VIROLOGY 176,403-4 12 (1990) Replication of Minute Virus of Mice DNA in Adenovirus-Infected or Adenovirus-Transformed Cells ELIZABETH FOX, PHILLIP T. MOEN, JR., AND JOHN W. BODNAR’ Department of Biology, Northeastern University* 360 Huntington Avenue, Boston, Massachusetts 02 115 Received September 26, 1989; accepted February 13, 1990 The effect of adenovirus infection or transformation on the DNA replication of Minute Virus of Mice (MVM) was studied in human fibroblast cell lines. In Wl38, HeLa, and 293 cells MVM infection allowed production of viral NS-1 and capsid proteins with or without adenovirus 2 (Ad2) co-infection. However, MVM DNA replication varied markedly. In HeLa cells MVM DNA was replicated weakly in host nucleoli, and replication was increased markedly by Ad2 co-infec- tion as well as recompartmentalized to Ad2 replication factories. In Ad-transformed 293 cells MVM DNA was replicated very efficiently when infected alone or with Ad2 co-infection although recompartmentalization from nucleoli to replica- tion factories was also seen. In WI38 cells MVM DNA was not replicated under any conditions. The variation in DNA replication in Wl38, HeLa, and 293 cells despite viral protein production in all cases suggests that MVM DNA replication is uncoupled from viral gene expression and that host factors required for MVM DNA replication are induced or recom- partmentalized by adenovirus infection or transformation. 0 1990 Academic Press, Inc. INTRODUCTION Parvoviruses are extremely sensitive to the growth state of the cells they infect, and in general will repli- cate their DNAs only in cells which are actively dividing or which have been induced to replicate their DNA by another means. In animal hosts parvoviruses grow mainly in tissues that are rapidly proliferating such as fetal cells, bone marrow cells, or gut epithelium (re- viewed by Cotmore and Tattersall, 1987). Targeting of proliferating cells can extend to tumor cells in that par- voviruses can be found in animal tumors (Cotmore and Tattersall, 1987); rodents chronically infected with par- voviruses and rodents injected with parvoviruses along with oncogenic viruses may have a reduced incidence of tumors (Toolan, 1967; Toolan and Ledinko, 1968; de la Maza and Carter, 1981; Toolan et al., 1982; Guetta et al., 1986a). This correlation of parvovirus growth with cell prolif- eration or DNA replication extends to studies in tissue culture. In normal hosts parvovirus DNA replication oc- curs only during cell S phase and depends heavily on host factors induced during S phase (Wolter et al., 1980; Cotmore and Tattersall, 1987). Parvovirus DNA replication or cell killing can extend to normally nonper- missive host cells if those cells are transformed by any of several agents including radiation, chemical carcino- gens, or oncogenic viruses (Cukor el al., 1975; Mous- set and Rommelaere, 1982; Heilbronn et al., 1984; Guetta et a/., 1986a; Mousset et al., 1986; Bantel- ’ To whom requests for reprints should be addressed. Schaal and zur Hausen, 1988; Cornelis eta/., 1988a,b). Finally, DNA replication of both defective and autono- mous parvoviruses can be induced in appropriate hosts by co-infection with an adenovirus or herpesvirus helpervirus (Ledinko eta/., 1969; Berns and Hauswirth, 1978; Janik et al., 1981). Parvovirus DNA replication is, therefore, tightly coupled to the proliferative state of the host requiring that the host be induced to be repli- cationally competent as par-t of the normal host cell proliferation, as part of cell transformation, or as part of lytic growth of a helper virus. Minute Virus of Mice, prototype strain (MVM{p}), normally grows in mouse fibroblasts, but its host range can be extended to human fibroblasts if they are trans- formed chemically, by radiation, or by SV40 (Mousset and Rommelaere, 1982; Mousset eta/., 1986; Cornelis et al., 1988a). Previous studies indicated that MVM DNA replication depends on a host factor expressed late in S phase (Wolteretal., 1980; Guetta eta/., 1986b; Cotmore and Tattersall, 1987) and that MVM(p) replica- tion occurs at substantial levels in mouse lymphocytes in spite of very low levels of viral transcription (Spalholz et al., 1983). Adenovirus can both transform cells and serve as a parvovirus helper virus, but few studies have been done with autonomous parvoviruses to investigate if there is any correlation between these two propenies. We, therefore, have begun to study human cell lines for their ability to replicate MVM DNA and for changes in MVM DNA replication induced by adenovirus coinfec- tion or by adenovirus transformation. We have concen- trated on standard well-studied cell lines since any data 403 0042.6822/90 $3.00 CopyrIght 0 1990 by Academic Press. Inc. All rtghis of reproductw I” any form reserved.
Transcript
VIROLOGY 176,403-4 12 (1990)
Replication of Minute Virus of Mice DNA in Adenovirus-Infected or Adenovirus-Transformed Cells
ELIZABETH FOX, PHILLIP T. MOEN, JR., AND JOHN W. BODNAR’
Department of Biology, Northeastern University* 360 Huntington Avenue, Boston, Massachusetts 02 115
Received September 26, 1989; accepted February 13, 1990
The effect of adenovirus infection or transformation on the DNA replication of Minute Virus of Mice (MVM) was studied in human fibroblast cell lines. In Wl38, HeLa, and 293 cells MVM infection allowed production of viral NS-1 and capsid proteins with or without adenovirus 2 (Ad2) co-infection. However, MVM DNA replication varied markedly. In HeLa cells MVM DNA was replicated weakly in host nucleoli, and replication was increased markedly by Ad2 co-infec- tion as well as recompartmentalized to Ad2 replication factories. In Ad-transformed 293 cells MVM DNA was replicated very efficiently when infected alone or with Ad2 co-infection although recompartmentalization from nucleoli to replica- tion factories was also seen. In WI38 cells MVM DNA was not replicated under any conditions. The variation in DNA replication in Wl38, HeLa, and 293 cells despite viral protein production in all cases suggests that MVM DNA replication is uncoupled from viral gene expression and that host factors required for MVM DNA replication are induced or recom- partmentalized by adenovirus infection or transformation. 0 1990 Academic Press, Inc.
INTRODUCTION
Parvoviruses are extremely sensitive to the growth state of the cells they infect, and in general will repli- cate their DNAs only in cells which are actively dividing or which have been induced to replicate their DNA by another means. In animal hosts parvoviruses grow mainly in tissues that are rapidly proliferating such as fetal cells, bone marrow cells, or gut epithelium (re- viewed by Cotmore and Tattersall, 1987). Targeting of proliferating cells can extend to tumor cells in that par- voviruses can be found in animal tumors (Cotmore and Tattersall, 1987); rodents chronically infected with par- voviruses and rodents injected with parvoviruses along with oncogenic viruses may have a reduced incidence of tumors (Toolan, 1967; Toolan and Ledinko, 1968; de la Maza and Carter, 1981; Toolan et al., 1982; Guetta et al., 1986a).
This correlation of parvovirus growth with cell prolif- eration or DNA replication extends to studies in tissue culture. In normal hosts parvovirus DNA replication oc- curs only during cell S phase and depends heavily on host factors induced during S phase (Wolter et al., 1980; Cotmore and Tattersall, 1987). Parvovirus DNA replication or cell killing can extend to normally nonper- missive host cells if those cells are transformed by any of several agents including radiation, chemical carcino- gens, or oncogenic viruses (Cukor el al., 1975; Mous- set and Rommelaere, 1982; Heilbronn et al., 1984; Guetta et a/., 1986a; Mousset et al., 1986; Bantel-
’ To whom requests for reprints should be addressed.
Schaal and zur Hausen, 1988; Cornelis eta/., 1988a,b). Finally, DNA replication of both defective and autono- mous parvoviruses can be induced in appropriate hosts by co-infection with an adenovirus or herpesvirus helpervirus (Ledinko eta/., 1969; Berns and Hauswirth, 1978; Janik et al., 1981). Parvovirus DNA replication is, therefore, tightly coupled to the proliferative state of the host requiring that the host be induced to be repli- cationally competent as par-t of the normal host cell proliferation, as part of cell transformation, or as part of lytic growth of a helper virus.
Minute Virus of Mice, prototype strain (MVM{p}), normally grows in mouse fibroblasts, but its host range can be extended to human fibroblasts if they are trans- formed chemically, by radiation, or by SV40 (Mousset and Rommelaere, 1982; Mousset eta/., 1986; Cornelis et al., 1988a). Previous studies indicated that MVM DNA replication depends on a host factor expressed late in S phase (Wolteretal., 1980; Guetta eta/., 1986b; Cotmore and Tattersall, 1987) and that MVM(p) replica- tion occurs at substantial levels in mouse lymphocytes in spite of very low levels of viral transcription (Spalholz et al., 1983).
Adenovirus can both transform cells and serve as a parvovirus helper virus, but few studies have been done with autonomous parvoviruses to investigate if there is any correlation between these two propenies. We, therefore, have begun to study human cell lines for their ability to replicate MVM DNA and for changes in MVM DNA replication induced by adenovirus coinfec- tion or by adenovirus transformation. We have concen- trated on standard well-studied cell lines since any data
403 0042.6822/90 $3.00 CopyrIght 0 1990 by Academic Press. Inc. All rtghis of reproductw I” any form reserved.
404 FOX, MOEN, AND BODNAR
we collect can be more easily correlated to known data about the cells or adenovirus infection of those cells. We have found that MVM DNA is replicated weakly in HeLa cells and that MVM DNA replication is increased and recompar-tmentalized by adenovirus 2 (Ad2) co-in- fection. MVM DNA is efficiently replicated in Ad5-trans- formed 293 cells and adenovirus co-infection does not increase MVM replication but does recompat-tmental- ize it. WI38 cells do not support MVM DNA replication with or without Ad2 co-infection and serve as our con- trol human cell line. Comparative studies were done in these cell lines of MVM protein production, MVM DNA replication, and intranuclear compartmentalization of viral and host proteins as well as MVM DNA. These studies indicated that MVM viral gene expression and DNA replication are uncoupled and that host factors can be induced or recompartmentalized by adenovirus infection or transformation to allow MVM DNA replica- tion in human cells.
MATERIALS AND METHODS
Cells and viruses
HeLa cells and 293 cells were provided by Dr. Marie Chow (Massachusetts Institute of Technology). Mouse A9 cells and 324K cells were provided by Dr. David Ward (Yale University). WI38 cells (ATCC CCL75) were obtained from the American Type Culture Collection. Cells were grown and infected with MVM(p} and/or Ad2 as previously described (Walton et al., 1989).
Localization of proteins and DNA in situ
Antisera against NS-1 (Cotmore and Tattersall, 1988) and MVM capsids were provided by Drs. Susan Cotmore and Peter Tattersall (Yale University). Antise- rum to Nucleolar Factor I (previously described as “MVM Terminal Protein”) was raised using purified MVM DNA-terminal protein complexes (Chow et a/., 1986). FITC-conjugated antibodies to adenovirus 2 viri- ons were obtained from Chemicon International. Cloned MVM DNA (pMM984) as described by Merch- linsky et al. (1983) was provided by Dr. David Ward (Yale University).
Cells were grown on microscope slides, infected with MVM(p) and/or Ad2, fixed at 24 hr postinfection, and proteins or DNA detected in situ as previously de- scribed (Walton et a/., 1989).
Extraction and detection of MVM and Ad2 DNA
Total DNA was extracted from infected cells as pre- viously described (Bodnar et al., 1989). Briefly cells were lysed with 0.19/o sodium dodecyl sulfate (SDS), and the cell lysates were incubated for 2 hr at 37” with
Proteinase K (200 pg/ml). Following extraction with phenol DNA preparations were further digested for 2 hr at 37” with Ribonuclease A (10 pg/ml). DNA samples were separated by agarose gel electrophoresis, trans- ferred to nitrocellulose, and the MVM DNA sequences detected by hybridization with cloned MVM DNA probes (pMM984) as described by Merchlinsky et a/. (1983). Ad2 DNA sequences were detected on identi- cal blots probed as before with Ad2 virion DNA.
Plaque assays for infectious MVM virions
Cells were grown in monolayers and infected with MVM (m.o.i. = 0.1) or co-infected with MVM (m.o.i. = 0.1) and Ad2 (m.o.i. = 0.2) as described above. In- fected cells were grown for 48 hr and the supernatants harvested. The supernatents were then assayed for in- fectious MVM virions on A9 cell monolayers as de- scribed by Merchlinsky et al. (1983). Note that Ad2 does not cause any cell lysis in A9 cells; therefore, MVM in MVM plus Ad2 co-infections could be titered directly.
RESULTS
Parvovirus DNA replication depends heavily on host factors (Wolter et al., 1980; Cotmore and Tattersall, 1987), and parvovirus replication can be extended to previously nonpermissive cell types by co-infection with a helper virus or by cell transformation by a variety of agents (Ledinko et a/., 1969; Berns and Hauswirth, 1978; Janik et al., 1981; Cukor et al., 1975; Mousset and Rommelaere, 1982; Heilbronn et al., 1984; Guetta et al., 1986a; Mousset et al., 1986; Bantel-Schaal and zur Hausen, 1988; Cornelis et al., 1988a,b). Therefore, identification of host factors required for patvovirus DNA replication may be possible by comparison of how it is modulated by a helper virus infection and viral transformation in similar cell types. We chose to begin such an investigation by studying the interactions of Minute Virus of Mice (MVM) DNA replication with ade- novirus. This system was chosen for the following rea- sons: (1) adenovirus has shown to be a helper virus for parvoviruses (Berns and Hauswirth, 1978) and can help viral growth of parvovirus H-l in at least one hu- man cell line (Ledinko et a/., 1969); (2) adenovirus is an oncogenic virus and might, therefore, induce MVM DNA replication in adenovirus-transformed cells; (3) both MVM and adenovirus 2 (Ad2) interact with host nucleoli (Walton et a/., 1989); and (4) preliminary stud- ies have shown that Ad2 can recompar-tmentalize MVM DNA replication in HeLa cells (Walton et a/., 1989). Studies on several cell lines (including mouse, hamster, monkey, and human lines-Fox and Bodnar, unpublished data) indicated changes in MVM DNA rep-
MVM-ADENOVIRUS INTERACTIONS 405
lication induced by Ad2 co-infection or by cell transfor- mation. We have chosen to study two human cell lines in detail to begin to investigate the effects of adenovi- rus coinfection and adenovirus transformation on MVM DNA replication.
Our test cell lines were HeLa cells (an extremelywell- studied human cervical carcinoma line which we used in our previous studies of Ad2 DNA replication- Bodnar and Pearson, 1980a,b; Walton eta/., 1989) and 293 cells (a standard human fibroblast line transformed by Ad5 containing multiple copies of the Ad5 El A/El B genes and exhibiting a highly transformed pheno- type-Graham et a/., 1974; Aiello et a/., 1979). As con- trol human cell lines we used WI38 cells (a human em- bryonic lung line exhibiting few characteristics of the transformed phenotype) and 324K cells (an SV40- transformed human line which has been studied for its ability to replicate MVM DNA and as a plaquing cell line for both MVM(p) and MVM(i) variants-Spalholz et al., 1983).
Experiments with HeLa and 293 cells (and the WI38 control) were designed to follow how far each cell line could support the MVM lytic cycle. The MVM lytic cy- cle follows sequentially: penetration, uncoating, tran- scription and translation of nonstructural proteins NS- 1 and NS-2 from the left (P4) promoter, transcription and translation of capsid proteins VP-l and VP-2 from the middle (P38) promoter, viral DNA replication, en- capsidation of single-stranded vDNA, cell lysis, and re- lease of infectious virions (reviewed in Cotmore and Tattersall, 1987). Transcription of the P38 promoter ap- pears to require transactivation by MVM protein NS-1 and P38 transcription follows almost immediately after P4 transcription (Cotmore and Tattersall, 1987; Doerig eta/., 1988; Tullis eta/., 1988). We were also interested in the intranuclear localization of MVM proteins and DNA since previous experiments have indicated that MVM DNA replication is associated with cell nucleoli and that this localization may be changed by Ad2 co- infection (Walton et al., 1989).
Biochemical data have indicated the presence of a protein covalently bound to the 5’termini of MVM DNA (Astell et a/., 1983). The isolation of antiserum to the MVM “terminal protein” was previously described (Chow et a/., 1986); this antiserum was raised against purified MVM DNA-terminal protein complexes and detects a 60-kDa antigen that is apparently host-en- coded (Chow et a/., 1986; Walton et al., 1989). Cot- more and Tattersall (1988) have found MVM-encoded protein NS-1 bound to MVM DNA termini. Therefore, there is apparently a complex of NS-1 and a host-en- coded 60-kDa protein bound to the MVM DNA termini.
Currently there is no direct proof whether NS-1 or the host-encoded protein is actually covalently attached to
MVM DNA. Therefore, to avoid confusion in the char- acterization of the proteins complexed with the MVM DNA termini we suggest the following nomenclature: (1) the host-encoded protein previously called “MVM Terminal Protein” will be called Nucleolar Factor I (NuFI) based on its characterization as a host nucleolar protein which is associated with the MVM DNA replica- tion origin and localized at sites of MVM DNA replica- tion (Chow et a/., 1986; Walton et a/., 1989); (2) the MVM-encoded nonstructural protein NS-1 will be used when functions are described which have been defini- tively attributed to it (Cotmore and Tattersall, 1987; Cotmore and Tattersall, 1988); and (3) the description “covalent terminal protein” will be used for functions assigned to a protein covalently attached to MVM DNA termini but not yet definitively shown to be either NS- 1 or NuFl (e.g., covalent attachment, nicking-closing during processing of MVM DNA termini, attachment of MVM DNA to the nuclear matrix-Astell et a/., 1983, Chow et a/., 1986, Bodnar et a/., 1989).
Synthesis and intranuclear localization of MVM gene products and Nucleolar Factor I
We first investigated the synthesis of MVM gene products and Nucleolar Factor I in Wl38, HeLa, and 293 cells. Cells grown on microscope slides were mock-infected, Ad2-infected, MVM-infected, or co-in- fected with MVM and Ad2 and fixed 24 hr postinfection with methanol:acetone as previously described (Wal- ton et al., 1989). The fixed cells were probed in situ for the presence and intranuclear localization of MVM nonstructural protein NS-1, MVM capsid proteins, Nu- cleolar Factor I, and Ad2 virions using appropriate anti- sera.
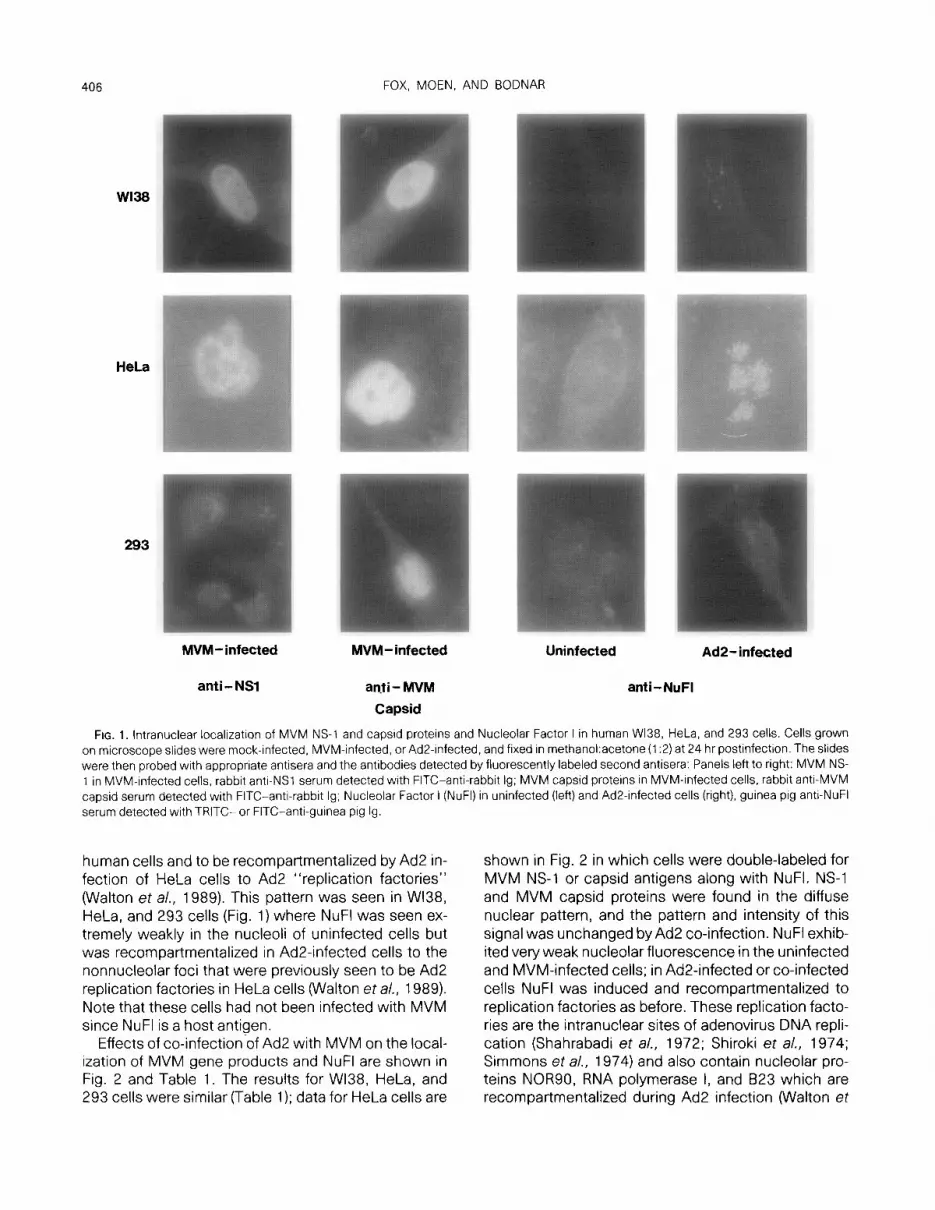
The results of the experiments to identify MVM pro- teins in situ are summarized in Fig. 1 and Table 1. In Wl38, HeLa, and 293 cells infected with MVM alone both MVM NS-1 and capsid proteins were found in a diffuse nuclear pattern often with nucleolar exclusion (Fig. 1). Therefore, experiments were not performed to measure MVM penetration, uncoating, and transcrip- tional regulation since all these steps must occur to produce the MVM gene products. Note that synthesis of both NS-1 and capsid proteins indicates that tran- scription from both P4 and P38 promoters occurs in the human cell lines tested. Also, there was no major difference in uptake of MVM virions in those cell types because the fraction of cells which were positive for MVM capsid fluorescence was between 26 and 52% (Table 1). Similar experiments with 324K cells gave similar results (Table 1).
NuFl (previously called “MVM Terminal Protein”) has been shown to localize to nucleoli in mouse and
406 FOX, MOEN, AND BODNAR
MVM - infected
anti - NSl
MVM- infected
anti - MVM
Capsid
Uninfected
anti-NuFI
Ad2- infected
FIG. 1. lntranuclear localizatron of MVM NS-1 and capsrd proteins and Nucleolar Factor I in human Wl38, HeLa, and 293 cells. Cells grown on microscope slides were mock-infected, MVM-infected. or Ad2-infected, and fixed in methanoLacetone (1:2) at 24 hr postinfection. The slides were then probed wrth appropriate antisera and the antibodies detected by fluorescently labeled second antisera: Panels left to right: MVM NS- 1 in MVM-infected cells, rabbit antr-NSl serum detected wrth FITC-anti-rabbit lg; MVM capsid proteins in MVM-Infected cells, rabbit anti-MVM capsid serum detected with FITC-anti-rabbit lg; Nucleolar Factor I (NuFI) in uninfected (left) and Ad2infected cells (right), guinea pig anti-NuFI serum detected with TRITC- or FITC-anti-guinea pig lg
human cells and to be recompartmentalized by Ad2 in- fection of HeLa cells to Ad2 “replication factories” (Walton et a/., 1989). This pattern was seen in Wl38, HeLa, and 293 cells (Fig. 1) where NuFl was seen ex- tremely weakly in the nucleoli of uninfected cells but was recompartmentalized in Ad2-infected cells to the nonnucleolar foci that were previously seen to be Ad2 replication factories in HeLa cells (Walton et al., 1989). Note that these cells had not been infected with MVM since NuFl is a host antigen.
Effects of co-infection-of Ad2 with MVM on the local- ization of MVM gene products and NuFl are shown in Fig. 2 and Table 1. The results for Wl38, HeLa, and 293 cells were similar (Table 1); data for HeLa cells are
shown in Fig. 2 in which cells were double-labeled for MVM NS-1 or capsid antigens along with NuFI. NS-1 and MVM capsid proteins were found in the diffuse nuclear pattern, and the pattern and intensity of this signal was unchanged by Ad2 co-infection. NuFl exhib- ited very weak nucleolar fluorescence in the uninfected and MVM-infected cells; in Ad2-infected or co-infected cells NuFl was induced and recompartmentalized to replication factories as before. These replication facto- ries are the intranuclear sites of adenovirus DNA repli- cation (Shahrabadi et al., 1972; Shiroki et al., 1974; Simmons et a/., 1974) and also contain nucleolar pro- teins NOR90, RNA polymerase I, and B23 which are recompartmentalized during Ad2 infection (Walton et
MVM-ADENOVIRUS INTERACTIONS 407
TABLE 1
PROTEIN PRODUCTION IN MVM-INFECTED HUMAN CELLS
NS-1 Capsid NuF-I % MVM-infected
WI38 U - - +- (4 A - - ++ (r) M ++ (d) ++ (4 +- (4 27 B ++ (4 ++ (4 ++ t)
HeLa U - - +- (4 A - - +++ (r) M +++(d) +++(d) +-(n) 56 B +++ (d) +++(d) +++(r)
293 U - - -
A - - ++ (d M ++ (4 ++ (4 - 26 B ++ (4 ++ (4 ++ (4
324K U - +- (n) A - +++ (r) M +++(d) +- (4 26 B +++(d)
Note. n. nucleolar; d, nuclear diffuse (nucleolar exclusion); r, repli- cation factories; U, uninfected; A, AdP-infected; M, MVM-infected; B, Ad2 + MVM-infected.
al., 1989). The Ad2 virion antigens were found in a diffuse nuclear pattern, and only in cells in which NuFl was found in replication factory patterns; time course experiments showed the appearance of Ad2 virion anti- gens later in infection (20-24 hr) than appearance of the NuFl foci (16-20 hr) (data not shown).
Replication of MVM DNA
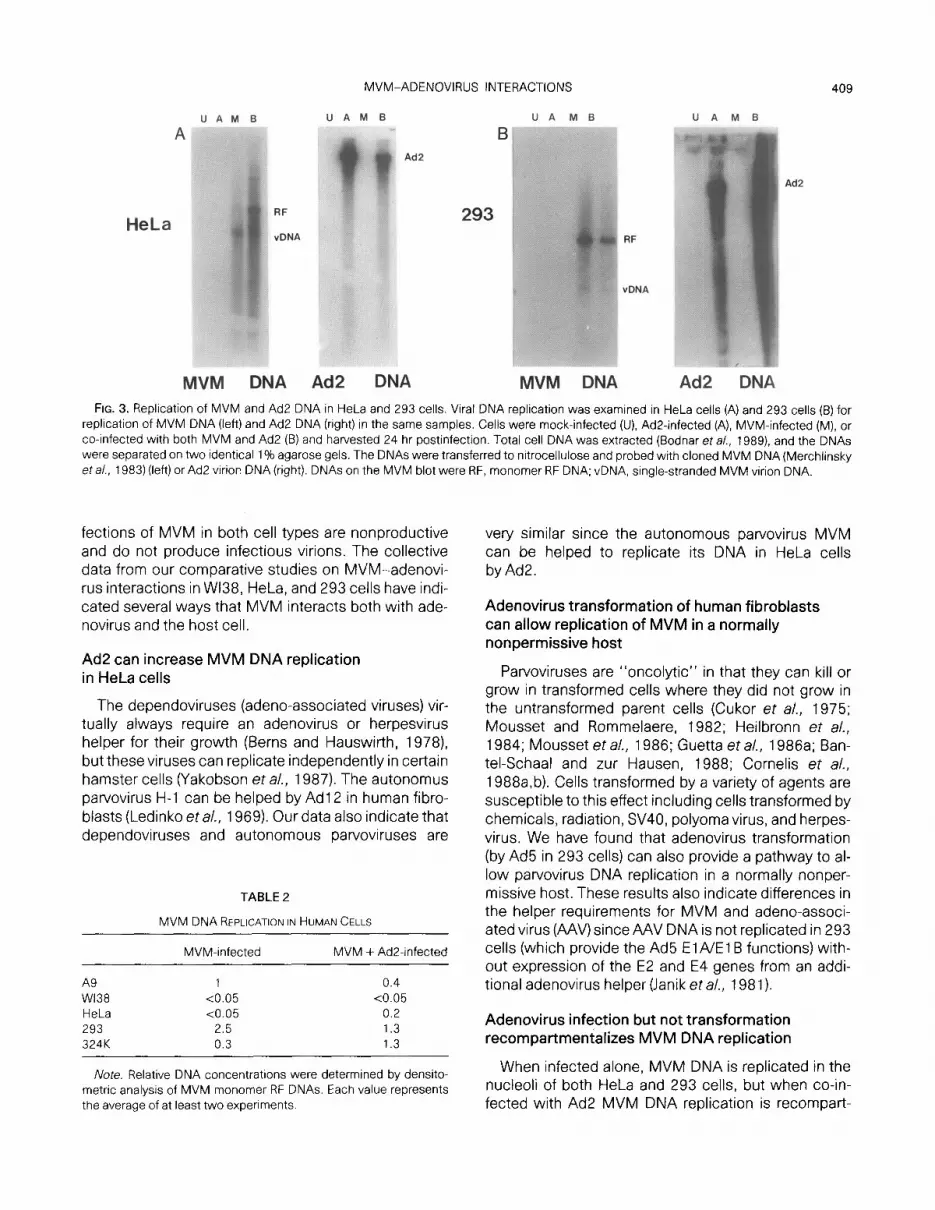
The cell lines were next tested for their ability to repli- cate with and without Ad2 co-infection. Cells were mock-infected, Ad2-infected, MVM-infected, or co-in- fected with MVM and Ad2 in cell monolayers, and total cell DNA was isolated using SDS/protease followed by phenol extraction and ethanol precipitation (Bodnar et al., 1989). DNA samples from equal numbers of cells were separated on two identical agarose gels and transferred to nitrocellulose; one gel was probed with cloned MVM DNA (Merchlinksy et al., 1983) while the second was probed with Ad2 virion DNA. Autoradio- grams of Southern blots for HeLa and 293 cells are shown in Fig. 3; densitometric analysis of the relative amount of MVM monomer Replicative Form (RF) DNA synthesized in all cells tested is shown in Table 2.
Major differences in replication of MVM DNA were observed in the test cell lines. In HeLa cells MVM DNA
was replicated weakly when infected alone, but MVM DNA replication was increased by Ad2 co-infection to almost 20% of levels seen in mouse A9 cells. In 293 cells and 324K cells both MVM and Ad2 were repli- cated well when infected alone or together, and accu- mulation of MVM DNA in 293 cells was over double that seen in A9 cells. WI38 cells did not support any MVM DNA replication with or without Ad2 co-infection.
lntranuclear localization of MVM DNA replication
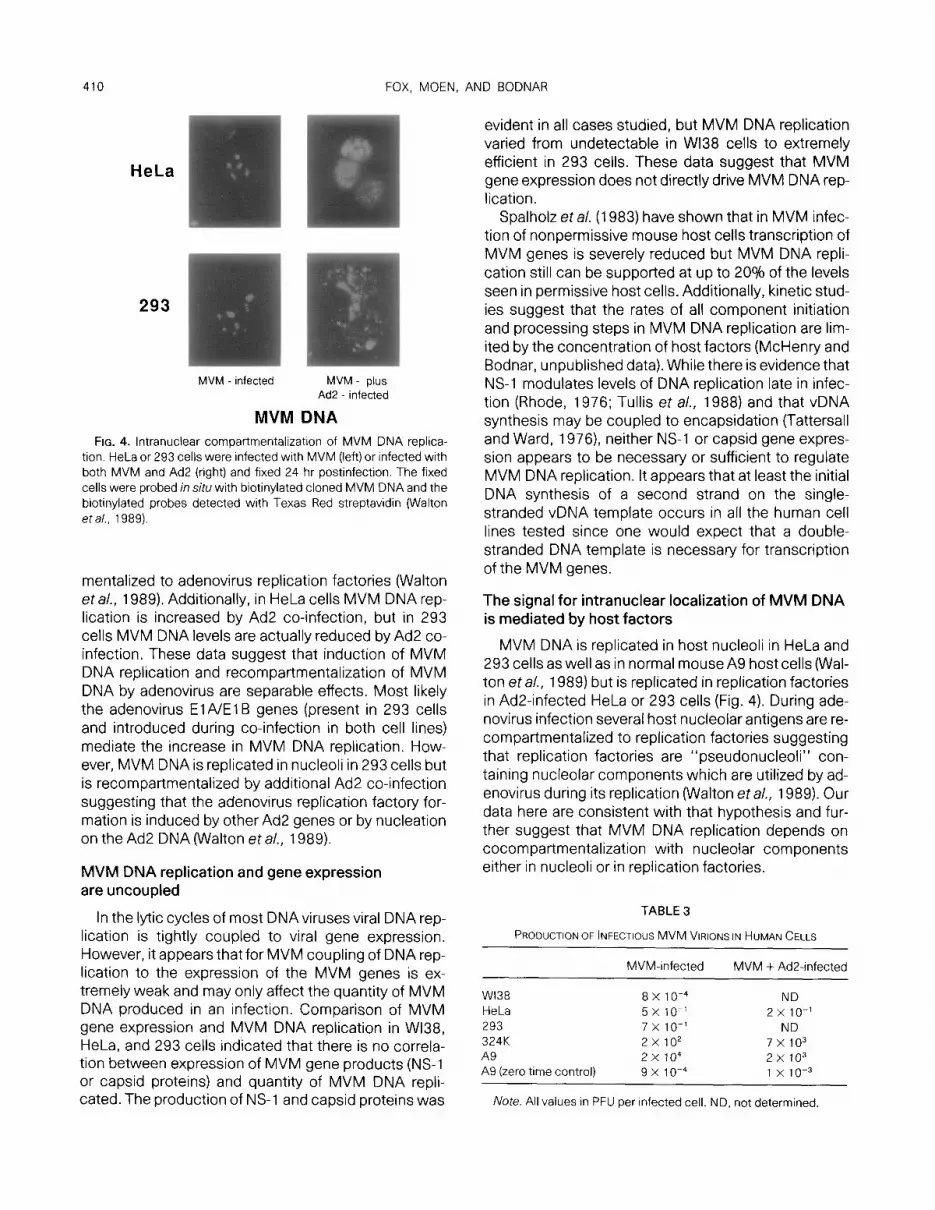
HeLa cells and 293 cells were further tested to deter- mine the intranuclear localization of MVM DNA replica- tion (Fig. 4). Cells infected on microscope slides were fixed 24 hr postinfection and probed in situ with biotin- ylated MVM DNA, and the probe DNA was detected with fluorescently labeled streptavidin as previouslyde- scribed (Walton et al., 1989). In cells infected with MVM alone the MVM DNA was found in a few foci within the nucleus; these foci were determined to be nucleoli by comparison of the fluorescent signals with phase contrast microscopy as previously shown for MVM infections in mouse A9 cells (Walton eta/., 1989). When HeLa and 293 cells were co-infected with MVM and Ad2, the fluorescence was found in multiple foci in the replication factory pattern seen for NuFI.
Production of infectious MVM virions
The final stage in MVM infection is encapsidation and release of infectious virions where only single- stranded DNA is packaged into MVM virions (Cotmore and Tattersall, 1987). Capsid proteins were synthe- sized in all cell lines tested (see Table l), and single- stranded DNA was seen in 293 cells and Ad2-infected HeLa cells (Fig. 3). However, we were interested in whether these cells could assemble these compo- nents into infectious virions.
Wl38, HeLa, and 293 cells (as well as mouse A9 and 324K controls) were tested by plaque assay for produc- tion of infectious MVM virions (Merchlinsky et al., 1983). Cells were MVM-infected or MVM- plus Ad2- infected (m.o.i.s: MVM, 0.1, Ad2, 0.2 PFU/cell), the ex- cess virions washed off after 30 minutes of adsorption, and the supernatents from the infected cells collected after 48 hr of infection. These supernatents were then used to infect A9 cells, and MVM plaques were counted after 7 days. As shown in Table 3 the output of MVM virions in A9 cells was about 20,000 PFU/in- fected cell and the residual adsorption to A9 cells was about 1 X 10m3 PFU/cell (as determined by harvesting supernatents from A9 cells directly following infection).
The production of infectious MVM virions was less than 1 PFU/infected cell for both HeLa cell and 293 cells; Ad2 co-infection of HeLa cells did not increase
FOX, MOEN. AND BODNAR
Anti - NuFl Anti- MVM Capsid
Anti-NuFI Anti -NSl
FIG. 2. lntranuclear localization of MVM proteins in HeLa cells. HeLa cells were Infected with both MVM and Ad2 and fixed 24 hr postrnfection. Cells were double-probed in situ with (A) anti-NuFI serum and anti-MVM capsid serum or(B) anti-NuFI serum and anti-N%1 serum. The antibod- res detected were with appropriate second antibodies conjugated with TRITC (left) or FITC (center) and compared with the same cells under phase contrast microscopy (right). Srnce NuFl recompartmentalization to replication factories is diagnostic of AD2 infection (data not shown), cells could be identified by the presence of NuFl and/or NS-1 and capsrds as U, uninfected; A, Ad2-infected; M, MVM-infected; B, infected with both MVM and Ad2
that yield. WI38 cells yielded less than 10e3 PFU/in- fected cell while the positive control 324K human line yielded 200 PFU/infected cell.
DISCUSSION
Parvovirus growth is highly dependent on the prolif- erative state of the host cell. In general parvoviruses grow in cells which are replicating or dividing normally (as in bone marrow or fetal cells) or aberrantly (as in tumor cells) or are triggered to replication competence by a helper virus (Berns and Hauswirth, 1978; Cotmore and Tattersall, 1987). Therefore, it appears that most of the major regulatory factors for parvovirus DNA repli- cation are host-encoded (Chow et al., 1986; Cotmore andTattersall, 1987; Guetta eta/., 1986b; Walton eta/., 1989). We have begun a systematic approach aimed toward identifying host factors which regulate Minute Virus of Mice (MVM) DNA replication by studying changes in host cells induced by adenovirus infection
or adenovirus transformation that allow MVM DNA rep- lication.
Here we have identified human host cells in which MVM DNA replication can be induced or recompart- mentalized by adenovirus infection or transformation. We have investigated the extent to which MVM(p) Iytic growth is supported in three human fibroblast cell lines (Wl38, HeLa, and 293). All three lines can support viral infection up to the production of viral NS-1 and capsid proteins. WI38 cells do not replicate MVM DNA with or without adenovirus co-infection. HeLa cells can repli- cate MVM DNA weakly in nucleoli, and MVM DNA replication is increased and recompartmentalized to adenovirus replication factories by adenovirus 2 (Ad2) co-infection. MVM DNA is replicated efficiently in ade- novirus-transformed 293 cells with or without Ad2 co- infection, but the co-infection causes recompartmen- talization of MVM DNAfrom 293 nucleoli to adenovirus replication factories. However, the adenovirus “helper” effect in HeLa and 293 cells is not complete since in-
MVM-ADENOVIRUS INTERACTIONS 409
HeLa
UAMB UA MB
Ad2
I3
293
U A M B
Ad2
vDNA
MVM DNA Ad2 DNA MVM DNA Ad2 DNA FIG. 3. Replication of MVM and Ad2 DNA in HeLa and 293 cells. Viral DNA replication was examined in HeLa cells (A) and 293 cells (6) for
replication of MVM DNA (left) and Ad2 DNA (right) in the same samples. Cells were mock-infected (U), Ad2infected (A), MVM-infected (M), or co-infected with both MVM and Ad2 (B) and harvested 24 hr postinfection. Total cell DNA was extracted (Bodnar et a/., 1989) and the DNAs were separated on two identical 1% agarose gels. The DNAs were transferred to nltrocellulose and probed with cloned MVM DNA (Merchlinsky eta/., 1983) (left) or Ad2 virion DNA (right). DNAs on the MVM blot were RF, monomer RF DNA; vDNA, single-stranded MVM virion DNA.
fections of MVM in both cell types are nonproductive and do not produce infectious virions. The collective data from our comparative studies on MVM-adenovi- rus interactions in Wl38, HeLa, and 293 cells have indi- cated several ways that MVM interacts both with ade- novirus and the host cell.
Ad2 can increase MVM DNA replication in HeLa cells
The dependoviruses (adeno-associated viruses) vir- tually always require an adenovirus or herpesvirus helper for their growth (Berns and Hauswirth, 1978) but these viruses can replicate independently in certain hamster cells (Yakobson et a/., 1987). The autonomus parvovirus H-l can be helped by Ad1 2 in human fibro- blasts (Ledinko eta/., 1969). Our data also indicate that dependoviruses and autonomous patvoviruses are
Ivote. Relative DNA concentrations were determined by densito- metric analysis of MVM monomer RF DNAs. Each value represents the average of at least two experiments.
very similar since the autonomous parvovirus MVM can be helped to replicate its DNA in HeLa cells by Ad2.
Adenovirus transformation of human fibroblasts can allow replication of MVM in a normally nonpermissive host
Parvoviruses are “oncolytic” in that they can kill or grow in transformed cells where they did not grow in the untransformed parent cells (Cukor et a/., 1975; Mousset and Rommelaere, 1982; Heilbronn et al., 1984; Mousset et al., 1986; Guetta et a/., 1986a; Ban- tel-Schaal and zur Hausen, 1988; Cornelis et al., 1988a,b). Cells transformed by a variety of agents are susceptible to this effect including cells transformed by chemicals, radiation, SV40, polyoma virus, and herpes- virus. We have found that adenovirus transformation (by Ad5 in 293 cells) can also provide a pathway to al- low parvovirus DNA replication in a normally nonper- missive host. These results also indicate differences in the helper requirements for MVM and adeno-associ- ated virus (AAV) since AAV DNA is not replicated in 293 cells (which provide the Ad5 El A/El B functions) with- out expression of the E2 and E4 genes from an addi- tional adenovirus helper (Janik et al., 1981).
Adenovirus infection but not transformation recompartmentalizes MVM DNA replication
When infected alone, MVM DNA is replicated in the nucleoli of both HeLa and 293 cells, but when co-in- fected with Ad2 MVM DNA replication is recompart-
410 FOX, MOEN, AND BODNAR
HeLa
293
I MVM - infected MVM plus
Ad2 - infected
MVM DNA FIG. 4. lntranuclear compartmentalizatron of MVM DNA replica-
tion. HeLa or 293 cells were infected with MVM (left) or infected with both MVM and Ad2 (right) and fixed 24 hr postinfection. The fixed cells were probed in situ with biotinylated cloned MVM DNA and the biotinylated probes detected with Texas Red streptavidin (Walton eta/., 1989).
mentalized to adenovirus replication factories (Walton eta/., 1989). Additionally, in HeLa cells MVM DNA rep- lication is increased by Ad2 co-infection, but in 293 cells MVM DNA levels are actually reduced by Ad2 co- infection. These data suggest that induction of MVM DNA replication and recompartmentalization of MVM DNA by adenovirus are separable effects. Most likely the adenovirus ElAIEl B genes (present in 293 cells and introduced during co-infection in both cell lines) mediate the increase in MVM DNA replication. How- ever, MVM DNA is replicated in nucleoli in 293 cells but is recompartmentalized by additional Ad2 co-infection suggesting that the adenovirus replication factory for- mation is induced by other Ad2 genes or by nucleation on the Ad2 DNA (Walton et a/., 1989).
MVM DNA replication and gene expression are uncoupled
In the lytic cycles of most DNAviruses viral DNA rep- lication is tightly coupled to viral gene expression. However, it appears that for MVM coupling of DNA rep- lication to the expression of the MVM genes is ex- tremely weak and may only affect the quantity of MVM DNA produced in an infection. Comparison of MVM gene expression and MVM DNA replication in Wl38, HeLa, and 293 cells indicated that there is no correla- tion between expression of MVM gene products (NS-1 or capsid proteins) and quantity of MVM DNA repli- cated. The production of NS-1 and capsid proteins was
evident in all cases studied, but MVM DNA replication varied from undetectable in WI38 cells to extremely efficient in 293 cells. These data suggest that MVM gene expression does not directly drive MVM DNA rep- lication.
Spalholz et al. (1983) have shown that in MVM infec- tion of nonpermissive mouse host cells transcription of MVM genes is severely reduced but MVM DNA repli- cation still can be supported at up to 20% of the levels seen in permissive host cells. Additionally, kinetic stud- ies suggest that the rates of all component initiation and processing steps in MVM DNA replication are lim- ited by the concentration of host factors (McHenry and Bodnar, unpublished data). While there is evidence that NS-1 modulates levels of DNA replication late in infec- tion (Rhode, 1976; Tullis et al., 1988) and that vDNA synthesis may be coupled to encapsidation (Tattersall and Ward, 1976) neither NS-1 or capsid gene expres- sion appears to be necessary or sufficient to regulate MVM DNA replication. It appears that at least the initial DNA synthesis of a second strand on the single- stranded vDNA template occurs in all the human cell lines tested since one would expect that a double- stranded DNA template is necessary for transcription of the MVM genes.
The signal for intranuclear localization of MVM DNA is mediated by host factors
MVM DNA is replicated in host nucleoli in HeLa and 293 cells as well as in normal mouse A9 host cells (Wal- ton et al., 1989) but is replicated in replication factories in Ad2-infected HeLa or 293 cells (Fig. 4). During ade- novirus infection several host nucleolar antigens are re- compartmentalized to replication factories suggesting that replication factories are “pseudonucleoli” con- taining nucleolar components which are utilized by ad- enovirus during its replication (Walton et al., 1989). Our data here are consistent with that hypothesis and fur- ther suggest that MVM DNA replication depends on cocompartmentalization with nucleolar components either in nucleoli or in replication factories.
TABLE 3
PRODUCTION OF INFECTIOUS MVM VIRIONS IN HUMAN CELLS
MVM-infected MVM + Ad2infected
WI38 8 X 1 O-4 ND HeLa 5x 10-l 2 x 10-l 293 7x 10-l ND 324K 2x lo* 7x 103 A9 2x lo4 2x 103 A9 (zero time control) 9 x 10-4 1 x 10-Z
Note. All values in PFU per infected cell. ND, not determined.
MVM-ADENOVIRUS INTERACTIONS 411
The nuclear matrix has been implicated in DNA repli- cation of both viral and cellular DNAs, and recent data indicate that MVM DNA is bound to the host nuclear matrix by the protein(s) associated with the MVM DNA termini (Bodnar et al., 1989). These data suggest that correct intranuclear localization of MVM DNA on the nuclear matrix for its replication is mediated by the pro- tein covalently bound to the MVM DNA termini.
It is unlikely that MVM gene products NS-1 or capsid proteins provide the signal for intranuclear localization of MVM DNA since both NS-1 and MVM capsid pro- teins are found in a diffuse pattern throughout MVM- and MVM-plus Ad2-infected cells. We suggest that any interactions of those proteins with MVM DNA are me- diated by diffusion of the proteins into the intranuclear compartments which contain MVM DNA. Therefore, the signal for intranuclear localization of MVM DNA is most likely provided by cellular factors.
Our data suggest that NuFl might provide that signal. NuFl colocalizes with MVM DNA in A9 cells (Walton et al., 1989) as well as in MVM- and MVM + Ad2-infected HeLa and 293 cells. NuFl is associated with MVM DNA termini (Chow eta/., 1986). While NuFl can be released from MVM-infected cell nuclei by extraction with non- ionic detergents (Chow et a/., 1986), it is associated with nucleolar chromatin in uninfected mouse cells (Walton and Bodnar, unpublished data). These data suggest that NuFl is a host protein which complexes with MVM DNA termini and MVM-encoded NS-1 at the termini (Cotmore and Tattersall, 1988) to localize MVM DNA to the proper intranuclear environment for replica- tion.
Our comparative study of MVM infections in adeno- virus-infected and adenovirus-transformed human cells has suggested mechanisms for MVM-host and MVM-helper virus interactions. Continued character- ization of parvovirus interactions with both transformed host cells and helper viruses should provide new in- sights into the mechanisms of both viral and host DNA replication.
ACKNOWLEDGMENTS
We thank Katherine McHenry for technical assistance and William Fowle and Dr. Daniel Scheirer for assistance with photography. We thank Drs. Susan Cotmore and Peter Tattersall (Yale University) for providing antisera. This work was supported by the National Insti- tutes of Health (GM35238).
REFERENCES
AIELLO, L., GUILFOYLE, R., HUEBNER, K., and WEINMANN, R. (1979). Ad- enovirus 5 DNA sequences present and RNA sequences tran- scribed in transformed human embryo kidney cells (HEK-Ad-5 or 293). Virology 94,460-469.
ASTELL, C. R., THOMSON, M., CHOW, M. B., and WARD, D. C. (1983). Structure and replication of minute virus of mice DNA. Co/d Spring Harbor Symp. &ant. Biol. 47,751-762.
BANTEL-SCHAAL, U., and ZUR HAUSEN, H. (1988). Adeno-associated viruses inhibit SV40 DNA amplification and replication of herpes simplex virus in SV40-transformed hamster cells. Virology 164, 64-74.
BERNS. K. I., and HAUSWIRTH, W. W. (1978). Parvovirus DNA structure and replication. ln “Replication of Mammalian Parvoviruses” (D. C. Ward and P. Tattersall, Eds.), pp. 13-32. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
BODNAR. J. W., HANSON, P. I., POLVINO-BODNAR, M., ZEMPSKY. W., and WARD, D. C. (1989). The terminal regions of adenovirus and minute virus of mice DNAs are preferentially associated with the nuclear matrix in infected cells. /. Viral. 63, 4344-4353.
BODNAR. 1. W., and PEARSON, G. D. (1980a). Kinetics of adenovirus DNA replication. I. Rate of adenovirus DNA replication. Virology 100,208-211.
BODNAR, J. W., and PEARSON, G. D. (1980b). Kinetics of adenovirus DNA replication. Il. Initiation of adenovirus DNA replication. Virol- ogy 105,357-370.
CHOW, M., BODNAR. J. W.. POLVINO-BODNAR. M., and WARD. D. C. (1986). Identification and characterization of a protein covalently bound to DNA of minute virus of mice. J. Viral. 57, 1094-l 104.
CORNELIS, J. J., BECQUART, P., DUPONCHEL, N., SALOM~, N., AVALOSSE, B. L., NAMBA, M., and ROMMELAERE, J. (1988a). Transformation of human fibroblasts by ionizing radiation, a chemical carcinogen, or simian virus 40 correlates with an increase in susceptibility to the autonomous parvoviruses H-l and minute virus of mice. J. Viral. 62, 1679-l 686.
CORNELIS, J. J., SPRUM, N., SPEGELAERE. P.. GUETTA, E., DARAWSHI, T., COTMORE, S. F., TAL, J.. and ROMMELAERE, J. (198813). Sensitization of transformed rat fibroblasts to killing by parvovirus minute virus of mice correlates with an increase in viral gene expression. 1. Viral. 62, 3438-3444.
COTMORE, S. F., and TA~ERSALL, P. (1987). The autonomously repli- cating parvoviruses of vertebrates. Adv. Virus Res. 33, 91-174.
COTMORE, S. F., and TA~ERSALL, P. (1988). The NS-1 polypeptide of minute virus of mice is covalently attached to the 5’termini of du- plex replicative form DNA and progeny single strands. J. Viral. 62, 851-860.
CUKOR, G.. BLACKLOW, N. R.. KIBRICK, S., and SWAN, I. C. (1975). Ad- eno-associated virus on cancer expression by herpevirus-trans- formed cells. 1. Nat/. Cancerhst. 55, 957-959.
DE LA MAZA, L. M., and CARTER, B. 1. (1981). Inhibition of adenovirus oncogenicity in hamsters by adeno-associated virus DNA. 1. Nat/. Cancerlnst. 67, 1323-1326.
DOERIG, C., HIRT, B., BEARD, P., and ANTONIEITI, J.-P. (1988). Minute virus of mice non-structural protein NS-1 is necessary and suffl- cient for rrans-activation of the viral P39 promoter. J. Gen. Viral. 69, 2563-2573.
GRAHAM, F. L., ABRAHAMS, P. S.. MULDER. C., HEIJNEKER, H. L., WAR- NAAR, S. O., DEVRIES, F. A., FIERS, W., and VAN DER EB, A. J. (1974). Studies on the in vitro transformation by DNA and by DNA frag- ments of human adenoviruses and simian virus 40. Cold Spring Harbor Symp. Quant. Biol. 39,637-650.
GUETTA, E., GRAZIANI, Y.. and TAL, J. (1986a). Suppression of Ehrlich ascites tumors In mice by minute virus of mice. J. Nat/. Cancer Inst. 76,1177-1180.
GUE~A, E., RON, D.. and TAL. J. (1986b). Developmental-dependent reptlcatlon of minute virus of mice in differentiated mouse testicu- lar cell lines. 1. Gen. Viral. 67, 2549-2554.
HEILBRONN, R., SCHLEHOFER. 1. R., and ZUR HAUSEN, H. (1984). SeleC- tive killing of carcinogen-treated SV40-transformed Chinese ham- ster cells by a defective parvovirus. Virology 136, 439-441.
412 FOX, MOEN. AND BODNAR
JANIK. J. E., HUSTON, M. M., and ROSE, J. A. (1981). LocatIons of ade- novlrus genes required for the replication of adeno-associated VI- rus. Proc. Nat/. Acad. Sci. USA 78, 1925-l 929.
LEDINKO. N., HOPKINS, S., and TOOLAN, H. (1969). Relatlonshtp be- tween potentiation of H-l growth by human adenovirus 12 and inhibition of the “helper” adenovlrus by H-l. /. Gen. viral. 5, 19- 31.
MERCHLINSKY, M. 1.. TATTERSALL, P. J., LEARY, J. J., COTMORE, S. F., GARDINER, E. M., and WARD, D. C. (1983). Construction of an infec- tious molecular clone of the autonomous parvovirus minute virus of mice. 1. Viral. 47, 227-232.
MOUSSET. S., CORNELIS, J.. SPRUYT. N., and ROMMECAERE, J. (1986). Transformation of established murine fibroblasts with an activated cellular Harvey-ras oncogene or the polyoma virus middle T gene increases cell permissiveness to parvovirus minute-virus-of-mice. Biochimie68, 951-955.
MOUSSET, S., and ROMMELAERE, J. (1982). Minute virus of mice inhib- its cell transformation by simian virus 40. Nature (London) 300, 537-539.
RHODE, S. L., Ill (1976). Replication process of the parvovtrus H-l. Isolation and characterization of temperature-sensitive H-l mu- tants defective in progeny DNA synthesis. /. Viral. 17, 659-667.
SHAHRABADI. M. S.. ROY, K. L., and YAMAMOTO, T. (1972). Macromo- lecular content of inclusions produced by a canine adenovirus. /. Viral. 10, 801-809.
SHIROKI. K., SHIMOJO, H.. and YAMAGUCHI, K. (1974). The viral DNA replication complex of adenovirus 12. Virology 60, 192-l 99.
SIMMONS, T.. HEYWOOD. P., and HODGE, L. D. (1974). lntranuclear site of replication of adenovirus DNA. /. MO/. Biol. 89, 423-433.
SPALHOU, B. A., BRAT~ON, J., WARD, D. C., and TATTERSALL, P. (1983). Intracellular restnction of parvovlrus replication in differentiated cells. In “Tumor Viruses and Differentiation,” pp. 315-328. A. R. LISS, New York.
TATTERSALL, P., and WARD, D. C. (1976). Rolling hairpln model for replication of parvovirus and linear chromosomal DNA. Nature (London) 263,106-l 09.
TOOLAN, H. W. (1967). Lack of oncogenic effect of the H-viruses for hamsters. Nature (London) 214, 1036.
TOOLAN, H. W., and LEDINKO, N. (1968). Inhibition by H-l virus of the incidence of tumors produced by adenovirus 12 in hamsters. /. Viral. 35, 475-478.
TOOLAN, H. W., RHODE, S. L., Ill, and GIERTHY, J. F. (1982). InhibitIon of 7,12-dlmethylbenz(a)anthraceneinduced tumors In Syrian ham- sters by prior infection with H-l parvovirus. CancerRes. 42,2552- 2555.
TULLIS, G. E., LABIENIEC-PINTEL, L.. CLEMENS, K. E., and PINTEL, D. (1988). Generation and characterization of a temperature-sensitive mutation in the NS-1 gene of the autonomous parvovlrus minute virus of mice. /. Viral. 62, 2736-2744.
WALTON, T. H., MOEN, P. T.. JR., Fox, E., and BODNAR, J. W. (1989). Interactions of Minute Virus of Mice and adenovirus with host nucleoli. J. Viral. 63, 3651-3660.
WOLTER. S., RICHARDS, R., and ARMENTROUT. R. W. (1980). Cell cycle- dependent replication of the DNA of minute virus of mice, a parvo- virus. Biochim. Biophys. Acta 607, 420-431.
YAKOBSON, B.. KOCH, T.. and WINOCOUR. E. (1987). Replication of ad- eno-associated virus in synchronized cells without the addition of a helper virus. J. Viral. 61, 972-981.