes may have a
complete hedgehog with both Hedge and Hint domains
present (Matus et al., 2008), it is most likely that the missing
Hedge in C. elegans was caused by a lineage-specific loss.
In summary, our results strongly suggest that the pearl
oyster P. fucata has retained the protostomian ancestral
condition of a complete single hedgehog gene with both
Hedge and Hint domains (Fig. 6).
Table 3. List of ESTs of Pinctada fucata’s annotated early devel-
opment-relevant signaling molecule gene models.
Family gene name
Number
of RNA
sequence
EST ID
FGF FGF8/17/18 0 –
FGF19/21/23 1 pfudmixc36137g12718
Headghog Hedgehog 0 –
PDGF/VEGF PVF1-like 0 –
TGFβ Dpp-BMP2/4 2 pfudmixc14451g01574
pfudmixc14449g01574
BMP3 1 pfudmixc32706g09287
Gbb-BMP5-8 3 pfudegg-4cellc02598g01138
pfud8-16cellc02367g00970
pfudmixc24496g05056
BMP9/10 0 –
Nodal 1 pfudmixc32568g09149
Myostatin 2 pfudmixc22489g04023
pfudtrochophorec00808g00102
Marverick-like 0 –
Wnt WntA 3 pfudmixc33241g09822
pfudtrochophorec02900g01620
pfudDshapec04488g02500
Wnt1 0 –
Wnt2 2 pfudegg-Dshapec05041g03189
pfudmixc38205g14786
Wnt4 3 pfud8-16cellc02776g01379
pfudDshapec04106g0211
pfudmixc08397g00554
Wnt6 0 –
Wnt7 0 –
Wnt9 0 –
Wnt10 0 –
Wnt11 2 pfudtrochophorec03235g01955
pfudmixc33657g10238
Wnt16 1 pfudmixc35143g11724
Page 7
D. H. E. Setiamarga et al.882
3. PDGF/VEGF
We found one PDGF/VEGF-related factor gene
model, which we call Pfupvf, in the P. fucata genome.
TBLASTN, BLASTP and phylogenetic analyses con-
firmed that Pifuc-pvf was a homolog of the fruit fly pvf1
(Fig 3). SMART predicted the presence of a PDGF
domain on Pifuc-PVF (Fig. 3A). However, this gene
model does not include a signal peptide, indicating that
it is probably not a full-length sequence. We also iden-
tified a single PDGF/VEGF-related factor receptor gene
model (Table 2).
The fruit fly D. melanogaster has three pvf-like
genes (Cho et al., 2002; Duchek et al., 2001; Heino et
al., 2001; Pietrokovski and Shilo, 2001), while our draft
genome searches of several other insects (e.g., Tribo-
lium castaneum, Acyrthosiphon pisum, and Nasonia vit-
ripennis) found variable pvf gene copy numbers: two in
T. castaneum, one in A. pisum, and one in N. vitripennis.
A genome survey of the nematode C. elegans sug-
gested that the nematode most likely has only one copy
(Popovici et al., 1999; Popovici et al., 2002). Our surveys
Fig. 1. Pifuc-Fgf protein structures and phylogenetic tree. (A) Domain structures of the two Pifuc-Fgf identified in this study. Both gene mod-
els have an Fgf protein domain. However, the signal peptide domain is not present in PfuFgf19-Like gene model, suggesting that this gene
model might be incomplete. (B) Phylogenetic tree of the Fgfs based on the amino acid sequences of their conserved Fgf domains. We included
all Fgf homologs and paralogs from fruit fly (Dm), human (Hs), limpet (Lg), and a polychaete (Ct). The two P. fucata gene models are placed
with their putative homologs, confirming BLAST results. Bootstrap supports below 40% are not shown.
Fig. 2. (A) The protein structures of Pifuc-Hh. Domain structures of
Pifuc-Hh clearly show a complete Hedgehog protein, containing a signal
peptide, a Hedge, and a Hint (or Hog) domains. (B) Molecular phylogeny
of Pifuc-Hh clustered it together with the hedgehog homologs from other
protostomes. Bootstrap supports below 40% are not shown.
Page 8
Signaling Molecule Genes in Pearl Oyster 883
on the two lophotrochozoan draft genomes also suggest a
variety of copy numbers (two in L. gigantea and three in C.
teleta). This variety of copy numbers suggest that a dynamic
evolutionary events involving local duplication and/or gene
losses have happened to pvf-like genes in the protostomes.
We were unable to confidently deduce the evolutionary
history of the PVDF/VEGF genes through the result of our
phylogenetic analysis, because most branches were not
supported by high bootstrap values. However, despite the
lack of a robust tree, one can infer that duplication and sub-
sequent sub-functionalization events have happened in ver-
tebrates independent from those that occurred in the protos-
tomes, causing the duplicated genes to differentiate into
PDGF and VEGF subfamilies (Holmes and Zachary, 2005).
Our observation here underlines the importance of improved
draft genomes and further molecular studies involving clon-
ing and functional studies, to conclusively elucidate the evo-
lutionary history of this gene family.
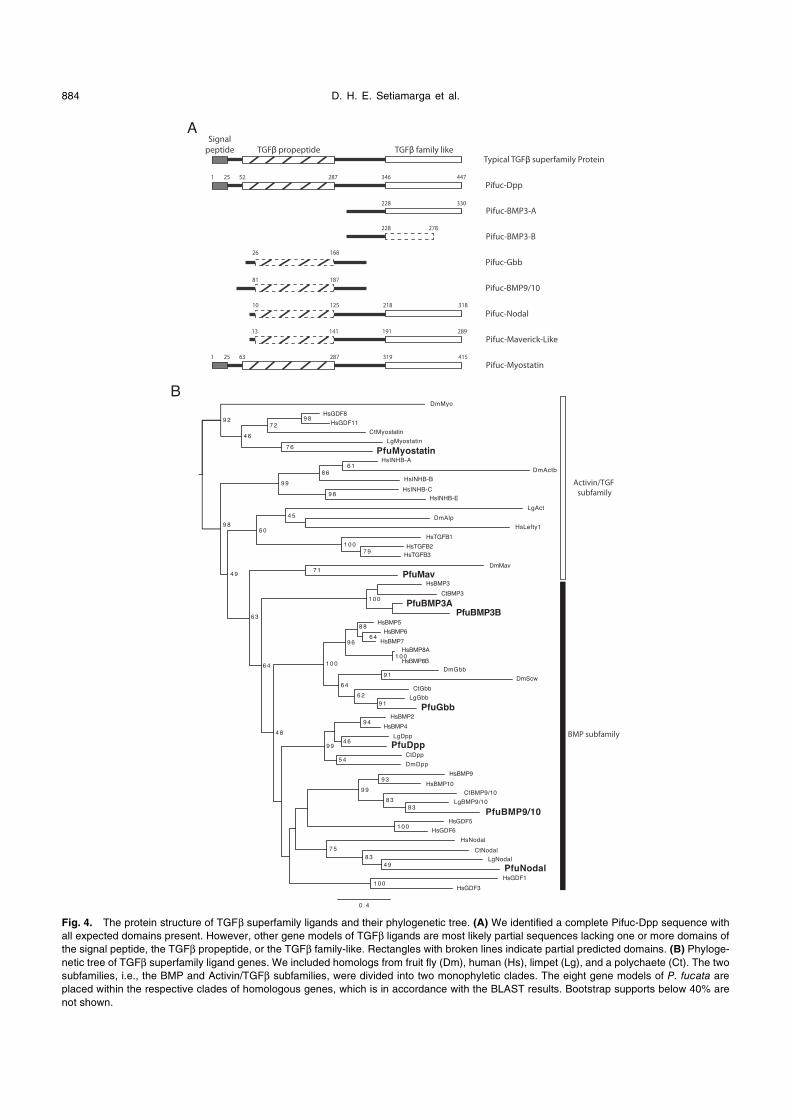
4. TGFββThe TGFβ superfamily consists of two subfamilies, the
BMP and the Activin/TGFβ subfamilies. A typical TGFβligand has two conserved domains, the TGFβ propeptide and
TGFβ-like domains (Fig. 4A). Previous molluscan studies
have identified three TGFβ ligand-coding genes (Nederbragt
et al., 2002; Grande and Patel, 2009; Kin et al., 2009). How-
ever, those previous studies suffered from the lack of
genomic data. Accordingly, we conducted a genomic survey
of TGFβ superfamily ligands in P. fucata to look for their
presence in the genome. We annotated five gene models as
homologs of BMP subfamily ligands (dpp-bmp2/4, bmp3,
gbb-bmp5-8, bmp9/10, and nodal), an Activin/TGFβ sub-
family ligand (myostatin), and maverick. The homology of
each copy was checked with TBLASTN, BLASTP, and phy-
logenetic analysis. The resulting phylogenetic tree (Fig. 4B)
confidently placed each copy with its tentative homologous
genes from other bilaterians, with reasonable bootstrap sup-
ports.
We did not find any gene model or EST for the other
three TGFβ family ligands (i.e., activin, tgfβ, and lefty) in the
currently available predicted gene models and the transcrip-
tome data. Our survey on the genome data of L. gigantea
and C. teleta also failed to find any gene model for tgfβ and
lefty. Previous studies have suggested that the absence of
tgfβ and lefty is probably synapomorphic to protostomes
(e.g., Van der Zee et al., 2008).
Our genomic survey also failed to identify any gene
model for activin in L. gigantea and C. teleta. Although
Grande and Patel (2009) reported the presence of an activin
homolog in L. gigantea, they did not detect its expression in
the developing embryo, leaving its function unclear, while its
affinity to other activin homologs was poorly supported. In
our phylogenetic analyses, the reported Lg activin grouped
with fruit fly’s activin-like protein with low support (Fig. 4B),
Fig. 3. (A) A comparison of Pifuc-Pvf protein with other VEGF and PDGF genes. PDGF/VEGF was thought to have later diverged into PDGF
and VEGF in vertebrates. Pifuc-Pvf identified in this report lacks a signal peptide, suggesting that it is an incomplete gene model. (B) Molecular
phylogenetic analyses of Pifuc-Pvf included PDGF and VEGF sequences from fruit fly (Dm), human (Hs), limpet (Lg), and a polychaete (Ct).
Bootstrap supports below 40% are not shown.
Page 9
D. H. E. Setiamarga et al.884
Fig. 4. The protein structure of TGFβ superfamily ligands and their phylogenetic tree. (A) We identified a complete Pifuc-Dpp sequence with
all expected domains present. However, other gene models of TGFβ ligands are most likely partial sequences lacking one or more domains of
the signal peptide, the TGFβ propeptide, or the TGFβ family-like. Rectangles with broken lines indicate partial predicted domains. (B) Phyloge-
netic tree of TGFβ superfamily ligand genes. We included homologs from fruit fly (Dm), human (Hs), limpet (Lg), and a polychaete (Ct). The two
subfamilies, i.e., the BMP and Activin/TGFβ subfamilies, were divided into two monophyletic clades. The eight gene models of P. fucata are
placed within the respective clades of homologous genes, which is in accordance with the BLAST results. Bootstrap supports below 40% are
not shown.
Page 10
Signaling Molecule Genes in Pearl Oyster 885
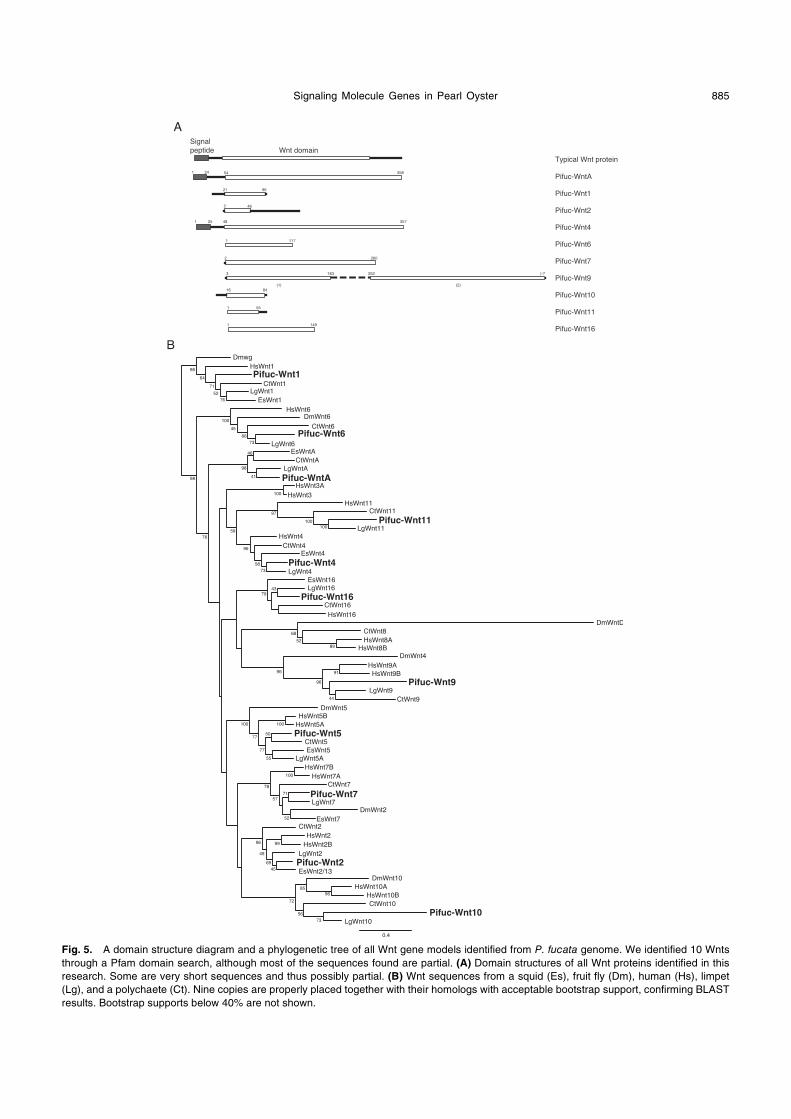
Fig. 5. A domain structure diagram and a phylogenetic tree of all Wnt gene models identified from P. fucata genome. We identified 10 Wnts
through a Pfam domain search, although most of the sequences found are partial. (A) Domain structures of all Wnt proteins identified in this
research. Some are very short sequences and thus possibly partial. (B) Wnt sequences from a squid (Es), fruit fly (Dm), human (Hs), limpet
(Lg), and a polychaete (Ct). Nine copies are properly placed together with their homologs with acceptable bootstrap support, confirming BLAST
results. Bootstrap supports below 40% are not shown.
Page 11
D. H. E. Setiamarga et al.886
but not with other activins. We also identified five gene
models for TGFβ receptors (bmp receptor type I, type II,
activin receptor type I, IB and type II) in the P. fucata
genome (Table 2, Supplementary Fig. S1). Since Myostatin
and probably other BMP ligands also bind to Activin recep-
tors (Huminiecki et al., 2009), the presence of these recep-
tors does not necessarily suggest the presence of activin.
With our current data, we are unable to confidently conclude
the presence of activin in mollusks, or lophotrochozoans.
Therefore, future genomic and molecular analyses will be
needed to conclusively show if activin is really present or
absent in P. fucata and other mollusks. However, since the
presence of two activin homologs has also been reported in
the fruit fly (Zhu et al., 2008), we can deduce that the pres-
ence of activin is probably ancestral to protostomes (Fig. 6).
5. Wnt
A Pfam domain search found 13 wnt candidate gene
models. Further analyses confirmed the presence of ten
PifucWnts gene models (wntA, 1, 2, 4, 6, 7, 9, 10, 11, 16)
(Table 2, Fig. 5). Most of the Pifuc-wnt sequences obtained
were short and probably partial, with Wnt11 as the shortest
(68 amino acids) (Fig. 5A). Phylogenetic analysis confidently
placed most of the gene models with their respective
human, fruit fly, squid (Euprymna scolopes), limpet, and
polychaete homologs, a result that is consistent with the ini-
tial BLASTP and TBLASTN results. We also found three
gene models for Wnt receptors, Frizzled, in the P. fucata
genome (Table 2, Supplementary Fig. S2).
The presence of multiple wnt genes in P. fucata is con-
sistent with our current knowledge of ancient wnt duplica-
tions, as most wnt genes and their receptors apparently
already existed in ancestral cnidarians (Kusserow et al.,
2005). In this study, we did not find any copy of wnt3 in the
P. fucata genome, or any EST sequence in the transcrip-
tome data. Previous studies have also indicated the
absence of wnt3 in the protostomes, suggesting that the
absence of the wnt3 homolog is possibly a synapomorphic
character of the protostomes, in agreement with previous
studies (Schubert et al., 2000; Kusserow et al., 2005; Cho
et al., 2010; Janssen et al., 2010). We also failed to detect
any gene model for wnt5 gene model in the present draft
genome of P. fucata. However, wnt5 is present in other
mollusks surveyed in our present study on wnt genes (E.
scolopes and L. gigantea). We also detected a wnt5 EST
sequence in the transcriptome data. From these observa-
tions, we can confidently deduce that wnt5 is present in P.
fucata, and its absence is most likely an artifact caused by
the current quality of the P. fucata draft genome. Our result
also suggests that wnt8 is absent in P. fucata, a conclusion
supported by its absence in both the draft genome and tran-
scriptome data. Meanwhile, our L. gigantea draft genome
search also indicated its absence, although we found the
gene in the draft genome of C. teleta. These results suggest
that wnt8 was probably lost in the molluscan lineage, in
agreement with previous studies (Kusserow et al., 2005;
Cho et al., 2011).
It should be noted that originally, Pfam domain search
Fig. 6. Reconstructions of gene copy numbers of the ancestral lophotrochozoans and ancestral protostome. We assumed that the greatest
common factor in which all gene copies present in any extant protostome must have had at least one copy in the ancestors, and if multiple cop-
ies are present in only one species, we assumed that it was duplicated locally. We also put the information of deuterostome genes into consid-
eration. For the ancestral deuterostome, we refer to findings from the basal chordate amphioxus and other vertebrates. Question marks
indicate uncertainties or lack of information; while cross marks on the ancestral protostome indicate possible absence.
Page 12
Signaling Molecule Genes in Pearl Oyster 887
found two wnt9 gene models (pfu_aug1.0_189.1_3639 and
pfu_aug1.0_189.1_3638), which were located in tandem on
the same scaffold. BLAST searches allied both genes to the
same wnt9a homolog in turkey (Meleagris gallopavo).
When both “genes” were subjected to SMART protein
domain searches, they were shown to be composed of only
partial Wnt domains. We retrieved the portion of the scaffold
sequence where the two gene models are located, and a
quick inspection of that sequence revealed that the
sequence of the region in-between the two original gene
models were undetermined (denoted by a series of N).
Therefore for our phylogenetic analysis, we simply concate-
nated the two gene models and treated them as a single
sequence.
When we plot the number of existing copies of wnt
genes in mollusks, annelids, insects, and nematodes, mol-
lusks and annelids show an almost complete number of wnt
copies, compared to model ecdysozoans (Fig. 6; Guder et
al., 2006; Cho et al., 2011). This suggests that mollusks
retain the ancient number of wnt genes, close to that pre-
dicted for the ancestral protostome. Future functional analy-
ses will be needed to see if both the copy number and the
gene functions are also conserved.
Insights into the evolution of signaling molecule genes
in protostomes
In this study, we conducted a general survey for genes
related to axis formation in early embryonic development.
We then focused our survey on signaling ligand-coding gene
families, namely FGF, Hedgehog, PDGF/VEGF, TGFβ, and
Wnt.
Our results allow us to predict the possible ancestral
condition of the gene copy numbers in protostomes, and
possible evolutionary processes that account for them. In
Fig. 6, we present a phylogenetic character mapping for the
presence/absence of FGF, Hedgehog, Wnt, TGFβ, and
PDGF/VEGF gene family members on the protostome tree.
The results suggested that the ancestral protostome had two
copies of fgf, a single copy of hedgehog, five copies of bmp-
related genes, one copy of activin, one copy of maverick,
and 12 copies of wnt genes. Figure 6 indicates that genomes
of D. melanogaster and C. elegans are possibly highly
derived because of extensive local duplications and lineage
specific losses. It appears that mollusks retain most gene
copies, probably indicating that molluscan genomes are
closer to that of the ancestral protostome.
Future expectations
The presence of a series of long N in various scaffolds
and the relatively short length of some scaffolds in the cur-
rent version of P. fucata draft genome may have been the
cause of our inability to identify complete sequences for
many gene models. Current transcriptome sequencing
method used to generate the currently available transcrip-
tome data may not be sufficiently sensitive to detect genes
expressed at very low levels, hence our inability to obtain
EST data for some signaling molecule genes. Therefore,
future work involving gene cloning and functional analyses
will be needed to confirm the in-silico results presented
here, while further refinements of P. fucata draft genome
and the transcriptome data will allow for better gene predic-
tions and synteny analyses to check gene homologies and
evolutionary histories conclusively (e.g., Bertrand et al.,
2010; Hui et al., 2012). For example, better sequence qual-
ity on longer scaffolds may enable us to see if the two wnt9
copies are actually sequences from a single gene, or if they
originated from two paralogous copies of wnt9 obtained
through a local tandem duplication event. Improved draft
genome and transcriptome data might also allow us to con-
fidently deduce the homology of lophotrochozoan and meta-
zoan non-fgf8 genes, and help us to elucidate the evolution
of the PDGF/VEGF gene family.
ACKNOWLEDGMENTS
For gene model annotations, three Genome Jamboree events
were held in 2011–2012 in Japan. These events were partially funded
by JAMBIO (Japanese Association for Marine Biology). The authors
would like to sincerely thank Takeshi Takeuchi, Takeshi Kawashima,
Ryo Koyanagi (OIST), and Hiroshi Wada (Univ. Tsukuba) for their
invaluable advice and supports. We are indebted to Christopher
Mah (Smithsonian National Muse. Nat. Hist.), Noah Ben-Aderet
(Scripps Inst. Ocean.) and Neil Aschliman (St. Ambrose Univ.) for
helping us to improve the English of our manuscript. The manu-
script’s English was then further improved by a professional editor.
We also thank two anonymous reviewers for their invaluable advice
to improve the manuscript.
REFERENCES
Aspöck G, Kagoshima H, Niklaus G, Bürglin TR (1999)
Caenorhabditis elegans has scores of hedgehog-related
genes: sequence and expression analysis. Genome Res 9:
909–923
Barolo S, Posakony JW (2002) Three habits of highly effective sig-
naling pathways: principles of transcriptional control by devel-
opmental cell signaling. Genes Dev 16: 1167–1181
Bertrand S, Camasses A, Somorjai I, Belgacem MR, Chabrol O,
Escande ML, et al. (2011) Amphioxus FGF signaling predicts
the acquisition of vertebrate morphological traits. Proc Natl
Acad Sci USA 108: 9160–9165
Cho NK, Keyes L, Johnson E, Heller J, Ryner L, Karim F, Krasnow
MA (2002) Developmental control of blood cell migration by the
Drosophila VEGF pathway. Cell 108: 865–876
Cho SJ, Vallés I, Giani Jr VG, Seaver EC, Weisblat DA (2010)
Evolutionary dynamics of the wnt gene family: A lophotro-
chozoan perspective. Mol Biol Evol 27: 1645–1658
Dray N, Tessmar-Raible K, Le Gouar M, Vibert L, Christodoulou F,
Schipany K, et al. (2010) Hedgehog signaling regulates seg-
ment formation in the annelid Platynereis. Science 329: 339–
342
Duchek P, Somogyi K, Jékely G, Beccari S, Rørth P (2001) Guidance
of cell migration by the Drosophila PDGF/VEGF receptor. Cell
107: 17–26
Fang D, Xu G, Hu Y, Pan C, Xie L, Zhang R (2011) Identification of
genes directly involved in shell formation and their functions in
pearl oyster, Pinctada fucata. PLoS One 6: e21860
Fujimura T, Wada K, Iwaki T (1995) Development and morphology
of the pearl oyster larvae, Pinctada fucata. Venus 54: 25–48 (in
Japanese)
Gazave E, Lapébie P, Richards GS, Brunet F, Ereskovsky AV,
Degnan BM, et al. (2009) Origin and evolution of the Notch sig-
nalling pathway: an overview from eukaryotic genomes. BMC
Evol Biol. 13: 249
Grande C, Patel NH (2009) Nodal signalling is involved in left-right
asymmetry in snails. Nature 457: 1007–1011
Guder C, Philipp I, Lengfeld T, Watanabe H, Hobmayer B, Holstein
TW (2006) The Wnt code: cnidarians signal the way. Oncogene
Page 13
D. H. E. Setiamarga et al.888
25: 7450–7460
Hao L, Johnsen R, Lauter G, Baillie D, Bürglin TR (2006)
Comprehensive analysis of gene expression patterns of hedge-
hog-related genes. BMC Genomics 7: 280
Heino TI, Kärpänen T, Wahlström G, Pulkkinen M, Eriksson U, Alitalo
K, Roos C (2001) The Drosophila VEGF receptor homolog is
expressed in hemocytes. Mech Dev 109: 69–77
Holmes DIR, Zachary I (2005) The vascular endothelial growth fac-
tor (VEGF) family: angiogenic factors in health and disease.
Genome Biol 6: 209
Huang P, Stern MJ (2005) FGF signaling in flies and worms: more
and more relevant to vertebrate biology. Cytokine Growth Factor
Rev 16: 151–158
Hui JHL, McDougall C, Monteiro AS, Holland PW, Arendt D,
Balavoine G, Ferrier DE (2012) Extensive chordate and annelid
macrosynteny reveals ancestral homeobox gene organization.
Mol Biol Evol 29: 157–165
Huminiecki L, Goldovsky L, Freilich S, Moustakas A, Ouzounis C,
Heldin CH (2009) Emergence, development and diversification
of the TGFβ signaling pathway within the animal kingdom. BMC
Evol Biol 9: 28
Ingham PW, Nakano Y, Seger C (2011) Mechanisms and functions
of Hedgehog signaling across the metazoan. Nat Rev Genet
12: 393–406
Janssen R, Le Gouar M, Pechmann M, Poulin F, Bolognesi R,
Schwager EE, et al. (2010) Conservation, loss, and redeploy-
ment of Wnt ligands in protostomes: implications for under-
standing the evolution of segment formation. BMC Evol Biol 10:
374
Kadam S, McMahon A, Tzou P, Stathopoulos A (2009) FGF ligands
in Drosophila have distinct activities required to support cell
migration and differentiation. Development 136: 739–747
Kin K, Kakoi S, Wada H (2009) Novel role for dpp in the shaping of
bivalve shells revealed in a conserved mollusk developmental
program. Dev Biol 329: 152–166
Kinoshita S, Wang N, Inoue H, Maeyama K, Okamoto K, Nagai K, et
al. (2011) Deep sequencing of ESTs from nacreous and pris-
matic layer producing tissues and a screen for novel shell for-
mation-related genes in the pearl oyster. PLoS One 6: e21238
Kusserow A, Pang K, Sturm C, Hrouda M, Lentfer J, Schmidt HA, et
al. (2005) Unexpected complexity of the Wnt gene family in a
sea anemone. Nature 433: 156–160
Letunic I, Doerks T, Bork P (2012) SMART 7: recent updates to the
protein domain annotation resource. Nucl Acids Res 40: D302–
D305
Maddison WP, Maddison DR (2011) Mesquite: a modular system for
evolutionary analysis. Version 2.75 http://mesquiteproject.org
Masaoka T, Kobayashi T (2007) Phylogeny and adaptive radiation
of the pearl oysters genus Pinctada. In “DNA Swimming in the
Water: Ontogeny, Phylogeny, Ecology, and Application of
Aquatic Organisms Revealed from DNA” Ed by Saruwatari S,
Tokai Univ Press, Tokyo pp 1–29 (in Japanese)
Matus DQ, Magie CR, Pang K, Martindale MQ, Thomsen GH (2008)
The Hedgehog gene family of the cnidarian, Nematostella
vectensis, and implications for understanding metazoan
Hedgehog pathway evolution. Dev Biol 313: 501–518
McGinty EL, Zenger KR, Jones DB, Jerry DR (2012) Transcriptome
analysis of biomineralisation-related genes within the pearl sac:
host and donor oyster contribution. Mar Genomics 5: 27–33
Nederbragt AJ, van Loon AE, Dictus WJ (2002) Expression of
Patella vulgata orthologs of engrailed and dpp-BMP2/4 in adja-
cent domains during mollusk shell development suggests a
conserved compartment boundary mechanism. Dev Biol 246:
341–355
Onai T, Takai A, Setiamarga DHE, Holland LZ (2012) Essential role
of Dkk3 for head formation by inhibiting Wnt/β-catenin and
Nodal/Vg1 signaling pathways in the basal chordate
amphioxus. Evol Dev 14: 338–350
Pani AM, Mullarkey EE, Aronowicz J, Assimacopoulos S, Grove EA,
Lowe CJ (2012) Ancient deuterostome origins of vertebrate
brain signalling centres. Nature 483: 289–294
Pei J, Kim BH, Grishin NV (2008) PROMALS3D: a tool for multiple
sequence and structure alignment. Nucleic Acid Res 36: 2295–
2300
Pietrokovski S, Shilo BZ (2001) Identification of new signaling com-
ponents in the Drosophila genome sequence. Funct Integr
Genomics 1: 250–255
Pires-daSilva A, Sommer RJ (2003) The evolution of signalling path-
ways in animal development. Nat Rev Genet 4: 39–49
Popovici C, Roubin R, Coulier F, Pontarotti P, Birnbaum D (1999)
The family of Caenorhabditis elegans tyrosine kinase recep-
tors: similarities and differences with mammalian receptors.
Genome Res 9: 1026–1039
Popovici C, Isnardon D, Birnbaum D, Roubin R (2002) Caenorhabditis
elegans receptors related to mammalian vascular endothelial
growth factor receptors are expressed in neural cells. Neurosci
Lett 329: 116–120
Saina M, Genikhovich G, Renfer E, Technau U (2009) BMPs and
chordin regulate patterning of the directive axis in a sea anem-
one. Proc Natl Acad Sci U S A 106: 18592–18597
Schubert MJ, Holland LZ, Holland ND, Jacobs DK (2000) A phyloge-
netic tree of the wnt genes based on all available full-length
sequences, including five from the cephalochordate amphioxus.
Mol Biol Evol 17: 1898–1903
Schultz J, Milpetz F, Bork P, Ponting CP (1998) SMART, a simple
modular architecture research tool: Identification of signaling
domains. Proc Natl Acad Sci U S A 95: 5857–5864
Simakov O, Marletaz F, Cho SJ, Edsinger-Gonzales E, Havlak P,
Hellsten U, et al. (2013) Insights into bilaterian evolution from
three spiralian genomes. Nature 493: 526–531
Stamatakis A, Hoover J, Rougemont J (2008) A rapid bootstrap
algorithm for the RAxML web-servers. Syst Biol 75: 758–771
Stathopoulos A, Tam B, Ronshaugen M, Frasch M, Levine M (2004)
pyramus and thisbe: FGF genes that patterns the mesoderm of
Drosophila embryos. Genes Dev 18: 687–699
Takeuchi T, Sarashina I, Iijima M, Endo K (2008) In vitro regulation
of CaCO3 crystal polymorphism by the highly acidic mollusk
shell protein Aspein. FEBS Lett 582: 591–596
Takeuchi T, Kawashima T, Koyanagi R, Gyoja F, Tanaka M, Ikuta T,
et al. (2012) Draft genome of the pearl oyster Pinctada fucata:
A platform for understanding bivalve biology. DNA Res [Epub
ahead of print]
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S
(2011) MEGA5: Molecular Evolutionary Genetics Analysis
using maximum likelihood, evolutionary distance, and maxi-
mum parsimony methods. Mol Biol Evol 28: 2731–2739
Van der Zee M, da Fonseca RN, Roth S (2008) TGF signaling in
Tribolium: vertebrate-like components in a beetle. Dev Genes
Evol 218: 203–213
Yasuoka Y, Kobayashi M, Kurokawa D, Akasaka K, Saiga H, Taira
M (2009) Evolutionary origins of blastoporal expression and
organizer activity of the vertebrate gastrula organizer gene lhx1
and its ancient metazoan paralog lhx3. Development 136:
2005–2014
Zhang G, Fang X, Guo X, Li L, Luo R, Xu F, et al. (2012) The oyster
genome reveals stress adaptation and complexity of shell for-
mation. Nature 490: 49–54
Zhu CC, Boonr JQ, Jensen PA, Hanna S, Podemski L, Locke J, et
al. (2008) Drosophila Activin-β and the Activin-like product
Dawdle function redundantly to regulate proliferation in the lar-
val brain. Development 135: 513–521
(Received October 23, 2012 / Accepted March 23, 2013)
























![CHARACTERIZATION OF VARIETIES OF FANO … · arxiv:1201.1133v1 [math.ag] 5 jan 2012 characterization of varieties of fano type via singularities of cox rings yoshinori gongyo, shinnosuke](https://static.documents.pub/doc/80x56/5b96f52509d3f2e10f8c057d/characterization-of-varieties-of-fano-arxiv12011133v1-mathag-5-jan-2012.jpg)


