JOURNAL OF BACTERIOLOGY, Aug. 1968, p. 374-382 Vol. 96, No. 2 Copyright @ 1968 American Society for Microbiology Printed in U.S.A. Reversal of the Vancomycin Inhibition of Peptidoglycan Synthesis by Cell Walls' RABINDRA K. SINHA2 AND FRANCIS C. NEUHAUS3 Biochemistry Division, Department of Chemistry, Northwestern University, Evanston, Illinois 60201 Received for publication 8 May 1968 Addition of cell walls to the peptidoglycan synthetase-acceptor system containing vancomycin (50 ,g/ml) prevented the inhibition by the antibiotic. In addition, the inhibition of incorporation of ['4C]muramyl-pentapeptide into peptidoglycan in the presence of vancomycin was reversed by the addition of cell walls to the assay mix- ture at 60 min. Cell walls previously saturated with vancomycin lost their ability to reverse the inhibition by the antibiotic. The inhibition of peptidoglycan synthesis by ristocetin was partially reversed by the addition of cell walls. The initial stage in peptidoglycan synthesis is catalyzed by phospho-N-acetyl(NAc)muramyl-penta- peptide translocase (uridine 5'-phosphate) according to the reaction: UDP-NAc-muramyl-pentapeptide + acceptor > acceptor- phospho-NAc-muramyl-pentapeptide + UMP where acceptor is C55-isoprenoid alcohol phosphate. Vancomycin stimulates the transfer of phospho-NAc-muramyl-pentapeptide to the acceptor, and the addition of cell walls to this assay mixture prevented the stimulation of transfer. In addition to the transfer reaction, the enzyme catalyzes the exchange of [3H]uridine monophosphate (UMP) with UDP-NAc-muramyl-pentapeptide. The exchange reaction is effectively inhibited by vancomycin. For example, 60 ,ug of vancomycin per ml inhibited the rate of exchange by 50%. Addition of cell walls restored the exchange of UMP with the UMP moiety of UDP-NAc-muramyl-pentapeptide. Thus, cell walls appeared to have a higher affinity for vancomycin than did either the peptidoglycan synthetase-acceptor system or phospho-NAc-muramyl-pentapeptide translocase. These results provide support for the proposal made by Best and Dur- ham that the effective binding of vancomycin to the cell wall could result in the in- hibition of transfer of membrane-associated peptidoglycan chains to the growing wall. Vancomycin inhibits the biosynthesis of bac- terial cell walls. The inhibition is accompanied by the accumulation of uridine diphosphate- N-acetyl-muramyl-L - alanyl- D - isoglutamyl- L- ly - syl-D-alanyl-D -alanine (UDP-NAc -muramyl-L- ala-D-isoglu-L-lys-D-ala-D-ala) in Staphylococcus aureus (17). This nucleotide and UDP-NAc- glucosamine are precursors of peptidoglycan, the major structural polymer of bacterial cell walls. The site of action of vancomycin has been investigated in several laboratories. Best and 1 Presented in part at the 67th Annual Meeting of the American Society for Microbiology, New York, N.Y., 30 April-4 May 1967. 2 Present address: Bose Institute, Calcutta, India. 3 Supported by U.S. Public Health Service Research Career Development Program Award 1-K3-AI-6950 from the National Institute of Allergy and Infectious Diseases. Durham (5) proposed that adsorption of vanco- mycin to acidic groups on the cell wall could inhibit the addition of new cell wall components to the existing peptidoglycan. The inhibition of teichoic acid biosynthesis by vancomycin in Bacillus subtilis and Bacillus licheniformis has been reported by Burger and Glaser (7). In 1965, Jordan (13) found that the inhibition of cell wall synthesis by vancomycin occurs earlier than that of membrane biosynthesis. It was concluded that the primary effect of vancomycin in intact cells of staphylococci is an inhibition of wall peptido- glycan synthesis and that inhibition of membrane biosynthesis is a secondary effect. Anderson et al. (1-3) established that peptidoglycan synthetase is effectively inhibited by both vancomycin and ristocetin. Moreover, vancomycin also affects phospho-N-acetyl-muramyl pentapeptide trans- locase (23, 24). Chatterjee and Perkins (8) 374 on June 17, 2018 by guest http://jb.asm.org/ Downloaded from
Transcript
JOURNAL OF BACTERIOLOGY, Aug. 1968, p. 374-382 Vol. 96, No. 2Copyright @ 1968 American Society for Microbiology Printed in U.S.A.
Reversal of the Vancomycin Inhibition ofPeptidoglycan Synthesis by Cell Walls'
RABINDRA K. SINHA2 AND FRANCIS C. NEUHAUS3
Biochemistry Division, Department ofChemistry, Northwestern University, Evanston, Illinois 60201
Received for publication 8 May 1968
Addition of cell walls to the peptidoglycan synthetase-acceptor system containingvancomycin (50 ,g/ml) prevented the inhibition by the antibiotic. In addition, theinhibition of incorporation of ['4C]muramyl-pentapeptide into peptidoglycan in thepresence of vancomycin was reversed by the addition of cell walls to the assay mix-ture at 60 min. Cell walls previously saturated with vancomycin lost their ability toreverse the inhibition by the antibiotic. The inhibition of peptidoglycan synthesis byristocetin was partially reversed by the addition of cell walls. The initial stage inpeptidoglycan synthesis is catalyzed by phospho-N-acetyl(NAc)muramyl-penta-peptide translocase (uridine 5'-phosphate) according to the reaction:
where acceptor is C55-isoprenoid alcohol phosphate. Vancomycin stimulatesthe transfer of phospho-NAc-muramyl-pentapeptide to the acceptor, and theaddition of cell walls to this assay mixture prevented the stimulation of transfer. Inaddition to the transfer reaction, the enzyme catalyzes the exchange of [3H]uridinemonophosphate (UMP) with UDP-NAc-muramyl-pentapeptide. The exchangereaction is effectively inhibited by vancomycin. For example, 60 ,ug of vancomycinper ml inhibited the rate of exchange by 50%. Addition of cell walls restored theexchange of UMP with the UMP moiety of UDP-NAc-muramyl-pentapeptide. Thus,cell walls appeared to have a higher affinity for vancomycin than did either thepeptidoglycan synthetase-acceptor system or phospho-NAc-muramyl-pentapeptidetranslocase. These results provide support for the proposal made by Best and Dur-ham that the effective binding of vancomycin to the cell wall could result in the in-hibition of transfer of membrane-associated peptidoglycan chains to the growingwall.
Vancomycin inhibits the biosynthesis of bac-terial cell walls. The inhibition is accompaniedby the accumulation of uridine diphosphate-N-acetyl-muramyl-L - alanyl-D - isoglutamyl- L- ly -syl-D-alanyl-D -alanine (UDP-NAc -muramyl-L-ala-D-isoglu-L-lys-D-ala-D-ala) in Staphylococcusaureus (17). This nucleotide and UDP-NAc-glucosamine are precursors of peptidoglycan,the major structural polymer of bacterial cellwalls. The site of action of vancomycin has beeninvestigated in several laboratories. Best and
1 Presented in part at the 67th Annual Meeting ofthe American Society for Microbiology, New York,N.Y., 30 April-4 May 1967.
2 Present address: Bose Institute, Calcutta, India.3 Supported by U.S. Public Health Service Research
Career Development Program Award 1-K3-AI-6950from the National Institute of Allergy and InfectiousDiseases.
Durham (5) proposed that adsorption of vanco-mycin to acidic groups on the cell wall couldinhibit the addition of new cell wall componentsto the existing peptidoglycan. The inhibition ofteichoic acid biosynthesis by vancomycin inBacillus subtilis and Bacillus licheniformis hasbeen reported by Burger and Glaser (7). In 1965,Jordan (13) found that the inhibition of cell wallsynthesis by vancomycin occurs earlier than thatof membrane biosynthesis. It was concluded thatthe primary effect of vancomycin in intact cellsof staphylococci is an inhibition of wall peptido-glycan synthesis and that inhibition of membranebiosynthesis is a secondary effect. Anderson et al.(1-3) established that peptidoglycan synthetaseis effectively inhibited by both vancomycin andristocetin. Moreover, vancomycin also affectsphospho-N-acetyl-muramyl pentapeptide trans-locase (23, 24). Chatterjee and Perkins (8)
discovered an additional nucleotide in whichvancomycin is bound to UDP-NAc-muramyl-pentapeptide when S. aureus, Micrococcuslysodeikticus, and Corynebacterium poinsettiaeare grown in the presence of the antibiotic.Recently, these investigators (9) reported thatthe vancomycin-nucleotide compound causes agreater inhibition of peptidoglycan formation incell-free systems than a corresponding amount offree vancomycin. It was suggested that the nucleo-tide-vancomycin adduct may be a necessaryintermediate for the function of the antibiotic (9).
Thus, there are a multiplicity of action sitesfor vancomycin. Inhibition at one or more ofthese sites could lead to the observed accumula-tion of UJDP-NAc-muramyl-pentapeptide andinhibition of peptidoglycan synthesis. This in-vestigation was undertaken to establish whichcomponent, the peptidoglycan synthetase-ac-ceptor system or the cell wall, has the higheraffinity for vancomycin. A study of the relativeaffinities for this antibiotic in in vitro experimentswill contribute to our understanding of theprimary site of action.
MATERIALS AND METHODSMaterials. D-[14C]alanine was the product of Cal-
biochem, Los Angeles, Calif. UDP-NAc-glucosamineand Sephadex G-25 were purchased from SigmaChemical Co., St. Louis, Mo., and Pharmacia FineChemicals, Inc., New Market, N.J., respectively.Triton X-100 and Antifoam 66 were the products ofRohm and Haas, Chicago, Ill., and General Electric(Silicone Products Department), Waterford, N.Y.,respectively. The plastic beads (styrene-divinyl ben-zene copolymer, 20 to 50 mesh, 8% cross-linked)were a gift from Dow Chemical Co. M. lysodeikticus(ATCC 4698) was purchased from American TypeCulture Collection, and S. aureus Copenhagen waskindly provided by J. L. Strominger. Vancomycin andristocetin (Spontin) were given by Eli Lilly & Co.,Indianapolis, Ind., and Abbott Laboratories, NorthChicago, Ill., respectively. Bacitracin was purchasedfrom Nutritional Biochemicals Corp., Cleveland,Ohio.
UDP-NAc-muramyl-L-ala-D-isoglu-L-lys and UDP-NAc -muramyl - L -ala-D-iSoglU-L-lys-D-[1 4C]ala-D-["4C]-ala were prepared as previously described (16) and bythe procedure of Stickgold and Neuhaus (25).Enzyme preparations. The preparation of phospho-
NAc-muramyl-pentapeptide translocase from S.aureus Copenhagen has been previously described(24). The membrane preparation which catalyzes thesynthesis of peptidoglycan was isolated from M.lysodeikticus (ATCC 4698). The bacteria were grownat 37 C with shaking for 24 hr in medium containing1% glucose, 0.5% K2HPO4, 1% yeast extract, 1%peptone, and 1%G of a salt solution containing 4%MgSO4 7H20, 0.2% FeSO4 -7H20, 0.16% MnSO4,and 0.2% NaCl (4). The yield of bacteria was 7 g(wet weight) per liter. The cells were washed in 0.02 MTris-chloride (pH 7.8) and resuspended to 15% (wetweight) in 0.005 M Tris-chloride (pH 7.8) containing
TABLE 1. Analysis of cell wall preparation fromM. lysodeikticusa
CzrasiGhuysenAmino acid (sugar) This work Czerkawski and SaltonI
a The results are expressed as molar ratios.b A wide variation of alanine-glutamic acid
ratios has been reported for M. lysodeikticus:Whitney and Grula (27), 2.1; Salton and Pavlik(20), 2.6; Sharon et al. (21), 2.7, in addition tothose reported above.
1 M KCI. The cells were disrupted according to theprocedure described by Struve et al. (25). The cellwalls and unbroken cells were removed at 4,340 X gfor 30 min, and the particulate enzyme fraction wassedimented by centrifugation at 144,000 X g for 30min. The sediment was suspended in 0.005 M Tris-chloride (pH 7.8) containing 1 M KCI, and the centrifu-gation between 4,340 X g for 30 min and 144,000 X gfor 30 min was repeated four times. The final pre-cipitate was suspended in three times its weight of0.005 M Tris-chloride (pH 7.8) containing 1 M KCI.The enzyme preparation was stored at -196 C.
Cell wall isolation. Cell walls from S. aureus Copen-hagen were prepared according to the method pre-viously described (24). For the preparation of cellwalls from M. lysodeikticus, the cells were disruptedas described above. Unbroken cells and cell wallswere sedimented at 12,000 X g for 30 min. The sedi-ment was suspended in 0.005 M Tris-chloride (pH 7.8)containing 1 M KCl and centrifuged at 3,000 X g for30 min. The supernatant fraction containing the cellwalls was centrifuged at 12,000 X g for 10 min. Thewalls were washed three times with the above buffercontaining 1 M KCl. The final precipitate (5.8 g, wetweight) was suspended in 18 ml of 0.005 M Tris-chloride (pH 7.8).The purity of the cell walls was established by
analyzing for amino acids, yellow pigmentation, andnucleic acid (18). For amino acid analysis, 0.7 mg(dry weight) of the walls was hydrolyzed in 0.5 ml of5.7 N HCl under nitrogen for 12 hr at 110 C. Theamino acids were determined on the Beckman model120 amino acid analyzer (Table 1) according to themethods described by Spackman et al. (22). Neitheryellow pigmentation nor nucleic acid was detected inthe wall preparations.
Assay for phospho-N-acetyl-muramyl-pentapeptide-translocase (UMP). This assay involves the transferof phospho-N-acetyl-muramyl-[14C]pentapeptide to alipid acceptor according to the following reaction:UDP-NAc-muramyl-[14C]pentapeptide+ acceptor > acceptor-phospho-NAc-
muramyl-['4C]pentapeptide + UMP (1)The amount of lipid intermediate was measured as a
perchloric acid precipitable fraction as describedpreviously (24). In addition, the translocase has beenassayed by the exchange of uridine monophosphate(UMP) with the [3H]UMP moiety of [3H]UDP-NAc-muramyl-pentapeptide according to the proceduredescribed by Heydanek et al. (in press).
Assay for peptidoglycan synthesis. This assay meas-ures the incorporation of muramyl-[14C]pentapeptidefrom UDP-NAc-muramyl-['4C]pentapeptide into pep-tidoglycan. The assay mixture contained: 0.05 M Tris-chloride (pH 7.8), 0.01 M MaC12 , 1.5 X 10-5 MUDP-NAc-muramyl-['4C]pentapeptide (7.5 X 103counts/min per nanomole), 5 X 10-5 M UDP-NAc-glucosamine, and the membrane preparation (1.1 mg,dry weight) from M. lysodeikticus in a total volume of0.1 ml. The incubation was carried out for 3 hr at25 C. The reaction was terminated by placing the tubein a boiling-water bath for 2 min, and the suspensionwas applied to Whatman 3MM paper and developedin a solvent system of isobutyric acid = NH40H-water(66:1:33, v/v). The peptidoglycan remained at theorigin of the chromatogram. It was excised andcounted in a Tri-Carb liquid scintillation spectrometer(Packard Instruments Co., Downers Grove, Ill.). Thescintillation fluid was toluene containing 0.3% 2,5-diphenyloxazole. The assay is similar to that describedby Anderson et al. (3), with the exception that we haveused Mn2+ and have omitted adenosine triphosphate(ATP).
Adsorption of vancomycin and ristocetin by cell walls.The adsorption of antibiotics to the cell walls wasassayed by a procedure similar to that described byBest and Durham (5). The assay mixture contained0.05 M Tris-chloride (pH 7.8), 0.01 M MnCl2, cellwalls, and either 1,000 lAg of vancomycin or 500 ,g ofristocetin per ml in a total volume of 1 ml. The adsorp-tion was carried out at 25 C for 30 min. The cell wallswere removed by centrifugation at 12,000 X g for 5min. The amount of antibiotic in the supernatantsolution was measured spectrophotometrically at280 mrn.
RESULTS
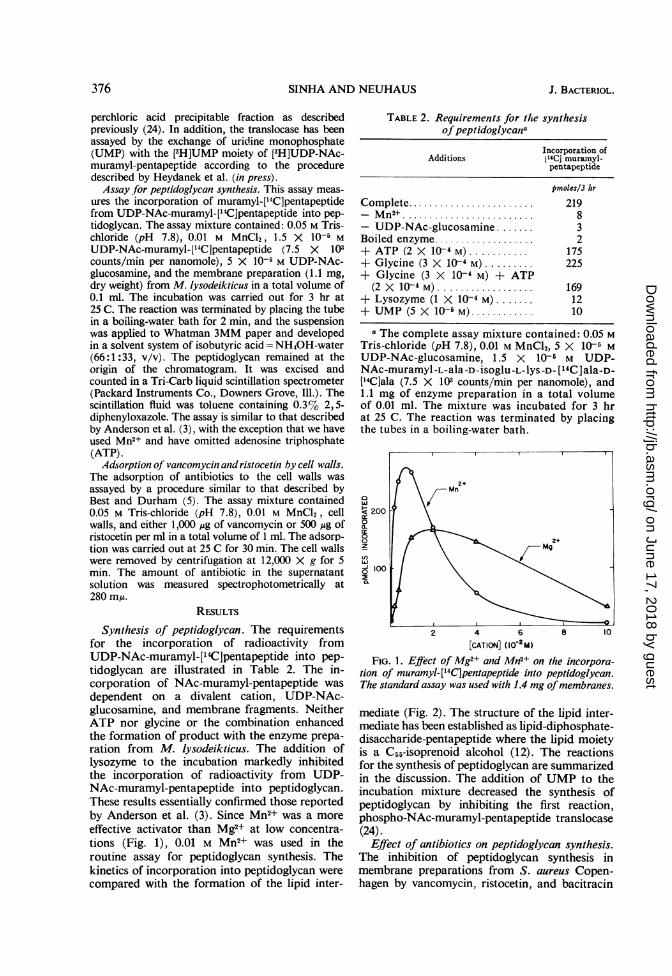
Synthesis of peptidoglycan. The requirementsfor the incorporation of radioactivity fromUDP-NAc-muramyl-['4C]pentapeptide into pep-tidoglycan are illustrated in Table 2. The in-corporation of NAc-muramyl-pentapeptide wasdependent on a divalent cation, UDP-NAc-glucosamine, and membrane fragments. NeitherATP nor glycine or the combination enhancedthe formation of product with the enzyme prepa-ration from M. lysodeikticus. The addition oflysozyme to the incubation markedly inhibitedthe incorporation of radioactivity from UDP-NAc-muramyl-pentapeptide into peptidoglycan.These results essentially confirmed those reportedby Anderson et al. (3). Since Mn2+ was a moreeffective activator than Mg2+ at low concentra-tions (Fig. 1), 0.01 M Mn2+ was used in theroutine assay for peptidoglycan synthesis. Thekinetics of incorporation into peptidoglycan werecompared with the formation of the lipid inter-
TABLE 2. Requirements for the synthesisofpeptidoglycana
Incorporation ofAdditions 114C1 muramyl-
pentapeptide
pmoles/3 hr
Complete ............. 219- Mn2.......................... 8- UDP-NAc-glucosamine ....... 3Boiled enzyme................... 2+ ATP (2X 10-4 M) ....... 175+ Glycine (3 X 10- M) ...... 225+ Glycine (3 X 10 4 M) + ATP
(2 X 104 M) ......... ......... 169+ Lysozyme (1 X 104 M) ...... 12+UMP (5X1-5M)............ 10
a The complete assay mixture contained: 0.05 MTris-chloride (pH 7.8), 0.01 M MnCl2, 5 X 105 MUDP-NAc-glucosamine, 1.5 X 105 M UDP-NAc-muramyl -L-ala -D- isoglu -L-lyS -D- [14C] ala-D-['4C]ala (7.5 X 103 counts/min per nanomole), and1.1 mg of enzyme preparation in a total volumeof 0.01 ml. The mixture was incubated for 3 hrat 25 C. The reaction was terminated by placingthe tubes in a boiling-water bath.
2 4 6 8 10[CATION] (10-2M)
FIG. 1. EjIect of Mg2+ and Mn2+ on the incorpora-tion of muramyl-[14C]pentapeptide into peptidoglycan.The standard assay was used with 1.4 mg ofmembran2es.
mediate (Fig. 2). The structure of the lipid inter-mediate has been established as lipid-diphosphate-disaccharide-pentapeptide where the lipid moietyis a C55-isoprenoid alcohol (12). The reactionsfor the synthesis of peptidoglycan are summarizedin the discussion. The addition of UMP to theincubation mixture decreased the synthesis ofpeptidoglycan by inhibiting the first reaction,phospho-NAc-muramyl-pentapeptide translocase(24).
Effect of antibiotics on peptidoglycan synthesis.The inhibition of peptidoglycan synthesis inmembrane preparations from S. aureus Copen-hagen by vancomycin, ristocetin, and bacitracin
was observed by Anderson et al. (1-3) and con-firmed by Struve et al. (23, 24). The effect of 250vancomycin, ristocetin, and bacitracin on thesynthesis of peptidoglycan with our enzymepreparation from M. lysodeikticus is shown in 200Fig. 3. The amount of antibiotic required for 50%
a
inhibition of the incorporation is as follows: @U VANCOMYCINristocetin, 7.5 ,ug/ml; vancomycin, 20 ,g/ml; 50bacitracin, 11 ,ug/ml. At a concentration of 50 M
0jug/ml, ristocetin, vancomycin, and bacitracin z RISTOCETINinhibited 96%, 85%, and 74%, respectively. In , \similar incubations, Anderson et al. (3) observed ilooethat the amount of antibiotic required for 50% 1inhibition was 15, 10, and 30 ,ug/ml for ristocetin, / BACITRACINvancomycin, and bacitracin, respectively. 50
Reversal of the inhibition by vancomycin and 50ristocetin. The effect of walls on the inhibition ofincorporation of NAc-muramyl-['4C]pentapep-tide into peptidoglycan by vancomycin, ristocetin, 50 100 150 200and bacitracin is shown in Fig. 4. Addition of ANTIBIOTIC (,Qg/mI)cell walls (0.7 mg) to the assay mixture containing FIG 3 Effect of vancomycin, ristocetin, and bacitra-vancomycin prevented most of the inhibition by cIn on thei nco mycin, ridocand sta-the antibiotic. In contrast, addition of walls (0.7 cmn on the incorporation into peptidoglycan. The stand-mg) to an incubation containing bacitracin and ard assay was used with 1.1 mg of the enzyme prepara-ristocetin did not reduce the inhibition to thesame extent.The inhibition of incorporation of NAc-
muramyl-[14C]pentapeptide into peptidoglycan oby vancomycin can be reversed by the addition CONTROLof cell walls. Vancomycin at a concentration of 20050 ,ug/ml inhibited the rate of incorporation by VANCOMYCIN
85% (Fig. 3). When cell walls were added after z1 hr to an assay mixture containing vancomycin, BACITRACINthe rate of incorporation was restored in propor- 0wtion to the amount of cell walls that were added(Fig. 5A). For example, in the presence of 0.70mg of cell walls, the velocity in the system con- RISTOCETIN
,, 0.2 0.4 0.6300 CELL WALLS (MG DRY WT)
FIG. 4. Effect of cell-wall concentration on theinhibition by antibiotics. The standard assay was usedwith 1.1 mg of the enzyme preparation. The following
$ 200 _ antibiotics were added to the assay mixture: (0) none;(AL) vancomycin (50 i.g/ml); (0) ristocetin (50 ,ug/ml);
o PEPTIDOGLYCAN and (a) bacitracin (50 gg/mm).
taining vancomycin is 80% of that observed in3 0 the absence of vancomycin. When 0.07 and 0.35
Amg of cell walls were added at 60 min, the veloci-ties were 16 and 56% of that observed in the
LIPID INTERMEDIATE absence of vancomycin. Cell walls previously,, saturated with vancomycin lost their ability to
60 120 180 reverse the inhibition by the antibiotic.MINUTES The inhibition of peptidoglycan synthesis by
FIG. 2. Kinetics of incorporation into peptidoglycan ristocetin can also be reversed by the addition ofand the lipid intermediate. The standard assay was used cell walls (Fig. 5B). For example, the addition ofwith 1.1 mg of membranes as described in the text. 0.35 mg and 0.70 mg of cell walls restored theThe lipid intermediate was measured by the method rate of incorporation to 30 and 45 % of thatdescribed by Struve et al. (24). observed in the absence of antibiotic. As in the
FIG. 5A. Reversal of the inhibition by vancomycinof peptidoglycan synthesis with cell walls from M.lysodeikticus. The standard assay was used with 1.1 mgof membranes. In the control (A), 0.35 mg of cellwalls from M. lysodeikticus was added to the standardassay (0). To the assay mixture containing 50 ,ug ofvancomycin per ml, the following additions of cell wallswere made at 60 min: (M) none; (@) 0.07 mg; (A)0.35 mg; and (0) 0.7 mg.
60 120MINUTES
FIG. 5B. Reversal of the inhibition by ristocetin ofpeptidoglycan synthesis with cell walls from M. lyso-deikticus. The procedure was identical to that describedfor Figure SA. To the assay mixture containing S0 ,ugof ristocetin per ml, the following additions of cell wallswere made at 60 min: (U) none; (0) 0.35 mg; (A)0.7mg.
case of vancomycin, the reversal was dependenton the amount of cell walls added to the assaymixture.
Effect of vancomycin and cell walls on phospho-NAc-muramyl-pentapeptide translocase (UMP).In previous studies (24), vancomycin was shownto stimulate the transfer of phospho-NAc-muramyl-['4C]pentapeptide from UDP-NAc-mu-ramyl-['4C]pentapeptide to a lipid acceptor ac-cording to the reaction:
The addition of cell walls to the assay mixturecontaining membrane fragments, UDP-NAc-muramyl-pentapeptide, and vancomycin pre-vented the stimulation of transfer by vancomycin.
200 400 600 800 1000VANCOMYCIN (&g/ml)
FIG. 6A. Reversal of the vancomycin effects (transferassay) on the translocase by cell walls. The reactionmixture for the transfer assay contained 1.5 X 105 MUDP-NAc-muramyl-[14C]pentapeptide (9.5 X 103counts/min per nanomole), 0.05 m Tris-chloride (pH7.8), 0.01 M MgCI2, and 0.55 mg ofmembrane prepara-tion from S. aureus Copenhagen in a total volume of0.1 ml. The mixture was incubated for 15 min at 25 C.The following additions of cell walls were made: (0)none, (0) 0.15 mg, (A) 0.30 mg, (A) 0.60 mg, (0)1.5mg.
200VANCOMYCIN (pg/ml)
FIG. 6B. Reversal of the vancomycin effects (ex-change assay) on the translocase by cell walls. The reac-tion mixture for the exchange assay contained 0.05 MTris-chloride (pH 7.8); 0.01 m MgC12, 3.2 X 105 m[3H]UDP-NAc-muramyl-pentapeptide (3.7 X 101counts/min per nmole), 3.3 X 10-4 m UMP, vancomy-cin, and 0.82 mg of membrane preparation from S.aureus Copenhagen in a total volume of 0.06 ml. Thefollowing additions of cell walls were made: (0) none,(0) 0.18 mg, and (0) 0.7 mg. The mixtures wereincubated at 25 C for 10 min and the reactions wereterminated by placing the tube in a boiling-water bathfor 2 min. The anount of [3H]UMP was quantitatedby the procedure describedby Heydanek et al. (in press).
The effect of increasing amounts of vancomycinon the translocase is shown in Fig. 6A. Withincreasing levels of cell walls, the concentrationrequired for maximal stimulation was shiftedtoward higher concentrations of the antibiotic.At a cell wall concentration of 1.5 mg/0.1 ml,the inhibitory and stimulatory effects of vanco-mycin were prevented. Vancomycin at low con-centrations was observed to enhance the transferassay (Fig. 6A). In contrast, low levels of theantibiotic inhibited the exchange assay (Fig. 6B).For example, 60 ,ug of vancomycin per ml in-hibited the rate of exchange by 50%. Additionof cell walls restored the exchange of UMP withthe UMP moiety of UDP-NAc-muramyl-penta-peptide.
Adsorption of vancomycin and ristocetin to cellwalls. Best and Durham (5, 6) described theadsorption of these antibiotics to cell walls iso-lated from B. subtilis. It was concluded thatelectrostatic interaction between acidic groupson the wall and basic groups on the antibioticwere involved. Mg'+ and other cations competedwith the antibiotic for the binding sites on thewall (5).To correlate the reversal of inhibition with
the extent of binding, adsorption of ristocetinand vancomycin to walls of M. lysodeikticus wasmeasured. Since divalent cations (Mn2+) werepresent in the assay mixtures for peptidoglycansynthesis, the adsorption of antibiotic was meas-ured in the presence and absence of Mn2+ (0.01M) .With vancomycin (Fig. 7A), 95% of the anti-
biotic was adsorbed in the absence of Mn2+,whereas in the presence of 0.01 M Mn2+, 82%of the antibiotic was adsorbed from the assaymixture. The concentrations of cell walls requiredfor half-maximal adsorption of 1,000 jAg of
A0180
0co 60-
~40 +0.O M Mn
20 ~
vancomycin were 0.28 and 0.68 mg/ml in theabsence and presence of 0.01 M Mn2+, respectively.With ristocetin (Fig. 7B), 83% of the antibioticwas adsorbed in the absence of Mn2+, whereasin the presence of 0.01 M Mn2+, 62% of the anti-biotic was adsorbed from the assay mixture. Theconcentration of cell walls required for half-maximal adsorption of 500 Mug of ristocetin was0.23 and 2.94 mg/ml in the absence and presenceof 0.01 M Mn2+, respectively.
DISCUSSIONThe elucidation of the enzymatic reactions
(Fig. 8) involved in the synthesis of peptidoglycanhas provided additional loci for testing the actionof vancomycin and ristocetin. Strominger andco-workers (1-3) concluded that the last stepin the cycle, transfer of the modified disaccharide-hexapeptide dimer from the lipid carrier to themembrane acceptor, is the component mostsensitive to ristocetin and vancomycin in M.lysodeikticus. The nature of the terminal acceptorassociated with the membrane has not beenestablished. Jordan and Reynolds (14) suggestedthat the in vitro experiments of Strominger andco-workers will not distinguish between theinhibition of peptidoglycan synthetase itself andthe blockage of addition sites on the acceptorassociated with the membrane.The effective binding of vancomycin and
ristocetin to the wall as described by Best andDurham (5, 6) could result in the inhibition oftransfer of membrane-associated peptidoglycanchains to the growing wall. Since vancomycinis readily adsorbed to the wall, Best and Durham(5) suggested that the inhibition by vancomycinof the membrane-associated reactions may be ofsecondary importance. This proposal is basedprimarily on the fact that vancomycin reaches
FIG. 8. Lipid cycle in the biosynthesis ofpeptidoglycan (15)
the membrane after the available sites on the cellwall have been saturated.
Thus, the experiments presented in this paperwere initiated in an attempt to establish whetherthe peptidoglycan synthetase-acceptor systemor the cell wall has the higher affinity for vanco-mycin. In addition to peptidoglycan synthetase,the initial enzyme of the cycle, phospho-NAc-muramyl-pentapeptide translocase, has beentested for the reversal of the effects by vanco-mycin.The addition of cell walls to an incubation
mixture containing vancomycin prevented theinhibition of incorporation of NAc-muramyl-['4C]pentapeptide into peptidoglycan. In thecase of ristocetin and bacitracin, the inhibitionwas partially prevented. Moreover, if cell wallswere added at 60 min to an incubation mixture
AC-nine
containing vancomycin, the inhibition of in-corporation was reversed. The extent of reversalwas dependent on the concentration of cell wallsadded to the incubation. Thus, cell walls appearto have a higher affinity for vancomycin thandoes the peptidoglycan synthetase-acceptorsystem associated with the membranes. Althoughthe concentration of walls required for reversalappears high (7 mg/ml), it must be emphasizedthat a higher concentration of membranes (11mg/ml) must be incorporated into the incubationmixture to observe the activity of peptidoglycansynthetase. On a percentage of dry weight com-parison, the ratio of wall to membrane in M.lysodeikticus is approximately one (18, 19). Thus,the concentration of walls that were added to anincubation mixture to observe reversal of theantibiotic effects is not excessive.
Best and Durham (5) observed that Mg2+ andother cations compete with vancomycin forbinding sites on the cell wall. This observationwas confirmed in the present investigation withwalls from M. lysodeikticus. An attempt wasmade to correlate the reversal of inhibitionof incorporation of NAc-muramyl-[114Cpenta-peptide into peptidoglycan by walls with theextent of binding of vancomycin to cell walls.However, it was not possible to perform theadsorption assay for vancomycin in the presenceof membranes. Thus, the concentrations of wallsrequired to adsorb a given amount of antibioticwas less in the absence of membranes than in thepresence. In the presence of 0.01 M Mn2+, only82% of the vancomycin was adsorbed from theassay mixture. The maximal reversal of inhibitionby vancomycin was 80% (Fig. 5A). These ob-servations are consistent with the experimentsdescribed by Best and Durham (5). They ob-served that only 80% of the vancomycin couldbe adsorbed from solution. (These results wereestablished in the absence of divalent cations.)Additional cell walls could not adsorb the re-maining 20%. On the basis of these results, itwas suggested (5) that vancomycin may existas two components in an 80:20 ratio.The first reaction in the cycle, phospho-NAc-
muramyl-pentapeptide translocase, is also affectedby vancomycin (24). The addition of low levelsof vancomycin to the assay mixture results in anenhancement of transfer while increasing concen-trations result in an inhibition of the transferreaction. In contrast, if the exchange assay isused, a pronounced inhibition is observed at lowconcentrations of vancomycin. Heydanek et al.(in press) proposed that the transfer of phospho-NAc-muramyl-pentapeptide to the C55-isoprenoidalcohol phosphate precedes via an enzyme-phospho-NAc-muramyl-pentapeptide intermedi-ate according to the following reaction sequence:
E + UMPPMp EUMPPMp (3)
EUMPPMp > EPMp + UMP (4)
EPMp + Acceptor Acceptor-PMp + E (5)
where E is enzyme, UMPPMp is UDP-NAc-muramyl-pentapeptide, and acceptor is C55-isoprenoid alcohol phosphate. The stimulation-inhibition observed in the transfer assay (reactions3, 4, and 5) results from a combination of thedetergent and inhibitory effects of the antibiotic.Thus, inhibition in the transfer assay is onlyobserved at high concentrations of vancomycin(500 Ag/ml). In contrast, the exchange reaction(reactions 3 and 4) is inhibited 50% by 60 ,ug/mlof antibiotic. For comparison, the peptidoglycan
synthetase-acceptor system is inhibited 50%by 20 ,ug of vancomycin per ml. Since the con-centrations of antibiotic required to inhibitthese enzymes differ by only threefold, thetranslocase may still be considered as a potentialsite of vancomycin action. It is apparent fromthe results that cell walls prevent the enhancementof transfer and the inhibition of exchange activity.When S. aureus is grown in the presence of
either vancomycin (17) or ristocetin (26), themajor nucleotide that accumulates is UDP-NAc-muramyl-pentapeptide. In addition, loweramounts of UDP-NAc-muramyl-L-ala were ob-served to accumulate in both cases. Chatterjeeand Perkins (8) observed an additional nucleotidein which vancomycin is bound to UDP-NAc-muramyl-pentapeptide. However, Best, Sinha,and Neuhaus (unpublished data) were not ableto detect a vancomycin-nucleotide adduct whenvancomycin was added to a culture of S. aureusCopenhagen. The synthesis of this complex iseffected by incubating vancomycin and UDP-NAc-muramyl-pentapeptide (H. R. Perkins,Biochem. J. 106:35P, 1968). This observationhas been confirmed by Best, Sinha, and Neuhaus(unpublished data). Although the exact role of thevancomycin-nucleotide complex has not beenestablished, this type of complex formation mayprovide the key to the mechanism of vancomycinaction. The minimal structure that is requiredfor complex formation is D-isOglu-L-lyS-D-ala-D-ala (H. R. Perkins, Biochem. J. 106:1 35P, 1968).This structure corresponds to the noncross-linked terminus where new peptidoglycan isbeing added. The binding of vancomycin to thissequence in the wall could prevent addition ofnew peptidoglycan strands according to theproposal of Best and Durham (5).
ACKNOWLEDGMENTSWe thank Rosemary Linzer and Judy Domin for
amino acid analyses on a Beckman Spinco aminoacid analyzer that was supported by Public HealthService grant HE-11 19 from the National HeartInstitute.
This investigation was supported by the PublicHealth Service grant AI-04615 from the NationalInstitute of Allergy and Infectious Diseases and by aPublic Health Service training grant GM-626 fromthe Division of General Medical Science.We thank Gary K. Best, William G. Struve, and
Menard G. Heydanek for discussions and methods.
LITERATURE CITED
1. Anderson, J. S., M. Matsuhashi, M. A. Haskin,and J. L. Strominger. 1965. Lipid-phospho-acetylmuramyl-pentapeptide and lipid-phospho-disaccharide-pentapeptide: presumed mem-brane transport intermediates in the biosyn-
thesis of bacterial cell walls. Proc. Natl. Acad.Sci. U.S. 53:881-889.
2. Anderson, J. S., M. Matsuhashi, M. A. Haskin,and J. L. Strominger. 1967. Biosynthesis of thepeptidoglycan of bacterial cell walls. II. Phos-pholipid carriersI in the reaction sequence. J.Biol. Chem. 242:3180-3190.
3. Anderson, J. S., P. M. Meadow, M. A. Haskin,and J. L. Strominger. 1966. Biosynthesis of thepeptidoglycan of bacterial cell walls. Arch.Biochem. Biophys. 116:487-515.
4. Beers, R. F., Jr. 1955. Submerged culture ofMicrococcus lysodeikticus for large-scale pro-duction of cells. Science 122:1016.
5. Best, G. K., and N. N. Durham. 1965. Vancomy-cin adsorption to Bacillus subtilis cell walls.Arch. Biochem. Biophys. 111:685-691.
6. Best, G. K., and N. N. Durham. 1966. Adsorp-tion of the ristocetins to Bacillus subtilis cellwalls. Antimicrobial Agents and Chemother-apy-1965, p. 334-338.
7. Burger, M. M., and L. Glaser. 1964. The synthesisof teichoic acids. I. Polyglycerophosphate. J.Biol. Chem. 239:3168-3177.
8. Chatterjee, A. N., and H. R. Perkins. 1966. Com-pounds formed between nucleotides related tothe biosynthesis of bacterial cell wall andvancomycin. Biochem. Biophys. Res. Commun.24:489-494.
9. Chatterjee, A. N., J. B. Ward, and H. R. Perkins.1967. Synthesis of mucopeptide by L-formmembranes. Nature 214:1311-1314.
10. Czerkawski, J. W., H. R. Perkins, and H. J.Rogers. 1963. A study of the composition andstructure of the cell wall mucopeptide ofMicrococcus lysodeikticus. Biochem. J. 86:468-474.
11. Ghuysen, J. M., and M. R. J. Salton. 1960. Acetyl-hexosamine compounds enzymically releasedfrom Micrococcus lysodeikticus cell walls. I.Isolation and composition of acetylhexosa-mine and acetylhexosamine-pentide complexes.Biochim. Biophys. Acta 40:462-472.
12. Higashi, Y., J. L. Strominger, and C. C. Sweeley.1967. Structure of a lipid intermediate in cellwall peptidoglycan synthesis: A derivative of a
Ca5 isoprenoid alcohol. Proc. Natl. Acad. Sci.U.S. 57:1878-1884.
13. Jordan, D. C. 1961. Effect of vancomycin on thesynthesis of the cell wall and cytoplasmicmembrane of Staphylococcus aureus. Can. J.Microbiol. 11:390-393.
14. Jordan, D. C., and P. E. Reynolds. 1967. Vanco-mycin, p. 102-116. In D. Gottlieb, and P. L.Shaw (ed.), Antibiotics: mechanism of action,vol. 1, Springer-Verlag, Heidelberg.
15. Katz, W., M. Matsuhashi, C. P. Dietrich, and
J. L. Strominger. 1967. Biosynthesis of thepeptidoglycan of bacterial cell walls. IV. In-corporation of glycine in Micrococcus lyso-deikticus. J. Biol. Chem. 242:3207-3217.
16. Neuhaus, F. C., and WV. G. Struve. 1965. En-zymatic synthesis of analogs of the cell wallprecursor. I. Kinetics and specificity of uridinediphospho - N - acetylmuramyl - L - alanyl - D-glutamyl-L-lysine: D-alanyl-D-alanine ligase(adenosine diphosphate) from Streptococcusfaecalis R. Biochemistry 4:120-131.
17. Reynolds, P. E. 1961. Studies on the mode ofaction of vancomycin. Biochim. Biophys. Acta52:403-405.
18. Salton, M. R. J. 1964. The bacterial cell wall.Elsevier Publishing Co., Amsterdam.
19. Salton, M. R. J. 1967. Bacterial membranes, p. 77.In B. D. Davis, and L. Warren (ed.), Thespecificity of cell surfaces. Prentice-Hall, Inc.,Englewood Cliffs, N.J.
20. Salton, M. R. J., and J. G. Pavlik. 1960. Studiesof the bacterial cell wall. VI. Wall compositionand sensitivity to lysozyme. Biochiim. Biophys.Acta 39:398-407.
21. Sharon, N., T. Osawa, H. M. Flowers, and R. W.Jeanloz. 1966. Isolation and study of the chemi-cal structure of a disaccharide from Micrococcuslysodeikticus cell walls. J. Biol. Chem. 241:223-230.
22. Spackman, D. H., W. H. Stein, and S. Moore.1958. Automatic recording apparatus for use inthe chromatography of amino acids. Anal.Chem. 30:1190-1206.
23. Struve, W. G., and F. C. Neuhaus. 1965. Evidencefor an initial acceptor of UDP-NAc-muramyl-pentapeptide in the synthesis of bacterial muco-peptide. Biochem. Biophys. Res. Commun.18:6-12.
24. Struve, W. G., R. K. Sinha, and F. C. Neuhaus.1966. On the initial stage in peptidoglycansynthesis. Phospho-N-acetyl-muramyl-penta-peptide translocase (uridine monophosphate).Biochemistry 5:82-93.
25. Stickgold, R. A., and F. C. Neuhaus. 1967. Onthe initial stage in peptidoglycan snythesis.Effect of 5-fluorouracil substitution on phospho-N-acetyl-muramyl-pentapeptide translocase(uridine 5'-phosphate). J. Biol. Chem. 242:1331-1337.
26. Wallas, C. H., and J. L. Strominger. 1963. Ristoce-tins, inhibitors of cell wall synthesis in Staphy-lococcus aureus. J. Biol. Chem. 238:2264-2266.
27. Whitney, J. G. and E. A. Grula. 1964. Incorpora-tion of D-serine into the cell wall mucopeptideof Micrococcus lysodeikticus. Biochem. Biophys.Res. Commun. 14:375-381.