Original Article Autophagy in Normal and Abnormal Early Human Pregnancies Laura Avagliano, MD, PhD 1 , Laura Terraneo, PhD 1 , Eleonora Virgili, BD 1 , Carla Martinelli PhD 2 , Patrizia Doi, MS 1 , Michele Samaja, PhD 1 , Gaetano Pietro Bulfamante, MD 1 , and Anna Maria Marconi, MD 1 Abstract Autophagy is an inducible catabolic process by which cells degrade and recycle materials to survive stress, starvation, and hypoxia. The aim of this study was to evaluate autophagy at the fetal–maternal interface, to assess autophagy involvement during the early phase of human gestation, and to explore autophagic modification in case of early abnormal pregnancy outcome. Specimens were collected from first-trimester normal gestations undergoing legal termination of pregnancy and first-trimester sporadic sponta- neous miscarriages. Autophagy was studied in villous and decidual samples by transmission electron microscopy, immunohisto- chemistry, immunofluorescence, and Western blotting. Autophagy markers were found in cytotrophoblast, syncytiotrophoblast, extravillous trophoblast, and decidual stromal cells. Autophagy is physiologically involved in early normal gestation. Compared with normal pregnancy, spontaneous miscarriage presents an increase in autophagy expression in villous specimens due to an increment in concentration of autophagic vacuole in syncytiotrophoblast, suggesting a cytoprotective mechanism of the cells to respond to microenvironmental challenge. Keywords autophagy, apoptosis, pregnancy, trophoblast, miscarriage Introduction Autophagy is an evolutionary conserved, intracellular inducible catabolic process by which injured organelles, damaged proteins, and pathogens are sequestered into autophagosomes, double membrane-bound vesicles that fuse with lysosomes for proteoly- tic degradation and material recycling. 1 Microtubule-associated protein light chain 3 (LC3), the mammalian homologue of yeast autophagy-related gene 8, intervenes in the late stages of autop- hagosome formation. Among LC3 isoforms, LC3-II, the phos- phatidylethanolamine conjugated product of LC3-I obtained after LC3 activation, 1,2 is currently used as a specific marker of autophagy for its role during autophagosome genesis. 3 Previous studies have investigated the occurrence of autop- hagy during the early stages of mammalian pregnancy in the preimplantation period for oocyte-to-embryo transition 4-6 but no extensive in vivo assessment of autophagy at the fetomater- nal interface after implantation has been performed in humans. At the site of implantation, apoptosis permits placenta develop- ment, 7 and a relatively hypoxic environment warrants an ade- quate embryonic and placental growth. 8 Although many cellular and molecular events are expected to regulate the var- ious steps of fetomaternal interactions, the mechanisms that regulate the early stage of pregnancy are not yet fully understood. The aim of the present study was to evaluate the occurrence of autophagy during the first trimester of pregnancy to verify whether it is involved in the early phase of gestation and whether autophagy expression differs between normal pregnancy (NP) and spontaneous miscarriage (SM). Materials and Methods Sample Collection Normal pregnancies. Tissues were obtained from 10 healthy, nonsmokers women (age 28.5 + 8.1 years) undergoing legal termination of pregnancy within 12 weeks of amenorrhea (mean gestational age 9.4 + 1.1 weeks; range 8-11 weeks); 1 Department of Health Sciences, San Paolo Hospital Medical School, Universita ` degli Studi di Milano, Milan, Italy 2 Department of Biomedical Sciences for Health, Universita ` degli Studi di Milano, Milan, Italy Corresponding Author: Laura Avagliano, Unit of Obstetrics and Gynecology, Department of Health Sciences, San Paolo Hospital Medical School. University of Milano, Via A. di Rudinı ` 8. 20142 Milano, Italy. Email: [email protected]Reproductive Sciences 2015, Vol. 22(7) 838-844 ª The Author(s) 2014 Reprints and permission: sagepub.com/journalsPermissions.nav DOI: 10.1177/1933719114565036 rs.sagepub.com by guest on June 24, 2015 rsx.sagepub.com Downloaded from
Transcript
Original Article
Autophagy in Normal and Abnormal EarlyHuman Pregnancies
Laura Avagliano, MD, PhD1, Laura Terraneo, PhD1,Eleonora Virgili, BD1, Carla Martinelli PhD2, Patrizia Doi, MS1,Michele Samaja, PhD1, Gaetano Pietro Bulfamante, MD1,and Anna Maria Marconi, MD1
AbstractAutophagy is an inducible catabolic process by which cells degrade and recycle materials to survive stress, starvation, and hypoxia.The aim of this study was to evaluate autophagy at the fetal–maternal interface, to assess autophagy involvement during the earlyphase of human gestation, and to explore autophagic modification in case of early abnormal pregnancy outcome. Specimens werecollected from first-trimester normal gestations undergoing legal termination of pregnancy and first-trimester sporadic sponta-neous miscarriages. Autophagy was studied in villous and decidual samples by transmission electron microscopy, immunohisto-chemistry, immunofluorescence, and Western blotting. Autophagy markers were found in cytotrophoblast, syncytiotrophoblast,extravillous trophoblast, and decidual stromal cells. Autophagy is physiologically involved in early normal gestation. Comparedwith normal pregnancy, spontaneous miscarriage presents an increase in autophagy expression in villous specimens due to anincrement in concentration of autophagic vacuole in syncytiotrophoblast, suggesting a cytoprotective mechanism of the cellsto respond to microenvironmental challenge.
maternal blood. Since autophagy can intervene in placental
trophoblasts during stressor conditions as a cytoprotective
process,16 we suggest that the increase of autophagy in syncy-
tiotrophoblast could reflect a prosurvival tentative of the cells
to respond to the environmental changes.
Moreover, we detected autophagy markers in decidua speci-
mens, trophoblastic-anchoring columns, and EVT. It is known
that EVT migrates from the columns through the decidual
stroma, reaches the spiral arteries, penetrates the wall of the
vessels, and colonizes the lumen forming the trophoblastic
plugs and conducting to vascular remodeling.17 Deficiency in
EVT invasion and inadequate vascular remodeling can lead
to adverse pregnancy outcome such as miscarriage, preeclamp-
sia, and intrauterine growth restriction.18 Previous studies per-
formed on trophoblastic cell lines suggested that autophagy is
involved in trophoblast invasion,14 even if the results are con-
flicting: Some authors observed that autophagy induced by
hypoxia supports trophoblast invasion,15,19 whereas other
authors observed an increase in autophagy but a decrease in tro-
phoblast invasion in relation to the inhibition of HIF1-a expres-
sion.20 Differences in autophagy expression between decidua
from NP and SM were not detected in the present study, while
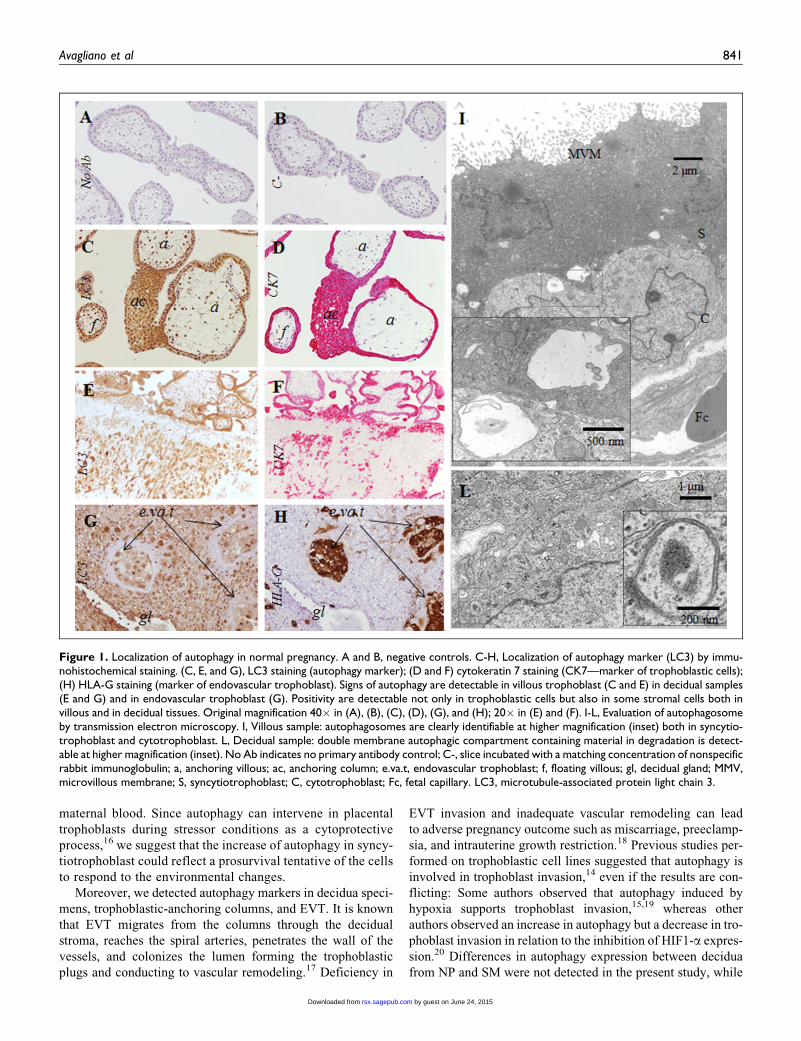
Figure 1. Localization of autophagy in normal pregnancy. A and B, negative controls. C-H, Localization of autophagy marker (LC3) by immu-nohistochemical staining. (C, E, and G), LC3 staining (autophagy marker); (D and F) cytokeratin 7 staining (CK7—marker of trophoblastic cells);(H) HLA-G staining (marker of endovascular trophoblast). Signs of autophagy are detectable in villous trophoblast (C and E) in decidual samples(E and G) and in endovascular trophoblast (G). Positivity are detectable not only in trophoblastic cells but also in some stromal cells both invillous and in decidual tissues. Original magnification 40� in (A), (B), (C), (D), (G), and (H); 20� in (E) and (F). I-L, Evaluation of autophagosomeby transmission electron microscopy. I, Villous sample: autophagosomes are clearly identifiable at higher magnification (inset) both in syncytio-trophoblast and cytotrophoblast. L, Decidual sample: double membrane autophagic compartment containing material in degradation is detect-able at higher magnification (inset). No Ab indicates no primary antibody control; C-, slice incubated with a matching concentration of nonspecificrabbit immunoglobulin; a, anchoring villous; ac, anchoring column; e.va.t, endovascular trophoblast; f, floating villous; gl, decidual gland; MMV,microvillous membrane; S, syncytiotrophoblast; C, cytotrophoblast; Fc, fetal capillary. LC3, microtubule-associated protein light chain 3.
Avagliano et al 841
by guest on June 24, 2015rsx.sagepub.comDownloaded from
apoptosis and hypoxia were increased in SM. Our finding of
increased apoptosis in first-trimester decidua of SM is in agree-
ment with a previous report where apoptosis was responsible
for the pregnancy loss.21 In our samples of SM, we found an
increased expression of autophagy in villous specimens but a
low expression of apoptosis; on the contrary, in decidua speci-
mens we found low levels of autophagy but an increased sus-
ceptibility to apoptosis. It is possible that the opposite trend
of expression of autophagy compared to hypoxia and apoptosis
markers in villi and decidua of SM might reflect the role of
autophagy in antagonizing hypoxia-induced apoptosis, but fur-
ther investigation is needed to clarify this issue.
This study has some limitations. First, samples obtained
from legal interruption of pregnancies include viable tissues,
whereas samples from SMs include tissues from nonviable
pregnancies; it is therefore not possible to determine whether
autophagy has been induced in vivo before or after fetal
demise. Second, samples were stored in liquid nitrogen before
the analysis, whereas a previous study suggested a role of cold
stress in the induction of autophagy,22 therefore our results
might be influenced by the freezing of samples before the anal-
ysis. Third, we could not exclude that the intersample variabil-
ity in villi affected our results: Autophagy is a dynamic and
continuous process and LC3-II is just a marker of one stage
of it. Different stages of the evolution during the maturation
from the phagophore through the autolysosome in our samples
could affect the presence or the level of LC3-II.
In conclusion, our study demonstrates the occurrence of
autophagy in vivo at the fetomaternal interface during the first
trimester of normal human gestation. A low level of autophagy
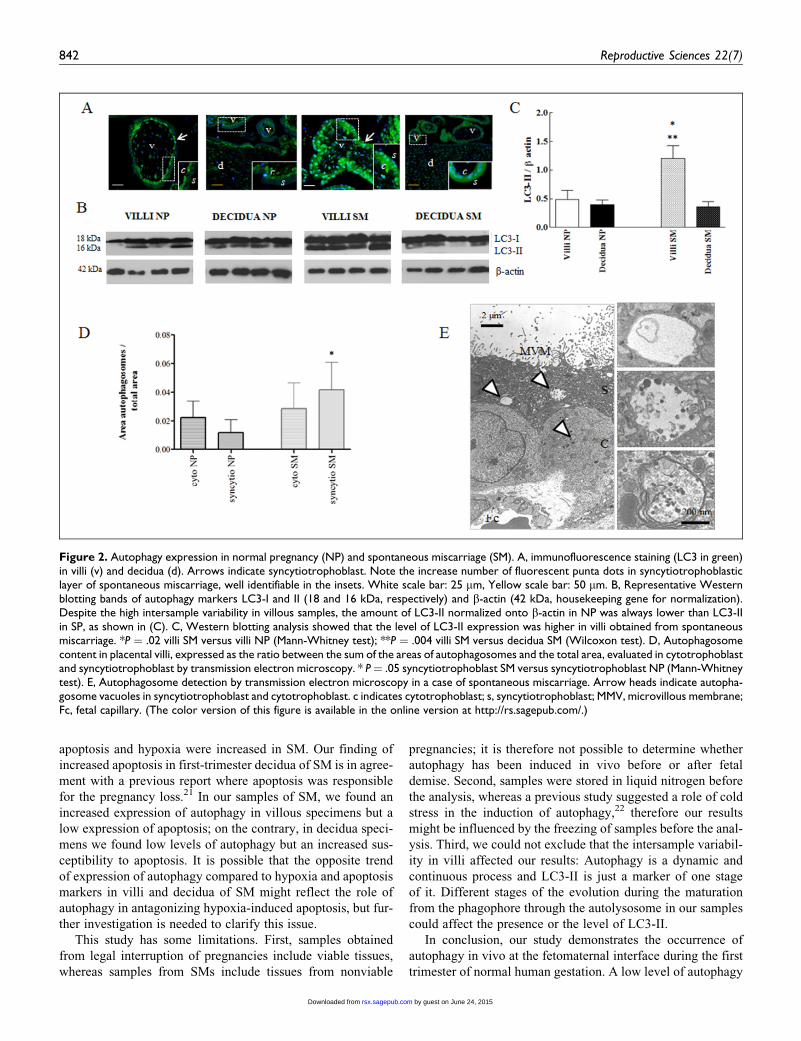
Figure 2. Autophagy expression in normal pregnancy (NP) and spontaneous miscarriage (SM). A, immunofluorescence staining (LC3 in green)in villi (v) and decidua (d). Arrows indicate syncytiotrophoblast. Note the increase number of fluorescent punta dots in syncytiotrophoblasticlayer of spontaneous miscarriage, well identifiable in the insets. White scale bar: 25 mm, Yellow scale bar: 50 mm. B, Representative Westernblotting bands of autophagy markers LC3-I and II (18 and 16 kDa, respectively) and b-actin (42 kDa, housekeeping gene for normalization).Despite the high intersample variability in villous samples, the amount of LC3-II normalized onto b-actin in NP was always lower than LC3-IIin SP, as shown in (C). C, Western blotting analysis showed that the level of LC3-II expression was higher in villi obtained from spontaneousmiscarriage. *P ¼ .02 villi SM versus villi NP (Mann-Whitney test); **P ¼ .004 villi SM versus decidua SM (Wilcoxon test). D, Autophagosomecontent in placental villi, expressed as the ratio between the sum of the areas of autophagosomes and the total area, evaluated in cytotrophoblastand syncytiotrophoblast by transmission electron microscopy. * P¼ .05 syncytiotrophoblast SM versus syncytiotrophoblast NP (Mann-Whitneytest). E, Autophagosome detection by transmission electron microscopy in a case of spontaneous miscarriage. Arrow heads indicate autopha-gosome vacuoles in syncytiotrophoblast and cytotrophoblast. c indicates cytotrophoblast; s, syncytiotrophoblast; MMV, microvillous membrane;Fc, fetal capillary. (The color version of this figure is available in the online version at http://rs.sagepub.com/.)
842 Reproductive Sciences 22(7)
by guest on June 24, 2015rsx.sagepub.comDownloaded from
2. Noda T, Fujita N, Yoshimori T. The late stages of autophagy: how
does the end begin? Cell Death Differ. 2009;16(7):984-990.
3. He C, Klionsky DJ. Regulation mechanisms and signaling path-
ways of autophagy. Annu Rev Genet. 2009;43:67-93.
4. Mizushima N, Levine B. Autophagy in mammalian development
and differentiation. Nat Cell Biol. 2010;12:823-830.
5. Tsukamoto S, Kuma A, Murakami M, Kishi C, Yamamoto A,
Mizushima N. Autophagy is essential for preimplantation deve-
lopment of mouse embryos. Science. 2008;321:117-120.
Figure 3. Evaluation of apoptosis and hypoxia by Western blotting analysis. A, Western blotting analysis showed that HIF-1a expression(hypoxia marker) was high in decidual samples obtained from spontaneous miscarriage (SM). *P ¼ .0002 decidua SM versus decidua normalpregnancy (NP; Mann-Whitney test); on the right, representative bands of HIF-1a (120 kDa) and b-actin (42 kDa, housekeeping gene for normal-ization) are shown. B, Western blotting analysis showed an increased trend in the Bax/Bcl2 ratio in decidua obtained from spontaneous mis-carriage. C, Western blotting analysis of cleaved caspase-3. The extent of the cleavage is comparable to the Bax/Bcl2 ratio, supportinggreater apoptosis in decidua SM compared to villi SM and all samples from NP. D, Immunohistochemical expression of cleaved caspase 3. Pos-itive cells are in brown. Note the increased number of positive cells in decidua from spontaneous miscarriage respect to decidua from normalpregnancy. HIF-1a indicates hypoxia-inducible factor. (The color version of this figure is available in the online version at http://rs.sagepub.com/.)
Avagliano et al 843
by guest on June 24, 2015rsx.sagepub.comDownloaded from