The Effect of Habitat Acoustics on Common Marmoset Vocal SignalTransmission
RYAN J. MORRILL1, A. WREN THOMAS1,2, NICOLA SCHIEL3, ANTONIO SOUTO4, AND CORY T. MILLER1,5*1Cortical Systems and Behavior Laboratory, Department of Psychology, University of California, San Diego, La Jolla,California2Hellen Wills Neuroscience Graduate Program, University of California, Berkeley, Berkeley, California3Laboratory of Applied and Theoretical Ethology, Department of Biology, Federal Rural University of Pernambuco, Recife,Brazil4Laboratory of Ethology, Department of Zoology, Federal University of Pernambuco, Recife, Brazil5Neurosciences Graduate Program, University of California, San Diego, La Jolla, California
INTRODUCTIONAnimal vocalizations evolved in response to a
myriad of selection pressures. These include con-straints imposed on signal structure by the species’vocal‐motor and auditory systems, as well asenvironmental constraints on effective exchangeof communicative information [Bradbury &Vehrencamp, 2011]. In noisy acoustic environments,interference fromboth biotic and abiotic sources, suchas the density of foliage, forest structure, humidityand heterospecific acoustic signals can distort andablate the spectro‐temporal structure of vocal signals[Morton, 1975; Wiley & Richards, 1978, 1982], there-by degrading the communicative content of vocalsignals. There are at least two dimensions alongwhich selection may act in order to improve commu-
nication in these environments. One is selection forauditory feedback‐mediated vocal‐motor control thatallows callers to monitor the acoustic environmentand initiate vocalizations during periods of lowacoustic interference [Roy et al., 2011; Waser &
Contract grant sponsor: NIH; contract grant numbers: K99/R00DC009007, R01 DC012087; contract grant sponsor: UCSDAcademic Senate Grant.�Correspondence to: Cory Miller, University of California, SanDiego, 9500 Gilman Dr. No. 0109, La Jolla, CA 92093. E‐mail:[email protected]
Received 30 October 2012; revised 1 March 2013; revisionaccepted 5 March 2013
DOI: 10.1002/ajp.22152Published online 16 April 2013 in Wiley Online Library(wileyonlinelibrary.com).
Brown, 1984]. Another is selection for acousticadaptations in the signal structure. Vocalizationsmay comprise acoustic characteristics that result indecreased interference by ambient sound or other-wise resist degradation in particular ecosystems[Waser & Brown, 1984, 1986; Waser & Waser,1977; Wiley & Richards, 1982]. Ultimately, a species’vocal communication system is likely to have evolvedboth behavioral and acoustic adaptations for improv-ing communicative efficacy to varying degrees. Theexact relationship between the two, however, is likelyspecies‐specific. The current study sought to comple-ment ongoing behavioral experiments investigatingcommon marmoset (Callithrix jacchus) feedback‐mediated vocal control [Roy et al., 2011] with acomprehensive characterization of the species’ habi-tat acoustics and vocal signal transmission. Broadlydefined, our aim was to better understand thepotential evolutionary and extant strategies em-ployed by marmosets to maintain effective communi-cation in their natural habitat.
The quantification of species’ habitat acousticshas occurred for several decades, with a wealth ofdata available [Brown et al., 1995; De la Torre &Snowdon, 2002; Kime et al., 2000; Marten & Marler,1977; Marten et al., 1977; Zimmerman, 1983]. Thesestudies have traditionally sought to characterize theacoustic landscape in which communication occurs,and, by comparison with extant species‐typicalvocalizations, determine the plausibility of selectionfor an acoustic structure that improves signaltransmission in this environment. Waser and Brown[1984] speculated that selection would favor nonhu-man primate long‐distance vocalizations that oc-curred within a specific spectral “sound window.” Inotherwords, long‐distance vocalizations comprising afundamental frequency (F0) in a particular frequencyrange may avoid acoustic interference from otherspecies, such as birds, thereby increasing transmis-sion efficacy. Alternatively, signalers could avoidacoustic interference by taking advantage of atemporal “sound window.” Callers could monitorthe ongoing acoustic events in the forest and controlthe timing and occurrence of vocal behaviors toinitiate during periods with little interferencefrom heterospecific vocalizations. Several Old Worldmonkeys, for example, show increased vocalizationrates early in the morning when other species areless acoustically active [Brown et al., 1995; Waser& Brown, 1986; Waser & Waser, 1977]. Exploitingwindows in both the frequency and time domainscan improve communication in noisy acousticenvironments.
The common marmoset (C. jacchus) is a small(�200–400 g), arboreal NewWorld monkey, endemicto Northeastern Brazil. This species potentiallyrepresents a unique case study for communicatingin complex acoustic environments for at least thefollowing reasons. Despite living in noisy forest
habitats, the vocal repertoire of the species isrelatively high in frequency, with most calls havinga F0 of �6–9 kHz [Bezera & Souto, 2008; Milleret al., 2010; Pistorio et al., 2006]. This includes theirlong‐distance contact call, known as the “phee”[Miller et al., 2010; Norcross & Newman, 1993].The physics of sound transmission suggest thatsignals of at this frequency are likely to significantlydegrade in forest habitats [Morton, 1975; Wiley &Richards, 1978]. One potential reason for the highfrequency of these calls may be related to the species’small body size (�200–400 g). Indeed many Calli-trichids of similar body size produce vocalizations inthis frequency range. There are, however, exceptions.The closely related and similarly sized cotton‐top andGeoffroy’s tamarins (Saguinus oedipus, S. geoffroyi)produce a long‐distance contact call with a F0 of�1.5–1.8 kHz [Cleveland & Snowdon, 1982; Weisset al., 2001], well below the F0 of common marmosetcalls. Furthermore, the high frequency of themarmoset vocal repertoire presumably places themin direct acoustic conflict with many of the avian,insect, and anuran species inhabiting the sameforests, but direct evidence to this end is limited. Asingle study of pygmy marmosets (Cebuella pyg-maea), another Callitrichid species that produceshigh frequency vocalizations [Elowson & Snowdon,1994; Pola & Snowdon, 1975], provided evidence ofsignificant acoustic competition from the naturalhabitat acoustics within the species‐typical vocalrange [De la Torre & Snowdon, 2002]. Comparablehabitat acoustic data are not available for otherCallitrichids. Field data on common marmosets haveindicated little change in calling rate over the courseof a day [Bezerra et al., 2009], suggesting thatmarmosets do not alter vocalization rate to takeadvantage of times of day with low acoustic interfer-ence. Nevertheless, other behavioral strategies oracoustic adaptations may serve to reduce acousticcompetition. More data are needed on the habitatacoustics of common marmosets in order to betterunderstand the scope of potential interference andthe presence of available spectral and temporal soundwindows.
The aim of this study was to characterize theeffects of habitat acoustics on vocal signal transmis-sion in common marmosets. The study consisted oftwo parts. First, we recorded the ambient acoustics ofa typical forest inhabited by the species in North-eastern Brazil during both the rainy and dry seasonsover a 2‐year period. The aim here was to quantita-tively characterize differences in biotic and abioticacoustic sources over different time intervals (i.e.,hours, seasons, and years). Given the strong seasonalvariability of this forest for precipitation and speciesdiversity [Santos et al., 2004] and density, as well asthe activity patterns of most vertebrates, we pre-dicted that significant differences in the ambientacoustics of the forest would be evident along
Am. J. Primatol.
Habitat Acoustics of Common Marmosets / 905
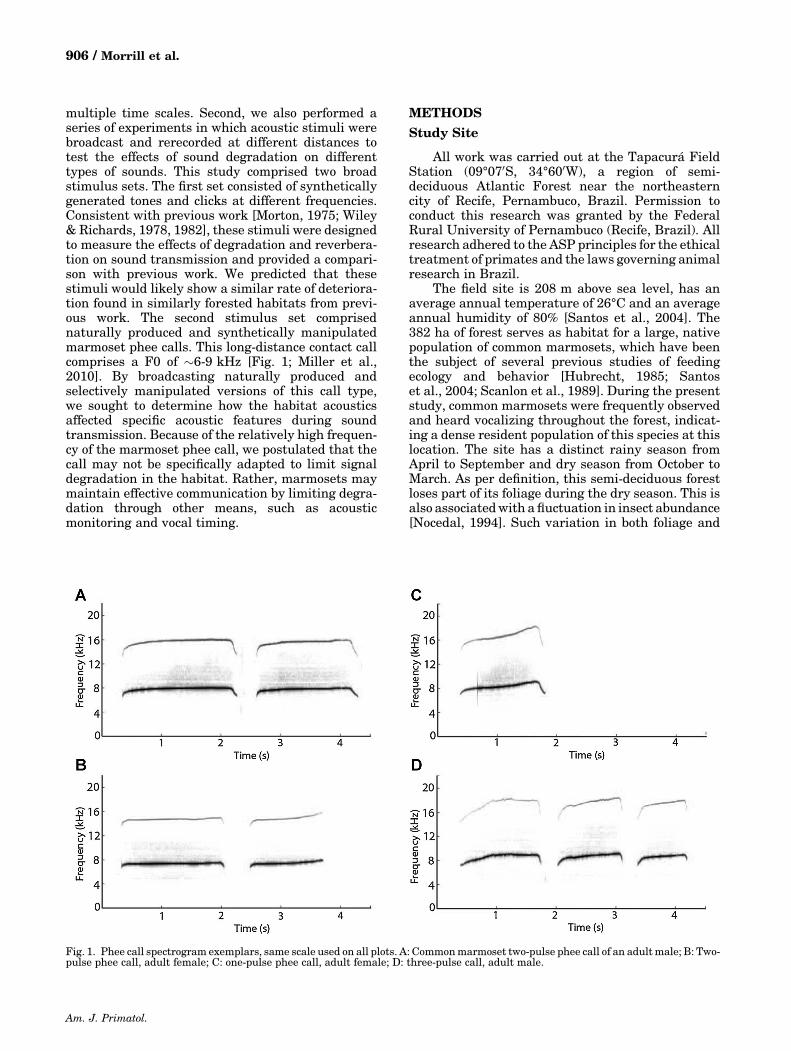
multiple time scales. Second, we also performed aseries of experiments in which acoustic stimuli werebroadcast and rerecorded at different distances totest the effects of sound degradation on differenttypes of sounds. This study comprised two broadstimulus sets. The first set consisted of syntheticallygenerated tones and clicks at different frequencies.Consistent with previous work [Morton, 1975; Wiley& Richards, 1978, 1982], these stimuli were designedto measure the effects of degradation and reverbera-tion on sound transmission and provided a compari-son with previous work. We predicted that thesestimuli would likely show a similar rate of deteriora-tion found in similarly forested habitats from previ-ous work. The second stimulus set comprisednaturally produced and synthetically manipulatedmarmoset phee calls. This long‐distance contact callcomprises a F0 of �6‐9 kHz [Fig. 1; Miller et al.,2010]. By broadcasting naturally produced andselectively manipulated versions of this call type,we sought to determine how the habitat acousticsaffected specific acoustic features during soundtransmission. Because of the relatively high frequen-cy of the marmoset phee call, we postulated that thecall may not be specifically adapted to limit signaldegradation in the habitat. Rather, marmosets maymaintain effective communication by limiting degra-dation through other means, such as acousticmonitoring and vocal timing.
METHODSStudy Site
All work was carried out at the Tapacurá FieldStation (09°070S, 34°600W), a region of semi‐deciduous Atlantic Forest near the northeasterncity of Recife, Pernambuco, Brazil. Permission toconduct this research was granted by the FederalRural University of Pernambuco (Recife, Brazil). Allresearch adhered to the ASP principles for the ethicaltreatment of primates and the laws governing animalresearch in Brazil.
The field site is 208 m above sea level, has anaverage annual temperature of 26°C and an averageannual humidity of 80% [Santos et al., 2004]. The382 ha of forest serves as habitat for a large, nativepopulation of common marmosets, which have beenthe subject of several previous studies of feedingecology and behavior [Hubrecht, 1985; Santoset al., 2004; Scanlon et al., 1989]. During the presentstudy, common marmosets were frequently observedand heard vocalizing throughout the forest, indicat-ing a dense resident population of this species at thislocation. The site has a distinct rainy season fromApril to September and dry season from October toMarch. As per definition, this semi‐deciduous forestloses part of its foliage during the dry season. This isalso associatedwith afluctuation in insect abundance[Nocedal, 1994]. Such variation in both foliage and
Fig. 1. Phee call spectrogram exemplars, same scale used on all plots. A: Commonmarmoset two‐pulse phee call of an adultmale; B: Two‐pulse phee call, adult female; C: one‐pulse phee call, adult female; D: three‐pulse call, adult male.
Am. J. Primatol.
906 / Morrill et al.
insect populations is expected to result in season‐specific habitat acoustic patterns. For this reason,four trips to the field site were made: twice duringthe wet season, in March 2010 and 2011, andtwice during the dry season, in late August–September 2010 and September 2011.
Ambient Recordings
Ambient recordings were performed throughoutthe day at 2 hr intervals from 0600 to 1800 hours atforested locations within the field site. Each ambientrecording was 10 min in length. Three full days ofrecordingswere gathered during each of the four tripsto the field site. Recordings weremade by an observerholding the microphone and recorder at arm heightwith the microphone pointed vertically. Great carewas taken to avoid recording human‐generatedsounds of any sort, and recording sessions wererestarted in the event of such sound interference.
Ambient recordings were made using a MarantzPMD660 digital recorder with a Sennheiser ME‐62omni‐directional condenser microphone (foam wind-screen attached) positioned vertically. All audio useda 44.1 kHz sample rate, stored in 16‐bit WAV formaton SD cards.
Ambient Acoustic Analysis
All analyses were performed in MATLAB v7.10and v7.11. To analyze changes in the frequencyspectrum throughout the day, we used a multi‐taperfast Fourier transform (FFT) method from theChronux MATLAB toolbox (www.chronux.org).Three sections of 1 min each were randomly sampledfrom the 10 min raw recordings, and the powerspectra were averaged to represent each recording.
To confirm similarities apparent the powerspectra, we used discriminant function analysis(DFA). This analysis builds a linear model frompast experience with a dataset, and then uses themodel to predict membership, based on a givenvariable, in naturally occurring data groupings.Here,DFA was used to determine if the spectral signatureof an ambient recording could predict the followingattributes of the recording: time of day, time of day byseason, season, season by year, and year. DFA wasperformed using algorithms from the MATLABStatistics Toolbox.
Sound BroadcastsTo investigate sound transmission properties of
the habitat, a set of sounds were broadcast and re‐recorded at three locations within the field site.Locations were chosen to represent possible varia-tions in vegetation density at the Tapucará FieldStation. Each location was marked using physicalmarkers andGPS during thefirst usage (March 2010)
and reused for the sound broadcasts during thesubsequent trips. Average vegetation density wasdetermined by employing the nearest neighbormethod, in which we measured the distance to thenearest neighbor of each tree (with at least 5 cmcircumference) at 1 m height, in two 5 m2 areas[Souto et al., 2007]. Broadcast location 1 had a hightree density of 3.7 trees/m2 and a high mean nearestneighbor distance of 86.2 cm. Location 2 had a lowerdensity of 2.3 trees/m2 and a nearest neighbordistance of 58.7 cm. Location 3 also had lower densityof 2.3 trees/m2 and a nearest neighbor distance of73.7 cm.
Broadcasts were made using a Roland Micro-Cube amplified speaker. The speaker was hung froma tree branch at a height of approximately 5 m, at alocation with no interfering trees or branchesimmediately in front of the speaker. The height ofthe speaker was positioned to be within the canopylayer at each location, as this is the level of the forestthat marmosets were observed to travel at thislocation. Before the beginning of each broadcastsession, the system was calibrated such that a 1 kHzsine wave tone measured 1 m distance from thespeaker registered 90 dB SPL on an Extech dual‐range sound level meter (Model 407732).
Recordings of broadcast sounds were made atfour distances from the sound source: 10, 20, 40, and80 m. All recording sites lay on an approximatelystraight line from the sound source, as confirmed byGPS mapping. To re‐record broadcast sounds, weused the Marantz PMD660 recorder and SennheiserME66 directional microphone (foam windscreenattached) aimed in the direction of the broadcastspeaker. Recordings were made using previouslymentioned recording settings.
Two playlists of sounds were broadcast. The firstplaylist, broadcast during the 2010 wet and dryseasons, consisted of the following components: (1)twenty‐five marmoset phee calls (recorded from acaptive colony atUniversity of California, SanDiego),(2) seven click train stimuli, each consisting ofthree 20‐cycle trains of 30 msec click and 70 msecsilence, with click F0s nearly logarithmically spacedat 125 Hz, 250 Hz, 500 Hz, 1 kHz, 2 kHz, 4 kHz,6 kHz, and 8 kHz, and (3) seven sine‐wavetones, 1 sec each, at the same frequencies as theclick trains.
The second playlist, broadcast only during the2010 dry season, consisted of acoustic manipulationsof marmoset phee calls. Manipulations were either inthe time domain—that is, calls were sped up orslowed down, whilemaintaining frequency—or in thefrequency domain—that is, the fundamental frequen-cy was shifted up or down, while maintaining theoriginal length of the call. This playlist consisted ofthe following components: (1) five unmodified mar-moset phee calls, (2) frequency manipulations ofthese five calls such that the mean F0 was at 250 Hz,
500 Hz, 1 kHz, 2 kHz, 4 kHz, 8 kHz, and 16 kHz,with harmonics scaled appropriately, and (3) timemanipulations of the five calls, such that frequencywas not altered, but call lengths were scaled byfactors of 0.2, 0.6, 0.8, 0.9, 1.1, 1.2, 1.4, and 1.8.
Analysis of Broadcast Sounds
Re‐recorded broadcast sounds were primarilyanalyzed using normalized cross‐correlation withthe original broadcast stimulus as the template.This analysis produces an array of correlationsas a function of time lag between the templatewaveform and the sample waveform. As such,the maximum cross‐correlation value, presumablylocated at the time lag where the originaltemplate stimulus is found in the recording, wasused for each broadcast sound, and all other valueswere discarded. Cross‐correlation values were com-pared using multivariate repeated measures AN-OVA, implemented with SPSS 20.0 software. Arepeated measures analysis was used because ofthe likelihood of dependency between correlationvalues at different distances during the samerecording session. As such, the within‐group variablewas distance from broadcast site (i.e., 10, 20, 40, or80 m). Between‐group variables analyzed were stim-ulus frequency, recording site, time‐of‐day, andseason. Wilks’ lambda was used to test for within‐group effects.
The click train is a signal with rapid fluctuationbetween acoustic energy and periods of silence. Thisallows for analysis of reverberative habitat effects asa function of both click frequency and distance. Tocalculate reverberation in the click train, first thererecorded signal was band‐pass filtered using anequiripple Parks‐McClellan FIR filter, pass bands�100 Hz from the click carrier frequency, designedwith MATLAB Signal Processing Toolbox. From thisfiltered signal, the ratio of the mean amplitudeduring the 70 msec period of post‐click silence to themean amplitude of the 30 msec click section was usedas a reverberation index [Naguib, 2003]. Thismeasurement indicates the amount of energy in thesilent sections that remains from the previous click asit reverberates. Thus, a reverberation index of 1.0indicates nomean amplitude difference between clickand nonclick sections. For each broadcast click train,the reverberation indices of all click‐silence cycleswere averaged to obtain a mean reverberation indexvalue.
RESULTS
Ambient AcousticsSpectral analysis of these data revealed trends in
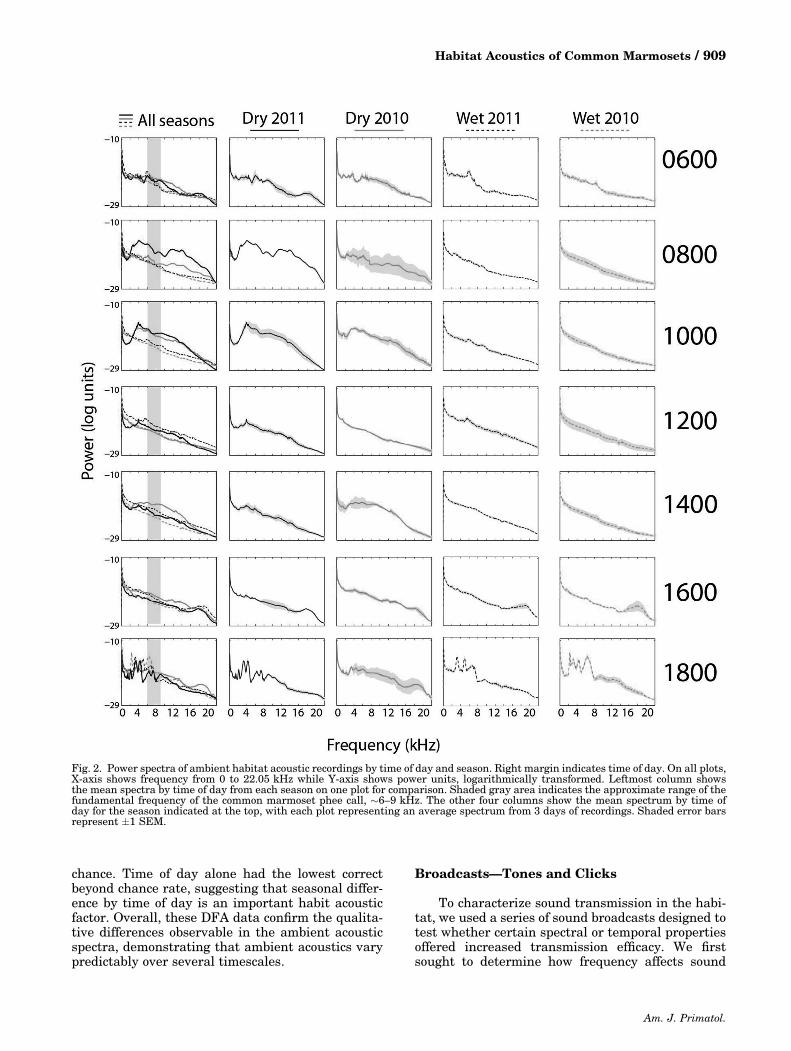
ambient noise by time of day and season (Fig. 2). Thedry seasonwas characterized by elevated biotic noise,
especially in the morning hours. The period from0600 to 1000 hours was marked by increased soundin a wide spectral band (2–16 kHz), likely due toinsect, bird, and anuran sounds in these morninghours. Noon (1200 hours) was relatively quiet,showing few peaks in sound frequency distribution,except for one near 4 kHz during the 2011 dry season.At 1400 hours in the 2010 dry season, soundattributable to insects such as crickets, locusts, orcicadas generated a peak between 2 and 14 kHz. Dueto decreased presence of these insects, this trend wasnot evident in the 2011 dry season, which onlyshowed a continued peak around 4 kHz. The lateafternoon (1600 hours) showed a relative lull inambient sound, although one high frequency soundaround 18 kHzwas consistently present. At the latestdaily time sampled, 1800 hours, there was a sharpincrease in 2–10 kHz sound due primarily to earlyevening anuran calling, as well as some insect‐generated sound.
The wet seasons were generally characterized byless overall biotic acoustic activity. This is mostevident during the morning periods when high levelsof vocalizations were typical in the dry season. In theearly morning (0600 hours), the 2011 wet seasonspectrum showed strong peak at 6 kHzwhile in 2010,a weaker peak was located near 8 kHz. Morningthrough midday (0800–1400 hours) was a period ofrelative quiet. At 1600 hours, the �18 kHz highfrequency sound, notable in dry season, was alsopresent in the wet season. Early evening(1600 hours) was marked by an increase in anuran‐and insect‐generated noise (2–8 kHz), which wasslightly decreased in bandwidthwhen compared withthe corresponding dry season recordings.
To explore the consistency of such trends, weused DFA, which uses a measured variable tocategorize members of a dataset into groups. Catego-rization is based on a linearmodel built from previousexperience with the data. In other words, DFA can beused to determine if a model can accurately catego-rize a dataset into its inherent categories. We usedDFA to determine the suitability of spectral signa-ture to classify each ambient acoustic recording bythe following categories: time of day, time of day byseason, season, season by year and year. Theaccuracy of the predictions shows whether the powerspectrum is a unique signature for categorization.Accuracy was calculated as the percentage of correctclassifications in a category minus the expectedcorrect categorization by chance. In the categoriza-tion of frequency spectra by season, for example, onewould expect 50% correct by chance, and thepercentage correct beyond this level is a measure ofthe suitability of spectral signature to classify in thiscategory. In all categories, DFA was able to classifycorrectly beyond chance (Fig. 3). Time of day byseason (e.g., “0600 hours wet season” vs. “0600 hoursdry season”) had the highest percentage correct above
Am. J. Primatol.
908 / Morrill et al.
chance. Time of day alone had the lowest correctbeyond chance rate, suggesting that seasonal differ-ence by time of day is an important habit acousticfactor. Overall, these DFA data confirm the qualita-tive differences observable in the ambient acousticspectra, demonstrating that ambient acoustics varypredictably over several timescales.
Broadcasts—Tones and Clicks
To characterize sound transmission in the habi-tat, we used a series of sound broadcasts designed totest whether certain spectral or temporal propertiesoffered increased transmission efficacy. We firstsought to determine how frequency affects sound
Fig. 2. Power spectra of ambient habitat acoustic recordings by time of day and season. Right margin indicates time of day. On all plots,X‐axis shows frequency from 0 to 22.05 kHz while Y‐axis shows power units, logarithmically transformed. Leftmost column showsthe mean spectra by time of day from each season on one plot for comparison. Shaded gray area indicates the approximate range of thefundamental frequency of the common marmoset phee call, �6–9 kHz. The other four columns show the mean spectrum by time ofday for the season indicated at the top, with each plot representing an average spectrum from 3 days of recordings. Shaded error barsrepresent �1 SEM.
Am. J. Primatol.
Habitat Acoustics of Common Marmosets / 909
transmission in the habitat by broadcasting a seriesof 1 sec sine wave tones at varied frequencies(250 Hz, 500 Hz, 1 kHz, 2 kHz, 4 kHz, 6 kHz, and8 kHz), and then testing correlation of rerecordedsounds with the original broadcasts. Generally, lowerfrequencies had a higher correlation coefficient thanhigher frequencies (Fig. 4), as verified by repeatedmeasures ANOVA (main effect of frequency, F[3,68] ¼ 6.31, P > 0.001). Post hoc analysis revealedthat lower frequency tones at 250 and 500 Hz differedsignificantly in correlation value from all others, butthose 1 kHz and higher did not differ significantly(Tukey’s HSD test, significance at P < 0.05 for alltests). The dry season showed higher mean correla-tion values (main effect of season, F[3, 1] ¼ 5.30,P ¼0.024) with more variability. Distance affectedcorrelation values during the dry season(F[3, 33] ¼ 6.26, P ¼ 0.001). Although 10 and 80 mwere not significantly different, 20 m was signifi-cantly higher than 40 m (Tukey’s test, P < 0.05).During the wet season, overall effect of distance wasnearly significant (F[3, 33] ¼ 2.54, P ¼ 0.073).
In addition to pure tones, we sought to examinewhether rapid amplitude fluctuations had an effecton transmission efficacy. We also broadcast clicktrains with pulses at varied fundamental frequen-
cies, with the same frequencies as used for tones.Clicks generally decreased in transmission efficacy asF0 increased (Fig. 4), which we again confirmedthrough repeated measures ANOVA (main effect offrequency, F[3, 6] ¼ 90.96, P < 0.001). Clicks at1 kHz and below differed significantly from all otherfrequencies, decreasing in correlation as frequencyincreased, while clicks 2 kHz and above did not differin correlation values from each other higher frequen-cies (Tukey’s test, P < 0.05). In the dry season, clicksdecreased in correlation over distance (F[3,33] ¼ 141.38, P < 0.001), and 10 m correlation val-ueswere significantly higher than 80 m (Tukey’s test,P < 0.05). A similar trend held for the wet season(F[3, 33] ¼ 92.32, P < 0.001), again with 10 mcorrelations significantly higher than 80 m (Tukey’stest, P < 0.05). Overall, variability of transmissionefficacy in the wet season was lower than in the dryseason. While the main effect of season was notsignificant (F[3, 1] ¼ 3.43, P ¼ 0.068), the interac-tion between season and distance was highly signifi-cant (F[3, 68] ¼ 20.54, P < 0.001).
Click train broadcasts were also analyzed forreverberation to determine how frequency anddistance from source affect this acoustic factor. As areverberation index, we measured the ratio of meansound pressure between click and silence sections ofeach click train broadcast. Reverberation indexpredictably increased with distance from sourceduring the wet and dry seasons together (Fig. 5;F[3, 68] ¼ 68.52, P < 0.001). Furthermore, reverber-ation index generally increased with increasingfrequency of click train during both seasons (maineffect of frequency, F[3, 6] ¼ 3.57, P ¼ 0.004). Clicksat 250 Hz did not differ significantly from 500 Hz,1 kHz, or 2 kHz, but differed significantly from allhigher frequencies. Likewise, the highest frequencyclicks, 4, 6, and 8 kHz, did not differ significantlyfrom each other (Tukey’s test, P < 0.05). Seasonaldifferences were not a significant factor for soundreverberation (main effect of season, F[3, 1] ¼ 1.822,P ¼ 0.181).
Broadcasts—Marmoset Phee Calls
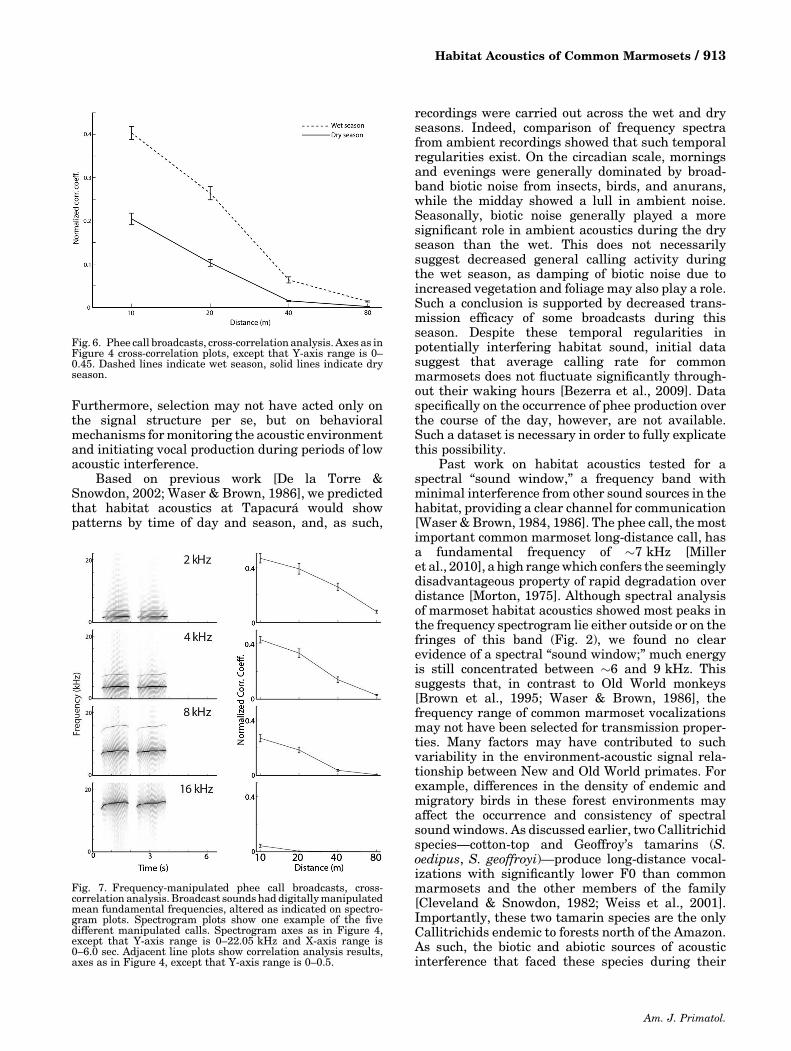
Recordings of naturally producedmarmoset pheecalls were broadcast during both wet and dry seasons(Fig. 6). For both seasons, cross‐correlation analysisindicated that phee call transmission efficacy degrad-ed predictably with increasing distance, as confirmedby repeated measures ANOVA (F[3, 248] ¼ 470.34,P < 0.001). Unlike tone and click stimuli, broadcastphee calls differed significantly at each distance fromall other distances (Tukey’s test, P < 0.05). General-ly, phee call correlations were less variable than theartificial sound broadcasts during both seasons. Pheecall broadcasts during the wet season showedsignificantly higher correlations than those in thedry season (main effect of season, F[3, 1] ¼ 161.59,
Fig. 3. Results of discriminant function analysis (DFA) inclassifying the mean spectral signature of each recordingcorrectly into various categorical variables, shown on the X‐axis. DFA is an analysis that shows the suitability of a givenvariable (here, power spectrum) in discriminating data intocategories. To compare results across categories of different size,plotted is the percentage correctly classified above the expectedcorrect classification by chance. For example, “season” representstwo groups, “wet” and “dry,” and correct classification, if left up tochance, would be 50%. DFA correctly classified 74% of samples byseason, so the 24% difference is plotted. Chance levels for eachgroup are indicated at the base of each bar.
Am. J. Primatol.
910 / Morrill et al.
P < 0.001). In addition, the interaction betweenseason and distance was significant (F[3, 248] ¼41.48, P < 0.001), suggesting that seasonal acousticdifferences may modulate the effects of distance oncall degradation.
Two characteristic properties of the phee call areits F0 and pulse length [Miller et al., 2010]. Todetermine whether these variables affect transmis-sion efficacy, we artificially manipulated frequency(while keeping length constant) or pulse length (whilekeeping frequency constant), and broadcast thesecalls using the same methods as tone and clickbroadcasts. As above, normalized cross‐correlationanalysis was carried out to determine transmissionefficacy. Due to equipment failure, this playlist wasonly broadcast during the 2010 dry season.
For frequency manipulations, we observed theexpected decrease in transmission efficacy as fre-quency increased (Fig. 7), again verified by repeatedmeasures ANOVA (main effect of frequency, F[3,3] ¼ 72.90,P < 0.001). Post hoc analysis showed thatall frequencies (2, 4, 8, and 16 kHz), differed in cross‐correlation values from all others (Tukey’s test,P < 0.05). Transmission efficacy also decreasedwith distance (F[3, 106] ¼ 240.49, P < 0.001), andthis pattern was significant at all distances (Tukey’stest, P < 0.05).
Betweenmanipulations of phee length, therewasno significant difference in transmission efficacy(Fig. 8; main effect of phee length, F[3, 7] ¼ 1.089,P ¼ 0.371). A consistent pattern of decreased corre-lation by distance was observed (F[3, 214] ¼ 675.82,
Fig. 4. Tone and click broadcasts, cross‐correlation analysis. Spectrograms are visualizations of original broadcast sounds, X‐axes showtime from 0 to 1.0 sec, and Y‐axes show frequency from 0 to 10 kHz. Columns to the right of spectrogram plots are correlations by distanceof adjacent broadcast sounds. Y‐axes of all plots show mean normalized correlations between original broadcast sounds and the re‐recorded sounds, where 0 would be uncorrelated and 1.0 would be perfect direct correlation. Actual Y‐axis scale is 0–0.55. Error barsindicate�1 SEM. Dashed lines indicate wet season, solid lines indicate dry season. X‐axes show measurement distance, logarithmicallyspaced at 10, 20, 40, and 80 m.
Am. J. Primatol.
Habitat Acoustics of Common Marmosets / 911
P < 0.001), which was significant at all distances(Tukey’s test, P < 0.05). For all phee manipulations(time and frequency), this decrease was highlypredictable, with lower variability than observedfor tone and click broadcasts in the dry season.
DISCUSSIONHere, we characterized the acoustic landscape of
a typical common marmoset habitat in NortheasternBrazil. Combining recordings of the ambient habitatacoustics with sound broadcast experiments, wesought to test whether the common marmoset long‐distance contact call—the phee call (Fig. 1)—com-
prises acoustic characteristics that may have beenselected to improve signal transmission, and there-fore communicative efficacy, in the species’ naturalhistory. Previous acoustic analyses indicated that,similar to those of many other Callitrichids, commonmarmoset vocalizations consist of a fundamentalfrequency (�6–9 kHz) that degrades quickly in forestenvironments [Bezera & Souto, 2008; Milleret al., 2010]. It is possible, however, that additionalacoustic characteristics augment signal transmis-sion. Our analyses indicated that phee call degrada-tion was much less variable relative to other acousticsignals, suggesting that the rate of acoustic distortionmay be predictable to conspecific signal receivers.
Fig. 5. Reverberation analysis on click broadcasts. Upper plot shows wet season, lower plot shows dry season. Click pulse fundamentalfrequency (F0) is indicated by colors, see inset figure legend. Y‐axis shows modulation depth, the ratio of mean amplitude during the70 msec silent inter‐pulse intervals to mean amplitude of the preceding 30 msec click pulse. This provides a measurement of how muchsound is left reverberating during the silence from the previous click pulse. To spectrally isolate the broadcast signal and itsreverberations, all recordings were bandpass filtered with filter corners�100 Hz from the click F0. X‐axis shows measurement distance.
Am. J. Primatol.
912 / Morrill et al.
Furthermore, selection may not have acted only onthe signal structure per se, but on behavioralmechanisms formonitoring the acoustic environmentand initiating vocal production during periods of lowacoustic interference.
Based on previous work [De la Torre &Snowdon, 2002; Waser & Brown, 1986], we predictedthat habitat acoustics at Tapacurá would showpatterns by time of day and season, and, as such,
recordings were carried out across the wet and dryseasons. Indeed, comparison of frequency spectrafrom ambient recordings showed that such temporalregularities exist. On the circadian scale, morningsand evenings were generally dominated by broad-band biotic noise from insects, birds, and anurans,while the midday showed a lull in ambient noise.Seasonally, biotic noise generally played a moresignificant role in ambient acoustics during the dryseason than the wet. This does not necessarilysuggest decreased general calling activity duringthe wet season, as damping of biotic noise due toincreased vegetation and foliage may also play a role.Such a conclusion is supported by decreased trans-mission efficacy of some broadcasts during thisseason. Despite these temporal regularities inpotentially interfering habitat sound, initial datasuggest that average calling rate for commonmarmosets does not fluctuate significantly through-out their waking hours [Bezerra et al., 2009]. Dataspecifically on the occurrence of phee production overthe course of the day, however, are not available.Such a dataset is necessary in order to fully explicatethis possibility.
Past work on habitat acoustics tested for aspectral “sound window,” a frequency band withminimal interference from other sound sources in thehabitat, providing a clear channel for communication[Waser &Brown, 1984, 1986]. The phee call, themostimportant common marmoset long‐distance call, hasa fundamental frequency of �7 kHz [Milleret al., 2010], a high rangewhich confers the seeminglydisadvantageous property of rapid degradation overdistance [Morton, 1975]. Although spectral analysisof marmoset habitat acoustics showed most peaks inthe frequency spectrogram lie either outside or on thefringes of this band (Fig. 2), we found no clearevidence of a spectral “sound window;” much energyis still concentrated between �6 and 9 kHz. Thissuggests that, in contrast to Old World monkeys[Brown et al., 1995; Waser & Brown, 1986], thefrequency range of common marmoset vocalizationsmay not have been selected for transmission proper-ties. Many factors may have contributed to suchvariability in the environment‐acoustic signal rela-tionship between New and Old World primates. Forexample, differences in the density of endemic andmigratory birds in these forest environments mayaffect the occurrence and consistency of spectralsoundwindows. As discussed earlier, two Callitrichidspecies—cotton‐top and Geoffroy’s tamarins (S.oedipus, S. geoffroyi)—produce long‐distance vocal-izations with significantly lower F0 than commonmarmosets and the other members of the family[Cleveland & Snowdon, 1982; Weiss et al., 2001].Importantly, these two tamarin species are the onlyCallitrichids endemic to forests north of the Amazon.As such, the biotic and abiotic sources of acousticinterference that faced these species during their
Fig. 6. Phee call broadcasts, cross‐correlation analysis. Axes as inFigure 4 cross‐correlation plots, except that Y‐axis range is 0–0.45. Dashed lines indicate wet season, solid lines indicate dryseason.
Fig. 7. Frequency‐manipulated phee call broadcasts, cross‐correlation analysis. Broadcast sounds had digitallymanipulatedmean fundamental frequencies, altered as indicated on spectro-gram plots. Spectrogram plots show one example of the fivedifferent manipulated calls. Spectrogram axes as in Figure 4,except that Y‐axis range is 0–22.05 kHz and X‐axis range is0–6.0 sec. Adjacent line plots show correlation analysis results,axes as in Figure 4, except that Y‐axis range is 0–0.5.
Am. J. Primatol.
Habitat Acoustics of Common Marmosets / 913
respective evolution are likely to be different from theother Callitrichid species. While this may not be thesole contributor to the observed differences in thisfamily, it is a reasonable factor. Furthermore, NewWorld primates seem to exhibit a higher degree ofvocal learning and plasticity than Old World pri-mates [Egnor & Hauser, 2004], which may allow forindividual callers to more readily modify their callingbehavior in response to the immediate acousticenvironment. More empirical work, however, isnecessary to confirm either of these possibleexplanations.
Analysis of phee call broadcast data generallyconfirmed our predictions for signal degradationtrends. Transmission efficacy for phee calls decreasedas a function of distance, as well as with increasingfundamental frequency. These trends were highlyconsistent with little variability between repeatedbroadcasts. Surprisingly, tones and clicks, our artifi-cial broadcast sounds, did not show the same trend bydistance. Instead, these sounds generally showedgreatest transmission efficacy at intermediate dis-tances of 20 and 40 m. Tones and clicks broadcast inthe dry season showed higher overall transmissionefficacy, but also higher variance, than in the wetseason. Taken together with the ambient acousticanalysis, these data suggest that the dry seasonprovides an environment for increased signal propa-gation but also higher interference due to biotic soundfrom heterospecific vocalizations, resulting in bothincreased transmission efficacy and variability. The
wet season, which is defined by increased foliagedensity, results in decreased and less variable signaltransmission.
The drastically different patterns found forphee broadcasts (a natural sound) versus tones andclicks (artificial sounds) may indicate at least someacoustic dimensions of the phee call were selected forsound transmission. Broadcast phee calls showedhighly predictable degradation over distance, aproperty that may be crucial for a long‐distancecontact call by providing the call receiver(s) withlocational information about the calling animal.Without these predictable degradation patterns,the receiver would not be able to aurally calculatethe position of the sender. In addition, this studydemonstrates that certain properties of the phee callmay be unrelated to sound transmission in thenaturalmarmoset habitat. For example, our analysesindicated that increasing or decreasing the length ofthe phee call pulse does not affect transmissionefficacy. Ruling out such a role, we may hypothesizethat pulse length variations instead play a social rolein communicating caller identity or some otherinformational feature of the phee call [Milleret al., 2010]. While the design of the current studyprecludes strong conclusions about the functionalrole of specific properties of the phee call, subsequentstudies in the field and laboratory will further testthis possibility.
Maintenance of effective communication in anoisy acoustic landscape is unlikely to rely entirely on
Fig. 8. Length‐manipulated phee call broadcasts, cross‐correlation analysis. Broadcast sounds had digitally manipulated pulse lengths,with lengths scaled as indicated on spectrogram plots. Inter‐pulse interval length was not altered. Spectrogram plots show one exampleof the five different manipulated calls. Spectrogram axes as in Figure 4, except that Y‐axis range is 0–22.05 kHz and X‐axis range is0–6.0 sec. Adjacent line plots show correlation analysis results, axes as in Figure 4, except that Y‐axis range is 0–0.4.
Am. J. Primatol.
914 / Morrill et al.
selection for the properties of a vocal signal. Soundtransmission can also be improved through comple-mentary behavioral control during vocal communica-tion. For example, a calling marmoset may monitorthe acoustic landscape and initiate vocal productionbased on temporal windows with little biotic inter-ference. This capacity requires mechanisms forauditory feedback and vocal control, two processeshistorically thought to be limited in nonhumanprimates. Recent work in common marmosets,however, has provided evidence to the contrary.Behavioral experiments indicate some degree of vocalcontrol over the acoustic structure of vocalizations[Miller et al., 2009], as well as an ability tocommunicate in the presence of experimentallycontrolled noise events [Roy et al., 2011]. Roy et al.[2011] broadcast bursts of white noise at differenttemporal patterns to pairs of visually occludedcommon marmosets. Subjects quickly began to onlyinitiate vocal production during silent periods. In onetest condition, consisting of alternating periods of8 sec white noise followed by 8 sec of silence, thecallers coordinated their vocal behavior in order tocomplete a vocal exchange within the 8 sec silentwindow. The capacity for vocal behavior mediated byenvironmental acoustic monitoring may be based onneural mechanisms in the auditory system forsensory‐motor integration [Eliades & Wang,2003, 2008]. Marmosets’ ability to exploit temporalwindows suggests that they may have evolvedmechanisms for a mixed communication strategy innoisy forest environments. Theymonitor the acousticenvironment for periods of silence and produce callswith highly predictable degradation rates, therebyemitting signals with the best opportunity foreffective communication. More data are needed,however, on the timing and occurrence of marmosetvocalizations in the presence of natural habitatacoustic interference.
Animal vocalizations, of course, are not evolu-tionarily selected solely for signal transmissionproperties in noisy environments. Communicationsystems evolve in response to a myriad of selectionforces ranging beyond purely ecological constraints,such as each signal’s social function, predationavoidance, etc. Relative to anuran and songbirdcommunication [Bradbury & Vehrencamp, 2011;Gerhardt & Huber, 2002; Ziegler & Marler, 2004],little remains known about the various ecological,social, and neural factors that affect nonhumanprimate communication. However, ongoing behavior-al and neurobiological research on the mechanismsunderlying common marmoset vocal perception andproduction has positioned this species as an emergingmodel system. Field research complimentary to suchlaboratory studies provides a powerful approachto enhance our understanding of the various forcesthat shaped the evolution of nonhuman primatecommunication.
ACKNOWLEDGMENTSThis work supported by grants to C.T.M. from
NIH (K99/R00 DC009007; R01 DC012087) and aUCSDAcademic SenateGrant.We thank theFederalRural University of Pernambuco for permission toconduct this research at Tapacurá Ecological FieldStation (Recife, Brazil). We also thank Camile Bioneand Monique Bastos for their help during fieldexperiments, and Paulo Martins for support at thefield station. All research adhered to the ASP ethicalprinciples for treatment of nonhuman primates inresearch and the Brazilian laws.
REFERENCES
Bezera BM, Souto A. 2008. Structure and usage of thevocal repertoire of Callithrix jacchus. Int J Primatol 29:671–701.
Bezerra BM, Souto A, de Oliveira MAB, Halsey LG. 2009.Vocalizations of wild common marmosets are influencedby diurnal and ontogenetic factors. Primates 50:231–237.
Bradbury JW, Vehrencamp SL. 2011. Principles of animalcommunication. Sunderland, MA: Sinauer Associates.
Brown CH, Gomez R, Waser PM. 1995. Old world monkeyvocalizations: adaptations to the local habitat? Anim Behav50:945–961.
Cleveland J, Snowdon CT. 1982. The complex vocal repertoireof the adult cotton‐top tamarin, Saguinus oedipus oedipus.Zeit Tierpsychol 58:231–270.
De la Torre S, Snowdon CT. 2002. Environmental correlates ofvocal communication of wild pygmy marmosets, Cebuellapygmaea. Anim Behav 63:847–856.
Egnor SER, Hauser MD. 2004. A paradox in the evolution ofprimate vocal learning. TINS 27:649–654.
Eliades SJ, Wang X. 2003. Sensory‐motor interaction in theprimate auditory cortex during self‐initiated vocalizations. JNeurophys 89:2185–2207.
Eliades SJ, Wang X. 2008. Neural substrates of vocalizationfeedback monitoring in primate auditory cortex. Nature453:1102–1106.
Elowson AM, Snowdon CT. 1994. Pygmy marmosets, Cebuellapygmaea, modify vocal structure in response to changedsocial environment. Anim Behav 47:1267–1277.
Gerhardt HC, Huber F. 2002. Acoustic communication ininsects and anurans: common problems and diverse sol-utions. Chicago: Chicago University Press.
Hubrecht RC. 1985. Home range size and use and territorialbehavior in the common marmoset, Callithrix jacchusjacchus, at the Tapacura Field Station, Recife, Brazil. IntJ Primatol 6:533–550.
Kime NM, Turner WR, Ryan MJ. 2000. The transmission ofadvertisement calls in Central American frogs. Behav Ecol11:71–83.
Marten K, Marler P. 1977. Sound transmission and itssignificance for animal vocalization. 1. Temperate habitats.Behav Ecol Sociobiol 2:271–290.
Marten K, Quine DB, Marler P. 1977. Sound transmission andits significance for animal vocalization. 2. Tropical habitats.Behav Ecol Sociobiol 2:291–302.
Miller CT, Eliades SJ, Wang X. 2009. Motor‐planning for vocalproduction in common marmosets. Anim Behav 78:1195–1203.
Miller CT, Mandel K, Wang X. 2010. The communicativecontent of the common marmoset phee call during antipho-nal calling. Am J Primatol 72:974–980.
Morton ES. 1975. Ecological sources of selection on aviansounds. Am Nat 109:17–34.
Am. J. Primatol.
Habitat Acoustics of Common Marmosets / 915
Naguib M. 2003. Reverberation of rapid and slow trills:implications for signal adaptations to long‐range communi-cation. J Acoustic Soc Am 113:1749–1756.
Nocedal J. 1994. Local migrations of insectivorous birds inwestern Mexico: implications for the protection and conser-vation of their habitats. In:Morton ES, YoungK, RamosMA,editors. Conservation international special edition: neotrop-ical migrants: growing points in neotropical migratory birdconservation. Cambridge: Birdlife International/CambridgeUniversity Press. p 129–142.
Norcross JL, Newman JD. 1993. Context and gender specificdifferences in the acoustic structure of common marmoset(Callithrix jacchus) phee calls. Am J Primatol 30:37–54.
Pistorio A, Vintch B, Wang X. 2006. Acoustic analyses ofvocal development in a New World primate, the commonmarmoset (Callithrix jacchus). J Acoustic Soc Am 120:1655–1670.
Pola Y, Snowdon CT. 1975. The vocalizations of pygmymarmosets (Cebuella pygmaea). Anim Behav 23:826–842.
Roy S,Miller CT, GottschD,Wang X. 2011. Vocal control by thecommonmarmoset in the presence of interfering noise. J ExpBiol 214:3619–3629.
Santos EM, Almeida AV, Vasconcelos SD. 2004. Feedinghabits of six anuran (Amphibia: Anura) species in arainforest fragment in northeastern Brazil. Iheringia94:933–948.
Scanlon CE, Chalmers NR, Monteiro da Cruz MAO. 1989.Home range use and exploitation of gum in the marmosetCallithrix jacchus jacchus. Int J Primatol 10:123–136.
Souto A, Bezerra BM, Schiel N, Huber L. 2007. Salutatorysearch in free‐living Callithrix jacchus: environmental andage influences. Int J Primatol 28:881–893.
Waser P, Brown CH. 1984. Is there a “sound window”for primate communication? Behav Ecol Sociobiol 15:73–76.
Waser PM, Brown CH. 1986. Habitat acoustics and primatecommunication. Am J Primatol 10:135–154.
Waser PM, Waser MS. 1977. Experimental studies of primatevocalization—specializations for long‐distance propagation.Zeit Tierpsych 43:239–263.
Weiss DJ, Garibaldi BT, HauserMD. 2001. The production andperception of long calls by cotton‐top tamarins (Saguinusoedipus): acoustic analyses and playback experiments. JComp Psychol 11:258–271.
Wiley RH, Richards DG. 1978. Physical constraints on acousticcommunication in the atmosphere: implications for theevolution of animal vocalizations. Behav Ecol Sociobiol 3:69–94.
Wiley RH, Richards DG. 1982. Adaptations for acousticcommunication in birds: sound propagation and signaldetection. In: Kroodsma DE, Miller EH, editors. Acousticcommunication in birds. vol 1. New York, NY: AcademicPress. p 131–181.
Ziegler HP, Marler P. 2004. Behavioral neurobiology ofbirdsong. New York, NY: New York Academy of Sciences.
Zimmerman BL. 1983. A comparison of structural features ofcalls of open and forest habitat frogs species in the centralAmazon. Herpetologica 39:235–246.