Pao-Hsien Chu3 Yu-Ying Kao4 Ching-Han Hsu2 and Kurt Ming-Chao Lin1
1 Institute of Biomedical Engineering and Nanomedicine National Health Research Institutes Zhunan Miaoli County Taiwan2Department of Biomedical Engineering and Environmental Sciences National Tsing Hua University Hsinchu Taiwan3Division of Cardiology Department of Internal Medicine Chang Gung Memorial Hospital College of MedicineChang Gung University Taipei Taiwan4Department of Biotechnology Chia Nan University of Pharmacy and Science Tainan Taiwan
Correspondence should be addressed to Kurt Ming-Chao Lin klinnhriorgtw
Received 27 June 2015 Accepted 26 August 2015
Academic Editor Bodh I Jugdutt
Copyright copy 2015 Tsung-Hsien Chen et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Mitochondrial heat shock proteins such as HSP60 are chaperones responsible for the folding transport and quality control ofmitochondrial matrix proteins and are essential for maintaining life Both prosurvival and proapoptotic roles have been proposedfor HSP60 and HSP60 is reportedly involved in the initiation of autoimmune metabolic and cardiovascular diseases The role ofHSP60 in pathogenesis of these diseases remains unclear partly because of the lack ofmousemodels expressingHSP60 In this studywe generated HSP60 conditional transgenic mice suitable for investigating in vivo outcomes by expressing HSP60 at the targetedorgan in disease models Ubiquitous HSP60 induction in the embryonic stage caused neonatal death in mice at postnatal day 1 Ahigh incidence of atrial septal defects was observed in HSP60-expressingmice with increased apoptosis andmyocyte degenerationthat possibly contributed to massive hemorrhage and sponge-like cardiac muscles Our results showed that neonatal heart failurethrough HSP60 induction likely involves developmental defects and excessive apoptosis The conditional HSP60 mouse model isuseful for studying crucial biological questions concerning HSP60
1 Introduction
Heat shock proteins (HSPs) are molecular chaperonesresponsible for critical functions such as protein foldingtransport and signaling Under stress HSPs preserve pro-teins in their native forms and refold denatured proteins [1]Mitochondrial chaperonins such as HSP60 together withHSP10 refold all proteins imported into the mitochondrialmatrix and facilitate the restoration of unfolded or misfoldedmatrix proteins under normal and stressed environmentshence they are vital for maintaining mitochondrial func-tions including ATP synthesis [2ndash4] Although the fun-damental biology of HSP60 particularly on mechanismsof substrate folding has been explored extensively HSP60-related studies are gaining increasing attention because newfindings about the roles of mitochondrial cytosolic and
extracellular HSP60 in a wide array of diseases have emerged[5] Most HSPs are prosurvival although both prosurvivaland prodeath roles of HSP60 have been reported Theprotective role of HSP60 in various cell types includingcardiac myocytes has been reported [6ndash13] Preservation ofmitochondrial capacity by HSP60 with reduced cytochromeC release during stress challenges and subsequent apoptosisreductionwere among the commonfindings of the protectionby HSP60 Upregulated HSP60 expression in several studiessuggested the prosurvival and antiapoptotic roles ofHSP60 incancers [14] such as cervical [15] prostate [16] and colorectalcancers [17] and Hodgkin lymphoma [18] The proapoptoticrole of HSP60 was documented in muscles [19] and invarious cancer cell lines [20 21] for example malignancy ofesophageal squamous cell carcinoma [22] and ovarian [23]and bladder cancers [24] was found inversely correlated with
Hindawi Publishing CorporationBioMed Research InternationalVolume 2015 Article ID 539805 11 pageshttpdxdoiorg1011552015539805
2 BioMed Research International
HSP60 expression The findings of HSP60 accumulation incytoplasm in various apoptosis models and HSP60 bindingto procaspase-3 supported the proapoptotic properties ofHSP60 [21 25] Besides the discrepancy among reports of invitro studies the roles of HSP60 in physiology and pathologycannot be fully elucidated without using transgenic (Tg)mouse models expressing HSP60 in vivo
The role of extracellular HSP60 in viral infection innateor adaptive immunity and atherosclerosis has been widelydocumented [26ndash28] Hepatitis B virus replication inducedHSP60 production and HSP60 has a critical role in regu-latory T cells functions particularly in IL-10 secretion [28]Innate immune cell surface receptors namely TLR2 TLR4and CD14 are known receptors for extracellular HSP60resulting in the release of mediators such as TNF-120572 IL-1120573 IL-6 and NO which in turn enhance inflammation intype 1 diabetes mellitus (DM) atherosclerosis arthritis andtransplant rejection
HSP60 expression in the heart is reduced in ageing andmetabolic diseases [29] Caloric restriction that is shownincreases lifespan restored aging-related decline of HSP60expression in the heart and improved cardiovascular func-tions [29] The reduction in HSP60 level contributing to lowinsulin sensitivity and to dysfunctions inmetabolic syndromeand type 2 DMwas proposed [30] Because of the importanceof HSP60 in many diseases Tg mouse models with inducibleand tissue-specific HSP60 expression can provide usefulinsights and expand the current understanding of thesediseases
Previously nonobese diabetic (NOD) HIIE120572-HSP60 Tgmice in which HSP60 is driven by the MHC class II-E120572(HIIE120572) promoter achieved HSP60 expression specifically inthe thymus and bone marrow and suppressed susceptibilityto autoimmunity induced DM [31] Tg mice expressingtruncated HSP60 instead of the entire HSP60 were alsoreported and the resultant cytosolic HSP60 Tg mice wereresistant to hepatic stress with increased cell survival [32]However Tg mice with HSP60 expression in other tissueshave not been reported Attempts to generate a conventionalHSP60mouse model did not succeed because Tg founders aschimeras failed to survive (personal communications) In thisreport we present a viable and healthy inducible HSP60 Tgmouse (G-Lox-HSP60) in FVB strain driven by a ubiquitousCMV early enhancerchicken 120573-actin promoter (CAGGS)This model overcame the early lethality of the conventionalTg approach After crossing the G-Lox-HSP60 mouse withthe Cre mouse we observed neonatal death in mice withHSP60 expression from the embryo stage
2 Materials and Methods
21 Generation of G-Lox-Hsp60 and EGFP-Cre TransgenicMice A G-Lox-Hsp60 mouse FVBN-Tg(ACTB-EGFPHSPD1)14Klin was generated by using the Tg vector(Figure 1(a)) and constructed by inserting the followingfragments CAGGS promoter 2 LoxP sites at the start andthe end separated by an EGFP coding sequence and an SV40polyA sequence (pEGFP-C2 Clontech Mountain ViewCA) and a human HSP60 cDNA (NM 002156 102ndash1920 bp)
inserted into the pDsRed-N1 vector (Clontech) in whichthe CMV enhancer was removed An ApaLI-AflII fragmentwas used for microinjection into the FVB blastocystsEGFP-Cre mouse FVBN-Tg(ACTB-EGFPCRE)21Klinwas constructed by inserting CAGGS and Cre (AF3348271767ndash2789 bp) into the pEGFP-C2 vector in which the CMVenhancer was removed [33] For unknown reasons the greenfluorescence of the EGFP-Cre mouse was extremely weakG-Lox-DsRed mice have been described previously [34]All animal experiments were conducted in accordance withthe accepted standards of animal care and were approvedby the Institutional Animal Care and Use Committee of theNational Health Research Institutes Taiwan The followingPCR primers pairs were used for genotyping for the resultsin Figure 1(b)(A) (I) 51015840-AATGCTCACCGTAAGCCTTT-31015840 51015840-CCATCTGAAAGTTTTGCAAG-31015840 119879
119898= 655∘C
407 bp (II) 51015840-CTGCTAACCATGTTCATGCC-31015840 51015840-ACCGTCAGTACGTGAGATATCTT-31015840 119879
119898= 58∘C
1443 bp and (III) 51015840-CTGCTAACCATGTTCATGCC-3101584051015840-CCATCTGAAAGTTTTGCAAG-31015840 119879
119898= 60∘C 1442 bp
22 Analysis of mRNA Semiquantitative RT-PCR and ProteinExpression Total RNA was isolated from heart homogenateswith a TRIzol reagent (Invitrogen Carlsbad CA) GenomicDNA contamination in the RNA samples was removedthrough DNase I digestion and cDNA of the first strandwas synthesized using ReverTra Ace reverse transcriptase(Toyobo Japan) and oligo(dT) as the primer The followingPCR primers were used for amplifying the target genes for theresults in Figure 1(b)(B) Human HSP60 the same as listedin (I) human and mouse HSP60 51015840-GTCAGAAATGTG-AATTCCAG-31015840 51015840-TTGACTGCCACAACCTGAAGAC-31015840 119879119898= 55∘C 200 bp and GAPDH 51015840-GTGGCAAAGTGG-
AGATTGCC-31015840 51015840-GATGATGACCCGTTTGGCTCC-31015840119879119898= 58∘C 290 bp Ten micrograms of total proteins isolated
from tissue homogenates was subjected to 10 SDS-PAGEfollowed by Western blotting using anti-HSP60 (ab5479Abcam) and anti-GFP (ab290) antibodies Ten microgramsof total proteins isolated fromMDA-MB231 cells was used forcomparison
23 Fluorescence Detection and Histological Analysis Heartsof neonatal mice and E17 mouse embryos were washed withPBS and fixed by 10 neutral formalin Stereo microscope(Olympus SZX10) with fluorescence module was used forobserving green fluorescence of neonatal mouse hearts Forinvestigating septum defects unfixed neonatal hearts wereimmersed in OCT matrix (Tissue-Tek USA) and frozenquickly in liquid nitrogen The samples were cryosectioned10 120583m thick followed by hematoxylin and eosin (HampE)staining to reveal the four-chamber view of hearts In parallelformalin-fixed hearts were embedded in paraffin and cutinto serial 5 120583m sections and stained with HampE to evaluatemorphology
24 Immunohistochemical and Immunofluorescence AssayParaffin-embedded tissue sectionswere dewaxed rehydratedand treated by 3 hydrogen peroxide inmethanol for 30min
BioMed Research International 3
Cre
CAGGS promoter EGFP HSP60
LoxP LoxP
SV40pA SV40pA
Bgl II Hind III Hind III
DsRed
SCG-Lox-HSP60
(I)
CAGGS promoter HSP60
LoxP
SV40pA
DsRed
SC
(III)
Ase I
Ase I
CAGGS promoter EGFP Cre SV40pAEGFP-Cre
(II)
SC
(a)
(A)
(B) (C)
CAGGS-hHSP60
hHSP60
G-Lox-HSP60EGFP-Cre
(I)
(II)
(III)
EGFP-Cre
HSP60 (m h)
hHSP60
GAPDH
G-Lox-HSP60
EGFP-Cre
HSP60
EGFP
G-Lox-HSP60
EGFP-Cre
+
+ +
++
+ +
+minus
minus+ + + +
+++ minus minus
minus
+
minus minus
minus
minus
minus
minus
minusminus
minus
minus
minus
MD
A231
MD
A231
(b)
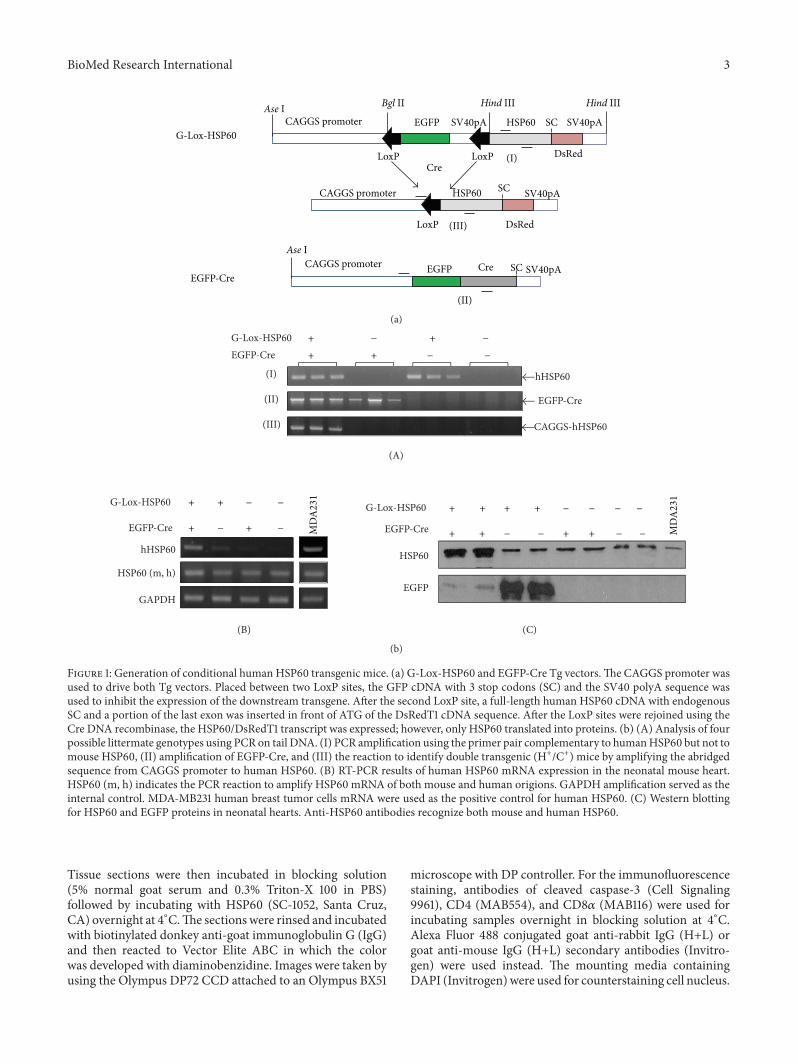
Figure 1 Generation of conditional human HSP60 transgenic mice (a) G-Lox-HSP60 and EGFP-Cre Tg vectorsThe CAGGS promoter wasused to drive both Tg vectors Placed between two LoxP sites the GFP cDNA with 3 stop codons (SC) and the SV40 polyA sequence wasused to inhibit the expression of the downstream transgene After the second LoxP site a full-length human HSP60 cDNA with endogenousSC and a portion of the last exon was inserted in front of ATG of the DsRedT1 cDNA sequence After the LoxP sites were rejoined using theCre DNA recombinase the HSP60DsRedT1 transcript was expressed however only HSP60 translated into proteins (b) (A) Analysis of fourpossible littermate genotypes using PCR on tail DNA (I) PCR amplification using the primer pair complementary to humanHSP60 but not tomouse HSP60 (II) amplification of EGFP-Cre and (III) the reaction to identify double transgenic (H+C+) mice by amplifying the abridgedsequence from CAGGS promoter to human HSP60 (B) RT-PCR results of human HSP60 mRNA expression in the neonatal mouse heartHSP60 (m h) indicates the PCR reaction to amplify HSP60 mRNA of both mouse and human origions GAPDH amplification served as theinternal control MDA-MB231 human breast tumor cells mRNA were used as the positive control for human HSP60 (C) Western blottingfor HSP60 and EGFP proteins in neonatal hearts Anti-HSP60 antibodies recognize both mouse and human HSP60
Tissue sections were then incubated in blocking solution(5 normal goat serum and 03 Triton-X 100 in PBS)followed by incubating with HSP60 (SC-1052 Santa CruzCA) overnight at 4∘CThe sections were rinsed and incubatedwith biotinylated donkey anti-goat immunoglobulin G (IgG)and then reacted to Vector Elite ABC in which the colorwas developed with diaminobenzidine Images were taken byusing the Olympus DP72 CCD attached to an Olympus BX51
microscope with DP controller For the immunofluorescencestaining antibodies of cleaved caspase-3 (Cell Signaling9961) CD4 (MAB554) and CD8120572 (MAB116) were used forincubating samples overnight in blocking solution at 4∘CAlexa Fluor 488 conjugated goat anti-rabbit IgG (H+L) orgoat anti-mouse IgG (H+L) secondary antibodies (Invitro-gen) were used instead The mounting media containingDAPI (Invitrogen)were used for counterstaining cell nucleus
4 BioMed Research International
25 TUNEL Assay Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay wasperformed for detecting nuclear DNA fragmentation inparaffin-embedded tissue sections as a measure of apoptosisFluorescein-12-dUTP was used as the substrate in TUNELreaction (DeadEnd Fluorometric TUNEL System Promega)the slides were counterstained by DAPI and the images weretaken by the Olympus microscope system
26 T2-Weighted Thoracic Region MR Images Magnetic res-onance imaging (MRI) was performed and images wereacquired using 94 Tesla MRI system (Bruker EttlingenGermany) with a maximum gradient strength of 140G cmminus1A quadrature volume coil (internal diameter = 10mm) wasused for RF transmission and reception We used a rapidacquisition with relaxation enhancement sequence to obtainthe T2-weighted images The repetition and echo times wereset at 2500 and 34 milliseconds respectively The field ofview was 10 times 10mm2 and the matrix size was 100 times 100 Theimages were sliced without gaps and the slice thickness was02mmThe voxel size for acquisitionwas 01times 01times 02mm3Finally the images were interpolated to 400times 400 using cubicinterpolation
27 Electron Micrograph Neonatal heart tissues were col-lected and fixed in 4 cold glutaraldehyde postfixed using1 osmium oxide and progressively dehydrated using alco-hol After resin embedding and polymerization 1 120583m thicksections were cut for initial observation Ultrathin (60ndash80 nmthick) sections were cut from the Epon-embedded blocksstained with uranyl acetate and lead citrate and examinedusing transmission electron microscopy (TEM Hitachi H-7500 Tokyo Japan) at 75 kV
3 Results and Discussion
31 Generation of HSP60 Conditional Transgenic MiceUnlike Tg mice expressing HSP10 [35] HSP20 [36] HSP27[37] or HSP70 [38] founders for establishing the ubiquitousHSP60 Tg mouse were not viable In this study we generateda conditional Tg mouse model using Cre-LoxP tools toallow inducible and tissue-specificHSP60 expression G-Lox-HSP60 mouse was generated by using the Tg vector illus-trated in Figure 1(a) CAGGSwas used to drive the expressionof transgenes enhanced green fluorescent protein (EGFP)or human HSP60 full-length cDNA Multiple stop codonsand the SV40 polyA sequences between the 2 LoxP sites ofthe Tg vector prevent the expression of downstream humanHSP60 sequence The G-Lox-HSP60 mice were normal indevelopment weight and reproduction with EGFP expres-sion in most organs During Cre-mediated recombination2 LoxP sites of the G-Lox-HSP60 vector are joined and theHSP60 expression is directly enhanced by CAGGS In thepresent study we generated an FVB ubiquitous Cre Tgmouseexpressing the EGFP-Cre fusion protein [33] We used thetail DNA-PCR method for studying the genotypes of the lit-termates in the crossing studies of G-Lox-HSP60 and EGFP-Cre mice the results are described in Figure 1(b) Deletion
Table 1 Summary of crossings between G-Lox-HSP60 (H) andEGFP-Cre (C) mice
Genotype H+C+ H+Cminus HminusC+ HminusCminus TotalNewborn number 72 76 69 86 303Neonatal death number 72 5 2 2 81Ratio 7272 576 269 286 81303H+C+ double Tg mouse H+Cminus HminusC+ single Tg mouse carrying eitherallele HminusCminus wild-type mouse
of the LoxP-flanked cassette was detected by the presence ofthe shorter PCR fragment in the double Tg mice Littermateswith all 4 possible genotypes were acquired in accordancewith the Mendelian frequency indicating no prenatal loss ofthe double Tgmice (H+C+) which were positive for both G-Lox-HSP60 and EGFP-Cre alleles (Table 1) Strong inductionof human HSP60 expression in the cardiac tissues of thedouble Tg mice was confirmed through RT-PCR Westernblotting (Figure 1(b)) and IHC staining (Figure 2)
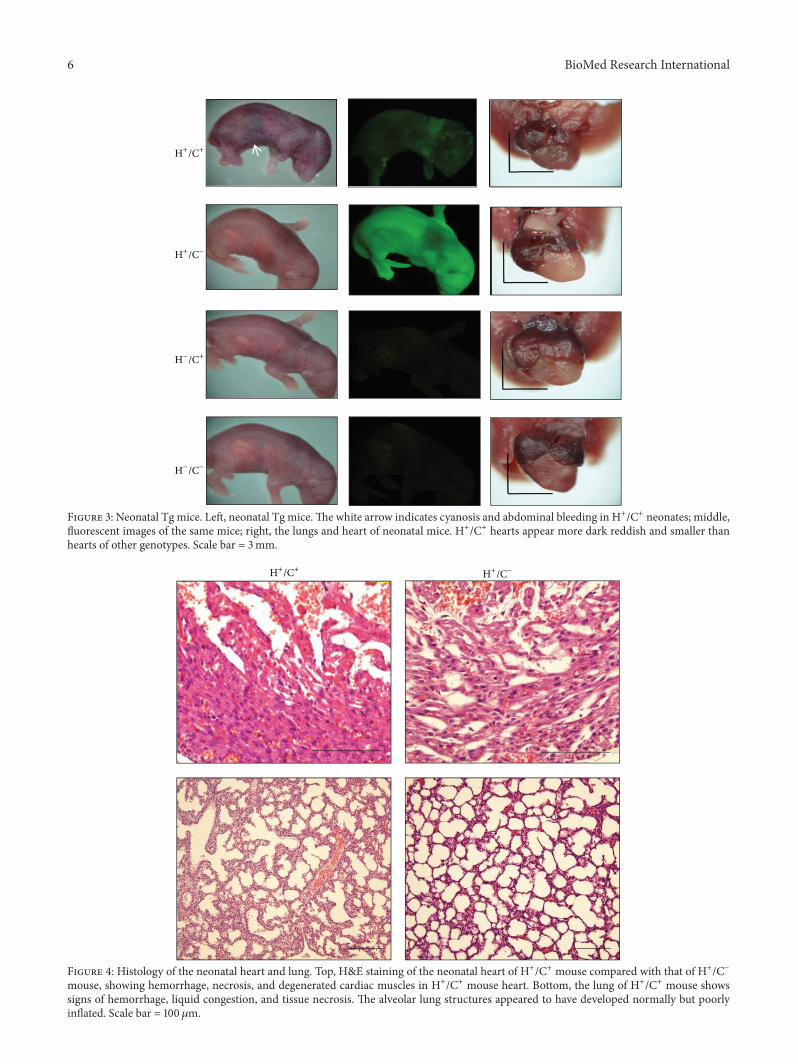
32 HSP60 Expression Led to Neonatal Deaths In more than20 crossing experiments using G-Lox-HSP60 and EGFP-Cremice all double TgHSP60mice litters diedwithin a fewhoursafter birth A few survived the first day but none survivedmore than a few days (7272 Table 1) Neonatal deaths wererare in single Tg micemdashH+Cminus (576) and HminusC+ (269)mdashwhich carried only G-Lox-HSP60 or EGFP-Cre and in thewild-type mice (HminusCminus 286) At birth neonatal HSP60mice were slightly smaller in size than average with signsof mild cyanosis and internal bleeding at the abdominalsegments before death (Figure 3) Severe cardiovascular orpulmonary abnormality was suspected to contribute to theneonatal deaths in the HSP60 mice H+Cminus mice exhibitedstrong whole-body green fluorescence whereas the HSP60mice displayed green fluorescent spots indicating remainingEGFP that was produced before recombination HminusC+ andHminusCminus mice did not exhibit fluorescence Thoracic chamberdissection of HSP60 mice revealed a smaller heart and thecolor of the ventricles was more reddish containing moreblood than the hearts of other genotypes (Figure 3)
Postmortem analysis of the HSP60 mice was performedthrough MRI and histology MRI revealed that the HSP60P1 mouse heart was smaller than that of H+Cminus mouse(Figure S1 in Supplementary Material available online athttpdxdoiorg1011552015539805) The HSP60 mouseheart chambers and lungs were filled with liquids that wereabsent in those of H+CminusmouseThe interventricular septumand chamber walls of the HSP60 mice were thinner thannormal (Figure S1) Histological examination revealed mas-sive hemorrhage at the heart and lungs of double Tg mouseThe cardiac muscle appeared degenerated or incompletelydifferentiated with sponge-like cardiac tissues fenestratedwith blood or blood vessels and signs of myocardiumnecrosis were observed (Figure 4) The lungs of the HSP60mice appeared to have developed normally except for thepresence of hemorrhage that potentially resulted in onlypartial inflation of the lungs Other organs such as thebrain kidneys and liver of the HSP60 mice appeared grossly
BioMed Research International 5
HampE HSP60
H+Cminus
HminusCminus
H+C+
Figure 2 HSP60 detection in neonatal H+C+ mice heart HampE and immunohistochemical staining of HSP60 induction in H+C+ heartsections but not in H+Cminus or HminusCminus heart sections Scale bar = 50 120583m
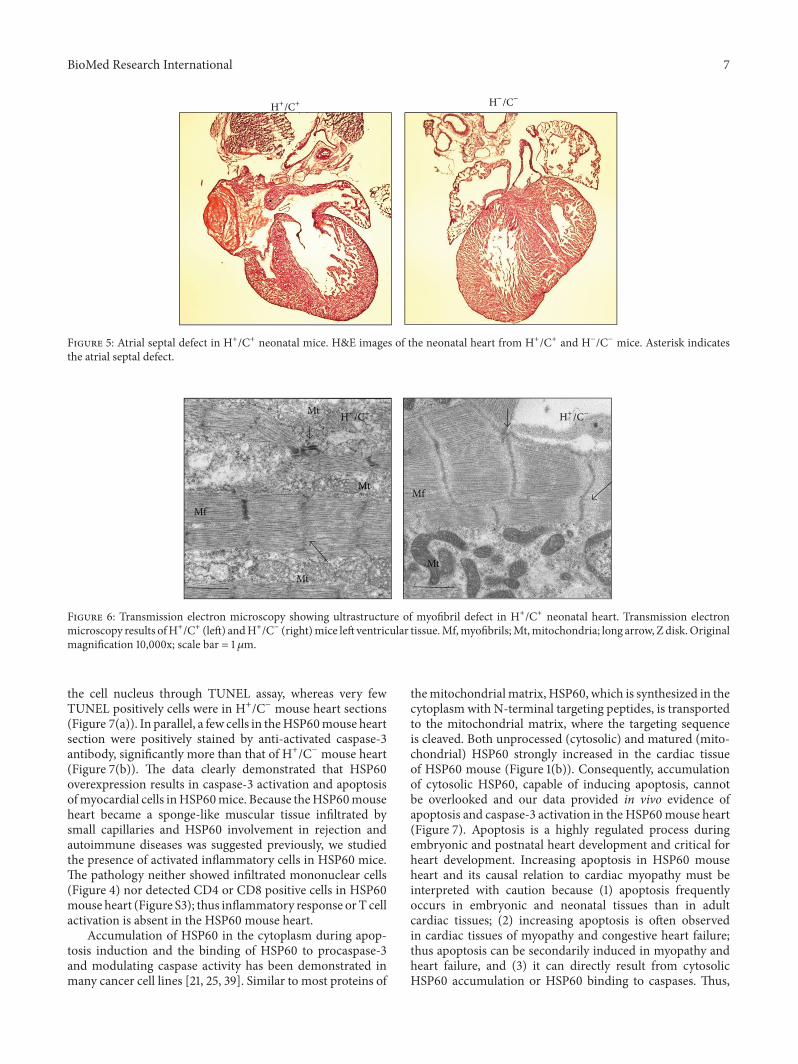
normal (data not shown) Spotty hemorrhage was observedin the skeletal muscles of the neck spine and between theribs however the hemorrhage in these organs was not assevere as that in the heart and lungs Because cardiovascularand respiratory stresses were observed and because the heartswere smaller than normal we speculate that congenitaldefects may contribute to the neonatal deaths of the HSP60mice Of the 10 HSP60 mice studied for septal defects 4had atrial septal defect (ASD) and one had ventricular septaldefect (VSD) as illustrated in Figure 5 Septum defects werenot observed in the control mice
Because the atrial septum is closed when the lungsare inflated during birthing and because ASD manifests asa serious problem only after birth we examined whetherthe HSP60 mice had developmental defects during theembryonic stage in the E17 embryos No apparent abnor-malities were visible externally in the HSP60 mouse E17embryos However the embryo sections revealed excessivehemorrhage vascularization and tissue necrosis in the heartsimilar to those in the postnatal heart Lungs of E17 ofHSP60 mouse appeared grossly normal (Figure S2) Thushistological evidence suggested that congenital heart diseasecaused by cardiomyopathy or incomplete heart development
in HSP60 mouse embryos results in ASD or VSD whichaggravates respiratory stress and congestive heart failure atpostnatal day 1
We searched for ultrastructural abnormality as featuresof cardiac myopathy in neonatal HSP60 mouse using elec-tron micrographs G-Lox-HSP60 mouse contained orga-nized arrays of myofibril filaments with intact mitochon-drion appearing in high contrast with dense laminaecristae throughout the mitochondrial cross-sectional areas(Figure 6) In contrast the myofibrils of HSP60 mouse heartexhibited pronounced fragmentation and disorganizationshown as ragged Z lines with shorter sarcomere lengthon average compared with G-Lox-HSP60 myofibrils Mito-chondrion between myofibrils was mostly broken losingboth outline and inner cristae indicating a disintegratedmitochondrial structureThus TEM results confirmed severecardiac myopathy in HSP60 mouse
33 Increasing Apoptosis in HSP60 Mouse Heart HSP60 waspreviously linked to apoptosis induction thus we studiedapoptotic cardiac myocytes in the HSP60 mouse by usingthe TUNEL assay Many cells in the HSP60 mouse heartsections were positively labeled by fluorescein-dUTP at
6 BioMed Research International
H+Cminus
HminusC+
HminusCminus
H+C+
Figure 3 Neonatal Tg mice Left neonatal Tg miceThe white arrow indicates cyanosis and abdominal bleeding in H+C+ neonates middlefluorescent images of the same mice right the lungs and heart of neonatal mice H+C+ hearts appear more dark reddish and smaller thanhearts of other genotypes Scale bar = 3mm
H+CminusH+C+
Figure 4 Histology of the neonatal heart and lung Top HampE staining of the neonatal heart of H+C+ mouse compared with that of H+Cminusmouse showing hemorrhage necrosis and degenerated cardiac muscles in H+C+ mouse heart Bottom the lung of H+C+ mouse showssigns of hemorrhage liquid congestion and tissue necrosis The alveolar lung structures appeared to have developed normally but poorlyinflated Scale bar = 100 120583m
BioMed Research International 7
H+C+ HminusCminus
Figure 5 Atrial septal defect in H+C+ neonatal mice HampE images of the neonatal heart from H+C+ and HminusCminus mice Asterisk indicatesthe atrial septal defect
H+C+ H+CminusMt
Mt
Mt
Mf
Mt
Mf
Figure 6 Transmission electron microscopy showing ultrastructure of myofibril defect in H+C+ neonatal heart Transmission electronmicroscopy results ofH+C+ (left) andH+Cminus (right)mice left ventricular tissueMfmyofibrilsMtmitochondria long arrow Z diskOriginalmagnification 10000x scale bar = 1 120583m
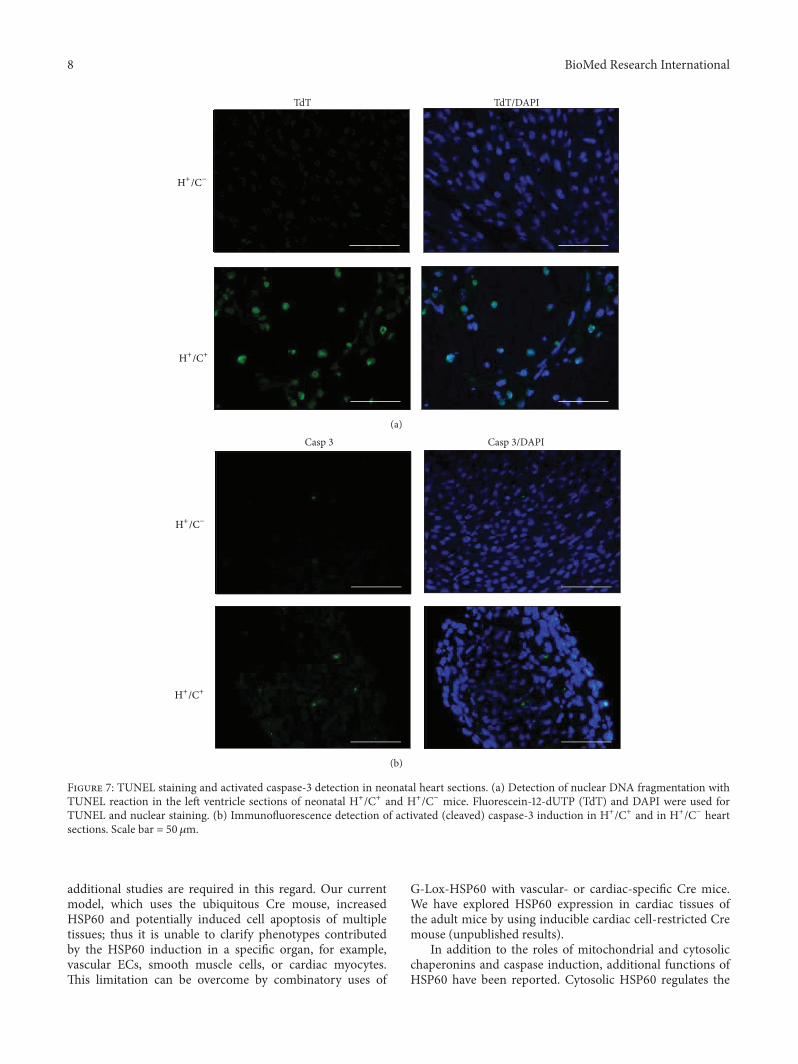
the cell nucleus through TUNEL assay whereas very fewTUNEL positively cells were in H+Cminus mouse heart sections(Figure 7(a)) In parallel a few cells in theHSP60mouse heartsection were positively stained by anti-activated caspase-3antibody significantly more than that of H+Cminus mouse heart(Figure 7(b)) The data clearly demonstrated that HSP60overexpression results in caspase-3 activation and apoptosisofmyocardial cells inHSP60mice Because theHSP60mouseheart became a sponge-like muscular tissue infiltrated bysmall capillaries and HSP60 involvement in rejection andautoimmune diseases was suggested previously we studiedthe presence of activated inflammatory cells in HSP60 miceThe pathology neither showed infiltrated mononuclear cells(Figure 4) nor detected CD4 or CD8 positive cells in HSP60mouse heart (Figure S3) thus inflammatory response orT cellactivation is absent in the HSP60 mouse heart
Accumulation of HSP60 in the cytoplasm during apop-tosis induction and the binding of HSP60 to procaspase-3and modulating caspase activity has been demonstrated inmany cancer cell lines [21 25 39] Similar to most proteins of
themitochondrialmatrix HSP60 which is synthesized in thecytoplasm with N-terminal targeting peptides is transportedto the mitochondrial matrix where the targeting sequenceis cleaved Both unprocessed (cytosolic) and matured (mito-chondrial) HSP60 strongly increased in the cardiac tissueof HSP60 mouse (Figure 1(b)) Consequently accumulationof cytosolic HSP60 capable of inducing apoptosis cannotbe overlooked and our data provided in vivo evidence ofapoptosis and caspase-3 activation in the HSP60mouse heart(Figure 7) Apoptosis is a highly regulated process duringembryonic and postnatal heart development and critical forheart development Increasing apoptosis in HSP60 mouseheart and its causal relation to cardiac myopathy must beinterpreted with caution because (1) apoptosis frequentlyoccurs in embryonic and neonatal tissues than in adultcardiac tissues (2) increasing apoptosis is often observedin cardiac tissues of myopathy and congestive heart failurethus apoptosis can be secondarily induced in myopathy andheart failure and (3) it can directly result from cytosolicHSP60 accumulation or HSP60 binding to caspases Thus
8 BioMed Research International
TdT TdTDAPI
H+Cminus
H+C+
(a)Casp 3DAPI
H+Cminus
H+C+
Casp 3
(b)
Figure 7 TUNEL staining and activated caspase-3 detection in neonatal heart sections (a) Detection of nuclear DNA fragmentation withTUNEL reaction in the left ventricle sections of neonatal H+C+ and H+Cminus mice Fluorescein-12-dUTP (TdT) and DAPI were used forTUNEL and nuclear staining (b) Immunofluorescence detection of activated (cleaved) caspase-3 induction in H+C+ and in H+Cminus heartsections Scale bar = 50120583m
additional studies are required in this regard Our currentmodel which uses the ubiquitous Cre mouse increasedHSP60 and potentially induced cell apoptosis of multipletissues thus it is unable to clarify phenotypes contributedby the HSP60 induction in a specific organ for examplevascular ECs smooth muscle cells or cardiac myocytesThis limitation can be overcome by combinatory uses of
G-Lox-HSP60 with vascular- or cardiac-specific Cre miceWe have explored HSP60 expression in cardiac tissues ofthe adult mice by using inducible cardiac cell-restricted Cremouse (unpublished results)
In addition to the roles of mitochondrial and cytosolicchaperonins and caspase induction additional functions ofHSP60 have been reported Cytosolic HSP60 regulates the
BioMed Research International 9
phosphorylation of IKK120572120573 and promotes NF-120581B prosur-vival pathway Transgenic expression of the cytosolic HSP60prevented stress-induced hepatic cell deaths in vivo [32]Overexpressing HSP60 in the human stem cells increasedHSP60 nuclear localization which suppressed ROS andp38JNK signaling and regulated genes relating to stem cellproliferation differentiation and stemness [40]Whether theaforementioned mechanisms are also involved in mediatingneonatal deaths remains unclear and future clarification isawaited
In this study congenital ASD was observed in a largenumber ofHSP60mice AlthoughASD exacerbated the exist-ing myopathy of HSP60 mice and was partially responsiblefor neonatal death ASD per sewas unlikely directly caused byHSP60 induction We speculate that ASD in neonatal HSP60mice may be the consequence of incomplete embryonicdevelopment existing myopathy and increasing apoptosisSimilar phenomenon has been demonstrated in the eNOSknockout mouse model in which high incidence of ASDor VSD was precipitated through increased apoptosis duringembryonic and postnatal stages [41] Our main findingsindicate that severe hemorrhage andmyocyte death occurredin mice with HSP60 expression and increased apoptosisin a macerated and spongy-like heart failed to meet thecardiovascular demands after birth and was the primarycause of HSP60-induced heart failure and neonatal death
4 Conclusions
HSP60 not only is the essentialmitochondrial chaperone pro-tein but also plays critical roles outside the mitochondrionAlthough numerous studies indicated the wide involvementof HSP60 in broad biological processes and many diseasesreports of transgenic mouse models of increasing or knock-down HSP60 expression are rare Thus the consequences ofaltering HSP60 levels in vivo in these disease settings remainunknown We used a floxed vector to establish conditionalHSP60 transgenic mice which allows studies on inducibleand organ-targeted HSP60 expression in adult mice and ispotentially useful in pursuing essential questions related toHSP60 in vivo In this study we show that ubiquitous HSP60expression from the embryonic stage results in increasedapoptosis myopathy high incidence of ASD and neonataldeaths
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Tsung-Hsien Chen and Shan-Wen Liu contribute equally tothis paper
Acknowledgments
The authors gratefully acknowledge the technical assistanceprovided by Tzu-Chin Wu and Mei-Yu Fan in NHRI-IBENand Functional and Micro-Magnetic Resonance ImagingCenter supported by NRPGM (NSC97-3112-B-001-013) Thisresearch was supported by grants from NHRI (BN-104-PP-02 BN-104-PP-24) and from Ministry of Science andTechnology (103-2320-B-400-018)
References
[1] I J Benjamin andD RMcMillan ldquoStress (heat shock) proteinsmolecular chaperones in cardiovascular biology and diseaserdquoCirculation Research vol 83 no 2 pp 117ndash132 1998
[2] F U Hartl ldquoMolecular chaperones in cellular protein foldingrdquoNature vol 381 no 6583 pp 571ndash580 1996
[3] J C Ranford and B Henderson ldquoChaperonins in diseasemechanismsmodels and treatmentsrdquoMolecular Pathology vol55 no 4 pp 209ndash213 2002
[4] CCDeocaris S CKaul andRWadhwa ldquoOn the brotherhoodof the mitochondrial chaperones mortalin and heat shockprotein 60rdquo Cell Stress and Chaperones vol 11 no 2 pp 116ndash128 2006
[5] F Cappello A M Gammazza A P Piccionello et al ldquoHsp60chaperonopathies and chaperonotherapy targets and agentsrdquoExpert Opinion on Therapeutic Targets vol 18 no 2 pp 185ndash208 2014
[6] S Lau N Patnaik M R Sayen and R Mestril ldquoSimultaneousoverexpression of two stress proteins in rat cardiomyocytes andmyogenic cells confers protection against ischemia-inducedinjuryrdquo Circulation vol 96 no 7 pp 2287ndash2294 1997
[7] K M Lin B Lin I Y Lian R Mestril I E Scheffler and WH Dillmann ldquoCombined and individual mitochondrial HSP60and HSP10 expression in cardiac myocytes protects mitochon-drial function and prevents apoptotic cell deaths induced bysimulated ischemia-reoxygenationrdquo Circulation vol 103 no 13pp 1787ndash1792 2001
[8] J J Malago J F J G Koninkx and J E van Dijk ldquoThe heatshock response and cytoprotection of the intestinal epitheliumrdquoCell Stress and Chaperones vol 7 no 2 pp 191ndash199 2002
[9] Y-X Shan T-L Yang R Mestril and P H Wang ldquoHsp10 andHsp60 suppress ubiquitination of insulin-like growth factor-1receptor and augment insulin-like growth factor-1 receptor sig-naling in cardiac muscle implications on decreased myocardialprotection in diabetic cardiomyopathyrdquoThe Journal of BiologicalChemistry vol 278 no 46 pp 45492ndash45498 2003
[10] H-S Chen Y-X Shan T-L Yang et al ldquoInsulin deficiencydownregulated heat shock protein 60 and IGF-1 receptorsignaling in diabetic myocardiumrdquo Diabetes vol 54 no 1 pp175ndash181 2005
10 BioMed Research International
[11] V Calabrese A Ravagna C Colombrita et al ldquoAcetylcarnitineinduces heme oxygenase in rat astrocytes and protects againstoxidative stress Involvement of the transcription factor Nrf2rdquoJournal of Neuroscience Research vol 79 no 4 pp 509ndash5212005
[12] V Veereshwarayya P Kumar K M Rosen R Mestril andH W Querfurth ldquoDifferential effects of mitochondrial heatshock protein 60 and related molecular chaperones to preventintracellular 120573-amyloid-induced inhibition of complex IV andlimit apoptosisrdquoThe Journal of Biological Chemistry vol 281 no40 pp 29468ndash29478 2006
[13] C Xu Y Lu Z Pan et al ldquoThe muscle-specific microRNAsmiR-1 and miR-133 produce opposing effects on apoptosis bytargeting HSP60 HSP70 and caspase-9 in cardiomyocytesrdquoJournal of Cell Science vol 120 part 17 pp 3045ndash3052 2007
[14] J C Ghosh T Dohi H K Byoung and D C Altieri ldquoHsp60regulation of tumor cell apoptosisrdquo The Journal of BiologicalChemistry vol 283 no 8 pp 5188ndash5194 2008
[15] P E Castle R Ashfaq F Ansari and C Y Muller ldquoImmuno-histochemical evaluation of heat shock proteins in normal andpreinvasive lesions of the cervixrdquo Cancer Letters vol 229 no 2pp 245ndash252 2005
[16] P A Cornford A R Dodson K F Parsons et al ldquoHeat shockprotein expression independently predicts clinical outcome inprostate cancerrdquoCancer Research vol 60 no 24 pp 7099ndash71052000
[17] F Cappello S David F Rappa et al ldquoThe expression of Hsp60and Hsp10 in large bowel carcinomas with lymph node metas-tasesrdquo BMC Cancer vol 5 article 39 2005
[18] P-L Hsu and S-M Hsu ldquoAbundance of heat shock proteins(hsp89 hsp60 and hsp27) in malignant cells of Hodgkinrsquosdiseaserdquo Cancer Research vol 58 no 23 pp 5507ndash5513 1998
[19] S Gupta and A A Knowlton ldquoHSP60 Bax apoptosis and theheartrdquo Journal of Cellular and Molecular Medicine vol 9 no 1pp 51ndash58 2005
[20] S Xanthoudakis R Sophie D Rasper et al ldquoHsp60 accel-erates the maturation of pro-caspase-3 by upstream activatorproteases during apoptosisrdquo The EMBO Journal vol 18 no 8pp 2049ndash2056 1999
[21] A Samali J Cai B Zhivotovsky D P Jones and S OrreniusldquoPresence of a pre-apoptotic complex of pro-caspase-3 Hsp60and Hsp10 in the mitochondrial fraction of Jurkat cellsrdquo TheEMBO Journal vol 18 no 8 pp 2040ndash2048 1999
[22] A Faried M Sohda M Nakajima TMiyazaki H Kato andHKuwano ldquoExpression of heat-shock protein Hsp60 correlatedwith the apoptotic index and patient prognosis in humanoesophageal squamous cell carcinomardquo European Journal ofCancer vol 40 no 18 pp 2804ndash2811 2004
[23] J Schneider E Jimenez K Marenbach H Romero D Marxand H Meden ldquoImmunohistochemical detection of HSP60-expression in human ovarian cancer Correlation with survivalin a series of 247 patientsrdquoAnticancer Research vol 19 no 3 pp2141ndash2146 1999
[24] T Lebret R W G Watson V Molinie et al ldquoHeat shockproteins HSP27 HSP60 HSP70 and HSP90 expression inbladder carcinomardquo Cancer vol 98 no 5 pp 970ndash977 2003
[25] A Samali and S Orrenius ldquoHeat shock proteins regulators ofstress response and apoptosisrdquo Cell Stress and Chaperones vol3 no 4 pp 228ndash236 1998
[26] W Van Eden G Wick S Albani and I Cohen ldquoStress heatshock proteins and autoimmunity how immune responses to
heat shock proteins are to be used for the control of chronicinflammatory diseasesrdquo Annals of the New York Academy ofSciences vol 1113 pp 217ndash237 2007
[27] C Habich and V Burkart ldquoHeat shock protein 60 regulatoryrole on innate immune cellsrdquo Cellular and Molecular LifeSciences vol 64 no 6 pp 742ndash751 2007
[28] Y Kondo Y Ueno K Kobayashi et al ldquoHepatitis B virus repli-cation could enhance regulatory T cell activity by producingsoluble heat shock protein 60 from hepatocytesrdquoThe Journal ofInfectious Diseases vol 202 no 2 pp 202ndash213 2010
[29] C Colotti G Cavallini R L Vitale et al ldquoEffects of agingand anti-aging caloric restrictions on carbonyl and heat shockprotein levels and expressionrdquo Biogerontology vol 6 no 6 pp397ndash406 2005
[30] P L Hooper ldquoInsulin signaling GSK-3 heat shock proteins andthe natural history of type 2 diabetes mellitus a hypothesisrdquoMetabolic Syndrome and RelatedDisorders vol 5 no 3 pp 220ndash230 2007
[31] O S Birk D C Douek D Elias et al ldquoA role of Hsp60 inautoimmune diabetes analysis in a transgenic modelrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 93 no 3 pp 1032ndash1037 1996
[32] J N Chun B Choi KW Lee et al ldquoCytosolicHsp60 is involvedin the NF-kappaB-dependent survival of cancer cells via IKKregulatiordquo PLoS ONE vol 5 no 3 Article ID e9422 2010
[33] M-R Chen S-W Liu T-C Wu et al ldquoRU486-induciblerecombination in the salivary glands of lactoferrin promoter-driven green fluorescent Cre transgenic micerdquo Genesis vol 48no 10 pp 585ndash595 2010
[34] H-Y Lin C-H Kao KM-C Lin V Kaartinen and L-T YangldquoNotch signaling regulates late-stage epidermal differentiationandmaintains postnatal hair cycle homeostasisrdquoPLoSONE vol6 no 1 Article ID e15842 2011
[35] A C Kayani G L Close W H Dillmann R Mestril M JJackson and A McArdle ldquoOverexpression of HSP10 in skeletalmuscle of transgenic mice prevents the age-related fall inmaximum tetanic force generation and muscle Cross-SectionalAreardquoThe American Journal of PhysiologymdashRegulatory Integra-tive and Comparative Physiology vol 299 no 1 pp R268ndashR2762010
[36] G-C Fan X Ren J Qian et al ldquoNovel cardioprotective role of asmall heat-shock protein Hsp20 against ischemiareperfusioninjuryrdquo Circulation vol 111 no 14 pp 1792ndash1799 2005
[37] J M Hollander J L Martin D D Belke et al ldquoOverex-pression of wild-type heat shock protein 27 and a nonphos-phorylatable heat shock protein 27 mutant protects againstischemiareperfusion injury in a transgenic mouse modelrdquoCirculation vol 110 no 23 pp 3544ndash3552 2004
[38] J-C L Plumier A M Krueger R W Currie D KontoyiannisG Kollias and G N Pagoulatos ldquoTransgenic mice expressingthe human inducible Hsp70 have hippocampal neurons resis-tant to ischemic injuryrdquo Cell Stress and Chaperones vol 2 no 3pp 162ndash167 1997
[39] D Chandra G Choy and D G Tang ldquoCytosolic accumulationof HSP60 during apoptosis with or without apparentmitochon-drial release Evidence that its pro-apoptotic or pro-survivalfunctions involve differential interactions with caspase-3rdquo TheJournal of Biological Chemistry vol 282 no 43 pp 31289ndash313012007
BioMed Research International 11
[40] J H Jang J S Jung J I Choi and S K Kang ldquoNuclearAgo2HSP60 contributes to broad spectrum of hATSCs func-tion via Oct4 regulationrdquoAntioxidants and Redox Signaling vol16 no 5 pp 383ndash399 2012
[41] Q Feng W Song X Lu et al ldquoDevelopment of heart failureand congenital septal defects in mice lacking endothelial nitricoxide synthaserdquo Circulation vol 106 no 7 pp 873ndash879 2002
HSP60 expression The findings of HSP60 accumulation incytoplasm in various apoptosis models and HSP60 bindingto procaspase-3 supported the proapoptotic properties ofHSP60 [21 25] Besides the discrepancy among reports of invitro studies the roles of HSP60 in physiology and pathologycannot be fully elucidated without using transgenic (Tg)mouse models expressing HSP60 in vivo
The role of extracellular HSP60 in viral infection innateor adaptive immunity and atherosclerosis has been widelydocumented [26ndash28] Hepatitis B virus replication inducedHSP60 production and HSP60 has a critical role in regu-latory T cells functions particularly in IL-10 secretion [28]Innate immune cell surface receptors namely TLR2 TLR4and CD14 are known receptors for extracellular HSP60resulting in the release of mediators such as TNF-120572 IL-1120573 IL-6 and NO which in turn enhance inflammation intype 1 diabetes mellitus (DM) atherosclerosis arthritis andtransplant rejection
HSP60 expression in the heart is reduced in ageing andmetabolic diseases [29] Caloric restriction that is shownincreases lifespan restored aging-related decline of HSP60expression in the heart and improved cardiovascular func-tions [29] The reduction in HSP60 level contributing to lowinsulin sensitivity and to dysfunctions inmetabolic syndromeand type 2 DMwas proposed [30] Because of the importanceof HSP60 in many diseases Tg mouse models with inducibleand tissue-specific HSP60 expression can provide usefulinsights and expand the current understanding of thesediseases
Previously nonobese diabetic (NOD) HIIE120572-HSP60 Tgmice in which HSP60 is driven by the MHC class II-E120572(HIIE120572) promoter achieved HSP60 expression specifically inthe thymus and bone marrow and suppressed susceptibilityto autoimmunity induced DM [31] Tg mice expressingtruncated HSP60 instead of the entire HSP60 were alsoreported and the resultant cytosolic HSP60 Tg mice wereresistant to hepatic stress with increased cell survival [32]However Tg mice with HSP60 expression in other tissueshave not been reported Attempts to generate a conventionalHSP60mouse model did not succeed because Tg founders aschimeras failed to survive (personal communications) In thisreport we present a viable and healthy inducible HSP60 Tgmouse (G-Lox-HSP60) in FVB strain driven by a ubiquitousCMV early enhancerchicken 120573-actin promoter (CAGGS)This model overcame the early lethality of the conventionalTg approach After crossing the G-Lox-HSP60 mouse withthe Cre mouse we observed neonatal death in mice withHSP60 expression from the embryo stage
2 Materials and Methods
21 Generation of G-Lox-Hsp60 and EGFP-Cre TransgenicMice A G-Lox-Hsp60 mouse FVBN-Tg(ACTB-EGFPHSPD1)14Klin was generated by using the Tg vector(Figure 1(a)) and constructed by inserting the followingfragments CAGGS promoter 2 LoxP sites at the start andthe end separated by an EGFP coding sequence and an SV40polyA sequence (pEGFP-C2 Clontech Mountain ViewCA) and a human HSP60 cDNA (NM 002156 102ndash1920 bp)
inserted into the pDsRed-N1 vector (Clontech) in whichthe CMV enhancer was removed An ApaLI-AflII fragmentwas used for microinjection into the FVB blastocystsEGFP-Cre mouse FVBN-Tg(ACTB-EGFPCRE)21Klinwas constructed by inserting CAGGS and Cre (AF3348271767ndash2789 bp) into the pEGFP-C2 vector in which the CMVenhancer was removed [33] For unknown reasons the greenfluorescence of the EGFP-Cre mouse was extremely weakG-Lox-DsRed mice have been described previously [34]All animal experiments were conducted in accordance withthe accepted standards of animal care and were approvedby the Institutional Animal Care and Use Committee of theNational Health Research Institutes Taiwan The followingPCR primers pairs were used for genotyping for the resultsin Figure 1(b)(A) (I) 51015840-AATGCTCACCGTAAGCCTTT-31015840 51015840-CCATCTGAAAGTTTTGCAAG-31015840 119879
119898= 655∘C
407 bp (II) 51015840-CTGCTAACCATGTTCATGCC-31015840 51015840-ACCGTCAGTACGTGAGATATCTT-31015840 119879
119898= 58∘C
1443 bp and (III) 51015840-CTGCTAACCATGTTCATGCC-3101584051015840-CCATCTGAAAGTTTTGCAAG-31015840 119879
119898= 60∘C 1442 bp
22 Analysis of mRNA Semiquantitative RT-PCR and ProteinExpression Total RNA was isolated from heart homogenateswith a TRIzol reagent (Invitrogen Carlsbad CA) GenomicDNA contamination in the RNA samples was removedthrough DNase I digestion and cDNA of the first strandwas synthesized using ReverTra Ace reverse transcriptase(Toyobo Japan) and oligo(dT) as the primer The followingPCR primers were used for amplifying the target genes for theresults in Figure 1(b)(B) Human HSP60 the same as listedin (I) human and mouse HSP60 51015840-GTCAGAAATGTG-AATTCCAG-31015840 51015840-TTGACTGCCACAACCTGAAGAC-31015840 119879119898= 55∘C 200 bp and GAPDH 51015840-GTGGCAAAGTGG-
AGATTGCC-31015840 51015840-GATGATGACCCGTTTGGCTCC-31015840119879119898= 58∘C 290 bp Ten micrograms of total proteins isolated
from tissue homogenates was subjected to 10 SDS-PAGEfollowed by Western blotting using anti-HSP60 (ab5479Abcam) and anti-GFP (ab290) antibodies Ten microgramsof total proteins isolated fromMDA-MB231 cells was used forcomparison
23 Fluorescence Detection and Histological Analysis Heartsof neonatal mice and E17 mouse embryos were washed withPBS and fixed by 10 neutral formalin Stereo microscope(Olympus SZX10) with fluorescence module was used forobserving green fluorescence of neonatal mouse hearts Forinvestigating septum defects unfixed neonatal hearts wereimmersed in OCT matrix (Tissue-Tek USA) and frozenquickly in liquid nitrogen The samples were cryosectioned10 120583m thick followed by hematoxylin and eosin (HampE)staining to reveal the four-chamber view of hearts In parallelformalin-fixed hearts were embedded in paraffin and cutinto serial 5 120583m sections and stained with HampE to evaluatemorphology
24 Immunohistochemical and Immunofluorescence AssayParaffin-embedded tissue sectionswere dewaxed rehydratedand treated by 3 hydrogen peroxide inmethanol for 30min
BioMed Research International 3
Cre
CAGGS promoter EGFP HSP60
LoxP LoxP
SV40pA SV40pA
Bgl II Hind III Hind III
DsRed
SCG-Lox-HSP60
(I)
CAGGS promoter HSP60
LoxP
SV40pA
DsRed
SC
(III)
Ase I
Ase I
CAGGS promoter EGFP Cre SV40pAEGFP-Cre
(II)
SC
(a)
(A)
(B) (C)
CAGGS-hHSP60
hHSP60
G-Lox-HSP60EGFP-Cre
(I)
(II)
(III)
EGFP-Cre
HSP60 (m h)
hHSP60
GAPDH
G-Lox-HSP60
EGFP-Cre
HSP60
EGFP
G-Lox-HSP60
EGFP-Cre
+
+ +
++
+ +
+minus
minus+ + + +
+++ minus minus
minus
+
minus minus
minus
minus
minus
minus
minusminus
minus
minus
minus
MD
A231
MD
A231
(b)
Figure 1 Generation of conditional human HSP60 transgenic mice (a) G-Lox-HSP60 and EGFP-Cre Tg vectorsThe CAGGS promoter wasused to drive both Tg vectors Placed between two LoxP sites the GFP cDNA with 3 stop codons (SC) and the SV40 polyA sequence wasused to inhibit the expression of the downstream transgene After the second LoxP site a full-length human HSP60 cDNA with endogenousSC and a portion of the last exon was inserted in front of ATG of the DsRedT1 cDNA sequence After the LoxP sites were rejoined using theCre DNA recombinase the HSP60DsRedT1 transcript was expressed however only HSP60 translated into proteins (b) (A) Analysis of fourpossible littermate genotypes using PCR on tail DNA (I) PCR amplification using the primer pair complementary to humanHSP60 but not tomouse HSP60 (II) amplification of EGFP-Cre and (III) the reaction to identify double transgenic (H+C+) mice by amplifying the abridgedsequence from CAGGS promoter to human HSP60 (B) RT-PCR results of human HSP60 mRNA expression in the neonatal mouse heartHSP60 (m h) indicates the PCR reaction to amplify HSP60 mRNA of both mouse and human origions GAPDH amplification served as theinternal control MDA-MB231 human breast tumor cells mRNA were used as the positive control for human HSP60 (C) Western blottingfor HSP60 and EGFP proteins in neonatal hearts Anti-HSP60 antibodies recognize both mouse and human HSP60
Tissue sections were then incubated in blocking solution(5 normal goat serum and 03 Triton-X 100 in PBS)followed by incubating with HSP60 (SC-1052 Santa CruzCA) overnight at 4∘CThe sections were rinsed and incubatedwith biotinylated donkey anti-goat immunoglobulin G (IgG)and then reacted to Vector Elite ABC in which the colorwas developed with diaminobenzidine Images were taken byusing the Olympus DP72 CCD attached to an Olympus BX51
microscope with DP controller For the immunofluorescencestaining antibodies of cleaved caspase-3 (Cell Signaling9961) CD4 (MAB554) and CD8120572 (MAB116) were used forincubating samples overnight in blocking solution at 4∘CAlexa Fluor 488 conjugated goat anti-rabbit IgG (H+L) orgoat anti-mouse IgG (H+L) secondary antibodies (Invitro-gen) were used instead The mounting media containingDAPI (Invitrogen)were used for counterstaining cell nucleus
4 BioMed Research International
25 TUNEL Assay Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay wasperformed for detecting nuclear DNA fragmentation inparaffin-embedded tissue sections as a measure of apoptosisFluorescein-12-dUTP was used as the substrate in TUNELreaction (DeadEnd Fluorometric TUNEL System Promega)the slides were counterstained by DAPI and the images weretaken by the Olympus microscope system
26 T2-Weighted Thoracic Region MR Images Magnetic res-onance imaging (MRI) was performed and images wereacquired using 94 Tesla MRI system (Bruker EttlingenGermany) with a maximum gradient strength of 140G cmminus1A quadrature volume coil (internal diameter = 10mm) wasused for RF transmission and reception We used a rapidacquisition with relaxation enhancement sequence to obtainthe T2-weighted images The repetition and echo times wereset at 2500 and 34 milliseconds respectively The field ofview was 10 times 10mm2 and the matrix size was 100 times 100 Theimages were sliced without gaps and the slice thickness was02mmThe voxel size for acquisitionwas 01times 01times 02mm3Finally the images were interpolated to 400times 400 using cubicinterpolation
27 Electron Micrograph Neonatal heart tissues were col-lected and fixed in 4 cold glutaraldehyde postfixed using1 osmium oxide and progressively dehydrated using alco-hol After resin embedding and polymerization 1 120583m thicksections were cut for initial observation Ultrathin (60ndash80 nmthick) sections were cut from the Epon-embedded blocksstained with uranyl acetate and lead citrate and examinedusing transmission electron microscopy (TEM Hitachi H-7500 Tokyo Japan) at 75 kV
3 Results and Discussion
31 Generation of HSP60 Conditional Transgenic MiceUnlike Tg mice expressing HSP10 [35] HSP20 [36] HSP27[37] or HSP70 [38] founders for establishing the ubiquitousHSP60 Tg mouse were not viable In this study we generateda conditional Tg mouse model using Cre-LoxP tools toallow inducible and tissue-specificHSP60 expression G-Lox-HSP60 mouse was generated by using the Tg vector illus-trated in Figure 1(a) CAGGSwas used to drive the expressionof transgenes enhanced green fluorescent protein (EGFP)or human HSP60 full-length cDNA Multiple stop codonsand the SV40 polyA sequences between the 2 LoxP sites ofthe Tg vector prevent the expression of downstream humanHSP60 sequence The G-Lox-HSP60 mice were normal indevelopment weight and reproduction with EGFP expres-sion in most organs During Cre-mediated recombination2 LoxP sites of the G-Lox-HSP60 vector are joined and theHSP60 expression is directly enhanced by CAGGS In thepresent study we generated an FVB ubiquitous Cre Tgmouseexpressing the EGFP-Cre fusion protein [33] We used thetail DNA-PCR method for studying the genotypes of the lit-termates in the crossing studies of G-Lox-HSP60 and EGFP-Cre mice the results are described in Figure 1(b) Deletion
Table 1 Summary of crossings between G-Lox-HSP60 (H) andEGFP-Cre (C) mice
Genotype H+C+ H+Cminus HminusC+ HminusCminus TotalNewborn number 72 76 69 86 303Neonatal death number 72 5 2 2 81Ratio 7272 576 269 286 81303H+C+ double Tg mouse H+Cminus HminusC+ single Tg mouse carrying eitherallele HminusCminus wild-type mouse
of the LoxP-flanked cassette was detected by the presence ofthe shorter PCR fragment in the double Tg mice Littermateswith all 4 possible genotypes were acquired in accordancewith the Mendelian frequency indicating no prenatal loss ofthe double Tgmice (H+C+) which were positive for both G-Lox-HSP60 and EGFP-Cre alleles (Table 1) Strong inductionof human HSP60 expression in the cardiac tissues of thedouble Tg mice was confirmed through RT-PCR Westernblotting (Figure 1(b)) and IHC staining (Figure 2)
32 HSP60 Expression Led to Neonatal Deaths In more than20 crossing experiments using G-Lox-HSP60 and EGFP-Cremice all double TgHSP60mice litters diedwithin a fewhoursafter birth A few survived the first day but none survivedmore than a few days (7272 Table 1) Neonatal deaths wererare in single Tg micemdashH+Cminus (576) and HminusC+ (269)mdashwhich carried only G-Lox-HSP60 or EGFP-Cre and in thewild-type mice (HminusCminus 286) At birth neonatal HSP60mice were slightly smaller in size than average with signsof mild cyanosis and internal bleeding at the abdominalsegments before death (Figure 3) Severe cardiovascular orpulmonary abnormality was suspected to contribute to theneonatal deaths in the HSP60 mice H+Cminus mice exhibitedstrong whole-body green fluorescence whereas the HSP60mice displayed green fluorescent spots indicating remainingEGFP that was produced before recombination HminusC+ andHminusCminus mice did not exhibit fluorescence Thoracic chamberdissection of HSP60 mice revealed a smaller heart and thecolor of the ventricles was more reddish containing moreblood than the hearts of other genotypes (Figure 3)
Postmortem analysis of the HSP60 mice was performedthrough MRI and histology MRI revealed that the HSP60P1 mouse heart was smaller than that of H+Cminus mouse(Figure S1 in Supplementary Material available online athttpdxdoiorg1011552015539805) The HSP60 mouseheart chambers and lungs were filled with liquids that wereabsent in those of H+CminusmouseThe interventricular septumand chamber walls of the HSP60 mice were thinner thannormal (Figure S1) Histological examination revealed mas-sive hemorrhage at the heart and lungs of double Tg mouseThe cardiac muscle appeared degenerated or incompletelydifferentiated with sponge-like cardiac tissues fenestratedwith blood or blood vessels and signs of myocardiumnecrosis were observed (Figure 4) The lungs of the HSP60mice appeared to have developed normally except for thepresence of hemorrhage that potentially resulted in onlypartial inflation of the lungs Other organs such as thebrain kidneys and liver of the HSP60 mice appeared grossly
BioMed Research International 5
HampE HSP60
H+Cminus
HminusCminus
H+C+
Figure 2 HSP60 detection in neonatal H+C+ mice heart HampE and immunohistochemical staining of HSP60 induction in H+C+ heartsections but not in H+Cminus or HminusCminus heart sections Scale bar = 50 120583m
normal (data not shown) Spotty hemorrhage was observedin the skeletal muscles of the neck spine and between theribs however the hemorrhage in these organs was not assevere as that in the heart and lungs Because cardiovascularand respiratory stresses were observed and because the heartswere smaller than normal we speculate that congenitaldefects may contribute to the neonatal deaths of the HSP60mice Of the 10 HSP60 mice studied for septal defects 4had atrial septal defect (ASD) and one had ventricular septaldefect (VSD) as illustrated in Figure 5 Septum defects werenot observed in the control mice
Because the atrial septum is closed when the lungsare inflated during birthing and because ASD manifests asa serious problem only after birth we examined whetherthe HSP60 mice had developmental defects during theembryonic stage in the E17 embryos No apparent abnor-malities were visible externally in the HSP60 mouse E17embryos However the embryo sections revealed excessivehemorrhage vascularization and tissue necrosis in the heartsimilar to those in the postnatal heart Lungs of E17 ofHSP60 mouse appeared grossly normal (Figure S2) Thushistological evidence suggested that congenital heart diseasecaused by cardiomyopathy or incomplete heart development
in HSP60 mouse embryos results in ASD or VSD whichaggravates respiratory stress and congestive heart failure atpostnatal day 1
We searched for ultrastructural abnormality as featuresof cardiac myopathy in neonatal HSP60 mouse using elec-tron micrographs G-Lox-HSP60 mouse contained orga-nized arrays of myofibril filaments with intact mitochon-drion appearing in high contrast with dense laminaecristae throughout the mitochondrial cross-sectional areas(Figure 6) In contrast the myofibrils of HSP60 mouse heartexhibited pronounced fragmentation and disorganizationshown as ragged Z lines with shorter sarcomere lengthon average compared with G-Lox-HSP60 myofibrils Mito-chondrion between myofibrils was mostly broken losingboth outline and inner cristae indicating a disintegratedmitochondrial structureThus TEM results confirmed severecardiac myopathy in HSP60 mouse
33 Increasing Apoptosis in HSP60 Mouse Heart HSP60 waspreviously linked to apoptosis induction thus we studiedapoptotic cardiac myocytes in the HSP60 mouse by usingthe TUNEL assay Many cells in the HSP60 mouse heartsections were positively labeled by fluorescein-dUTP at
6 BioMed Research International
H+Cminus
HminusC+
HminusCminus
H+C+
Figure 3 Neonatal Tg mice Left neonatal Tg miceThe white arrow indicates cyanosis and abdominal bleeding in H+C+ neonates middlefluorescent images of the same mice right the lungs and heart of neonatal mice H+C+ hearts appear more dark reddish and smaller thanhearts of other genotypes Scale bar = 3mm
H+CminusH+C+
Figure 4 Histology of the neonatal heart and lung Top HampE staining of the neonatal heart of H+C+ mouse compared with that of H+Cminusmouse showing hemorrhage necrosis and degenerated cardiac muscles in H+C+ mouse heart Bottom the lung of H+C+ mouse showssigns of hemorrhage liquid congestion and tissue necrosis The alveolar lung structures appeared to have developed normally but poorlyinflated Scale bar = 100 120583m
BioMed Research International 7
H+C+ HminusCminus
Figure 5 Atrial septal defect in H+C+ neonatal mice HampE images of the neonatal heart from H+C+ and HminusCminus mice Asterisk indicatesthe atrial septal defect
H+C+ H+CminusMt
Mt
Mt
Mf
Mt
Mf
Figure 6 Transmission electron microscopy showing ultrastructure of myofibril defect in H+C+ neonatal heart Transmission electronmicroscopy results ofH+C+ (left) andH+Cminus (right)mice left ventricular tissueMfmyofibrilsMtmitochondria long arrow Z diskOriginalmagnification 10000x scale bar = 1 120583m
the cell nucleus through TUNEL assay whereas very fewTUNEL positively cells were in H+Cminus mouse heart sections(Figure 7(a)) In parallel a few cells in theHSP60mouse heartsection were positively stained by anti-activated caspase-3antibody significantly more than that of H+Cminus mouse heart(Figure 7(b)) The data clearly demonstrated that HSP60overexpression results in caspase-3 activation and apoptosisofmyocardial cells inHSP60mice Because theHSP60mouseheart became a sponge-like muscular tissue infiltrated bysmall capillaries and HSP60 involvement in rejection andautoimmune diseases was suggested previously we studiedthe presence of activated inflammatory cells in HSP60 miceThe pathology neither showed infiltrated mononuclear cells(Figure 4) nor detected CD4 or CD8 positive cells in HSP60mouse heart (Figure S3) thus inflammatory response orT cellactivation is absent in the HSP60 mouse heart
Accumulation of HSP60 in the cytoplasm during apop-tosis induction and the binding of HSP60 to procaspase-3and modulating caspase activity has been demonstrated inmany cancer cell lines [21 25 39] Similar to most proteins of
themitochondrialmatrix HSP60 which is synthesized in thecytoplasm with N-terminal targeting peptides is transportedto the mitochondrial matrix where the targeting sequenceis cleaved Both unprocessed (cytosolic) and matured (mito-chondrial) HSP60 strongly increased in the cardiac tissueof HSP60 mouse (Figure 1(b)) Consequently accumulationof cytosolic HSP60 capable of inducing apoptosis cannotbe overlooked and our data provided in vivo evidence ofapoptosis and caspase-3 activation in the HSP60mouse heart(Figure 7) Apoptosis is a highly regulated process duringembryonic and postnatal heart development and critical forheart development Increasing apoptosis in HSP60 mouseheart and its causal relation to cardiac myopathy must beinterpreted with caution because (1) apoptosis frequentlyoccurs in embryonic and neonatal tissues than in adultcardiac tissues (2) increasing apoptosis is often observedin cardiac tissues of myopathy and congestive heart failurethus apoptosis can be secondarily induced in myopathy andheart failure and (3) it can directly result from cytosolicHSP60 accumulation or HSP60 binding to caspases Thus
8 BioMed Research International
TdT TdTDAPI
H+Cminus
H+C+
(a)Casp 3DAPI
H+Cminus
H+C+
Casp 3
(b)
Figure 7 TUNEL staining and activated caspase-3 detection in neonatal heart sections (a) Detection of nuclear DNA fragmentation withTUNEL reaction in the left ventricle sections of neonatal H+C+ and H+Cminus mice Fluorescein-12-dUTP (TdT) and DAPI were used forTUNEL and nuclear staining (b) Immunofluorescence detection of activated (cleaved) caspase-3 induction in H+C+ and in H+Cminus heartsections Scale bar = 50120583m
additional studies are required in this regard Our currentmodel which uses the ubiquitous Cre mouse increasedHSP60 and potentially induced cell apoptosis of multipletissues thus it is unable to clarify phenotypes contributedby the HSP60 induction in a specific organ for examplevascular ECs smooth muscle cells or cardiac myocytesThis limitation can be overcome by combinatory uses of
G-Lox-HSP60 with vascular- or cardiac-specific Cre miceWe have explored HSP60 expression in cardiac tissues ofthe adult mice by using inducible cardiac cell-restricted Cremouse (unpublished results)
In addition to the roles of mitochondrial and cytosolicchaperonins and caspase induction additional functions ofHSP60 have been reported Cytosolic HSP60 regulates the
BioMed Research International 9
phosphorylation of IKK120572120573 and promotes NF-120581B prosur-vival pathway Transgenic expression of the cytosolic HSP60prevented stress-induced hepatic cell deaths in vivo [32]Overexpressing HSP60 in the human stem cells increasedHSP60 nuclear localization which suppressed ROS andp38JNK signaling and regulated genes relating to stem cellproliferation differentiation and stemness [40]Whether theaforementioned mechanisms are also involved in mediatingneonatal deaths remains unclear and future clarification isawaited
In this study congenital ASD was observed in a largenumber ofHSP60mice AlthoughASD exacerbated the exist-ing myopathy of HSP60 mice and was partially responsiblefor neonatal death ASD per sewas unlikely directly caused byHSP60 induction We speculate that ASD in neonatal HSP60mice may be the consequence of incomplete embryonicdevelopment existing myopathy and increasing apoptosisSimilar phenomenon has been demonstrated in the eNOSknockout mouse model in which high incidence of ASDor VSD was precipitated through increased apoptosis duringembryonic and postnatal stages [41] Our main findingsindicate that severe hemorrhage andmyocyte death occurredin mice with HSP60 expression and increased apoptosisin a macerated and spongy-like heart failed to meet thecardiovascular demands after birth and was the primarycause of HSP60-induced heart failure and neonatal death
4 Conclusions
HSP60 not only is the essentialmitochondrial chaperone pro-tein but also plays critical roles outside the mitochondrionAlthough numerous studies indicated the wide involvementof HSP60 in broad biological processes and many diseasesreports of transgenic mouse models of increasing or knock-down HSP60 expression are rare Thus the consequences ofaltering HSP60 levels in vivo in these disease settings remainunknown We used a floxed vector to establish conditionalHSP60 transgenic mice which allows studies on inducibleand organ-targeted HSP60 expression in adult mice and ispotentially useful in pursuing essential questions related toHSP60 in vivo In this study we show that ubiquitous HSP60expression from the embryonic stage results in increasedapoptosis myopathy high incidence of ASD and neonataldeaths
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Tsung-Hsien Chen and Shan-Wen Liu contribute equally tothis paper
Acknowledgments
The authors gratefully acknowledge the technical assistanceprovided by Tzu-Chin Wu and Mei-Yu Fan in NHRI-IBENand Functional and Micro-Magnetic Resonance ImagingCenter supported by NRPGM (NSC97-3112-B-001-013) Thisresearch was supported by grants from NHRI (BN-104-PP-02 BN-104-PP-24) and from Ministry of Science andTechnology (103-2320-B-400-018)
References
[1] I J Benjamin andD RMcMillan ldquoStress (heat shock) proteinsmolecular chaperones in cardiovascular biology and diseaserdquoCirculation Research vol 83 no 2 pp 117ndash132 1998
[2] F U Hartl ldquoMolecular chaperones in cellular protein foldingrdquoNature vol 381 no 6583 pp 571ndash580 1996
[3] J C Ranford and B Henderson ldquoChaperonins in diseasemechanismsmodels and treatmentsrdquoMolecular Pathology vol55 no 4 pp 209ndash213 2002
[4] CCDeocaris S CKaul andRWadhwa ldquoOn the brotherhoodof the mitochondrial chaperones mortalin and heat shockprotein 60rdquo Cell Stress and Chaperones vol 11 no 2 pp 116ndash128 2006
[5] F Cappello A M Gammazza A P Piccionello et al ldquoHsp60chaperonopathies and chaperonotherapy targets and agentsrdquoExpert Opinion on Therapeutic Targets vol 18 no 2 pp 185ndash208 2014
[6] S Lau N Patnaik M R Sayen and R Mestril ldquoSimultaneousoverexpression of two stress proteins in rat cardiomyocytes andmyogenic cells confers protection against ischemia-inducedinjuryrdquo Circulation vol 96 no 7 pp 2287ndash2294 1997
[7] K M Lin B Lin I Y Lian R Mestril I E Scheffler and WH Dillmann ldquoCombined and individual mitochondrial HSP60and HSP10 expression in cardiac myocytes protects mitochon-drial function and prevents apoptotic cell deaths induced bysimulated ischemia-reoxygenationrdquo Circulation vol 103 no 13pp 1787ndash1792 2001
[8] J J Malago J F J G Koninkx and J E van Dijk ldquoThe heatshock response and cytoprotection of the intestinal epitheliumrdquoCell Stress and Chaperones vol 7 no 2 pp 191ndash199 2002
[9] Y-X Shan T-L Yang R Mestril and P H Wang ldquoHsp10 andHsp60 suppress ubiquitination of insulin-like growth factor-1receptor and augment insulin-like growth factor-1 receptor sig-naling in cardiac muscle implications on decreased myocardialprotection in diabetic cardiomyopathyrdquoThe Journal of BiologicalChemistry vol 278 no 46 pp 45492ndash45498 2003
[10] H-S Chen Y-X Shan T-L Yang et al ldquoInsulin deficiencydownregulated heat shock protein 60 and IGF-1 receptorsignaling in diabetic myocardiumrdquo Diabetes vol 54 no 1 pp175ndash181 2005
10 BioMed Research International
[11] V Calabrese A Ravagna C Colombrita et al ldquoAcetylcarnitineinduces heme oxygenase in rat astrocytes and protects againstoxidative stress Involvement of the transcription factor Nrf2rdquoJournal of Neuroscience Research vol 79 no 4 pp 509ndash5212005
[12] V Veereshwarayya P Kumar K M Rosen R Mestril andH W Querfurth ldquoDifferential effects of mitochondrial heatshock protein 60 and related molecular chaperones to preventintracellular 120573-amyloid-induced inhibition of complex IV andlimit apoptosisrdquoThe Journal of Biological Chemistry vol 281 no40 pp 29468ndash29478 2006
[13] C Xu Y Lu Z Pan et al ldquoThe muscle-specific microRNAsmiR-1 and miR-133 produce opposing effects on apoptosis bytargeting HSP60 HSP70 and caspase-9 in cardiomyocytesrdquoJournal of Cell Science vol 120 part 17 pp 3045ndash3052 2007
[14] J C Ghosh T Dohi H K Byoung and D C Altieri ldquoHsp60regulation of tumor cell apoptosisrdquo The Journal of BiologicalChemistry vol 283 no 8 pp 5188ndash5194 2008
[15] P E Castle R Ashfaq F Ansari and C Y Muller ldquoImmuno-histochemical evaluation of heat shock proteins in normal andpreinvasive lesions of the cervixrdquo Cancer Letters vol 229 no 2pp 245ndash252 2005
[16] P A Cornford A R Dodson K F Parsons et al ldquoHeat shockprotein expression independently predicts clinical outcome inprostate cancerrdquoCancer Research vol 60 no 24 pp 7099ndash71052000
[17] F Cappello S David F Rappa et al ldquoThe expression of Hsp60and Hsp10 in large bowel carcinomas with lymph node metas-tasesrdquo BMC Cancer vol 5 article 39 2005
[18] P-L Hsu and S-M Hsu ldquoAbundance of heat shock proteins(hsp89 hsp60 and hsp27) in malignant cells of Hodgkinrsquosdiseaserdquo Cancer Research vol 58 no 23 pp 5507ndash5513 1998
[19] S Gupta and A A Knowlton ldquoHSP60 Bax apoptosis and theheartrdquo Journal of Cellular and Molecular Medicine vol 9 no 1pp 51ndash58 2005
[20] S Xanthoudakis R Sophie D Rasper et al ldquoHsp60 accel-erates the maturation of pro-caspase-3 by upstream activatorproteases during apoptosisrdquo The EMBO Journal vol 18 no 8pp 2049ndash2056 1999
[21] A Samali J Cai B Zhivotovsky D P Jones and S OrreniusldquoPresence of a pre-apoptotic complex of pro-caspase-3 Hsp60and Hsp10 in the mitochondrial fraction of Jurkat cellsrdquo TheEMBO Journal vol 18 no 8 pp 2040ndash2048 1999
[22] A Faried M Sohda M Nakajima TMiyazaki H Kato andHKuwano ldquoExpression of heat-shock protein Hsp60 correlatedwith the apoptotic index and patient prognosis in humanoesophageal squamous cell carcinomardquo European Journal ofCancer vol 40 no 18 pp 2804ndash2811 2004
[23] J Schneider E Jimenez K Marenbach H Romero D Marxand H Meden ldquoImmunohistochemical detection of HSP60-expression in human ovarian cancer Correlation with survivalin a series of 247 patientsrdquoAnticancer Research vol 19 no 3 pp2141ndash2146 1999
[24] T Lebret R W G Watson V Molinie et al ldquoHeat shockproteins HSP27 HSP60 HSP70 and HSP90 expression inbladder carcinomardquo Cancer vol 98 no 5 pp 970ndash977 2003
[25] A Samali and S Orrenius ldquoHeat shock proteins regulators ofstress response and apoptosisrdquo Cell Stress and Chaperones vol3 no 4 pp 228ndash236 1998
[26] W Van Eden G Wick S Albani and I Cohen ldquoStress heatshock proteins and autoimmunity how immune responses to
heat shock proteins are to be used for the control of chronicinflammatory diseasesrdquo Annals of the New York Academy ofSciences vol 1113 pp 217ndash237 2007
[27] C Habich and V Burkart ldquoHeat shock protein 60 regulatoryrole on innate immune cellsrdquo Cellular and Molecular LifeSciences vol 64 no 6 pp 742ndash751 2007
[28] Y Kondo Y Ueno K Kobayashi et al ldquoHepatitis B virus repli-cation could enhance regulatory T cell activity by producingsoluble heat shock protein 60 from hepatocytesrdquoThe Journal ofInfectious Diseases vol 202 no 2 pp 202ndash213 2010
[29] C Colotti G Cavallini R L Vitale et al ldquoEffects of agingand anti-aging caloric restrictions on carbonyl and heat shockprotein levels and expressionrdquo Biogerontology vol 6 no 6 pp397ndash406 2005
[30] P L Hooper ldquoInsulin signaling GSK-3 heat shock proteins andthe natural history of type 2 diabetes mellitus a hypothesisrdquoMetabolic Syndrome and RelatedDisorders vol 5 no 3 pp 220ndash230 2007
[31] O S Birk D C Douek D Elias et al ldquoA role of Hsp60 inautoimmune diabetes analysis in a transgenic modelrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 93 no 3 pp 1032ndash1037 1996
[32] J N Chun B Choi KW Lee et al ldquoCytosolicHsp60 is involvedin the NF-kappaB-dependent survival of cancer cells via IKKregulatiordquo PLoS ONE vol 5 no 3 Article ID e9422 2010
[33] M-R Chen S-W Liu T-C Wu et al ldquoRU486-induciblerecombination in the salivary glands of lactoferrin promoter-driven green fluorescent Cre transgenic micerdquo Genesis vol 48no 10 pp 585ndash595 2010
[34] H-Y Lin C-H Kao KM-C Lin V Kaartinen and L-T YangldquoNotch signaling regulates late-stage epidermal differentiationandmaintains postnatal hair cycle homeostasisrdquoPLoSONE vol6 no 1 Article ID e15842 2011
[35] A C Kayani G L Close W H Dillmann R Mestril M JJackson and A McArdle ldquoOverexpression of HSP10 in skeletalmuscle of transgenic mice prevents the age-related fall inmaximum tetanic force generation and muscle Cross-SectionalAreardquoThe American Journal of PhysiologymdashRegulatory Integra-tive and Comparative Physiology vol 299 no 1 pp R268ndashR2762010
[36] G-C Fan X Ren J Qian et al ldquoNovel cardioprotective role of asmall heat-shock protein Hsp20 against ischemiareperfusioninjuryrdquo Circulation vol 111 no 14 pp 1792ndash1799 2005
[37] J M Hollander J L Martin D D Belke et al ldquoOverex-pression of wild-type heat shock protein 27 and a nonphos-phorylatable heat shock protein 27 mutant protects againstischemiareperfusion injury in a transgenic mouse modelrdquoCirculation vol 110 no 23 pp 3544ndash3552 2004
[38] J-C L Plumier A M Krueger R W Currie D KontoyiannisG Kollias and G N Pagoulatos ldquoTransgenic mice expressingthe human inducible Hsp70 have hippocampal neurons resis-tant to ischemic injuryrdquo Cell Stress and Chaperones vol 2 no 3pp 162ndash167 1997
[39] D Chandra G Choy and D G Tang ldquoCytosolic accumulationof HSP60 during apoptosis with or without apparentmitochon-drial release Evidence that its pro-apoptotic or pro-survivalfunctions involve differential interactions with caspase-3rdquo TheJournal of Biological Chemistry vol 282 no 43 pp 31289ndash313012007
BioMed Research International 11
[40] J H Jang J S Jung J I Choi and S K Kang ldquoNuclearAgo2HSP60 contributes to broad spectrum of hATSCs func-tion via Oct4 regulationrdquoAntioxidants and Redox Signaling vol16 no 5 pp 383ndash399 2012
[41] Q Feng W Song X Lu et al ldquoDevelopment of heart failureand congenital septal defects in mice lacking endothelial nitricoxide synthaserdquo Circulation vol 106 no 7 pp 873ndash879 2002
Figure 1 Generation of conditional human HSP60 transgenic mice (a) G-Lox-HSP60 and EGFP-Cre Tg vectorsThe CAGGS promoter wasused to drive both Tg vectors Placed between two LoxP sites the GFP cDNA with 3 stop codons (SC) and the SV40 polyA sequence wasused to inhibit the expression of the downstream transgene After the second LoxP site a full-length human HSP60 cDNA with endogenousSC and a portion of the last exon was inserted in front of ATG of the DsRedT1 cDNA sequence After the LoxP sites were rejoined using theCre DNA recombinase the HSP60DsRedT1 transcript was expressed however only HSP60 translated into proteins (b) (A) Analysis of fourpossible littermate genotypes using PCR on tail DNA (I) PCR amplification using the primer pair complementary to humanHSP60 but not tomouse HSP60 (II) amplification of EGFP-Cre and (III) the reaction to identify double transgenic (H+C+) mice by amplifying the abridgedsequence from CAGGS promoter to human HSP60 (B) RT-PCR results of human HSP60 mRNA expression in the neonatal mouse heartHSP60 (m h) indicates the PCR reaction to amplify HSP60 mRNA of both mouse and human origions GAPDH amplification served as theinternal control MDA-MB231 human breast tumor cells mRNA were used as the positive control for human HSP60 (C) Western blottingfor HSP60 and EGFP proteins in neonatal hearts Anti-HSP60 antibodies recognize both mouse and human HSP60
Tissue sections were then incubated in blocking solution(5 normal goat serum and 03 Triton-X 100 in PBS)followed by incubating with HSP60 (SC-1052 Santa CruzCA) overnight at 4∘CThe sections were rinsed and incubatedwith biotinylated donkey anti-goat immunoglobulin G (IgG)and then reacted to Vector Elite ABC in which the colorwas developed with diaminobenzidine Images were taken byusing the Olympus DP72 CCD attached to an Olympus BX51
microscope with DP controller For the immunofluorescencestaining antibodies of cleaved caspase-3 (Cell Signaling9961) CD4 (MAB554) and CD8120572 (MAB116) were used forincubating samples overnight in blocking solution at 4∘CAlexa Fluor 488 conjugated goat anti-rabbit IgG (H+L) orgoat anti-mouse IgG (H+L) secondary antibodies (Invitro-gen) were used instead The mounting media containingDAPI (Invitrogen)were used for counterstaining cell nucleus
4 BioMed Research International
25 TUNEL Assay Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay wasperformed for detecting nuclear DNA fragmentation inparaffin-embedded tissue sections as a measure of apoptosisFluorescein-12-dUTP was used as the substrate in TUNELreaction (DeadEnd Fluorometric TUNEL System Promega)the slides were counterstained by DAPI and the images weretaken by the Olympus microscope system
26 T2-Weighted Thoracic Region MR Images Magnetic res-onance imaging (MRI) was performed and images wereacquired using 94 Tesla MRI system (Bruker EttlingenGermany) with a maximum gradient strength of 140G cmminus1A quadrature volume coil (internal diameter = 10mm) wasused for RF transmission and reception We used a rapidacquisition with relaxation enhancement sequence to obtainthe T2-weighted images The repetition and echo times wereset at 2500 and 34 milliseconds respectively The field ofview was 10 times 10mm2 and the matrix size was 100 times 100 Theimages were sliced without gaps and the slice thickness was02mmThe voxel size for acquisitionwas 01times 01times 02mm3Finally the images were interpolated to 400times 400 using cubicinterpolation
27 Electron Micrograph Neonatal heart tissues were col-lected and fixed in 4 cold glutaraldehyde postfixed using1 osmium oxide and progressively dehydrated using alco-hol After resin embedding and polymerization 1 120583m thicksections were cut for initial observation Ultrathin (60ndash80 nmthick) sections were cut from the Epon-embedded blocksstained with uranyl acetate and lead citrate and examinedusing transmission electron microscopy (TEM Hitachi H-7500 Tokyo Japan) at 75 kV
3 Results and Discussion
31 Generation of HSP60 Conditional Transgenic MiceUnlike Tg mice expressing HSP10 [35] HSP20 [36] HSP27[37] or HSP70 [38] founders for establishing the ubiquitousHSP60 Tg mouse were not viable In this study we generateda conditional Tg mouse model using Cre-LoxP tools toallow inducible and tissue-specificHSP60 expression G-Lox-HSP60 mouse was generated by using the Tg vector illus-trated in Figure 1(a) CAGGSwas used to drive the expressionof transgenes enhanced green fluorescent protein (EGFP)or human HSP60 full-length cDNA Multiple stop codonsand the SV40 polyA sequences between the 2 LoxP sites ofthe Tg vector prevent the expression of downstream humanHSP60 sequence The G-Lox-HSP60 mice were normal indevelopment weight and reproduction with EGFP expres-sion in most organs During Cre-mediated recombination2 LoxP sites of the G-Lox-HSP60 vector are joined and theHSP60 expression is directly enhanced by CAGGS In thepresent study we generated an FVB ubiquitous Cre Tgmouseexpressing the EGFP-Cre fusion protein [33] We used thetail DNA-PCR method for studying the genotypes of the lit-termates in the crossing studies of G-Lox-HSP60 and EGFP-Cre mice the results are described in Figure 1(b) Deletion
Table 1 Summary of crossings between G-Lox-HSP60 (H) andEGFP-Cre (C) mice
Genotype H+C+ H+Cminus HminusC+ HminusCminus TotalNewborn number 72 76 69 86 303Neonatal death number 72 5 2 2 81Ratio 7272 576 269 286 81303H+C+ double Tg mouse H+Cminus HminusC+ single Tg mouse carrying eitherallele HminusCminus wild-type mouse
of the LoxP-flanked cassette was detected by the presence ofthe shorter PCR fragment in the double Tg mice Littermateswith all 4 possible genotypes were acquired in accordancewith the Mendelian frequency indicating no prenatal loss ofthe double Tgmice (H+C+) which were positive for both G-Lox-HSP60 and EGFP-Cre alleles (Table 1) Strong inductionof human HSP60 expression in the cardiac tissues of thedouble Tg mice was confirmed through RT-PCR Westernblotting (Figure 1(b)) and IHC staining (Figure 2)
32 HSP60 Expression Led to Neonatal Deaths In more than20 crossing experiments using G-Lox-HSP60 and EGFP-Cremice all double TgHSP60mice litters diedwithin a fewhoursafter birth A few survived the first day but none survivedmore than a few days (7272 Table 1) Neonatal deaths wererare in single Tg micemdashH+Cminus (576) and HminusC+ (269)mdashwhich carried only G-Lox-HSP60 or EGFP-Cre and in thewild-type mice (HminusCminus 286) At birth neonatal HSP60mice were slightly smaller in size than average with signsof mild cyanosis and internal bleeding at the abdominalsegments before death (Figure 3) Severe cardiovascular orpulmonary abnormality was suspected to contribute to theneonatal deaths in the HSP60 mice H+Cminus mice exhibitedstrong whole-body green fluorescence whereas the HSP60mice displayed green fluorescent spots indicating remainingEGFP that was produced before recombination HminusC+ andHminusCminus mice did not exhibit fluorescence Thoracic chamberdissection of HSP60 mice revealed a smaller heart and thecolor of the ventricles was more reddish containing moreblood than the hearts of other genotypes (Figure 3)
Postmortem analysis of the HSP60 mice was performedthrough MRI and histology MRI revealed that the HSP60P1 mouse heart was smaller than that of H+Cminus mouse(Figure S1 in Supplementary Material available online athttpdxdoiorg1011552015539805) The HSP60 mouseheart chambers and lungs were filled with liquids that wereabsent in those of H+CminusmouseThe interventricular septumand chamber walls of the HSP60 mice were thinner thannormal (Figure S1) Histological examination revealed mas-sive hemorrhage at the heart and lungs of double Tg mouseThe cardiac muscle appeared degenerated or incompletelydifferentiated with sponge-like cardiac tissues fenestratedwith blood or blood vessels and signs of myocardiumnecrosis were observed (Figure 4) The lungs of the HSP60mice appeared to have developed normally except for thepresence of hemorrhage that potentially resulted in onlypartial inflation of the lungs Other organs such as thebrain kidneys and liver of the HSP60 mice appeared grossly
BioMed Research International 5
HampE HSP60
H+Cminus
HminusCminus
H+C+
Figure 2 HSP60 detection in neonatal H+C+ mice heart HampE and immunohistochemical staining of HSP60 induction in H+C+ heartsections but not in H+Cminus or HminusCminus heart sections Scale bar = 50 120583m
normal (data not shown) Spotty hemorrhage was observedin the skeletal muscles of the neck spine and between theribs however the hemorrhage in these organs was not assevere as that in the heart and lungs Because cardiovascularand respiratory stresses were observed and because the heartswere smaller than normal we speculate that congenitaldefects may contribute to the neonatal deaths of the HSP60mice Of the 10 HSP60 mice studied for septal defects 4had atrial septal defect (ASD) and one had ventricular septaldefect (VSD) as illustrated in Figure 5 Septum defects werenot observed in the control mice
Because the atrial septum is closed when the lungsare inflated during birthing and because ASD manifests asa serious problem only after birth we examined whetherthe HSP60 mice had developmental defects during theembryonic stage in the E17 embryos No apparent abnor-malities were visible externally in the HSP60 mouse E17embryos However the embryo sections revealed excessivehemorrhage vascularization and tissue necrosis in the heartsimilar to those in the postnatal heart Lungs of E17 ofHSP60 mouse appeared grossly normal (Figure S2) Thushistological evidence suggested that congenital heart diseasecaused by cardiomyopathy or incomplete heart development
in HSP60 mouse embryos results in ASD or VSD whichaggravates respiratory stress and congestive heart failure atpostnatal day 1
We searched for ultrastructural abnormality as featuresof cardiac myopathy in neonatal HSP60 mouse using elec-tron micrographs G-Lox-HSP60 mouse contained orga-nized arrays of myofibril filaments with intact mitochon-drion appearing in high contrast with dense laminaecristae throughout the mitochondrial cross-sectional areas(Figure 6) In contrast the myofibrils of HSP60 mouse heartexhibited pronounced fragmentation and disorganizationshown as ragged Z lines with shorter sarcomere lengthon average compared with G-Lox-HSP60 myofibrils Mito-chondrion between myofibrils was mostly broken losingboth outline and inner cristae indicating a disintegratedmitochondrial structureThus TEM results confirmed severecardiac myopathy in HSP60 mouse
33 Increasing Apoptosis in HSP60 Mouse Heart HSP60 waspreviously linked to apoptosis induction thus we studiedapoptotic cardiac myocytes in the HSP60 mouse by usingthe TUNEL assay Many cells in the HSP60 mouse heartsections were positively labeled by fluorescein-dUTP at
6 BioMed Research International
H+Cminus
HminusC+
HminusCminus
H+C+
Figure 3 Neonatal Tg mice Left neonatal Tg miceThe white arrow indicates cyanosis and abdominal bleeding in H+C+ neonates middlefluorescent images of the same mice right the lungs and heart of neonatal mice H+C+ hearts appear more dark reddish and smaller thanhearts of other genotypes Scale bar = 3mm
H+CminusH+C+
Figure 4 Histology of the neonatal heart and lung Top HampE staining of the neonatal heart of H+C+ mouse compared with that of H+Cminusmouse showing hemorrhage necrosis and degenerated cardiac muscles in H+C+ mouse heart Bottom the lung of H+C+ mouse showssigns of hemorrhage liquid congestion and tissue necrosis The alveolar lung structures appeared to have developed normally but poorlyinflated Scale bar = 100 120583m
BioMed Research International 7
H+C+ HminusCminus
Figure 5 Atrial septal defect in H+C+ neonatal mice HampE images of the neonatal heart from H+C+ and HminusCminus mice Asterisk indicatesthe atrial septal defect
H+C+ H+CminusMt
Mt
Mt
Mf
Mt
Mf
Figure 6 Transmission electron microscopy showing ultrastructure of myofibril defect in H+C+ neonatal heart Transmission electronmicroscopy results ofH+C+ (left) andH+Cminus (right)mice left ventricular tissueMfmyofibrilsMtmitochondria long arrow Z diskOriginalmagnification 10000x scale bar = 1 120583m
the cell nucleus through TUNEL assay whereas very fewTUNEL positively cells were in H+Cminus mouse heart sections(Figure 7(a)) In parallel a few cells in theHSP60mouse heartsection were positively stained by anti-activated caspase-3antibody significantly more than that of H+Cminus mouse heart(Figure 7(b)) The data clearly demonstrated that HSP60overexpression results in caspase-3 activation and apoptosisofmyocardial cells inHSP60mice Because theHSP60mouseheart became a sponge-like muscular tissue infiltrated bysmall capillaries and HSP60 involvement in rejection andautoimmune diseases was suggested previously we studiedthe presence of activated inflammatory cells in HSP60 miceThe pathology neither showed infiltrated mononuclear cells(Figure 4) nor detected CD4 or CD8 positive cells in HSP60mouse heart (Figure S3) thus inflammatory response orT cellactivation is absent in the HSP60 mouse heart
Accumulation of HSP60 in the cytoplasm during apop-tosis induction and the binding of HSP60 to procaspase-3and modulating caspase activity has been demonstrated inmany cancer cell lines [21 25 39] Similar to most proteins of
themitochondrialmatrix HSP60 which is synthesized in thecytoplasm with N-terminal targeting peptides is transportedto the mitochondrial matrix where the targeting sequenceis cleaved Both unprocessed (cytosolic) and matured (mito-chondrial) HSP60 strongly increased in the cardiac tissueof HSP60 mouse (Figure 1(b)) Consequently accumulationof cytosolic HSP60 capable of inducing apoptosis cannotbe overlooked and our data provided in vivo evidence ofapoptosis and caspase-3 activation in the HSP60mouse heart(Figure 7) Apoptosis is a highly regulated process duringembryonic and postnatal heart development and critical forheart development Increasing apoptosis in HSP60 mouseheart and its causal relation to cardiac myopathy must beinterpreted with caution because (1) apoptosis frequentlyoccurs in embryonic and neonatal tissues than in adultcardiac tissues (2) increasing apoptosis is often observedin cardiac tissues of myopathy and congestive heart failurethus apoptosis can be secondarily induced in myopathy andheart failure and (3) it can directly result from cytosolicHSP60 accumulation or HSP60 binding to caspases Thus
8 BioMed Research International
TdT TdTDAPI
H+Cminus
H+C+
(a)Casp 3DAPI
H+Cminus
H+C+
Casp 3
(b)
Figure 7 TUNEL staining and activated caspase-3 detection in neonatal heart sections (a) Detection of nuclear DNA fragmentation withTUNEL reaction in the left ventricle sections of neonatal H+C+ and H+Cminus mice Fluorescein-12-dUTP (TdT) and DAPI were used forTUNEL and nuclear staining (b) Immunofluorescence detection of activated (cleaved) caspase-3 induction in H+C+ and in H+Cminus heartsections Scale bar = 50120583m
additional studies are required in this regard Our currentmodel which uses the ubiquitous Cre mouse increasedHSP60 and potentially induced cell apoptosis of multipletissues thus it is unable to clarify phenotypes contributedby the HSP60 induction in a specific organ for examplevascular ECs smooth muscle cells or cardiac myocytesThis limitation can be overcome by combinatory uses of
G-Lox-HSP60 with vascular- or cardiac-specific Cre miceWe have explored HSP60 expression in cardiac tissues ofthe adult mice by using inducible cardiac cell-restricted Cremouse (unpublished results)
In addition to the roles of mitochondrial and cytosolicchaperonins and caspase induction additional functions ofHSP60 have been reported Cytosolic HSP60 regulates the
BioMed Research International 9
phosphorylation of IKK120572120573 and promotes NF-120581B prosur-vival pathway Transgenic expression of the cytosolic HSP60prevented stress-induced hepatic cell deaths in vivo [32]Overexpressing HSP60 in the human stem cells increasedHSP60 nuclear localization which suppressed ROS andp38JNK signaling and regulated genes relating to stem cellproliferation differentiation and stemness [40]Whether theaforementioned mechanisms are also involved in mediatingneonatal deaths remains unclear and future clarification isawaited
In this study congenital ASD was observed in a largenumber ofHSP60mice AlthoughASD exacerbated the exist-ing myopathy of HSP60 mice and was partially responsiblefor neonatal death ASD per sewas unlikely directly caused byHSP60 induction We speculate that ASD in neonatal HSP60mice may be the consequence of incomplete embryonicdevelopment existing myopathy and increasing apoptosisSimilar phenomenon has been demonstrated in the eNOSknockout mouse model in which high incidence of ASDor VSD was precipitated through increased apoptosis duringembryonic and postnatal stages [41] Our main findingsindicate that severe hemorrhage andmyocyte death occurredin mice with HSP60 expression and increased apoptosisin a macerated and spongy-like heart failed to meet thecardiovascular demands after birth and was the primarycause of HSP60-induced heart failure and neonatal death
4 Conclusions
HSP60 not only is the essentialmitochondrial chaperone pro-tein but also plays critical roles outside the mitochondrionAlthough numerous studies indicated the wide involvementof HSP60 in broad biological processes and many diseasesreports of transgenic mouse models of increasing or knock-down HSP60 expression are rare Thus the consequences ofaltering HSP60 levels in vivo in these disease settings remainunknown We used a floxed vector to establish conditionalHSP60 transgenic mice which allows studies on inducibleand organ-targeted HSP60 expression in adult mice and ispotentially useful in pursuing essential questions related toHSP60 in vivo In this study we show that ubiquitous HSP60expression from the embryonic stage results in increasedapoptosis myopathy high incidence of ASD and neonataldeaths
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Tsung-Hsien Chen and Shan-Wen Liu contribute equally tothis paper
Acknowledgments
The authors gratefully acknowledge the technical assistanceprovided by Tzu-Chin Wu and Mei-Yu Fan in NHRI-IBENand Functional and Micro-Magnetic Resonance ImagingCenter supported by NRPGM (NSC97-3112-B-001-013) Thisresearch was supported by grants from NHRI (BN-104-PP-02 BN-104-PP-24) and from Ministry of Science andTechnology (103-2320-B-400-018)
References
[1] I J Benjamin andD RMcMillan ldquoStress (heat shock) proteinsmolecular chaperones in cardiovascular biology and diseaserdquoCirculation Research vol 83 no 2 pp 117ndash132 1998
[2] F U Hartl ldquoMolecular chaperones in cellular protein foldingrdquoNature vol 381 no 6583 pp 571ndash580 1996
[3] J C Ranford and B Henderson ldquoChaperonins in diseasemechanismsmodels and treatmentsrdquoMolecular Pathology vol55 no 4 pp 209ndash213 2002
[4] CCDeocaris S CKaul andRWadhwa ldquoOn the brotherhoodof the mitochondrial chaperones mortalin and heat shockprotein 60rdquo Cell Stress and Chaperones vol 11 no 2 pp 116ndash128 2006
[5] F Cappello A M Gammazza A P Piccionello et al ldquoHsp60chaperonopathies and chaperonotherapy targets and agentsrdquoExpert Opinion on Therapeutic Targets vol 18 no 2 pp 185ndash208 2014
[6] S Lau N Patnaik M R Sayen and R Mestril ldquoSimultaneousoverexpression of two stress proteins in rat cardiomyocytes andmyogenic cells confers protection against ischemia-inducedinjuryrdquo Circulation vol 96 no 7 pp 2287ndash2294 1997
[7] K M Lin B Lin I Y Lian R Mestril I E Scheffler and WH Dillmann ldquoCombined and individual mitochondrial HSP60and HSP10 expression in cardiac myocytes protects mitochon-drial function and prevents apoptotic cell deaths induced bysimulated ischemia-reoxygenationrdquo Circulation vol 103 no 13pp 1787ndash1792 2001
[8] J J Malago J F J G Koninkx and J E van Dijk ldquoThe heatshock response and cytoprotection of the intestinal epitheliumrdquoCell Stress and Chaperones vol 7 no 2 pp 191ndash199 2002
[9] Y-X Shan T-L Yang R Mestril and P H Wang ldquoHsp10 andHsp60 suppress ubiquitination of insulin-like growth factor-1receptor and augment insulin-like growth factor-1 receptor sig-naling in cardiac muscle implications on decreased myocardialprotection in diabetic cardiomyopathyrdquoThe Journal of BiologicalChemistry vol 278 no 46 pp 45492ndash45498 2003
[10] H-S Chen Y-X Shan T-L Yang et al ldquoInsulin deficiencydownregulated heat shock protein 60 and IGF-1 receptorsignaling in diabetic myocardiumrdquo Diabetes vol 54 no 1 pp175ndash181 2005
10 BioMed Research International
[11] V Calabrese A Ravagna C Colombrita et al ldquoAcetylcarnitineinduces heme oxygenase in rat astrocytes and protects againstoxidative stress Involvement of the transcription factor Nrf2rdquoJournal of Neuroscience Research vol 79 no 4 pp 509ndash5212005
[12] V Veereshwarayya P Kumar K M Rosen R Mestril andH W Querfurth ldquoDifferential effects of mitochondrial heatshock protein 60 and related molecular chaperones to preventintracellular 120573-amyloid-induced inhibition of complex IV andlimit apoptosisrdquoThe Journal of Biological Chemistry vol 281 no40 pp 29468ndash29478 2006
[13] C Xu Y Lu Z Pan et al ldquoThe muscle-specific microRNAsmiR-1 and miR-133 produce opposing effects on apoptosis bytargeting HSP60 HSP70 and caspase-9 in cardiomyocytesrdquoJournal of Cell Science vol 120 part 17 pp 3045ndash3052 2007
[14] J C Ghosh T Dohi H K Byoung and D C Altieri ldquoHsp60regulation of tumor cell apoptosisrdquo The Journal of BiologicalChemistry vol 283 no 8 pp 5188ndash5194 2008
[15] P E Castle R Ashfaq F Ansari and C Y Muller ldquoImmuno-histochemical evaluation of heat shock proteins in normal andpreinvasive lesions of the cervixrdquo Cancer Letters vol 229 no 2pp 245ndash252 2005
[16] P A Cornford A R Dodson K F Parsons et al ldquoHeat shockprotein expression independently predicts clinical outcome inprostate cancerrdquoCancer Research vol 60 no 24 pp 7099ndash71052000
[17] F Cappello S David F Rappa et al ldquoThe expression of Hsp60and Hsp10 in large bowel carcinomas with lymph node metas-tasesrdquo BMC Cancer vol 5 article 39 2005
[18] P-L Hsu and S-M Hsu ldquoAbundance of heat shock proteins(hsp89 hsp60 and hsp27) in malignant cells of Hodgkinrsquosdiseaserdquo Cancer Research vol 58 no 23 pp 5507ndash5513 1998
[19] S Gupta and A A Knowlton ldquoHSP60 Bax apoptosis and theheartrdquo Journal of Cellular and Molecular Medicine vol 9 no 1pp 51ndash58 2005
[20] S Xanthoudakis R Sophie D Rasper et al ldquoHsp60 accel-erates the maturation of pro-caspase-3 by upstream activatorproteases during apoptosisrdquo The EMBO Journal vol 18 no 8pp 2049ndash2056 1999
[21] A Samali J Cai B Zhivotovsky D P Jones and S OrreniusldquoPresence of a pre-apoptotic complex of pro-caspase-3 Hsp60and Hsp10 in the mitochondrial fraction of Jurkat cellsrdquo TheEMBO Journal vol 18 no 8 pp 2040ndash2048 1999
[22] A Faried M Sohda M Nakajima TMiyazaki H Kato andHKuwano ldquoExpression of heat-shock protein Hsp60 correlatedwith the apoptotic index and patient prognosis in humanoesophageal squamous cell carcinomardquo European Journal ofCancer vol 40 no 18 pp 2804ndash2811 2004
[23] J Schneider E Jimenez K Marenbach H Romero D Marxand H Meden ldquoImmunohistochemical detection of HSP60-expression in human ovarian cancer Correlation with survivalin a series of 247 patientsrdquoAnticancer Research vol 19 no 3 pp2141ndash2146 1999
[24] T Lebret R W G Watson V Molinie et al ldquoHeat shockproteins HSP27 HSP60 HSP70 and HSP90 expression inbladder carcinomardquo Cancer vol 98 no 5 pp 970ndash977 2003
[25] A Samali and S Orrenius ldquoHeat shock proteins regulators ofstress response and apoptosisrdquo Cell Stress and Chaperones vol3 no 4 pp 228ndash236 1998
[26] W Van Eden G Wick S Albani and I Cohen ldquoStress heatshock proteins and autoimmunity how immune responses to
heat shock proteins are to be used for the control of chronicinflammatory diseasesrdquo Annals of the New York Academy ofSciences vol 1113 pp 217ndash237 2007
[27] C Habich and V Burkart ldquoHeat shock protein 60 regulatoryrole on innate immune cellsrdquo Cellular and Molecular LifeSciences vol 64 no 6 pp 742ndash751 2007
[28] Y Kondo Y Ueno K Kobayashi et al ldquoHepatitis B virus repli-cation could enhance regulatory T cell activity by producingsoluble heat shock protein 60 from hepatocytesrdquoThe Journal ofInfectious Diseases vol 202 no 2 pp 202ndash213 2010
[29] C Colotti G Cavallini R L Vitale et al ldquoEffects of agingand anti-aging caloric restrictions on carbonyl and heat shockprotein levels and expressionrdquo Biogerontology vol 6 no 6 pp397ndash406 2005
[30] P L Hooper ldquoInsulin signaling GSK-3 heat shock proteins andthe natural history of type 2 diabetes mellitus a hypothesisrdquoMetabolic Syndrome and RelatedDisorders vol 5 no 3 pp 220ndash230 2007
[31] O S Birk D C Douek D Elias et al ldquoA role of Hsp60 inautoimmune diabetes analysis in a transgenic modelrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 93 no 3 pp 1032ndash1037 1996
[32] J N Chun B Choi KW Lee et al ldquoCytosolicHsp60 is involvedin the NF-kappaB-dependent survival of cancer cells via IKKregulatiordquo PLoS ONE vol 5 no 3 Article ID e9422 2010
[33] M-R Chen S-W Liu T-C Wu et al ldquoRU486-induciblerecombination in the salivary glands of lactoferrin promoter-driven green fluorescent Cre transgenic micerdquo Genesis vol 48no 10 pp 585ndash595 2010
[34] H-Y Lin C-H Kao KM-C Lin V Kaartinen and L-T YangldquoNotch signaling regulates late-stage epidermal differentiationandmaintains postnatal hair cycle homeostasisrdquoPLoSONE vol6 no 1 Article ID e15842 2011
[35] A C Kayani G L Close W H Dillmann R Mestril M JJackson and A McArdle ldquoOverexpression of HSP10 in skeletalmuscle of transgenic mice prevents the age-related fall inmaximum tetanic force generation and muscle Cross-SectionalAreardquoThe American Journal of PhysiologymdashRegulatory Integra-tive and Comparative Physiology vol 299 no 1 pp R268ndashR2762010
[36] G-C Fan X Ren J Qian et al ldquoNovel cardioprotective role of asmall heat-shock protein Hsp20 against ischemiareperfusioninjuryrdquo Circulation vol 111 no 14 pp 1792ndash1799 2005
[37] J M Hollander J L Martin D D Belke et al ldquoOverex-pression of wild-type heat shock protein 27 and a nonphos-phorylatable heat shock protein 27 mutant protects againstischemiareperfusion injury in a transgenic mouse modelrdquoCirculation vol 110 no 23 pp 3544ndash3552 2004
[38] J-C L Plumier A M Krueger R W Currie D KontoyiannisG Kollias and G N Pagoulatos ldquoTransgenic mice expressingthe human inducible Hsp70 have hippocampal neurons resis-tant to ischemic injuryrdquo Cell Stress and Chaperones vol 2 no 3pp 162ndash167 1997
[39] D Chandra G Choy and D G Tang ldquoCytosolic accumulationof HSP60 during apoptosis with or without apparentmitochon-drial release Evidence that its pro-apoptotic or pro-survivalfunctions involve differential interactions with caspase-3rdquo TheJournal of Biological Chemistry vol 282 no 43 pp 31289ndash313012007
BioMed Research International 11
[40] J H Jang J S Jung J I Choi and S K Kang ldquoNuclearAgo2HSP60 contributes to broad spectrum of hATSCs func-tion via Oct4 regulationrdquoAntioxidants and Redox Signaling vol16 no 5 pp 383ndash399 2012
[41] Q Feng W Song X Lu et al ldquoDevelopment of heart failureand congenital septal defects in mice lacking endothelial nitricoxide synthaserdquo Circulation vol 106 no 7 pp 873ndash879 2002
25 TUNEL Assay Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay wasperformed for detecting nuclear DNA fragmentation inparaffin-embedded tissue sections as a measure of apoptosisFluorescein-12-dUTP was used as the substrate in TUNELreaction (DeadEnd Fluorometric TUNEL System Promega)the slides were counterstained by DAPI and the images weretaken by the Olympus microscope system
26 T2-Weighted Thoracic Region MR Images Magnetic res-onance imaging (MRI) was performed and images wereacquired using 94 Tesla MRI system (Bruker EttlingenGermany) with a maximum gradient strength of 140G cmminus1A quadrature volume coil (internal diameter = 10mm) wasused for RF transmission and reception We used a rapidacquisition with relaxation enhancement sequence to obtainthe T2-weighted images The repetition and echo times wereset at 2500 and 34 milliseconds respectively The field ofview was 10 times 10mm2 and the matrix size was 100 times 100 Theimages were sliced without gaps and the slice thickness was02mmThe voxel size for acquisitionwas 01times 01times 02mm3Finally the images were interpolated to 400times 400 using cubicinterpolation
27 Electron Micrograph Neonatal heart tissues were col-lected and fixed in 4 cold glutaraldehyde postfixed using1 osmium oxide and progressively dehydrated using alco-hol After resin embedding and polymerization 1 120583m thicksections were cut for initial observation Ultrathin (60ndash80 nmthick) sections were cut from the Epon-embedded blocksstained with uranyl acetate and lead citrate and examinedusing transmission electron microscopy (TEM Hitachi H-7500 Tokyo Japan) at 75 kV
3 Results and Discussion
31 Generation of HSP60 Conditional Transgenic MiceUnlike Tg mice expressing HSP10 [35] HSP20 [36] HSP27[37] or HSP70 [38] founders for establishing the ubiquitousHSP60 Tg mouse were not viable In this study we generateda conditional Tg mouse model using Cre-LoxP tools toallow inducible and tissue-specificHSP60 expression G-Lox-HSP60 mouse was generated by using the Tg vector illus-trated in Figure 1(a) CAGGSwas used to drive the expressionof transgenes enhanced green fluorescent protein (EGFP)or human HSP60 full-length cDNA Multiple stop codonsand the SV40 polyA sequences between the 2 LoxP sites ofthe Tg vector prevent the expression of downstream humanHSP60 sequence The G-Lox-HSP60 mice were normal indevelopment weight and reproduction with EGFP expres-sion in most organs During Cre-mediated recombination2 LoxP sites of the G-Lox-HSP60 vector are joined and theHSP60 expression is directly enhanced by CAGGS In thepresent study we generated an FVB ubiquitous Cre Tgmouseexpressing the EGFP-Cre fusion protein [33] We used thetail DNA-PCR method for studying the genotypes of the lit-termates in the crossing studies of G-Lox-HSP60 and EGFP-Cre mice the results are described in Figure 1(b) Deletion
Table 1 Summary of crossings between G-Lox-HSP60 (H) andEGFP-Cre (C) mice
Genotype H+C+ H+Cminus HminusC+ HminusCminus TotalNewborn number 72 76 69 86 303Neonatal death number 72 5 2 2 81Ratio 7272 576 269 286 81303H+C+ double Tg mouse H+Cminus HminusC+ single Tg mouse carrying eitherallele HminusCminus wild-type mouse
of the LoxP-flanked cassette was detected by the presence ofthe shorter PCR fragment in the double Tg mice Littermateswith all 4 possible genotypes were acquired in accordancewith the Mendelian frequency indicating no prenatal loss ofthe double Tgmice (H+C+) which were positive for both G-Lox-HSP60 and EGFP-Cre alleles (Table 1) Strong inductionof human HSP60 expression in the cardiac tissues of thedouble Tg mice was confirmed through RT-PCR Westernblotting (Figure 1(b)) and IHC staining (Figure 2)
32 HSP60 Expression Led to Neonatal Deaths In more than20 crossing experiments using G-Lox-HSP60 and EGFP-Cremice all double TgHSP60mice litters diedwithin a fewhoursafter birth A few survived the first day but none survivedmore than a few days (7272 Table 1) Neonatal deaths wererare in single Tg micemdashH+Cminus (576) and HminusC+ (269)mdashwhich carried only G-Lox-HSP60 or EGFP-Cre and in thewild-type mice (HminusCminus 286) At birth neonatal HSP60mice were slightly smaller in size than average with signsof mild cyanosis and internal bleeding at the abdominalsegments before death (Figure 3) Severe cardiovascular orpulmonary abnormality was suspected to contribute to theneonatal deaths in the HSP60 mice H+Cminus mice exhibitedstrong whole-body green fluorescence whereas the HSP60mice displayed green fluorescent spots indicating remainingEGFP that was produced before recombination HminusC+ andHminusCminus mice did not exhibit fluorescence Thoracic chamberdissection of HSP60 mice revealed a smaller heart and thecolor of the ventricles was more reddish containing moreblood than the hearts of other genotypes (Figure 3)
Postmortem analysis of the HSP60 mice was performedthrough MRI and histology MRI revealed that the HSP60P1 mouse heart was smaller than that of H+Cminus mouse(Figure S1 in Supplementary Material available online athttpdxdoiorg1011552015539805) The HSP60 mouseheart chambers and lungs were filled with liquids that wereabsent in those of H+CminusmouseThe interventricular septumand chamber walls of the HSP60 mice were thinner thannormal (Figure S1) Histological examination revealed mas-sive hemorrhage at the heart and lungs of double Tg mouseThe cardiac muscle appeared degenerated or incompletelydifferentiated with sponge-like cardiac tissues fenestratedwith blood or blood vessels and signs of myocardiumnecrosis were observed (Figure 4) The lungs of the HSP60mice appeared to have developed normally except for thepresence of hemorrhage that potentially resulted in onlypartial inflation of the lungs Other organs such as thebrain kidneys and liver of the HSP60 mice appeared grossly
BioMed Research International 5
HampE HSP60
H+Cminus
HminusCminus
H+C+
Figure 2 HSP60 detection in neonatal H+C+ mice heart HampE and immunohistochemical staining of HSP60 induction in H+C+ heartsections but not in H+Cminus or HminusCminus heart sections Scale bar = 50 120583m
normal (data not shown) Spotty hemorrhage was observedin the skeletal muscles of the neck spine and between theribs however the hemorrhage in these organs was not assevere as that in the heart and lungs Because cardiovascularand respiratory stresses were observed and because the heartswere smaller than normal we speculate that congenitaldefects may contribute to the neonatal deaths of the HSP60mice Of the 10 HSP60 mice studied for septal defects 4had atrial septal defect (ASD) and one had ventricular septaldefect (VSD) as illustrated in Figure 5 Septum defects werenot observed in the control mice
Because the atrial septum is closed when the lungsare inflated during birthing and because ASD manifests asa serious problem only after birth we examined whetherthe HSP60 mice had developmental defects during theembryonic stage in the E17 embryos No apparent abnor-malities were visible externally in the HSP60 mouse E17embryos However the embryo sections revealed excessivehemorrhage vascularization and tissue necrosis in the heartsimilar to those in the postnatal heart Lungs of E17 ofHSP60 mouse appeared grossly normal (Figure S2) Thushistological evidence suggested that congenital heart diseasecaused by cardiomyopathy or incomplete heart development
in HSP60 mouse embryos results in ASD or VSD whichaggravates respiratory stress and congestive heart failure atpostnatal day 1
We searched for ultrastructural abnormality as featuresof cardiac myopathy in neonatal HSP60 mouse using elec-tron micrographs G-Lox-HSP60 mouse contained orga-nized arrays of myofibril filaments with intact mitochon-drion appearing in high contrast with dense laminaecristae throughout the mitochondrial cross-sectional areas(Figure 6) In contrast the myofibrils of HSP60 mouse heartexhibited pronounced fragmentation and disorganizationshown as ragged Z lines with shorter sarcomere lengthon average compared with G-Lox-HSP60 myofibrils Mito-chondrion between myofibrils was mostly broken losingboth outline and inner cristae indicating a disintegratedmitochondrial structureThus TEM results confirmed severecardiac myopathy in HSP60 mouse
33 Increasing Apoptosis in HSP60 Mouse Heart HSP60 waspreviously linked to apoptosis induction thus we studiedapoptotic cardiac myocytes in the HSP60 mouse by usingthe TUNEL assay Many cells in the HSP60 mouse heartsections were positively labeled by fluorescein-dUTP at
6 BioMed Research International
H+Cminus
HminusC+
HminusCminus
H+C+
Figure 3 Neonatal Tg mice Left neonatal Tg miceThe white arrow indicates cyanosis and abdominal bleeding in H+C+ neonates middlefluorescent images of the same mice right the lungs and heart of neonatal mice H+C+ hearts appear more dark reddish and smaller thanhearts of other genotypes Scale bar = 3mm
H+CminusH+C+
Figure 4 Histology of the neonatal heart and lung Top HampE staining of the neonatal heart of H+C+ mouse compared with that of H+Cminusmouse showing hemorrhage necrosis and degenerated cardiac muscles in H+C+ mouse heart Bottom the lung of H+C+ mouse showssigns of hemorrhage liquid congestion and tissue necrosis The alveolar lung structures appeared to have developed normally but poorlyinflated Scale bar = 100 120583m
BioMed Research International 7
H+C+ HminusCminus
Figure 5 Atrial septal defect in H+C+ neonatal mice HampE images of the neonatal heart from H+C+ and HminusCminus mice Asterisk indicatesthe atrial septal defect
H+C+ H+CminusMt
Mt
Mt
Mf
Mt
Mf
Figure 6 Transmission electron microscopy showing ultrastructure of myofibril defect in H+C+ neonatal heart Transmission electronmicroscopy results ofH+C+ (left) andH+Cminus (right)mice left ventricular tissueMfmyofibrilsMtmitochondria long arrow Z diskOriginalmagnification 10000x scale bar = 1 120583m
the cell nucleus through TUNEL assay whereas very fewTUNEL positively cells were in H+Cminus mouse heart sections(Figure 7(a)) In parallel a few cells in theHSP60mouse heartsection were positively stained by anti-activated caspase-3antibody significantly more than that of H+Cminus mouse heart(Figure 7(b)) The data clearly demonstrated that HSP60overexpression results in caspase-3 activation and apoptosisofmyocardial cells inHSP60mice Because theHSP60mouseheart became a sponge-like muscular tissue infiltrated bysmall capillaries and HSP60 involvement in rejection andautoimmune diseases was suggested previously we studiedthe presence of activated inflammatory cells in HSP60 miceThe pathology neither showed infiltrated mononuclear cells(Figure 4) nor detected CD4 or CD8 positive cells in HSP60mouse heart (Figure S3) thus inflammatory response orT cellactivation is absent in the HSP60 mouse heart
Accumulation of HSP60 in the cytoplasm during apop-tosis induction and the binding of HSP60 to procaspase-3and modulating caspase activity has been demonstrated inmany cancer cell lines [21 25 39] Similar to most proteins of
themitochondrialmatrix HSP60 which is synthesized in thecytoplasm with N-terminal targeting peptides is transportedto the mitochondrial matrix where the targeting sequenceis cleaved Both unprocessed (cytosolic) and matured (mito-chondrial) HSP60 strongly increased in the cardiac tissueof HSP60 mouse (Figure 1(b)) Consequently accumulationof cytosolic HSP60 capable of inducing apoptosis cannotbe overlooked and our data provided in vivo evidence ofapoptosis and caspase-3 activation in the HSP60mouse heart(Figure 7) Apoptosis is a highly regulated process duringembryonic and postnatal heart development and critical forheart development Increasing apoptosis in HSP60 mouseheart and its causal relation to cardiac myopathy must beinterpreted with caution because (1) apoptosis frequentlyoccurs in embryonic and neonatal tissues than in adultcardiac tissues (2) increasing apoptosis is often observedin cardiac tissues of myopathy and congestive heart failurethus apoptosis can be secondarily induced in myopathy andheart failure and (3) it can directly result from cytosolicHSP60 accumulation or HSP60 binding to caspases Thus
8 BioMed Research International
TdT TdTDAPI
H+Cminus
H+C+
(a)Casp 3DAPI
H+Cminus
H+C+
Casp 3
(b)
Figure 7 TUNEL staining and activated caspase-3 detection in neonatal heart sections (a) Detection of nuclear DNA fragmentation withTUNEL reaction in the left ventricle sections of neonatal H+C+ and H+Cminus mice Fluorescein-12-dUTP (TdT) and DAPI were used forTUNEL and nuclear staining (b) Immunofluorescence detection of activated (cleaved) caspase-3 induction in H+C+ and in H+Cminus heartsections Scale bar = 50120583m
additional studies are required in this regard Our currentmodel which uses the ubiquitous Cre mouse increasedHSP60 and potentially induced cell apoptosis of multipletissues thus it is unable to clarify phenotypes contributedby the HSP60 induction in a specific organ for examplevascular ECs smooth muscle cells or cardiac myocytesThis limitation can be overcome by combinatory uses of
G-Lox-HSP60 with vascular- or cardiac-specific Cre miceWe have explored HSP60 expression in cardiac tissues ofthe adult mice by using inducible cardiac cell-restricted Cremouse (unpublished results)
In addition to the roles of mitochondrial and cytosolicchaperonins and caspase induction additional functions ofHSP60 have been reported Cytosolic HSP60 regulates the
BioMed Research International 9
phosphorylation of IKK120572120573 and promotes NF-120581B prosur-vival pathway Transgenic expression of the cytosolic HSP60prevented stress-induced hepatic cell deaths in vivo [32]Overexpressing HSP60 in the human stem cells increasedHSP60 nuclear localization which suppressed ROS andp38JNK signaling and regulated genes relating to stem cellproliferation differentiation and stemness [40]Whether theaforementioned mechanisms are also involved in mediatingneonatal deaths remains unclear and future clarification isawaited
In this study congenital ASD was observed in a largenumber ofHSP60mice AlthoughASD exacerbated the exist-ing myopathy of HSP60 mice and was partially responsiblefor neonatal death ASD per sewas unlikely directly caused byHSP60 induction We speculate that ASD in neonatal HSP60mice may be the consequence of incomplete embryonicdevelopment existing myopathy and increasing apoptosisSimilar phenomenon has been demonstrated in the eNOSknockout mouse model in which high incidence of ASDor VSD was precipitated through increased apoptosis duringembryonic and postnatal stages [41] Our main findingsindicate that severe hemorrhage andmyocyte death occurredin mice with HSP60 expression and increased apoptosisin a macerated and spongy-like heart failed to meet thecardiovascular demands after birth and was the primarycause of HSP60-induced heart failure and neonatal death
4 Conclusions
HSP60 not only is the essentialmitochondrial chaperone pro-tein but also plays critical roles outside the mitochondrionAlthough numerous studies indicated the wide involvementof HSP60 in broad biological processes and many diseasesreports of transgenic mouse models of increasing or knock-down HSP60 expression are rare Thus the consequences ofaltering HSP60 levels in vivo in these disease settings remainunknown We used a floxed vector to establish conditionalHSP60 transgenic mice which allows studies on inducibleand organ-targeted HSP60 expression in adult mice and ispotentially useful in pursuing essential questions related toHSP60 in vivo In this study we show that ubiquitous HSP60expression from the embryonic stage results in increasedapoptosis myopathy high incidence of ASD and neonataldeaths
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Tsung-Hsien Chen and Shan-Wen Liu contribute equally tothis paper
Acknowledgments
The authors gratefully acknowledge the technical assistanceprovided by Tzu-Chin Wu and Mei-Yu Fan in NHRI-IBENand Functional and Micro-Magnetic Resonance ImagingCenter supported by NRPGM (NSC97-3112-B-001-013) Thisresearch was supported by grants from NHRI (BN-104-PP-02 BN-104-PP-24) and from Ministry of Science andTechnology (103-2320-B-400-018)
References
[1] I J Benjamin andD RMcMillan ldquoStress (heat shock) proteinsmolecular chaperones in cardiovascular biology and diseaserdquoCirculation Research vol 83 no 2 pp 117ndash132 1998
[2] F U Hartl ldquoMolecular chaperones in cellular protein foldingrdquoNature vol 381 no 6583 pp 571ndash580 1996
[3] J C Ranford and B Henderson ldquoChaperonins in diseasemechanismsmodels and treatmentsrdquoMolecular Pathology vol55 no 4 pp 209ndash213 2002
[4] CCDeocaris S CKaul andRWadhwa ldquoOn the brotherhoodof the mitochondrial chaperones mortalin and heat shockprotein 60rdquo Cell Stress and Chaperones vol 11 no 2 pp 116ndash128 2006
[5] F Cappello A M Gammazza A P Piccionello et al ldquoHsp60chaperonopathies and chaperonotherapy targets and agentsrdquoExpert Opinion on Therapeutic Targets vol 18 no 2 pp 185ndash208 2014
[6] S Lau N Patnaik M R Sayen and R Mestril ldquoSimultaneousoverexpression of two stress proteins in rat cardiomyocytes andmyogenic cells confers protection against ischemia-inducedinjuryrdquo Circulation vol 96 no 7 pp 2287ndash2294 1997
[7] K M Lin B Lin I Y Lian R Mestril I E Scheffler and WH Dillmann ldquoCombined and individual mitochondrial HSP60and HSP10 expression in cardiac myocytes protects mitochon-drial function and prevents apoptotic cell deaths induced bysimulated ischemia-reoxygenationrdquo Circulation vol 103 no 13pp 1787ndash1792 2001
[8] J J Malago J F J G Koninkx and J E van Dijk ldquoThe heatshock response and cytoprotection of the intestinal epitheliumrdquoCell Stress and Chaperones vol 7 no 2 pp 191ndash199 2002
[9] Y-X Shan T-L Yang R Mestril and P H Wang ldquoHsp10 andHsp60 suppress ubiquitination of insulin-like growth factor-1receptor and augment insulin-like growth factor-1 receptor sig-naling in cardiac muscle implications on decreased myocardialprotection in diabetic cardiomyopathyrdquoThe Journal of BiologicalChemistry vol 278 no 46 pp 45492ndash45498 2003
[10] H-S Chen Y-X Shan T-L Yang et al ldquoInsulin deficiencydownregulated heat shock protein 60 and IGF-1 receptorsignaling in diabetic myocardiumrdquo Diabetes vol 54 no 1 pp175ndash181 2005
10 BioMed Research International
[11] V Calabrese A Ravagna C Colombrita et al ldquoAcetylcarnitineinduces heme oxygenase in rat astrocytes and protects againstoxidative stress Involvement of the transcription factor Nrf2rdquoJournal of Neuroscience Research vol 79 no 4 pp 509ndash5212005
[12] V Veereshwarayya P Kumar K M Rosen R Mestril andH W Querfurth ldquoDifferential effects of mitochondrial heatshock protein 60 and related molecular chaperones to preventintracellular 120573-amyloid-induced inhibition of complex IV andlimit apoptosisrdquoThe Journal of Biological Chemistry vol 281 no40 pp 29468ndash29478 2006
[13] C Xu Y Lu Z Pan et al ldquoThe muscle-specific microRNAsmiR-1 and miR-133 produce opposing effects on apoptosis bytargeting HSP60 HSP70 and caspase-9 in cardiomyocytesrdquoJournal of Cell Science vol 120 part 17 pp 3045ndash3052 2007
[14] J C Ghosh T Dohi H K Byoung and D C Altieri ldquoHsp60regulation of tumor cell apoptosisrdquo The Journal of BiologicalChemistry vol 283 no 8 pp 5188ndash5194 2008
[15] P E Castle R Ashfaq F Ansari and C Y Muller ldquoImmuno-histochemical evaluation of heat shock proteins in normal andpreinvasive lesions of the cervixrdquo Cancer Letters vol 229 no 2pp 245ndash252 2005
[16] P A Cornford A R Dodson K F Parsons et al ldquoHeat shockprotein expression independently predicts clinical outcome inprostate cancerrdquoCancer Research vol 60 no 24 pp 7099ndash71052000
[17] F Cappello S David F Rappa et al ldquoThe expression of Hsp60and Hsp10 in large bowel carcinomas with lymph node metas-tasesrdquo BMC Cancer vol 5 article 39 2005
[18] P-L Hsu and S-M Hsu ldquoAbundance of heat shock proteins(hsp89 hsp60 and hsp27) in malignant cells of Hodgkinrsquosdiseaserdquo Cancer Research vol 58 no 23 pp 5507ndash5513 1998
[19] S Gupta and A A Knowlton ldquoHSP60 Bax apoptosis and theheartrdquo Journal of Cellular and Molecular Medicine vol 9 no 1pp 51ndash58 2005
[20] S Xanthoudakis R Sophie D Rasper et al ldquoHsp60 accel-erates the maturation of pro-caspase-3 by upstream activatorproteases during apoptosisrdquo The EMBO Journal vol 18 no 8pp 2049ndash2056 1999
[21] A Samali J Cai B Zhivotovsky D P Jones and S OrreniusldquoPresence of a pre-apoptotic complex of pro-caspase-3 Hsp60and Hsp10 in the mitochondrial fraction of Jurkat cellsrdquo TheEMBO Journal vol 18 no 8 pp 2040ndash2048 1999
[22] A Faried M Sohda M Nakajima TMiyazaki H Kato andHKuwano ldquoExpression of heat-shock protein Hsp60 correlatedwith the apoptotic index and patient prognosis in humanoesophageal squamous cell carcinomardquo European Journal ofCancer vol 40 no 18 pp 2804ndash2811 2004
[23] J Schneider E Jimenez K Marenbach H Romero D Marxand H Meden ldquoImmunohistochemical detection of HSP60-expression in human ovarian cancer Correlation with survivalin a series of 247 patientsrdquoAnticancer Research vol 19 no 3 pp2141ndash2146 1999
[24] T Lebret R W G Watson V Molinie et al ldquoHeat shockproteins HSP27 HSP60 HSP70 and HSP90 expression inbladder carcinomardquo Cancer vol 98 no 5 pp 970ndash977 2003
[25] A Samali and S Orrenius ldquoHeat shock proteins regulators ofstress response and apoptosisrdquo Cell Stress and Chaperones vol3 no 4 pp 228ndash236 1998
[26] W Van Eden G Wick S Albani and I Cohen ldquoStress heatshock proteins and autoimmunity how immune responses to
heat shock proteins are to be used for the control of chronicinflammatory diseasesrdquo Annals of the New York Academy ofSciences vol 1113 pp 217ndash237 2007
[27] C Habich and V Burkart ldquoHeat shock protein 60 regulatoryrole on innate immune cellsrdquo Cellular and Molecular LifeSciences vol 64 no 6 pp 742ndash751 2007
[28] Y Kondo Y Ueno K Kobayashi et al ldquoHepatitis B virus repli-cation could enhance regulatory T cell activity by producingsoluble heat shock protein 60 from hepatocytesrdquoThe Journal ofInfectious Diseases vol 202 no 2 pp 202ndash213 2010
[29] C Colotti G Cavallini R L Vitale et al ldquoEffects of agingand anti-aging caloric restrictions on carbonyl and heat shockprotein levels and expressionrdquo Biogerontology vol 6 no 6 pp397ndash406 2005
[30] P L Hooper ldquoInsulin signaling GSK-3 heat shock proteins andthe natural history of type 2 diabetes mellitus a hypothesisrdquoMetabolic Syndrome and RelatedDisorders vol 5 no 3 pp 220ndash230 2007
[31] O S Birk D C Douek D Elias et al ldquoA role of Hsp60 inautoimmune diabetes analysis in a transgenic modelrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 93 no 3 pp 1032ndash1037 1996
[32] J N Chun B Choi KW Lee et al ldquoCytosolicHsp60 is involvedin the NF-kappaB-dependent survival of cancer cells via IKKregulatiordquo PLoS ONE vol 5 no 3 Article ID e9422 2010
[33] M-R Chen S-W Liu T-C Wu et al ldquoRU486-induciblerecombination in the salivary glands of lactoferrin promoter-driven green fluorescent Cre transgenic micerdquo Genesis vol 48no 10 pp 585ndash595 2010
[34] H-Y Lin C-H Kao KM-C Lin V Kaartinen and L-T YangldquoNotch signaling regulates late-stage epidermal differentiationandmaintains postnatal hair cycle homeostasisrdquoPLoSONE vol6 no 1 Article ID e15842 2011
[35] A C Kayani G L Close W H Dillmann R Mestril M JJackson and A McArdle ldquoOverexpression of HSP10 in skeletalmuscle of transgenic mice prevents the age-related fall inmaximum tetanic force generation and muscle Cross-SectionalAreardquoThe American Journal of PhysiologymdashRegulatory Integra-tive and Comparative Physiology vol 299 no 1 pp R268ndashR2762010
[36] G-C Fan X Ren J Qian et al ldquoNovel cardioprotective role of asmall heat-shock protein Hsp20 against ischemiareperfusioninjuryrdquo Circulation vol 111 no 14 pp 1792ndash1799 2005
[37] J M Hollander J L Martin D D Belke et al ldquoOverex-pression of wild-type heat shock protein 27 and a nonphos-phorylatable heat shock protein 27 mutant protects againstischemiareperfusion injury in a transgenic mouse modelrdquoCirculation vol 110 no 23 pp 3544ndash3552 2004
[38] J-C L Plumier A M Krueger R W Currie D KontoyiannisG Kollias and G N Pagoulatos ldquoTransgenic mice expressingthe human inducible Hsp70 have hippocampal neurons resis-tant to ischemic injuryrdquo Cell Stress and Chaperones vol 2 no 3pp 162ndash167 1997
[39] D Chandra G Choy and D G Tang ldquoCytosolic accumulationof HSP60 during apoptosis with or without apparentmitochon-drial release Evidence that its pro-apoptotic or pro-survivalfunctions involve differential interactions with caspase-3rdquo TheJournal of Biological Chemistry vol 282 no 43 pp 31289ndash313012007
BioMed Research International 11
[40] J H Jang J S Jung J I Choi and S K Kang ldquoNuclearAgo2HSP60 contributes to broad spectrum of hATSCs func-tion via Oct4 regulationrdquoAntioxidants and Redox Signaling vol16 no 5 pp 383ndash399 2012
[41] Q Feng W Song X Lu et al ldquoDevelopment of heart failureand congenital septal defects in mice lacking endothelial nitricoxide synthaserdquo Circulation vol 106 no 7 pp 873ndash879 2002
Figure 2 HSP60 detection in neonatal H+C+ mice heart HampE and immunohistochemical staining of HSP60 induction in H+C+ heartsections but not in H+Cminus or HminusCminus heart sections Scale bar = 50 120583m
normal (data not shown) Spotty hemorrhage was observedin the skeletal muscles of the neck spine and between theribs however the hemorrhage in these organs was not assevere as that in the heart and lungs Because cardiovascularand respiratory stresses were observed and because the heartswere smaller than normal we speculate that congenitaldefects may contribute to the neonatal deaths of the HSP60mice Of the 10 HSP60 mice studied for septal defects 4had atrial septal defect (ASD) and one had ventricular septaldefect (VSD) as illustrated in Figure 5 Septum defects werenot observed in the control mice
Because the atrial septum is closed when the lungsare inflated during birthing and because ASD manifests asa serious problem only after birth we examined whetherthe HSP60 mice had developmental defects during theembryonic stage in the E17 embryos No apparent abnor-malities were visible externally in the HSP60 mouse E17embryos However the embryo sections revealed excessivehemorrhage vascularization and tissue necrosis in the heartsimilar to those in the postnatal heart Lungs of E17 ofHSP60 mouse appeared grossly normal (Figure S2) Thushistological evidence suggested that congenital heart diseasecaused by cardiomyopathy or incomplete heart development
in HSP60 mouse embryos results in ASD or VSD whichaggravates respiratory stress and congestive heart failure atpostnatal day 1
We searched for ultrastructural abnormality as featuresof cardiac myopathy in neonatal HSP60 mouse using elec-tron micrographs G-Lox-HSP60 mouse contained orga-nized arrays of myofibril filaments with intact mitochon-drion appearing in high contrast with dense laminaecristae throughout the mitochondrial cross-sectional areas(Figure 6) In contrast the myofibrils of HSP60 mouse heartexhibited pronounced fragmentation and disorganizationshown as ragged Z lines with shorter sarcomere lengthon average compared with G-Lox-HSP60 myofibrils Mito-chondrion between myofibrils was mostly broken losingboth outline and inner cristae indicating a disintegratedmitochondrial structureThus TEM results confirmed severecardiac myopathy in HSP60 mouse
33 Increasing Apoptosis in HSP60 Mouse Heart HSP60 waspreviously linked to apoptosis induction thus we studiedapoptotic cardiac myocytes in the HSP60 mouse by usingthe TUNEL assay Many cells in the HSP60 mouse heartsections were positively labeled by fluorescein-dUTP at
6 BioMed Research International
H+Cminus
HminusC+
HminusCminus
H+C+
Figure 3 Neonatal Tg mice Left neonatal Tg miceThe white arrow indicates cyanosis and abdominal bleeding in H+C+ neonates middlefluorescent images of the same mice right the lungs and heart of neonatal mice H+C+ hearts appear more dark reddish and smaller thanhearts of other genotypes Scale bar = 3mm
H+CminusH+C+
Figure 4 Histology of the neonatal heart and lung Top HampE staining of the neonatal heart of H+C+ mouse compared with that of H+Cminusmouse showing hemorrhage necrosis and degenerated cardiac muscles in H+C+ mouse heart Bottom the lung of H+C+ mouse showssigns of hemorrhage liquid congestion and tissue necrosis The alveolar lung structures appeared to have developed normally but poorlyinflated Scale bar = 100 120583m
BioMed Research International 7
H+C+ HminusCminus
Figure 5 Atrial septal defect in H+C+ neonatal mice HampE images of the neonatal heart from H+C+ and HminusCminus mice Asterisk indicatesthe atrial septal defect
H+C+ H+CminusMt
Mt
Mt
Mf
Mt
Mf
Figure 6 Transmission electron microscopy showing ultrastructure of myofibril defect in H+C+ neonatal heart Transmission electronmicroscopy results ofH+C+ (left) andH+Cminus (right)mice left ventricular tissueMfmyofibrilsMtmitochondria long arrow Z diskOriginalmagnification 10000x scale bar = 1 120583m
the cell nucleus through TUNEL assay whereas very fewTUNEL positively cells were in H+Cminus mouse heart sections(Figure 7(a)) In parallel a few cells in theHSP60mouse heartsection were positively stained by anti-activated caspase-3antibody significantly more than that of H+Cminus mouse heart(Figure 7(b)) The data clearly demonstrated that HSP60overexpression results in caspase-3 activation and apoptosisofmyocardial cells inHSP60mice Because theHSP60mouseheart became a sponge-like muscular tissue infiltrated bysmall capillaries and HSP60 involvement in rejection andautoimmune diseases was suggested previously we studiedthe presence of activated inflammatory cells in HSP60 miceThe pathology neither showed infiltrated mononuclear cells(Figure 4) nor detected CD4 or CD8 positive cells in HSP60mouse heart (Figure S3) thus inflammatory response orT cellactivation is absent in the HSP60 mouse heart
Accumulation of HSP60 in the cytoplasm during apop-tosis induction and the binding of HSP60 to procaspase-3and modulating caspase activity has been demonstrated inmany cancer cell lines [21 25 39] Similar to most proteins of
themitochondrialmatrix HSP60 which is synthesized in thecytoplasm with N-terminal targeting peptides is transportedto the mitochondrial matrix where the targeting sequenceis cleaved Both unprocessed (cytosolic) and matured (mito-chondrial) HSP60 strongly increased in the cardiac tissueof HSP60 mouse (Figure 1(b)) Consequently accumulationof cytosolic HSP60 capable of inducing apoptosis cannotbe overlooked and our data provided in vivo evidence ofapoptosis and caspase-3 activation in the HSP60mouse heart(Figure 7) Apoptosis is a highly regulated process duringembryonic and postnatal heart development and critical forheart development Increasing apoptosis in HSP60 mouseheart and its causal relation to cardiac myopathy must beinterpreted with caution because (1) apoptosis frequentlyoccurs in embryonic and neonatal tissues than in adultcardiac tissues (2) increasing apoptosis is often observedin cardiac tissues of myopathy and congestive heart failurethus apoptosis can be secondarily induced in myopathy andheart failure and (3) it can directly result from cytosolicHSP60 accumulation or HSP60 binding to caspases Thus
8 BioMed Research International
TdT TdTDAPI
H+Cminus
H+C+
(a)Casp 3DAPI
H+Cminus
H+C+
Casp 3
(b)
Figure 7 TUNEL staining and activated caspase-3 detection in neonatal heart sections (a) Detection of nuclear DNA fragmentation withTUNEL reaction in the left ventricle sections of neonatal H+C+ and H+Cminus mice Fluorescein-12-dUTP (TdT) and DAPI were used forTUNEL and nuclear staining (b) Immunofluorescence detection of activated (cleaved) caspase-3 induction in H+C+ and in H+Cminus heartsections Scale bar = 50120583m
additional studies are required in this regard Our currentmodel which uses the ubiquitous Cre mouse increasedHSP60 and potentially induced cell apoptosis of multipletissues thus it is unable to clarify phenotypes contributedby the HSP60 induction in a specific organ for examplevascular ECs smooth muscle cells or cardiac myocytesThis limitation can be overcome by combinatory uses of
G-Lox-HSP60 with vascular- or cardiac-specific Cre miceWe have explored HSP60 expression in cardiac tissues ofthe adult mice by using inducible cardiac cell-restricted Cremouse (unpublished results)
In addition to the roles of mitochondrial and cytosolicchaperonins and caspase induction additional functions ofHSP60 have been reported Cytosolic HSP60 regulates the
BioMed Research International 9
phosphorylation of IKK120572120573 and promotes NF-120581B prosur-vival pathway Transgenic expression of the cytosolic HSP60prevented stress-induced hepatic cell deaths in vivo [32]Overexpressing HSP60 in the human stem cells increasedHSP60 nuclear localization which suppressed ROS andp38JNK signaling and regulated genes relating to stem cellproliferation differentiation and stemness [40]Whether theaforementioned mechanisms are also involved in mediatingneonatal deaths remains unclear and future clarification isawaited
In this study congenital ASD was observed in a largenumber ofHSP60mice AlthoughASD exacerbated the exist-ing myopathy of HSP60 mice and was partially responsiblefor neonatal death ASD per sewas unlikely directly caused byHSP60 induction We speculate that ASD in neonatal HSP60mice may be the consequence of incomplete embryonicdevelopment existing myopathy and increasing apoptosisSimilar phenomenon has been demonstrated in the eNOSknockout mouse model in which high incidence of ASDor VSD was precipitated through increased apoptosis duringembryonic and postnatal stages [41] Our main findingsindicate that severe hemorrhage andmyocyte death occurredin mice with HSP60 expression and increased apoptosisin a macerated and spongy-like heart failed to meet thecardiovascular demands after birth and was the primarycause of HSP60-induced heart failure and neonatal death
4 Conclusions
HSP60 not only is the essentialmitochondrial chaperone pro-tein but also plays critical roles outside the mitochondrionAlthough numerous studies indicated the wide involvementof HSP60 in broad biological processes and many diseasesreports of transgenic mouse models of increasing or knock-down HSP60 expression are rare Thus the consequences ofaltering HSP60 levels in vivo in these disease settings remainunknown We used a floxed vector to establish conditionalHSP60 transgenic mice which allows studies on inducibleand organ-targeted HSP60 expression in adult mice and ispotentially useful in pursuing essential questions related toHSP60 in vivo In this study we show that ubiquitous HSP60expression from the embryonic stage results in increasedapoptosis myopathy high incidence of ASD and neonataldeaths
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Tsung-Hsien Chen and Shan-Wen Liu contribute equally tothis paper
Acknowledgments
The authors gratefully acknowledge the technical assistanceprovided by Tzu-Chin Wu and Mei-Yu Fan in NHRI-IBENand Functional and Micro-Magnetic Resonance ImagingCenter supported by NRPGM (NSC97-3112-B-001-013) Thisresearch was supported by grants from NHRI (BN-104-PP-02 BN-104-PP-24) and from Ministry of Science andTechnology (103-2320-B-400-018)
References
[1] I J Benjamin andD RMcMillan ldquoStress (heat shock) proteinsmolecular chaperones in cardiovascular biology and diseaserdquoCirculation Research vol 83 no 2 pp 117ndash132 1998
[2] F U Hartl ldquoMolecular chaperones in cellular protein foldingrdquoNature vol 381 no 6583 pp 571ndash580 1996
[3] J C Ranford and B Henderson ldquoChaperonins in diseasemechanismsmodels and treatmentsrdquoMolecular Pathology vol55 no 4 pp 209ndash213 2002
[4] CCDeocaris S CKaul andRWadhwa ldquoOn the brotherhoodof the mitochondrial chaperones mortalin and heat shockprotein 60rdquo Cell Stress and Chaperones vol 11 no 2 pp 116ndash128 2006
[5] F Cappello A M Gammazza A P Piccionello et al ldquoHsp60chaperonopathies and chaperonotherapy targets and agentsrdquoExpert Opinion on Therapeutic Targets vol 18 no 2 pp 185ndash208 2014
[6] S Lau N Patnaik M R Sayen and R Mestril ldquoSimultaneousoverexpression of two stress proteins in rat cardiomyocytes andmyogenic cells confers protection against ischemia-inducedinjuryrdquo Circulation vol 96 no 7 pp 2287ndash2294 1997
[7] K M Lin B Lin I Y Lian R Mestril I E Scheffler and WH Dillmann ldquoCombined and individual mitochondrial HSP60and HSP10 expression in cardiac myocytes protects mitochon-drial function and prevents apoptotic cell deaths induced bysimulated ischemia-reoxygenationrdquo Circulation vol 103 no 13pp 1787ndash1792 2001
[8] J J Malago J F J G Koninkx and J E van Dijk ldquoThe heatshock response and cytoprotection of the intestinal epitheliumrdquoCell Stress and Chaperones vol 7 no 2 pp 191ndash199 2002
[9] Y-X Shan T-L Yang R Mestril and P H Wang ldquoHsp10 andHsp60 suppress ubiquitination of insulin-like growth factor-1receptor and augment insulin-like growth factor-1 receptor sig-naling in cardiac muscle implications on decreased myocardialprotection in diabetic cardiomyopathyrdquoThe Journal of BiologicalChemistry vol 278 no 46 pp 45492ndash45498 2003
[10] H-S Chen Y-X Shan T-L Yang et al ldquoInsulin deficiencydownregulated heat shock protein 60 and IGF-1 receptorsignaling in diabetic myocardiumrdquo Diabetes vol 54 no 1 pp175ndash181 2005
10 BioMed Research International
[11] V Calabrese A Ravagna C Colombrita et al ldquoAcetylcarnitineinduces heme oxygenase in rat astrocytes and protects againstoxidative stress Involvement of the transcription factor Nrf2rdquoJournal of Neuroscience Research vol 79 no 4 pp 509ndash5212005
[12] V Veereshwarayya P Kumar K M Rosen R Mestril andH W Querfurth ldquoDifferential effects of mitochondrial heatshock protein 60 and related molecular chaperones to preventintracellular 120573-amyloid-induced inhibition of complex IV andlimit apoptosisrdquoThe Journal of Biological Chemistry vol 281 no40 pp 29468ndash29478 2006
[13] C Xu Y Lu Z Pan et al ldquoThe muscle-specific microRNAsmiR-1 and miR-133 produce opposing effects on apoptosis bytargeting HSP60 HSP70 and caspase-9 in cardiomyocytesrdquoJournal of Cell Science vol 120 part 17 pp 3045ndash3052 2007
[14] J C Ghosh T Dohi H K Byoung and D C Altieri ldquoHsp60regulation of tumor cell apoptosisrdquo The Journal of BiologicalChemistry vol 283 no 8 pp 5188ndash5194 2008
[15] P E Castle R Ashfaq F Ansari and C Y Muller ldquoImmuno-histochemical evaluation of heat shock proteins in normal andpreinvasive lesions of the cervixrdquo Cancer Letters vol 229 no 2pp 245ndash252 2005
[16] P A Cornford A R Dodson K F Parsons et al ldquoHeat shockprotein expression independently predicts clinical outcome inprostate cancerrdquoCancer Research vol 60 no 24 pp 7099ndash71052000
[17] F Cappello S David F Rappa et al ldquoThe expression of Hsp60and Hsp10 in large bowel carcinomas with lymph node metas-tasesrdquo BMC Cancer vol 5 article 39 2005
[18] P-L Hsu and S-M Hsu ldquoAbundance of heat shock proteins(hsp89 hsp60 and hsp27) in malignant cells of Hodgkinrsquosdiseaserdquo Cancer Research vol 58 no 23 pp 5507ndash5513 1998
[19] S Gupta and A A Knowlton ldquoHSP60 Bax apoptosis and theheartrdquo Journal of Cellular and Molecular Medicine vol 9 no 1pp 51ndash58 2005
[20] S Xanthoudakis R Sophie D Rasper et al ldquoHsp60 accel-erates the maturation of pro-caspase-3 by upstream activatorproteases during apoptosisrdquo The EMBO Journal vol 18 no 8pp 2049ndash2056 1999
[21] A Samali J Cai B Zhivotovsky D P Jones and S OrreniusldquoPresence of a pre-apoptotic complex of pro-caspase-3 Hsp60and Hsp10 in the mitochondrial fraction of Jurkat cellsrdquo TheEMBO Journal vol 18 no 8 pp 2040ndash2048 1999
[22] A Faried M Sohda M Nakajima TMiyazaki H Kato andHKuwano ldquoExpression of heat-shock protein Hsp60 correlatedwith the apoptotic index and patient prognosis in humanoesophageal squamous cell carcinomardquo European Journal ofCancer vol 40 no 18 pp 2804ndash2811 2004
[23] J Schneider E Jimenez K Marenbach H Romero D Marxand H Meden ldquoImmunohistochemical detection of HSP60-expression in human ovarian cancer Correlation with survivalin a series of 247 patientsrdquoAnticancer Research vol 19 no 3 pp2141ndash2146 1999
[24] T Lebret R W G Watson V Molinie et al ldquoHeat shockproteins HSP27 HSP60 HSP70 and HSP90 expression inbladder carcinomardquo Cancer vol 98 no 5 pp 970ndash977 2003
[25] A Samali and S Orrenius ldquoHeat shock proteins regulators ofstress response and apoptosisrdquo Cell Stress and Chaperones vol3 no 4 pp 228ndash236 1998
[26] W Van Eden G Wick S Albani and I Cohen ldquoStress heatshock proteins and autoimmunity how immune responses to
heat shock proteins are to be used for the control of chronicinflammatory diseasesrdquo Annals of the New York Academy ofSciences vol 1113 pp 217ndash237 2007
[27] C Habich and V Burkart ldquoHeat shock protein 60 regulatoryrole on innate immune cellsrdquo Cellular and Molecular LifeSciences vol 64 no 6 pp 742ndash751 2007
[28] Y Kondo Y Ueno K Kobayashi et al ldquoHepatitis B virus repli-cation could enhance regulatory T cell activity by producingsoluble heat shock protein 60 from hepatocytesrdquoThe Journal ofInfectious Diseases vol 202 no 2 pp 202ndash213 2010
[29] C Colotti G Cavallini R L Vitale et al ldquoEffects of agingand anti-aging caloric restrictions on carbonyl and heat shockprotein levels and expressionrdquo Biogerontology vol 6 no 6 pp397ndash406 2005
[30] P L Hooper ldquoInsulin signaling GSK-3 heat shock proteins andthe natural history of type 2 diabetes mellitus a hypothesisrdquoMetabolic Syndrome and RelatedDisorders vol 5 no 3 pp 220ndash230 2007
[31] O S Birk D C Douek D Elias et al ldquoA role of Hsp60 inautoimmune diabetes analysis in a transgenic modelrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 93 no 3 pp 1032ndash1037 1996
[32] J N Chun B Choi KW Lee et al ldquoCytosolicHsp60 is involvedin the NF-kappaB-dependent survival of cancer cells via IKKregulatiordquo PLoS ONE vol 5 no 3 Article ID e9422 2010
[33] M-R Chen S-W Liu T-C Wu et al ldquoRU486-induciblerecombination in the salivary glands of lactoferrin promoter-driven green fluorescent Cre transgenic micerdquo Genesis vol 48no 10 pp 585ndash595 2010
[34] H-Y Lin C-H Kao KM-C Lin V Kaartinen and L-T YangldquoNotch signaling regulates late-stage epidermal differentiationandmaintains postnatal hair cycle homeostasisrdquoPLoSONE vol6 no 1 Article ID e15842 2011
[35] A C Kayani G L Close W H Dillmann R Mestril M JJackson and A McArdle ldquoOverexpression of HSP10 in skeletalmuscle of transgenic mice prevents the age-related fall inmaximum tetanic force generation and muscle Cross-SectionalAreardquoThe American Journal of PhysiologymdashRegulatory Integra-tive and Comparative Physiology vol 299 no 1 pp R268ndashR2762010
[36] G-C Fan X Ren J Qian et al ldquoNovel cardioprotective role of asmall heat-shock protein Hsp20 against ischemiareperfusioninjuryrdquo Circulation vol 111 no 14 pp 1792ndash1799 2005
[37] J M Hollander J L Martin D D Belke et al ldquoOverex-pression of wild-type heat shock protein 27 and a nonphos-phorylatable heat shock protein 27 mutant protects againstischemiareperfusion injury in a transgenic mouse modelrdquoCirculation vol 110 no 23 pp 3544ndash3552 2004
[38] J-C L Plumier A M Krueger R W Currie D KontoyiannisG Kollias and G N Pagoulatos ldquoTransgenic mice expressingthe human inducible Hsp70 have hippocampal neurons resis-tant to ischemic injuryrdquo Cell Stress and Chaperones vol 2 no 3pp 162ndash167 1997
[39] D Chandra G Choy and D G Tang ldquoCytosolic accumulationof HSP60 during apoptosis with or without apparentmitochon-drial release Evidence that its pro-apoptotic or pro-survivalfunctions involve differential interactions with caspase-3rdquo TheJournal of Biological Chemistry vol 282 no 43 pp 31289ndash313012007
BioMed Research International 11
[40] J H Jang J S Jung J I Choi and S K Kang ldquoNuclearAgo2HSP60 contributes to broad spectrum of hATSCs func-tion via Oct4 regulationrdquoAntioxidants and Redox Signaling vol16 no 5 pp 383ndash399 2012
[41] Q Feng W Song X Lu et al ldquoDevelopment of heart failureand congenital septal defects in mice lacking endothelial nitricoxide synthaserdquo Circulation vol 106 no 7 pp 873ndash879 2002
Figure 3 Neonatal Tg mice Left neonatal Tg miceThe white arrow indicates cyanosis and abdominal bleeding in H+C+ neonates middlefluorescent images of the same mice right the lungs and heart of neonatal mice H+C+ hearts appear more dark reddish and smaller thanhearts of other genotypes Scale bar = 3mm
H+CminusH+C+
Figure 4 Histology of the neonatal heart and lung Top HampE staining of the neonatal heart of H+C+ mouse compared with that of H+Cminusmouse showing hemorrhage necrosis and degenerated cardiac muscles in H+C+ mouse heart Bottom the lung of H+C+ mouse showssigns of hemorrhage liquid congestion and tissue necrosis The alveolar lung structures appeared to have developed normally but poorlyinflated Scale bar = 100 120583m
BioMed Research International 7
H+C+ HminusCminus
Figure 5 Atrial septal defect in H+C+ neonatal mice HampE images of the neonatal heart from H+C+ and HminusCminus mice Asterisk indicatesthe atrial septal defect
H+C+ H+CminusMt
Mt
Mt
Mf
Mt
Mf
Figure 6 Transmission electron microscopy showing ultrastructure of myofibril defect in H+C+ neonatal heart Transmission electronmicroscopy results ofH+C+ (left) andH+Cminus (right)mice left ventricular tissueMfmyofibrilsMtmitochondria long arrow Z diskOriginalmagnification 10000x scale bar = 1 120583m
the cell nucleus through TUNEL assay whereas very fewTUNEL positively cells were in H+Cminus mouse heart sections(Figure 7(a)) In parallel a few cells in theHSP60mouse heartsection were positively stained by anti-activated caspase-3antibody significantly more than that of H+Cminus mouse heart(Figure 7(b)) The data clearly demonstrated that HSP60overexpression results in caspase-3 activation and apoptosisofmyocardial cells inHSP60mice Because theHSP60mouseheart became a sponge-like muscular tissue infiltrated bysmall capillaries and HSP60 involvement in rejection andautoimmune diseases was suggested previously we studiedthe presence of activated inflammatory cells in HSP60 miceThe pathology neither showed infiltrated mononuclear cells(Figure 4) nor detected CD4 or CD8 positive cells in HSP60mouse heart (Figure S3) thus inflammatory response orT cellactivation is absent in the HSP60 mouse heart
Accumulation of HSP60 in the cytoplasm during apop-tosis induction and the binding of HSP60 to procaspase-3and modulating caspase activity has been demonstrated inmany cancer cell lines [21 25 39] Similar to most proteins of
themitochondrialmatrix HSP60 which is synthesized in thecytoplasm with N-terminal targeting peptides is transportedto the mitochondrial matrix where the targeting sequenceis cleaved Both unprocessed (cytosolic) and matured (mito-chondrial) HSP60 strongly increased in the cardiac tissueof HSP60 mouse (Figure 1(b)) Consequently accumulationof cytosolic HSP60 capable of inducing apoptosis cannotbe overlooked and our data provided in vivo evidence ofapoptosis and caspase-3 activation in the HSP60mouse heart(Figure 7) Apoptosis is a highly regulated process duringembryonic and postnatal heart development and critical forheart development Increasing apoptosis in HSP60 mouseheart and its causal relation to cardiac myopathy must beinterpreted with caution because (1) apoptosis frequentlyoccurs in embryonic and neonatal tissues than in adultcardiac tissues (2) increasing apoptosis is often observedin cardiac tissues of myopathy and congestive heart failurethus apoptosis can be secondarily induced in myopathy andheart failure and (3) it can directly result from cytosolicHSP60 accumulation or HSP60 binding to caspases Thus
8 BioMed Research International
TdT TdTDAPI
H+Cminus
H+C+
(a)Casp 3DAPI
H+Cminus
H+C+
Casp 3
(b)
Figure 7 TUNEL staining and activated caspase-3 detection in neonatal heart sections (a) Detection of nuclear DNA fragmentation withTUNEL reaction in the left ventricle sections of neonatal H+C+ and H+Cminus mice Fluorescein-12-dUTP (TdT) and DAPI were used forTUNEL and nuclear staining (b) Immunofluorescence detection of activated (cleaved) caspase-3 induction in H+C+ and in H+Cminus heartsections Scale bar = 50120583m
additional studies are required in this regard Our currentmodel which uses the ubiquitous Cre mouse increasedHSP60 and potentially induced cell apoptosis of multipletissues thus it is unable to clarify phenotypes contributedby the HSP60 induction in a specific organ for examplevascular ECs smooth muscle cells or cardiac myocytesThis limitation can be overcome by combinatory uses of
G-Lox-HSP60 with vascular- or cardiac-specific Cre miceWe have explored HSP60 expression in cardiac tissues ofthe adult mice by using inducible cardiac cell-restricted Cremouse (unpublished results)
In addition to the roles of mitochondrial and cytosolicchaperonins and caspase induction additional functions ofHSP60 have been reported Cytosolic HSP60 regulates the
BioMed Research International 9
phosphorylation of IKK120572120573 and promotes NF-120581B prosur-vival pathway Transgenic expression of the cytosolic HSP60prevented stress-induced hepatic cell deaths in vivo [32]Overexpressing HSP60 in the human stem cells increasedHSP60 nuclear localization which suppressed ROS andp38JNK signaling and regulated genes relating to stem cellproliferation differentiation and stemness [40]Whether theaforementioned mechanisms are also involved in mediatingneonatal deaths remains unclear and future clarification isawaited
In this study congenital ASD was observed in a largenumber ofHSP60mice AlthoughASD exacerbated the exist-ing myopathy of HSP60 mice and was partially responsiblefor neonatal death ASD per sewas unlikely directly caused byHSP60 induction We speculate that ASD in neonatal HSP60mice may be the consequence of incomplete embryonicdevelopment existing myopathy and increasing apoptosisSimilar phenomenon has been demonstrated in the eNOSknockout mouse model in which high incidence of ASDor VSD was precipitated through increased apoptosis duringembryonic and postnatal stages [41] Our main findingsindicate that severe hemorrhage andmyocyte death occurredin mice with HSP60 expression and increased apoptosisin a macerated and spongy-like heart failed to meet thecardiovascular demands after birth and was the primarycause of HSP60-induced heart failure and neonatal death
4 Conclusions
HSP60 not only is the essentialmitochondrial chaperone pro-tein but also plays critical roles outside the mitochondrionAlthough numerous studies indicated the wide involvementof HSP60 in broad biological processes and many diseasesreports of transgenic mouse models of increasing or knock-down HSP60 expression are rare Thus the consequences ofaltering HSP60 levels in vivo in these disease settings remainunknown We used a floxed vector to establish conditionalHSP60 transgenic mice which allows studies on inducibleand organ-targeted HSP60 expression in adult mice and ispotentially useful in pursuing essential questions related toHSP60 in vivo In this study we show that ubiquitous HSP60expression from the embryonic stage results in increasedapoptosis myopathy high incidence of ASD and neonataldeaths
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Tsung-Hsien Chen and Shan-Wen Liu contribute equally tothis paper
Acknowledgments
The authors gratefully acknowledge the technical assistanceprovided by Tzu-Chin Wu and Mei-Yu Fan in NHRI-IBENand Functional and Micro-Magnetic Resonance ImagingCenter supported by NRPGM (NSC97-3112-B-001-013) Thisresearch was supported by grants from NHRI (BN-104-PP-02 BN-104-PP-24) and from Ministry of Science andTechnology (103-2320-B-400-018)
References
[1] I J Benjamin andD RMcMillan ldquoStress (heat shock) proteinsmolecular chaperones in cardiovascular biology and diseaserdquoCirculation Research vol 83 no 2 pp 117ndash132 1998
[2] F U Hartl ldquoMolecular chaperones in cellular protein foldingrdquoNature vol 381 no 6583 pp 571ndash580 1996
[3] J C Ranford and B Henderson ldquoChaperonins in diseasemechanismsmodels and treatmentsrdquoMolecular Pathology vol55 no 4 pp 209ndash213 2002
[4] CCDeocaris S CKaul andRWadhwa ldquoOn the brotherhoodof the mitochondrial chaperones mortalin and heat shockprotein 60rdquo Cell Stress and Chaperones vol 11 no 2 pp 116ndash128 2006
[5] F Cappello A M Gammazza A P Piccionello et al ldquoHsp60chaperonopathies and chaperonotherapy targets and agentsrdquoExpert Opinion on Therapeutic Targets vol 18 no 2 pp 185ndash208 2014
[6] S Lau N Patnaik M R Sayen and R Mestril ldquoSimultaneousoverexpression of two stress proteins in rat cardiomyocytes andmyogenic cells confers protection against ischemia-inducedinjuryrdquo Circulation vol 96 no 7 pp 2287ndash2294 1997
[7] K M Lin B Lin I Y Lian R Mestril I E Scheffler and WH Dillmann ldquoCombined and individual mitochondrial HSP60and HSP10 expression in cardiac myocytes protects mitochon-drial function and prevents apoptotic cell deaths induced bysimulated ischemia-reoxygenationrdquo Circulation vol 103 no 13pp 1787ndash1792 2001
[8] J J Malago J F J G Koninkx and J E van Dijk ldquoThe heatshock response and cytoprotection of the intestinal epitheliumrdquoCell Stress and Chaperones vol 7 no 2 pp 191ndash199 2002
[9] Y-X Shan T-L Yang R Mestril and P H Wang ldquoHsp10 andHsp60 suppress ubiquitination of insulin-like growth factor-1receptor and augment insulin-like growth factor-1 receptor sig-naling in cardiac muscle implications on decreased myocardialprotection in diabetic cardiomyopathyrdquoThe Journal of BiologicalChemistry vol 278 no 46 pp 45492ndash45498 2003
[10] H-S Chen Y-X Shan T-L Yang et al ldquoInsulin deficiencydownregulated heat shock protein 60 and IGF-1 receptorsignaling in diabetic myocardiumrdquo Diabetes vol 54 no 1 pp175ndash181 2005
10 BioMed Research International
[11] V Calabrese A Ravagna C Colombrita et al ldquoAcetylcarnitineinduces heme oxygenase in rat astrocytes and protects againstoxidative stress Involvement of the transcription factor Nrf2rdquoJournal of Neuroscience Research vol 79 no 4 pp 509ndash5212005
[12] V Veereshwarayya P Kumar K M Rosen R Mestril andH W Querfurth ldquoDifferential effects of mitochondrial heatshock protein 60 and related molecular chaperones to preventintracellular 120573-amyloid-induced inhibition of complex IV andlimit apoptosisrdquoThe Journal of Biological Chemistry vol 281 no40 pp 29468ndash29478 2006
[13] C Xu Y Lu Z Pan et al ldquoThe muscle-specific microRNAsmiR-1 and miR-133 produce opposing effects on apoptosis bytargeting HSP60 HSP70 and caspase-9 in cardiomyocytesrdquoJournal of Cell Science vol 120 part 17 pp 3045ndash3052 2007
[14] J C Ghosh T Dohi H K Byoung and D C Altieri ldquoHsp60regulation of tumor cell apoptosisrdquo The Journal of BiologicalChemistry vol 283 no 8 pp 5188ndash5194 2008
[15] P E Castle R Ashfaq F Ansari and C Y Muller ldquoImmuno-histochemical evaluation of heat shock proteins in normal andpreinvasive lesions of the cervixrdquo Cancer Letters vol 229 no 2pp 245ndash252 2005
[16] P A Cornford A R Dodson K F Parsons et al ldquoHeat shockprotein expression independently predicts clinical outcome inprostate cancerrdquoCancer Research vol 60 no 24 pp 7099ndash71052000
[17] F Cappello S David F Rappa et al ldquoThe expression of Hsp60and Hsp10 in large bowel carcinomas with lymph node metas-tasesrdquo BMC Cancer vol 5 article 39 2005
[18] P-L Hsu and S-M Hsu ldquoAbundance of heat shock proteins(hsp89 hsp60 and hsp27) in malignant cells of Hodgkinrsquosdiseaserdquo Cancer Research vol 58 no 23 pp 5507ndash5513 1998
[19] S Gupta and A A Knowlton ldquoHSP60 Bax apoptosis and theheartrdquo Journal of Cellular and Molecular Medicine vol 9 no 1pp 51ndash58 2005
[20] S Xanthoudakis R Sophie D Rasper et al ldquoHsp60 accel-erates the maturation of pro-caspase-3 by upstream activatorproteases during apoptosisrdquo The EMBO Journal vol 18 no 8pp 2049ndash2056 1999
[21] A Samali J Cai B Zhivotovsky D P Jones and S OrreniusldquoPresence of a pre-apoptotic complex of pro-caspase-3 Hsp60and Hsp10 in the mitochondrial fraction of Jurkat cellsrdquo TheEMBO Journal vol 18 no 8 pp 2040ndash2048 1999
[22] A Faried M Sohda M Nakajima TMiyazaki H Kato andHKuwano ldquoExpression of heat-shock protein Hsp60 correlatedwith the apoptotic index and patient prognosis in humanoesophageal squamous cell carcinomardquo European Journal ofCancer vol 40 no 18 pp 2804ndash2811 2004
[23] J Schneider E Jimenez K Marenbach H Romero D Marxand H Meden ldquoImmunohistochemical detection of HSP60-expression in human ovarian cancer Correlation with survivalin a series of 247 patientsrdquoAnticancer Research vol 19 no 3 pp2141ndash2146 1999
[24] T Lebret R W G Watson V Molinie et al ldquoHeat shockproteins HSP27 HSP60 HSP70 and HSP90 expression inbladder carcinomardquo Cancer vol 98 no 5 pp 970ndash977 2003
[25] A Samali and S Orrenius ldquoHeat shock proteins regulators ofstress response and apoptosisrdquo Cell Stress and Chaperones vol3 no 4 pp 228ndash236 1998
[26] W Van Eden G Wick S Albani and I Cohen ldquoStress heatshock proteins and autoimmunity how immune responses to
heat shock proteins are to be used for the control of chronicinflammatory diseasesrdquo Annals of the New York Academy ofSciences vol 1113 pp 217ndash237 2007
[27] C Habich and V Burkart ldquoHeat shock protein 60 regulatoryrole on innate immune cellsrdquo Cellular and Molecular LifeSciences vol 64 no 6 pp 742ndash751 2007
[28] Y Kondo Y Ueno K Kobayashi et al ldquoHepatitis B virus repli-cation could enhance regulatory T cell activity by producingsoluble heat shock protein 60 from hepatocytesrdquoThe Journal ofInfectious Diseases vol 202 no 2 pp 202ndash213 2010
[29] C Colotti G Cavallini R L Vitale et al ldquoEffects of agingand anti-aging caloric restrictions on carbonyl and heat shockprotein levels and expressionrdquo Biogerontology vol 6 no 6 pp397ndash406 2005
[30] P L Hooper ldquoInsulin signaling GSK-3 heat shock proteins andthe natural history of type 2 diabetes mellitus a hypothesisrdquoMetabolic Syndrome and RelatedDisorders vol 5 no 3 pp 220ndash230 2007
[31] O S Birk D C Douek D Elias et al ldquoA role of Hsp60 inautoimmune diabetes analysis in a transgenic modelrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 93 no 3 pp 1032ndash1037 1996
[32] J N Chun B Choi KW Lee et al ldquoCytosolicHsp60 is involvedin the NF-kappaB-dependent survival of cancer cells via IKKregulatiordquo PLoS ONE vol 5 no 3 Article ID e9422 2010
[33] M-R Chen S-W Liu T-C Wu et al ldquoRU486-induciblerecombination in the salivary glands of lactoferrin promoter-driven green fluorescent Cre transgenic micerdquo Genesis vol 48no 10 pp 585ndash595 2010
[34] H-Y Lin C-H Kao KM-C Lin V Kaartinen and L-T YangldquoNotch signaling regulates late-stage epidermal differentiationandmaintains postnatal hair cycle homeostasisrdquoPLoSONE vol6 no 1 Article ID e15842 2011
[35] A C Kayani G L Close W H Dillmann R Mestril M JJackson and A McArdle ldquoOverexpression of HSP10 in skeletalmuscle of transgenic mice prevents the age-related fall inmaximum tetanic force generation and muscle Cross-SectionalAreardquoThe American Journal of PhysiologymdashRegulatory Integra-tive and Comparative Physiology vol 299 no 1 pp R268ndashR2762010
[36] G-C Fan X Ren J Qian et al ldquoNovel cardioprotective role of asmall heat-shock protein Hsp20 against ischemiareperfusioninjuryrdquo Circulation vol 111 no 14 pp 1792ndash1799 2005
[37] J M Hollander J L Martin D D Belke et al ldquoOverex-pression of wild-type heat shock protein 27 and a nonphos-phorylatable heat shock protein 27 mutant protects againstischemiareperfusion injury in a transgenic mouse modelrdquoCirculation vol 110 no 23 pp 3544ndash3552 2004
[38] J-C L Plumier A M Krueger R W Currie D KontoyiannisG Kollias and G N Pagoulatos ldquoTransgenic mice expressingthe human inducible Hsp70 have hippocampal neurons resis-tant to ischemic injuryrdquo Cell Stress and Chaperones vol 2 no 3pp 162ndash167 1997
[39] D Chandra G Choy and D G Tang ldquoCytosolic accumulationof HSP60 during apoptosis with or without apparentmitochon-drial release Evidence that its pro-apoptotic or pro-survivalfunctions involve differential interactions with caspase-3rdquo TheJournal of Biological Chemistry vol 282 no 43 pp 31289ndash313012007
BioMed Research International 11
[40] J H Jang J S Jung J I Choi and S K Kang ldquoNuclearAgo2HSP60 contributes to broad spectrum of hATSCs func-tion via Oct4 regulationrdquoAntioxidants and Redox Signaling vol16 no 5 pp 383ndash399 2012
[41] Q Feng W Song X Lu et al ldquoDevelopment of heart failureand congenital septal defects in mice lacking endothelial nitricoxide synthaserdquo Circulation vol 106 no 7 pp 873ndash879 2002
Figure 5 Atrial septal defect in H+C+ neonatal mice HampE images of the neonatal heart from H+C+ and HminusCminus mice Asterisk indicatesthe atrial septal defect
H+C+ H+CminusMt
Mt
Mt
Mf
Mt
Mf
Figure 6 Transmission electron microscopy showing ultrastructure of myofibril defect in H+C+ neonatal heart Transmission electronmicroscopy results ofH+C+ (left) andH+Cminus (right)mice left ventricular tissueMfmyofibrilsMtmitochondria long arrow Z diskOriginalmagnification 10000x scale bar = 1 120583m
the cell nucleus through TUNEL assay whereas very fewTUNEL positively cells were in H+Cminus mouse heart sections(Figure 7(a)) In parallel a few cells in theHSP60mouse heartsection were positively stained by anti-activated caspase-3antibody significantly more than that of H+Cminus mouse heart(Figure 7(b)) The data clearly demonstrated that HSP60overexpression results in caspase-3 activation and apoptosisofmyocardial cells inHSP60mice Because theHSP60mouseheart became a sponge-like muscular tissue infiltrated bysmall capillaries and HSP60 involvement in rejection andautoimmune diseases was suggested previously we studiedthe presence of activated inflammatory cells in HSP60 miceThe pathology neither showed infiltrated mononuclear cells(Figure 4) nor detected CD4 or CD8 positive cells in HSP60mouse heart (Figure S3) thus inflammatory response orT cellactivation is absent in the HSP60 mouse heart
Accumulation of HSP60 in the cytoplasm during apop-tosis induction and the binding of HSP60 to procaspase-3and modulating caspase activity has been demonstrated inmany cancer cell lines [21 25 39] Similar to most proteins of
themitochondrialmatrix HSP60 which is synthesized in thecytoplasm with N-terminal targeting peptides is transportedto the mitochondrial matrix where the targeting sequenceis cleaved Both unprocessed (cytosolic) and matured (mito-chondrial) HSP60 strongly increased in the cardiac tissueof HSP60 mouse (Figure 1(b)) Consequently accumulationof cytosolic HSP60 capable of inducing apoptosis cannotbe overlooked and our data provided in vivo evidence ofapoptosis and caspase-3 activation in the HSP60mouse heart(Figure 7) Apoptosis is a highly regulated process duringembryonic and postnatal heart development and critical forheart development Increasing apoptosis in HSP60 mouseheart and its causal relation to cardiac myopathy must beinterpreted with caution because (1) apoptosis frequentlyoccurs in embryonic and neonatal tissues than in adultcardiac tissues (2) increasing apoptosis is often observedin cardiac tissues of myopathy and congestive heart failurethus apoptosis can be secondarily induced in myopathy andheart failure and (3) it can directly result from cytosolicHSP60 accumulation or HSP60 binding to caspases Thus
8 BioMed Research International
TdT TdTDAPI
H+Cminus
H+C+
(a)Casp 3DAPI
H+Cminus
H+C+
Casp 3
(b)
Figure 7 TUNEL staining and activated caspase-3 detection in neonatal heart sections (a) Detection of nuclear DNA fragmentation withTUNEL reaction in the left ventricle sections of neonatal H+C+ and H+Cminus mice Fluorescein-12-dUTP (TdT) and DAPI were used forTUNEL and nuclear staining (b) Immunofluorescence detection of activated (cleaved) caspase-3 induction in H+C+ and in H+Cminus heartsections Scale bar = 50120583m
additional studies are required in this regard Our currentmodel which uses the ubiquitous Cre mouse increasedHSP60 and potentially induced cell apoptosis of multipletissues thus it is unable to clarify phenotypes contributedby the HSP60 induction in a specific organ for examplevascular ECs smooth muscle cells or cardiac myocytesThis limitation can be overcome by combinatory uses of
G-Lox-HSP60 with vascular- or cardiac-specific Cre miceWe have explored HSP60 expression in cardiac tissues ofthe adult mice by using inducible cardiac cell-restricted Cremouse (unpublished results)
In addition to the roles of mitochondrial and cytosolicchaperonins and caspase induction additional functions ofHSP60 have been reported Cytosolic HSP60 regulates the
BioMed Research International 9
phosphorylation of IKK120572120573 and promotes NF-120581B prosur-vival pathway Transgenic expression of the cytosolic HSP60prevented stress-induced hepatic cell deaths in vivo [32]Overexpressing HSP60 in the human stem cells increasedHSP60 nuclear localization which suppressed ROS andp38JNK signaling and regulated genes relating to stem cellproliferation differentiation and stemness [40]Whether theaforementioned mechanisms are also involved in mediatingneonatal deaths remains unclear and future clarification isawaited
In this study congenital ASD was observed in a largenumber ofHSP60mice AlthoughASD exacerbated the exist-ing myopathy of HSP60 mice and was partially responsiblefor neonatal death ASD per sewas unlikely directly caused byHSP60 induction We speculate that ASD in neonatal HSP60mice may be the consequence of incomplete embryonicdevelopment existing myopathy and increasing apoptosisSimilar phenomenon has been demonstrated in the eNOSknockout mouse model in which high incidence of ASDor VSD was precipitated through increased apoptosis duringembryonic and postnatal stages [41] Our main findingsindicate that severe hemorrhage andmyocyte death occurredin mice with HSP60 expression and increased apoptosisin a macerated and spongy-like heart failed to meet thecardiovascular demands after birth and was the primarycause of HSP60-induced heart failure and neonatal death
4 Conclusions
HSP60 not only is the essentialmitochondrial chaperone pro-tein but also plays critical roles outside the mitochondrionAlthough numerous studies indicated the wide involvementof HSP60 in broad biological processes and many diseasesreports of transgenic mouse models of increasing or knock-down HSP60 expression are rare Thus the consequences ofaltering HSP60 levels in vivo in these disease settings remainunknown We used a floxed vector to establish conditionalHSP60 transgenic mice which allows studies on inducibleand organ-targeted HSP60 expression in adult mice and ispotentially useful in pursuing essential questions related toHSP60 in vivo In this study we show that ubiquitous HSP60expression from the embryonic stage results in increasedapoptosis myopathy high incidence of ASD and neonataldeaths
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Tsung-Hsien Chen and Shan-Wen Liu contribute equally tothis paper
Acknowledgments
The authors gratefully acknowledge the technical assistanceprovided by Tzu-Chin Wu and Mei-Yu Fan in NHRI-IBENand Functional and Micro-Magnetic Resonance ImagingCenter supported by NRPGM (NSC97-3112-B-001-013) Thisresearch was supported by grants from NHRI (BN-104-PP-02 BN-104-PP-24) and from Ministry of Science andTechnology (103-2320-B-400-018)
References
[1] I J Benjamin andD RMcMillan ldquoStress (heat shock) proteinsmolecular chaperones in cardiovascular biology and diseaserdquoCirculation Research vol 83 no 2 pp 117ndash132 1998
[2] F U Hartl ldquoMolecular chaperones in cellular protein foldingrdquoNature vol 381 no 6583 pp 571ndash580 1996
[3] J C Ranford and B Henderson ldquoChaperonins in diseasemechanismsmodels and treatmentsrdquoMolecular Pathology vol55 no 4 pp 209ndash213 2002
[4] CCDeocaris S CKaul andRWadhwa ldquoOn the brotherhoodof the mitochondrial chaperones mortalin and heat shockprotein 60rdquo Cell Stress and Chaperones vol 11 no 2 pp 116ndash128 2006
[5] F Cappello A M Gammazza A P Piccionello et al ldquoHsp60chaperonopathies and chaperonotherapy targets and agentsrdquoExpert Opinion on Therapeutic Targets vol 18 no 2 pp 185ndash208 2014
[6] S Lau N Patnaik M R Sayen and R Mestril ldquoSimultaneousoverexpression of two stress proteins in rat cardiomyocytes andmyogenic cells confers protection against ischemia-inducedinjuryrdquo Circulation vol 96 no 7 pp 2287ndash2294 1997
[7] K M Lin B Lin I Y Lian R Mestril I E Scheffler and WH Dillmann ldquoCombined and individual mitochondrial HSP60and HSP10 expression in cardiac myocytes protects mitochon-drial function and prevents apoptotic cell deaths induced bysimulated ischemia-reoxygenationrdquo Circulation vol 103 no 13pp 1787ndash1792 2001
[8] J J Malago J F J G Koninkx and J E van Dijk ldquoThe heatshock response and cytoprotection of the intestinal epitheliumrdquoCell Stress and Chaperones vol 7 no 2 pp 191ndash199 2002
[9] Y-X Shan T-L Yang R Mestril and P H Wang ldquoHsp10 andHsp60 suppress ubiquitination of insulin-like growth factor-1receptor and augment insulin-like growth factor-1 receptor sig-naling in cardiac muscle implications on decreased myocardialprotection in diabetic cardiomyopathyrdquoThe Journal of BiologicalChemistry vol 278 no 46 pp 45492ndash45498 2003
[10] H-S Chen Y-X Shan T-L Yang et al ldquoInsulin deficiencydownregulated heat shock protein 60 and IGF-1 receptorsignaling in diabetic myocardiumrdquo Diabetes vol 54 no 1 pp175ndash181 2005
10 BioMed Research International
[11] V Calabrese A Ravagna C Colombrita et al ldquoAcetylcarnitineinduces heme oxygenase in rat astrocytes and protects againstoxidative stress Involvement of the transcription factor Nrf2rdquoJournal of Neuroscience Research vol 79 no 4 pp 509ndash5212005
[12] V Veereshwarayya P Kumar K M Rosen R Mestril andH W Querfurth ldquoDifferential effects of mitochondrial heatshock protein 60 and related molecular chaperones to preventintracellular 120573-amyloid-induced inhibition of complex IV andlimit apoptosisrdquoThe Journal of Biological Chemistry vol 281 no40 pp 29468ndash29478 2006
[13] C Xu Y Lu Z Pan et al ldquoThe muscle-specific microRNAsmiR-1 and miR-133 produce opposing effects on apoptosis bytargeting HSP60 HSP70 and caspase-9 in cardiomyocytesrdquoJournal of Cell Science vol 120 part 17 pp 3045ndash3052 2007
[14] J C Ghosh T Dohi H K Byoung and D C Altieri ldquoHsp60regulation of tumor cell apoptosisrdquo The Journal of BiologicalChemistry vol 283 no 8 pp 5188ndash5194 2008
[15] P E Castle R Ashfaq F Ansari and C Y Muller ldquoImmuno-histochemical evaluation of heat shock proteins in normal andpreinvasive lesions of the cervixrdquo Cancer Letters vol 229 no 2pp 245ndash252 2005
[16] P A Cornford A R Dodson K F Parsons et al ldquoHeat shockprotein expression independently predicts clinical outcome inprostate cancerrdquoCancer Research vol 60 no 24 pp 7099ndash71052000
[17] F Cappello S David F Rappa et al ldquoThe expression of Hsp60and Hsp10 in large bowel carcinomas with lymph node metas-tasesrdquo BMC Cancer vol 5 article 39 2005
[18] P-L Hsu and S-M Hsu ldquoAbundance of heat shock proteins(hsp89 hsp60 and hsp27) in malignant cells of Hodgkinrsquosdiseaserdquo Cancer Research vol 58 no 23 pp 5507ndash5513 1998
[19] S Gupta and A A Knowlton ldquoHSP60 Bax apoptosis and theheartrdquo Journal of Cellular and Molecular Medicine vol 9 no 1pp 51ndash58 2005
[20] S Xanthoudakis R Sophie D Rasper et al ldquoHsp60 accel-erates the maturation of pro-caspase-3 by upstream activatorproteases during apoptosisrdquo The EMBO Journal vol 18 no 8pp 2049ndash2056 1999
[21] A Samali J Cai B Zhivotovsky D P Jones and S OrreniusldquoPresence of a pre-apoptotic complex of pro-caspase-3 Hsp60and Hsp10 in the mitochondrial fraction of Jurkat cellsrdquo TheEMBO Journal vol 18 no 8 pp 2040ndash2048 1999
[22] A Faried M Sohda M Nakajima TMiyazaki H Kato andHKuwano ldquoExpression of heat-shock protein Hsp60 correlatedwith the apoptotic index and patient prognosis in humanoesophageal squamous cell carcinomardquo European Journal ofCancer vol 40 no 18 pp 2804ndash2811 2004
[23] J Schneider E Jimenez K Marenbach H Romero D Marxand H Meden ldquoImmunohistochemical detection of HSP60-expression in human ovarian cancer Correlation with survivalin a series of 247 patientsrdquoAnticancer Research vol 19 no 3 pp2141ndash2146 1999
[24] T Lebret R W G Watson V Molinie et al ldquoHeat shockproteins HSP27 HSP60 HSP70 and HSP90 expression inbladder carcinomardquo Cancer vol 98 no 5 pp 970ndash977 2003
[25] A Samali and S Orrenius ldquoHeat shock proteins regulators ofstress response and apoptosisrdquo Cell Stress and Chaperones vol3 no 4 pp 228ndash236 1998
[26] W Van Eden G Wick S Albani and I Cohen ldquoStress heatshock proteins and autoimmunity how immune responses to
heat shock proteins are to be used for the control of chronicinflammatory diseasesrdquo Annals of the New York Academy ofSciences vol 1113 pp 217ndash237 2007
[27] C Habich and V Burkart ldquoHeat shock protein 60 regulatoryrole on innate immune cellsrdquo Cellular and Molecular LifeSciences vol 64 no 6 pp 742ndash751 2007
[28] Y Kondo Y Ueno K Kobayashi et al ldquoHepatitis B virus repli-cation could enhance regulatory T cell activity by producingsoluble heat shock protein 60 from hepatocytesrdquoThe Journal ofInfectious Diseases vol 202 no 2 pp 202ndash213 2010
[29] C Colotti G Cavallini R L Vitale et al ldquoEffects of agingand anti-aging caloric restrictions on carbonyl and heat shockprotein levels and expressionrdquo Biogerontology vol 6 no 6 pp397ndash406 2005
[30] P L Hooper ldquoInsulin signaling GSK-3 heat shock proteins andthe natural history of type 2 diabetes mellitus a hypothesisrdquoMetabolic Syndrome and RelatedDisorders vol 5 no 3 pp 220ndash230 2007
[31] O S Birk D C Douek D Elias et al ldquoA role of Hsp60 inautoimmune diabetes analysis in a transgenic modelrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 93 no 3 pp 1032ndash1037 1996
[32] J N Chun B Choi KW Lee et al ldquoCytosolicHsp60 is involvedin the NF-kappaB-dependent survival of cancer cells via IKKregulatiordquo PLoS ONE vol 5 no 3 Article ID e9422 2010
[33] M-R Chen S-W Liu T-C Wu et al ldquoRU486-induciblerecombination in the salivary glands of lactoferrin promoter-driven green fluorescent Cre transgenic micerdquo Genesis vol 48no 10 pp 585ndash595 2010
[34] H-Y Lin C-H Kao KM-C Lin V Kaartinen and L-T YangldquoNotch signaling regulates late-stage epidermal differentiationandmaintains postnatal hair cycle homeostasisrdquoPLoSONE vol6 no 1 Article ID e15842 2011
[35] A C Kayani G L Close W H Dillmann R Mestril M JJackson and A McArdle ldquoOverexpression of HSP10 in skeletalmuscle of transgenic mice prevents the age-related fall inmaximum tetanic force generation and muscle Cross-SectionalAreardquoThe American Journal of PhysiologymdashRegulatory Integra-tive and Comparative Physiology vol 299 no 1 pp R268ndashR2762010
[36] G-C Fan X Ren J Qian et al ldquoNovel cardioprotective role of asmall heat-shock protein Hsp20 against ischemiareperfusioninjuryrdquo Circulation vol 111 no 14 pp 1792ndash1799 2005
[37] J M Hollander J L Martin D D Belke et al ldquoOverex-pression of wild-type heat shock protein 27 and a nonphos-phorylatable heat shock protein 27 mutant protects againstischemiareperfusion injury in a transgenic mouse modelrdquoCirculation vol 110 no 23 pp 3544ndash3552 2004
[38] J-C L Plumier A M Krueger R W Currie D KontoyiannisG Kollias and G N Pagoulatos ldquoTransgenic mice expressingthe human inducible Hsp70 have hippocampal neurons resis-tant to ischemic injuryrdquo Cell Stress and Chaperones vol 2 no 3pp 162ndash167 1997
[39] D Chandra G Choy and D G Tang ldquoCytosolic accumulationof HSP60 during apoptosis with or without apparentmitochon-drial release Evidence that its pro-apoptotic or pro-survivalfunctions involve differential interactions with caspase-3rdquo TheJournal of Biological Chemistry vol 282 no 43 pp 31289ndash313012007
BioMed Research International 11
[40] J H Jang J S Jung J I Choi and S K Kang ldquoNuclearAgo2HSP60 contributes to broad spectrum of hATSCs func-tion via Oct4 regulationrdquoAntioxidants and Redox Signaling vol16 no 5 pp 383ndash399 2012
[41] Q Feng W Song X Lu et al ldquoDevelopment of heart failureand congenital septal defects in mice lacking endothelial nitricoxide synthaserdquo Circulation vol 106 no 7 pp 873ndash879 2002
Figure 7 TUNEL staining and activated caspase-3 detection in neonatal heart sections (a) Detection of nuclear DNA fragmentation withTUNEL reaction in the left ventricle sections of neonatal H+C+ and H+Cminus mice Fluorescein-12-dUTP (TdT) and DAPI were used forTUNEL and nuclear staining (b) Immunofluorescence detection of activated (cleaved) caspase-3 induction in H+C+ and in H+Cminus heartsections Scale bar = 50120583m
additional studies are required in this regard Our currentmodel which uses the ubiquitous Cre mouse increasedHSP60 and potentially induced cell apoptosis of multipletissues thus it is unable to clarify phenotypes contributedby the HSP60 induction in a specific organ for examplevascular ECs smooth muscle cells or cardiac myocytesThis limitation can be overcome by combinatory uses of
G-Lox-HSP60 with vascular- or cardiac-specific Cre miceWe have explored HSP60 expression in cardiac tissues ofthe adult mice by using inducible cardiac cell-restricted Cremouse (unpublished results)
In addition to the roles of mitochondrial and cytosolicchaperonins and caspase induction additional functions ofHSP60 have been reported Cytosolic HSP60 regulates the
BioMed Research International 9
phosphorylation of IKK120572120573 and promotes NF-120581B prosur-vival pathway Transgenic expression of the cytosolic HSP60prevented stress-induced hepatic cell deaths in vivo [32]Overexpressing HSP60 in the human stem cells increasedHSP60 nuclear localization which suppressed ROS andp38JNK signaling and regulated genes relating to stem cellproliferation differentiation and stemness [40]Whether theaforementioned mechanisms are also involved in mediatingneonatal deaths remains unclear and future clarification isawaited
In this study congenital ASD was observed in a largenumber ofHSP60mice AlthoughASD exacerbated the exist-ing myopathy of HSP60 mice and was partially responsiblefor neonatal death ASD per sewas unlikely directly caused byHSP60 induction We speculate that ASD in neonatal HSP60mice may be the consequence of incomplete embryonicdevelopment existing myopathy and increasing apoptosisSimilar phenomenon has been demonstrated in the eNOSknockout mouse model in which high incidence of ASDor VSD was precipitated through increased apoptosis duringembryonic and postnatal stages [41] Our main findingsindicate that severe hemorrhage andmyocyte death occurredin mice with HSP60 expression and increased apoptosisin a macerated and spongy-like heart failed to meet thecardiovascular demands after birth and was the primarycause of HSP60-induced heart failure and neonatal death
4 Conclusions
HSP60 not only is the essentialmitochondrial chaperone pro-tein but also plays critical roles outside the mitochondrionAlthough numerous studies indicated the wide involvementof HSP60 in broad biological processes and many diseasesreports of transgenic mouse models of increasing or knock-down HSP60 expression are rare Thus the consequences ofaltering HSP60 levels in vivo in these disease settings remainunknown We used a floxed vector to establish conditionalHSP60 transgenic mice which allows studies on inducibleand organ-targeted HSP60 expression in adult mice and ispotentially useful in pursuing essential questions related toHSP60 in vivo In this study we show that ubiquitous HSP60expression from the embryonic stage results in increasedapoptosis myopathy high incidence of ASD and neonataldeaths
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Tsung-Hsien Chen and Shan-Wen Liu contribute equally tothis paper
Acknowledgments
The authors gratefully acknowledge the technical assistanceprovided by Tzu-Chin Wu and Mei-Yu Fan in NHRI-IBENand Functional and Micro-Magnetic Resonance ImagingCenter supported by NRPGM (NSC97-3112-B-001-013) Thisresearch was supported by grants from NHRI (BN-104-PP-02 BN-104-PP-24) and from Ministry of Science andTechnology (103-2320-B-400-018)
References
[1] I J Benjamin andD RMcMillan ldquoStress (heat shock) proteinsmolecular chaperones in cardiovascular biology and diseaserdquoCirculation Research vol 83 no 2 pp 117ndash132 1998
[2] F U Hartl ldquoMolecular chaperones in cellular protein foldingrdquoNature vol 381 no 6583 pp 571ndash580 1996
[3] J C Ranford and B Henderson ldquoChaperonins in diseasemechanismsmodels and treatmentsrdquoMolecular Pathology vol55 no 4 pp 209ndash213 2002
[4] CCDeocaris S CKaul andRWadhwa ldquoOn the brotherhoodof the mitochondrial chaperones mortalin and heat shockprotein 60rdquo Cell Stress and Chaperones vol 11 no 2 pp 116ndash128 2006
[5] F Cappello A M Gammazza A P Piccionello et al ldquoHsp60chaperonopathies and chaperonotherapy targets and agentsrdquoExpert Opinion on Therapeutic Targets vol 18 no 2 pp 185ndash208 2014
[6] S Lau N Patnaik M R Sayen and R Mestril ldquoSimultaneousoverexpression of two stress proteins in rat cardiomyocytes andmyogenic cells confers protection against ischemia-inducedinjuryrdquo Circulation vol 96 no 7 pp 2287ndash2294 1997
[7] K M Lin B Lin I Y Lian R Mestril I E Scheffler and WH Dillmann ldquoCombined and individual mitochondrial HSP60and HSP10 expression in cardiac myocytes protects mitochon-drial function and prevents apoptotic cell deaths induced bysimulated ischemia-reoxygenationrdquo Circulation vol 103 no 13pp 1787ndash1792 2001
[8] J J Malago J F J G Koninkx and J E van Dijk ldquoThe heatshock response and cytoprotection of the intestinal epitheliumrdquoCell Stress and Chaperones vol 7 no 2 pp 191ndash199 2002
[9] Y-X Shan T-L Yang R Mestril and P H Wang ldquoHsp10 andHsp60 suppress ubiquitination of insulin-like growth factor-1receptor and augment insulin-like growth factor-1 receptor sig-naling in cardiac muscle implications on decreased myocardialprotection in diabetic cardiomyopathyrdquoThe Journal of BiologicalChemistry vol 278 no 46 pp 45492ndash45498 2003
[10] H-S Chen Y-X Shan T-L Yang et al ldquoInsulin deficiencydownregulated heat shock protein 60 and IGF-1 receptorsignaling in diabetic myocardiumrdquo Diabetes vol 54 no 1 pp175ndash181 2005
10 BioMed Research International
[11] V Calabrese A Ravagna C Colombrita et al ldquoAcetylcarnitineinduces heme oxygenase in rat astrocytes and protects againstoxidative stress Involvement of the transcription factor Nrf2rdquoJournal of Neuroscience Research vol 79 no 4 pp 509ndash5212005
[12] V Veereshwarayya P Kumar K M Rosen R Mestril andH W Querfurth ldquoDifferential effects of mitochondrial heatshock protein 60 and related molecular chaperones to preventintracellular 120573-amyloid-induced inhibition of complex IV andlimit apoptosisrdquoThe Journal of Biological Chemistry vol 281 no40 pp 29468ndash29478 2006
[13] C Xu Y Lu Z Pan et al ldquoThe muscle-specific microRNAsmiR-1 and miR-133 produce opposing effects on apoptosis bytargeting HSP60 HSP70 and caspase-9 in cardiomyocytesrdquoJournal of Cell Science vol 120 part 17 pp 3045ndash3052 2007
[14] J C Ghosh T Dohi H K Byoung and D C Altieri ldquoHsp60regulation of tumor cell apoptosisrdquo The Journal of BiologicalChemistry vol 283 no 8 pp 5188ndash5194 2008
[15] P E Castle R Ashfaq F Ansari and C Y Muller ldquoImmuno-histochemical evaluation of heat shock proteins in normal andpreinvasive lesions of the cervixrdquo Cancer Letters vol 229 no 2pp 245ndash252 2005
[16] P A Cornford A R Dodson K F Parsons et al ldquoHeat shockprotein expression independently predicts clinical outcome inprostate cancerrdquoCancer Research vol 60 no 24 pp 7099ndash71052000
[17] F Cappello S David F Rappa et al ldquoThe expression of Hsp60and Hsp10 in large bowel carcinomas with lymph node metas-tasesrdquo BMC Cancer vol 5 article 39 2005
[18] P-L Hsu and S-M Hsu ldquoAbundance of heat shock proteins(hsp89 hsp60 and hsp27) in malignant cells of Hodgkinrsquosdiseaserdquo Cancer Research vol 58 no 23 pp 5507ndash5513 1998
[19] S Gupta and A A Knowlton ldquoHSP60 Bax apoptosis and theheartrdquo Journal of Cellular and Molecular Medicine vol 9 no 1pp 51ndash58 2005
[20] S Xanthoudakis R Sophie D Rasper et al ldquoHsp60 accel-erates the maturation of pro-caspase-3 by upstream activatorproteases during apoptosisrdquo The EMBO Journal vol 18 no 8pp 2049ndash2056 1999
[21] A Samali J Cai B Zhivotovsky D P Jones and S OrreniusldquoPresence of a pre-apoptotic complex of pro-caspase-3 Hsp60and Hsp10 in the mitochondrial fraction of Jurkat cellsrdquo TheEMBO Journal vol 18 no 8 pp 2040ndash2048 1999
[22] A Faried M Sohda M Nakajima TMiyazaki H Kato andHKuwano ldquoExpression of heat-shock protein Hsp60 correlatedwith the apoptotic index and patient prognosis in humanoesophageal squamous cell carcinomardquo European Journal ofCancer vol 40 no 18 pp 2804ndash2811 2004
[23] J Schneider E Jimenez K Marenbach H Romero D Marxand H Meden ldquoImmunohistochemical detection of HSP60-expression in human ovarian cancer Correlation with survivalin a series of 247 patientsrdquoAnticancer Research vol 19 no 3 pp2141ndash2146 1999
[24] T Lebret R W G Watson V Molinie et al ldquoHeat shockproteins HSP27 HSP60 HSP70 and HSP90 expression inbladder carcinomardquo Cancer vol 98 no 5 pp 970ndash977 2003
[25] A Samali and S Orrenius ldquoHeat shock proteins regulators ofstress response and apoptosisrdquo Cell Stress and Chaperones vol3 no 4 pp 228ndash236 1998
[26] W Van Eden G Wick S Albani and I Cohen ldquoStress heatshock proteins and autoimmunity how immune responses to
heat shock proteins are to be used for the control of chronicinflammatory diseasesrdquo Annals of the New York Academy ofSciences vol 1113 pp 217ndash237 2007
[27] C Habich and V Burkart ldquoHeat shock protein 60 regulatoryrole on innate immune cellsrdquo Cellular and Molecular LifeSciences vol 64 no 6 pp 742ndash751 2007
[28] Y Kondo Y Ueno K Kobayashi et al ldquoHepatitis B virus repli-cation could enhance regulatory T cell activity by producingsoluble heat shock protein 60 from hepatocytesrdquoThe Journal ofInfectious Diseases vol 202 no 2 pp 202ndash213 2010
[29] C Colotti G Cavallini R L Vitale et al ldquoEffects of agingand anti-aging caloric restrictions on carbonyl and heat shockprotein levels and expressionrdquo Biogerontology vol 6 no 6 pp397ndash406 2005
[30] P L Hooper ldquoInsulin signaling GSK-3 heat shock proteins andthe natural history of type 2 diabetes mellitus a hypothesisrdquoMetabolic Syndrome and RelatedDisorders vol 5 no 3 pp 220ndash230 2007
[31] O S Birk D C Douek D Elias et al ldquoA role of Hsp60 inautoimmune diabetes analysis in a transgenic modelrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 93 no 3 pp 1032ndash1037 1996
[32] J N Chun B Choi KW Lee et al ldquoCytosolicHsp60 is involvedin the NF-kappaB-dependent survival of cancer cells via IKKregulatiordquo PLoS ONE vol 5 no 3 Article ID e9422 2010
[33] M-R Chen S-W Liu T-C Wu et al ldquoRU486-induciblerecombination in the salivary glands of lactoferrin promoter-driven green fluorescent Cre transgenic micerdquo Genesis vol 48no 10 pp 585ndash595 2010
[34] H-Y Lin C-H Kao KM-C Lin V Kaartinen and L-T YangldquoNotch signaling regulates late-stage epidermal differentiationandmaintains postnatal hair cycle homeostasisrdquoPLoSONE vol6 no 1 Article ID e15842 2011
[35] A C Kayani G L Close W H Dillmann R Mestril M JJackson and A McArdle ldquoOverexpression of HSP10 in skeletalmuscle of transgenic mice prevents the age-related fall inmaximum tetanic force generation and muscle Cross-SectionalAreardquoThe American Journal of PhysiologymdashRegulatory Integra-tive and Comparative Physiology vol 299 no 1 pp R268ndashR2762010
[36] G-C Fan X Ren J Qian et al ldquoNovel cardioprotective role of asmall heat-shock protein Hsp20 against ischemiareperfusioninjuryrdquo Circulation vol 111 no 14 pp 1792ndash1799 2005
[37] J M Hollander J L Martin D D Belke et al ldquoOverex-pression of wild-type heat shock protein 27 and a nonphos-phorylatable heat shock protein 27 mutant protects againstischemiareperfusion injury in a transgenic mouse modelrdquoCirculation vol 110 no 23 pp 3544ndash3552 2004
[38] J-C L Plumier A M Krueger R W Currie D KontoyiannisG Kollias and G N Pagoulatos ldquoTransgenic mice expressingthe human inducible Hsp70 have hippocampal neurons resis-tant to ischemic injuryrdquo Cell Stress and Chaperones vol 2 no 3pp 162ndash167 1997
[39] D Chandra G Choy and D G Tang ldquoCytosolic accumulationof HSP60 during apoptosis with or without apparentmitochon-drial release Evidence that its pro-apoptotic or pro-survivalfunctions involve differential interactions with caspase-3rdquo TheJournal of Biological Chemistry vol 282 no 43 pp 31289ndash313012007
BioMed Research International 11
[40] J H Jang J S Jung J I Choi and S K Kang ldquoNuclearAgo2HSP60 contributes to broad spectrum of hATSCs func-tion via Oct4 regulationrdquoAntioxidants and Redox Signaling vol16 no 5 pp 383ndash399 2012
[41] Q Feng W Song X Lu et al ldquoDevelopment of heart failureand congenital septal defects in mice lacking endothelial nitricoxide synthaserdquo Circulation vol 106 no 7 pp 873ndash879 2002
phosphorylation of IKK120572120573 and promotes NF-120581B prosur-vival pathway Transgenic expression of the cytosolic HSP60prevented stress-induced hepatic cell deaths in vivo [32]Overexpressing HSP60 in the human stem cells increasedHSP60 nuclear localization which suppressed ROS andp38JNK signaling and regulated genes relating to stem cellproliferation differentiation and stemness [40]Whether theaforementioned mechanisms are also involved in mediatingneonatal deaths remains unclear and future clarification isawaited
In this study congenital ASD was observed in a largenumber ofHSP60mice AlthoughASD exacerbated the exist-ing myopathy of HSP60 mice and was partially responsiblefor neonatal death ASD per sewas unlikely directly caused byHSP60 induction We speculate that ASD in neonatal HSP60mice may be the consequence of incomplete embryonicdevelopment existing myopathy and increasing apoptosisSimilar phenomenon has been demonstrated in the eNOSknockout mouse model in which high incidence of ASDor VSD was precipitated through increased apoptosis duringembryonic and postnatal stages [41] Our main findingsindicate that severe hemorrhage andmyocyte death occurredin mice with HSP60 expression and increased apoptosisin a macerated and spongy-like heart failed to meet thecardiovascular demands after birth and was the primarycause of HSP60-induced heart failure and neonatal death
4 Conclusions
HSP60 not only is the essentialmitochondrial chaperone pro-tein but also plays critical roles outside the mitochondrionAlthough numerous studies indicated the wide involvementof HSP60 in broad biological processes and many diseasesreports of transgenic mouse models of increasing or knock-down HSP60 expression are rare Thus the consequences ofaltering HSP60 levels in vivo in these disease settings remainunknown We used a floxed vector to establish conditionalHSP60 transgenic mice which allows studies on inducibleand organ-targeted HSP60 expression in adult mice and ispotentially useful in pursuing essential questions related toHSP60 in vivo In this study we show that ubiquitous HSP60expression from the embryonic stage results in increasedapoptosis myopathy high incidence of ASD and neonataldeaths
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
Tsung-Hsien Chen and Shan-Wen Liu contribute equally tothis paper
Acknowledgments
The authors gratefully acknowledge the technical assistanceprovided by Tzu-Chin Wu and Mei-Yu Fan in NHRI-IBENand Functional and Micro-Magnetic Resonance ImagingCenter supported by NRPGM (NSC97-3112-B-001-013) Thisresearch was supported by grants from NHRI (BN-104-PP-02 BN-104-PP-24) and from Ministry of Science andTechnology (103-2320-B-400-018)
References
[1] I J Benjamin andD RMcMillan ldquoStress (heat shock) proteinsmolecular chaperones in cardiovascular biology and diseaserdquoCirculation Research vol 83 no 2 pp 117ndash132 1998
[2] F U Hartl ldquoMolecular chaperones in cellular protein foldingrdquoNature vol 381 no 6583 pp 571ndash580 1996
[3] J C Ranford and B Henderson ldquoChaperonins in diseasemechanismsmodels and treatmentsrdquoMolecular Pathology vol55 no 4 pp 209ndash213 2002
[4] CCDeocaris S CKaul andRWadhwa ldquoOn the brotherhoodof the mitochondrial chaperones mortalin and heat shockprotein 60rdquo Cell Stress and Chaperones vol 11 no 2 pp 116ndash128 2006
[5] F Cappello A M Gammazza A P Piccionello et al ldquoHsp60chaperonopathies and chaperonotherapy targets and agentsrdquoExpert Opinion on Therapeutic Targets vol 18 no 2 pp 185ndash208 2014
[6] S Lau N Patnaik M R Sayen and R Mestril ldquoSimultaneousoverexpression of two stress proteins in rat cardiomyocytes andmyogenic cells confers protection against ischemia-inducedinjuryrdquo Circulation vol 96 no 7 pp 2287ndash2294 1997
[7] K M Lin B Lin I Y Lian R Mestril I E Scheffler and WH Dillmann ldquoCombined and individual mitochondrial HSP60and HSP10 expression in cardiac myocytes protects mitochon-drial function and prevents apoptotic cell deaths induced bysimulated ischemia-reoxygenationrdquo Circulation vol 103 no 13pp 1787ndash1792 2001
[8] J J Malago J F J G Koninkx and J E van Dijk ldquoThe heatshock response and cytoprotection of the intestinal epitheliumrdquoCell Stress and Chaperones vol 7 no 2 pp 191ndash199 2002
[9] Y-X Shan T-L Yang R Mestril and P H Wang ldquoHsp10 andHsp60 suppress ubiquitination of insulin-like growth factor-1receptor and augment insulin-like growth factor-1 receptor sig-naling in cardiac muscle implications on decreased myocardialprotection in diabetic cardiomyopathyrdquoThe Journal of BiologicalChemistry vol 278 no 46 pp 45492ndash45498 2003
[10] H-S Chen Y-X Shan T-L Yang et al ldquoInsulin deficiencydownregulated heat shock protein 60 and IGF-1 receptorsignaling in diabetic myocardiumrdquo Diabetes vol 54 no 1 pp175ndash181 2005
10 BioMed Research International
[11] V Calabrese A Ravagna C Colombrita et al ldquoAcetylcarnitineinduces heme oxygenase in rat astrocytes and protects againstoxidative stress Involvement of the transcription factor Nrf2rdquoJournal of Neuroscience Research vol 79 no 4 pp 509ndash5212005
[12] V Veereshwarayya P Kumar K M Rosen R Mestril andH W Querfurth ldquoDifferential effects of mitochondrial heatshock protein 60 and related molecular chaperones to preventintracellular 120573-amyloid-induced inhibition of complex IV andlimit apoptosisrdquoThe Journal of Biological Chemistry vol 281 no40 pp 29468ndash29478 2006
[13] C Xu Y Lu Z Pan et al ldquoThe muscle-specific microRNAsmiR-1 and miR-133 produce opposing effects on apoptosis bytargeting HSP60 HSP70 and caspase-9 in cardiomyocytesrdquoJournal of Cell Science vol 120 part 17 pp 3045ndash3052 2007
[14] J C Ghosh T Dohi H K Byoung and D C Altieri ldquoHsp60regulation of tumor cell apoptosisrdquo The Journal of BiologicalChemistry vol 283 no 8 pp 5188ndash5194 2008
[15] P E Castle R Ashfaq F Ansari and C Y Muller ldquoImmuno-histochemical evaluation of heat shock proteins in normal andpreinvasive lesions of the cervixrdquo Cancer Letters vol 229 no 2pp 245ndash252 2005
[16] P A Cornford A R Dodson K F Parsons et al ldquoHeat shockprotein expression independently predicts clinical outcome inprostate cancerrdquoCancer Research vol 60 no 24 pp 7099ndash71052000
[17] F Cappello S David F Rappa et al ldquoThe expression of Hsp60and Hsp10 in large bowel carcinomas with lymph node metas-tasesrdquo BMC Cancer vol 5 article 39 2005
[18] P-L Hsu and S-M Hsu ldquoAbundance of heat shock proteins(hsp89 hsp60 and hsp27) in malignant cells of Hodgkinrsquosdiseaserdquo Cancer Research vol 58 no 23 pp 5507ndash5513 1998
[19] S Gupta and A A Knowlton ldquoHSP60 Bax apoptosis and theheartrdquo Journal of Cellular and Molecular Medicine vol 9 no 1pp 51ndash58 2005
[20] S Xanthoudakis R Sophie D Rasper et al ldquoHsp60 accel-erates the maturation of pro-caspase-3 by upstream activatorproteases during apoptosisrdquo The EMBO Journal vol 18 no 8pp 2049ndash2056 1999
[21] A Samali J Cai B Zhivotovsky D P Jones and S OrreniusldquoPresence of a pre-apoptotic complex of pro-caspase-3 Hsp60and Hsp10 in the mitochondrial fraction of Jurkat cellsrdquo TheEMBO Journal vol 18 no 8 pp 2040ndash2048 1999
[22] A Faried M Sohda M Nakajima TMiyazaki H Kato andHKuwano ldquoExpression of heat-shock protein Hsp60 correlatedwith the apoptotic index and patient prognosis in humanoesophageal squamous cell carcinomardquo European Journal ofCancer vol 40 no 18 pp 2804ndash2811 2004
[23] J Schneider E Jimenez K Marenbach H Romero D Marxand H Meden ldquoImmunohistochemical detection of HSP60-expression in human ovarian cancer Correlation with survivalin a series of 247 patientsrdquoAnticancer Research vol 19 no 3 pp2141ndash2146 1999
[24] T Lebret R W G Watson V Molinie et al ldquoHeat shockproteins HSP27 HSP60 HSP70 and HSP90 expression inbladder carcinomardquo Cancer vol 98 no 5 pp 970ndash977 2003
[25] A Samali and S Orrenius ldquoHeat shock proteins regulators ofstress response and apoptosisrdquo Cell Stress and Chaperones vol3 no 4 pp 228ndash236 1998
[26] W Van Eden G Wick S Albani and I Cohen ldquoStress heatshock proteins and autoimmunity how immune responses to
heat shock proteins are to be used for the control of chronicinflammatory diseasesrdquo Annals of the New York Academy ofSciences vol 1113 pp 217ndash237 2007
[27] C Habich and V Burkart ldquoHeat shock protein 60 regulatoryrole on innate immune cellsrdquo Cellular and Molecular LifeSciences vol 64 no 6 pp 742ndash751 2007
[28] Y Kondo Y Ueno K Kobayashi et al ldquoHepatitis B virus repli-cation could enhance regulatory T cell activity by producingsoluble heat shock protein 60 from hepatocytesrdquoThe Journal ofInfectious Diseases vol 202 no 2 pp 202ndash213 2010
[29] C Colotti G Cavallini R L Vitale et al ldquoEffects of agingand anti-aging caloric restrictions on carbonyl and heat shockprotein levels and expressionrdquo Biogerontology vol 6 no 6 pp397ndash406 2005
[30] P L Hooper ldquoInsulin signaling GSK-3 heat shock proteins andthe natural history of type 2 diabetes mellitus a hypothesisrdquoMetabolic Syndrome and RelatedDisorders vol 5 no 3 pp 220ndash230 2007
[31] O S Birk D C Douek D Elias et al ldquoA role of Hsp60 inautoimmune diabetes analysis in a transgenic modelrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 93 no 3 pp 1032ndash1037 1996
[32] J N Chun B Choi KW Lee et al ldquoCytosolicHsp60 is involvedin the NF-kappaB-dependent survival of cancer cells via IKKregulatiordquo PLoS ONE vol 5 no 3 Article ID e9422 2010
[33] M-R Chen S-W Liu T-C Wu et al ldquoRU486-induciblerecombination in the salivary glands of lactoferrin promoter-driven green fluorescent Cre transgenic micerdquo Genesis vol 48no 10 pp 585ndash595 2010
[34] H-Y Lin C-H Kao KM-C Lin V Kaartinen and L-T YangldquoNotch signaling regulates late-stage epidermal differentiationandmaintains postnatal hair cycle homeostasisrdquoPLoSONE vol6 no 1 Article ID e15842 2011
[35] A C Kayani G L Close W H Dillmann R Mestril M JJackson and A McArdle ldquoOverexpression of HSP10 in skeletalmuscle of transgenic mice prevents the age-related fall inmaximum tetanic force generation and muscle Cross-SectionalAreardquoThe American Journal of PhysiologymdashRegulatory Integra-tive and Comparative Physiology vol 299 no 1 pp R268ndashR2762010
[36] G-C Fan X Ren J Qian et al ldquoNovel cardioprotective role of asmall heat-shock protein Hsp20 against ischemiareperfusioninjuryrdquo Circulation vol 111 no 14 pp 1792ndash1799 2005
[37] J M Hollander J L Martin D D Belke et al ldquoOverex-pression of wild-type heat shock protein 27 and a nonphos-phorylatable heat shock protein 27 mutant protects againstischemiareperfusion injury in a transgenic mouse modelrdquoCirculation vol 110 no 23 pp 3544ndash3552 2004
[38] J-C L Plumier A M Krueger R W Currie D KontoyiannisG Kollias and G N Pagoulatos ldquoTransgenic mice expressingthe human inducible Hsp70 have hippocampal neurons resis-tant to ischemic injuryrdquo Cell Stress and Chaperones vol 2 no 3pp 162ndash167 1997
[39] D Chandra G Choy and D G Tang ldquoCytosolic accumulationof HSP60 during apoptosis with or without apparentmitochon-drial release Evidence that its pro-apoptotic or pro-survivalfunctions involve differential interactions with caspase-3rdquo TheJournal of Biological Chemistry vol 282 no 43 pp 31289ndash313012007
BioMed Research International 11
[40] J H Jang J S Jung J I Choi and S K Kang ldquoNuclearAgo2HSP60 contributes to broad spectrum of hATSCs func-tion via Oct4 regulationrdquoAntioxidants and Redox Signaling vol16 no 5 pp 383ndash399 2012
[41] Q Feng W Song X Lu et al ldquoDevelopment of heart failureand congenital septal defects in mice lacking endothelial nitricoxide synthaserdquo Circulation vol 106 no 7 pp 873ndash879 2002
[11] V Calabrese A Ravagna C Colombrita et al ldquoAcetylcarnitineinduces heme oxygenase in rat astrocytes and protects againstoxidative stress Involvement of the transcription factor Nrf2rdquoJournal of Neuroscience Research vol 79 no 4 pp 509ndash5212005
[12] V Veereshwarayya P Kumar K M Rosen R Mestril andH W Querfurth ldquoDifferential effects of mitochondrial heatshock protein 60 and related molecular chaperones to preventintracellular 120573-amyloid-induced inhibition of complex IV andlimit apoptosisrdquoThe Journal of Biological Chemistry vol 281 no40 pp 29468ndash29478 2006
[13] C Xu Y Lu Z Pan et al ldquoThe muscle-specific microRNAsmiR-1 and miR-133 produce opposing effects on apoptosis bytargeting HSP60 HSP70 and caspase-9 in cardiomyocytesrdquoJournal of Cell Science vol 120 part 17 pp 3045ndash3052 2007
[14] J C Ghosh T Dohi H K Byoung and D C Altieri ldquoHsp60regulation of tumor cell apoptosisrdquo The Journal of BiologicalChemistry vol 283 no 8 pp 5188ndash5194 2008
[15] P E Castle R Ashfaq F Ansari and C Y Muller ldquoImmuno-histochemical evaluation of heat shock proteins in normal andpreinvasive lesions of the cervixrdquo Cancer Letters vol 229 no 2pp 245ndash252 2005
[16] P A Cornford A R Dodson K F Parsons et al ldquoHeat shockprotein expression independently predicts clinical outcome inprostate cancerrdquoCancer Research vol 60 no 24 pp 7099ndash71052000
[17] F Cappello S David F Rappa et al ldquoThe expression of Hsp60and Hsp10 in large bowel carcinomas with lymph node metas-tasesrdquo BMC Cancer vol 5 article 39 2005
[18] P-L Hsu and S-M Hsu ldquoAbundance of heat shock proteins(hsp89 hsp60 and hsp27) in malignant cells of Hodgkinrsquosdiseaserdquo Cancer Research vol 58 no 23 pp 5507ndash5513 1998
[19] S Gupta and A A Knowlton ldquoHSP60 Bax apoptosis and theheartrdquo Journal of Cellular and Molecular Medicine vol 9 no 1pp 51ndash58 2005
[20] S Xanthoudakis R Sophie D Rasper et al ldquoHsp60 accel-erates the maturation of pro-caspase-3 by upstream activatorproteases during apoptosisrdquo The EMBO Journal vol 18 no 8pp 2049ndash2056 1999
[21] A Samali J Cai B Zhivotovsky D P Jones and S OrreniusldquoPresence of a pre-apoptotic complex of pro-caspase-3 Hsp60and Hsp10 in the mitochondrial fraction of Jurkat cellsrdquo TheEMBO Journal vol 18 no 8 pp 2040ndash2048 1999
[22] A Faried M Sohda M Nakajima TMiyazaki H Kato andHKuwano ldquoExpression of heat-shock protein Hsp60 correlatedwith the apoptotic index and patient prognosis in humanoesophageal squamous cell carcinomardquo European Journal ofCancer vol 40 no 18 pp 2804ndash2811 2004
[23] J Schneider E Jimenez K Marenbach H Romero D Marxand H Meden ldquoImmunohistochemical detection of HSP60-expression in human ovarian cancer Correlation with survivalin a series of 247 patientsrdquoAnticancer Research vol 19 no 3 pp2141ndash2146 1999
[24] T Lebret R W G Watson V Molinie et al ldquoHeat shockproteins HSP27 HSP60 HSP70 and HSP90 expression inbladder carcinomardquo Cancer vol 98 no 5 pp 970ndash977 2003
[25] A Samali and S Orrenius ldquoHeat shock proteins regulators ofstress response and apoptosisrdquo Cell Stress and Chaperones vol3 no 4 pp 228ndash236 1998
[26] W Van Eden G Wick S Albani and I Cohen ldquoStress heatshock proteins and autoimmunity how immune responses to
heat shock proteins are to be used for the control of chronicinflammatory diseasesrdquo Annals of the New York Academy ofSciences vol 1113 pp 217ndash237 2007
[27] C Habich and V Burkart ldquoHeat shock protein 60 regulatoryrole on innate immune cellsrdquo Cellular and Molecular LifeSciences vol 64 no 6 pp 742ndash751 2007
[28] Y Kondo Y Ueno K Kobayashi et al ldquoHepatitis B virus repli-cation could enhance regulatory T cell activity by producingsoluble heat shock protein 60 from hepatocytesrdquoThe Journal ofInfectious Diseases vol 202 no 2 pp 202ndash213 2010
[29] C Colotti G Cavallini R L Vitale et al ldquoEffects of agingand anti-aging caloric restrictions on carbonyl and heat shockprotein levels and expressionrdquo Biogerontology vol 6 no 6 pp397ndash406 2005
[30] P L Hooper ldquoInsulin signaling GSK-3 heat shock proteins andthe natural history of type 2 diabetes mellitus a hypothesisrdquoMetabolic Syndrome and RelatedDisorders vol 5 no 3 pp 220ndash230 2007
[31] O S Birk D C Douek D Elias et al ldquoA role of Hsp60 inautoimmune diabetes analysis in a transgenic modelrdquo Proceed-ings of the National Academy of Sciences of the United States ofAmerica vol 93 no 3 pp 1032ndash1037 1996
[32] J N Chun B Choi KW Lee et al ldquoCytosolicHsp60 is involvedin the NF-kappaB-dependent survival of cancer cells via IKKregulatiordquo PLoS ONE vol 5 no 3 Article ID e9422 2010
[33] M-R Chen S-W Liu T-C Wu et al ldquoRU486-induciblerecombination in the salivary glands of lactoferrin promoter-driven green fluorescent Cre transgenic micerdquo Genesis vol 48no 10 pp 585ndash595 2010
[34] H-Y Lin C-H Kao KM-C Lin V Kaartinen and L-T YangldquoNotch signaling regulates late-stage epidermal differentiationandmaintains postnatal hair cycle homeostasisrdquoPLoSONE vol6 no 1 Article ID e15842 2011
[35] A C Kayani G L Close W H Dillmann R Mestril M JJackson and A McArdle ldquoOverexpression of HSP10 in skeletalmuscle of transgenic mice prevents the age-related fall inmaximum tetanic force generation and muscle Cross-SectionalAreardquoThe American Journal of PhysiologymdashRegulatory Integra-tive and Comparative Physiology vol 299 no 1 pp R268ndashR2762010
[36] G-C Fan X Ren J Qian et al ldquoNovel cardioprotective role of asmall heat-shock protein Hsp20 against ischemiareperfusioninjuryrdquo Circulation vol 111 no 14 pp 1792ndash1799 2005
[37] J M Hollander J L Martin D D Belke et al ldquoOverex-pression of wild-type heat shock protein 27 and a nonphos-phorylatable heat shock protein 27 mutant protects againstischemiareperfusion injury in a transgenic mouse modelrdquoCirculation vol 110 no 23 pp 3544ndash3552 2004
[38] J-C L Plumier A M Krueger R W Currie D KontoyiannisG Kollias and G N Pagoulatos ldquoTransgenic mice expressingthe human inducible Hsp70 have hippocampal neurons resis-tant to ischemic injuryrdquo Cell Stress and Chaperones vol 2 no 3pp 162ndash167 1997
[39] D Chandra G Choy and D G Tang ldquoCytosolic accumulationof HSP60 during apoptosis with or without apparentmitochon-drial release Evidence that its pro-apoptotic or pro-survivalfunctions involve differential interactions with caspase-3rdquo TheJournal of Biological Chemistry vol 282 no 43 pp 31289ndash313012007
BioMed Research International 11
[40] J H Jang J S Jung J I Choi and S K Kang ldquoNuclearAgo2HSP60 contributes to broad spectrum of hATSCs func-tion via Oct4 regulationrdquoAntioxidants and Redox Signaling vol16 no 5 pp 383ndash399 2012
[41] Q Feng W Song X Lu et al ldquoDevelopment of heart failureand congenital septal defects in mice lacking endothelial nitricoxide synthaserdquo Circulation vol 106 no 7 pp 873ndash879 2002
[40] J H Jang J S Jung J I Choi and S K Kang ldquoNuclearAgo2HSP60 contributes to broad spectrum of hATSCs func-tion via Oct4 regulationrdquoAntioxidants and Redox Signaling vol16 no 5 pp 383ndash399 2012
[41] Q Feng W Song X Lu et al ldquoDevelopment of heart failureand congenital septal defects in mice lacking endothelial nitricoxide synthaserdquo Circulation vol 106 no 7 pp 873ndash879 2002