92


REVISTA MEXICANA DE CIENCIAS FORESTALESAntes Ciencia Forestal en México
ISSN: 2007-1132
M.C. Carlos Mallén Rivera EDITOR EN JEFE
Dr. Victor Javier Arriola Padilla SECRETARIO TÉCNICO
Dra. Cecilia Nieto de Pascual Pola M.C. Marisela C. Zamora MartínezCOORDINADORA EDITORIAL CURADORA DE PUBLICACIÓN
CONSEJO CONSULTIVO INTERNACIONAL
Dr. Celedonio Aguirre Bravo.- Forest Service, United States Department of Agriculture. Estados Unidos de NorteaméricaDra. Amelia Capote Rodríguez.- Instituto de Investigaciones Fundamentales en Agricultura Tropical. La Habana, Cuba
Dr. Carlos Rodriguez Franco.- US Forest Service Research and Development. Estados Unidos de NorteaméricaIng. Martín Sánchez Acosta.- Instituto Nacional de Tecnología Agropecuaria. Argentina
Dra. Laura K. Snook.- International Plant Genetic Resources Institute. Roma, Italia Dr. Santiago Vignote Peña.- E.T.S.I. de Montes, Universidad Politécnica de Madrid. España
CONSEJO CONSULTIVO NACIONAL
Dr. Salvador Fernández Rivera.- Coordinación de Investigación, Innovación y Vinculación, INIFAPDr. Miguel Caballero Deloya.- Fundador de la Revista Ciencia Forestal en México
Dr. Oscar Alberto Aguirre Calderón.- Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo LeónDr. Carlos Héctor Ávila Bello.- Vicerrectoría, Universidad Veracruzana
Dr. Francisco Becerra Luna, Centro de Investigación Regional – Centro, INIFAPDr. Robert Bye Boetler.- Jardín Botánico, Instituto de Biología, Universidad Nacional Autónoma de México
Dra. Amparo Borja de la Rosa.- División de Ciencias Forestales, Universidad Autónoma ChapingoDra. Patricia Koleff Osorio.- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad
Ing. Francisco Javier Musálem López.- Academia Nacional de Ciencias ForestalesDr. Juan Bautista Rentería Ánima.- Dirección de Soporte Forestal, INIFAP
Dra. María Valdés Ramírez.- Escuela Nacional de Ciencias Biológicas, Instituto Politécnico NacionalDr. Alejandro Velázquez Martínez.- Especialidad Forestal, Colegio de Postgraduados
La Revista Mexicana de Ciencias Forestales es una publicación científica del sector forestal del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Centro Público de Investigación y Organismo Público Descentralizado de la Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA). Tiene como objetivo difundir los resultados de la investigación que realiza el propio Instituto, así como la comunidad científica nacional e internacional en el ámbito de los recursos forestales. El contenido de las contribuciones que conforman cada número es responsabilidad de los autores y su aceptación quedará a criterio del Comité Editorial, con base en los arbitrajes técnicos y de acuerdo a las normas editoriales. Se autoriza la reproducción de los trabajos si se otorga el debido crédito tanto a los autores como a la revista. Los nombres comerciales citados en las contribuciones, no implican patrocinio o recomendación a las empresas referidas, ni crítica a otros productos, herramientas o instrumentos similares.
La Revista Mexicana de Ciencias Forestales está inscrita en el Índice de Revistas Mexicanas de Investigación Científica y Tecnológica, del Consejo Nacional de Ciencia y Tecnología (CONACYT). Es referida en el servicio de CABI Publishing (Forestry Abstracts y Forest Products Abstracts) de CAB International, así como en el Catálogo de Revistas
del Sistema Regional de Información en Línea para Revistas Científicas de América y El Caribe, España y Portugal (LATINDEX); en el Índice de Revistas Latinoamericanas en Ciencias (PERIÓDICA); en el Catálogo Hemerográfico de Revistas Latinoamericanas, Sección de Ciencias Exactas y Naturales (HELA) y en la Scientific Electronic Library Online (SciELO-México).
La Revista Mexicana de Ciencias Forestales Volumen 2, Número 8, noviembre-diciembre 2011, es una publicación bimestral editada por el Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). Av. Progreso No. 5, Barrio de Santa Catarina, Delegación Coyoacán, C. P. 04010, México D. F. www.inifap.gob.mx, [email protected]. Distribuida por el Centro Nacional de Investigación Disciplinaria en Conservación y Mejoramiento de Ecosistemas Forestales (CENID-COMEF). Editor Responsable. Carlos Mallén Rivera. Reservas de Derechos al Uso Exclusivo No. 04-2010-012512434400-102. ISSN: 2007-1132, otorgados por el Instituto Nacional del Derecho de Autor (INDAUTOR). Certificado de Licitud de Título y Licitud de Contenido: En trámite por la Comisión Calificadora de Publicaciones y Revistas Ilustradas de la Secretaría de Gobernación. Impreso por: Graphx, S.A. de C.V., Tacuba 40-205, Col. Centro Histórico, C.P. 06010, México, D.F. Este número se terminó de imprimir el 17 de noviembre de 2011, con un tiraje de 1,000 ejemplares.
Portada: Wangari Muta Maathai. Dominio público.

REVISTA MEXICANA DE CIENCIAS FORESTALES
Vol. 2 Núm. 8 noviembre - diciembre 2011
CONTENIDO
Página
EDITORIAL “WANGARI MUTA MAATHAI. “REMEMBRANZA POR SU CONTRIBUCIÓN AL DESARROLLO SOSTENIBLE, LA DEMOCRACIA Y LA PAZ”.
ARTÍCULOS MODELOS PARA ESTIMACIÓN Y DISTRIBUCIÓN DE BIOMASA DE Abies religiosa (Kunth) Schltdl. et Cham.EN PROCESO DE DECLINACIÓNBIOMASS ESTIMATION AND DISTRIBUTION MODELS OF Abies religiosa (Kunth) Schltdl. et Cham. IN DECLINEPatricia Flores-Nieves, Miguel Ángel López-López, Gregorio Ángeles–Pérez, María de Lourdes de la Isla-Serrano yGermán Calva-Vásquez
CARACTERÍSTICAS MORFOLÓGICAS DE PLÁNTULAS DE DOS ESPECIES FORESTALES TROPICALESPROPAGADAS EN CONTENEDORES BIODEGRADABLES Y CHAROLAS STYROBLOCKMORPHOLOGICAL CHARACTERISTICS OF SEEDLINGS OF TWO TROPICAL FOREST SPECIES PROPAGATED INBIODEGRADABLE CONTAINERS AND STYROBLOCK TRAYSH. Jesús Muñoz Flores, J. Jesús García Magaña, Víctor Manuel Coria Ávalos, Gabriela Orozco Gutiérrez yYadira Yolanda Muñoz Vega
HONGOS OPHIOSTOMATOIDES DE GALERIAS DE Dendroctonus adjunctus Blandford ENPinus hartwegii Lindl.OPHIOSTOMATOID FUNGI OF GALLERIES OF Dendroctonus adjunctus Blandford IN Pinus hartwegii Lindl.Omar Alejandro Pérez Vera, Dionicio Alvarado Rosales, David Cibrián Tovar, Armando Equihua Martínez yElizabeth Cárdenas Soriano
EVALUACIÓN DEL CARBONO ORGÁNICO EN SUELOS DE LOS MANGLARES DE NAYARITORGANIC CARBON ASSESSMENT IN MANGROVE SOILS OF NAYARITEduardo Valdés Velarde, Juan Ignacio Valdez Hernández, Víctor Manuel Ordaz Chaparro, Juan Fernando Gallardo Lancho,Joel Pérez Nieto y Ciriaco Ayala Sánchez
GERMINACIÓN DE ESPECIES DE CACTÁCEAS EN CATEGORÍA DE RIESGO DEL DESIERTO CHIHUAHUENSEGERMINATION OF CACTI SPECIES UNDER RISK STATUS OF THE CHIHUAHUAN DESERT Joel Flores y Enrique Jurado
LOS SUELOS DEL ESTADO DE MÉXICO Y SU ACTUALIZACIÓN A LA BASE REFERENCIAL MUNDIAL DELRECURSO SUELO 2006THE SOILS OF MEXICO STATE AND THEIR UPDATE TO THE WORLD REFERENCE BASE FOR SOIL RESOURCES 2006Erasto Domingo Sotelo Ruíz, Antonio González Hernández, Gustavo Cruz Bello, Francisco Moreno Sánchezy Gustavo Cruz Cárdenas
CONSEJO ARBITRAL
3
9
21
35
47
59
71
85

4
Carlos Galindo, (2011). Flor de Tulipan africano.

3
Editorial
“Wangari Muta Maathai. Remembranza por su contribución al
desarrollo sostenible, la democracia y la paz”.
«Un árbol tiene raíces en el suelo y ramas que rozan el cielo, recordándonos que
para prosperar debemos saber nuestro origen. Al igual que los árboles, por mucho
que lleguemos lejos, son nuestras raíces las que nos alimentan»
Wangari Muta Maathai
Wangari Muta Maathai, ganadora del premio Nobel de la Paz 2004, falleció el domingo 25 de septiembre de 2011, justo cuando en
la capital de México se clausuraba la más importante exposición forestal. Ese día lejos de guardarse un minuto de silencio, prevaleció
un profundo silencio sobre tan lamentable deceso. Hecho que en general fue compartido en el mundo -si quien hubiera
muerto hubiese sido un futbolista millonario o una popular cantante aficionada a las drogas, los medios se hubieran volcado
en la noticia-. Sin embargo, su influencia se ha sentido en todo el orbe, la Revista Time la declaró Héroe del Planeta, al sumar con
valentía a su causa a miles de africanos y millones de voluntades. Impulsó la participación colectiva, preponderantemente femenina,
en torno a los proyectos de reforestación como un instrumento para mejorar la calidad de vida de las comunidades rurales. Dedicó
su vida a defender el bosque, a otras mujeres y a la democracia. Pero sobre todo, ayudó a construir el concepto sostenible,
formulado por otra mujer: Norman Brundtland. De hecho su activismo político estuvo ligado a su labor de conservación. En diversos
foros preparatorios a la reunión de Johannesburgo, que marcaba los diez años de su similar de Río de Janeiro, se
expresaba la inquietud de no “africanizar” la cumbre. Qué lejos se estaba de imaginar que la solución a la crisis ambiental estaba en el
continente de donde precisamente surgió el género humano.
Michelle Bachelet, Directora Ejecutiva de ONU Mujeres, en homenaje a Wangari Muta Maathai declaró ese domingo
25 septiembre: “Nos unimos a muchos en África y en todo el mundo para llorar su fallecimiento y para celebrar su vida como líder
excepcional, primera mujer africana en recibir el Premio Nobel de la Paz. La Profesora Maathai se declaró, con valentía y siendo
víctima de acosos y ataques, a favor de la protección del medio ambiente y del progreso de los derechos de las mujeres, de la
lucha contra la desertificación, la escasez de agua y el hambre en el medio rural.” Bachelet exaltó sobre todo su figura de líder
extraordinaria que a través del impulso de la reforestación empoderó a miles de mujeres y alentó con pasión una nueva manera de
pensar y actuar, que combina la democracia y el desarrollo sostenible. Líder intrépida, fue donde nadie había osado ir y desafió a
autoridades a quienes pocos osaban desafiar. Rehusando ser intimidada, se mantuvo firme sobre la plena participación de las mujeres
en la vida cívica y pública: hoy nos deja un legado que permanecerá para siempre con nosotros. Sus ideas innovadoras sobre la
creación de empleo, gracias al restablecimiento medioambiental, forman parte de la agenda mundial de desarrollo en materia de
trabajos y de una economía verdes, en el contexto del desarrollo sostenible y de la erradicación de la pobreza. Para
Bachelet, la Dra. Maathai inspirará, especialmente, la preparación para la Conferencia de la ONU sobre Desarrollo Sostenible
que se celebrará en Rio de Janeiro en junio de 2012.
Madame Wangari fue una pionera desde su época universitaria, cuando obtuvo la licenciatura en Biología en Atchison, Kansas,
EE.UU. y al ampliar sus estudios en Pittsburgh, Alemania y en la Universidad de Nairobi, siendo en 1971, la primera mujer de
África Central y Oriental en obtener un doctorado. Nació en Nyeri, Kenia, madre de tres hijos, diputada y ministra adjunta para Medio
ambiente, Recursos Naturales y Vida Silvestre. También en el ámbito privado, rompió con una sociedad que relega a la mujer. Su
marido, un antiguo parlamentario, se divorció de ella en 1980 con el argumento de que “era demasiada educada, con amplio carácter
y éxito para poder controlarla”. Su mayor contribución fue el Movimiento Cinturón Verde de Kenia, un proyecto que impulsó

4
en 1977 y que combina la promoción de la biodiversidad
con la del empleo a mujeres. Productos de esta iniciativa son
30 millones de árboles plantados en su país y la ocupación
de 50 mil mujeres pobres en diferentes viveros. Desde 1986,
dicho movimiento originó una gran red panafricana que ha
llevado proyectos similares a países como Tanzania
y Etiopía. “Si uno desea salvar el entorno, primero hay que
proteger al pueblo. Si somos incapaces de preservar la especie
humana, ¿qué objeto tiene salvaguardar las especies
vegetales?”, declaró al resumir su filosofía, que expuso
más de una vez en la tribuna de la sede de la Organización
de las Naciones Unidas (ONU).
En su época como directora del Departamento de Anatomía
Veterinaria en Nairobi (1976-1977), empezó su actividad
en el Consejo Nacional de Mujeres de Kenia, organización
que presidió entre 1981 y 1987. Promotora de cancelar
la deuda externa del Tercer Mundo, destacó como decidida
opositora al régimen dictatorial de Daniel Arap Moi, por
lo cual, durante la década de 1990, se le detuvo y encarceló.
En 1997 fue candidata a la presidencia de Kenia, pero su
partido retiró su candidatura días antes de las elecciones. Un
año después, lideró la oposición a un proyecto gubernamental
de construcción en la selva lo que desencadenó una revuelta
popular que fue duramente reprimida por el gobierno, acto
que recibió el repudio nacional e internacional, en respuesta
a su convocatoria. Su compromiso se recompensó con
una profusión de premios, como el de Mujeres del Mundo
de Women Aid (1989), el de la Fundación Ecologista
Goldman (1991) –el llamado Nobel de los ecologistas–,
el Premio África de Naciones Unidas (1991) o el Petra Kelly
(2004). Para el Comité Nobel, la paz en la tierra depende
de la capacidad para asegurar el ambiente, y Maathai
se sitúa al frente de la lucha en la promoción del desarrollo
económico, cultural y ecológicamente viable de África,
con una visión planetaria de lo sostenible que abraza la
democracia, los derechos humanos y, en particular, los de
la mujer.
Pensaba de forma global y actuaba en el ámbito local, el
movimiento Cinturón Verde, es un programa que combina
existencia comunitaria y protección ambiental, al propagar entre
campesinos la idea básica de que plantar árboles mejora sus
vidas y la de sus hijos. Su quehacer en el mundo subdesarrollado
combinó ciencia, compromiso social y política activa. Proteger
bosques a través de la educación, la planificación familiar,
la nutrición y la lucha contra la corrupción. Su propuesta
también compete el otorgamiento de la responsabilidad de un
proceso de autogestión de las mujeres, que ni derecho a la
propiedad tenían en Kenia. Su lucha contribuyó a generar una
sensibilización de la población sobre su derecho a oponerse
al abuso del poder. La campaña contra la apropiación
ilegal de terrenos públicos y bosques, para su urbanización, le
granjeó la enemistad del presidente Daniel Arap Moi, quien la
calificó de “perturbada y amenaza a la seguridad del país”.
Y en otra vertiente, Wangari Muta Maathai es una expresión
de la poesía africana, una oportunidad para apurar en la
espiritualidad y la sabiduría de un continente que es, en sí
mismo, erupción milenaria -selva, desierto, ríos- almas que danzan
y veneran a sus antepasados. Pero también, muy lamentablemente,
una tierra ultrajada por la violencia y el racismo. Empero, a
pesar del intenso dolor y de las heridas continuas subsiste el
canto de sus poetas –y Wangari lo es–, que preservan
la belleza, la religiosidad y el nativo que nos vuelve a
enseñar a la nueva humanidad. En su misión llevó siempre el
mejorar la situación de las mujeres rurales pobres. Creyó en
la inmensa fuerza femenina y trabajó duro para movilizar a las
mujeres pobres. Esta creencia idealizada, la experta en
ciencias ambientales Lali tha la identifica en el poema
“Las mujeres se siembran” de Halldis Moren:
La mujer es la plantación de un árbol en el mundo.De rodillas, como si alguien en la oración,entre los restos de los muchos árbolesque la tormenta se ha roto.Ella debe intentarlo de nuevo, tal vez uno al fin se dejará de crecer en paz. Ella ve las manos extendidas sobre la tierra como si estuviera tratando de imponer la calma Por su temor. Oh muerte! La Tierra, estar quieto, Estad quietos, así que mi árbol puede crecer.
En su conferencia nobel esta mujer africana, primera en
recibir el Premio, declara que espera animar a mujeres y niñas
a levantar la voz y tener más espacio para el liderazgo.
Reconoce el trabajo de innumerables personas y grupos en
todo el mundo. Quienes trabajan en silencio y, a menudo, sin el
menor reconocimiento, protegiendo el ambiente,
promoviendo la democracia, defendiendo los derechos
humanos y garantizando la igualdad, sembrando semillas

5
de paz, a todos ellos les obsequia el premio, no solo para
que se sientan representados, sino para que lo utilicen en promover
su misión y cumplir con las expectativas de este mundo,
que será suyo. También agradece a sus conciudadanos
“…que permanecieron tercamente con la esperanza de que
la democracia pudiera hacerse realidad y su entorno gestionado
de forma sostenible…” reconociéndose producto de esta lucha.
Resalta Wangari en el Comité Nobel un reconocimiento de
que la paz pasa obligadamente por el desarrollo sostenible y
este se finca en la democracia, es una idea cuyo momento
ha llegado.
Recuerda como fue testigo, desde su infancia, de la
devastación de los bosques y la privatización de las tierras,
que acaban con la biodiversidad local y sus funciones
ecológicas. En 1977, cuando comenzó el Movimiento del
Cinturón Verde, en parte fue una respuesta a las necesidades
de las mujeres rurales, la falta de leña, agua potable, una
alimentación equilibrada, vivienda y de ingresos. A lo largo
de África, las mujeres son las cuidadoras primarias, con
responsabilidad significativa para el cultivo de la tierra y
alimentar a sus familias. Son las primeras en tomar conciencia
del daño ambiental, que vuelve a los recursos escasos y
originan que no puedan sostener a sus familias. Las respuestas
capitalistas, como la introducción de la agricultura comercial
y el comercio internacional controlado por el precio de las
exportaciones, no garantizan de forma razonable y justa el
ingreso de los agricultores a pequeña escala. Narra cómo llegó
a entender que cuando el ambiente es destruido o saqueado
se socava el futuro de las generaciones venideras.
Así, la plantación de árboles se convirtió en una opción
natural para hacer frente a necesidades básicas iniciales
identificadas por las mujeres. Además, la reforestación
es simple, asequible y garantiza resultados rápidos y exitosos
dentro de un plazo razonable. Esto mantiene el interés y el
compromiso. Así que, las mujeres plantaron más de 30 millones
de árboles que proveen de combustible, alimentos, vivienda
y los ingresos para apoyar la educación de sus hijos y las
necesidades del hogar. La actividad también generó
empleos y mejoró los suelos y las cuencas hidrográficas. A
través de su participación, las mujeres adquieren poder
sobre sus vidas y la de sus familias, especialmente de su
posición social y económica.
Prosigue en su conferencia, el trabajo era difícil porque
históricamente el pueblo había sido persuadido de que por
su condición de pobre, no solo carecía de capital, sino del
conocimiento y las habilidades para hacer frente a
sus desafíos; y en cambio, estaba condicionado a creer que
las soluciones a sus problemas debían venir de “afuera”.
Sin embargo, las mujeres se percataban de que la solución
de sus requerimientos depende de que su entorno esté sano y
bien gestionado; siendo conscientes –en carne propia- de que
un ambiente degradado conduce a una lucha por los recursos
escasos, que culmina en la guerra. Incluso intuían las injusticias de
los acuerdos económicos internacionales. Con el fin de ayudar
a las comunidades para entender estas relaciones, la galardonada
explica como se desarrolló un programa de educación
ciudadana, en el cual las personas identifican sus problemas,
las causas y posibles soluciones. Progresivamente, establecen
conexiones entre sus propias acciones y los problemas del medio
social. Se enteran como afrontar problemas: la corrupción, la
violencia contra las mujeres y los niños, la interrupción y
ruptura de las familias; así como, la desintegración de las
culturas y comunidades. También identifican el abuso
de drogas y sustancias enervantes, especialmente entre los
jóvenes, así como enfermedades devastadoras y que ocurren en
proporciones epidémicas, de particular preocupación son el SIDA,
el paludismo y la desnutrición. Así mismo, llama la atención sobre
la grave afectación humana por la destrucción generalizada
de los ecosistemas, empero de manera preponderante
por la inestabilidad climática y la contaminación en los suelos
y las aguas que acentúan una atroz pobreza. En el proceso,
se descubren parte de las soluciones, descubren su potencial
oculto encontrando amplias facultades para superar la inercia
y entrar en acción. Llegan a reconocer que los habitantes
rurales son los principales guardianes y beneficiarios del medio
ambiente que los sostienen. Comunidades enteras también
llegan a entender que, si bien, es necesario responsabilizar a
sus gobiernos, es igualmente importante, que en sus relaciones
con los demás, ejemplifiquen los valores de liderazgo que
desean ver en sus propios dirigentes, es decir, la justicia, la
integridad y la confianza. Aunque inicialmente las actividades
del Movimiento del Cinturón Verde no se refirieron a
los problemas de la democracia y la paz, pronto quedó claro
que el manejo responsable del ambiente era imposible sin un
amplio espacio democrático. Por lo tanto, el árbol se convirtió
en un símbolo de la lucha democrática en Kenia.

6
Los ciudadanos se movilizaron para desafiar los abusos
generalizados, verbigracia, por la incompetente gestión de
la energía. Simbólicamente en el parque Uhuru de Nairobi,
se plantó un árbol de la paz para exigir la liberación de los
prisioneros de conciencia y de una transición pacífica hacia la
democracia. Miles fueron movilizados, facultados para tomar
medidas y aprendiendo a superar el miedo para defender sus
derechos ciudadanos. Con el tiempo, el árbol también
se convirtió en un símbolo de la paz, especialmente, durante los
conflictos étnicos en Kenia. El árbol como símbolo de
armonía está en consonancia con una tradición generalizada
de África. Por ejemplo, los ancianos de Kikuyu llevaban un
bastón del árbol thigi que, cuando se coloca entre dos partes
contendientes, los lleva a la pacificación y la reconciliación.
Estas prácticas son parte de un rico patrimonio cultural, que
contribuye tanto a la conservación de los hábitats como
de las culturas. Pero con la introducción de nuevos valores,
la biodiversidad local no es valorada y, en consecuencia, se
degrada y desaparece. Por esta razón, el Movimiento Cinturón
Verde explora el concepto de la biodiversidad cultural,
especialmente, con respecto a las semillas autóctonas y a las
plantas medicinales.
A medida que se entendieron las causas de la degradación
ecológica, se vió la necesidad de una buena gobernanza,
de hecho el estado ambiental en cualquier localidad es su
reflejo. Naciones que cuentan con sistemas de mala gestión es
probable que tengan conflictos y leyes pobres para proteger
el medio ambiente. En 2002, el valor, resistencia, paciencia y
compromiso de los miembros del Movimiento Cinturón Verde, de
otras organizaciones de la sociedad civil y del público en Kenia
culminó en la transición pacífica a un gobierno democrático y
sentaron las bases para una sociedad más estable.
Cuando se otorgó el premio Nobel de la Paz en 2008
se estaba a 30 años de que se inició este trabajo. Sin
embargo, las actividades que devastan el medio ambiente y
las sociedades continuaban sin disminuir. La profesora Maathai
sentencia: “Hoy en día nos enfrentamos a un desafío que
exige un cambio en nuestra forma de pensar, de modo que la
humanidad deje de amenazar su sistema de soporte vital.
Estamos llamados a ayudar a la tierra a curar sus heridas
y en el proceso de sanar las nuestras - de hecho- para abrazar
la creación en toda su diversidad, belleza y maravilla. Esto
sucederá si vemos la necesidad de reactivar nuestro sentido de
pertenencia a una familia más grande de la vida, con
los que hemos compartido nuestro proceso evolutivo. En el
curso de la historia, llega un momento en que la humanidad
debe cambiar a un nuevo nivel de conciencia, para llegar a
un grado moral más alto. Un momento en que tenemos que
arrojar el miedo y dar esperanza a los demás. Ese momento es
ahora. El Comité Noruego del Nobel ha desafiado al mundo
para ampliar la comprensión de la paz: no puede haber paz
sin desarrollo equitativo, y no puede haber desarrollo sin una
gestión sostenible del medio ambiente en un espacio democrático
y pacífico. Este cambio es una idea cuyo momento ha llegado.”
Pidió a los líderes, especialmente de África, ampliar el
espacio democrático y construir sociedades justas y equitativas
que permitan la creatividad y la energía de sus ciudadanos a
prosperar: “Aquellos de nosotros que hemos tenido el privilegio
de recibir educación, habilidades y experiencias e
incluso alimentación debemos ser un modelo para la próxima
generación de liderazgo. La cultura juega un papel central
en la vida política, económica y social de las comunidades,
puede ser el eslabón perdido en el desarrollo de África. La
cultura es dinámica y evoluciona con el tiempo, descartando las
tradiciones retrógradas, como la mutilación genital femenina”.
Y no puede dejar de recordar sus orígenes y alecciona a los
africanos, a descubrir los aspectos positivos de su cultura. En
la aceptación de ellos, se daría un sentido de pertenencia,
identidad y confianza en sí mismo. Hace un llamado a
los gobiernos a reconocer el papel de los movimientos
sociales en la construcción de una masa crítica de
ciudadanos responsables, que ayuden a mantener
el equilibrio de poderes en la sociedad. Por su parte, la
sociedad civil debe abarcar no solo sus derechos, sino también
sus responsabilidades. Además, la industria y las instituciones
globales deben apreciar que garantizar la justicia económica,
la equidad y la integridad ecológica son de mayor
valor que las ganancias a cualquier costo. Las desigualdades
mundiales extremas, los patrones dominantes de consumo
siguen a expensas del medio ambiente y la coexistencia pacífica.
También ruega la atención de los jóvenes a comprometerse en
actividades que contribuyan a alcanzar sus sueños de largo
plazo. “Ellos tienen la energía y creatividad para dar forma a
un futuro sostenible. A los jóvenes les digo, que son un regalo
para sus comunidades y para el mundo. Ustedes son nuestra

7
esperanza y nuestro futuro. El enfoque holístico del desarrollo,
como lo demuestra el Movimiento Cinturón Verde, podría ser
adoptado y replicado en varias partes de África, y más allá.”
Y volvió a su niñez cuando bebía agua de los ríos, jugaba
entre las hojas de “arrurruz” y trataba de atrapar
renacuajos. Ahora ese mundo de sus padres ya no es el de sus
nietos. Las corrientes se han secado, las mujeres caminan
largas distancias para conseguir agua, no siempre limpia, y los
niños nunca sabrán lo que han perdido. “El desafío consiste en
restaurar la casa de los renacuajos y devolver a nuestros hijos
un mundo bello y maravilloso”.
En una entrevista esta noble científica social declaró: “Es a
causa de la conducta inmoral, que hay contaminación en todas
partes; el aire, el agua, la tierra y los alimentos, se han visto
gravemente corrompidos debido a una conducta impropia.
La contaminación se podría haber evitado. La fuerte
caída de las virtudes como el amor, la compasión y la
tolerancia es directamente responsable de la degradación
que uno ve hoy en día. De hecho, incluso se podría decir que
estos elementos tienen miedo del hombre. La impureza
se refleja en la contaminación exterior. La buena conducta debe ser
la base real para la vida. Sin embargo, el hombre moderno
carece totalmente de carácter y virtudes. No es de extrañar la
paz y la felicidad se le escapan.”
La Dra.Wangari Maathai promovió iniciativas que
incluyeron la protección de los ecosistemas tropicales de la
cuenca del Congo, siendo embajadora itinerante de
esta región de acceso global para la diversidad biológica
y penosamente amenazados por la tala ilegal, la exploración
minera, la caza furtiva y el comercio de animales silvestres.
En la construcción de su cruzada para proteger los
bosques, la profesora Maathai se enteró de un plan del
gobierno para privatizar grandes áreas de tierra en el bosque
Karura. Ella protestó en contra de un plan de industrialización
del Congo, como lo hizo por muchos otros proyectos de este
talante, incluso con el visto bueno de gobiernos y sociedad,
decía: “No podemos desarrollar nuestros países, si vamos
a seguir la corrupción en ambos lados”. Ella tenía notable
tolerancia, paciencia y perseverancia, tomó enormes
riesgos personales para luchar por la justicia, así como por
los derechos humanos y del medio ambiente. Ella consideró
que los problemas primero se resolvían con la fuerza mental,
también creía en la asociación humana y que la felicidad se
encontraba en la unión. Una vez le preguntaban a un científico
que la conoció ¿cuál creía que era el secreto de esa fuente
de inspiración interminable? Y él respondió: La profesora
Maathai retrata el amor en cada acción suya. El amor era
el aliento mismo del nudo de unión natural con la progenie
verde. Cada lucha que acometió pone de relieve el amor puro
e incondicional y sin mancha que formó la base de sus obras
altruistas. Su vida y su obra representan la frase “El amor es el
desinterés y el yo es falta de amor ‘.
Cuando recibió la llamada, con la que le comunicaron que
había recibido el premio nobel, Wangari detuvo el automóvil
en un hotel que encontró de camino a uno de tantos pueblos
que apoyaba con sus proyectos de conservación forestal y
pidió permiso para plantar un árbol que celebrara el
acontecimiento. Ese árbol fue un Tulipán Africano.
Carlos Mallén Rivera
Editor en Jefe
Carlos Galindo (2010) Flor de Tulipan africano.
Spathodea campanulata Deauv
“The living conditions of the poor must be improved if we
really want to save our environment”
Wangari Maathai, Premio Nobel de la Paz 2004.

8
Wangari Muta Maathai. Dominio público.

MODELOS PARA ESTIMACIÓN Y DISTRIBUCIÓN DE BIOMASA DEAbies religiosa (Kunth) Schltdl. et Cham. EN PROCESO DE DECLINACIÓN
BIOMASS ESTIMATION AND DISTRIBUTION MODELS OF
Abies religiosa (Kunth) Schltdl. et Cham. IN DECLINE
Patricia Flores-Nieves 1, Miguel Ángel López-López 1, Gregorio Ángeles–Pérez 1,María de Lourdes de la Isla-Serrano 2 y Germán Calva-Vásquez 3
RESUMEN
Actualmente, el gobierno federal promueve el pago por captura de carbono en los bosques para mitigar el calentamiento global. Por ello, existe la necesidad de estimar la biomasa y tasas de captura de carbono en los bosques de Abies religiosa. En este estudio se generaron modelos para calcular la biomasa de fuste, de madera de ramas y de acículas de árboles completos y de ramas de oyamel en rodales del Cerro Tláloc, Texcoco, Estado de México. Para el caso de árboles completos, la biomasa estuvo en función del diámetro normal (DN), mientras que para ramas individuales fue una función del diámetro basal (DB). El uso de los modelos generados a nivel árbol se recomienda para árboles con DN entre 12 y 105 cm, n cambio los modelos a nivel de ramas se recomiendan para diámetros basales entre 1 y 120 mm. El modelo para fustes fue de tipo potencial y los de madera de ramas y follaje de árboles completos de tipo exponencial. Los coeficientes de determinación de los modelos a nivel árbol indican que existe una considerable variabilidad, principalmente, en las biomasas de ramas, lo que puede ser consecuencia del proceso de declinación en la zona de estudio. La biomasa total de los árboles se distribuye en forma atípica entre los compartimentos aéreos: 97 % en los fustes, 3 % en madera de ramas y 0.07 % en follaje. Este patrón de distribución de biomasa refleja el efecto del deterioro de los bosques de A. religiosa.
Palabras clave: Fuste, rama, acículas, ecuación alométrica, oyamel, productividad primaria neta.
ABSTRACT
At present, the federal government promotes a payment for carbon sequestration by forests to mitigate global change. This is why there exists a need for estimating biomass and carbon sequestration rates in Abies religiosa forests. In this study stem, branch wood and needle biomass estimation models were generated for whole A. religiosa trees and branches in stands at Cerro Tláloc, Texcoco, Estado de México. For whole trees biomass was a function of breast height diameter (BHD), and for individual branches, biomass was a function of basal diameter (BD). Use of tree-level models is suitable for trees with BHD within the range of 12 and 105 cm, and branch-level models are recommended for BD within 1 and 120 mm. Stem models were potential while tree-level branch wood and foliage ones were exponential. Determination coefficients for whole-tree models show that there exists a large variability, especially for branch biomass, which is likely to be a result of the decline process taking place in the study area. Total tree biomass is atypically distributed within aboveground tree components: 97 % corresponds to stems, 3 % to branch-wood and 0.07 % to foliage. This biomass distribution pattern may be a reflection of the decline process affecting A. religiosa in the study area.
Key words: Stem, branch, needle, allometric equation, Sacred-fir, net primary productivity.
Fecha de recepción: 5 de febrero 2010Fecha de aceptación: 25 de julio de 2011
1 Posgrado Forestal, Colegio de Posgraduados. Correo-e: [email protected] Posgrado de Hidrociencias, Colegio de Posgraduados.3 Facultad de Estudios Superiores Zaragoza, UNAM.

Rev. Mex. Cien. For. Vol. 2 Núm. 8
10
INTRODUCCIÓN
La conversión de los bosques maduros en áreas abiertas para diferentes usos económicos del suelo, como agricultura o ganadería, es un fenómeno que se ha presentado a lo largo de los últimos siglos (Herrera et al., 2001). Los bosques están dentro de los ecosistemas naturales con mayor producción primaria neta. Se calcula que producen de 400 a 1,000 g m-2 año-1 de carbono, cantidad dos veces superior a la de los pastizales y varias veces mayor que la de los océanos (Waring y Schlesinger, 1985). Una parte de esta producción se acumula como biomasa y humus, que constituyen la producción primaria neta. La otra parte se destina a procesos de mantenimiento del ecosistema, a través de la respiración (Escandón et al., 1999).
Garzuglia y Saket (2003) definen la biomasa aérea como la cantidad total de materia orgánica aérea presente en los árboles, e incluyen hojas, ramas, tronco principal y corteza. Para la estimación de biomasa, el procedimiento más común consiste en el muestreo destructivo de unos cuantos árboles para después modelar el contenido de biomasa en función de variables fáciles de medir, para ello se utilizan métodos de regresión (Díaz et al., 2007).
La biomasa se ha convertido en un elemento importante en los estudios sobre los cambios que ocurren a escala mundial, dado el posible efecto atenuador (sumidero de carbono) que los bosques pueden tener al secuestrar los excedentes de los gases de efecto invernadero, de un modo temporal (biomasa) y permanente (suelo) y a las consecuencias que se derivan de la modificación de las condiciones climáticas sobre la salud, estructura y biodiversidad de un sistema forestal (Vidal et al . , 2004).
Las ecuaciones de biomasa permiten estimar, con bastante exactitud, el peso de las especies forestales a partir de un número reducido de parámetros de los árboles en pie (López y Keyes, 1987; Castellanos et al., 1996; Rojo et al., 2005). Sin embargo, la disponibilidad de modelos para coníferas como Abies religiosa (Kunth) Schltdl. et Cham. es reducida. En México se han desarrollado ecuaciones para Pinus montezumae Lamb. (Garcidueñas, 1987; Manzano et al., 2007; López-López et al., 2009), Pinus cembroides Zucc. (López y Keyes, 1987), Pinus patula Schltdl. et Cham. (Castellanos et al., 1996), Alnus glabrata Fernald, Clethra hartwegii Britt., Rapanea myricoides (Schltdl.) Lundell, Quercus peduncularis Née, Liquidambar macrophylla Oerst e Inga sp. (Acosta et al., 2002), Hevea brasiliensis Müll. Arg. (Rojo et al., 2005) y para Abies religiosa (Avendaño et al., 2009).
Los modelos para el cálculo de la biomasa de árboles están influenciados por los cambios en la estructura, como consecuencia del manejo, presencia de plagas y enfermedades, condiciones climáticas, de suelo o incluso factores genéticos.
INTRODUCTION
The conversion of old forests into open areas for different economic land use such as agriculture or livestock farming has occurred during the last centuries (Herrera et al., 2001). Forests are one of the natural ecosystems with highest net primary production. It has been calculated that they produce from 400 to 1,000 g m-2 year-1 C, which is an amount twice higher than grasslands and several more than that of oceans (Waring and Schlesinger, 1985). One part of production accumulates as biomass and humus, which constitute the net primary production. The other part is bound to maintenance process of the ecosystem, through respiration (Escandón et al., 1999).
Garzuglia and Saket (2003) define aerial biomass as the total amount of aerial organic matter in trees, including leaves, branches main stem and bark. The most regular procedure for biomass estimation is destructive sampling of some trees for a later modeling of biomass content in terms of easily measurable variables, in which regression methods are used (Díaz et al., 2007).
Biomass has become an important element in studies about changes that occur world-wide, from the possible mitigating effect (carbon sink) of forests when they perform the sequestration of the exceeding greenhouse- effect gases, in a temporary (biomass) or permanent (soil) way and to the consequences of the changing climate conditions upon health, structure and biodiversity of a forest system (Vidal et al., 2004).
Biomass equations allow a rather accurate estimation of the weight of forest species from a small number of standing-tree parameters (López and Keyes, 1987; Castellanos et al., 1996; Rojo et al., 2005). However, the availability of models for softwoods such as Abies religiosa (Kunth) Schltdl. et Cham. is rather small. Some equations have been developed in Mexico for Pinus montezumae Lamb. (Garcidueñas, 1987; Manzano et al., 2007; López-López et al., 2009), Pinus cembroides Zucc. (López y Keyes, 1987), Pinus patula Schltdl. et Cham. (Castellanos et al., 1996), Alnus glabrata Fernald, Clethra hartwegii Britt., Rapanea myricoides (Schltdl.) Lundell, Quercus peduncularis Née, Liquidambar macrophylla Oerst e Inga sp. (Acosta et al., 2002), Hevea brasiliensis Müll. Arg. (Rojo et al., 2005), as well as for Abies religiosa (Avendaño et al., 2009).
Models for tree biomass are influenced by changes in structure, as a consequence of management, plagues and diseases, climate, soil and even some genetic factors.
Forest decline is defined as a multifactor sickness caused by biotic and abiotic agents that generates a decrease of vigor and survival of trees (Granados et al., 2001 and Vázquez et al., 2004). Ciesla (1989) and Granados et al. (2001) point out that there are symptoms related to this phenomenon, among

Flores-Nieves et al., Modelos para estimación y distribución...
11
La declinación forestal se define como una enfermedad multifactorial causada tanto por agentes abióticos como bióticos, lo que implica la reducción del vigor y supervivencia de los árboles (Granados et al., 2001 y Vázquez et al., 2004). Ciesla (1989) y Granados et al. (2001) y López et al. (2006) señalan que son varios los síntomas relacionados con dicho fenómeno, tales como la reducción del crecimiento, degeneración de los sistemas radicales, clorosis en el follaje, reducción de las reservas fotosintéticas, mortalidad de brotes y ramas e incremento de la incidencia de ataques de insectos, que se presentan de manera secuencial o simultánea. Alvarado et al. (1993) citan que puede alterarse la forma cónica de la copa y generarse defoliación severa y muerte de ramas en las partes bajas del árbol. Al momento de modelar las variables de los árboles o ecosistemas sujetos a este proceso de declinación, se espera que todos los síntomas mencionados tengan un efecto directo en los parámetros de los modelos generados.
El objetivo de este estudio fue aportar nuevos modelos para la estimación de biomasa de los componentes aéreos de A. religiosa en bosques afectados por el proceso de declinación, como es el caso de algunos bosques cercanos al Valle de México. Se pretende que las ecuaciones sirvan como herramienta para la realización de estudios de productividad primaria neta, captura de carbono y de distribución de biomasa en bosques que se encuentren en dicha condición.
MATERIALES Y MÉTODOS
Área de estudio
El presente trabajo se llevó a cabo en el Cerro Tláloc, en la región fisiográfica conocida como Sierra Nevada, que se ubica al oriente del Estado de México (Figura 1). La elevación del cerro es de 4,120 m y el área de estudio se localiza en la vertiente occidental del mismo, en dónde la especie dominante es A. religiosa.
Clima.- A lo largo del declive occidental del Cerro Tláloc se distinguen tres subtipos climáticos: en las áreas planas más cercanas a los lomeríos el clima es C (w
0) (w) b(i´); templado
subhúmedo con una precipitación media anual de 700 mm, con régimen de lluvia en verano, temperatura media anual entre 12 y 18 °C y con una oscilación térmica de 5 a 7 °C. En la zona de lomeríos, hacia las estribaciones de la Sierra de Río Frío el clima es C (w
1) (w) b(i´); templado subhúmedo, con una
precipitación media anual entre 800 y 900 mm, régimen de lluvias en verano, con temperatura media anual entre 12 y 18 °C y con una oscilación térmica de 5 a 7 °C. En las laderas montañosas el clima es de tipo C(w
2) (w) b(i´); templado subhúmedo, con una
precipitación media anual entre 900 y 1,200 mm, régimen de lluvias en verano, temperatura media anual de 10 y 14 °C, con una oscilación térmica de 5 a 7 °C. Las fluctuaciones climáticas se deben a la orografía (Ortíz y Cuanalo, 1977).
which are a reduction in growth, degeneration of the root system, foliage chlorosis, diminishment of photosynthetic reserves, shoot and branch mortality and an increment of insect attack, all of which could act sequentially or simultaneously. Alvarado et al. (1993) stated that the conic shape of the crown can change and generate an intensive defoliation and death of the lower branches. At the time of modeling the tree variables or ecosystems subjected to decline, it is expected that all the aforementioned symptoms have a direct effect on the parameter of the generated models.
The aim of this study was to provide new models for biomass estimation of the aerial components of A. religiosa in forests in decline, as some nearby Valle de Mexico are. It is expected that these equations might be useful to accomplish net primary productivity, carbon sequestration and of biomass distribution studies in forests that are in this condition.
MATERIALS AND METHODS
Study area
The study here described was carried out in Cerro Tláloc (Tlaloc Hill), in Sierra Nevada that lies at the East of Estado de México (Figure 1). The highest altitude is 4,120 m and the study area is located in the western hillside, where A. religiosa is the dominant species.
Climate.- Along the western fall of Cerro Tlaloc, three climate subtypes can be found: in the closest plains to the hills, the type of weather is C (w
0) (w) b(i´), which is a subhumid temperate
climate with 700 mm of annual mean precipitation, summer rainfall, annual average temperature between 12 and 18 °C and a thermic oscilation from 5 to 7 °C. In the hill zone, nearby Río Frio Mountain Range, the weather is C (w
1)
(w) b(i´) which is a subhumid temperate climate with 800-900 mm of annual mean precipitation, summer rainfall, annual average temperature between 12 and 18 °C and a thermic oscilation from 5 to 7 °C. In the hillsides, the weather is of the C(w
2) (w) b(i´) type,
which is a subhumid temperate climate with 900 and 1200 mm of annual mean precipitation, summer rainfall, annual average temperature between 10 and 18 °C and a thermic oscilation from 5 to 7 °C Climatic fluctuations are due to orography (Ortíz and Cuanalo, 1977).
Soils.- According to Mooser (1975), the edaphic material of Valle de México was the result of the volcanic activity of the Tertiary and Quaternary periods. The formations of the middle Tertiary (Oligocene and Miocene) are located from Tláloc up to near San Pablo Ixayoc town. Soils are incipient, of coarse texture near the cineritic cone of Tláloc, and in the resting areas, they are black, deep, of medium texture and rich in organic matter and with medium texture (migajones o francos).

Rev. Mex. Cien. For. Vol. 2 Núm. 8
12
Suelos.- De acuerdo con Mooser (1975) el material edáfico que constituye el Valle de México fue producto de erupciones ocurridas en el terciario y cuaternario. Las formaciones del terciario medio (oligoceno-mioceno) se localizan desde el Tláloc hasta cerca del poblado de San Pablo Ixáyoc. Los suelos son incipientes, de textura gruesa en las proximidades del cono cinerítico del Tláloc y en las áreas restantes son negros, profundos, muy ricos en materia orgánica y de textura media (migajones o francos).
Vegetación.- En la zona alta de la montaña se desarrolla el bosque y el pastizal de altura. En la parte más alta de esta zona se tiene bosque de Pinus hartwegii Lindl., Garrya laurifolia Benth. y Juniperus monticola Martínez. Más abajo, se observa el bosque de Abies religiosa y Quercus spp. (Palma, 1996). En la zona media de lomeríos se desarrolla bosque de Pinus leiophylla Schiede ex Schltdl. et Cham., Pinus montezumae Lamb., Pinus pseudostrobus Lindl., Pinus rudis Endl. y Cupressus lindleyi Klotzsch ex Endl. En sitios húmedos la vegetación está constituida por Alnus jorullensis Benth., Salix bonplandiana Kunth, Fraxinus udhei (Wenz.) Lingelsh., Buddleia cordata Kunth, Prunus serotina Ehrh., Taxodium mucronatum Ten. y Senecio salignus DC.
Vegetation.- In the mountain zone forest and highland grasslands grow. In the highest part of the zone, there is a forest of Pinus hartwegii Lindl., Garrya laurifolia Benth. and Juniperus monticola Martínez. In a lower area Abies religiosa and Quercus spp. forests are found (Palma, 1996). In the middle of hills Pinus leiophylla Schiede ex Schltdl. et Cham., Pinus montezumae Lamb., Pinus pseudostrobus Lindl., Pinus rudis Endl. and Cupressus lindleyi Klotzsch ex Endl. forests grow. In humid places, vegetation is made-up by Alnus jorullensis Benth., Salix bonplandiana Kunth, Fraxinus udhei (Wenz.) Lingelsh., Buddleia cordata Kunth, Prunus serotina Ehrh., Taxodium mucronatum Ten. and Senecio salignus DC.
Models for total tree biomass
In order to generate the total tree biomass models 10 individuals of different size were selected, that included the greatest diameter variability in the zone (12 to 105 cm). Normal diameter was measured to the selected trees with a diametric tape.
Afterwards, total heights were measured with a 50 m longimeter branches were removed and the stem was divided into logs of commercial size (2.55 m de long) by using a Truper
Figura 1. Localización del área de estudio. Cerro Tláloc, Texcoco, Estado de México.Figure 1. Location of the study area (Cerro Tlaloc, Texcoco, Estado de Mexico State).

Flores-Nieves et al., Modelos para estimación y distribución...
13
Modelos para biomasa de árboles completos
Para generar los modelos de biomasa de árboles completos se seleccionaron 10 individuos de diferentes dimensiones, que incluyeron la mayor variabilidad de diámetros presentes en la zona (12 a 105 cm). A los árboles ejemplares, se les midió el diámetro normal (DN) con una cinta diamétrica y posteriormente fueron derribados.
En seguida se le determinó su altura total con un longímetro de 50 m, se desramó y el fuste fue seccionado en trozas de dimensiones comerciales (2.55 m de longitud), con una usando una motosierra marca Truper con espada de 20”. Durante el troceo del fuste se obtuvo una rodaja de aproximadamente cinco centímetros de espesor como muestra de cada troza. A cada troza se le midieron los diámetros inferior y superior con una cinta diamétrica marca Forestry Suppliers Metric Fabric Diameter Tape, modelo 283D/160 cm, con el objeto de estimar su volumen. El diámetro de la base de cada una de las ramas del árbol se midió con un vernier digital marca Mitutoyo.
En campo se obtuvo el peso húmedo de cada una de las trozas (cuando fue posible), mediante una báscula marca Ohaus de 100 kg de capacidad y 50 g de precisión. Cuando estas pesaban más de 100 kg en húmedo, solo se determinó su volumen con la fórmula de Smalian:
V = (B + b) / 2*L
Donde:V = Volumen de la troza (m3)B, b = Área de la sección mayor y menor de la troza (m2)L = Longitud de la troza (m)
Las rodajas del fuste se trasladaron en bolsas de polietileno al laboratorio del Postgrado Forestal del Campus Montecillo del Colegio de Postgraduados, para secarlas a 80 °C, hasta alcanzar peso constante en un horno de circulación forzada marca Felisa modelo F20. Posteriormente se determinó la relación entre el peso seco y peso húmedo. Esta proporción se aplicó al peso húmedo total de las trozas previamente pesadas en campo, para determinar su biomasa.
Cuando las trozas pesaron más de 100 kg, el cálculo de su biomasa se realizó de manera indirecta, a partir de la estimación de la densidad de la madera. Las rodajas extraídas de cada troza se utilizaron para calcular la densidad de la madera de la troza correspondiente.
En el laboratorio, las rodajas (o muestras de ellas en el caso de rodajas muy grandes) se impermeabilizaron con parafina para obtener su volumen mediante el principio de Arquímedes. Para ello se usaron recipientes con agua de tamaño adecuado
power saw with a 20” blade. During these operations, a 5 cm thick slice from each log was taken as a sample. To each log were measured the lower and upper diameters with a diametric tape, in order to estimate its volume. The diameter of the base of each one of the branches was measured with a Mitutoyo digital vernier.
In the field was obtained the fresh weight of each log (when it was possible), with an Ohaus 100 kg scale and 50 g precision. When their wet weight was over 100 kg, volume was only determined with the Smalian formula:
V = (B + b) / 2*L
Where:
V = Volume of the long (m3)B, b = Area of the greatest and smallest section of the log (m2)L = Length of the log (m)
Stem slices were kept into plastic bags and were taken to the Forest Graduate Studies Laboratory at Montecillo Campus that belongs to the Graduate Studies School (Colegio de Postgraduados), where they were dried at 80 °C, until they reached a constant weight in a forced- circulation kiln marca y modelo. Y por cuánto tiempo Afterwards, the dry weight/wet weight relation was determined. This ratio was applied to the total wet weight of the logs previously weighted at the field, in order to determine biomass.
When logs were weighed more than 100 kg, the determination of biomass was carried out in an indirect way, from the estimation of wood density. The slices taken from each log were used to calculate the wood density of the corresponding log.
In the laboratory, the slices (or samples of them when slices were too big) were waterproofed with paraffin in order to get their volume by the Archimedes principle. For that ending, containers with water of a right size for the sample volume, which were placed on the 100 kg Ohaus scale of 50 g precision, when big samples are involved. When samples were small, a 2 kg Ohaus and precision of tenth of gram was used. Once the volume is had, weight was determined in order to estimate their density by the following formula:
δm = m/v
Where:δm = Density of wet wood (kg m-3)m = Mass (kg)v = Volume (m3)

Rev. Mex. Cien. For. Vol. 2 Núm. 8
14
para el volumen de las muestras, se colocaron sobre una báscula Ohaus de 100 kg de capacidad y 50 g de precisión, en el caso de muestras grandes. En el caso de las muestras pequeñas se utilizó una balanza Ohaus de dos kilogramos de capacidad y un décimo de gramo de precisión. Una vez que se obtuvo el volumen se procedió a determinar su peso, para estimar su densidad mediante la fórmula:
δm = m/v
Donde:
δm = Densidad de la madera en húmedo (kg m-3)m = Masa (kg)v = Volumen (m3)
La biomasa de cada muestra o rodaja se determinó colocando las muestras en un horno de secado de circulación forzada marca Felisa modelo F20 a 80 °C, para eliminar la parafina. Una vez conocidos la densidad de las rodajas y el volumen de las trozas correspondientes, se calculó la biomasa de las trozas mediante la fórmula:
Bt = δt vt
Donde:
Bt = Biomasa de troza (kg) δt = Densidad (kg m-3)vt = Volumen de la troza (m3)
Con los datos de biomasa de fustes se corrió un procedimiento de regresión, para generar un modelo de estimación de biomasa de ese compartimento en función del diámetro normal.
Bf = f (DN)
Donde:
Bf = Biomasa de fuste (kg)DN = Diámetro normal (cm)
La biomasa de madera de ramas y del follaje se estimó a partir del diámetro basal de la rama, para lo cual se utilizaron modelos previamente elaborados por el procedimiento que se describe a continuación.
Modelos para biomasa a nivel de ramas
El modelo para estimar la biomasa de madera de ramas individuales se elaboró a partir de 35 ramas de diversos diámetros basales (1 a 120 mm), procedentes de varios árboles del área de estudio. A las ramas recolectadas se les determinó
*
The biomass of every sample or slice was determined by placing the samples into a marca y modelo drying forced-circulation kiln at 80 °C in order to remove the paraffin. Once the slice density and the volume of their logs were known, the biomass of the logs was calculated by the following formula:
Bt = δt vt
Where:
Bt = Log biomass (kg)δt = Density (kg m-3)vt = Volume of the log (m3)
With the stem biomass data a regression was run to generate a biomass estimation model of that compartment in terms of the normal diameter:
Bf = f (DN)
Where:
Bf = Stem biomass (kg)DN = Normal diameter (cm)
Branch wood and foliage biomass was estimated from the basal branch diameter; thus, previously made models were used by the procedure was described as follows.
Models for biomass at a branch level
The model to estimate wood biomass of individual branches was designed from 35 branches of different basal diameters (1 to 120 mm), taken from several trees of the study area. To the collected branches, basal diameter was calculated with a Mitutoyo digital vernier caliper, and from them, leaves were pulled apart in the field, and were weighted by a Ohaus ES100L scale in wet, wood and foliage separately. When branches surpassed the capacity of the Felisa F20 drying kiln, slices of wood were taken and several representative samples, 10 at the most, were weighted in wet. This material was taken to the laboratory to be dried at a Felisa F20 forced-circulation kiln at 80 y 70 °C, respectively, in order to determine its moister content by the following equation:
CHm = (PHm - PSm) 100) / PHm
Where:CHm =Moisture content of the sample or slice (%)PHm = Wet weight of the sample (g)PSm = Dry weight of the sample (g)
*
*

Flores-Nieves et al., Modelos para estimación y distribución...
15
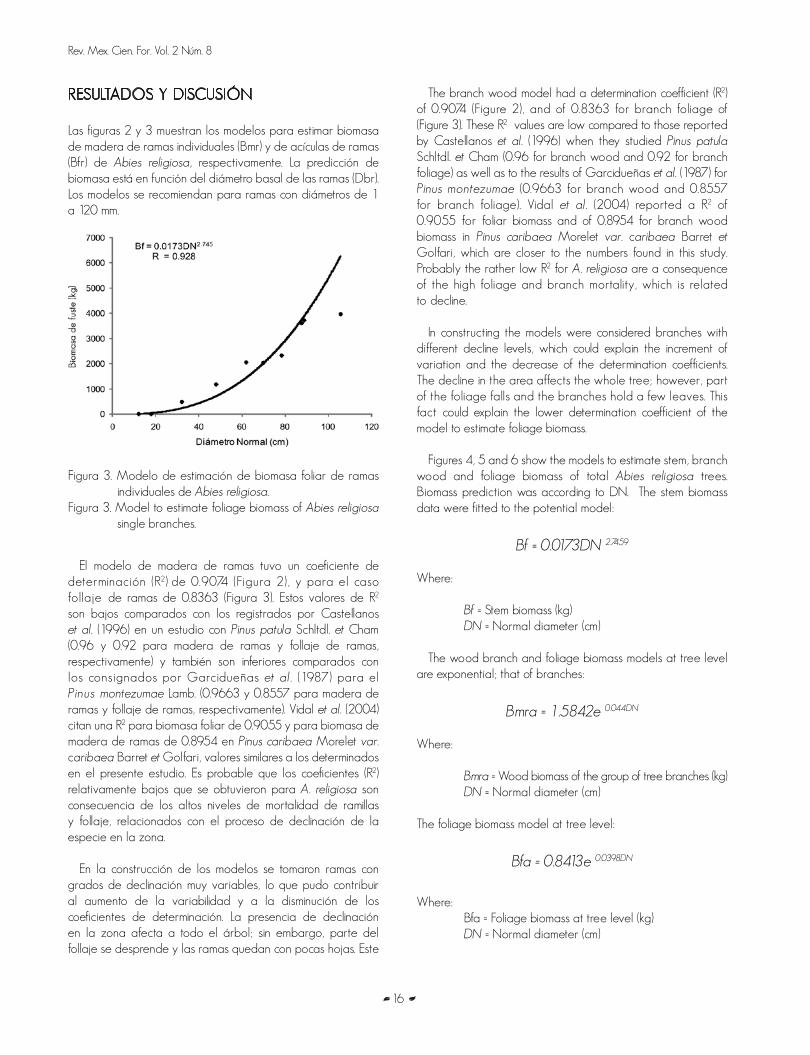
Figura 2. Modelo de estimación de biomasa de madera de ramas individuales de Abies religiosa
Figure 2. Model to estimate wood biomass of Abies religiosa single branches.
su diámetro basal con un vernier digital marca Mitutoyo y posteriormente fueron deshojadas en campo y se pesaron en húmedo la madera y el follaje por separado, en una balanza Ohaus ES100L. De la madera, cuando las ramas superaron la capacidad del horno de secado marca Felisa F20, se tomaron rodajas, y del follaje se obtuvo un máximo de 10 muestras representativas que también fueron pesadas en húmedo. El material se llevó al laboratorio para su secado en un horno de circulación forzada marca Felisa F20 a 80 y 70 °C, respectivamente, para determinar su contenido de humedad mediante la ecuación:
CHm = ((PHm - PSm) 100) / PHm
Donde:
CHm = Contenido de humedad de la muestra o rodaja (%)PHm = Peso húmedo de la muestra (g)PSm = Peso seco de la muestra (g)
El contenido de humedad se aplicó al total de madera de las ramas y follaje para obtener su peso seco, a partir de su peso húmedo.
Bcr = PHcr – (PHcr CHcr / 100)
Donde:
Bcr = Biomasa del compartimiento de la rama (madera o follaje, kg)PHcr = Peso húmedo del compartimiento de la rama (madera o follaje, kg)CHcr = Contenido de humedad del compartimiento de la rama (madera o follaje, %)
Mediante el uso de la regresión no lineal se procedió a generar los modelos para estimar la biomasa de madera de ramas y la de follaje, las cuales fueron las variables dependientes estimadas, en función del diámetro basal de la rama.
Bmr = f (Dbr)
Bfr = f (Dbr)
Donde:
Bmr = biomasa de madera de ramas individuales (kg)Bfr = biomasa de acículas de ramas (kg)Dbr = diámetro basal de rama (mm)
*
*
Moisture content was applied to the total branch wood and foliage to get their dry weight from their wet weight.
Bcr = PHcr – (PHcr CHcr / 100)
Where:
Bcr = Biomass of compartment of the branch (wood or foliage, kg)PHcr = Wet weight of the branch compartment (wood or foliage, kg)CHcr = Moisture content of the branch compartment (wood or foliage, %)
By non-linear regression, a model was designed to estimate
branch wood and foliage biomass, which were de dependent estimated variables, in terms of the basal diameter of the branch.
Bmr = f (Dbr)Bfr = f (Dbr)
Where:
Bmr = Single branch wood biomass biomass (kg)Bfr = Branch needle biomass (kg)Dbr = Basal diameter of the branch (mm)
RESULTS AND DISCUSION
Figures 2 and 3 show the models to estimate wood biomass of single branches (Bmr) and of branch needles (Bfr) of Abies religiosa. Biomass prediction is according to the basal diameter of branches (Dbr). Models are recommended for branches of 1 to 120 mm in diameter.
*
Rev. Mex. Cien. For. Vol. 2 Núm. 8
16
RESULTADOS Y DISCUSIÓN
Las figuras 2 y 3 muestran los modelos para estimar biomasa de madera de ramas individuales (Bmr) y de acículas de ramas (Bfr) de Abies religiosa, respectivamente. La predicción de biomasa está en función del diámetro basal de las ramas (Dbr). Los modelos se recomiendan para ramas con diámetros de 1 a 120 mm.
The branch wood model had a determination coefficient (R2)
of 0.9074 (Figure 2), and of 0.8363 for branch foliage of (Figure 3). These R2 values are low compared to those reported by Castellanos et al. (1996) when they studied Pinus patula Schltdl. et Cham (0.96 for branch wood and 0.92 for branch foliage) as well as to the results of Garcidueñas et al. (1987) for Pinus montezumae (0.9663 for branch wood and 0.8557 for branch foliage). Vidal et al. (2004) reported a R2 of 0.9055 for foliar biomass and of 0.8954 for branch wood biomass in Pinus caribaea Morelet var. caribaea Barret et Golfari, which are closer to the numbers found in this study. Probably the rather low R2 for A. religiosa are a consequence of the high foliage and branch mortality, which is related to decline.
In constructing the models were considered branches with different decline levels, which could explain the increment of variation and the decrease of the determination coefficients. The decline in the area affects the whole tree; however, part of the foliage falls and the branches hold a few leaves. This fact could explain the lower determination coefficient of the model to estimate foliage biomass.
Figures 4, 5 and 6 show the models to estimate stem, branch wood and foliage biomass of total Abies religiosa trees. Biomass prediction was according to DN. The stem biomass data were fitted to the potential model:
Bf = 0.0173DN 2.7459
Where:
Bf = Stem biomass (kg) DN = Normal diameter (cm)
The wood branch and foliage biomass models at tree level are exponential; that of branches:
Bmra = 1.5842e 0.044DN
Where:
Bmra = Wood biomass of the group of tree branches (kg) DN = Normal diameter (cm)
The foliage biomass model at tree level:
Bfa = 0.8413e 0.0398DN
Where: Bfa = Foliage biomass at tree level (kg) DN = Normal diameter (cm)
Figura 3. Modelo de estimación de biomasa foliar de ramas individuales de Abies religiosa.
Figura 3. Model to estimate foliage biomass of Abies religiosa single branches.
El modelo de madera de ramas tuvo un coeficiente de determinación (R2) de 0.9074 (Figura 2), y para el caso follaje de ramas de 0.8363 (Figura 3). Estos valores de R2 son bajos comparados con los registrados por Castellanos et al. (1996) en un estudio con Pinus patula Schltdl. et Cham (0.96 y 0.92 para madera de ramas y follaje de ramas, respectivamente) y también son inferiores comparados con los consignados por Garcidueñas et al . (1987) para el Pinus montezumae Lamb. (0.9663 y 0.8557 para madera de ramas y follaje de ramas, respectivamente). Vidal et al. (2004) citan una R2 para biomasa foliar de 0.9055 y para biomasa de madera de ramas de 0.8954 en Pinus caribaea Morelet var. caribaea Barret et Golfari, valores similares a los determinados en el presente estudio. Es probable que los coeficientes (R2) relativamente bajos que se obtuvieron para A. religiosa son consecuencia de los altos niveles de mortalidad de ramillas y follaje, relacionados con el proceso de declinación de la especie en la zona.
En la construcción de los modelos se tomaron ramas con grados de declinación muy variables, lo que pudo contribuir al aumento de la variabilidad y a la disminución de los coeficientes de determinación. La presencia de declinación en la zona afecta a todo el árbol; sin embargo, parte del follaje se desprende y las ramas quedan con pocas hojas. Este

Flores-Nieves et al., Modelos para estimación y distribución...
17
hecho explicaría el menor coeficiente de determinación del modelo para estimar la biomasa foliar.
Las figuras 4, 5 y 6 muestran los modelos para estimar biomasa de fuste, de madera de ramas y de follaje para árboles completos de A. religiosa . La predicción de biomasa estuvo en función del DN. Los datos de biomasa de fuste se ajustaron al modelo potencial:
Bf = 0.0173DN 2.7459
Donde:
Bf = Biomasa de fuste (kg) DN = Diámetro normal (cm)
Los modelos de biomasa de madera de ramas y follaje a nivel árbol son de tipo exponencial, el de ramas:
Bmra = 1.5842e 0.044DN
Donde:
Bmra = Biomasa de madera del conjunto de ramas del árbol (kg) DN = Diámetro normal (cm)
El modelo de biomasa de follaje a nivel árbol:
Bfa = 0.8413e 0.0398DN
Donde:
Bfa = Biomasa de follaje a nivel árbol (kg) DN = Diámetro normal (cm)
Estas ecuaciones son recomendables para valores de DN
en el intervalo de 12 a 106 cm y son confiables para usarse en condiciones similares, en cuanto al grado de declinación.
Los coeficientes de determinación para los modelos de estimación de biomasa a nivel árbol fueron 0.928, 0.6176 y 0.588 para fuste, madera de ramas y follaje, respectivamente. Aunque los valores de R2 son superiores a los registrados por algunos otros autores, hubiera sido deseable lograr valores más altos; sin embargo, los altos niveles de mortalidad a nivel fuste, ramas y follaje, estos a su vez causados por el proceso de declinación en la zona, posiblemente originaron estos resultados, especialmente en los casos de ramas y follaje (figuras 5 y 6).
Díaz (2007) en un estudio con Pinus patula consigna valores de R2 = 0.9828 para biomasa de fuste cuyo diámetro normal
These equations are advisable for DN values between 12 and 106 cm and are reliable to be used under similar conditions, in terms of decline.
The determination coefficients for the biomass estimation models were 0.928 for the stem, 0.6176 for branch wood and 0.588 for foliage. Even if the R2 values reported by several authors are higher, it would have been good to get higher numbers in this study; however, the high mortality levels at stem, branch and foliage level caused by decline in the area are the reasons why better results were not accomplished, specially in branch and foliage (figures 5 and 6).
In a Pinus patula study, Díaz (2007) reported R2 values of 0.9828 for stem biomass with a normal diameter interval from 6.7 to 64.1 cm. This number in higher than that found for A. religiosa; however, the range of sizes of the stem considered in this study might have helped to reduce the R2 value in the research.
For total A. religiosa trees, Avendaño et al. (2009) found R2 = 0.99. Geron and Ruark (1988) estimated an R2 of 0.62 for Pinus radiata D. Don needle biomass. Navar et al. (2001) cite 0.83 for R2 of stem biomass, 0.66 for branch wood biomass and 0.38 for needle biomass of Pinus durangensis Ehren. and Pinus cooperi Blanco. The determination coefficients of these studies for the branch wood and needle biomass are low, in general, and are coincidental to those here documented. In the case of pines, the low determination coefficients for biomass models of branch wood and foliage can respond to the mortality of branches causes by self-pruning, from the intolerant feature of this taxonomic group; in regard to A. religiosa, a totally shadow-tolerant species, the scarce self-pruning does not explain the low determination coefficients. Thus, it is only attributable to branch mortality, as a result of decline to which this specie is subject in this study area and in other places of is natural distribution area (Alvarado-Rosales et al. 2002, López 1997, Hernández-et al. 2001).
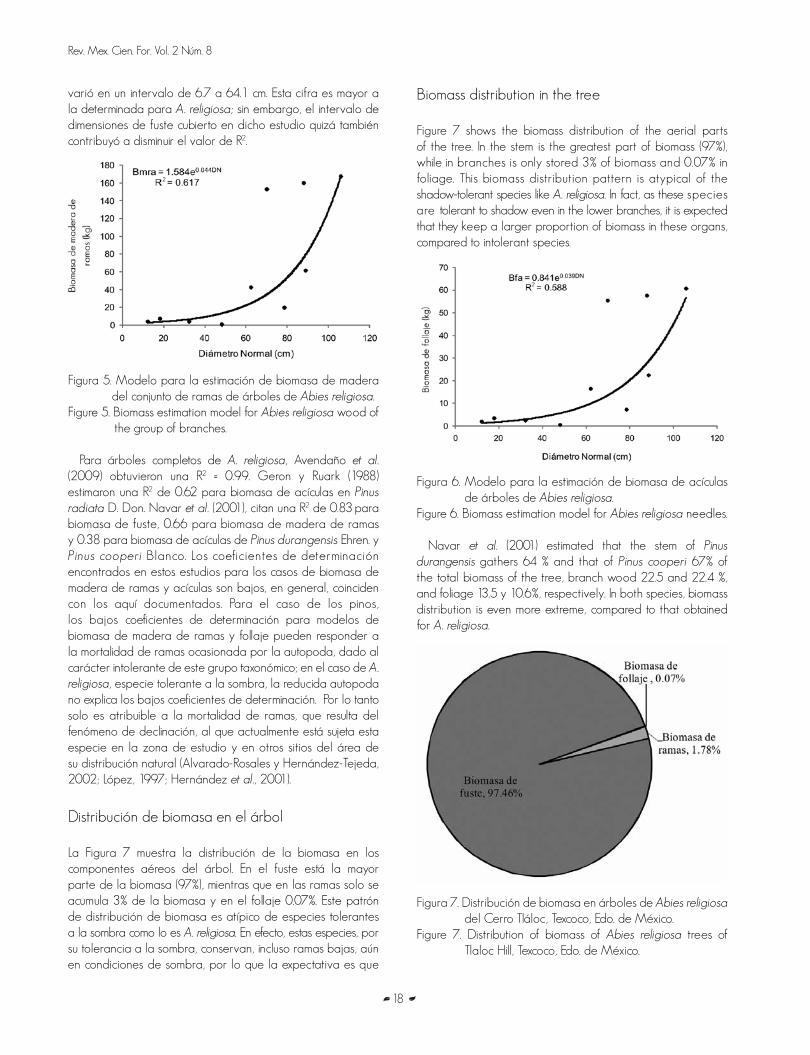
Figura 4. Modelo para la estimación de biomasa de fustes de Abies religiosa.
Figure 4. Biomass estimation model for Abies religiosa stems.
Rev. Mex. Cien. For. Vol. 2 Núm. 8
18
varió en un intervalo de 6.7 a 64.1 cm. Esta cifra es mayor a la determinada para A. religiosa; sin embargo, el intervalo de dimensiones de fuste cubierto en dicho estudio quizá también contribuyó a disminuir el valor de R2.
Figura 5. Modelo para la estimación de biomasa de madera del conjunto de ramas de árboles de Abies religiosa.
Figure 5. Biomass estimation model for Abies religiosa wood of the group of branches.
Para árboles completos de A. religiosa, Avendaño et al. (2009) obtuvieron una R2 = 0.99. Geron y Ruark (1988) estimaron una R2 de 0.62 para biomasa de acículas en Pinus radiata D. Don. Navar et al. (2001), citan una R2 de 0.83 para biomasa de fuste, 0.66 para biomasa de madera de ramas y 0.38 para biomasa de acículas de Pinus durangensis Ehren. y Pinus cooperi Blanco. Los coeficientes de determinación encontrados en estos estudios para los casos de biomasa de madera de ramas y acículas son bajos, en general, coinciden con los aquí documentados. Para el caso de los pinos, los bajos coeficientes de determinación para modelos de biomasa de madera de ramas y follaje pueden responder a la mortalidad de ramas ocasionada por la autopoda, dado al carácter intolerante de este grupo taxonómico; en el caso de A. religiosa, especie tolerante a la sombra, la reducida autopoda no explica los bajos coeficientes de determinación. Por lo tanto solo es atribuible a la mortalidad de ramas, que resulta del fenómeno de declinación, al que actualmente está sujeta esta especie en la zona de estudio y en otros sitios del área de su distribución natural (Alvarado-Rosales y Hernández-Tejeda, 2002; López, 1997; Hernández et al., 2001).
Distribución de biomasa en el árbol
La Figura 7 muestra la distribución de la biomasa en los componentes aéreos del árbol. En el fuste está la mayor parte de la biomasa (97%), mientras que en las ramas solo se acumula 3% de la biomasa y en el follaje 0.07%. Este patrón de distribución de biomasa es atípico de especies tolerantes a la sombra como lo es A. religiosa. En efecto, estas especies, por su tolerancia a la sombra, conservan, incluso ramas bajas, aún en condiciones de sombra, por lo que la expectativa es que
Biomass distribution in the tree
Figure 7 shows the biomass distribution of the aerial parts of the tree. In the stem is the greatest part of biomass (97%), while in branches is only stored 3% of biomass and 0.07% in foliage. This biomass distribution pattern is atypical of the shadow-tolerant species like A. religiosa. In fact, as these species are tolerant to shadow even in the lower branches, it is expected that they keep a larger proportion of biomass in these organs, compared to intolerant species.
Figura 6. Modelo para la estimación de biomasa de acículas de árboles de Abies religiosa.
Figure 6. Biomass estimation model for Abies religiosa needles.
Navar et al. (2001) estimated that the stem of Pinus durangensis gathers 64 % and that of Pinus cooperi 67% of the total biomass of the tree, branch wood 22.5 and 22.4 %, and foliage 13.5 y 10.6%, respectively. In both species, biomass distribution is even more extreme, compared to that obtained for A. religiosa.
Figura 7. Distribución de biomasa en árboles de Abies religiosa del Cerro Tláloc, Texcoco, Edo. de México.
Figure 7. Distribution of biomass of Abies religiosa trees of Tlaloc Hill, Texcoco, Edo. de México.

Flores-Nieves et al., Modelos para estimación y distribución...
19
Nevertheless, they are consistent with the present look of the trees in the study area. Avendaño et al. (2009) reported that in firs, 81.3 % of biomass is stored in the stem, 6.9% in branches and 8.6% in foliage. The branch biomass value is similar to that determined in the actual study, but the foliar biomass is clearly lower. The difference in the biomass distribution pattern, is probably, the result of the effect of decline.
CONCLUSIONS
The models here presented are applicable to Abies religiosa in decline. The crown structure of the trees in the study area bear signs of distortion. This increases the variability of the biomass of crowns and makes modeling, difficult.
The biomass distribution pattern of the aerial parts of A. religiosa trees in the study area is atypical, compared to individuals without decline, as a reaction to the high mortality o foliage and branches.
ACKNOWLEDGEMENTS
To CONACYT for the scholarship granted to the first author for his Master in Science studies and for the financial support provided to the 61559, 2006 project. To the Priority Research Line No. 8 (LPI-8) of the Graduate Studies School for having partially sponsored this study
End of the English version
Ciesla, W. M. 1989. Aerial photos for assessment on forest decline. A multinational overview. Journal of Forestry. 87 (2): 37-41.
Díaz F., R., M. Acosta M., F. Carrillo A., E. Buendía R., E. Flores A. y J. Etchevers D. 2007. Determinación de ecuaciones alométricas para estimar biomasa y carbono en Pinus Patula Schl. et. Cham. Madera y Bosques 13(1): 25-34.
Escandón C., J., J. B. H. J. de Jong., S. Ochoa O., I. March M. y M. A. Castillo. 1999. Evaluación de dos métodos para la estimación de biomasa arbórea a través de datos LANDSAT TM en Jusnajab La Laguna, Chiapas, México: estudio de caso. Boletín del Instituto de Geografia Núm.40: 71-84.
Garcidueñas M., A. R. 1987. Producción de biomasa y acumulación de nutrientes en un rodal de Pinus montezumae Lamb. Tesis de Maestría en Ciencias. Colegio de Postgraduados. Texcoco, Edo. de México. México. 243 p.
Geron C., D. and G. Ruark A. 1988. Comparison of constant and variable allometric ratios for predicting foliar biomass of various trees in general. Canadian Journal of Forestry Research. 18(10): 1298-1304.
Garzuglia, M. and M. Saket. 2003. Wood volume and woody biomass: review of FRA 2000 estimates. Forest Resources Assessment WP 68. Food and Agriculture Organization of the United Nations. Rome. Italy. 30 p.
Granados S., D. y G. F. López R. 2001. Declinación forestal. Revista Chapingo. Ciencias Forestales y del Ambiente 7(1): 5-13.
Hernández T., T., R. M. Cox, J. Malcolm, M. L. de la Isla B, J. Vargas H., A. Velázquez M. y D. Alvarado R. 2001 Impacto del ozono sobre el polen de Pinus hartwegii Lindl. Rev. Cien. For. en Mex. 26: 5-18.
Herrera A., M. A., I. del Valle J. y O. Alonso S. 2001. Biomasa de la vegetación herbácea y leñosa pequeña y necromasa en bosques tropicales primarios y secundarios de colombia. In: Memorias del Simposio Internacional Medición y Monitoreo de la Captura de Carbono en Ecosistemas Forestales. Valdivia, Chile.18 p.
tengan una proporción más grande de biomasa en estos órganos, en comparación con las especies intolerantes.
Para Pinus durangensis y Pinus cooperi, Navar et al. (2001) estimaron que el fuste contiene 64 y 67% de la biomasa total del árbol, la madera de ramas 22.5 y 22.4 % y el follaje 13.5 y 10.6%, respectivamente. En ambas especies, la distribución de biomasa es todavía más extrema, comparada con la que se obtuvo para A. religiosa.
No obstante, son congruentes con la apariencia actual del arbolado en el área de estudio. Avendaño et al. (2009), mencionan que en los árboles de oyamel 81.3 % de la biomasa se acumula en los fustes, 6.9% en ramas y 8.6 % en follaje. El valor de biomasa de ramas es similar al determinado en el presente estudio, pero la proporción de biomasa foliar es claramente menor. La diferencia en el patrón de distribución de biomasa, probablemente, resulta del efecto del fenómeno de declinación.
CONCLUSIONES
Los modelos desarrollados en el presente estudio son aplicables a los árboles de Abies religiosa en proceso de declinación. La estructura de copa de los árboles de A. religiosa en dicha condición en el área de interés muestra señales de alteración. Esto aumenta la variabilidad de la biomasa de copas y dificulta su modelación.
El patrón de distribución de biomasa en los componentes aéreos de árboles de A. religiosa en el área de estudio es atípico, en comparación con individuos sin declinación, en respuesta a la alta mortalidad de follaje y ramas.
AGRADECIMIENTOS
Al CONACYT por la beca otorgada al primer autor para sus estudios de Maestría en Ciencias y por el financiamiento a través del proyecto 61559, 2006. A la Línea Prioritaria de Investigación 8 (LPI-8) del Colegio de Postgraduados, por el financiamiento parcial al estudio.
REFERENCIAS
Acosta M., M., J. J. Vargas H., A. Velázquez M. y J. Etchevers B. 2002. Estimación de la biomasa aérea mediante el uso de relaciones alométricas en seis especies arbóreas en Oaxaca, México. Agrociencia. 36: 725-736.
Alvarado R., D., L. I. De Bauer and A. J. Galindo. 1993. Decline of sacred fir (Abies religiosa) in a forest park south of Mexico City. Environ. Pollut. 80: 115–121.
Alvarado-Rosales, D. and T. Hernández-Tejeda. 2002. Decline of Sacred-fir in the Desierto de los Leones Nacional Park. In: Fenn, M. E., L. I. de Bauer, T. Hernández-T. (Eds.) Urban air pollution and forests. Resources at risk in the Mexico City air basin. Springer. New York, NY. USA. pp. 243-260.
Avendaño H., D., M. Acosta M., F. Carrillo A. y J. Etchevers B. 2009. Estimación de biomasa y carbono en un bosque de Abies religiosa. Fitotecnia Mexicana. 32 (3): 233-238.
Castellanos F., J., A. Velázquez M., J. J. Vargas H., C. Rodríguez F. y A. M. Fierros G. 1996. Producción de biomasa en un rodal de Pinus patula Schl. et Cham. Agrociencia. 30: 123-128.

Rev. Mex. Cien. For. Vol. 2 Núm. 8
20
López L., M. A. 1997. Efectos del ambiente aéreo y del suelo sobre el desarrollo de síntomas de declinación de oyamel. Terra 15: 287-293.
López-L., M. A., R. Reich M., C. Aguirre-Bravo. and A. Velázquez M. 2009. Pine growth and nutrient status as related to pine/alder ratio in mixed stands. Journal of Biological Sciences. 9(7): 637-647.
López-López M.. A., A. Velázquez M., J. Acosta M. y E. Estañol B. 2006. Biomass distribution in declining sacred-fir seedlings. Interciencia 31 (6): 451-455.
López R., C. y R. Keyes. 1987. Modelos para estimación de biomasa de Pinus cembroides Zucc. In: Memorias del II Simposio Nacional sobre Pinos Piñoneros. México, D. F. México. pp: 211-220.
Manzano M., D., M. A. López L., F. Manzano M. y E. Pineda H. 2007. Productividad primaria neta de una plantación de recuperación de suelos. In: Memorias del VIII Congreso Mexicano de Recursos Forestales. Morelia, Mich., México. 11 p.
Mooser, F. 1975. Historia geológica de la Cuenca de México. In: Departamento del Distrito Federal (Eds). Memorias de las obras del sistema del drenaje profundo del Distrito Federal. Tomo 1. Talleres Gráficos de la Nación. México, D. F. México. pp. 7-38.
Navar , J., N. González. y J. Graciano. 2001. Ecuaciones para estimar componentes de biomasa en plantaciones forestales de Durango. México. In: Memorias del Simposio Internacional Medición y Monitoreo de la Captura de Carbono en Ecosistemas Forestales. Valdivia, Chile. pp. 1– 12.
Ortíz S., C. y H. Cuanalo de la Cerda. 1977. Levantamiento fisiográfico del área de influencia de Chapingo. Colegio de Postgraduados. Escuela Nacional de Agricultura, Chapingo, Edo. de Méx., México. 83 p.
Palma T., A. 1996. Tipología del uso forestal de la tierra de la región norte de la Sierra Nevada y su cartografía. Tesis de Maestría. Colegio de Postgraduados. Montecillo. Edo. de México. México. 103 p.
Rojo M., G. E., J. Jasso M., J. J. Vargas H., D. J. Palma L. y A. Velázquez M. 2005. Biomasa aérea en plantaciones comerciales de hule (Hevea brasiliensis Müll. Arg.) en el estado de Oaxaca, México. Agrociencia 39: 449-456.
Vázquez S., L., J. Tamarit C. y J. Quintanar O. 2004. Caracterización de la declinación de bosques de encino en “Sierra de los Lobos” Guanajuato, México. Polibotánica. 17: 1-14.
Vidal, A., J. Benítez Y., J. Rodríguez., R. Carlos. y H. Gra. 2004. Estimación de la biomasa de copa para árboles en pie de Pinus caribaea var caribaea en la E.F.I. La Palma de la provincia de Pinar del Río, Cuba. Quebracho. Revista de Ciencias Forestales (11): 60-66.
Waring E., H. and W. H. Schlesinger. 1985. Forest ecosystems. Academic Press. Orlando, FL. USA. 340 p.

CARACTERÍSTICAS MORFOLÓGICAS DE PLÁNTULAS DE DOS ESPECIES FORESTALES TROPICALES PROPAGADAS EN CONTENEDORES BIODEGRADABLES
Y CHAROLAS STYROBLOCK
MORPHOLOGICAL CHARACTERISTICS OF SEEDLINGS OF TWO TROPICAL FOREST SPECIES
PROPAGATED IN BIODEGRADABLE CONTAINERS AND STYROBLOCK TRAYS
H. Jesús Muñoz Flores 1, J. Jesús García Magaña 1, Víctor Manuel Coria Ávalos 1,Gabriela Orozco Gutiérrez 1 y Yadira Yolanda Muñoz Vega2
RESUMEN
La función de los envases utilizados en la propagación de especies forestales es contener el sustrato que abastece a las raíces de agua, aire, nutrimentos minerales y provee de soporte físico a la planta durante su permanencia en el vivero. El objetivo de este trabajo fue evaluar la factibilidad técnica de utilizar contenedores biodegradables (de fibra de coco), en comparación con charolas Styroblock de 60 y 77 cavidades para la producción de Enterolobium cyclocarpum y Tabebuia rosea. Se aplicó un diseño completamente al azar, con seis tratamientos y cuatro repeticiones. A los tres meses de estancia en vivero, los individuos de E. cyclocarpum en los contenedores de fibra de coco lograron una altura de 24.8 cm, que fue significativamente superior (Pr > F= <0.0001) a los valores registrados cuando se usaron charolas de 60 y 77 cavidades (14.9 cm). El diámetro del cuello de la raíz de T. rosea y E. cyclocarpum en contenedores de fibra de coco mostraron diferencias significativas (Pr>F= <0.0001), que resultaron en dos grupos (Tukey α=0.05); el primero con los mayores diámetros (6.2 a 5.5 mm) para ambas especies en los envases biodegradables y el segundo que incluye las charolas de 60 y 77 cavidades (2.7 a 3.3 mm). En los dos taxa, la biomasa total (aérea y radical) fue superior en contenedores de fibra de coco, aunque los valores para E. cyclocarpum (4.69 g) fueron superiores significativamente respecto a los de T. rosea (3.66 g). Estos envases no contaminan y en ellos se producen plantas de alta calidad.
Palabras clave: Contenedores de poliestireno, contenedores de fibra de coco, Enterolobium cyclocarpum (Jacq.) Griseb., propagación de plantas, Tabebuia rosea (Bertol) DC., viveros forestales.
ABSTRACT
The function of any container for the propagation of forest species is to contain the substrate that supplies roots with water, air, mineral nutrients and provides physical support to the plant while it stays in the nursery. The aim of this study was to assess the technical feasibility of using biodegradable containers (made from coconut fiber) compared with trays of 60 and 77 Styroblock cavities for the propagation of two tropical forest species, Enterolobium cyclocarpum and Tabebuia rosea. A completely randomized design with six treatments and four repetitions was used. After three months in the nursery, the plants of E. cyclocarpum in coconut fiber containers reached a height of 24.8 cm, which was significantly higher (Pr> F = <0.0001) in regard to plants of the same species that were placed into trays of 60 and 77 cavities (14.9 cm). The diameter of the neck of the root of T. rosea and E. cyclocarpum in the first kind of containers showed significant differences (Pr> F = <0.0001), which resulted in two groups (Tukey α = 0.05); the first one, with the largest diameters (6.2 to 5.5 mm) in the two species that were in the biodegradable containers and the second, which includes the trays of 60 and 77 cavities (2.7 to 3.3 mm). In both taxa the total biomass (aerial and root) was higher in coconut fiber containers, although the values obtained in E. cyclocarpum (4.69 g) were significantly higher compared to those of T. rosea (3.66 g). These containers do not pollute, and produce high quality plants.
Key words: Polystyrene containers, coconut fiber containers, Enterolobium cyclocarpum (Jacq.) Griseb., plants propagation, Tabebuia rosea (Bertol) DC, forest nursery.
Fecha de recepción: 22 de febrero de 2010.Fecha de aceptación:23 de diciembre de 2011.
1 Campo Experimental Uruapan, Centro de Investigación Regional Pacífico Centro, INIFAP. Correo-e: [email protected] 2 Facultad de Agrobiología “Presidente Juárez”, Universidad Michoacana de San Nicolás de Hidalgo

Rev. Mex. Cien. For. Vol. 2 Núm. 8
22
INTRODUCCIÓN
Para la propagación de especies forestales en vivero, se necesita contar con información que permita ubicar, dimensionar e implementar las acciones contenidas en los planes de trabajo que posibiliten el cumplimiento de los objetivos y metas planteados. Independientemente del tipo de germoplasma a propagar, una de las principales decisiones del viverista es la determinación del tipo del envase o contenedor a emplear, el cual se elegirá en función de la especie a propagar, así mismo deberán considerarse algunas características de tipo biológico y económico (Mexal et al.,1993). Las primeras incluyen el tamaño de la semilla o estaquilla, la altura final de la plántula y su calidad, así como, las condiciones ambientales del lugar definitivo de la plantación. Desde el punto de vista económico, se tomaran en cuenta el costo, la capacidad, posibilidad de reuso, disponibilidad en el mercado, densidad de plantas por unidad de superficie, facilidad de llenado, manejo y transporte (Fisher, 1992; Landis et al., 1994; Ruano, 2002; Toral et al., 2000). También son importantes el tipo y tamaño de envase, ya que si las plantas crecen muy juntas y se entrelaza su follaje son más susceptibles a daños por plagas o enfermedades. Esto se puede solucionar si se conoce el hábito de crecimiento de cada una de las especies, con la finalidad de manejar la densidad óptima, la cual incide en la lignificación de los tallos (Landis et al., 1994; Peterson y Sutherland, 1989; Timmis y Tanaka, 1976).
El tipo y tamaño del contenedor tiene relación directa con la morfología radical y el desarrollo aéreo de las plantas en vivero, esta, a su vez influye enormemente en el éxito de la plantación (Burdett et al., 1983; Landis et al.,1994; Ritchie, 1984; Wenny et al., 1988;); por lo tanto los viveristas se asegurarán de emplear el tipo de contenedor apropiado, en cuanto a diseño, capacidad, espacio entre celdas, durabilidad, costo, dificultad de manejo en el vivero, transporte y operación de la plantación. Además, se integrarán, en la toma de decisiones, de acuerdo a su destino final: plantaciones forestales de conservación o comerciales (Fisher, 1992).
Las propiedades del contenedor ideal se han investigado desde la década de 1930 en Estados Unidos de América (Strachan, 1974) y aunque pueden ser comparados en distintas formas, la más apropiada es en relación a su funcionalidad.
La función primaria de cualquier envase es la de contener una cantidad de sustrato, que abastece a las raíces con nutrimentos minerales y provee soporte físico a la planta mientras está en el vivero (Mexal et al., 1993). El contenedor ideal es el que permite obtener la mejor calidad de plantas, en el menor tiempo posible (Liegel y Venator, 1987).
En Estados Unidos de América, los contenedores de mayor uso en los viveros son de poliestireno comprimido (51% del total nacional), plástico rígido 26%, tipo libro o funda
INTRODUCTION
In order to propagate forest species at the nursery it is necessary to have the information that allows to locate, measure and implement the actions in work-plans that make it possible to accomplish the considered aims and goals. Regardless of the kind of germ plasm that will be propagated, one of the most important decisions that the nursery-man will make is to determine the kind of container to use, which will be selected according to the species to be handled, as well as some biological and economic elements (Mexal et al., 1993). The first include the size of the seed or cutting, the final height of the seedling and their quality, as well as the environmental conditions of the final plantation place. From the economic viewpoint, costs, capacity, reuse possibility, market availability, plant density per area, filling possibilities, handling and transportation (Fisher, 1992; Landis et al., 1994; Ruano, 2002; Toral et al., 2000). The kind and size of the container are important too, since if the plants grow very close to each other and their foliage mixes between them, they are more vulnerable to damages caused by plagues and diseases. This can be solved if the growth habits of each of the species are known, in order to manage the optimal density, which affects the lignifications of stems (Landis et al., 1994; Peterson and Sutherland, 1989; Timmis and Tanaka, 1976).
The kind and size of the container has a direct relation with root morphology and the aerial development of plants in the nursery, which affects enormously the success of the plantation (Burdett et al., 1983; Landis et al., 1994; Ritchie, 1984; Wenny et al., 1988), as well; thus, the nursery workers will make sure to use the right type of container in terms of design, capacity, space between cells, durability, cost, management difficulty at the nursery, transportation and operation of the plantation. In addition, they will be taken into account for decision making (Fisher, 1992), in regard to their final destination, commercial forest plantations or conservation.
The properties of the ideal container have been investigated since the 1930’s in the United States of America (Strachan, 1974) and in spite of being compared in many different ways, the most suitable considers its functional qualities.
The basic function of any container is to keep some amount of substrate, which provides mineral nutrients to roots and provides physical support to the plant while it is in the nursery (Mexal et al., 1993). The ideal container favors to get the best quality of plants in the least time (Liegel and Venator, 1987).
In the United States of America, the most regularly used containers at the nursery are made of compressed polystyrene (51% of the total at national scale), 26% rigid plastic, 10% book or dustjacket type, 10% paper and 3% of different kind (Landis et al., 1994). Those of polystyrene, once their life span has ended, become pollutant and those of rigid or flexible plastic

Muñoz et al., Características morfológicas de plántulas...
23
10%, de papel 10% y de otros tipos 3% (Landis et al., 1994). Los de poliestireno, una vez terminada su vida útil representan materiales contaminantes y los de plástico rígido o flexible tardan 100 años o más en degradarse; ante este problema es necesario explorar otros materiales para su fabricación.
Los contenedores elaborados a base de fibra de coco se proponen como una tecnología alternativa, amable con el ambiente, cuyas características favorecen la producción de individuos de alta calidad en el vivero; además no es necesario eliminarlo durante el trasplante en campo, por lo que esta actividad se realiza en menos tiempo. En sitios donde ocurren temperaturas extremas y se dispone de escasa humedad, las paredes del contenedor protegen el sistema radical de las plántulas, lo que aumenta el porcentaje de sobrevivencia en la plantación definitiva.
Los envases empleados en la propagación de plantas en vivero, en general, se pueden clasificar en dos tipos: los que se dejan al plantar (Paperpot MR, cartón asfaltado Jiffy MR y pellets), y los que se remueven al momento del trasplante, algunos de estos (charolas de poliestireno comprimido, de plástico rígido, tipo libro, y los de polietileno negro) presentan restricciones después de la plantación (Barnett y McGilvray, 1981; Wright et al., 1999).
Los contenedores de papel se usan ampliamente en el centro y este de Canadá, pero en el oeste se dejaron de utilizar porque las raíces tenían problemas para salir una vez que eran trasplantadas las plántulas, especialmente, en localidades frías y húmedas (Sims, 1988). Entre las ventajas que ofrecen estos envases es que están hechos de material biodegradable, lo que es importante si se considera que del 60 a 70% de las operaciones en el vivero se semi-mecanizan.
En zonas de baja precipitación se ha registrado que algunos permanecen casi intactos después de cuatro años de que la plantación se realizó, por lo que es recomendable romperlos al momento del trasplante (Mexal et al., 1993). Las presentaciones varían en número de cavidades de 84 a 1,400 por metro cuadrado. Una desventaja importante es que no tienen una pared interna que evite el crecimiento de la raíz en espiral, lo que resulta en la formación de un anillo radical que limita el desarrollo del resto del sistema de soporte y de absorción de agua y nutrimentos.
En Canadá se empezó a emplear un nuevo sistema de producción de plantas de fácil manejo, denominado Jiffy Pellets, envase y sustrato a la vez, con el cual el uso, la compra o recolección de medios de crecimiento o substratos se hacen innecesarios, así como la adquisición y llenado de envases. En consecuencia, su manipulación y plantación se realizan con facilidad y economía; no obstante, presenta desventajas, tales como, que una vez humedecidos, por ningún motivo debe
take 100 years or more to decompose; in order to face this problem, it is necessary to explore other materials for their manufacturing.
Containers made from coconut fiber are put forward as an alternative technology, eco-friendly, that favor the production of high quality individuals at the nursery; in addition, it is not necessary to destroy them while transplantation is carried out in the field, which reduces time. In places with extreme temperatures and low moisture, the container walls protect the roots of seedlings, which increases survival per cent in the final plantation.
In general, the containers used for the propagation of plants in the nursery, can be classified in two types: those left while planting (Paperpot [TM], Jiffy Asphalted Cardboard [TM] and pellets) and those that are removed when transplanting is made, some of which compressed polystyrene trays, rigid plastic book-like and of black polyethylene) show some limitations after plantation (Barnett and McGilvray, 1981; Wright et al., 1999).
Paper containers (Paperpot [TM]) are widely used in Central and Eastern Canada, but they stopped to be used at the West since the roots had problems to come out once the seedlings were transplanted, specially in cold and moist locations (Sims, 1988). One advantage is that they are made-up by biodegradable material, which is important as 60 to 70% of the operations in the nursery are semi-mechanical.
In low-precipitation places, some keep almost intact after four years from the time the plantation was made, which makes it necessary to break them when transplanting takes place (Mexal et al., 1993). Presentations vary in their number of cavities, from 84 to 1,400 m-2. A problem with them is that they do not have an internal wall that avoids spiral root growth, which provokes the formation of a root ring that limits the development of the rest of the support system as well as of water and nutrient absorption.
A new system of plant production, easy to handle, has started to be used in Canada; it is known as Jiffy Pellets [TM], which is a container and substrate at the same time, with which the use, purchase or collect of growing media or substrate become unnecessary, as well as buying and filling containers. Consequently, their handling and plantation are easier and cheaper; in spite of it, it has some limitations, such as, that once they get damp, they cannot lack water, which suggests a greater water expense; another is that the space to develop the radical system is small; in addition, species of grass habit and wide crown have some health problems and routines to provide specific nutriments to each taxon must be formulated.
The mostly used containers at the nurseries of Mexico are of black polyethylene, although they have been of incorrect

Rev. Mex. Cien. For. Vol. 2 Núm. 8
24
faltarles el suministro hídrico, lo que hace que se tenga un mayor gasto de agua; otra, es que el espacio para desarrollar el sistema radical es reducido; además, las especies de hábito cespitoso y copa amplia tienen problemas de sanidad y se deben desarrollar rutinas de aporte de nutrimentos específicos para cada taxón.
El tipo de envase que más se emplea en los viveros de México es el de polietileno negro, aunque han sido de dimensiones inapropiadas para la producción de plántulas vigorosas. En general, para la propagación de pinos se recomienda un tamaño de bolsa de 7 × 20 cm o 10 × 20 cm y para latifoliadas 10 × 23 cm, en ambos casos con fondo (Patiño y Marín, 1983). Entre sus principales desventajas están la dificultad para mecanizar las operaciones y el crecimiento de la raíz en espiral dentro del envase (González, 1995; Liegel y Venator, 1987).
Otro tipo de envases individuales son Ray Leach [MR]., para pinos se recomienda el tamaño de 2.5 × 16 cm (1,076 celdas m-2) o 3.8 × 20 cm (527 celdas m-2). Estos tienen las siguientes ventajas: los espacios de crecimiento se utilizan eficientemente, se requiere menor cantidad de substrato, son reutilizables y dependiendo del uso y exposición al sol duran de 8 a 10 años (Mexal et al., 1993).
Los contenedores múltiples de tipo Styroblock [MR] o Styrofoam blocks [MR] son de diversas capacidades. En función del hábito de crecimiento de las plantas a propagar existen envases con características definidas; por ejemplo, los de diseño que permiten la poda radical (Stuewe, 2006). Para la propagación de coníferas se recomiendan celdas tamaño 3 × 22.9 cm (764 celdas m-2) y 4.1 × 15.2 cm (441 celdas m-2) y para latifoliadas de 6.1 × 15.2 cm (215 celdas m-2) (Mexal et al., 1993).
Para Pinus pseudostrobus Lindl., P. douglasiana Martínez y P. herrerae Martínez y en general para las especies que no tienen crecimiento inicial cespitoso se pueden utilizar mayores densidades por metro cuadrado. Por el contrario, para P. michoacana Martínez y P. montezumae Lamb, por sus características de crecimiento inicial, se usan densidades menores (García y Muñoz, 1993).
La característica principal de las plantas a considerar durante su producción en vivero es el desarrollo del sistema radical, del que depende la calidad de la planta y, en consecuencia su capacidad desde su sobrevivencia después del trasplante hasta las tasas de crecimiento inicial y de desarrollo subsecuente del árbol (Duryea y McClain, 1984; Ruano, 2002; Toral et al., 2000).
Los fabricantes de contenedores siguen trabajando en el desarrollo de nuevos diseños, con el propósito de mejorar la morfología de las plantas (radical y aéreo), reducir
dimensions for the production of vigorous plants. In general, for the propagation of pines it is advisable a 7 × 20 cm or 10 × 20 cm bag and of 10 × 23 cm for broadleaves, in both cases with bottom (Patiño and Marín, 1983). Among their major limitations are the difficulties to make the operations mechanical and spiral root growth inside the container (González, 1995; Liegel and Venator, 1987).
Another type of individual containers are Ray Leach [TM]. For pines are recommended those of 2.5 × 16 cm (1,076 cells m-2) or 3.8 × 20 cm (527 cells m-2). They are convenient because their spaces are efficiently used, a smaller amount of substrate is necessary, they can be used again, and according to the use they have had and their exposure to sunlight, they can last for 8 to 10 years (Mexal et al., 1993).
The Styroblock [TM] o Styrofoam blocks [TM] multiple containers have different capacities. According to growth habits of plants that will be propagated, there are containers with specific characteristics; for example, those for radical pruning (Stuewe, 2006). For conifers, 3 x 22.9 cm (764 cells m-2) and 4.1 x 15.2 cm (441 cells m-2) cells are recommended and of 6.1 x 15.2 cm (215 cells m-2) for broadleaves (Mexal et al., 1993).
For Pinus pseudostrobus Lindl., P. douglasiana Martínez and P. herrerae Martínez as well as for species that do not have a grass-like initial growth, greater densities per square meter. On the other hand, for P. michoacana Martínez and P. montezumae Lamb, lower densities are used (García and Muñoz, 1993).
The main characteristic of plants to take into account during their production at the nursery is the development of the radical system, from which the quality of plant depends and, consequently, their ability to survive after transplanting up to the initial growth rates and the subsequent development of the tree (Duryea and McClain, 1984; Ruano, 2002; Toral et al., 2000).
The manufacturers of containers are still working in the development of new designs in order to improve the morphology of plants (root and air), reduce efforts both in operation at the nursery as well at the time of transplanting, and at the same time, to consider the possibilities of reusing the container, costs, and adjustments in the nursery and irrigation, among other situations (Cuny, 1996).
The aim of this study was to assess the technical feasibility of the use of containers made from coconut-fiber, compared to two types of expanded polystyrene trays for the propagation of two tropical forest species at the nursery.

Muñoz et al., Características morfológicas de plántulas...
25
esfuerzos tanto en la operación en el vivero, como al momento del trasplante y al mismo tiempo considerar posibilidades de reuso del contenedor, costos, adecuaciones en el vivero e irrigación, entre otros (Cuny, 1996).
El objetivo del presente trabajo fue evaluar la factibilidad técnica del uso de contenedores elaborados a base de fibra de coco, en comparación con dos tipos de charolas de poliestireno expandido para la propagación en vivero de dos especies forestales de clima tropical.
MATERIALES Y MÉTODOS
El estudio se llevó a cabo del 15 de abril al 15 de julio de 2008 en el vivero forestal del patronato de la Escuela Nacional Forestal de Uruapan, Michoacán A.C. ubicado en las instalaciones del Centro de Educación y Capacitación Forestal 01-CONAFOR, en la ciudad de Uruapan.
Las especies utilizadas fueron Enterolobium cyclocarpum (Jacq.) Griseb (Leguminosae) y Tabebuia rosea (Bertol) DC (Bignoniaceae).
Los frutos de E. cyclocarpum y T. rosea se recolectaron en la región de Nuevo Urecho, Mich., de árboles adultos y sanos. El material se secó a la sombra y se le extrajeron los embriones, mismos que se limpiaron de impurezas y se desinfectaron en una solución compuesta por tres partes de hipoclorito de sodio y siete de agua, en la que se sumergieron durante 10 min; se enjuagaron en agua y se secaron a la sombra. Los embriones cigóticos de E. cyclocarpum se escarificaron manualmente, con el propósito de acelerar la germinación. La siembra se realizó en almácigos, 1000 semillas, que contenían 50% de turba de musgo de pantano y 50% de agrolita. Del total de semillas germinadas se seleccionaron las plantas más vigorosas y de tamaño uniforme; se extrajeron e inmediatamente se sometieron a un riego abundante.
Durante toda la etapa de vivero se aplicó diariamente una lámina de agua de aproximadamente 1 cm, de lunes a sábado. El control de malezas se realizó en forma manual, al ejecutar la revisión diaria de las plantas. No se presentaron daños por plagas o enfermedades.
El sustrato consistió de una mezcla de 50 % de Peat moss (turba de musgo de pantano) + 25 % de Agrolita + 25 % de Vermiculita, complementada con fertilizante sólido de liberación lenta: Osmocote (18-6-12) a razón de 5 kg por metro cúbico de sustrato (Mexal et al., 1993).
Se utilizaron tres tipos de contenedores: a) contenedores elaborados con 65% de fibra del fruto de la palmera Cocus nucifera L., y 35% de látex natural de savia de Hevea brasiliensis (Willd. Ex A.Juss.) Müll. Arg. (árbol del hule),
MATERIALS AND METHODS
The study was carried out from April 15th to July 15th, 2008 at the forest nursery of the National Forest School board of Uruapan, Michoacán State located in the Education and Forest Training Center Number 1 of CONAFOR at Uruapan city.
The species that were tested were Enterolobium cyclocarpum (Jacq.) Griseb (Leguminosae) and Tabebuia rosea (Bertol) DC (Bignoniaceae).
The fruits of these species were collected in the Nuevo Urecho region, Michoacán State, from healthy adult trees. The material was dried under shadow and the embryos were extracted from them, to which all impurities were removed and were disinfected with a compound solution of three parts of sodium hypochlorite by seven of water, in which they were immersed for 10 min; they were rinsed with water and were put to dry under shadow. The zygotic embryos of E. cyclocarpum were scarified by hand in order to make germination faster. Sowing of 1,000 seeds was made in a mixture of 50% Peat moss + 50% agrolite. Of the total germinated seeds were selected the most vigorous and uniform -size plants; they were extracted and immediately were given abundant watering.
During the whole stage at the nursery, about a 1 cm water plate was applied every day, from Monday to Saturday. Weed control was hand-made, and was carried out while the daily review of plants was made. No damages were found from plagues or disease.
Substrate was a mixture of 50 % Peat moss + 25 % Agrolite + 25 % Vermiculite completed by a solid fertilizer with slow liberation: Osmocote (18-6-12) at a rate of 5 kg/m3 of substrate (Mexal et al., 1993).
Three types of containers were used: a) Containers made from 65% of fiber of Cocus nucifera L. palm and 35% of natural latex from Hevea brasiliensis (Willd. Ex A. Juss.) Müll. Arg. (rubber tree), with 314 cm3 and 205 plants m-2; b) 60 × 35 cm Styroblock (polystyrene) trays with 60 pots of 220 cm3 and 284 m-2cells (5.1 cm diameter and 12 cm deep; and c) 60 × 35 cm Styroblock (polystyrene) trays with 77 pots of 175 cm3 and 364 m-2cells (4.2 cm diameter and 11.7 cm long).
Treatments were allocated in a completely at random design with four replications; each unit was formed by 77 plants and consisted of a matrix species-type of container; they were the following: 1) E. cyclocarpum in cococut fiber container; 2) T. rosea in 77- pot tray; 3) T. rosea in 60 -pot tray; 4) E. cyclocarpum in –pot tray; 5) T. rosea in cococut fiber container and 6) E. cyclocarpum in 60 -pot tray. The statistical model to analyze the resulting information was proposed by Martínez (1988).

Rev. Mex. Cien. For. Vol. 2 Núm. 8
26
con una capacidad de 314 cm3 y densidad de 205 plantas m-2; b) charolas Styroblock (poliestireno) con dimensiones de 60 x 35 cm, con 60 cavidades de 220 cm3 y una capacidad de 284 celdas m-2, (5.1 cm de diámetro y 12 cm de profundidad); y c) charola de Styroblock (poliestireno) con dimensiones de 60 x 35 cm, con 77 cavidades de 175 cm3 y una capacidad de 364 celdas m-2, (4.2 cm de diámetro y 11.7 cm de longitud).
Los tratamientos se distribuyeron en un diseño completamente al azar con cuatro repeticiones, cada unidad estuvo compuesta por 77 plantas, y consistieron en una matriz especie-tipo de contenedor; fueron los siguientes: 1) E. cyclocarpum en contenedor de fibra de coco, 2) T. rosea en charola de 77 cavidades, 3) T. rosea en charola de 60 cavidades, 4) E. cyclocarpum en charola de 77 cavidades, 5) T. rosea en contenedor de fibra de coco y 6) E. cyclocarpum en charola de 60 cavidades.
El modelo estadístico para el análisis de la información fue el propuesto por Martínez (1988).
Yij = µ + T
i + Є
ij
Donde:
Yij = Variable respuesta
µ = Promedio general que considera las diferentes fuentes de variaciónT
i = Efecto del i-ésimo tratamientoЄij = Error aleatorio ij
Se hizo un análisis de varianza con el paquete Statistical Analysis System (SAS, 2004) mediante el procedimiento PROC GLM. Cuando se observaron diferencias entre tratamientos (p <0.05), se aplicaron pruebas de comparación de medias de Tukey. Se evaluó la supervivencia, altura, diámetro del cuello de la raíz, las biomasas secas total, aérea y radical; además del volumen y número de raíces primarias.
Supervivencia. Se consideró como el número de plantas vivas. Para efectos de comparación analítica, la variable se expresó en porcentaje.
Altura. Se midió desde el cuello de la raíz hasta la yema terminal, medida con una regla en centímetros.
Diámetro del cuello de la raíz. Esta dimensión se tomó en la parte baja del epicótilo, para lo cual se utilizó un vernier Scala modelo 222a en milímetros con aproximación de una décima.
Biomasa seca total. Se determinó con la materia húmeda del total de la planta colocada en una estufa de secado MAPSA
Yij = µ + T
i + Є
ij
Where:
Yij = Response variable
µ = General average that takes into account the different sources of variationT
i = Effect of the i-esim treatment
Єij = ij Random error
An analysis of variance was performed with the Statistical Analysis System (SAS, 2004) through PROC GLM. When differences between treatments were observed (p <0.05) Tukey’s mean tests were applied. Survival, height, diameter of the root neck, total, aerial and root dry biomass, as well as volume and number of primary roots, were assessed.
Survival. The number of live plants were considered. For analytical comparison endings, the variable was expressed in per cent.
Height. It was taken from the neck of the root to the terminal bud, measured by a ruler (cm).
Diameter of root neck. This size was taken in the lower part of the epicotyl with a 222a Model Scala vernier caliper (mm), with 1/10th approach.
Total dry biomass. It was determined with the moist material of the whole plant that was placed into a MAPSA HDT-27 kiln for 72 h at 70 oC until a constant weight was obtained, in grams with 1/100th approach.
Aerial dry biomass. The aerial part of the plant, from the terminal bud to the neck where it joins to the root; as the total dry biomass, it was placed into a MAPSA HDT-27 kiln for 72 h at 70 oC until a constant weight was obtained, in grams with 1/100th approach.
Root dry biomass. The root was taken to a constant weight as it was placed, too, into a MAPSA HDT-27 kiln for 72 h at 70oC until a constant weight was obtained, in grams with 1/100th approach.
Number of primary roots. Only those over 0.5 mm were counted.
Volume estimator. d2 h was used as an estimator for the volume of plants in the six treatments. This is a combined variable that comes from the square of the value of the diameter of the root neck in cm (d2) and such value is multiplied by height in cm (h).
Data were taken from 308 plants by treatment: height was determined every two weeks during the three months that their cultivation lasted; the diameter of the neck, the aerial total dry matter, as well as the aerial and root, the volume variable and

Muñoz et al., Características morfológicas de plántulas...
27
HDT-27, durante 72 h a 70 °C hasta obtener un peso constante, en gramos con una aproximación de una centésima.
Biomasa seca aérea. La parte aérea de la planta, desde la yema terminal hasta el cuello donde se une con la raíz, se colocó en una estufa de secado MAPSA HDT-27, durante 72 h a 70 °C, hasta obtener un peso constante, en gramos y con una aproximación de una centésima.
Biomasa seca radical. La raíz se llevó a peso constante en una estufa de secado MAPSA HDT-27, durante 72 h a 70 °C, las unidades fueron en gramos con una aproximación de una centésima.
Número de raíces primarias. Se contabilizaron aquéllas que presentaron un diámetro mayor a 0.5 mm.
Estimador del volumen. Se empleó la expresión d2 h como estimador del volumen de las plantas en los seis tratamientos. Esta es una variable combinada que resulta de elevar al cuadrado el valor del diámetro del cuello de la raíz en cm (d2)
y dicho valor se multiplica por la altura en cm (h).
La toma de datos se llevo a cabo en 308 plantas por tratamiento: la altura se determinó cada quince días durante los tres meses de cultivo; el diámetro de cuello, la materia seca aérea total, aérea y radical, la variable volumen y el número de raíces primarias se evaluaron al final de la etapa de cultivo en el vivero.
RESULTADOS Y DISCUSIÓN
Los resultados del experimento se resumen en el Cuadro 1.
Tratamiento/Variable Supervivencia
(%)
Altura (cm)
Diámetro del
cuello (mm)
Estimador del
volumen (cc)
Número de raíces primarias
Materia seca
radical(g)
Materia seca
radical(g)
Materiaseca
radical(g)
1. Fibra de coco- Enterolobium 100 24.8 a 5.5 a 7.5 a 15.7 b 3.76 a 0.93 b 4.69 a
2. (77) Cavidades-Tabebuia 100 16.1 b 2.9 b 1.3 c 5.7 d 105 d 0.23 c 1.28 d
3. (60) Cavidades-Tabebuia 100 11.4 b 3.3 b 1.3 c 5.0 d 1.28 d 0.59 b 1.87 d
4. (77) Cavidades-Enterolobium 100 15.2 b 2.7 b 1.1 c 6.5 d 1.80 c 0.31 c 2.11 c
5. Fibra de coco-Tabebuia 100 12.0 b 6.2 a 4.7 b 18.7 a 2.17 b 1.49 a 3.66 b
6. (60) Cavidades-Enterolobium 100 14.9 b 3.2 b 1.5 c 8.7 c 1.74 c 0.28 c 2.02 c
the number of primary roots were assessed at the end of the cultivation stage at the nursery.
RESULTS AND DISCUSSION
The results of the experiment are in Table 1.
Survival. After three month in the nursery, survival in both species was 100%, which means that this variable was not affected either by the type of container or the roughness of the selected species. Similar results were quoted by Fisher (1992) and Topic et al. (2006). The loss of plants at the nursery is generally related to the quality of seed, the lack of water supply, the overheating of containers, low temperatures (under 0 oC), hail, the drying wind, the lack or excess of minerals and the attack of plagues and diseases (Alvarado et al., 2004; Barnett and McGilvray, 1997; James et al., 1995; Mexal et al., 1993; Ruano, 2002; Toral et al., 2000) and in the actual study, seedlings were not exposed to adverse conditions during production.
Total height. One of the indicators of plant quality is height, which is correlated with the amount of foliage, which provides photosynthetic potential and transpiration area. The tallest plants have some advantages upon the vegetation with which they compete, but are in disadvantage in places with low moisture, since they are unbalanced individuals, difficult to plant and susceptible to wind damage (Haase, 2006). The height reaches by E. cyclocarpum and T. rosea in the nursery stage,
Cuadro 1. Resultados de la evaluación de las variables consideradas y prueba de separación de medias de Tukey.Table 1. Results of the assessments of variables and Tukey’s mean separation test.
En la separación de medias con la prueba de Tukey, los valores sin letras en común en las columnas difieren significativamente a un α = 0.05.In Tukey’s mean separation test, values without common letters in the columns are significantly different at with α = 0.05.

Rev. Mex. Cien. For. Vol. 2 Núm. 8
28
Supervivencia. Después de tres meses en el vivero, la supervivencia en ambas especies fue del 100%, lo que indica que esta variable no fue afectada por el tipo de contenedor, ni por la rusticidad de las especies estudiadas. Resultados similares fueron citados por Fisher (1992) y Topic et al. (2006). La pérdida de plántulas en vivero generalmente se relaciona con la calidad de la semilla, la falta de suministro hídrico, el sobrecalentamiento de los contenedores, las bajas temperaturas (por abajo de cero grados), las granizadas, el viento desecante, la falta o el exceso de minerales y el ataque de plagas y enfermedades (Alvarado et al . , 2004; Barnett y McGilvray, 1997; James et al.,1995; Mexal et al.,1993; Ruano, 2002; Toral et al., 2000) y en el presente estudio las plántulas no estuvieron expuestas a condiciones adversas de producción.
Altura total. Uno de los indicadores de calidad de planta es la altura, que se correlaciona con la cantidad de follaje, el cual provee una capacidad fotosintética y es el área de transpiración. Las plantas más altas t ienen ventajas sobre la vegetación con la que compiten, pero están en desventaja en sit ios con baja humedad, ya que son individuos desbalanceados, difíciles de plantar y sujetos a daños por el viento (Haase, 2006). La altura alcanzada por E. cyclocarpum y T. rosea en la fase de vivero les permite ser competitivas al ser trasplantadas al lugar definitivo, donde la vegetación es agresiva, si se trata de plantaciones de conservación. Durante el periodo en el vivero, las especies E. cyclocarpum y T. rosea mostraron diferente altura por efecto del envase (Cuadro 1). Así en el contenedor de fibra de coco la de E. cyclocarpum fue mayor que la de T. rosea (Figura 1); así mismo, resultó significativamente superior (Pr > F= <0.0001) en relación con las alturas en las charolas de poliestireno de 60 y 77 cavidades (Cuadro 1). Respecto a T. rosea, los valores más altos se lograron en el contenedor de 77 cavidades, probablemente, porque los de fibra de coco y los de 60 cavidades ofrecen menor densidad, lo que significa más espacio de crecimiento y menor competencia (Figura 1).
. En particular, las plantas de T. rosea registraron una velocidad
de crecimiento que no fue significativamente diferente en los tres tipos de contenedores; sin embargo, durante la última evaluación se observó un ligero efecto de la densidad con plantas con alturas superiores, cuando se usaron charolas de 77 cavidades (Cuadro 1). Se consigna que las densidades altas de plantas por metro cuadrado promueven mayor crecimiento en altura, en este caso T. rosea mostró esa tendencia en los contenedores de 77 cavidades (385 plantas m-2), pero E. cyclocarpum tuvo valores más altos cuando la densidad fue menor (205 plantas m-2) en contenedores de fibra de coco (Mexal et al., 1993; Landis et al., 1994). Las especies de latifoliadas, generalmente, requieren volúmenes más grandes de sustrato, lo que resulta en densidades bajas de plantas por unidad de área, en comparación con las coníferas, porque el follaje de las latifoliadas absorbe más agua y nutrimentos y
allows them to be competitive when being transplanted to the final place, where vegetation is aggressive, if they are conservation plantations. During the nursery period, E. cyclocarpum and T. rosea showed different height due to effect of the container (Table 1). Thus, in the coconut-fiber container, E. cyclocarpum became taller than T. rosea (Figure 1); also, it was significantly higher (Pr > F= <0.0001) compared to the heights of seedlings from polystyrene trays of 60 and 77 pots (Table 1). In regard to T. rosea, the highest values were accomplished in the 77 pot containers, probably, because those from coconut-fiber and of 60 pots offer a lower density, which means broader space and less competence (Figure 1).
T. rosea plants in particular, did not record a growth speed that was not significantly different in the three type of containers; however, during the last assessment, a light effect of the density of plants with greater heights was observed, when 77 pot trays were used (Table 1). It is in the record that high density of plants per square meter promotes a better heoght growth; in this case, T. rosea showed this tendency in the 77 pot containers (385 plants m-2), but E. cyclocarpum had higher values when density was lower (205 plants m-2) in coconut-fiber containers (Mexal et al., 1993; Landis et al., 1994). Broadleaf species, regularly require grater substrate volumes, which gives way to low plant densities per area, compared to softwoods, since the foliage of the first absorbs more water and nutriments and produce more shadow (Landis et al., 1994). This competence for light favors a greater height, in regard to density and affects the morphology and weight of plants, as quoted by Timmis and Tanaka (1976). At the end of the assessment period, this behavior was observed in Tabebuia, but not in E. cyclocarpum.
Diameter of neck root. This is considered the best feature to predict plant survival and growth once they get established in the field; as the stem diameter is thicker, the amount of roots increases too (Haase, 2006). In the actual study, E. cyclocarpum and T. rosea plants in the coconut-fiber containers registered values within quality standards (Table 1). It has been quoted that a lower density, neck diameter is thicker, which has been proved for both species in 205, 284 and 364 plants m-2 (Timmis and Tanaka, 1976). At the end of the three month period at the nursery, the value of neck diameter had significant differences (Pr>F= <0.0001) among the six treatments; mean separation (Tukey’s test α=0.05) favored the formation of two groups. The first with the greatest records for both species from coconut-fiber containers, and the second, with smaller dimensions, for plants from 60 and 77 pot trays (Table 1).
The diameter of the root neck of E. cyclocarpum cultivated in coconut-fiber containers was 42% and 51% greater that the plants produced in 60 and 77 pot trays. For T. rosea they were 59% and 47% higher, in regard to those obtained with the same species in the polystyrene trays of 77 and 60 pots. In some studies it has been recorded that plants that grow
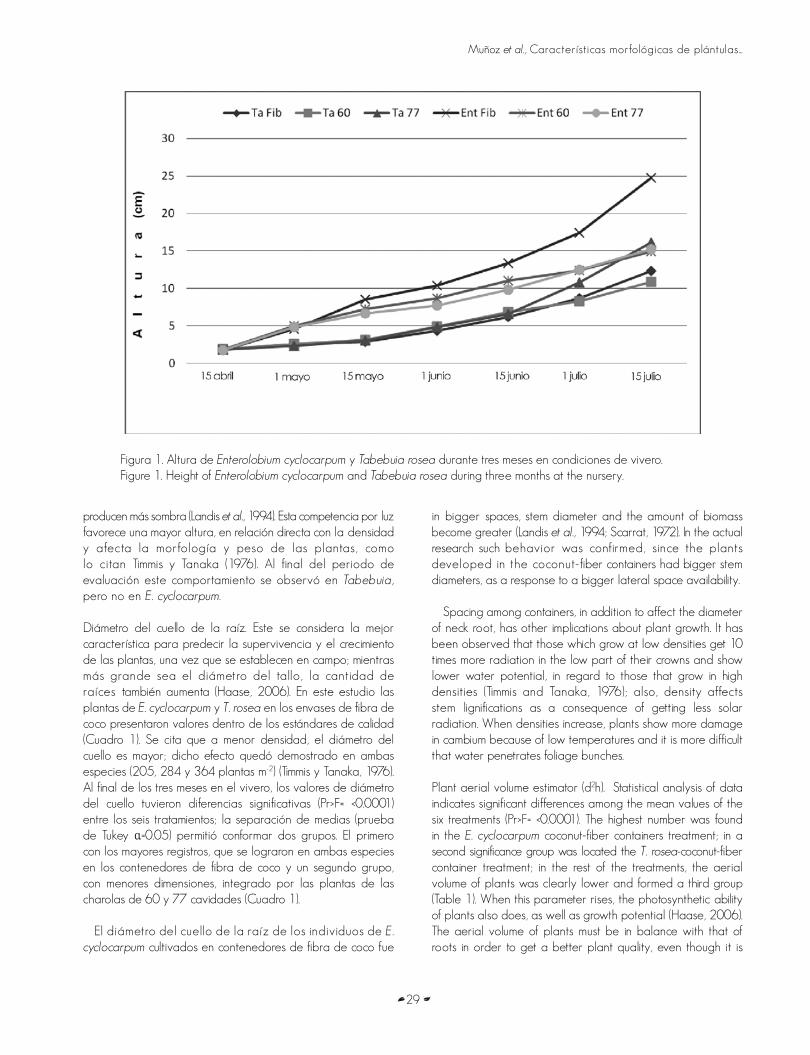
Muñoz et al., Características morfológicas de plántulas...
29
producen más sombra (Landis et al., 1994). Esta competencia por luz favorece una mayor altura, en relación directa con la densidad y afecta la morfología y peso de las plantas, como lo citan Timmis y Tanaka (1976). Al final del periodo de evaluación este comportamiento se observó en Tabebuia, pero no en E. cyclocarpum.
Diámetro del cuello de la raíz. Este se considera la mejor característica para predecir la supervivencia y el crecimiento de las plantas, una vez que se establecen en campo; mientras más grande sea el diámetro del tallo, la cantidad de raíces también aumenta (Haase, 2006). En este estudio las plantas de E. cyclocarpum y T. rosea en los envases de fibra de coco presentaron valores dentro de los estándares de calidad (Cuadro 1). Se cita que a menor densidad, el diámetro del cuello es mayor; dicho efecto quedó demostrado en ambas especies (205, 284 y 364 plantas m-2) (Timmis y Tanaka, 1976). Al final de los tres meses en el vivero, los valores de diámetro del cuello tuvieron diferencias significativas (Pr>F= <0.0001) entre los seis tratamientos; la separación de medias (prueba de Tukey α=0.05) permitió conformar dos grupos. El primero con los mayores registros, que se lograron en ambas especies en los contenedores de fibra de coco y un segundo grupo, con menores dimensiones, integrado por las plantas de las charolas de 60 y 77 cavidades (Cuadro 1).
El diámetro del cuello de la raíz de los individuos de E. cyclocarpum cultivados en contenedores de fibra de coco fue
in bigger spaces, stem diameter and the amount of biomass become greater (Landis et al., 1994; Scarrat, 1972). In the actual research such behavior was confirmed, since the plants developed in the coconut-fiber containers had bigger stem diameters, as a response to a bigger lateral space availability.
Spacing among containers, in addition to affect the diameter of neck root, has other implications about plant growth. It has been observed that those which grow at low densities get 10 times more radiation in the low part of their crowns and show lower water potential, in regard to those that grow in high densities (Timmis and Tanaka, 1976); also, density affects stem lignifications as a consequence of getting less solar radiation. When densities increase, plants show more damage in cambium because of low temperatures and it is more difficult that water penetrates foliage bunches.
Plant aerial volume estimator (d2h). Statistical analysis of data indicates significant differences among the mean values of the six treatments (Pr>F= <0.0001). The highest number was found in the E. cyclocarpum coconut-fiber containers treatment; in a second significance group was located the T. rosea-coconut-fiber container treatment; in the rest of the treatments, the aerial volume of plants was clearly lower and formed a third group (Table 1). When this parameter rises, the photosynthetic ability of plants also does, as well as growth potential (Haase, 2006). The aerial volume of plants must be in balance with that of roots in order to get a better plant quality, even though it is
Figura 1. Altura de Enterolobium cyclocarpum y Tabebuia rosea durante tres meses en condiciones de vivero.Figure 1. Height of Enterolobium cyclocarpum and Tabebuia rosea during three months at the nursery.

Rev. Mex. Cien. For. Vol. 2 Núm. 8
30
42% y 51% mayor que el de las plantas producidas en charolas de 60 y 77 cavidades. Para T. rosea fueron de 59% y 47% superior, respecto a los obtenidos con la misma especie en las charolas de poliestireno de 77 y 60 cavidades. En algunos estudios se registra que las plantas que crecen en espacios más grandes, el diámetro de tallo y la cantidad de biomasa son mayores (Landis et al., 1994; Scarrat, 1972). En el presente estudio, se comprobó este comportamiento, ya que las plantas desarrolladas en los envases de fibra de coco tuvieron diámetros de tallo más grandes, como respuesta a una mayor disponibilidad de espacio lateral.
El espaciamiento entre contenedores, además de influir en el diámetro del cuello de la raíz tiene otras implicaciones sobre el crecimiento de la planta. Se ha observado que aquéllas que crecen a bajas densidades reciben 10 veces más radiación en la parte baja de sus copas y muestran un potencial hídrico más bajo, en relación a las que se desarrollan en densidades altas (Timmis y Tanaka, 1976); así mismo, la densidad afecta la lignificación de los tallos como consecuencia de recibir menos radiación solar. Cuando la densidad aumenta las plantas presentan más daños en el cambium por las bajas temperaturas, y es más difícil que penetre el agua de riego en los manchones de follaje.
Estimador del volumen aéreo de las plantas (d2h). El análisis estadístico de los datos mostró diferencias significativas entre los valores medios de los seis tratamientos (Pr>F= <0.0001). La cifra más alta se registró en el tratamiento E. cyclocarpum-contenedores de fibra de coco; en un segundo grupo de significancia se ubicó el de T. rosea-contenedor de fibra de coco; en el resto de los tratamientos el volumen aéreo de las plantas fue claramente inferior y conformó un tercer grupo (Cuadro 1). Cuando este parámetro aumenta, también lo hace la capacidad fotosintética de las plantas y, por lo tanto, el potencial de crecimiento se incrementa (Haase, 2006). El volumen aéreo de las plantas debe estar en balance con el de las raíces, a fin de obtener una mayor calidad de planta, aunque es preciso tomar en cuenta que un área de transpiración demasiado grande conduce a estrés en sitios secos, antes de que el sistema radical se establezca. En dichos ambientes se requerirán riegos de auxilio o bien proteger la raíz temporalmente para evitar la total desecación.
Número de raíces primarias. El número de raíces primarias está correlacionado con la sobrevivencia de las plantas en campo. El régimen de cultivo de plantas en vivero y las características del contenedor inciden en la emisión de raíces y en el volumen radical. El análisis de la información del efecto de los tres tipos de contenedores evaluados determinó diferencias significativas (Pr>F= <0.0001). El mayor número de raíces se registró en los contenedores de fibra de coco. T. rosea tuvo la cifra más alta, aunque las dos especies estudiadas registraron una respuesta positiva respecto a la emisión de raíces primarias. En relación con las charolas de poliestireno,
necessary to take into account that a rather big transpiration area favors stress in dry places, before the radical system is fixed. In such environments help watering will be demanded or otherwise, to protect the root temporarily in order to avoid total desiccation.
Number of primary roots. The number of primary roots is correlated with the cultivation of plants at the nursery. The regime of plant cultivation at the nursery and the details of the container have an impact upon the formation of roots and root volume. The analysis of the information about the effect of the three type of containers that were evaluated determined significant difference (Pr>F= <0.0001). The greatest number of roots was found in the coconut-fiber containers. T. rosea had the highest number, even though the two selected species, had a positive reaction of primary root formation. In regard to the polystyrene trays, T. rosea had similar results in all cases, while E. cyclocarpum showed a better behavior in container of 60 pots, but the difference was not significant compared to what was observed in trays of 77 pots (Table 1).
Aerial, root and total dry biomass. These variables have related positively with survival in the field and the following growth (Haase, 2006). In the three types of containers there were significant differences (Pr>F= <0.0001).
In aerial biomass, coconut-fiber treatments were outstanding; nevertheless, E. cyclocarpum overpassed T. rosea in the expanded polystyrene containers. The values of the 60 and 77 pot trays were similar (Table 1). In regard to root biomass, the most important results came from coconut-fiber containers, but T. rosea significantly went ahead E. cyclocarpum. The walls of the coconut-fiber container widely favor the elongation of roots and their pruning, which avoids their spiral-shape growth, in such a way that they increase ramification, and, consequently, a great amount of roots is produced, as these results prove compared to what happened with polystyrene trays. Coconut-fiber containers have been used for years since they have several advantages over plastic containers: they promote vigorous root systems, they avoid root curling, they keep growth media fresh through wall evaporation, which promotes the proliferation of roots, protect the root system from heat and cold, they help to produce bigger and more vigorous plants, and they are bio-degradable (Ruter, 1995; Ruter, 2000).
Tabebuia rosea in Styroblock trays has a better response in containers of greater capacity and space, those of 60 pots, and the reaction of E. cyclocarpum was similar in both containers. These trays are easily damaged with handling and if plants remain at the nursery for long times, roots penetrate polystyrene walls, a fact that makes it difficult to extract them. (Barnett and McGilvray, 1997).
The value of the total biomass variable is the result of the addition of the aerial and root biomass, and consequently,

Muñoz et al., Características morfológicas de plántulas...
31
T. rosea tuvo resultados similares en todos los casos; mientras que E. cyclocarpum presentó un mejor comportamiento en contenedores de 60 cavidades, pero la diferencia no fue significativa respecto a los observados en las charolas de 77 cavidades (Cuadro 1).
Biomasa seca aérea, radical y total. Estas variables se han relacionado positivamente con la supervivencia en el campo y el crecimiento subsecuente (Haase, 2006). En los tres tipos de contenedores se obtuvieron diferencias significativas (Pr>F= <0.0001).
En la biomasa aérea sobresalieron los tratamientos que incluyeron los envases de fibra de coco; no obstante, E. cyclocarpum superó a T. rosea en los de poliestireno expandido. Los valores observados en las charolas de 60 y 77 cavidades fueron muy semejantes (Cuadro 1). En relación a la biomasa radical, los mayores registros se lograron en los contenedores de fibra de coco, solo que T. rosea superó a E. cyclocarpum significativamente. Las paredes del contenedor de fibra de coco favorecen ampliamente la extensión de raíces y su poda, lo que evita su crecimiento en espiral; de tal manera que se incrementan las ramificaciones y en consecuencia se obtiene una gran cantidad de raíces, como se demuestra al comparar los resultados obtenidos con las charolas de poliestireno. Los envases de fibra de coco se han empleado por años porque presentan varias ventajas sobre los contenedores de plástico: promueven los sistemas radicales vigorosos, evitan el enrollamiento de las raíces, mantienen fresco el medio de crecimiento a través de la evaporación por las paredes, lo que a su vez promueve la proliferación de las raíces, protegen al sistema radical del calor y del frío, ayudan a producir plantas más grandes y vigorosas, además de ser biodegradables (Ruter, 1995; Ruter, 2000).
Tabebuia rosea en charolas Styroblock respondió mejor en los contenedores de mayor capacidad y espacio, los de 60 cavidades, y la respuesta de E. cyclocarpum fue similar en ambos envases. Estas charolas se dañan fácilmente con el manejo y si las plantas permanecen en el vivero por largos periodos, las raíces penetran en las paredes de poliestireno lo que dificulta la extracción del cepellón (Barnett y McGilvray, 1997).
El valor de la variable biomasa total es el resultado de la suma de las biomasas aérea y la radical, y en consecuencia, se logran los mayores valores para las dos especies en contenedores de fibra de coco, aunque los de E. cyclocarpum fueron significativamente superiores con respecto a los de T. rosea. En las charolas de 60 cavidades T. rosea registró crecimiento significativamente superior al observado en las de 77 cavidades. Los resultados en E. cyclocarpum los fueron similares en ambos tipos de charolas (Cuadro 1).
the greatest values for the two species in coconut fiber containers are obtained, even though E. cyclocarpum were significantly higher to those of T. rosea. In the 60 pot trays, T. rosea had a significantly greater growth than that observed in those of 77 pots. E. cyclocarpum results were similar in both types of trays (Table 1).
CONCLUSIONS
Enterolobium cyclocarpum and Tabebuia rosea showed a different growth rhythm; Enterolobium became higher and Tabebuia rosea formed a greater root neck.
The best height growth results came from the coconut-fiber containers for Enterolobium cyclocarpum and in root neck diameter for T. rosea. Both were significantly higher than those obtained in polystyrene trays of 60 and 77 pots, and they also developed greater biomass values than those of polystyrene trays.
Starting from the materials used in their manufacturing, their structure and plant density by square meter, coconut-fiber containers allow the production of plants with high morphological quality (healthier and more abundant root systems) compared to those plants propagated in expanded polystyrene trays of 60 and 77 pots.
Coconut-fiber containers keep their structure during the nursery stage, a most-wanted feature for high quality plant production. In addition, plant roots penetrate the walls of the container, which promotes a greater fiber formation, and that, on the other hand, guarantees a better fixation of the plant in the field.
ACKNOWLEDGEMENTS
Our gratitude to Artifibras S. A de C. V. Company for their economic support to accomplish this study, as well as to the Education and Forest Training Center No. 1 of CONAFOR at Uruapan city, for letting us to fulfill the experiments at their forest nursery.
End of the English version

Rev. Mex. Cien. For. Vol. 2 Núm. 8
32
CONCLUSIONES
Enterolobium cyclocarpum y Tabebuia rosea mostraron diferente ritmo de crecimiento, Enterolobium presentó mayor altura y Tabebuia rosea un diámetro del cuello de la raíz superior.
En contenedores de fibra de coco se obtuvieron los mejores crecimientos en altura para Enterolobium cyclocarpum y en diámetro del cuello de la raíz, para T. rosea. Ambos fueron significativamente superiores a los obtenidos en charolas de poliestireno de 60 y 77 cavidades. Las dos especies también registraron los valores de biomasa significativamente más grandes a los de las charolas de poliestireno.
Por los materiales empleados en su fabricación, su estructura y la densidad de planta por metro cuadrado, los contenedores de fibra de coco permiten obtener plantas de alta calidad morfológica (sistemas radicales más abundantes y sanos) en comparación con los de las plantas propagadas en charolas de poliestireno expandido de 60 y 77 cavidades.
Los envases de fibra de coco mantienen su estructura durante la etapa de vivero, característica deseable para la producción de planta de calidad. Además las raíces de las plantas penetran por las paredes del contenedor lo que promueve mayor fibrosidad, que a su vez garantiza un mejor anclaje de las plantas en campo.
AGRADECIMIENTOS
Se agradece a la Empresa ARTIFIBRAS S.A de C.V. por el apoyo económico para la realización del presente estudio, así como al Centro de Educación y Capacitación Forestal 01-CONAFOR Uruapan, Michoacán, por permitirnos llevar a cabo el estudio en vivero forestal a su cargo.
REFERENCIAS
Alvarado R., D., S. Castro., C. Cigarrero C., R. Álvarez R. y L. de L. Saavedra R. 2004. Manual de detección y manejo de enfermedades bajo el sistema de contenedor. Caso Vivero San Luis Tlaxialtemalco. CONAFOR. México, D. F. México. 74 p.
Barnett, P. and J. M. McGilvray. 1981. Container planting systems for the South. Southern Forest Experimental Station. USDA Forest Service. New Orleans, LA. USA. 18 p.
Barnett, P. and J. M. McGilvray. 1997. Practical guidelines for producing longleaf pine seedlings in containers. General Technical Report SRS-14. Southern Research Station. USDA Forest Service. Asheville, NC. USA. 28 p.
Burdett, A. N., D. G. Simpson; and C. F. Thompson. 1983. Root development and plantation establishment success. Plant and Soil 71(1-3): 103-110.
Cuny, H. 1996. The container evolution. American Association of Nurserymen. Nursery Management & Production 12(8): 56-61.
Duryea, M. L. and K. L. McClain. 1984. Altering seedling physiology to improve reforestation success. In: Duryea, M. L. and G. N. Brown (Eds.). Seedling physiology and reforestation success. In: Proceedings of the Physiology Working Group, Technical Session: Society American Foresters National Convention, Portland. OR USA October 16th-20th, 1983. Martinus Nijhoff Publishers. Dordrecht, The Netherlands. 114 p.
Fisher, J. T. 1992. Container plant propagation. Forest nursery training course. New Mexico State University. Las Cruces, NM. USA. 210 p.
García M., J. J. y H. J. Muñoz F. 1993. Guía para la producción de planta forestal en envases. Guía Técnica Núm. 3. CIRPAC. INIFAP. Uruapan, Mich. 47 p.
González K., V. 1995. Viveros forestales. Centro Nacional de Investigación Disciplinaria en Conservación y Mejoramiento de Ecosistemas Forestales. INIFAP. México, D.F. México. Publicación Especial No.3. pp. 3-7.
Haase, D. L. 2006. Morphological and physiological evaluations of seedling quality. In: Riley, L. E., R. K. Dumroese, T. D., Landis (Coords.). 2007. National Proceedings: Forest and Conservation Nursery Associations. 2006. RMRS-P-50. USDA, Forest Service. Rocky Mountain Research Station. Fort Collins, CO. USA. pp: 3-8. http://www.rngr.net/nurseries/publicatons/proceedings (22 de junio de 2008).
James, R. L., R. K. Dumroese and D. L. Wenny. 1995. Botrytis cinerea carried by adult fungus gnat (Diptera: Sciaridae) in container nurseries. Tree Panters Notes, Winter, 48(2): 53.
Landis, T. D., R. W; Tinus., S. E. McDonald and J. P., Barnett. 1994. The Container Tree Nursery Manual. Agricultural Handbook 674. U.S.D.A. Forest Service. Washington, DC. USA. Vol. 2. 88 p.
Liegel, L. H. and C. R. Venator. 1987. A technical guide for forest nursery management in the Caribbean and Latin America. G.T.R. SO-67. Southern Forest Experimental Station. USDA Forest Service. New Orleans, LA. USA. 156 p.
Martínez G., A. 1988. Diseños experimentales. Editorial Trillas. México, D. F. México. pp. 118-160.
Mexal, J. G., R. W. Newman, J. T. Fisher, R. Phillips, T. Sammis and T. D. Landis. 1993. Forest nurseries: planning, establishment and seedling growth. USDA, Forest Service, New Mexico State University. International training course. 73 p.
Patiño, V. F. y J. Marín Ch. 1983. Viveros forestales. Planeación, establecimiento y producción de planta. Libro Técnico. SARH. CIR Sureste INIFAP.Mérida, Yuc. México. 159 p.
Peterson, M. and J. R. Sutherland. 1989. Grey mould control by seedling canopy humidity reduction through under-bench ventilation and Styroblock aeration. FRDA Rep. 077. Forestry Canada, Pacific Forestry Centre. Victoria, BC. Canada. 16 p.
Ritchie, G. A. 1984. Root growth potential. Principles procedures, and predictive ability. In: Duryea, M. L. (Ed.).Proceedings. Evaluating seedling quality: principles, procedures and predictive abilities of major tests. Forest Research Laboratory, Oregon State Corvallis University, OR. USA. p. 93-105.
Ruano M., J. R. 2002. Viveros forestales. Manual de cultivo y proyectos. Mundi Prensa. México, D. F. México, 281 p.
Ruter, M. R. 1995. Growth of coreopsis and plumbago in plastic and Cu (OH)
2 impregnated fiber containers. Hort Technology 5(4):
300-302.Ruter, J. M. 2000. Cross-country containers. American Nurseryman 191
(3): 26-30.Scarrat, J. B.1972. Effect of tube diameter and spacing of tubed seedling
planting stock. Rep. O-X-170. Great Lakes Forest Research Centre. Canadian Forest Service. Sault St. Marie, Ontario Canada. 16 p.
Statistical Analysis System Institute.2004. SAS / ETS® 9.1. Users Guide. SAS Institute Inc. Cary. NC.USA. 2170 p.
Sims, J. 1988. Shifting from paperpots to ecopots. Silviculture Magazine 3(2): 10-12.

Muñoz et al., Características morfológicas de plántulas...
33
Strachan, M. D. 1974. Tar paper containers: Engineering the container. Great Plains Agricultural Council Publication No.68. In: Tinus, R. W., W. I. Stein and W. E. Balmer. (Eds.). Proceedings of the North American Containerized Forest Tree Seedling Symposium. Denver, CO. USA. pp. 209-210.
Stuewe, E. 2006. Trends in container types. In: Riley, L. E., R. K. Dumroese and T. D. Landis. (Coords.). National Proceedings: Forest and Conservation Nursery Associations-2005. RMRS-P-43- Rocky Mountain Research Station. USDA. Forest Service.Fort Collins, CO. USA. 160 p. http://www.rngr.net/nurseries/publications/proceedings. (15 de julio de 2008).
Timmis, R. and Y. Tanaka. 1976. Effects of container density and plant water stress on growth and cold hardiness of Douglas-Fir seedlings. Forest Science 22(2): 167-172.
Topic, V., L. Butorac, G. Jelic and S. Peric. 2006. Influence of container type on growth and development of holm oak (Quercus ilex L.) seedlings in a nursery. Periodicum Biologorum 108(6): 643-648.
Toral I., F. M., D. Campos R., A. Fratti B. y R. Varela O. 2000. Manual de producción de plantas forestales en contenedores. PRODEFO. Documento Técnico 25. Guadalajara, Jal., México. 219 p.
Wenny, D. L., Y. Liu, R. K. Dumroese and H. L. Osborne. 1988. First year field growth of chemically root pruned containerized seedlings. New Forests 2(2): 111-118.
Wright, J. A., J. Escobar and G. Henderson. 1999. Utilization of Jiffy pellets in the production of pine and eucalyptus seedlings, pine rooted cuttings and native species propagation. Nursery and Field Comparisons. In: Landis, T. D., and J. P Barnett. (Coords.). National Proceedings: Forest and Conservation Nursery Association. Gen. Tech. Rep. SRS-25. Forest Service, Southern Research Station. USDA. Department of Agriculture. Asheville, NC. USA. p. 54-56.

Rev. Mex. Cien. For. Vol. 2 Núm. 8
34
Wangari Muta Maathai. Dominio público.

HONGOS OPHIOSTOMATOIDES DE GALERIAS DE Dendroctonus adjunctus Blandford EN Pinus hartwegii Lindl.
OPHIOSTOMATOID FUNGI OF GALLERIES OF Dendroctonus adjunctus Blandford IN
Pinus hartwegii Lindl.
Omar Alejandro Pérez Vera 1, Dionicio Alvarado Rosales 1, David Cibrián Tovar 2,Armando Equihua Martínez 3 y Elizabeth Cárdenas Soriano 1
RESUMEN
Los descortezadores (Coleoptera: Curculionidae: Scolytinae) son vectores del hongo Ophiostoma spp., causante del manchado azul de la madera que contribuye a la muerte de árboles en coníferas y angiospermas. En México, Dendroctonus adjunctus es una de las principales plagas de los bosques de pino, pero se desconocen los hongos simbiontes que introduce en sus galerías. El objetivo del presente estudio fue identificar los hongos ophiostomatoides asociados al pino de las alturas (Pinus hartwegii). Se colectaron 20 muestras de corteza con galerías del insecto en la Estación Forestal Experimental Zoquiapan (EFEZ) de la Universidad Autónoma Chapingo, Estado de México. Las muestras se desinfestaron con hipoclorito de sodio, se colocaron en cámaras con alta humedad relativa e incubaron a 25 ± 2 °C en oscuridad durante 30 días, para el desarrollo de estructuras reproductivas. Los hongos fueron aislados en extracto-malta-agar al 2%; se hizo su caracterización morfológica y molecular, esta mediante la amplificación de la región intergénica (ITS) de los genes ribosomales rADN, con la técnica de la reacción en cadena de la polimerasa (PCR). Se registraron cinco hongos: Leptographium guttulatum, Ophiostoma angusticollis, O. nigrocarpum, O. olivaceapini y Pesotum sp., con la presencia del teleomorfo y del anamorfo en el medio de cultivo. Los análisis moleculares corroboraron las identificaciones, sin embargo, O. nigrocarpum y O.angusticollis mostraron una homología con el género Sporothrix (anamorfo) en un 99% y Pesotum un 98% con Ophiostoma (teleomorfo). Las secuencias se depositaron en el Banco de Genes del NCBI.
Palabras clave: Descortezador, Leptographium guttulatum M.J. Wingf. & K. Jacobs, Ophiostoma angusticollis (E.F. Wright & H.D. Griffin) M. Villarreal, Ophiostoma nigrocarpum (R.W. Davidson) de Hoog, Ophiostoma olivaceapini (R.W. Davidson) Seifert & G. Okada, Pesotum J.L. Crane & Schokn.
ABSTRACT
Bark beetles (Coleoptera: Curculionidae: Scolytinae) are vectors of Ophiostoma fungi species which cause the blue-stained wood disease and contribute to the death of conifer and angiosperm trees. In Mexico, Dendroctonus adjunctus is one of the major pests of pine forests; however, most of the symbiotic fungi introduced into its galleries are unknown. The main objective of the present study was to identify and characterize ophiostomatoid fungi associated with hartwegii pine (Pinus hartwegii). Twelve bark beetle galleries were collected in the Zoquiapan Experimental Forest Station (EFEZ by its acronym in Spanish) from the Universidad Autonoma Chapingo in Zoquiapan, State of Mexico. Samples were disinfected with sodium hypochlorite, placed in chambers with high relative humidity and incubated at 25 ± 2 °C in darkness during 30 days for the development of reproductive structures. Fungi were isolated on 2% agar-malt extract; its identification was done using morphological characters and were molecularly characterized by means of amplification of the internal transcribes spacer (ITS) region of the ribosomal rDNA genes using the Polymerase chain reaction (PCR) technique. Five fungi were identified: Leptographium guttulatum, Ophiostoma nigrocarpum, O. angusticollis O. olivaceapinii and Pesotum sp. The molecular analysis confirmed that O. nigrocarpum and O.angusticollis showed 99% homology levels with the genus Sporothrix (anamorph), whereas Pesotum had 98% homology levels with Ophiostoma (teleomorph). The sequences were deposited in the Gene Bank of the NCBI.
Key words: Bark beetle, Leptographium guttulatum M.J. Wingf. & K. Jacobs, Ophiostoma angusticollis (E.F. Wright & H.D. Griffin) M. Villarreal, Ophiostoma nigrocarpum (R.W. Davidson) de Hoog, Ophiostoma olivaceapini (R.W. Davidson) Seifert & G. Okada, Pesotum J.L. Crane & Schokn.
Fecha de recepción: 20 de agosto de 2010.Fecha de aceptación: 23 de diciembre de 2011.1 Programa de Fitopatología. Colegio de Postgraduados. Campus Montecillo. Correo-e: [email protected] División de Ciencias Forestales (DICIFO), Universidad Autónoma Chapingo.3 Programa de Entomología y Acarología. Colegio de Postgraduados, Campus Montecillo.

Rev. Mex. Cien. For. Vol. 2 Núm. 8
36
INTRODUCTION
Ceratocystis sensu lato Ellis & Halst. belong to the ophiostomatoid group and include more than 110 species of ascomycetes that have similar morphological features and are adapted for dispersion by arthropods, such as the Dendroctonus insects (Upadhyay, 1993; Malloch and Blackwell, 1993). Several taxa of ophiostomatoides are important softwood and hardwood pathogens (Wingfield, 1993; Jacobs and Wingfield, 2001), and others can cause staining of logs or wood recently cut (Wingfield, 1993) or damage crops such as cotton, sweet potato, cacao, coffee, sugar cane, mango, pineapple, banana, tobacco and carrot (Kile, 1993).
The spores of these fungi are produced in the galleries of insects and are transporte in the exoskeleton, mycangia or digestive system of the barkbeetle (Coleoptera: Curculionidae: Scolytinae); such fungi group has a nutritional mutualistic or anthagonistic effect, or non over the insect (Paine et al., 1997; Ayres et al., 2000). It has been put in the record that they help to kill a tree as they shorten the availability of water, sugar and monotherpenoid accumulation potentially toxic for the phloem and xylem (Popp et al., 1995; Croisé et al., 2001).
In North America and Europe Ophiostoma ulmi (Buisman) Nannf. and O. novo-ulmi Brasier have been detected as responsible for the Dutch elm disease (DED) (Wingfield, 1993). Leptographium wageneri var. wageneri (W. B. Kendr.) M. J. Wingf. has been observed in Pinus monophylla Torr. & Frém. and P. edulis Engelm.); L. wageneri (W. B. Kendr.) M. J. Wingf. var. pseudotsugae T. C. Harr. & F.W. Cobb in Pseudotsuga menziesii (Mirb.) Franco and L. wageneri (T. C. Harr. & F. W. Cobb) var. ponderosum T. C. Harr. & F. W. Cobb in P. ponderosa Laws., P. contorta (Balf.) Critchfield and P. jeffreyi Murr. Also, they are the causing agents of the black stain root disease in the United States of America and Canada (Harrington, 1988; Wingfield and Knox- Davies, 1980).
In Mexico, 17 ophiostomatoid fungi associated with Conophthorus cembroides Woods, Dendroctonus adjunctus Blandford, D. mexicanus Hopkins, D. valens LeConte, Ips calligraphus Germar and Pseudohylesinus spp. Blandford, which are insects that feed from the phloem and vascular cambium, have been reported by Marmolejo, 1991; Marmolejo and Butin, 1993; Zhou et al., 2004; Cibrián et al., 2007 and Pérez et al., 2009.
The most common taxa that cause wood staining are Ophiostoma piliferum (Fr.) Syd. & P. Syd. and O. ips (Rumbold) Nannf. (Marmolejo and Butin, 1993; Marmolejo, 1991; Pérez et al., 2009). Ceratocystis fimbriata Ellis & Halst. has been quoted as the vascular fungus of Hevea brasiliensis Muell. Arg., and its presence leads to the death of the tree, or recently cut wood staining and lowers gum harvest in forest plantations (Cibrián et al., 2007). The three species have been reported
INTRODUCCIÓN
Ceratocystis sensu lato Ellis & Halst. pertenecen al grupo de los ophiostomatoides e incluyen más de 110 especies de ascomicetes que presentan características morfológicas similares y están adaptados a dispersarse por artrópodos, como los insectos del género Dendroctonus (Upadhyay, 1993; Malloch y Blackwell, 1993). Varios taxa de ophiostomatoides son patógenos importantes de coníferas y angiospermas (Wingfield, 1993; Jacobs y Wingfield, 2001), otros pueden causar un manchado en troncos y en la madera recién cortada (Wingfield, 1993) o afectar cultivos como algodón, camote, cacao, café, caña de azúcar, guayaba, mango, piña, plátano, tabaco y zanahoria (Kile, 1993).
Las esporas de estos hongos se producen en las galerías de insectos y son transportadas en el exoesqueleto, micangios o aparato digestivo del descortezador (Coleoptera: Curculionidae: Scolytinae); dicho grupo fúngico tiene un efecto nutricional mutualista o antagonista, o bien ninguno sobre el insecto (Paine et al., 1997; Ayres et al., 2000). Se ha consignado que ayudan a matar al árbol, por la reducción de agua, azúcares y acumulación de monoterpenoides potencialmente tóxicos en el floema y xilema (Popp et al., 1995; Croisé et al., 2001).
En América del Norte y Europa se tiene identificado a Ophiostoma ulmi (Buisman) Nannf. y O. novo-ulmi Brasier como responsables de la enfermedad del olmo Holandés (Wingfield, 1993). Leptographium wageneri var. wageneri (W. B. Kendr). M. J. Wingf. se han observado en pinos piñoneros (Pinus monophylla Torr. & Frém. y P. edulis Engelm), L. wageneri (W.B. Kendr) M. J. Wingf. var. pseudotsugae T. C. Harr. & F. W. Cobb en Pseudotsuga menziesii (Mirb) Franco y L. wageneri (T. C. Harr. & F. W. Cobb) var. ponderosum T.C. Harr. & F.W. Cobb en pinos duros (P. ponderosa Laws., P. contorta (Balf) Critchfield y P. jeffreyi Murr). Además son los agentes causales de la enfermedad del teñido oscuro de raíces, en Estados Unidos y Canadá (Harrington, 1988; Wingfield y Knox- Davies, 1980).
En México se han registrado 17 hongos ophiostomatoides asociados con insectos que se alimentan del floema y cambium vascular: Conophthorus cembroides Woods, Dendroctonus adjunctus Blandford, D. mexicanus Hopkins, D. valens LeConte, Ips calligraphus Germar y Pseudohylesinus spp. Blandford (Marmolejo, 1991; Marmolejo y Butin, 1993; Zhou et al., 2004; Cibrián et al., 2007; Pérez et al., 2009).
Los taxa más frecuentes que provocan manchado en la madera son Ophiostoma piliferum (Fr.) Syd. & P. Syd. y O. ips (Rumbold) Nannf. (Marmolejo y Butin, 1993; Marmolejo, 1991; Pérez et al., 2009). Ceratocystis fimbriata Ellis & Halst. se cita como un hongo vascular en Hevea brasiliensis Muell. Arg., cuya presencia induce la muerte del árbol, o manchados en la madera recién cortada y reduce la cosecha de goma en

Pérez et al., Hongos ophiostomastoides de...
37
as pathogens in many parts of the world (Seifert, 1993). They have been isolated from insects and their galleries and their identification has been based on their morphological characteristics, even though the absence of a sexual stage makes it difficult for taxonomic endings (Zhou et al., 2001).
The ITS sequences of the ribosomal genes (rDNA) have been successfully used to determine the phylogenetic relations among ophiostomatoid fung, since the morphology o their ascospores is a non-reliable taxonomic feature (Hausner et al., 1993; Wingfield et al., 1994). Zhou et al. (2001) increased the internal ITS regions of the rRNA genes and with that method identified Ophiostoma pulvinisporum X. D. Zhou & M. J . Wingf . and Sporothrix sp. Hektoen & C. F. Perkins) in México. The aim of this study was to identify ophiostomatoid fungi associated with Pinus hartwegii Lindl.
MATERIALS AND METHODS
Sampling in the field
20 Pinus hartwegii bark samples with insect galleries of Dendroctonus adjunctus were collected at the Zoquiapan Forest Experimental Station (EFEZ) that belongs to Universidad Autónoma Chapingo, at Mexico State. The material was labeled and placed into polypaper bags and was taken to the Forest Pathology Laboratory off the Graduate Studies College, Montecillo-Texcoco, where they were preserved at 4�C for its further analysis.
Fungus isolation
Samples were washed with distilled water, disinfected with 1.5% commercial sodium hypochloride for 2 min; they were rinsed thrice with sterile distilled water, were dried on sterile paper and finally were put into moist chambers. They were incubated at 25 ± 2 °C in darkness for 30 days in order to induce sexual or asexual structure formation. Ascospore masses or conidia present on the apex of reproductive structures were transferred with a sterile dissection needle to Petri boxes with 2% MAE (malt-agar-extract), 0.1g cycloheximide and 0.02 g streptomycin sulfate and incubated at 25 ± 2 °C under natural light for 20 days. Then, fragments of the fungus developed in Petri boxes with 2% MAE were transferred and incubated under the same isolation conditions. Monoconidial or monoascosporic culture were made in Petri boxes with water-agar and were incubated at 25 ± 2 °C for 24 h. Germinated conidia and ascospores were individually put into Petri boxes with 2% MAE and were incubated until the sexual or asexual structure was formed. All of the isolations were preserved in leaning assay tubes with 2% MAE and covered with mineral oil.
plantaciones forestales (Cibrián et al., 2007). Las tres especies han sido registradas como patógenas en muchas partes del mundo (Seifert,1993). Se han aislado de insectos y de sus galerías y su identificación se ha hecho con base en sus características morfológicas aunque, la ausencia del estado sexual dificulta su separación taxonómica (Zhou et al., 2001).
Las secuencias de las regiones ITS de los genes ribosomales (rADN) se han usado con éxito para determinar las relaciones filogenéticas entre los hongos ophiostomatoides, ya que la morfología de sus ascosporas es un carácter taxonómico no confiable (Hausner et al., 1993; Wingfield et al., 1994). Zhou et al. (2001) amplificaron las regiones internas ITS de los genes rRNA y con ese método identificaron a Ophiostoma pulvinisporum X. D. Zhou & M. J. Wingf. y Sporothrix sp. Hektoen & C. F. Perkins) en México. El objetivo del presente trabajo fue identificar hongos ophiostomatoides asociados a Pinus hartwegii Lindl.
MATERIALES Y MÉTODOS
Muestreo en campo
Se colectaron 20 muestras de corteza de Pinus hartwegii con galerías del insecto descortezador Dendroctonus adjunctus en la Estación Forestal Experimental Zoquiapan (EFEZ) de la Universidad Autónoma Chapingo, Estado de México. El material se etiquetó, colocó en bolsas de polipapel y se trasladó al Laboratorio de Patología Forestal del Colegio de Postgraduados, Campus Montecillo-Texcoco, Estado de México, donde se conservaron a 4 °C hasta su posterior análisis.
Aislamiento del hongo
Las muestras se lavaron con agua destilada, se desinfestaron con hipoclorito de sodio comercial al 1.5% durante 2 min, se lavaron tres veces con agua destilada estéril, se secaron con papel estéril y finalmente se colocaron en cámaras húmedas. Estas se incubaron a 25 ± 2 °C en oscuridad durante 30 días para inducir la formación de estructuras sexuales o asexuales. Las masas de ascosporas o conidios presentes en el ápice de las estructuras reproductivas se transfirieron con una aguja de disección estéril a cajas de Petri con extracto-malta-agar al 2% (EMA), cicloeximida (0.1g) y sulfato de estreptomicina (0.02g) e incubaron a 25 ± 2 °C en luz natural durante 20 días. Se transfirieron fragmentos del hongo desarrollado a cajas Petri con EMA (2%) e incubaron en las mismas condiciones del aislamiento. Los cultivos monoconidiales o monoascospóricos se realizaron en cajas Petri con agar-agua (AA) y se incubaron a 25 ± 2 °C durante 24 h. Las conidias y ascosporas germinadas se colocaron, individualmente, en cajas de Petri con EMA (2%) y se incubaron hasta que se formara la estructura sexual o asexual. Todos los aislamientos se conservaron en tubos de ensayo inclinados con EMA (2%) y cubiertos con aceite mineral.

Rev. Mex. Cien. For. Vol. 2 Núm. 8
38
Identificación morfológica
A partir de 10 cultivos puros (monoconidiales o monoascopóricos) en EMA (2%) a 25 ± 2 °C e incubados durante 40 días se identificaron los hongos a nivel de género con las claves taxonómicas de Barnett y Hunter (1972) y Hanlin (2001), y para las especies se emplearon las de Upadhyay (1981) y Jacobs y Wingfield (2001). Además, se observó la forma de crecimiento, color de la colonia, tipo de micelio, formación de estructuras reproductivas, tipo y forma de conidios o ascosporas, en las que se midieron en micras 200 ascosporas y conidios y 30 peritecios o sinemas. El seguimiento al desarrollo de conidios y tipo de ascosporas se hizo con microscopía de barrido; para ello se tomaron discos con crecimiento fungal de 5 mm de diámetro. Las muestras se fijaron en glutaraldehído al 3% con amortiguador de fosfatos Sorensen 0.1M, pH 7.2, durante 24 h. Posteriormente, se lavaron tres veces con el amortiguador de fosfatos Sorensen por un minuto y se deshidrataron en una serie de soluciones de etanol (30, 40, 50, 60, 70, 80, 90 y 100%, las últimas tres soluciones se usaron dos veces) por 15 min. en cada cambio. Los discos se colocaron en una secadora de punto crítico de CO
2 Samdri-780A® (TOUSIMIS Research
Corporation, Rockville, EUA) a 31.1 °C y 1073 psi; enseguida, los discos se pegaron en un portamuestras de latón de 9 mm de diámetro con cinta adhesiva-conductiva de cobre y se recubrieron con oro durante 4 min en una ionizadora JFC-1100® (JEOL LTD, Tokio, Japón). Las muestras se revisaron en un microscopio electrónico de barrido JSM-35C® (JEOL LTD, Tokio, Japón) operado a 15 Kv en el laboratorio de Microscopia Electrónica del Colegio de Postgraduados.
Caracterización molecular
Los 10 aislamientos de hongos puros utilizados para extraer el ADN se cultivaron en matraces con 50 mL de medio líquido de extracto de malta (EM: 20 g de extracto de malta, 1,000 mL de agua destilada estéril), a una temperatura de 25 °C y oscuridad por 25 días. La extracción del ADN se hizo de acuerdo al método AP (Sambrook y Russell, 2001). Su calidad se evaluó por electroforesis en gel de agarosa al 1%. La amplificación de las regiones internas ITS1 e ITS2 de los genes ribosomales (rADN), localizados entre la subunidad pequeña 18S.5.8S y la subunidad larga 5.8S-28S se realizó mediante la técnica de PCR con los iniciadores universales ITS1-F (5´-CTTGGTCATTTAGAGGGAAGTAA-3´) e ITS4 (5´-TCC TCCGCTTATTGATATGC-3´).
La mezcla de reacción de PCR estuvo compuesta de: 12.5 µL agua libre de nucleasas, 2.5 µL solución amortiguadora Buffer 1X, 2.5 µL de MgCl
2 a 2.5 mM, 0.5 µL de dNTPs a 10 mM,
0.5 µL de cada oligonucleótido, 2 U de Taq-DNA polimerasa (Invitrogen) y 2 µL de DNA (10 ng), con un volumen final de 25 µL. Se utilizó un termociclador Applied BiosystemsMR (Mod. Termal Cycler 2720) con el siguiente programa: desnaturalización
Morpholgic identification
From the 10 pure cultures (monoconidial or monoascoporic) in 2% MAE at 25 ± 2 °C and incubated for 40 days, at a genus level, fungi were determined by following the taxonomic keys of Barnett and Hunter (1972) and Hanlin (2001), and for species those of Upadhyay (1981) and Jacobs and Wingfield (2001). Also, the way they grew, the color of the colony, the type of mycelium, reproductive structure formation, type and form of conidia and ascospores were observed, in which 200 ascospores and conidia and 30 peritecia or synnemas were measured in microns. The follow-up of the development of conidia and type of ascospores was made by electron microscopy; for that ending, fungal growth discs of 5 mm in diameter were taken. Samples were fixed in 3% glutaraldehyde with 0.1M Sorensen phoshate buffer, pH 7.2 or 24 h. Afterwards, they were rinsed thrice with the Sorensen fosfate buffer for one minute and were dehydrated with a series of ethanol solutions (30, 40, 50, 60, 70, 80, 90 and 100%; the three last were used twice) for 15 min in every change.. The discs were placed into a Samdri-780A® (TOUSIMIS Research Corporation, Rockville, USA) kiln at the critical point of CO
2 at 31.1 °C and 1073 psi; right
away, the discs were fixed on a 9 mm diameter brass sampler with a copper conductive-adhesive tape and were covered with gold during 4 min in a JFC-1100® (JEOL LTD, Tokio, Japan ioniziner). Samples were reviewed with a JSM-35C® (JEOL LTD, Tokio, Japan) electron microscope operated at 15 Kv in the Electronic Microscopy laboratory of the Graduate Studies College.
Molecular Description
The ten pure fungi isolations used to extract DNA were cultivated in 50 mL flacks of liquid medium of malt extract (ME: 20 g of malt extract + 1,000 mL of sterile distilled water) at 25 oC in darkness for 25 days. DNA extraction was made according to the AP method (Sambrook and Russell, 2001). Its quality was assessed by electrophoresis in agrose gel at 1%. The amplification of the ITS1 and ITS2 internal regions of the rhibozomal (rDNA) genes, located between the 18S.5.8S small subunit and the 5.8S-28S long subunit was made by the PCR technique with ITS1-F (5´-CTTGGTCATTTAGAGGGAAGTAA-3´) and ITS4 (5´-TCC TCCGCTTATTGATATGC-3´) universal initiators.
The reaction mixture of PCR was made up by 12.5 µL water free of nucleases, 2.5 µL Buffer solution of 1X, 2.5 µL de MgCl
2 at 2.5 mM, 0.5 µL of dNTPs at 10 mM, 0.5 µL of each
oligonucleotide, 2 U of Taq-DNA polymerase (Invitrogen) and 2 µL of DNA (10 ng), with a final volume of 25 µL. An Applied BiosystemsTM (Thermal Cycler 2720 model) thermal cycler was used with the following program: initial desnaturalization at 95 °C for 3 min; 35 desnaturalization cycles of 94 °C for 59 sec; alignment at 55 °C for 59 sec and extension at 72 °C for 1 min and a final extension of 72 °C for 8 min (Zhou et al., 2004).

Pérez et al., Hongos ophiostomastoides de...
39
inicial a 95 °C por 3 min; 35 ciclos de desnaturalización a 94 °C por 59 seg; alineamiento a 55 °C por 59 seg y extensión a 72 °C por 1 min, y una extensión final de 72 °C por 8 min. (Zhou et al., 2004).
Los productos amplificados se analizaron por electroforesis en gel de agarosa al 1.0%, se tiñeron con bromuro de etidio y las bandas se visualizaron en un fotodocumentador (Bio-Imagen Systems Mini Bis Pro). Los productos de la PCR fueron tomados directamente del gel y purificados con Wizard SV (Promega, USA).
Estos últimos fueron secuenciados en ambas direcciones (5’ a 3’ y 3’ a 5’) con los mismos iniciadores ITS, en el Instituto de Fisiología Celular de la Universidad Nacional Autónoma de México con un secuenciador Genetic Analizer 3100 (Applied Biosystem Corp). Las secuencias se compararon por alineamiento con la información disponible en la base de datos del GenBank (NCBI: National Center for Biotechnology Information), mediante el método BLAST (http://www.ncbi.nlm.nih.gov/BLAST/).
RESULTADOS Y DISCUSIÓN
Aislamientos e identificación
Se obtuvieron 10 aislamientos de galerías del insecto descortezador Dendroctonus adjunctus de Pinus hartwegii. Se identificaron cuatro hongos a nivel especie: Leptographium guttulatum M.J. Wingf. & K. Jacobs, Ophiostoma angusticollis (E.F. Wright & H.D. Griffin) M. Villarreal, O. nigrocarpum (R.W. Davidson) de Hoog y O. olivaceapini (R.W. Davidson) Seifert & G. Okada y una del género Pesotum J. L. Crane & Schokn.
Leptographium guttulatum tuvo un crecimiento de 80 mm a los 10 - 12 días en EMA (2%), a una temperatura de 26 ± 1 °C, en condiciones de laboratorio; coloración de color café claro a café oliváceo y al reverso de la caja tonalidad café oscuro. El micelio del hongo inmerso en el medio de cultivo fue de color café claro a café oliváceo y micelio aéreo hialino, cuando joven (Figura 1A). Conidióforos simples de color café oliváceo de (210-) 500.33 (-815.60) µm de longitud y sin rizoides emergiendo del medio de cultivo. Estípite café oliváceo, cilíndrico con siete septas de (115.20-) 270.40 (-650.60) µm de longitud y 5.5 a 11 µm de ancho. Células apical y basal no hinchadas. Aparato conidiogénico de (62.55-) 150 (-195.80) µm de longitud con ramificación tipo B con 2 a 4 ramas (Figura 1B). Células conidiogénicas de proliferación percurrente, cilíndricas de 2 a 3, con el ápice ligeramente angosto y redondeado. Conidias hialinas, aseptadas, oblongas a ovoides con gútulas prominentes de (5.59-) 6.74 (-11.62) × 2.0-3.0 µm, de apariencia simpodial en la base de las células conidiogénicas (Figura 1C). Las conidias están contenidas en una gota mucilaginosa y transparente que cubre todo el aparato
The amplified products were analyzed by agarose gel electrophoresis at 1%; they were stained with ethidium bromide and bands were visualized in a photo-documenter (Bio-Imagen Systems Mini Bis Pro). The PCR products were directly taken from gel and purified with SV Wizard (Promega).
The latter were sequenced in both directions (5’ to 3’ and 3’ to 5’) with the same ITS initiators at the Cell Physiology Institute of the National University of Mexico (UNAM) with a Genetic Analizer 3100 (Applied Biosystem Corp) sequencer. Sequences were compared by alignment with available information in the data base of GenBank (NCBI: National Center for Biotechnology Information) by the BLAST method (http://www.ncbi.nlm.nih.gov/BLAST/).
RESULTS AND DISCUSSION
Isolation and identification
Ten isolations of insect galleries of Dendroctonus adjunctus bark-beetle of Pinus hartwegii were obtained. At the species level four fungi were identified: Leptographium guttulatum M.J. Wingf. & K. Jacobs, Ophiostoma angusticollis (E. F. Wright & H. D. Griffin) M. Villarreal, O. nigrocarpum (R. W. Davidson) de Hoog and O. olivaceapini (R. W. Davidson) Seifert & G. Okada and one of the Pesotum J. L. Crane & Schokn genus.
Leptographium guttulatum grew 80 mm after 10 to 12 days in 2% MAE at 26 ± 1 °C, in laboratory conditions; from light brown to olive brown and at the other side of the box, dark brown. The mycelium of the fungi immersed in the culture medium, light brown to olive- brown and hyaline aerial mycelium, when young (Figure 1A). Olive-brown simple conidiophores (210-) 500.33 (-815.60) µm long and without rhizoids emerging from the culture medium. Olive- brown stipe, cylinder shaped, with seven septa (115.20-) 270.40 (-650.60) µm long and 5.5 to 11 µm wide. Non-swollen apical and basal cells. Conidiogenic apparatus of (62.55-) 150 (-195.80) µm long with B type ramification of 2 to 4 branches (Figure 1B) of percurrent proliferation, cylinder shaped from 2 to 3, with the apex slightly narrow and rounded. Hyaline conidia, asepted, oblong to ovoid with (5.59-) 6.74 (-11.62) × 2.0-3.0 µm prominent guttules, of sympodial aspect at the base of the conidiogenic cells (Figure 1C). Conidias are contained in a mucilaginous and transparent drop that covers all the conidiogenic apparatus, in young isolations of 10 to 20 days and in isolations elder than 30 days, the mucilaginous drops become cream color.
These morphologic characteristics agree with the descriptions of Jacobs et al. (2001) and Jacobs and Wingfield (2001). Barras and Perry (1971) isolated Leptographium sp. (Lagerb and Melin Svenska) from the micangio of Dendroctonus adjunctus. From their morphology, Six and Paine (1996) determined mtDNA sequencies and RFLP of the micangial fungus Leptographium

Rev. Mex. Cien. For. Vol. 2 Núm. 8
40
conidiogénico, en aislamientos jóvenes de 10 a 20 días y en aislamientos mayores de 30 días las gotas mucilaginosas se tornan de color crema.
Estas características morfológicas correspondieron a las descripciones de Jacobs et al. (2001) y Jacobs y Wingfield (2001). Barras y Perry (1971) aislaron del micangio de Dendroctonus adjunctus a Leptographium sp. (Lagerb y Melin Svenska). Six y Paine (1996) identificaron por características morfológicas, secuencias de ADNmt y RFLP al hongo micangial Leptographium pyrinum R.W. Davidson asociado a D. adjunctus. L. pyrinum puede distinguirse de otras especies de Leptographium por la conidia ovoide y la presencia de material granular sobre las hifas (Jacobs y Wingfield, 2001).
Ophiostoma angusticollis registró un crecimiento radial de 26 mm de diámetro a los 15 días, a una temperatura de 26 ± 1 °C, en medio de cultivo EMA (2%); coloración blanca cremosa y al reverso de la caja de Petri, blanca (Figura 1D). Los peritecios crecen superficiales o ligeramente inmersos en el medio de cultivo a los 15 o 20 días. Su base es globosa, oscura, de (84.60-) 96.30 (-115.20) μm de diámetro. El cuello del peritecio es café oscuro a negro, de 225 a 507 μm de largo y (12.00-) 15.60 (-21.30) μm de ancho de la base. El ápice del cuello romo, más claro, de (1.85-) 3.24 (-6.30) μm de ancho e hifas ostiolares ausentes (Figura 1E). Las ascas son evanescentes. Ascosporas hialinas, unicelulares, reniformes de (2.00-) 3.50 (-4.70) × (1.0-) 1.6 (-2.0) μm (Figura 1F). La masa de ascosporas está contenida en un mucílago blanco cremoso, en el ápice del cuello del peritecio (Figura 1G). La descripción anterior concuerda con las de Olchowecki y Reid (1974), Upadhyay (1981) y Aghayeva et al. (2004).
En el micelio de O. angusticollis se observó un tipo de célula conidiogénica de apariencia denticulada, similar a su anamorfo Sporothrix y con conidios hialinos, unicelulares, ovoides, de (2.0-) 3.5 (-7.0) × (1.0-) 1.2 (-2.0) μm, con desarrollo simpodial (Upadhyay, 1981). Morfológicamente Ophiostoma angusticollis (E.F. Wright & H.D. Griffin) M. Villarreal M. Villarreal, Arenal, V. Rubio & De Troya son muy parecidos. Sin embargo, el anamorfo de O. sejunctum es Hyalorhinocladiella H.P. Upadhyay & W.B. Kendrick (Villarreal et al., 2005); además, O. angusticollis contiene la masa de ascosporas en un mucílago, a diferencia de O. sejunctum (Olchowecki y Reid, 1974). Se ha aislado O. angusticollis y otros Ceratocystis sensu lato de la cámara pupal de un cerambícido asociado con Bursaphelenchus xylophilus Steiner et Buhrer; así como, del nemátodo de la madera del pino en Pinus resinosa Aiton y P. banksiana Lamb. (Wingfield, 1987).
Ophiostoma nigrocarpum presentó un crecimiento radial en medio de cultivo EMA (2%) y alcanzó un diámetro de 30 mm, a los 15 días. La coloración de la colonia fue de blanca a gris oscuro, en función la edad del cultivo
pyrinum R.W. Davidson associated to D. adjunctus. L. pyrinum can be distinguished from other Leptographium species for the ovoid conidia and granular matter over hyphae (Jacobs and Wingfield, 2001).
Ophiostoma angusticollis had a radial growth of 26 mm of diameter after 15 days at 26 ± 1 °C in MAE at 2%; creamy white color and on the other side of Petri boxes, white color (Figure 1D). Perithecia have a superficial growth or lightly immerse in the culture medium after 15 to 20 days. Its base is globose, dark, (84.60-) 96.30 (-115.20) μm in diameter. The neck of the perithecium is dark brown to black, 225 to 507 μm long and (12.00-) 15.60 (-21.30) μm wide at the base. The apex of the neck is blunt, lighter in color, (1.85-) 3.24 (-6.30) μm wide and absent ostiolary hyphae (Figure 1E). Ascae are evanescent. Hyaline ascospores, unicelular, reniform, (2.00-) 3.50 (-4.70) × (1.0-) 1.6 (-2.0) μm (Figure 1F). The ascospore mass is contained in a creamy white mucilage in the apex of the neck of the perithecium (Figure 1G). The previous description agrees with those of Olchowecki and Reid (1974), Upadhyay (1981) and Aghayeva et al. (2004).
In the mycelium of O. angusticollis it was observed a type of conidiogenic cell of denticulate appearance, similar to its anamorphic Sporothrix and with hyaline conidia, one-cell, ovoid, (2.0-) 3.5 (-7.0) × (1.0-) 1.2 (-2.0) μm with sympodial development (Upadhyay, 1981). Morphologically, Ophiostoma angusticollis (E. F. Wright & H. D. Griffin) M. Villarreal M. Villarreal, Arenal, V. Rubio & De Troya are very similar. However, the anamorphic of O. sejunctum is Hyalorhinocladiella H.P. Upadhyay & W.B. Kendrick (Villarreal et al., 2005); also, O. angusticollis has the ascospore mass in a mucilage, in contrast to O. sejunctum (Olchowecki and Reid, 1974). O. angusticollis and other Ceratocystis sensu lato have been isolated from the pupal chamber of a cerambycidae associated with Bursaphelenchus xylophilus Steiner et Buhrer, as well as from the nematode of the wood of Pinus resinosa Aiton and P. banksiana Lamb. (Wingfield, 1987).
Ophiostoma nigrocarpum showed a radial growth in 2% MAE and reached a 30 mm diameter after 15 days. The colony was white to dark grey, depending on the age of the culture (Figure 1H). One-cell conidia, ovoid, ellipsoidal or cylindric, (2.0-) 3.5 (-7.0) × (1.0-) 1.2 (-2.0) μm and in a white mycelium, similar to the Sporothrix genus (Figure 1I). Perithecia grew on the surface or lightly immersed in the culture and develop after 10 to 15 days (Figure 1). Globose dark base, (188-) 107.82 (-84.6) μm diameter (Figure 1K). Perithecium neck dark brown to black, 180 to 210 μm long and (18-) 22 (-36) μm wide at the base; apex is lighter, (15-) 13 (-21) μm wide and ostiolary hyphae, asepted, light brown, (1.5-) 3.2 (-3.8) μm wide. Ascae are evanescent. Hyaline ascosporeas, unicelular, moon-shape, (2.90-) 3.50 (-4.90) × (1.0-) 1.6 (-2.0) μm (Figure 1K and L). The ascospore mass is contained in a creamy

Pérez et al., Hongos ophiostomastoides de...
41
(Figura 1H). Conidios unicelulares, ovoides, elipsoides a cilíndricos de (2.0-) 3.5 (-7.0) × (1.0-) 1.2 (-2.0) μm y contenidos en micelio blanco, similar al del género Sporothrix (Figura 1I). Los peritecios crecen superficiales o ligeramente inmersos en el medio de cultivo, desarrollándose a los 10 - 15 días (Figura 1J). Su base es globosa, oscura, de (188-) 107.82 (-84.6) μm de diámetro (Figura 1K). Cuello del peritecio café oscuro a negro, de 180 a 210 μm de largo y (18-) 22 (-36) μm de ancho de la base; el ápice es más claro, de (15-) 13 (-21) μm de ancho e hifas ostiolares, aceptadas, de color café claro de (1.5-) 3.2 (-3.8) μm de ancho. Ascas de tipo evanescente. Ascosporas hialinas, unicelulares, de forma lunada, de (2.90-) 3.50 (-4.90) × (1.0-) 1.6 (-2.0) μm (Figura 1K y L). La masa de ascosporas está contenida en un mucilago blanco cremoso en el ápice del cuello del peritecio. Esta descripción morfológica coincide con las de Upadhyay (1981) y Aghayeva et al. (2004)
En Ophiostoma olivaceapinii se registró un crecimiento radial en medio EMA (2%), con un diámetro de 45 mm, a los 15 días y a una temperatura de 26 ± 1 °C. La coloración de la colonia fue café claro, café oscuro en cultivos jóvenes y café grisáceo a los 30 días. El micelio inmerso de color café (Figura 2A). Conidióforos sinematosos café a café oscuro, de 480 a 560 μm de largo incluyendo el aparato conidiogénico. Sinemas solitarios o agrupados (2 a 6), de color café a café oscuro, a los 10 o 12 días (Figura 2B). Células conidiogénicas aparentemente con proliferación percurrente y anelaciones de apariencia simpodial, cilíndrica y hialina. Conidios hialinos, unicelulares, elipsoides a ovoides, de (7.64-) 4.82 (-1.97) × (5.33-) 2.65 (-1.37) μm. La masa de conidios contenida en un mucílago hialino a grisáceo en el ápice del sinema, dependiendo de la edad del cultivo. Los peritecios crecen superficiales o ligeramente inmersos en el medio de cultivo y se desarrollan a los 25 días, solitarios o agrupados (Figura 2C); base globosa, oscuros, de 190 a 200 μm de diámetro. El cuello del peritecio café oscuro a negro de 250 a 618 μm de largo sin incluir hifas ostiolares, de (20-) 26 (-38) μm de ancho de la base, de (13-) 16 (-21) μm de ancho del ápice y ápice del cuello ligeramente curvo. Hifas hialinas, de 20 a 25 μm de largo y de (1-) 1.5 (-2.0) μm de ancho. Las ascas son de tipo evanescentes. Ascosporas hialinas, reniformes (Figura 2D). Estas características concuerdan con las descripciones de Barnett y Hunter (1972); Upadhyay (1981) y Mouton et al. (1993).
Pesotum sp. (sinónimo de Graphium corda creció 80 mm en 15 días en EMA (2%), a una temperatura de 26 ± 1 °C; su coloración fue café, café olivo a gris a los 70 días, al reverso de la caja es café oscuro (Figura 2E). Micelio inmerso café y micelio aéreo hialino. Sinemas café oscuro a negro oscuro de (450.16-) 236.99 (-150.40) × (37.60-) 22.65 (-9.40) μm (Figura 1F). Célula conidiogénica percurrente, anélida, cilíndrica y hialina. Conidia holoblástica, hialina, unicelular, elipsoide a ovoide de (7.64-) 4.82 (-1.97) × (6.33-) 2.65 (-1.37) μm y la masa de conidias se acumula en el ápice del sinema (Figura 1G). La descripción anterior coincide con las descripciones de Barnett y Hunter (1972) y Upadhyay (1981).
white mucilage in the apex of the neck of the perithecium. This morphologic description is consistent with that of Upadhyay (1981) and Aghayeva et al. (2004).
Ophiostoma olivaceapinii had a radial growth on 2% MAE, media with a diameter of 45 mm, after 15 days at 26 ± 1 °C. The colony was light brown, dark brown in young cultures and greyish brown after 30 days. Immersed mycelium, brown (Figure 2A). Synnematous conidiaphore brown to dark brown, of 480 to 560 μm long including the conidiogenic apparatus. Single and clumped synnemas (2 to 6), brown to dark brown, after 10 to 12 days (Figure 2B). Conidiogenic cells apparently with percurrent proliferation and annellides of sympodial, barrel-shaped and hyaline look. (7.64-) 4.82 (-1.97) × (5.33-) 2.65 (-1.37) μm hyaline, one-celled, ellipsoidal or ovoid conidia. Conidia mass in a hyaline or greyish mucilage at the apex of the synnema, according to the age of the culture. Perithecia grow at the surface or lightly immersed into the culture medium and develop after 25 days, single or clumped (Figure 2C); globose base, dark, 190 a 200 μm of diameter. Neck of the perithecium, dark brown to black, 250 to 618 μm long, without ostiolary hyphae (20-) 26 (-38) μm wide at the base, (13-) 16 (-21) μm wide at the apex and apex of the neck lightly curved. Hyaline hyphae, 20 to 25 μm long and (1-) 1.5 (-2.0) μm wide. Ascae are evanescent. Hyaline ascospores, reniform (Figura 2D). These descriptions agree with those of Barnett and Hunter (1972), Upadhyay (1981) and Mouton et al. (1993).
Pesotum sp. (Graphium corda synonym ) grew 80 mm after 15 days in 2% MAE, at 26 ± 1 °C; brown, olive-brown to grey after 70 days, and at the back of the box, dark brown (Figure 2E). Immersed mycelium, brown and aerial mycelium, hyaline. Dark brown to dark black synnema, (450.16-) 236.99 (-150.40) × (37.60-) 22.65 (-9.40) μm (Figure 1F). Percurrent conidiogenic cell, annelid, cylindric and hyaline. Holoblastic conidia, hyaline, unicelular, ellipsoid to ovoid, (7.64-) 4.82 (-1.97) × (6.33-) 2.65 (-1.37) μm and the condidia mass clumped at the apex of the synnema (Figure 1G). The former description agrees with those of Barnett and Hunter (1972) and Upadhyay (1981).
Molecular description
The PCR product of the ITS1 e ITS2 internal regions of the (rADN) rhibozomal genes was 700 pb (pair bases). The ZOQ2 y ZOQ8 sequences of Leptographium guttulatum; ZOQ5, ZOQ15, ZOQ17 of Ophiostoma olivaceapini; ZOQ9 and ZOQ16 of O. angusticollis aligned with the sequences of the same species at the NCBI Gene Bank (Table 1), with a 98 to 100% homology, except for the Pesotum (ZOQ6) genus that aligned with Ophiostoma sp. (Access number: AY649781), with a 98% homology, considering its teleomorph (Harrington et al., 2001).

Rev. Mex. Cien. For. Vol. 2 Núm. 8
42
Figura 1. Características morfológicas de los hongos en cultivo y microscópicas de Leptographium guttulatum, Ophiostoma angusticollis y O. nigrocarpum en EMA al 2%. A) Colonia de L. guttulatum de color café claro a café oliváceo a los 12 días. B) Conidióforos solitarios. C) Conidias unicelulares. D) Colonia blanco cremoso de O. angusticollis a los 15 días. E) Peritecio. F) Ascosporas reniformes. G) Masas de ascosporas en una matriz gelatinosa. H) Colonia blanco cremoso con peritecios de O. nigrocarpum. I) Conidias elipsoides a cilíndricas. J y K) Peritecios y corte de la base. L) Ascosporas reniformes.
Figure 1. Morphologic and microscopic characteristics of the fungi cultures of Leptographium guttulatum, Ophiostoma angusticollis and O. nigrocarpum in MAE at 2%. A) L. guttulatum colony, from light brown to olive brown after 12 days. B) Lonely conidiophores. C) One-cell conidia. D) Creamy white colony of O. angusticollis after 15 days. E) Perithecium. F) Reniform ascospores. G) Ascospore masses in a gelly matrix. H) Creamy white colony with perithecia of O. nigrocarpum. I) Ellypsoid to cylindric conidia. J and K) Perithecia and cutting at the base. L) Reniform ascospores.
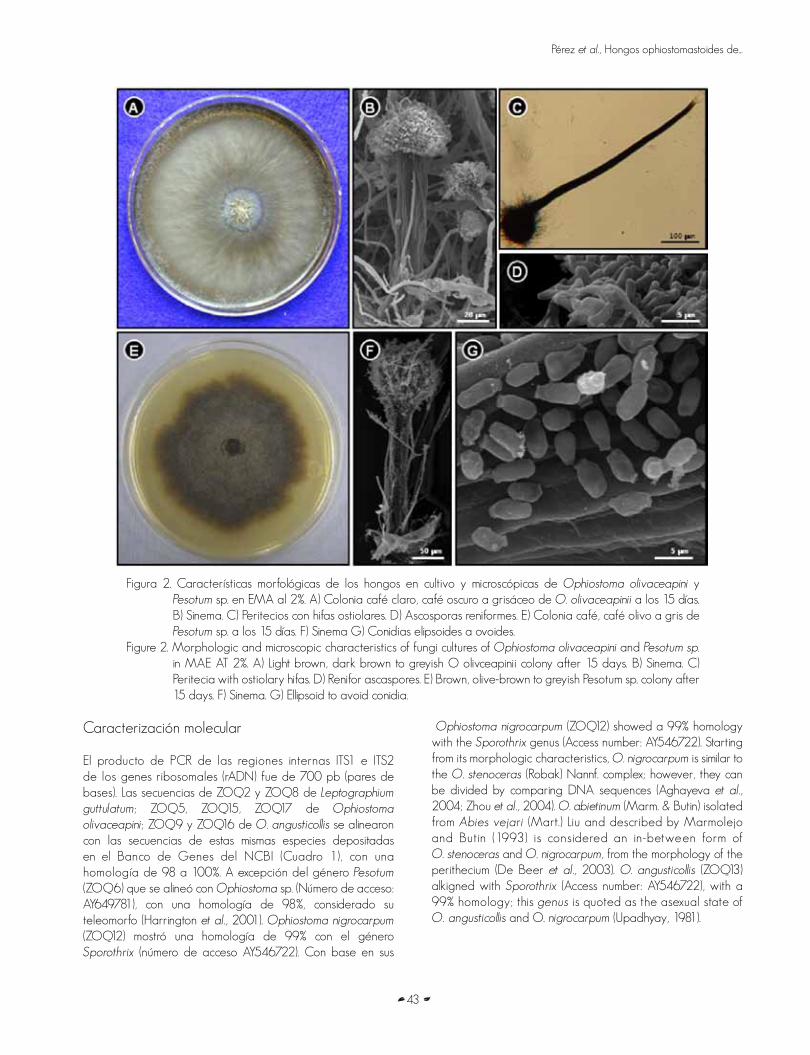
Pérez et al., Hongos ophiostomastoides de...
43
Figura 2. Características morfológicas de los hongos en cultivo y microscópicas de Ophiostoma olivaceapini y Pesotum sp. en EMA al 2%. A) Colonia café claro, café oscuro a grisáceo de O. olivaceapinii a los 15 días. B) Sinema. C) Peritecios con hifas ostiolares. D) Ascosporas reniformes. E) Colonia café, café olivo a gris de Pesotum sp. a los 15 días. F) Sinema G) Conidias elipsoides a ovoides.
Figure 2. Morphologic and microscopic characteristics of fungi cultures of Ophiostoma olivaceapini and Pesotum sp. in MAE AT 2%. A) Light brown, dark brown to greyish O olivceapinii colony after 15 days. B) Sinema. C) Peritecia with ostiolary hifas. D) Renifor ascaspores. E) Brown, olive-brown to greyish Pesotum sp. colony after 15 days. F) Sinema. G) Ellipsoid to avoid conidia.
Caracterización molecular
El producto de PCR de las regiones internas ITS1 e ITS2 de los genes ribosomales (rADN) fue de 700 pb (pares de bases). Las secuencias de ZOQ2 y ZOQ8 de Leptographium guttulatum; ZOQ5, ZOQ15, ZOQ17 de Ophiostoma olivaceapini; ZOQ9 y ZOQ16 de O. angusticollis se alinearon con las secuencias de estas mismas especies depositadas en el Banco de Genes del NCBI (Cuadro 1), con una homología de 98 a 100%. A excepción del género Pesotum (ZOQ6) que se alineó con Ophiostoma sp. (Número de acceso: AY649781), con una homología de 98%, considerado su teleomorfo (Harrington et al., 2001). Ophiostoma nigrocarpum (ZOQ12) mostró una homología de 99% con el género Sporothrix (número de acceso AY546722). Con base en sus
Ophiostoma nigrocarpum (ZOQ12) showed a 99% homology with the Sporothrix genus (Access number: AY546722). Starting from its morphologic characteristics, O. nigrocarpum is similar to the O. stenoceras (Robak) Nannf. complex; however, they can be divided by comparing DNA sequences (Aghayeva et al., 2004; Zhou et al., 2004). O. abietinum (Marm. & Butin) isolated from Abies vejari (Mart.) Liu and described by Marmolejo and Butin (1993) is considered an in-between form of O. stenoceras and O. nigrocarpum, from the morphology of the perithecium (De Beer et al., 2003). O. angusticollis (ZOQ13) alkigned with Sporothrix (Access number: AY546722), with a 99% homology; this genus is quoted as the asexual state of O. angusticollis and O. nigrocarpum (Upadhyay, 1981).
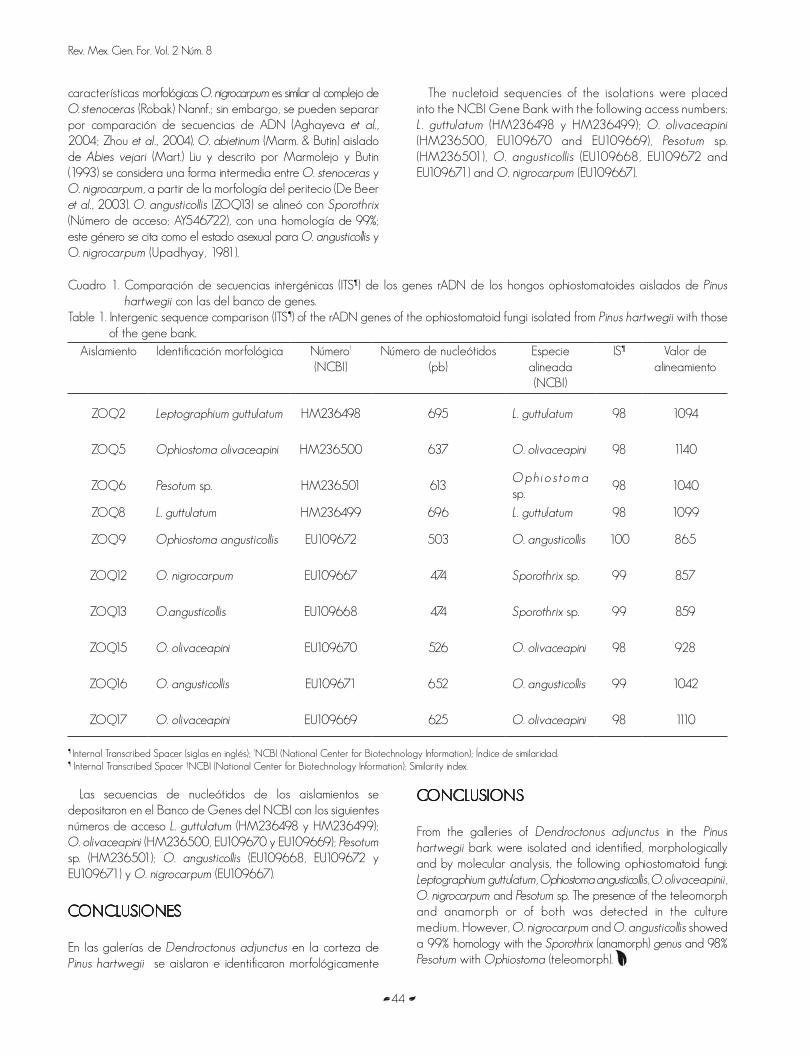
Rev. Mex. Cien. For. Vol. 2 Núm. 8
44
The nucletoid sequencies of the isolations were placed into the NCBI Gene Bank with the following access numbers: L. guttulatum (HM236498 y HM236499); O. olivaceapini (HM236500, EU109670 and EU109669), Pesotum sp. (HM236501), O. angusticollis (EU109668, EU109672 and EU109671) and O. nigrocarpum (EU109667).
características morfológicas O. nigrocarpum es similar al complejo de O. stenoceras (Robak) Nannf.; sin embargo, se pueden separar por comparación de secuencias de ADN (Aghayeva et al., 2004; Zhou et al., 2004). O. abietinum (Marm. & Butin) aislado de Abies vejari (Mart.) Liu y descrito por Marmolejo y Butin (1993) se considera una forma intermedia entre O. stenoceras y O. nigrocarpum, a partir de la morfología del peritecio (De Beer et al., 2003). O. angusticollis (ZOQ13) se alineó con Sporothrix (Número de acceso: AY546722), con una homología de 99%; este género se cita como el estado asexual para O. angusticollis y O. nigrocarpum (Upadhyay, 1981).
Cuadro 1. Comparación de secuencias intergénicas (ITS¶) de los genes rADN de los hongos ophiostomatoides aislados de Pinus hartwegii con las del banco de genes.
Table 1. Intergenic sequence comparison (ITS¶) of the rADN genes of the ophiostomatoid fungi isolated from Pinus hartwegii with those of the gene bank.
Aislamiento Identificación morfológica Número†
(NCBI)Número de nucleótidos
(pb)Especie alineada(NCBI)
IS¶ Valor de alineamiento
ZOQ2 Leptographium guttulatum HM236498 695 L. guttulatum 98 1094
ZOQ5 Ophiostoma olivaceapini HM236500 637 O. olivaceapini 98 1140
ZOQ6 Pesotum sp. HM236501 613Ophio s toma sp.
98 1040
ZOQ8 L. guttulatum HM236499 696 L. guttulatum 98 1099
ZOQ9 Ophiostoma angusticollis EU109672 503 O. angusticollis 100 865
ZOQ12 O. nigrocarpum EU109667 474 Sporothrix sp. 99 857
ZOQ13 O.angusticollis EU109668 474 Sporothrix sp. 99 859
ZOQ15 O. olivaceapini EU109670 526 O. olivaceapini 98 928
ZOQ16 O. angusticollis EU109671 652 O. angusticollis 99 1042
ZOQ17 O. olivaceapini EU109669 625 O. olivaceapini 98 1110
¶ Internal Transcribed Spacer (siglas en inglés); †NCBI (National Center for Biotechnology Information); Índice de similaridad.¶ Internal Transcribed Spacer †NCBI (National Center for Biotechnology Information); Similarity index.
Las secuencias de nucleótidos de los aislamientos se depositaron en el Banco de Genes del NCBI con los siguientes números de acceso L. guttulatum (HM236498 y HM236499); O. olivaceapini (HM236500, EU109670 y EU109669); Pesotum sp. (HM236501); O. angusticollis (EU109668, EU109672 y EU109671) y O. nigrocarpum (EU109667).
CONCLUSIONES
En las galerías de Dendroctonus adjunctus en la corteza de Pinus hartwegii se aislaron e identificaron morfológicamente
CONCLUSIONS
From the galleries of Dendroctonus adjunctus in the Pinus hartwegii bark were isolated and identified, morphologically and by molecular analysis, the following ophiostomatoid fungi: Leptographium guttulatum, Ophiostoma angusticollis, O. olivaceapinii, O. nigrocarpum and Pesotum sp. The presence of the teleomorph and anamorph or of both was detected in the culture medium. However, O. nigrocarpum and O. angusticollis showed a 99% homology with the Sporothrix (anamorph) genus and 98% Pesotum with Ophiostoma (teleomorph).

Pérez et al., Hongos ophiostomastoides de...
45
y por análisis molecular los hongos ophiostomatoides Leptographium guttulatum, Ophiostoma angusticollis, O. olivaceapinii, O. nigrocarpum y Pesotum sp. Se registró la presencia del teleomorfo y anamorfo o ambos en el medio de cultivo. Sin embargo, O. nigrocarpum y O. angusticollis mostraron una homología con el género Sporothrix (anamorfo) en 99% y Pesotum en 98% con Ophiostoma (teleomorfo).
REFERENCIAS
Aghayeva, D. N., Z. W. De Beer and T. Kirisitis. 2004. Two new Ophiostoma species with Sporothrix anamorphs from Austri and Azerbaijan. Mycologia 96 (4): 866-828.
Ayres, M. P., R. T. Wilkens, J. J. Ruel, M. J. Lombardero, and E. Vallery. 2000. Nitrogen budgets of phloem-feeding bark beetles with and without symbiotic fungi (Coleoptera: Scolytidae). Ecology 81 (8): 2198-2210.
Barnett, H. L. and B. B. Hunter, 1972. Illustrated genera of imperfect fungi. Third edition. Burgess Publishing Company. Minnesota, MN. USA. 241 p.
Barras S., J. and T. Perry. 1971. Gland cells and fungi associated with prothoracic mycangium of Dendroctonus adjunctus (Coleoptera: Scolytidae). Annals Entomological Society of America 64 (1): 123-126.
Cibrián T., D., D. A. Rosales and S. E. G. Díaz. 2007. Enfermedades forestales en México/Forest diseases in Mexico. Universidad Autónoma Chapingo; CONAFOR-SEMARNAT, México; Forest Service USDA, EUA; NRCAN Forest Service, Canadá y Comisión Forestal de América del Norte, COFAN, FAO. Chapingo, Edo. de México., México. 587 p.
Croisé, L., F. Lieutier, H. Cochard and E. Dreyer. 2001. Effect of drought stress and high density stem inoculations with Leptographium wingfieldii on hydraulic properties of young Scots pine trees. Tree Physiology 21 (7): 427-436.
De Beer, Z. W., T. C. Harrington, H. F. Vismer, B. D. Wingfield and M. J. Wingfield. 2003. Phylogeny of the Ophiostoma stenoceras-Sporothrix schenckii complex. Mycologia 95 (3): 434-441.
Hanlin, R. T. 2001. Illustrated genera of ascomycetes. Volume II. The American Phytopathological Society, St. Paul, MN. USA. 113 p.
Harrington, T. C., D. McNew, J. Steimel, D. Hofstra and R. Farrel. 2001. Phylogeny and taxonomy of the Ophiostoma piceae complex and the Dutch elm disease fungi. Mycologia 93 (1): 111-136.
Harrington, T. C. 1988. Leptographium species, their distributions, hosts and insect vectors. In: Harrington, T. C. and F. W. Cobb. (Eds.). Leptographium root diseases on conifers. The American Phytopathological Society Press. St. Paul, MN. USA. pp: 1-39.
Hausner, G., J. Reid and G. R. Klassen. 1993. On the subdivision of Ceratocystis s.l., based on partial ribosomal DNA sequences. Canadian Journal of Botany 71 (1): 52-63.
Jacobs, K. and M. J. Wingfield. 2001. Leptographium species. Tree pathogens, insect associates, and agents of blue-stain. The American Phytopathological Society. St. Paul, MN. USA. 207 p.
Jacobs, K., M. J. Wingfield, C. Coetsee, T. Kirisitis and B. D. Wingfield. 2001. Leptographium guttulatum sp. nov., a new species from spruce and pine in Europe. Mycologia 93 (2): 380-388.
Kile, G. A. 1993. Plant diseases caused by species of Ceratocystis sensu stricto and Chalara. In: Wingfield, M. J., K. A. Seifert and J. F. Webber (Eds.). Ceratocystis and Ophiostoma: Taxonomy, ecology and pathogenicity. The American Phytopathological Society. St. Paul, MN USA. pp: 173-184.
Malloch, D. and M. Blackwell. 1993. Dispersal biology of the ophiostomatoid fungi. In: Wingfield, M. J., K. A. Seifert, J. F. Webber (Eds.). Ceratocystis and Ophiostoma: Taxonomy, ecology and pathogenicity. The American Phytopathological Society. St. Paul, MN. USA. pp: 173-184.
Marmolejo J., G. 1991. El manchado azul de la Madera en Nuevo León. Reporte científico No. 27. Facultad de Ciencias Forestales. Universidad Autónoma de Nuevo León. Linares, N. L. México. 59 p.
Marmolejo J., G. y H. Butin. 1993. Las especies de Ophiostoma y Ceratocystiopsis (Ascomycetes, Microascales) conocidas de Nuevo León, México. Reporte científico especial No. 13. Facultad de Ciencias Forestales. Universidad Autónoma de Nuevo León. Linares, N. L. México. pp: 155-170.
Mouton, M., M. J. Wingfield and P. S. Van-Wyk. 1993. Conidium development in the synnematous anamorphs of Ophiostoma. Mycotaxon 40 (6): 371-379.
Olchowecki, A. and J. Reid. 1974. Taxonomy of the genus Ceratocystis in Manitoba. Canadian Journal of Botany 52 (7): 1675-1711.
Paine, T. D., K. F. Raffa and T. C. Harrington. 1997. Interactions among scolytid bark beetles, their associated fungi, and live host conifers. Annual Review of Entomology 42: 179-206.
Pérez V., O. A., D. Alvarado R., E. Cárdenas S., A. Equihua M., D. Cibrián, T., J. G. Álvarez M., D. Mejía S. y T. C. Harrington. 2009. Ophiostoma ips asociado al insecto descortezador (Dendroctonus adjunctus) del pino de las alturas (Pinus hartwegii). Revista Mexicana de Micología 30: 9-18.
Popp, M. P., J. D. Johnson and M. S. Lesney. 1995. Characterization of the induced response of slash pine to inoculation with bark beetle vectored fungi. Tree Physiology 15 (9): 619-623.
Sambrook, J. and D. W. Russell. 2001. Molecular cloning. A laboratory manual. Third edition. 1:1.32-1.34. Cold Spring Harbour Laboratory Press. New York, NY. USA. 2344 p.
Seifert, K. A. 1993. Sapstain of commercial lumber by species of Ophiostoma and Ceratocystis. In: Wingfield, M. J., K. A. Seifert and J. F. Webber (Eds.). Ceratocystis and Ophiostoma. Taxonomy, Ecology, and Pathogenicity. The American Phytopathological Society. St. Paul, MN. USA. pp: 141-151.
Six, D. and T. D. Paine. 1996. Leptographium pyrinum is a mycangial fungus of Dendroctonus adjunctus. Mycologia 88 (5): 739-744.
Upadhyay, H. P. 1981. A monograph of Ceratocystis and Ceratocystiopsis. University of Georgia Press. Athens, GA. USA. 176 p.
Upadhyay, H. P. 1993. Classification of the ophiostomatoid fungi. In: Wingfield, M. J., K. A. Seifert and J. F. Webber (Eds). Ceratocystis and Ophiostoma: Taxonomy, ecology and pathogenicity. The American Phytopathological Society. St. Paul, MN. USA. pp: 7-13.
Villarreal, M., V. Rubio, M. T. De Troya and F. Arenal. 2005. A new Ophiostoma species isolated from Pinus pinaster in the Iberian Peninsula. Mycotaxon 92: 259-268.
Wingfield, B. D., W. S. Grant, J. F. Wolfaardt and M. J. Wingfield. 1994. Ribosomal RNA sequence phylogeny is not congruent with ascospore morphology among species in Ceratocystis sensu stricto. Molecular Biology and Evolution 11 (3): 376-383.
Wingfield, M. J. 1987. Fungi associated with the pine wood nematode, Bursaphelenchus xylophilus, and cerambycid beetles. Mycologia 79 (2): 325-328.
Wingfield, M. J. 1993. Leptographium species as anamorphs of Ophiostoma: progress in establishing acceptable generic and species concepts. In: Wingfield, M. J., K. A. Seifert and J. F. Webber (Eds). Ceratocystis and Ophiostoma: Taxonomy, ecology and pathogenicity. The American Phytopathological Society. St. Paul, MN. USA. pp: 43-51.
Wingfield, M. J. and P. S. Knox-Davies. 1980. Root disease, associated with Verticicladiella alacris, of pines in South Africa. Plant Disease 64 (6): 569-571.
Zhou, X. D., W. Z. De Beer, B. D. Wingfield and M.J. Wingfield, 2001. Ophiostomatoid fungi associated with three pine-infesting bark beetles in South Africa. Sydowia 53 (2): 290-300.
Zhou, X. D., Z. W. De Beer, D. C. Tovar, B. D. Wingfield and M. J. Wingfield. 2004 Characterisation of Ophiostoma species associated with pine bark beetles from Mexico, including O. pulvinisporum sp. nov. Mycological Research 108 (6): 690-6

Rev. Mex. Cien. For. Vol. 2 Núm. 8
46
Wangari Muta Maathai. Dominio público.

EVALUACIÓN DEL CARBONO ORGÁNICO EN SUELOS DE LOSMANGLARES DE NAYARIT
ORGANIC CARBON ASSESSMENT IN MANGROVE SOILS OF NAYARIT
Eduardo Valdés Velarde 1, Juan Ignacio Valdez Hernández2, Víctor Manuel Ordaz Chaparro3, Juan Fernando Gallardo Lancho4, Joel Pérez Nieto 1 y Ciriaco Ayala Sánchez 1
RESUMEN
Los manglares son un recurso ecológico y económico muy valioso para la reproducción de especies acuáticas y la alimentación de aves, peces, crustáceos, reptiles y mamíferos, además de un reservorio de carbono. En diez asociaciones de manglar y nueve clases de geoformas de la región de Marismas Nacionales, en el estado de Nayarit, se recolectaron un total de 254 muestras compuestas en los primeros -20 cm de suelo, en julio-agosto de 2004, en ellas se determinó el contenido de carbono orgánico (COS) en la fracción mineral, por el método del analizador. En la época de lluvias, los suelos de la clase, de geoforma Islote tuvieron el mayor contenido de COS, con 128.5 t C ha-1; mientras que a los de la Barra arcillosa les correspondió el menor con 54.6 t C ha-1; estas diferencias fueron significativas (F = 4.02, gl = 8, p = 0.0003). Respecto a la asociación de manglar, el valor más alto de COS se es t imó para la combinación Laguncular ia racemosa – Rhizophora mangle , con 106 .7 t C ha -1 y más ba jo en la de R. mangle –L. racemosa –A. germinans, con 73.7 t C ha-1; aunque no fueron significativas (F = 2.28, gl = 9, p = 0.0204). De acuerdo con los resultados obtenidos, los manglares de Marismas Nacionales que más contribuyeron al contenido de C en el suelo se encontraron en las geoformas de clase orilla y con predominio de L. racemosa y A. germinans.
Palabras clave: Asociación de manglar, Avicennia germinans L. Stearn, clase de geoforma, Laguncularia racemosa (L.) Gaertn., Marismas Nacionales, Rhizophora mangle L.
ABSTRACT
In 10 mangrove associations and nine landform classes analyzed of the Marismas Nacionales region, state of Nayarit, Mexico a total of 254 soil mixed samples on the first -20 cm were collected, in July and August of 2004, in which the soil organic carbon (SOC) content was determined through the analyzer method. During the rain season, soils in the landform Islote showed the highest SOC content with 128.5 t C ha-1 while the Barra arcillosa landform the lowest with 54.6 t C ha-1; these differences were significant (F = 4.02, gl = 8, p = 0.0003); about mangrove association, the highest SOC content was in the Laguncularia racemosa – Rhizophora mangle combination with 106.7 t C ha-1 and the lowest in the R. mangle –L. racemosa –A. germinans combination with 73.7 t C ha-1; these differences were not significant (F = 2.28, gl = 9, p = 0.0204). According with these results, the mangroves of Marismas Nacionales that had a greater contribution to the C content C in soil were in landform it borders and prevalence of L. racemosa and A. germinans.
Key words: Mangrove association, Avicennia germinans L. Stearn, Landform Class, Laguncularia racemosa (L.) Gaertn., Marismas Nacionales, Rhizophora mangle L.
Fecha de recepción: 22 de marzo de 2010.Fecha de aceptación: 23 de diciembre de 2011.
1Academia de Ecología. Departamento de Fitotecnia, Universidad Autónoma Chapingo. Correo-e: [email protected] Forestal. Colegio de Postgraduados.3Programa de Edafología. Colegio de Postgraduados.4Instituto de Recursos Naturales y Agrobiología de Salamanca/Colegio Superior de Investigaciones Científicas y Universidad de Salamanca, España.

48
Rev. Mex. Cien. For. Vol. 2 Núm. 8
INTRODUCTION
The chemical characteristics of carbon (C) give it a great versatility to form molecules for the basis components of living beings. Carbon cycle is one of the mainstays of the biochemical equilibrium of any ecosystem and, in addition to the hydrologic cycle, is responsible of the ecologic maintenance and of the atmosphere-lithosphere-biosphere interactions (Post et al., 1990; Osnaya, 2003).
The CO2content of the atmosphere increased from 270 to
290 ppm during the XIX century, from 315 ppm in 1956 from 335 ppm in 1976 and up to 379 ppm in 2005 (De la Cruz, 1986; Bravo, 2007). This tendency keeps a rate of 1 ppm per year. The carbon balance (C) is one the most important in nature, since it determines the organic matter flows (MO) and also, in a great proportion, that of other bioelements such as N, P and S. Thus, taking them as a starting point, it is possible to control the atmospheric CO
2 content (Gallardo and González,
2004). In the United Nations Intergovernmental Panel on Climate Change (IPCC) it has been underlined that the storage of C by plants (also named “carbon sequestration”) in a low cost option to reduce O
2 content.
The information about atmospheric C capture through photosynthesis and the increment of primary production in biomass is, mainly, bound to forest and agroforest ecosystems. However, little is documented about the efficiency of such process in mangroves and their forest management, which are an opportunity to store C. Mangrove soils have very low OM decomposition average numbers and its C storage potential is high, which makes them an option for such ending (Ponce-Hernández, 1999; UNFCCC, 2000; Moreno et al., 2002a).
Almost ¾ of C in ecosystems is concentrated in soil as root biomass or in stable forms of humusified organic compounds (Rosenzweig and Hillel, 2000; Gleason and Ewel, 2002; Mitra et al., 2005). In spite of the fact that their estimation is not easy, it is necessary to better understand the distribution and relative importance of the aerial and underground C deposits in terrestrial ecosystems (Acosta et al., 2001; Etchevers et al., 2002).
According to Armentano (1981), since the beginning of the 80’s humid lands were already acknowledged as an important C source and their contribution as crucial in its cycle. The sequestration of this element in the soil of marshes is regularly removed from the atmospheric circulation for long periods, even thousands of years. Thus, this scientist has suggested that the role of these ecosystems as natural deposits of C helped in the last centuries, to balance the loss of such element, as it was dispersed in the atmosphere, which had their origin in the reduction of the forest mass, as a consequence of human activities over time. Even though these formations do not cover more than 3% of the Earth, they gather great amounts of C that
INTRODUCCIÓN
Las características químicas del carbono (C) le confieren una gran versatilidad para la construcción de moléculas que forman los componentes básicos de los seres vivos. El ciclo del carbono es uno de los pilares del equilibrio biogeoquímico en cualquier ecosistema y, junto con el ciclo hidrológico, es responsable del mantenimiento ecológico y de las interacciones atmósfera-hidrosfera-litosfera-biosfera (Post et al., 1990; Osnaya, 2003).
El contenido de CO2 de la atmósfera se aumentó de 270
a 290 ppm durante el siglo XIX, de 315 ppm en 1956 a 335 ppm en 1976 y hasta 379 ppm en 2005 (De la Cruz, 1986; Bravo, 2007). Esta tendencia continúa a una tasa de 1 ppm por año. El balance de carbono (C) es uno de los más importantes en la naturaleza, puesto que determina los flujos de materia orgánica (MO) y, también en gran parte, el de otros bioelementos, como por ejemplo N, P y S. Por lo tanto a partir de ellos es posible controlar el contenido de CO
2 atmosférico
(Gallardo y González, 2004). En el Panel Intergubernamental de las Naciones Unidas sobre el Cambio Climático (IPCC, por sus siglas en inglés) se ha indicado que el almacenamiento de C por las plantas (también llamado secuestro de C) es una alternativa de bajo costo para reducir el contenido de CO
2 atmosférico.
La información acerca de la captura de C atmosférico mediante la fotosíntesis y el incremento de la productividad primaria en biomasa está, primordialmente, dirigida a ecosistemas forestales y agroforestales. Sin embargo, poco se tiene documentado sobre la eficiencia de dicho proceso en manglares y de su manejo forestal, los cuales representan una oportunidad para el almacenamiento de C. Los suelos de manglar tienen promedios de descomposición de MO bajos y su potencial de almacenamiento de C es alto, por lo que pueden ser una alternativa para tal fin (Ponce-Hernández, 1999; UNFCCC, 2000; Moreno et al., 2002a).
Casi tres cuartas partes del C en los ecosistemas se concentran en el suelo como biomasa de raíces o en formas estables de compuestos orgánicos humificados (Rosenzweig y Hillel, 2000; Gleason y Ewel, 2002; Mitra et al., 2005). Pese a que su estimación no es fácil, es necesaria para entender mejor la distribución e importancia relativa de los depósitos aéreos y subterráneos de C en los ecosistemas terrestres (Acosta et al., 2001; Etchevers et al., 2002).
Según Armentano (1981), desde principios de la década de los años 80 ya se reconocía a los humedales como una fuente importante de C y su contribución clave en el ciclo del mismo. El secuestro de este elemento en los suelos de cenagales es removido regularmente de circulación de la atmósfera por largos periodos, hasta de miles de años. Por ello, este autor ha sugerido que la función de las ciénagas como

49
Valdés et al., Evaluación del carbono orgánico en suelos de los Manglares de Nayarit
almacenes naturales de C contribuyó en los últimos siglos a balancear las pérdidas de dicho elemento, por su liberación a la atmósfera, las cuales tuvieron su origen en la reducción de la masa forestal, a consecuencia de las actividades antrópicas a lo largo de la historia. Aunque dichas formaciones no ocupan más del 3% de la extensión total del planeta, contienen grandes cantidades de C que aportarían más que otros sistemas a los intercambios del ciclo global del C (Armentano, 1981).
Los manglares son un recurso ecológico y económico muy valioso, pues ofrecen sitios para la reproducción de varias especies acuáticas y de alimento para aves, peces, crustáceos, reptiles y mamíferos; representan una fuente renovable de madera y son lugares de acumulación de C, nutrimentos, sedimentos y contaminantes; además de, que son una barrera protectora contra la erosión costera (Alongi, 2002; Moreno et al., 2002b).
Los bosques de mangles están sujetos a disturbios, directa e indirectamente. El efecto indirecto es quizás el más generalizado y se debe a la urbanización e infraestructura hotelera, así como a las granjas camaroneras. A estas causas se agregan las políticas económicas orientadas hacia la obtención de ganancias a corto plazo, la modificación de la hidrología por desviaciones de los cauces fluviales con fines agrícolas y la sustitución de áreas de manglar por obras de acuacultura (López y Ezcurra, 2002; Valdez, 2002).
A principios de los años 80, Armentano (1981) determinó que 230, 000 ha de manglares en los trópicos habían sido convertidos en estanques piscícolas. Con esa tasa de destrucción, calculó que 86’ 250,000 t de C se liberarían en los años subsiguientes, como consecuencia de la exposición de los suelos y el restablecimiento del equilibrio natural. Se estima que después de 10 a 20 años esos depósitos habrían liberado a la atmósfera de cuatro a nueve millones de toneladas de C.
La tasa de liberación de CO2 de los manglares, y de los
humedales, en general, es posible cuantificarla con precisión, si se tienen disponibles los siguientes datos:
a) El área de humedales o el volumen del sustrato orgánico alterado, drenado o excavado.
b) La densidad aparente del suelo orgánico.
c) Las concentraciones de C orgánico del material de sustrato.
d) La tasa de oxidación de MO por combustión o descomposición natural.
Adicionalmente, se requiere conocer la cantidad de cobertura vegetal removida o reducida para establecer el flujo neto de C en un sitio de manglar dado (De la Cruz, 1986).
would provide more than other systems to the interchange of the global C cycle.
Mangroves are very valuable ecological and economic resources as they offer convenient spaces for the reproduction of several aquatic species as well as to feed birds, fish, crustaceans, reptiles and mammals; they are a renewable source if wood and are deposits for C accumulation, nutriments, sediments and pollutants; also, they act as a protection wall against coast erosion (Alongi, 2002; Moreno et al., 2002b).
Mangrove forests are subject to disturb, directly or indirectly. The indirect effect is the most extended and it is a result of urbanization and hotel infrastructure, as well as shrimp farms. To these causes are added the economic policies bound to obtain short-term profits, the modification of hydrology from river bank deviations with agriculture purposes and the conversion of mangroves into aquaculture works (López and Ezcurra, 2002; Valdez, 2002).
At the beginning of the 80’s, Armentano (1981) determined that 230, 000 ha of mangroves in the tropics had been transformed into fish ponds. With that destruction rate, he made a calculation of 86’ 250,000 C t would be lost in the following years, as a consequence of the exposure of soils and the re establishment of natural balance. It is presumed that after 10 to 20 years, these deposits would have dispersed to the atmosphere from 4 to 9 million tons of C ((Armentano, 1981).
It is possible to quantify the CO2 liberation rate of mangroves,
and of humid lands, in general, with precision, if the following data are available:
a) The humid land area or the altered, drained or excavated volume of organic substrate.
b) The apparent density of organic soil.
c) The organic C concentrations of the substrate matter.
d) The OM oxidation rate for combustion or natural decomposition.
Additionally, it is necessary to know the amount of vegetation cover that has been removed or diminished to establish the net C flow in a specific mangrove site (De la Cruz, 1986).
The aim of the actual study was to determine the organic C content in the first -20 cm of soil in the mangroves of Marismas Nacionales, Nayarit State, by mangrove association and geoform class.

50
Rev. Mex. Cien. For. Vol. 2 Núm. 8
El objetivo del presente estudio fue determinar el contenido de C orgánico en los primeros -20 cm del suelo en los manglares de Marismas Nacionales, Nayarit, por asociación de manglar y clase de geoforma.
MATERIALES Y MÉTODOS
Área de estudio
Se localiza en la planicie costera del Pacífico, hacia la parte NO del estado de Nayarit, entre los 21° 56’ y 22° 15’ N, y los 105° 19’ y 105° 39’ O y ocupa gran parte de la región conocida como Marismas Nacionales (Figura 1). El área de estudio muestreada (23,596 ha) forma parte de los municipios de Rosamorada, Santiago Ixcuintla, Tecuala y Tuxpan; el tipo de clima predominante es Aw
1(w)(i’): cálido subhúmedo con
temperatura media anual mayor a 22 °C y precipitación media anual de 1,000 y 1,500 mm (García, 2004).
La región de Marismas Nacionales posee suelos poco evolucionados, de aporte coluvial-marino no consolidado, con erosionabilidad moderada, salinización o sodificación (IUSS Working Group WRB, 2006). Bojórquez et al. (2008) identificaron en esta región como principales suelos a los Cambisoles, Fluvisoles, Feozems, Gleysoles, Regosoles, Arenosoles y Solonchaks y para la llanura de inundación mareal, en las barras paralelas, playas y dunas costeras son los Solonchaks, Regosoles y Arenosoles. El IUSS Working Group WRB (2006) confirmó que los suelos que se distribuyen más ampliamente en la zona Cambisol crómico (72%) y Feozem háplico (28%), con más del 15% de saturación de sodio en alguna porción situada a menos de -125 cm de profundidad y una clase textural fina en los -30 cm superficiales (INEGI, 2002).
El tipo de vegetación dominante corresponde al manglar, que está conformado por las especies: Avicennia germinans L. Stearn, conocido como “puyeque”, árbol de hasta 25 m de altura cuya madera se aprovecha localmente en la construcción de casas (soleras, colgados); Laguncularia racemosa (L.) Gaertn. f. o “mangle”, que supera los 30 m de altura, su madera se utiliza con fines comerciales en la fabricación de galeras para el secado de tabaco y Rhizophora mangle L. o “candelón”, que alcanza 25 m de altura y su madera se usa en la región para construir casas (soleras, cadenas) (Valdez, 2002; Valdez, 2004).
Obtención y preparación de muestras
De julio a agosto de 2004 se establecieron 88 unidades de muestreo (UM) de 300 m2 (30 × 10 m), cada una de ellas con tres subunidades de muestreo (SUM) de 100 m2 (10 × 10 m) (Valdez, 2002). En el centro de las SUM se sacó una muestra de suelo, a una profundidad de 0 a 20 cm, para un total de 264 (Acosta et al., 2001).
MATERIALS AND METHODS
Study area
It is found in the coastal plains of the Pacific Ocean, to the northwest of Nayarit State, between 21° 56’ and 22° 15’ N, and 105° 19’ and 105° 39’ W and covers great part of the region know as Marismas Nacionales (Figure 1). The study area that was sampled (23, 596 ha) belongs to Rosamorada, Santiago Ixcuintla, Tecuala and Tuxpan municipalities; the kind of climate that prevails is Aw
1(w)(i’): warm sub-humid with
annual average temperature over 22 °C and annual average precipitation of 1,000 and 1,500 mm (García, 2004).
Marismas Nacionales region has a few poorly evolved soils, of semi- consolidated coluvial-marine supplywith medium erodability, salinisation or sodification (IUSS Working Group WRB, 2006). Bojórquez et al. (2008) found Cambisols, Fluvisols, Feozems, Gleysols, Regosols, Arenosols and Solonchaks as the main types of soils in this region and for the marine floods moorland, parallel bars, beaches and coastal dunes, Solonchanks, Regosols and Arenosols. The IUSS Working Group WRB (2006) confirmed that the soils that are distributed at great extent are chromic Cambisol (72%) and haplic Feozem (28%), with more than 15% of sodium saturation in some fraction located at -125 cm deep and a fine texture class in the -30 cm (INEGI, 2002).
The type of dominant vegetation is mangrove, that is the made up by Avicennia germinans L. Stearn, known as “puyeque”, tree up to 25 m high; its wood is locally used in house building (“soleras”, “colgados”); Laguncularia racemosa (L.) Gaertn. f. or “mangle”, tree over 30 m tall; its wood is used with commercial purposes in making the structures to dry tobacco and Rhizophora mangle L. or “candelón”, that can measure 25 m high and its wood is used in the region as building material (“soleras”, chains) (Valdez, 2002; Valdez, 2004).
Procurement and sample preparation
From July to August, 2004, 88 sampling units were established (UM) of 300 m2 (30 × 10 m), with three sampling subunits (SUM) of 100 m2 (10 × 10 m) each (Valdez, 2002). At the center of every SUM, a soil sample was taken from 0 to 20 cm deep, with a total number of 264 (Acosta et al., 2001).
Also, apparent density (Da) was determined by a cylindric drill at 0 to 20 cm deep (Porta et al., 2003).
To quantify edaphic C, the 264 samples taken from the soil were dried at room temperature and weighed; afterwards, slow combustion of OM by heat at 750 °C (USDA, 1973) was applied to be weighed once more. That is, the amount of OM of the soil (MOS) was obtained; to calculate the organic

51
Valdés et al., Evaluación del carbono orgánico en suelos de los Manglares de Nayarit
Fuente: RHARAP, 2008.Source: RHARAP, 2008.
Figura 1. Área de estudio con los sitios de muestreo en Marismas Nacionales, Nayarit.Figure 1. Study area with the sampling sites at Marismas Nacionales, Nayarit State.
Además, se determinó la densidad aparente (Da) mediante una barrena cilíndrica a una profundidad de 0 a 20 cm (Porta et al., 2003).
Para cuantificar el C edáfico, las 264 muestras de suelo extraídas fueron secadas a temperatura ambiente y pesadas; a continuación se aplicó una combustión lenta de la MO por calentamiento a 750 °C (USDA, 1973) y se volvieron a pesar. Es decir, se obtuvo la cantidad de MO del suelo (MOS); para calcular el contenido de carbono orgánico del suelo (COS) ese valor se dividió entre 1.74. Por último se hizo la conversión a toneladas de carbono por hectárea.
Todos los datos procedieron de la misma profundidad del suelo (0 a 20 cm) y se analizaron considerando las 10 asociaciones de manglar y nueve clases de geoforma existentes en los sitios de muestreo.
Las asociaciones de manglar son combinaciones de las tres especies antes citadas (Valdez, 2004), y se hace referencia a ellas de la siguiente manera: A =A. germinans, L = L. racemosa
carbon content in soil (COS) that value was divided into 1.74. Finally, a transformation of carbon tons per hectare was made.
All of the data came from the same soil depth (0 to 20 cm) and were analyzed considering the 10 mangrove associations and nine geoform classes present in the sampling site.
Mangrove associations are combinations of the three previously quoted species (Valdez, 2004), and are referred to as: A =A. germinans, L = L. racemosa and R = R. mangle if they are dominant; with the same letters but small (a, l, r), when they are not. For example, a “Lra” key means prevalence of L . racemosa over R. mangle , and of the latter over A. germinans.
Geoform classes are different coastal landscape morphologies with mangrove (Bird, 2003) and the following were detected: Bra = Loamy bar, Bar = Sandy bar, Cnp = Non-marsh basin, Cpt = Marsh basin, Isa = Island, Ise = Islet, Orc = Channel edge, Orl = Lagoon edge, Orr = River bank (Figure 2).

52
Rev. Mex. Cien. For. Vol. 2 Núm. 8
y R = R. mangle, si son dominantes; se indican con las mismas letras, pero en minúsculas (a, l, r), cuando no lo son. Por ejemplo, una clave “Lra” significará predominancia de L. racemosa sobre R. mangle y de esta última sobre A. germinans.
Las clases de geoforma son distintas morfologías del paisaje costero con presencia de manglar (Bird, 2003) y se identificaron las siguientes: Bra = Barra arcillosa, Bar = Barra arenosa, Cnp = Cuenca no pantanosa, Cpt = Cuenca pantanosa, Isa = Isla, Ise = Islote, Orc = Orilla de canal, Orl = Orilla de laguna, Orr = Orilla de río (Figura 2).
Con la finalidad de reconocer otras relaciones estadísticas significativas, se formaron combinaciones de clases de geoforma: Barras (“B”), Cuencas (“C”), Islas (“I”) y Orillas (“O”); así como, combinaciones de asociaciones de manglar: manglares con predominancia del género Avicennia (“A”), de Laguncularia (“L”) y de Rhizophora (“R”).
Análisis estadístico
La identificación de las clases de geoformas y asociaciones de manglar se hizo mediante polígonos con el programa ArcView GIS versión 3.2 (ESRI, 1999); además, para distinguir texturas y tonos de grises se usaron ortofotos digitales escala 1:20,000 (INEGI, 2005) del área de estudio (Figura 2).
Los análisis estadísticos y los datos de laboratorio y gabinete se llevaron a cabo con los programas Minitab 14 (MINITAB, 2003) y SAS (SAS, 1999) e incluyeron los que se enlistan a continuación (Rebolledo, 1999; Infante y Zárate, 2003):
a) Pruebas de normalidad para identificar el grado de heterogeneidad u homogeneidad de los datos, mediante el uso s is temát ico de la prueba de residuales.
b) Análisis de varianza (ANOVA) con niveles de significancia < 0.05 o mejores.
c) Pruebas de medias (Tukey) para identificar posibles diferencias significativas entre grupos (clases de geoforma y asociaciones de manglar).
En principio se obtuvieron 264 datos, de los que se eliminaron 10 atípicos que, según la prueba de residuales, no contribuyeron de forma significativa al modelo analizado (p > 0.05).
RESULTADOS Y DISCUSIÓN
Las nueve clases de geoforma y las 10 asociaciones de manglar se muestran en el Cuadro 1, y en el Cuadro 2 se agrupan las cuatro clases principales de geoforma y tres asociaciones de manglar.
El potencial para el secuestro de carbono (C) en los trópicos húmedos radica en la biomasa aérea, no en el suelo (Palm et al., 2000; Van Noordwijk et al., 1997). Sin
In order to find different statistical relations, combinations of geoforms were formed: Bars (“B”), Basins (“C”), Islands (“I”) and Edges (“O”), as well as combinations of mangrove associations: mangroves where Avicennia genus prevailed (“A”), where Laguncularia (“L”) prevailed or Rhizophora (“R”) did.
Statistical Analysis
The identification of geoform classes and mangrove associations was made by polygons with the 3.2 ArcView GIS program (ESRI, 1999); to detect textures and grey tones, 1:20,000 digital orthophotos of the study area were used (INEGI, 2005) (Figure 2).
Fuente: INEGI, 2005).Source: INEGI, 2005.
Figura 2. Ejemplo de delimitación de clases de geoforma (polígonos color claro) y asociaciones de manglar (polígonos color oscuro).
Figure 2. Example of the geoform classes demarcation (light color polygons) and mangrove associations
Statistical analysis and laboratory and office data were carried out with the following programs Minitab 14 (MINITAB, 2003) and SAS (SAS, 1999), which included the following (Rebolledo, 1999; Infante and Zárate, 2003):
a) Normality tests to identify the heterogeneity or homogeneity degree of the data by the systematic use of residuals test.
b) Analysis of variance (ANOVA) with < 0.05 significance levels or better.
c) Mean tests (Tukey) to identify the possible significant differences among groups (geoform classes and mangrove associations).

53
Valdés et al., Evaluación del carbono orgánico en suelos de los Manglares de Nayarit
At first, 264 data were obtained, from which 10 were eliminated, since, according to the residuals test, made no significant contribution to the analyzed model (p > 0.05).
RESULTS AND DISCUSSION
The nine geoform classes and the 10 mangrove classes are shown in Table 1 and in Table 2 are grouped the four main geoform classes and three mangrove associations.
The potential for carbon sequestration (C) in the humid tropics lies in aerial biomass, not in the soil (Palm et al., 2000; Van Noordwijk et al., 1997). However, in the case of wetlands, and particularly mangroves, the stored edaphic C is very important, if it is taken into account the great amount that is released when they are destructed by land use change. De la Cruz (1986) states that that number may be around 375 t C ha-1
of the edaphic layer of a mangrove, when it is deforested and dug out to build fish-tanks.
The Islote geoform class (Ise) has a significantly higher COS content (F = 4.02, gl = 8, p < 0.05) than the Sandy bar (Bar) and Loamy bar (Bra) classes, but from the statistical view point, it was the same (p > 0.05) than the Island (Isa) class (Table 1, Figure 3).
The geoform classes grouped into Islands were significant (F = 2.26, df = 4, p < 0.05) in regard to the higher average COS contents than the Edges, Basins and Bar groups (Table 2).
Cuadro 2. Contenido promedio de carbono orgánico del suelo (t ha-1) por combinación de clase de geoforma y asociación de manglar.
Table 2. Average organic soil carbon content (COS) (t ha-1) by a combination of geoform class and mangrove association.
Clase de geoforma (combinada)1
Descripción Clave COS (t ha-1) NBarra B 57.9c 12Orilla O 91.0b 172Isla I 125.9ª 12Cuenca C 85.7b 58
Asociación de manglar (combinado)2
Especie predominante
ClaveCOS (t ha-1) N
A. germinans A 81.4b 73L. racemosa L 96.6ª 136R. mangle R 81.5b 45
1Las clases de geoforma están ordenadas mar-tierra adentro.2Las asociaciones de manglar están ordenados alfabéticamente.N = Cantidad de muestras.Medias seguidas con letras distintas indican diferencias significativas (Tukey, p < 0.05) entre hileras.1 Geoform clases are ordered in a sea-inland distribution2 Mangrove associations follow an alphabetical order. N = Sample numberMeans followed by different letters show significant differences (Tukey, p < 0.05) between rows.
embargo, en el caso de los humedales, y en particular de los manglares, el C edáfico almacenado es de gran trascendencia, si se considera la enorme cantidad que se libera cuando son destruidos por cambio de uso. De la Cruz (1986) afirma que esta cifra puede ser de alrededor de 375 t C ha-1 de la capa edáfica de un manglar, cuando es deforestado y excavado para construir estanques acuícolas.
Cuadro 1. Contenido promedio de carbono orgánico del suelo (COS) (t ha-1) por clase de geoforma y asociación de manglar (año 2004).
Table 1. Average organic soil carbon content (COS) (t ha-1) by geoform class and mangrove association (2004 year).
Clase de geoforma 1
Descripción Clave COS
(t ha-1)
N
Barra arenosa Bar 61.1 (c) 6Barra arcillosa Bra 54.6 (c) 6Orilla de laguna Orl 85.3 (abc) 59Orilla de río Orr 97.9 (abc) 84Orilla de canal Orc 75.3 (bc) 29Isla Isa 117.6 (ab) 6Islote Ise 128.5 (a) 6Cuenca pantanosa Cpt 85.7 (abc) 27Cuenca no pantanosa Cnp 91.8 (abc) 31
Asociación de manglar2
Descripción Clave COS
(t ha-1)
N
A. germinans con L. racemosa Al 80.0 (a) 46A. germinans con L. racemosa
y R. mangle
Alr
85.4 (a) 27
L. racemosa L 100.3 (a) 7L. racemosa con A. germinans La 94.4 (a) 47L. racemosa con A. germinans y R. mangle
Lar89.5 (a) 33
L. racemosa con R. mangle Lr 106.7 (a) 19L. racemosa con R. mangle y A. germinans
Lra102.7 (a) 30
R. mangle con A. germinans y L. racemosa
Ral
94.4 (a) 6
R. mangle y L. racemosa Rl 85.3 (a) 13R. mangle con L. racemosa y A. germinans
Rla73.7 (a) 26
Las clases de geoforma están ordenadas mar-tierra adentro.2 Las asociaciones de manglar están ordenados alfabéticamente.N = Cantidad de muestrasMedias seguidas con letras distintas indican diferencias significativas (Tukey, p < 0.05) entre hileras.1 Geoform clases are ordered in a sea-inland distribution2 Mangrove associations follow an alphabetical order. N = Sample numberMeans followed by different letters show significant differences (Tukey, p < 0.05) between row

54
Rev. Mex. Cien. For. Vol. 2 Núm. 8
Las líneas en las barras indican el error estándar.The lines in the bars refer to standard error.
Figura 3. Contenido promedio de carbono orgánico del suelo por clase de geoforma.
Figure 3. Average organic soil carbon content by geoform class
Fort the Laguncularia racemosa – Rhizophora mangle (Lr.) was observed a higher significant average organic soil carbon content (COS) (F = 2.28, gl = 9, p < 0.0204) than that of the Avicennia germinans – L. racemosa (Al.) and R. mangle – L. racemosa – A. germinans (Rla.) associations, but in the statistical scope, it is the same (p > 0.05) as that of L. racemosa – R. mangle – A. germinans (Lra.) and L. racemosa (L.) (Figure 4).
In the mangroves of Nayarit State, the higher average COS contents were in the Island and Islet geoforms (117.6 and 128.5 t C ha-1, respectively), as a response to the high loam content in soils and the important accumulation of OM that comes, mainly, from R. mangle leaves, which is the dominant species at river banks and lagoons, and that have the least decomposition rate of litter, compared to other mangrove taxa (Yáñez-Araceiba et al., 1998). Fujimoto (2000) determined that R. mangle forests are better to capture C than forests with different composition.
Geoform class is related, as well, with the greatest COS content, since it is affected by hydrology that favors OM dragging and accumulation, as it is a medium size area surrounded by channels and lagoons or estuaries. Pool et al. (1975) acknowledged that the most productive mangroves are the best protected estuaries and more regularly fed by soft currents, in contrast to those of the coast in which stronger currents occur and sediments are more unstable. Fujimoto (2000) confirmed higher C concentrations (39 and 49 kg m-2) in geoforms of small deltas and estuaries that in mega deltas or basins (27 and 33 kg m-2). There are many conditions that may control or determine productivity and diversity of vegetal associations, such as climate, geomorphology, tide range, fresh water and other factors (Pool et al., 1975). Thus, it is surprising that their edaphic characteristics have received a rather poor attention from scientific researchers up to now.
La clase de geoforma “Islote” (Ise) tuvo un contenido de COS significativamente (F = 4.02, gl = 8, p < 0.05) mayor que las clases “Barra arenosa” (Bar) y “Barra arcillosa” (Bra), pero desde el punto de vista estadístico igual (p > 0.05) que la clase “Isla” (Isa) (Cuadro 1, Figura 3).
Las clases de geoforma agrupadas en “Islas” tuvieron significancia (F = 2.26, gl = 4, p < 0.05) respecto a los mayores contenidos promedio de COS, que los grupos “Orillas”, “Cuencas” y “Barras” (Cuadro 2).
Para las asociaciones de manglar Laguncularia racemosa – Rhizophora mangle (Lr.) se observó un contenido promedio de COS con significancia (F = 2.28, gl = 9, p < 0.0204) mayor que las asociaciones Avicennia germinans – L. racemosa (Al.) y R. mangle – L. racemosa – A. germinans (Rla.), (Lra.) y L. racemosa (L.) (Figura 4).
En los manglares de Nayarit, los contenidos más altos en promedio de COS estuvieron en las geoformas Isla e Islote (117.6 y 128.5 t C ha-1, respectivamente), en respuesta al alto contenido arcilloso del suelo y a la acumulación importante de MO procedente de las hojas de R. mangle, en mayor medida, especie predominante en orillas de ríos y lagunas y que posee menor tasa de descomposición de la hojarasca, en comparación con otros taxa de mangle (Yáñez-Araceiba et al., 1998) . Fuj imoto (2000) determinó que los bosques de R. mangle son mejores para capturar C, que los bosques con diferente composición.
La clase de geoforma está relacionada también con el mayor contenido de COS, pues al tratarse de una superficie de tamaño moderado, rodeada de canales por un lado y de la laguna o el estero por el otro, es influenciada por una hidrología que favorece el acarreo y acumulación de MO. Pool et al. (1975) reconocieron que los manglares más productivos son los de estuarios mejor protegidos y alimentados en forma más regular por corrientes suaves, en oposición a los de la costa en los que se presentan corrientes de agua más fuertes y sedimentos más inestables. Fujimoto (2000) confirmó concentraciones superiores de C (39 y 49 kg m-2) en geoformas de deltas pequeños y estuarios que en mega deltas o cuencas (27 y 33 kg m-2). Existen muchas condiciones que pueden controlar o determinar la productividad y diversidad en estas asociaciones vegetales, entre ellas el clima, la geomorfología, rango de mareas, entrada de agua dulce y otros factores (Pool et al., 1975). Por lo tanto, es sorprendente que las características edáficas de las mismas hayan recibido, relativamente, poca atención de los investigadores hasta el momento.
En el presente trabajo, las geoformas orilla y cuenca no tuvieron los valores más altos de COS, pero su importancia radica en que son las zonas con aprovechamiento forestal más intenso en la región y están amenazadas por el cambio de uso del suelo para infraestructura turística o acuicultura.

55
Valdés et al., Evaluación del carbono orgánico en suelos de los Manglares de Nayarit
En particular, la clase de geoforma “Orilla de río” (Orr) está destinada a la protección de cauces (Valdez, 2004), lo cual señala su papel ecológico en la retención de C en el suelo.
El contenido promedio de COS, en general, fue más alto en manglares con predominio de L. racemosa (Figura 4), que parece contradecir lo planteado por Betoulle et al., (2001), quienes determinaron mayor contenido de CO en mantillo de A. germinans y R. mangle, que en L. racemosa. Probablemente, el contenido más grande de COS en las asociaciones de manglar donde dominaba L. racemosa se deba a que estas se ubicaron, por lo regular, en las geoformas de orillas, posición que se favoreció por el régimen hidrológico de corrientes suaves (Pool et al., 1975).
Los manglares con predominancia de L. racemosa, en especial aquellos con presencia de A. germinans, y que tuvieron los más altos contenidos de COS son los más amenazados por futuros cambios en uso de suelo para la región de Marismas Nacionales. Por ello, es necesario continuar con la elaboración de planes de manejo a escala local que permitan unaprovechamiento sustentable de los recursos naturales (Valdez, 2004).
Los manglares de Nayarit tuvieron un contenido de COS de 89,7 t C ha-1 superior a lo que se conoce en manglares templados de Japón, que fue de 57 t C ha-1 (Khan et al., 2007) en una profundidad similar de muestreo, pero menor contenido que en otros de tipo tropical de México y del mundo, como en Tabasco, con 132 t C ha-1 (Moreno et al., 2002a), Micronesia con 97 t C ha-1 (Fujimoto et a.l, 1999) y Senegal con 91 t C ha-1
(Woomer et al., 2004).
Las líneas en las barras indican el error estándar.The lines in the bars refer to standard error.
Figura 4. Carbono orgánico del suelo por asociación de manglar.
Figure 4. Soil organic carbon by mangrove association
In the actual work, Edge and Basin geoforms did not get the highest COS values, but they are important as they are the most intense harvested forest zones of the region and they are threatened by land use change for tourism or aquaculture infrastructure. The River bank geform class (Orr) in particular, is intended to protect the river beds (Valdez, 2004), which appoints an ecological role in soil C retention.
Average COS content, in general, was higher in mangroves where L. racemosa prevails (Figure 4), which seems to be contrary to Betoulle et al. (2001), who determined a higher CO content in the litter of A. germinans and R. mangle over L. racemosa. Probably, the great COS content in the mangrove associations where L. racemosa dominated is due to the place where they were located, as they were regularly in the Edge geoforms, which were favored by the light currents regime (Pool et al., 1975).
Mangroves where L. racemosa prevails, especially those with A. germinans, and that had the highest COS contents, are the mos t threatened by fu ture changes in land use for the Marismas Nacionales regions. Thus, it is necessary to continue with local management plans that allow the sustainable use of natural resource (Valdez, 2004).
The mangroves of Nayarit had a COS content of 89,7 t C ha-1 which is over the records of Japan, 57 t C ha-1 (Khan et al., 2007) at a similar sampling depth, but lower that in other tropical soils of Mexico as Tabasco with 132 t C ha-1 (Moreno et al., 2002a); and other parts of the world, like Micronesia with 97 t C ha-1 (Fujimoto et al., 1999) and Senegal with 91 t C ha-1 (Woomer et al., 2004).
In terms of per cent, in the mangrove forests of Nayarit, more COS (7%) was estimated than what Fujimoto (2000) determined in Vietnam (3%), Sukardjo (1994) in Indonesia (4%) and Van Noordwijk et al. (1997) in wetlands of Sumatra (6%); but lower than those described by Boto and Wellington (1984) in mangrove of Northern Australia (8%), Betoulle et al. (2001) in the French Guyana (33%) and Khan et al. (2007) in Japan (47%).
With the same sampling depth (0 a 20 cm) and compared to other natural tropical systems, in the studied mangroves was found a lower amount of COS than what Callo-Concha et al. (2001) determined in a primary forest of the Peruvian Amazonia (99 t C ha-1), and in a grassland of the same region (93 t C ha-1).
When the studied mangroves of Nayarit State were measured against the natural subtropical forest of the alluvial plains of Bajo Paraná, Argentina, Aceñolaza et al. (2008) determined high COS concentrations in Salix humboldtiana Willd. forests (93 t C ha-1), but lower than 34 t C ha-1 in Tessaria integrifolia Ruiz & Pavón forests and 33 t C ha-1in Albizia inundata (Mart.) Barneby & J. W. Grimes forests.

56
Rev. Mex. Cien. For. Vol. 2 Núm. 8
En porcentaje, en los bosques nayaritas de mangle se estimó más COS (7%) que lo calculado por Fujimoto (2000) en Vietnam (3%), Sukardjo (1994) en Indonesia (4%), y Van Noordwijk et al. (1997) en humedales de Sumatra (6%); pero menor que lo descrito por Boto y Wellington (1984) en manglares del norte de Australia (8%), Betoulle et al. (2001) en Guyana Francesa (33%) y Khan et al. (2007) en Japón (47%).
Con la misma profundidad de muestreo (0 a 20 cm) y en comparación con otros sistemas naturales tropicales, en los manglares estudiados se obtuvo menor cantidad de COS que lo determinado por Callo-Concha et al. (2001) en un bosque primario de la Amazonia Peruana, con 99 t C ha-1 y en un pastizal de la misma región, con 93 t C ha-1. Al confrontar los manglares de Nayarit con los bosques naturales subtropicales de la llanura aluvial de Bajo Paraná en Argentina, Aceñolaza et al. (2008) cuantificaron concentraciones altas de COS, con 93 t C ha-1 en bosques de sauce (Salix humboldtiana Willd.), pero menor a 34 t C ha-1 en bosques de aliso del río (Tessaria integrifolia Ruiz & Pavón) y en bosques de timbó blanco (Albizia inundata (Mart.) Barneby & J. W. Grimes) con 33 t C ha-1.
A partir del intervalo promedio de contenido de COS surgido de este trabajo, así como la superficie total de la región de Marismas Nacionales en el estado de Nayarit (3,876,873 ha) (CONABIO, 2008) se estimaron de 81.8 a 89.7 t C ha-1 totales en los primeros 20 cm del suelo. Por lo tanto, es evidente que los suelos de los manglares de Nayarit contienen cantidades notables de COS, lo cual representa un gran valor natural y juega un rol significativo en el ciclo del C a escala local.
A pesar de que la biomasa aérea de cualquier bosque posee mayor potencial para la captura de C, el suelo ejerce un papel clave en la regulación de su balance y ciclo. La trascendencia de lo anterior se acentúa en el caso de los humedales, en particular de los manglares, donde las condiciones de anaerobiosis por efecto de la inundación parcial o total, permanente o intermitente retardan la descomposición de la MO, que a su vez evita la liberación del C a la atmósfera y lo conserva atrapado en el suelo, incluso por milenios. Es por ello, que la deforestación de un manglar por cambio de uso de suelo con fines, por ejemplo, turísticos o de producción acuícola comercial, libera grandes cantidades de C a la atmósfera.
CONCLUSIONES
Los suelos en las clases de geoforma combinadas de Isla tuvieron valores superiores de COS con 125.9 t ha-1, seguidos de los suelos en las geoformas de Cuencas y Orillas con 85.7 y 91.0 t ha-1, respectivamente; mientras que, en las geoformas de Barras se estimaron los contenidos menores (57.9 t ha-1). Sin embargo, debe centrarse la mayor atención en las clases de geoformas de Orilla, dado que son las más vulnerables por estar en contacto directo con los cambios de regímenes hidrológicos diarios (intermareales) y estacionales
From the COS average content interval from this work as well as the total area of the Marismas Nacionales region in Nayarit State (3,876,873 ha) (CONABIO, 2008), 81.8 to 89.7 t C ha-1 were calculated for the first 20 cm of soil depth. Therefore, it is evident that the soils of the mangroves of Nayarit
have remarkable amounts of COS, which has a great natural value and plays a significant role in the C cycle at a local scale.
In spite of the fact that the aerial biomass of any forest has a greater potential for C sequestration, soil plays a crucial role in its balance and cycle regulation. The relevance of the former statement is emphasized in wetlands, mangroves in particular, where anaerobic conditions from the partial or total flooding, permanent or intermittent, delay OM decomposition, which avoid C release to the atmosphere and keeps it trapped in the soils, even for millennia. Thus, mangrove deforestation from land use change with tourism or commercial fish production, for example, releases great C amounts to the atmosphere.
CONCLUSIONS
The soils in the geoform classes with the Island combination reported COS 125.9 t ha-1 followed by the soils in the Basins (85.7 t ha-1) and Edges (91.0 t ha-1) geoforms, while in the Bars geoforms contents were lower (57 .9 t ha-1). However, the greatest attention must be focused on the Edge geoforms, since they are the most vulnerable as they are in direct contact with daily (intertidal) or seasonable (rains and droughts) hydrologic changes as well as for being where the most intense activities of wood harvest take place.
In regard to the mangrove associations, the highest COS content was confirmed in the Laguncularia racemosa – Rhizophora mangle (106.7 t ha-1) combination, and the lowest in R. mangle –L. racemosa –A. germinans (73.7 t ha-1). In general, in the concentrated mangrove associations where L. racemosa prevails, the maximal COS contents (96.6 t ha-1) were found. Since this mangrove association is more frequent in the Edge geoforms, it is concluded than in the management and conservation plans for mangroves of Marismas Nacionales, the conservation of the Edge geoforms, in particular where this species dominates, must be considered a priority.
Compared to other mangroves of Mexico and the world, the soils of Marismas Nacionales confirmed to be essential as COS storage, with values near to those of Histosols. The mangroves of Nayarit rank as one of the richest of planet Earth in terms of their CO regulatory function in nature, as they have a greater ability to store huge volumes of C in the soil, than other world-famous ecosystems for their OM content, such as grasslands and tropical forests.

57
Valdés et al., Evaluación del carbono orgánico en suelos de los Manglares de Nayarit
(lluvias y sequía), y por ser, en ellas, donde se desarrolla la actividad más intensa de aprovechamiento maderero.
Respecto a las asociaciones de manglar, el contenido de COS más alto se verificó en la combinación Laguncularia racemosa – Rhizophora mangle (106.7 t ha-1) y el menor en la de R. mangle –L. racemosa –A. germinans (73.7 t ha-1). En general, las asociaciones de manglar concentradas con predominancia de L. racemosa registraron los mayores contenidos de COS, con 96.6 t ha-1. Dado que esta asociación de manglares es más frecuente en las geoformas de Orilla, se concluye que los planes de manejo y conservación orientados a los manglares de Marismas Nacionales deben considerar como prioritaria la preservación de estas geoformas, en particular aquellas donde domine dicha especie.
En comparación con otros manglares de México y del mundo, los suelos de los de Marismas Nacionales demostraron ser fundamentales como almacenadores de COS, con valores cercanos a los de Histosoles. Los manglares de Nayarit figuran entre los biomas más ricos del planeta, en cuanto a su función reguladora del CO en la naturaleza, por tener mayor capacidad para almacenar grandes volúmenes de C en el suelo, que otros ecosistemas del mundo famosos por su riqueza en MO, como los pastizales y las selvas.
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca otorgada al primer autor y por financiar el inicio del estudio; a los ejidatarios de las comunidades de Mexcaltitán, Los Morillos, Palma Grande, Santa Cruz y Unión de Corrientes en el estado de Nayarit por su apoyo en el trabajo de campo; al Ing. Alfonso Solís Venegas por la ayuda técnica y logística; al Consejo Superior de Investigaciones Científicas (CSIC) de Salamanca y a la Universidad de Salamanca (España) por las facilidades brindadas durante el análisis en laboratorio de las muestras colectadas y al World Wildlife Fond (WWF) por la beca “Príncipe Bernhard” para el primer autor y por financiar la conclusión de la investigación.
REFERENCIAS
Aceñolaza, P., L. P. Zamboni y J. F. Gallardo L. 2008. Estimación de carbono en tres bosques de la llanura aluvial del Bajo Paraná (R. Argentina). In: Gallardo., L., J. F. (Comp.). La captura de carbono en ecosistemas terrestres iberoamericanos. Red Iberoamericana de Física y Química Ambiental. Sociedad Iberoamericana de Física y Química Ambiental. Salamanca, España. pp. 39-53.
Acosta, M., J. D. Etchevers B., C. Monreal, K. Quednow y C. Hidalgo M. 2001. Un método para la medición del carbono en los compartimentos subterráneos (raíces y suelo) de sistemas forestales y agrícolas de ladera en México. In: Memorias del Simposio Internacional Medición y Monitoreo de la Captura de Carbono en Ecosistemas Forestales, Universidad Austral de Chile, Valdivia, Chile. p. 15.
Alongi, D. M. 2002. Present state and future of the world’s mangrove forests. Environmental Conservation 29:331-349.
Armentano, T. V. 1981. The potential significance of tropical organic soils for the carbon balance of the terrestrial biosphere. http://www.osti.gov/energyci tat ions/product .biblio . j sp?ost i_id=5468757. (6 de agosto de 2008).
Betoulle, J. L., F. Fromard, A. Fabre et H. Puig. 2001. Caractérisation des chutes de litière et des apports au sol en nutriments dans une mangrove de Guyane française. Canadian Journal of Botany 79:238-249.
ACKNOWLEDGEMENTS
To the Consejo Nacional de Ciencia y Tecnología (CONACYT) for the scholarship granted to the first author and for financing the initial stage of the actual study; to the people of; Mexcaltitán, Los Morillos, Palma Grande, Santa Cruz y Unión de Corrientes in Nayarit State for their help in field work; to Eng. Alfonso Solís Venegas for his technical and logistic help; to the Consejo Superior de Investigaciones Científicas (CSIC) of Salamanca, and the Universidad de Salamanca (Spain) for the facilities provided during the laboratory analysis of the collected samples and to the World Wildlife Found (WWF) for the “Príncipe Bernhard” scholarship awarded to the first author and for sponsoring the conclusion of this research study.
End of the English version
Bird, E. 2003. Coastal geomorphology: an introduction. Wiley and Sons, Inc. West Sussex, UK. 340 p.
Bojórquez, I., A. Hernández, D. García, O. Nájera, F. Flores, A. Madueño y R. Bugarín. 2008. Características de los suelos de las barras paralelas, playas y dunas de la llanura costera norte del estado de Nayarit, México. Cultivos Tropicales 29(1):37-42.
Boto, K. G. and J. T. Wellington. 1984. Soil characteristics and nutrient status in a Northern Australia Mangrove Forest. http://www.jstor.org/stable/1351957. Estuarine Research Federation. Estuaries 7(1):61-69. (6 de agosto de 2008).
Bravo, F. (Coord.) 2007. El papel de los bosques en la mitigación del cambio climático. Fundación Gas Natural, Barcelona, España. 320 p.
Callo-Concha, D., L. Krishnamurthy y J. Alegre. 2001. Estimación del carbono secuestrado por algunos sistemas agroforestales y testigos en tres pisos de la Amazonia Peruana. In: Memorias del Simposio Internacional Medición y Monitoreo de la Captura de Carbono en Ecosistemas Forestales . Universidad Austral de Chile (UACH). Valdivia, Chile. p. 23.
Comisión Nacional para la Biodiversidad (CONABIO). 2008. 22. Río Baluarte–Marismas Nacionales. http://www.conabio.gob.mx/conocimiento/regionalizacion/doctos/rhp_022.html. (4 de julio de 2008).
De la Cruz A., A. 1986. Tropical wetlands as a carbon source. Aquatic Botany 25:109-115.
Environmental System Research Institute (ESRI). 1999. ArcView GIS 3.2. Neuron Data, Inc. EUA. Sistema de Información Geográfica. Seattle, WA. USA. s/p.
Etchevers B., J. D., C. Hidalgo M., J. Padilla, R. M. López, C. Monreal, C. Iturralde, B. Rapidez, F. De León, M. Acosta M., A. Vergara, K. Quednow, A. Fernández y C. Figueroa. 2002. Subproyecto II: Metodología de la medición de la captura de carbono. Proyecto Manejo Sostenible de Laderas (PMSL). Regiones Cuicateca, Mazateca y Mixe, Oaxaca, México. Colegio de Postgraduados de Montecillo. SAGARPA-GEF-Banco Mundial. México, D. F. México. pp. 23-45.
Fujimoto, K., A. Imaya, R. Tabuchi, S. Kuramoto, H. Utsugi and T. Murofushi. 1999. Belowground carbon storage of Micronesian mangrove forests. Ecological Research 14:409-413.
Fujimoto, K. 2000. Below-ground carbon sequestration of mangrove forests in the Asia-Pacific Region. In: International Workshop Asia-Pacific Cooperation. Research for Conservation of Mangroves. 26 - 30 de marzo, 2000; Okinawa, Japón. http://landbase.hq.unu.edu/Workshops/OkinawaMarch2000/Papers/Fujimotopapermar2000.htm. LandBase. Environmental Monitoring and Governance in the East Asian Hydrosphere 2008. s/p. (22 de agosto de 2008).
Gallardo L., J. F. and M. I. González. 2004. Sequestration of C in a Spanish chestnut coppice. Investigación Agraria. Sistema de Recursos Forestales:108-113.
García, E. 2004. Modificaciones al sistema climático de Köppen para la Republica Mexicana. 5ta Edición. Instituto de Geografía. Serie de libros No. 6. Universidad Nacional Autónoma de México. México, D. F. México. 292 p.

58
Rev. Mex. Cien. For. Vol. 2 Núm. 8
Gleason, S. M. and K. C. Ewel. 2002. Organic matter dynamics on the forest floor of a Micronesian mangrove forest: an investigation of species composition shifts. Biotropica 34(2):190-198.
Instituto Nacional de Estadística, Geografía e Informática (INEGI). 2002. Síntesis de información geográfica del estado de Nayarit. Aguascalientes, Ags. México. 140 p.
Instituto Nacional de Estadística, Geografía e Informática (INEGI). 2005. Ortofotos digitales de la región de Marismas Nacionales, Nayarit, esc. 1:20 000 F13A88(E), F13C18(E). México.
Infante G., S. y G. P. Zárate de Lara. 2003. Métodos estadísticos; un enfoque interdisciplinario. 2 ed. Ed. Trillas. México, D. F. México. 643 p.
International Union of Soil Science (IUSS) Working Group World Reference Base (WRB). 2006. World reference base for soil resources 2006. 2 ed. World Soil Resources Reports No. 103. FAO, Rome. 128 p.
Khan, M., S. Suwa and A. Hagihara. 2007. Carbon and nitrogen pools in a mangrove stand of Kandelia obovata (S. L.) Yong: distribution in the soil and vegetation system. Wetlands Ecology and Management 15(2): 141-153.
López P., J. y E. Ezcurra. 2002. Los manglares de México: una revisión. Madera y Bosques Número Especial 1:25-51.
MINITAB. 2003. Minitab Release 14 (Version 14.10). Statistical Software. USA. s/p.
Mitra, S.; R. Wassmann and P. L. G. Vlek. 2005. An appraisal of global wetland area and its organic carbon stock. Current Science 88(1):25-35.
Moreno C., E., A. Guerrero, M. C. Gutiérrez C., C. A. Ortiz S. y D. J. Palma. 2002a. Los manglares de Tabasco: una reserva natural de carbono. Madera y Bosques. Número especial 1:115-128.
Moreno C., P., J. L. Rojas, D. Zárate, M. A. Ortiz, A. L. Lara, y T. Saavedra. 2002b. Diagnóstico de los manglares de Veracruz; distribución, vínculo con los recursos pesqueros y su problemática. Madera y Bosques. Número especial 1:61-88.
Osnaya, P. 2003. Avances de México en materia de cambio climático 2001-2002. Osnaya, P. (Comp.), Fernández, A. y J. Martínez (Coord.). Instituto Nacional de Ecología-Semarnat. México. 104 p.
Palm, C. A., P. L. Woomer, J. Alegre, L. Arévalo, C. Castilla, D. G. Cordeiro, B. Feigl , K . Hairiah, J . Kotto-Same, A. Mendes, A. Moukam, D. Murdiyarso, R. Njomgang, W. J. Parton, A. Ricse, V. Rodrigues, S. M. Sitompul and M. Van Noordwijk. 2000. Carbon sequestration and trace emissions in slash and burn and alternative land uses in the humid tropics. ASB Climatic Change Working Group Report. Final Report, Phase 2. Nairobi, Kenya. 25 p.
Ponce-H.,R. 1999. Assessing the carbon stock and carbon sequestration potential of current and potential land use systems and the economic rationality of land use conversions . In : Food and Agriculture Organization (FAO) (Comp). Prevention of land degradation, enhancement of carbon sequestration and conservation of biodiversity thorough land use change and sustainable land management wit focus in Latin America and the Caribbean. FAO World Soil Resources Reports 86. pp:79-94.
Porta, J., M. López A. y C. Roquero. 2003. Edafología para la Agricultura y el Medio Ambiente. 3 Ed. Mundi-Prensa. Madrid, España. 917 p.
Post, W. M., T. H. Peng, W. R. Emanuel, A. W. King, V. H. Dale, and D. L. De Angelis. 1990. The global carbon cycle. American Scientist 78:310–326.
Pool, D. J., A. E. Lugo and S. C. Snedeker. 1975. Litter production in mangrove forests of Southern Florida and Puerto Rico. In: Walsh, G. E., S. C. Snedeker and H. J. Teas. (Comp.) Proccedings of the International Symposium of Biology an Management of Mangroves. Institute of Food and Agriculture Sciences, University of Florida, Gainesville, FL, USA. pp. 213 - 237.
Rebolledo H., H. 1999. Statistical Analysis System SAS en microcomputadora; Análisis estadístico de datos experimentales. Departamento de Suelos. Universidad Autónoma Chapingo. Chapingo, Edo. de Méx. México. 176 p.
Red Hemisférica de Reservas para Aves Playeras (RHARAP). 2008. Mapa de Marismas Nacionales. Galería de Fotos. http://www.whsrn.org/data/Unsorted/mapa_grande-21919-1.jpg. (1 de diciembre de 2008).
Rosenzweig, C. and D. Hillel. 2000. Soils and global climate change: Challenges and opportunities. Soil Science, 165(1):47-56.
SAS - Institute. 1999. The SAS System. (Versión 5.1 para Windows). Cary, NC, EUA. s/p.
Sukardjo, S. 1994. Soils in the mangrove forests of the Apar Nature Reserve, Tanah Grogot, East Kalimantan, Indonesia. Southeast Asian Studies. 32(3):385-398.
United Nations Framework Convention on Climate Change (UNFCCC). 2000. Sexto periodo de sesiones de la conferencia de las partes en la convención marco de las Naciones Unidas sobre el cambio climático. In: Carpeta de prensa. Convención - Marco sobre el Cambio Climático. ONU. La Haya. Países Bajos. 14 p.
United States Departament of Agriculture (USDA). 1973. Investigación de suelos; métodos de laboratorio y procedimientos para recoger muestras. Trad. por A. Contín. Soil Conservation Service. U. S. Departament of Agriculture. Ed. Trillas. México, D .F., México. pp. 41-45.
Valdez-H., J. L. 2002. Aprovechamiento forestal de manglares en el estado de Nayarit, costa Pacífica de México. Madera y Bosques. Número Especial 1:129-145.
Valdez-H., J. L. 2004. Manejo forestal de un manglar al sur de Marismas Nacionales, Nayarit. Madera y Bosques Número Especial 2:93-104.
Van Noordwijk, M., C. Cerri, P.L. Woomer, K. Nugroho and M. Bemoux. 1997. Soil carbon dynamics in the humid tropical forest zone. Geoderma 79:187-225.
Woomer, P. L., L.L.Tieszen, G. Tappan, A. Touré and M. Sall. 2004. Land use change and terrestrial carbon stocks in Senegal. Journal of Arid Environments 59(3):625-642.
Yáñez-Araceiba, A., R. R. Twillwy y A. L. Lara-Domínguez. 1998. Los ecosistemas de manglar frente al Cambio Climático Global. Madera y Bosques 4(2):3-19.

NOTA DE INVESTIGACIÓN
GERMINACIÓN DE ESPECIES DE CACTÁCEAS EN CATEGORÍA DE RIESGO DEL DESIERTO CHIHUAHUENSE
GERMINATION OF CACTI SPECIES UNDER RISK STATUS OF THE CHIHUAHUAN DESERT
Joel Flores1 y Enrique Jurado 2
RESUMEN
Se presenta una síntesis de los principales resultados sobre germinación de semillas de 28 cactáceas con estatus de conservación y cuatro sin estatus, nativas del desierto chihuahuense. Se incluye la información referente a la presencia de latencia y la forma de romperla, requerimientos de luz, impacto del tiempo de almacenamiento, competencia entre semillas y el efecto del tamaño de las semillas en la germinación. En tres de 17 taxa del género Turbinicarpus se documenta la existencia de latencia y en dos de ellas su pérdida con el paso del tiempo; algo similar se observa en dos especies de Opuntia, pero en combinación con la ayuda de hongos que escarifican la cubierta de las semillas. Entre los taxa con estatus, 17 tienen latencia (germinación ≤ 70%) y las restantes fueron fotoblásticas positivas, con semillas pequeñas cuyo peso fue menor de 1 mg. De dos cactáceas columnares endémicas de México, Isolatocereus dumortieri mostró menor porcentaje de germinación con el aumento de la densidad de semillas; mientras que la germinación de Myrtillocactus geometrizans no fue afectada por este factor. Las semillas pequeñas de cuatro años de edad de Astrophytum myriostigma registraron valores de germinación mayores y esta fue más rápida que en las grandes. Se discuten futuras líneas de investigación que contribuyan a entender la dinámica de las semillas de cactáceas en condiciones ambientales actuales y futuras para fomentar su conservación.
Palabras clave: Cactaceae, escarificación, germinación, latencia, plantas desérticas, semillas.
ABSTRACT
A synthesis of the main results of seed germination of 28 cacti under risk status and 4 without risk from the Chihuahuan Desert is presented here. The effect on germination of dormancy, light, seed age, seed competition and seed size were assessed. Three out of 17 taxa of the genus Turbinicarpus showed dormancy and two of them lost dormancy with age. Two Opuntia species also lost dormancy with age, aided by the presence of fungus that scarified the seed coat. From 28 cacti species at risk, 17 were dormant (germination ≤ 70%); and 11 non-dormant species were positive photoblastic that had small seeds (<1 mg). From the germination of two endemic columnar species, Isolatocereus dumortieri reduced germination at higher seed densities, while that of Myrtillocactus geometrizans was unaffected by seed density. 4 yr old small seeds from threatened Astrophytum myriostigma germinated faster and at higher per cent than large seeds. Future research lines are suggested to get a better understanding of cacti seed dynamics in current and future conditions in order to contribute to the preservation of these plants.
Key words: Cactaceae, seed dormancy, germination, seed scarification, desert plants, seeds.
Fecha de recepción: 16 de mayo de 2011.Fecha de aceptación: 8 de diciembre de 2011.
1División de Ciencias Ambientales. Instituto Potosino de Investigación Científica y Tecnológica, A.C. Correo-e: [email protected] de Ciencias Forestales, Universidad Autónoma de Nuevo León.

60
Rev. Mex. Cien. For. Vol. 2 Núm. 8
The Chihuahuan Desert gathers a great richness of non-wood forest species such as cacti, many of which have been classified into several risk categories as a result of the illegal collection to which they are subjected (Hernández and Godínez, 1994; Hernández and Bárcenas, 1995, 1996; Gómez-Hinostrosa and Hernández, 2000; Hernández et al., 2001). It has been proposed that its ex situ propagation may be an option to reduce its illegal extraction (Hernández and Bárcenas, 1995; Rojas-Aréchiga and Vázquez-Yanes, 2000; Boyle and Anderson, 2002; Robbins, 2003). However, in spite of the importance of the biology of the germination of seeds in cacti reproduction, the first studies in this regard are dated in 1959 (Rojas-Aréchiga and Vázquez-Yanes, 2000; Godínez-Álvarez et al., 2003).
The actual study is a synthesis of the main results obtained about the germination of the species of the Cactaceae family that are classified in some of the risk categories and that grow in the Chihuahuan Desert. Seed dormancy and the way to beak it, light demand, competition among seeds, and storage time are considered basic factors that affect germination.
Seed dormancy and the way to beak it
Seed dormancy is an adaptive strategy of plants very frequent in hostile and unpredictable environments, such as arid and semi-arid (Jurado and Moles, 2003; Jurado and Flores, 2005). Several cacti have them and many treatments have been tested to break it, such as, acid immersion to simulate their pass through the digestive tube in vertebrates; seed washing to dilute or remove chemical compounds present at the test and that inhibit germination; mechanical scarification (similar to natural abrasion) and stratification periods (over-wintering) and after ripening (Rojas-Aréchiga and Vázquez-Yanes, 2000).
Through several experimental studies the germination of 17 taxa of the Turbinicarpus genus in a risk category at least, and included in the Appendix I of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) was assessed. Seeds were put to germinate at 25 oC with a 12 h light x 12 h darkness photoperiod; the material consisted on seeds collected during the previous 10 months and for five species, others stored during some years. They were placed in Petri boxes with sterile sand; the substrate was selected from preliminary tests in which it was probed that it keeps moisture better that filter paper. Five replications with 20 seeds each and without germination promotion treatments were tested.
Only T. pseudopectinatus (Backeberg) Glass et Foster, T. lophophoroides (Werdermann) Buxbaum et Backeberg y T. jauernigii Frank showed dormancy (Table 1). As time went by, three germination patterns were found: a) seeds loose their viability after one year; b) seeds keep viable and germinate in a similar way for two years and c) as dormancy breaks, germination increases in seeds of 1 to 2 years old (Flores et al., 2005). Recent studies of Opuntia leucotricha DC.
En el Desierto Chihuahuense existe una gran riqueza de especies forestales no maderables, como las cactáceas, muchas de ellas clasificadas en diversas categorías de riesgo debido, entre otras causas, a la recolecta ilegal a la que están sujetas (Hernández y Godínez, 1994; Hernández y Bárcenas, 1995, 1996; Gómez-Hinostrosa y Hernández, 2000; Hernández et al., 2001). Se ha sugerido que su propagación ex situ puede ser un medio para reducir su extracción ilegal (Hernández y Bárcenas, 1995; Rojas-Aréchiga y Vázquez-Yanes, 2000; Boyle y Anderson, 2002; Robbins, 2003). Sin embargo, a pesar de la importancia de la biología de la germinación de semillas en la reproducción de las cactáceas, los estudios pioneros sobre este tema datan de 1959 (Rojas-Aréchiga y Vázquez-Yanes, 2000; Godínez-Álvarez et al., 2003).
El presente trabajo es una síntesis de los principales resultados obtenidos sobre la germinación de especies de la familia Cactaceae clasificada en alguna categoría de riesgo y que se distribuyen en el Desierto Chihuahuense. Se consideran como factores fundamentales la presencia de latencia en las semillas y la forma de romperla, el requerimiento de luz, la competencia entre semillas, el efecto del tiempo de almacenamiento, para germinar y del tamaño de las semillas.
Latencia de semillas y formas de romperla
La latencia de las semillas es una estrategia adaptativa de las plantas muy común en ambientes hostiles e impredecibles, tales como los áridos y semiáridos (Jurado y Moles, 2003; Jurado y Flores 2005). Varias cactáceas la presentan, y se han usado muchos tratamientos para tratar de romperla; por ejemplo, la inmersión en ácido para simular el paso de las semillas por el tracto digestivo de vertebrados, el lavado de las semillas para diluir o remover compuestos químicos existentes en la testa y que inhiben la germinación, la escarificación mecánica (parecida a la abrasión natural) y los períodos de estratificación (over-wintering) y postmaduración (after-ripening) (Rojas-Aréchiga y Vázquez-Yanes, 2000).
En diversas experiencias se evaluó la germinación de 17 taxa del género Turbinicarpus, el cual se caracteriza por tener todas sus especies con alguna categoría de riesgo e incluidas en el Apéndice I de la Convención sobre el Comercio Internacional de Especies Amenazas de Flora y Fauna Silvestres (CITES). Las semillas se pusieron a germinar a 25 °C con fotoperiodo de 12 horas luz por 12 horas de oscuridad; el material consistió en semillas de todos los taxa recolectadas en los 10 meses previos al experimento y semillas almacenadas por varios años para cinco especies. Las semillas se depositaron en cajas Petri con arena estéril; el sustrato se escogió con base en pruebas preliminares en las cuales se demostró que retiene mayor humedad que el papel filtro. Se tuvieron cinco repeticiones, con 20 semillas en cada una y sin pre-tratamientos promotores de la germinación.

61
Flores y Jurado. Germinación de Especies de Cactáceas en Categoría de riesgo...
Solamente T. pseudopectinatus (Backeberg) Glass et Foster, T. lophophoroides (Werdermann) Buxbaum et Backeberg y T. jauernigii Frank (Cuadro 1) presentaron latencia. Con el paso del tiempo se determinaron tres patrones de germinación (Cuadro 2): a) las semillas pierden viabilidad al año, b) las semillas permanecen viables y germinan de manera similar hasta por dos años y c) la germinación aumenta en semillas de 1 y 2 años de edad, al romperse
and O. streptacantha Lem. (with no conservation status) showed that dormancy is broken with time combined with fungi that scarify seed cover (Delgado-Sánchez et al., 2010, 2011). In arid and semi-arid environments, these organisms are not relevant (Porras-Alfaro et al., 2008), but considering the previous results, they should be taken into account. Thus, it would be important to carry out studies in this regard, with different taxa of the Cactacea family.
Cuadro 1. Porcentaje de germinación de las semillas (media ± desviación estándar) de especies del género Turbinicarpus.Table 1. Seed germination per cent (mean ± standard deviation) of Turbinicarpus species.
Especie (%)
T. pseudopectinatus (Backeberg) Glass & Foster 8 ± 5.70T. lophophoroides (Werdermann) Buxbaum & Backeberg 12 ± 2.73T. jauernigii Frank 38 ± 6.70T. swobodae Diers 67 ± 5.70T. laui Glass & Foster 71±11.40T. knuthianus (Boedecker) John & Riha 72±18.50T. vierecki subsp. major (Glass & Foster) Glass 84 ± 4.18T. schmiedickeanus subsp. flaviflorus (Frank & Lau) Glass 86 ± 6.51T. alonsoi Glass & Arias 86 ± 7.40T. rioverdensis (Frank) X T. schmiedickeanus subsp. klinkerianus (Backeber & Jacobsen) Taylor
89 ± 6.51
T. gielsdorfianus (Werdermann) John & Riha 91 ± 6.51T. macrochele var. polaskii Lechner y Jantschgi 92 ± 4.47T. macrochele subsp. frailensis Lechner & Jantschgi 93 ± 5.70T. valdezianus (H. Moeller) Glass & Foster 94 ± 4.18T. schmiedickeanus subsp. macrochele (Werdermann) Taylor 95 ± 3.53T. schmiedickeanus subsp. klinkerianus (Backeber & Jacobsen) Taylor 96 ± 4.18T. schmiedickeanus subsp. schwarzii (Shurly) Taylor 97 ± 2.73
Fuente: Flores et al., 2005.Source: Flores et al., 2005.
Also, putresceines were added to Turbinicarpus, which are polycations of low molecular weight used as germination promoters, without positive results; however, it was confirmed that the dormancy of T. pseudopectinatus and T. lophophoroides is broken with time (Flores et al., 2008).
Photoblastism in cacti
There are three possible germination reactions to light (Baskin y Baskin, 1998): a) species with absolute light demand (positive photoblastism), b) those that germinate in a similar way to light and to darkness (neutral photoblastism); c) those with an absolute darkness demand (negative photoblastism). The following essay describes the effect of light upon the germination of 28 cacti species in risk status. Seeds were put to germinate at 25 oC with two treatments: a) 14 light h/10 darkness h photoperiod and b) germination under total darkness. The material was displayed in Petri boxes with sterile sand. For the darkness treatment, Petri boxes were covered by a two layer aluminum foil; five replications with 20 seeds each were tested, with a green light for darkness. Watering was applied every day.
su latencia (Flores et al., 2005). En estudios recientes con Opuntia leucotricha DC. y O. streptacantha Lem. (sin status de conservación), se determinó que la latencia se rompe con el paso del tiempo en combinación con hongos que escarifican la cubierta de las semillas (Delgado-Sánchez et al., 2010, 2011). En ambientes áridos y semiáridos no se considera que estos organismos sean relevantes (Porras-Alfaro et al., 2008), pero a partir de los resultados anteriores debiesen tomarse en cuenta. Por lo tanto, sería importante realizar estudios sobre el tema, con otros taxa de la familia Cactaceae.
Así mismo, se han aplicado putrescinas en Turbinicarpus, policationes de bajo peso molecular usados como promotores de la germinación, sin resultados positivos; sin embargo se corroboró que la latencia de T. pseudopectinatus y de T. lophophoroides se rompe con el paso del tiempo (Flores et al., 2008).
Fotoblastismo en cactáceas
Existen tres posibles respuestas de germinación por efecto de la luz (Baskin y Baskin, 1998): a) especies con requerimiento
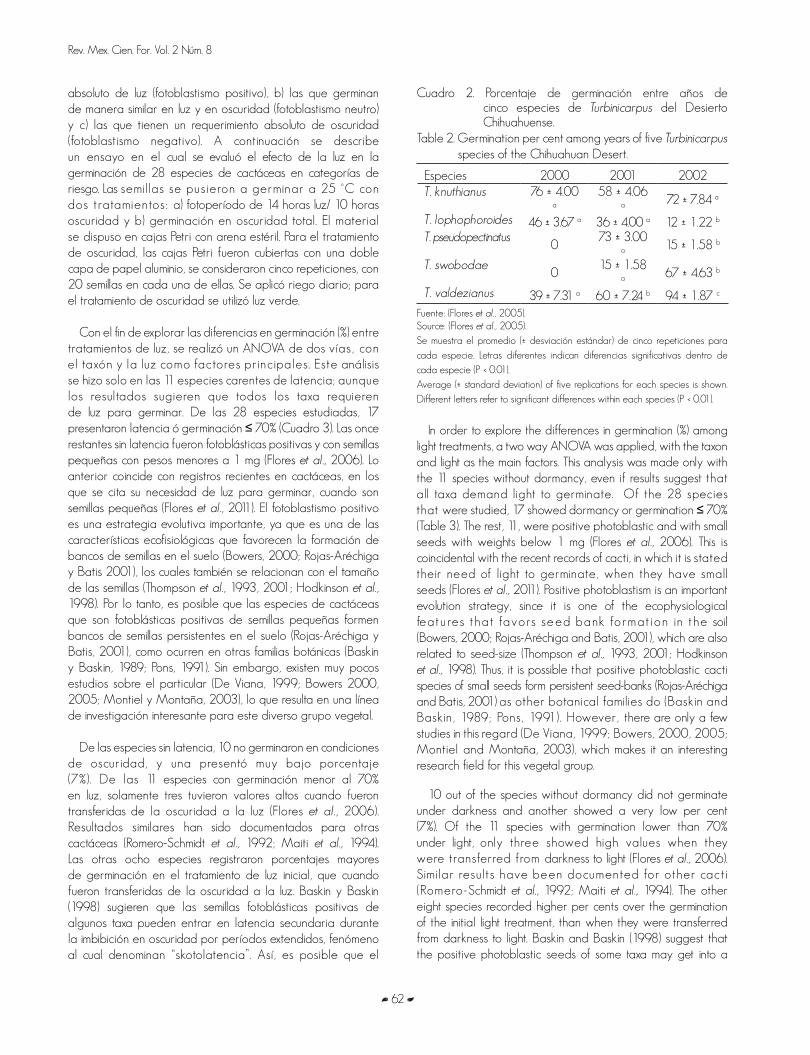
62
Rev. Mex. Cien. For. Vol. 2 Núm. 8
absoluto de luz (fotoblastismo positivo), b) las que germinan de manera similar en luz y en oscuridad (fotoblastismo neutro) y c) las que tienen un requerimiento absoluto de oscuridad (fotoblastismo negativo). A continuación se describe un ensayo en el cual se evaluó el efecto de la luz en la germinación de 28 especies de cactáceas en categorías de riesgo. Las semillas se pusieron a germinar a 25 °C con dos tratamientos: a) fotoperíodo de 14 horas luz/ 10 horas oscuridad y b) germinación en oscuridad total. El material se dispuso en cajas Petri con arena estéril. Para el tratamiento de oscuridad, las cajas Petri fueron cubiertas con una doble capa de papel aluminio, se consideraron cinco repeticiones, con 20 semillas en cada una de ellas. Se aplicó riego diario; para el tratamiento de oscuridad se utilizó luz verde.
Con el fin de explorar las diferencias en germinación (%) entre tratamientos de luz, se realizó un ANOVA de dos vías, con el taxón y la luz como factores principales . Este análisis se hizo solo en las 11 especies carentes de latencia; aunque los resultados sugieren que todos los taxa requieren de luz para germinar. De las 28 especies estudiadas, 17 presentaron latencia ó germinación ≤ 70% (Cuadro 3). Las once restantes sin latencia fueron fotoblásticas positivas y con semillas pequeñas con pesos menores a 1 mg (Flores et al., 2006). Lo anterior coincide con registros recientes en cactáceas, en los que se cita su necesidad de luz para germinar, cuando son semillas pequeñas (Flores et al., 2011). El fotoblastismo positivo es una estrategia evolutiva importante, ya que es una de las características ecofisiológicas que favorecen la formación de bancos de semillas en el suelo (Bowers, 2000; Rojas-Aréchiga y Batis 2001), los cuales también se relacionan con el tamaño de las semillas (Thompson et al., 1993, 2001; Hodkinson et al., 1998). Por lo tanto, es posible que las especies de cactáceas que son fotoblásticas positivas de semillas pequeñas formen bancos de semillas persistentes en el suelo (Rojas-Aréchiga y Batis, 2001), como ocurren en otras familias botánicas (Baskin y Baskin, 1989; Pons, 1991). Sin embargo, existen muy pocos estudios sobre el particular (De Viana, 1999; Bowers 2000, 2005; Montiel y Montaña, 2003), lo que resulta en una línea de investigación interesante para este diverso grupo vegetal.
De las especies sin latencia, 10 no germinaron en condiciones de oscuridad, y una presentó muy bajo porcentaje (7%). De las 11 especies con germinación menor al 70% en luz, solamente tres tuvieron valores altos cuando fueron transferidas de la oscuridad a la luz (Flores et al., 2006). Resultados similares han sido documentados para otras cactáceas (Romero-Schmidt et al., 1992; Maiti et al., 1994). Las otras ocho especies registraron porcentajes mayores de germinación en el tratamiento de luz inicial, que cuando fueron transferidas de la oscuridad a la luz. Baskin y Baskin (1998) sugieren que las semillas fotoblásticas positivas de algunos taxa pueden entrar en latencia secundaria durante la imbibición en oscuridad por períodos extendidos, fenómeno al cual denominan “skotolatencia”. Así, es posible que el
Cuadro 2. Porcentaje de germinación entre años de cinco especies de Turbinicarpus del Desierto Chihuahuense.
Table 2. Germination per cent among years of five Turbinicarpus species of the Chihuahuan Desert.
Especies 2000 2001 2002T. knuthianus 76 ± 4.00
a
58 ± 4.06 a 72 ± 7.84 a
T. lophophoroides 46 ± 3.67 a 36 ± 4.00 a 12 ± 1.22 b
T. pseudopectinatus0
73 ± 3.00 a 15 ± 1.58 b
T. swobodae0
15 ± 1.58 a 67 ± 4.63 b
T. valdezianus 39 ± 7.31 a 60 ± 7.24 b 94 ± 1.87 c
Fuente: (Flores et al., 2005).Source: (Flores et al., 2005).Se muestra el promedio (± desviación estándar) de cinco repeticiones para cada especie. Letras diferentes indican diferencias significativas dentro de cada especie (P < 0.01).Average (± standard deviation) of five replications for each species is shown. Different letters refer to significant differences within each species (P < 0.01).
In order to explore the differences in germination (%) among light treatments, a two way ANOVA was applied, with the taxon and light as the main factors. This analysis was made only with the 11 species without dormancy, even if results suggest that all taxa demand light to germinate. Of the 28 species that were studied, 17 showed dormancy or germination ≤ 70% (Table 3). The rest, 11, were positive photoblastic and with small seeds with weights below 1 mg (Flores et al., 2006). This is coincidental with the recent records of cacti, in which it is stated their need of light to germinate, when they have small seeds (Flores et al., 2011). Positive photoblastism is an important evolution strategy, since it is one of the ecophysiological features that favors se ed bank format ion in the soil (Bowers, 2000; Rojas-Aréchiga and Batis, 2001), which are also related to seed-size (Thompson et al., 1993, 2001; Hodkinson et al., 1998). Thus, it is possible that positive photoblastic cacti species of small seeds form persistent seed-banks (Rojas-Aréchiga and Batis, 2001) as other botanical families do (Baskin and Baskin, 1989; Pons, 1991). However, there are only a few studies in this regard (De Viana, 1999; Bowers, 2000, 2005; Montiel and Montaña, 2003), which makes it an interesting research field for this vegetal group.
10 out of the species without dormancy did not germinate under darkness and another showed a very low per cent (7%). Of the 11 species with germination lower than 70% under light, only three showed high values when they were transferred from darkness to light (Flores et al., 2006). Similar results have been documented for other cacti (Romero-Schmidt et al., 1992; Maiti et al., 1994). The other eight species recorded higher per cents over the germination of the initial light treatment, than when they were transferred from darkness to light. Baskin and Baskin (1998) suggest that the positive photoblastic seeds of some taxa may get into a

63
Flores y Jurado. Germinación de Especies de Cactáceas en Categoría de riesgo...
tratamiento de oscuridad haya provocado la pérdida de fotosensibilidad en las semillas bajo estudio.
Cuadro 3. Porcentaje de germinación de semillas de especies de cactus bajo tratamientos de luz.Table 3. Seed germination per cent of cacti species under light treatments.
Especie Luz OscuridadLuz después de
obscuridadTurbinicarpus schmiedickeanus subsp. macrochele (Werdermann) Taylor* 98.00 a 7.00 b 11.80 b
Mammillaria crinita DC. * 97.00 a 0.00 b 94.00 a
Ariocarpus retusus subsp. trigonus (Weber) Anderson & Maurice* 94.00 a 0.00 b 57.00 c
Turbinicarpus schmiedickeanus subsp. flaviflorus (Frank & Lau) Glass* 86.00 a 0.00 b 6.00 b
Turbinicarpus schmiedickeanus subsp. schwarzii (Shurly) Taylor* 84.00 a 0.00 b 38.00 c
Mammillaria bocasana Poselg.* 83.00 a 0.00 b 41.00 c
Turbinicarpus schmiedickeanus subsp. klinkerianus (Backeber & Jacobsen) Taylor*
80.00 a 0.00 b 8.00 b
Mammillaria longimamma DC.* 75.00 a 0.00 b 35.00 c
Pelecyphora strobiliformis (Werderm.) Fric & Schelle * 74.00 a 0.00 b 84.00 a
Thelocactus conothelos subsp. flavus (Mosco & Zanovelo)* 74.00 a 0.00 b 78.00 a
Turbinicarpus macrochele subsp. frailensis Lechner & Jantschgi* 74.00 a 0.00 b 1.00 b
Turbinicarpus schmiedickeanus subsp. machrochele (Werd.) Taylor var. rubriflorus (Frank) Zachar
69.00 0.00 0.00
Mammillaria orcuttii Boed. 68.00 0.00 34.00Turbinicarpus gielsdorfianus (Werdermann) John & Riha 68.00 0.00 2.00Ariocarpus fissuratus subsp. bravoanus (Hernández & Anderson) 66.00 0.00 45.00Ariocarpus kotschoubeyanus (Lem.) Schum. 56.00 1.00 1.00Ariocarpus retusus subsp. retusus (Scheidw.) 53.00 0.00 10.00Mammillaria plumosa F.A.C. Weber 51.00 0.00 1.00Turbinicarpus laui Glass & Foster 48.00 0.00 3.00Mammilloydia candida (Scheidw.) Buxb. 46.00 0.00 21.00Mammillaria aureilanata Backeberg 42.00 0.00 6.00Obregonia denegrii Fric, Zivot & Prírode 41.00 0.00 4.00Turbinicarpus alonsoi Frank 41.00 0.00 25.00Turbinicarpus jauernigii Frank 37.00 0.00 0.00Ariocarpus fissuratus subsp. hintonii (Stuppy & Taylor) Halda 36.00 5.00 31.00Epithelantha micromeris (Engelm.) Weber ex Britton & Rose 33.00 0.00 4.00Turbinicarpus lophophoroides (Werdermann) Buxbaum & Backeberg 8.00 0.00 1.00Turbinicarpus pseudopectinatus (Backeberg) Glass & Foster 5.00 5.00 5.40
Fuente: Flores et al., 2006.Source: Flores et al., 2006.Letras diferentes muestran diferencias significativas entre tratamientos (P < 0.0001) para las 11 especies que no presentaron latencia (indicadas con asterisco). Las especies marcadas con asterisco se consideran “skotolatentes”. Different letters refer to significant differences within each species (P < 0.0001) for 11 species that did not show dormancy (*); species with * are considered “skotodormant”
Competencia entre semillas
La densidad de semillas en el suelo depende de su forma de dispersión; por ejemplo, los frutos que poseen pulpa son consumidos principalmente por vertebrados frugívoros, quienes dispersan las semillas en grupos al regurgitar o al defecar (Traveset, 1998). La ingesta de semillas promueve la germinación en muchas plantas (Traveset, 1998; Verdú y Traveset, 2004) entre ellas algunas cactáceas (Cortés-Figueira
secondary dormancy during the imbibition in darkness for long period, which is known as “skotodormancy”. Thus, it is possible that the darkness treatment has caused the loss of photosensibility in the studied seeds.
Competition among seeds
Seed density in soil depends on its dispersal; for example, fruits with pulp are eaten, mainly, by frugivore vertebrates, which disperse seeds in groups or by regurgitation or defecation (Traveset, 1998). Seed consumption promotes germination in many plants (Traveset, 1998; Verdú y Traveset, 2004), some of which are cacti (Cortés-Figueira et al., 1994; Pedroni and Sánchez, 1997; Godínez-Álvarez and Valiente-Banuet, 1998;
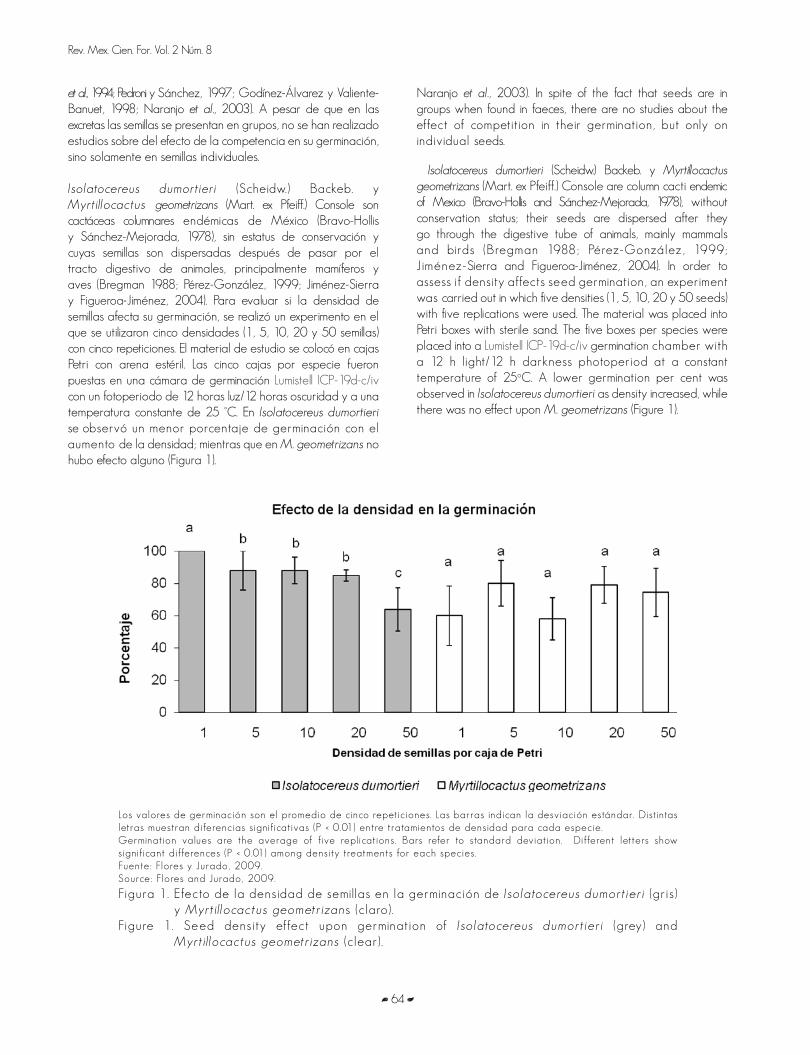
64
Rev. Mex. Cien. For. Vol. 2 Núm. 8
et al., 1994; Pedroni y Sánchez, 1997; Godínez-Álvarez y Valiente-Banuet, 1998; Naranjo et al., 2003). A pesar de que en las excretas las semillas se presentan en grupos, no se han realizado estudios sobre del efecto de la competencia en su germinación, sino solamente en semillas individuales.
Isolatocereus dumortieri (Scheidw.) Backeb. y Myrtillocactus geometrizans (Mart. ex Pfeiff.) Console son cactáceas columnares endémicas de México (Bravo-Hollis y Sánchez-Mejorada, 1978), sin estatus de conservación y cuyas semillas son dispersadas después de pasar por el tracto digestivo de animales, principalmente mamíferos y aves (Bregman 1988; Pérez-González, 1999; Jiménez-Sierra y Figueroa-Jiménez, 2004). Para evaluar si la densidad de semillas afecta su germinación, se realizó un experimento en el que se utilizaron cinco densidades (1, 5, 10, 20 y 50 semillas) con cinco repeticiones. El material de estudio se colocó en cajas Petri con arena estéril. Las cinco cajas por especie fueron puestas en una cámara de germinación Lumistell ICP-19d-c/iv con un fotoperiodo de 12 horas luz/12 horas oscuridad y a una temperatura constante de 25 °C. En Isolatocereus dumortieri se observó un menor porcentaje de germinación con el aumento de la densidad; mientras que en M. geometrizans no hubo efecto alguno (Figura 1).
Naranjo et al., 2003). In spite of the fact that seeds are in groups when found in faeces, there are no studies about the effect of competition in their germination, but only on individual seeds.
Isolatocereus dumortieri (Scheidw.) Backeb. y Myrtillocactus geometrizans (Mart. ex Pfeiff.) Console are column cacti endemic of Mexico (Bravo-Hollis and Sánchez-Mejorada, 1978), without conservation status; their seeds are dispersed after they go through the digestive tube of animals, mainly mammals and birds (Bregman 1988; Pérez-González, 1999; J iménez-Sierra and Figueroa-Jiménez, 2004). In order to assess if density affects seed germination, an experiment was carried out in which five densities (1, 5, 10, 20 y 50 seeds) with five replications were used. The material was placed into Petri boxes with sterile sand. The five boxes per species were placed into a Lumistell ICP-19d-c/iv germination chamber with a 12 h light/12 h darkness photoperiod at a constant temperature of 25oC. A lower germination per cent was observed in Isolatocereus dumortieri as density increased, while there was no effect upon M. geometrizans (Figure 1).
Los valores de germinación son el promedio de cinco repeticiones. Las barras indican la desviación estándar. Dist intas letras muestran diferencias significativas (P < 0.01) entre tratamientos de densidad para cada especie.Germination values are the average of five replications . Bars refer to standard deviation. Different letters show significant differences (P < 0.01) among density treatments for each species . Fuente: Flores y Jurado, 2009.Source: Flores and Jurado, 2009.
Figura 1. Efecto de la densidad de semillas en la germinación de Isolatocereus dumortieri (gris ) y Myrtillocactus geometrizans (claro) .
Figure 1. Seed density effect upon germination of Isolatocereus dumortieri (grey) and Myrtillocactus geometrizans (clear) .

65
Flores y Jurado. Germinación de Especies de Cactáceas en Categoría de riesgo...
These results suggest that I. dumortieri seedlings would compete for resources, or they might be relating to the size of fruits, since in cacti they usually include a greater number of seeds (Ayala-Cordero et al., 2004). Those of M. geometrizans are small (1 - 2 cm diameter) if compared to those of I . dumortieri (3-3.5 cm diameter) (Bravo-Hollis and Sánchez-Mejorada, 1978). However, a dispersal animal could eat only pieces of a big fruit or many small ones, and thus, the effect of the number of seeds per fruit would be neutralized.
Probably the former species have different dispersal agents, which puts one of them at a greater risk of competition with its siblings. It would be expected that the seeds of M. geometrizans would disperse in a single way or in low density, or that it may have a secondary dispersal agent (ants, for example) that removes seeds from those originally grouped in the faeces, while those of I. dumortieri are more frequently disseminate as clump dispersal. Nevertheless, to have a better understanding of the implications of seed density on germination, it is necessary to implement detailed research about seed dispersal of these species (Flores and Jurado, 2009).
It is also possible to explain the differences of the effects of seed density upon germimation from the view point of the competitive capacity (Silvertown and Charlesworth, 2001). I. dumortieri might show a low competition ability (r selection) as its germination diminishes as seed density increases; on the other hand, M. geometrizans has a greater advantage (k selection) as it does not show an effect of seed density on germination.
Effect of seed size on germination
Literature reports that variations on seed weight of the same species reveal different biological abilities. For example, big seeds has a greater ability to germinate than the small ones, when they are buried and later emerge from the ground (Brown et al., 2003), since they have a greater nutriment reservoir (Leishman et al., 2000). Differences of 1 mg imply variations of germination capacity (Baloch et al., 2001).
Astrophytum myriostigma Lem. is an endmic threatened cacti of the Chihuahuan Desert, that grows in different locations of that region; one of them is “El Sarnoso” mountain range in Lerdo municipio, Durango State (Sánchez-Salas and Romero-Méndez, 2003). A. myriostigma individuals are extracted to be used as ornamental plants, which has favored the thinning of their populations and even the total elimination of some populations, which suffered the loss of their habitat from unrestricted collection, in addition to mining. In 2001 there were around 100 individuals with 55% and their germination per cent for seeds immersed in water for 24 h and of 42.5% for seeds with no treatment (Rioja-Paradela and Romero-Méndez, 2002); thus, it can be inferred that the species has seed dormancy as has been recorded for other
Estos resultados sugieren que las plántulas de I . dumortieri estarían compitiendo por recursos, o bien relacionarse con el tamaño de los frutos, pues en cactáceas los más grandes suelen incluir un mayor número de semillas (Ayala-Cordero et al . , 2004) . En ese caso los de M. geometrizans son pequeños (1 a 2 cm de diámetro) en comparación con los de I. dumortieri (de 3 a 3.5 cm de diámetro) (Bravo-Hollis y Sánchez-Mejorada, 1978). Sin embargo, un animal dispersor podría consumir solo porciones de un fruto grande o muchos pequeños, y con ello se neutralizaría el efecto del número de semillas por fruto.
Es probable que las especies estudiadas tengan diferentes agentes dispersores, por lo que una de ellas enfrente mayor riesgo de competencia con plántulas hermanas. Se esperaría que las semillas de M. geometrizans se dispersen de manera solitaria o en baja densidad, o bien que tengan un agente dispersor secundario (por ejemplo hormigas) que separe las semillas originalmente agregadas en las heces; mientras que las de I. dumortieri sean dispersadas de manera agregada con más frecuencia (clump dispersal). No obstante, para entender mejor las implicaciones de la densidad de semillas en la germinación se requiere de investigaciones detalladas sobre su dispersión de las semillas de estas especies (Flores y Jurado, 2009).
También es posible interpretar las diferencias en el efecto de la densidad de semillas en la germinación desde el punto de vista de la capacidad competitiva (Silvertown y Charlesworth, 2001) . Tal vez I . dumortieri presente baja capacidad competitiva (selección r) al disminuir su germinación conforme aumenta la densidad de semillas; en cambio M. geometrizans es más favorecida (selección k) a l no presentar un efecto de la dens idad de semillas en la germinación.
Efecto del tamaño de semillas en la germinación
En la literatura se consigna que las variaciones en peso entre semillas de una misma especie reflejan capacidades biológicas diferentes . Por ejemplo, las semillas grandes t ienen una capacidad superior para germinar que las pequeñas, cuando están enterradas y posteriormente emergen del suelo (Brown et al . , 2003), ya que t ienen mayor reserva de nutrimentos (Leishman et al., 2000). Diferencias de 1 mg implican variaciones en la capacidad germinativa (Baloch et al., 2001).
Astrophytum myriostigma Lem. es una cactácea amenazada y endémica del Desierto Chihuahuense, que se distribuye en distintas localidades de dicha región, entre ellas la sierra “El Sarnoso” en el municipio Lerdo, Durango (Sánchez-Salas y Romero-Méndez, 2003). Los individuos de A. myriostigma son extraídos para su uso como plantas de ornato, lo que ha

66
Rev. Mex. Cien. For. Vol. 2 Núm. 8
originado la disminución de las poblaciones e incluso la pérdida de una población entera, cuyo hábitat desapareció por la recolecta indiscriminada, además de las actividades mineras. En el 2001, existían alrededor de 100 individuos y su porcentaje de germinación era de 55% para semillas sumergidas en agua por 24h y de 42.5% para semillas sin tratamiento (Rioja-Paradela y Romero-Méndez, 2002); por lo que se infiere que la especie posee latencia de semillas como la registrada para otras especies de la familia Cactaceae (Rojas-Aréchiga y Vázquez-Yanes 2000; Flores et al., 2005, 2006).
A partir de un banco de semillas que se conservó ex
situ se obtuvo material de A. myriostigma de cuatro años de almacenamiento; se le sometió a distintos tratamientos promotores de la germinación para conocer su mejor respuesta y determinar así su capacidad de conservar el germoplasma de esta forma
.Lamb (1959), en una clasificación que realizó sobre el
tamaño de semillas de distintas especies de cactáceas consignó que A. myriost igma posee dos tamaños de semilla, por lo que se decidió evaluar su impacto en la germinación. En varios taxa, incluso algunos de la familia Cactaceae, se ha documentado dicho efecto tanto entre especies (Bowers y Pierson, 2001), como dentro de un mismo taxón (Ayala-Cordero et al., 2004).
Las semillas frescas de A. myriostigma germinan bajo distintos tratamientos de manera similar o menor que sin tratamiento alguno (De La Rosa-Ibarra y García, 1994; Arredondo-Gómez y Camacho-Morfín, 1995; Rioja-Paradela y Romero-Méndez, 2002). Sin embargo, no se había evaluado la germinación de semillas “viejas”. El material se sembró en cajas de Petri y se colocó en una germinadora automática Lumistell ICP-19d-c/iv con fotoperiodo de 12 h luz/ 12 h oscuridad y temperatura de 25 °C. Se utilizó una mezcla de vermiculita de grano medio (60%) y arena esterilizada (40%). Se evaluaron el porcentaje y la velocidad de germinación en semillas de dos clases de tamaño, significativamente distintas en longitud (P = 0.003) y peso seco (P < 0.0001), pero no en diámetro. Se aplicaron cinco tratamientos para romper la posible latencia tanto en semillas grandes, como en semillas pequeñas; con cinco repeticiones y 30 semillas por repetición, de tal manera que el total de semillas para cada tamaño fue de 750. Los tratamientos consistieron en: 1) inmersión en H
2SO
4 100%, por
5min; 2) inmersión en agua destilada por 24h; 3) escarificación mecánica, en cuyo caso se retiró con pinzas la testa del embrión, sin dañarlo; 4) enfriamiento, las semillas se mantuvieron a 5 °C por 12 h antes de la siembra y 5) un tratamiento control o testigo, en el cual las semillas se usaron sin ningún tratamiento promotor de la germinación.
En el porcentaje de germinación incidieron los tratamientos, el tamaño de las semillas y la interacción tratamiento x tamaño de semilla. Los mejores tratamientos fueron agua destilada
species of cacti (Rojas-Aréchiga and Vázquez-Yanes 2000; Flores et al., 2005, 2006).
From the seed bank of A. myriostigma that was kept in situ, seeds were taken after four years of storage; they were tested by several germination treatments in order to know their best response and determine its ability to preserve germ plasm in this way.
In the classification Lamb (1959) made of seed size of several cacti species, he found that those of A. myriostigma have two sizes, and so he assessed the impact of that feature on germination. In several taxa, even some of Cactaceae, it has been documented that effect among species as well as in the same taxon (Ayala-Cordero et al., 2004).
Fresh seeds of A. myriostigma germinate in different treatments in a similar way or less than with no treatment at all (De la Rosa-Ibarra and García, 1994; Arredondo-Gómez and Camacho-Morfín, 1995; Rioja-Paradela and Romero-Méndez, 2002). However, germination of “old” seeds had not been assessed. They were sowed in Petri boxes and were placed in an Lumistell ICP-19d-c/iv automatic germination chamber with a 12 h light/12 h darkness photoperiod, at 25 °C. A medium grain vermiculite (60%) and sterilized sand (40%) mixture was used as substrate. Per cent and speed of germination of seeds of two sizes with a significant difference in length (P = 0.003) and dry weight (P < 0.0001), but not in diameter. Five treatments were tested to break the possible dormancy in big and small seeds, with five replications and 30 seeds per replication, in such a way that the total number of seeds for each size was 750. There were the following treatments: 1) a 5 min immersion into 100% H
2SO
4; 2) a 24 h immersion into
distilled water; 3) mechanical scarification, where the test of the embryo was removed by tweezers without harming it; 4) cooling, seeds were kept at 5 °C by 12 h previous to sowing and 5) a control treatment, in which seeds were used with no germination promotion treatment.
Treatments, seed size and the treatment x seed size interaction affected germination per cent. The best treatments were distilled water and cooling. Small seeds showed a greater germination, in regard to control in all cases, except for scarification, in which a low germination was obtained, regardless of size (Table 4).
Size and the treatment x seed size interaction influenced germination speed. Small seeds germinated faster (3.8 seeds day-1) than big ones (1.7 seeds day-1). With the H
2SO
4 treatment, germination speed in small seeds was higher
and with scarification it was higher in the bigger ones (Table 4). The result of small seeds is coincidental with what Kikuzawa and Koyama (1999) found, as they studied the necessary scaling of water absorption for the germination of 14 species of different seed size. These scientists reported that the small ones

67
Flores y Jurado. Germinación de Especies de Cactáceas en Categoría de riesgo...
y enfriamiento. Las semillas pequeñas mostraron mayor germinación, con respecto al testigo en todos los casos, excepto la escarificación, en el que se obtuvo baja germinación, independientemente del tamaño (Cuadro 4).
Cuadro 4. Promedio del porcentaje y tiempo medio ó velocidad germinación (± desviación estándar) de semillas grandes y pequeñas de A. myriostigma bajo diferentes tratamientos.
Table 4. Per cent average and mean time or germination speed (±standard deviation) of big and small A. myriostigma seeds under different treatments.
Tratamiento Semillas grandes Semillas pequeñasGerminación (%) Velocidad (días) Germinación (%) Velocidad (días)
H2SO
420.67 a ± 5.67 0.78 a ± 0.42 92.00 b ± 4.29 4.80b ± 0.28
Agua destilada 42.67 a ± 3.55 1.77 a ± 0.29 96.00 b ± 2.44 4.39a ±0.42Escarificación mecánica 22.00 a ± 4.78 3.14 a ± 0.71 12.67 a ± 5.61 0.81b ±0.33Escarificación 43.33 a ± 3.80 1.32 a ± 0.28 92.67 b ± 3.55 4.29a ±0.39Testigo 58.00 a ± 0.81 1.25 a ± 0.06 90.00 b ± 2.78 4.60a ± 0.20
Fuente: (Sánchez-Salas et al., 2006).Source: (Sánchez-Salas et al., 2006).Diferentes letras dentro de cada tratamiento significan diferencias significativas (P < 0.0001) entre tamaño de semillas.Different letters show significant differences (P < 0.0001) among density treatments for each species among seed size.
absorbed water faster than the big ones and that get into small cracks on the soil surface, which lets them get to a microsite where moisture is kept and prevents from loosing water.
El tamaño y la interacción tratamiento × tamaño de semilla afectó la velocidad de germinación. Las semillas pequeñas germinaron más rápido (3.8 semillas día-1) que las grandes (1.7 semillas día-1). Con el tratamiento de H
2SO
4 la
velocidad de germinación en semillas pequeñas fue más alta y con la escarificación, fue mayor en las semillas grandes (Cuadro 4). El resultado de las semillas pequeñas coincide con el consignado por Kikuzawa y Koyama (1999), quienes estudiaron el escalamiento en la absorción de agua necesario para la germinación de 14 especies con distintos tamaños de semilla. Estos investigadores registraron que las pequeñas absorbieron agua más rápidamente que las grandes y que penetran a través de pequeñas grietas en la superficie del suelo, lo que les permite acceder a un micrositio en donde se mantiene la humedad y previene la pérdida de agua.
Las semillas grandes de A. myriostigma, de 4 años de edad, presentaron un porcentaje de germinación parecido al del material de recolecta reciente de la misma población, aunque de tamaño no especificado (Rioja-Paradela y Romero-Méndez, 2002). Sin embargo, las pequeñas de 4 años mostraron una germinación mayor. Así, es probable que algunas de estas tengan embriones inmaduros, cuando jóvenes y que alcancen su madurez con el paso del tiempo. También se ha observado en algunas cactáceas mayor germinación en semillas de 1 año o más, que en semillas recién obtenidas; así mismo hay otras cuya capacidad germinativa es semejante a la de semillas frescas y con semillas de 1 o 2 años de edad (Flores et al., 2005; 2008), esto sugiere que pueden formar bancos de semillas en el suelo. En conclusión, en las semillas pequeñas de cuatro años de edad se determinaron porcentajes y velocidades de germinación superiores en comparación con las más grandes, lo que podría estar relacionado con su capacidad para absorber agua más rápidamente, o bien tratarse de una estrategia para permanecer en el banco de semillas del suelo por varios años.
A. myriostigma big seeds of four-years old showed a per cent of germination similar to the material of recent collection from the same population, even if size was not specified (Rioja-Paradela and Romero-Méndez, 2002). However, the small ones of four years showed a higher germination. Thus, probably some of these have immature embryos when they are young and that reach their maturity with time. It has also been observed in some cacti of 1 year old or elder than in seeds recently collected; also, there are others whose germination ability is similar to that of fresh ones or with 1 to 2 year-old seeds (Flores et al., 2005; 2008), which suggests that they might form seed banks in the soil. Finally, in small four year-old seeds germination per cents and speeds were determined as higher than those of big seeds, a fact that might be related to their ability to absorb water faster, or as a strategy to stay in the seed bank of the soil for several years.
CONCLUSIONS
Three out of the 17 taxa of the Turbinicarpus have dormancy.
Seeds of two Opuntia species lose their dormancy with time, combined with fungi that scarify their cover.
In 17 out of the 28 species with conservation status of the Ariocarpus, Epithelantha, Mammillaria, Mammilloydia, Obregonia, Pelecyphora, Thelocactus and Turbinicarpus genus dormancy was found (germination ≤ 70%), and the rest (11) were positive photoblastic and with small seeds (< 1 mg), that most probably form a natural germ plasm bank.
Three germination patterns were defined as time went by: a) seeds that loose their viability after the first year; b) seeds keep viable and germinate in a similar way for two years and c) germination increases in 1 to 2 year-old seeds when dormancy is broken.

68
Rev. Mex. Cien. For. Vol. 2 Núm. 8
CONCLUSIONES
Tres de 17 taxa del género Turbinicarpus tienen latencia.
Las semillas de dos especies de Opuntia pierden latencia con el paso del tiempo, en combinación con la presencia de hongos que escarifican su cubierta.
De 28 especies con estatus de conservación de los géneros Ariocarpus, Epithelantha, Mammillaria, Mammilloydia, Obregonia, Pelecyphora, Thelocactus y Turbinicarpus en 17 se observó latencia (germinación ≤ 70%), y las once restantes fueron fotoblásticas positivas y de semillas pequeñas, con peso menor a 1 mg, que muy probablemente forman un banco de germoplasma natural.
Se determinaron tres patrones de germinación con el paso del tiempo: a) las semillas pierden viabilidad al año, b) las semillas permanecen viables y germinan de manera similar hasta por dos años y c) la germinación aumenta en semillas de 1 y 2 años de edad al romperse la latencia.
Isolatocereus dumortieri mostró menor porcentaje de germinación con el aumento de la densidad; mientras que el de M. geometrizans no fue afectado por la densidad de semillas.
Las semillas pequeñas de cuatro años de edad de Astrophytum myriostigma, cactácea amenazada de extinción, tuvieron porcentajes y velocidades de germinación mayores en comparación con las grandes.
AGRADECIMIENTOS
Esta investigación fue financiada por CONAFOR 9769 y por apoyos complementarios que se obtuvieron entre los que destacan: PAICYT (de la UANL); PIFI en estancias de investigación, Millenium Seed Bank (organismo perteneciente a Kew Botanical Gardens) y SEMARNAT 2006-23818. Estudiantes, colegas y personal de apoyo facilitaron esta investigación con invaluables contribuciones de trabajo en campo y laboratorio.
REFERENCIAS
Arredondo-Gómez, A. y F. Camacho-Morfín. 1995. Germinación de Astrophytum myriostigma (Lemaire) en relación con la procedencia de las semillas y la temperatura de incubación. Cactáceas y Suculentas Mexicanas 40: 34-38.
Ayala-Cordero, G., T. Terrazas, L. López-Mata y C. Trejo. 2004. Variación en el tamaño y peso de la semilla y su relación con la germinación en una población de Stenocereus beneckei. Interciencia 29: 692-697.
Baloch, H. A, A. DiTomasso and A. K. Watson. 2001. Intrapopulation variation in Abutilon theophrasti seed mass and its relationship to seed germinability. Seed Science Research 11: 335-343.
Baskin, J. M. and C. C. Baskin. 1989. Physiology of dormancy and germination in relation to seed bank ecology. In: M. A. Leck, V. T. Parker and R. L. Simpson (Eds.). Ecology of Soil Seed Banks. Academic Press. San Diego, CA. USA. pp: 53-66.
Baskin, J. M. and C. C. Baskin. 1998. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination. Academic Press. San Diego, CA. USA. 666 p.
Isolatocereus dumortieri showed a lower germination per cent as density increased, while was not affected by seed density.
Four-year old small seeds of Astrophytum myriostigma, a threatened cactus, had higher germination per cents and speeds than those that are bigger in size.
AKNOWLEDGEMENTS
This investigation was sponsored by CONAFOR Project No. 9769 and by complementary supports such as PAICYT (of the UANL); research stay in PIFI, Millenium Seed Bank (organization of the Kew Botanical Gardens) and SEMARNAT 2006-23818. Students, colleagues, and support personnel helped to accomplish this study by providing their invaluable help in field and laboratory work.
End of the English version
Boyle, T. H. and E. F. Anderson. 2002. Biodiversity and conservation. In: Nobel P. S. (Ed.). Cacti: Biology and Uses. University of California Press. Berkeley, CA. USA. pp. 125-141.
Bowers, J. E. 2005. New evidence for persistent or transient seed banks in three Sonoran Desert cacti. Southwestern Naturalist 50: 482-487.
Bowers, J. E. and E. A. Pierson. 2001. Implications of seed size for seedling survival in Carnegiea gigantea and Ferocactus wislizeni (Cactaceae). Southwestern Naturalist 46: 272-281.
Bravo-Hollis, H. y H. Sánchez-Mejorada. 1978. Las cactáceas de México, Vol. I. Universidad Nacional Autónoma de México. México, D.F. México. 743 p.
Bregman, R. 1988. Forms of seed dispersal in Cactaceae. Acta Botanica Neerlandica 37: 395-402.
Brown, J., N. J. Enright and B. P. Miller. 2003. Seed production and germination in two rare and three common co-occurring Acacia species from southeast Australia. Austral Ecology 28: 271-280.
Cortés-Figueira, J. E., J. Vasconcellos-Neto, M. A. García and A. L. Teixeira de Souza. 1994. Saurocory in Melocactus violaceus (Cactaceae). Biotropica 26: 295-301.
De la Rosa-Ibarra, M. y H. García. 1994. Estimulación de la germinación de cinco especies de cactáceas consideradas en peligro de extinción. Phyton-International Journal of Experimental Botany 56: 147-150.
Delgado-Sánchez, P., M. A. Ortega-Amaro, J. F. Jimenez-Bremont and J. Flores. 2011. Are fungi important for breaking seed dormancy in desert species? Experimental evidence in Opuntia streptacantha (Cactaceae). Plant Biology 13: 154-159.
Delgado-Sánchez, P., M. A. Ortega-Amaro, A. A. Rodríguez-Hernández, J. F. Jiménez-Bremont and J. Flores. 2010. Further evidence from the effect of fungi on breaking Opuntia seed dormancy. Plant Signaling & Behavior 5(10): 1229-1230.
De Viana, M. L. 1999. Seed production and seed bank of Trichocereus pasacana (Cactaceae) in northwestern Argentina. Tropical Ecology 40: 79-84.
Flores, J., A. Arredondo and E. Jurado. 2005. Comparative seed germination in species of Turbinicarpus: An endangered cacti genus. Natural Areas Journal 25: 183-187.
Flores, J., E. Jurado and A. Arredondo. 2006. Effect of light on germination of seeds of Cactaceae from the Chihuahuan Desert, México. Seed Science Research 16: 149-155.
Flores, J., E. Jurado and J. F. Jiménez-Bremont. 2008. Breaking seed dormancy in specially protected Turbinicarpus lophophoroides and Turbinicarpus pseudopectinatus (Cactaceae). Plant Species Biology 23: 44-47.
Flores, J. y E. Jurado. 2009. Efecto de la densidad de semillas en la germinación de Isolatocereus dumortieri y Myrtillocactus geometrizans, cactáceas columnares endémicas de México. Revista Mexicana de Biodiversidad 80: 141-144.

69
Flores y Jurado. Germinación de Especies de Cactáceas en Categoría de riesgo...
Flores, J., E. Jurado, L. Chapa-Vargas, A. Ceroni-Stuva, P. Dávila-Aranda, G. Galíndez, D. Gurvich , P . León-Lobos , C. Ordóñez, P . Ortega-Baes, N. Ramírez-Bullón, A. Sandoval, C.E. Seal, T. Ulian and H.W. Pritchard. 2011. Seeds photoblastism and its relationship with some plant traits in 136 cacti species. Environmental and Experimental Botany 71: 79-88.
Godínez-Álvarez, H., T. Valverde and P. Ortega-Baes. 2003. Demographic trends in the Cactaceae. Botanical Review 69, 173-203.
Gómez-Hinostrosa, C. and H. M. Hernández. 2000. Diversity, geographic distribution, and conservation of Cactaceae in the Mier y Noriega region, Mexico. Biodiversity & Conservation 9: 403-418.
Hernández, H. M., C. Gómez-Hinostrosa and R. T. Bárcenas. 2001. Diversity, spatial arrangement, and endemism of Cactaceae in the Huizache area, a hot spot in the Chihuahuan Desert. Biodiversity & Conservation 10: 1097-1112.
Hernández, H. M. and R. T. Bárcenas.1995. Endangered cacti in the Chihuahuan Desert. I. Distribution patterns. Conservation Biology 9: 1176-1188.
Hernández, H. M. and R. T. Bárcenas. 1996. Endangered cacti in the Chihuahuan Desert. II. Biogeography and conservation. Conservation Biology 10: 1200-1209.
Hernández, H. M. y H. Godínez. 1994. Contribución al conocimiento de las cactáceas mexicanas amenazadas. Acta Botánica Mexicana 26: 33-52.
Hodkinson, D. J., A. P. Askew, K. Thompson, J. G. Hodgson, J. P. Bakker and R. M. Bakker. 1998. Ecological correlates of seed size in the British flora. Functional Ecology 12: 762-766.
Jiménez-Sierra, C. y L. Figueroa-Jiménez. 2004. Isolatocereus dumortieri (Scheidw.) Backeb. (Stenocereus dumortieri). Cactáceas y Suculentas Mexicanas 49: 96.
Jurado, E. and J. Flores. 2005. Is seed dormancy under environmental control or bound to plant traits? Journal of Vegetation Science 16: 559-564.
Jurado, E. and A. Moles. 2003. Germination deferment strategies. In: Nicolas, G., N. K. J. Bradford, D. Come, and H. W. Pritchard (Eds.). The Biology of Seeds: Recent Research Advances. CAB International. Wallingford, UK. pp: 381-388.
Kikuzawa, K. and H. Koyama. 1999. Scaling of soil water absorption by seeds: an experiment using seed analogues. Seed Science Research 9: 171-178.
Lamb, E. 1959. Cacti from seed - the easy way. Bradford Press. London, UK. 64 p.
Leishman, M. R., I. J. Wright, A.T. Moles, and M. Westoby. 2000. The Evolutionary Ecology of Seed Size. In: Fenner, M. (Ed.). Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed. Cabi. Wallingford, UK. pp. 31-57.
Maiti,R. K., J. L. Hernández-Piñero and M. Valdés-Marroquín. 1994. Seed ultrastructure and germination of some species of Cactaceae. Phyton - International Journal of Experimental Botany 55: 97-105.
Montiel, S. and C. Montaña. 2003. Seed bank dynamics of the desert cactus Opuntia rastrera in two habitats from the Chihuahuan Desert. Plant Ecology 166: 241-248.
Naranjo, M. E., C. Rengifo and P. J. Soriano. 2003. Effect of ingestion by bats and birds on seed germination of Stenocereus griseus and Subpilocereus repandus (Cactaceae). Journal of Tropical Ecology 19: 19-25.
Pedroni, F. and M. Sánchez. 1997. Seed dispersal of Pereskia aculeata Muller (Cactaceae) in a forest fragment in southeast Brazil. Revista Brasileira do Biologia 57: 479-486.
Pérez-G., S. 1999. Estudio etnobotánico, ecológico y de usos potenciales del garambullo (Myrtillocactus geometrizans) como base para su domesticación y cultivo. Cuaderno de Trabajo, Sistema de Investigación Miguel Hidalgo, CONACYT. Querétaro, Qro. México. 31 p.
Pons, T. L. 1991. Induction of dark dormancy in seeds: its importance for the seed bank in the soil. Functional Ecology 5: 669-675.
Porras-A., A., J. Herrera, R. L. Sinsabaugh, K. J. Odenbach, T. Lowrey and D. O. Natvig. 2008. Novel root fungal consortium associated with a dominant desert grass. Applied and Environmental Microbiology 74: 2805-2813.
Rioja-P. T. M. y U. Romero-M. 2002. Efecto del remojo sobre el potencial reproductivo sexual de Astrophytum myriostigma Lem. (Cactaceae) en condiciones controladas. Nakari 13: 21-34.
Robbins, C. S. 2003. Prickly trade: trade and conservation of Chihuahuan desert cacti, In: Robbins, C. S. and R. T. Bárcenas-Luna (Eds.). TRAFFIC North America. World Wildlife Fund. Washington, DC., USA. pp: 1-48.
Rojas-A., M. y A. Batis. 2001. Las semillas de cactáceas ¿Forman bancos en el suelo? Cactáceas y Suculentas Mexicanas 46: 76-82.
Rojas-A., M. and C. Vázquez-Y. 2000. Cactus seed germination: a review. Journal of Arid Environments 44: 85-104.
Romero-S., H. L., F. Vega-V., H. Nolasco and C. Montaño. 1992. The effect of darkness, freezing, acidity and salinity on seed germination of Ferocactus peninsulae (Cactaceae). Journal of Arid Environments 23: 389-395.
Sánchez-S., J. y U. Romero-M. 2003. Listado de la flora cactológica de la Sierra El Sarnoso, Durango, México. Nakari 14: 27-36.
Sánchez-S., J., J. Flores y E. Martínez-G. 2006. Efecto del tamaño de semilla en la germinación de Astrophytum myriostigma Lemaire. (Cactaceae), especie amenazada de extinción. Interciencia 31: 371-375.
Silvertown and Charlesworth, 2001. p. 7Thompson, K., S. R. Band and J. G. Hodgson. 1993. Seed size and shape
predict persistence in soil. Functional Ecology 7: 236-241.Thompson, K., A. Jalili, J. G. Hodgson, B. Hamzeh’ee, Y. Asri, S. Shaw, A. Shirvany,
S. Yazdani, M. Khoshnevis, F. Zarrinkamar, M. Ghahramani and R. Safavi. 2001. Seed size, shape and persistence in the soil in an Iranian flora. Seed Science Research 11: 345-355.
Traveset, A. 1998. Effect of seed passage through vertebrate frugivores´guts on germination: a review. Perspectives in Plant Ecology, Evolution and Systematics 1(2): 151-190.
Verdú, M. and A. Traveset. 2004. Bridging meta-analysis and the comparative method: a test of seed size effect on germination after frugivores’ gut passage. Oecologia 138: 414-418.

70
Rev. Mex. Cien. For. Vol. 2 Núm. 8
Wangari Muta Maathai. Dominio público.

NOTA DE INVESTIGACIÓN
LOS SUELOS DEL ESTADO DE MÉXICO Y SU ACTUALIZACIÓN A LA BASE REFERENCIAL MUNDIAL DEL RECURSO SUELO 2006
THE SOILS OF MEXICO STATE AND THEIR UPDATE TO THE WORLD REFERENCE BASEFOR SOIL RESOURCES 2006
Erasto Domingo Sotelo Ruíz 1, Antonio González Hernández2, Gustavo Cruz Bello3, Francisco Moreno Sánchez2 y Gustavo Cruz Cárdenas4
RESUMEN
El suelo es uno de los principales recursos naturales, en el cual se sustentan los bosques y la producción de alimentos para una población que crece cada día. Contar con información actualizada sobre este recurso y su distribución facilita la planeación y clasificación de las actividades agropecuarias y de manejo de los bosques. La clasificación de suelos de la Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO) denominada Base Referencial Mundial del Recurso Suelo (WRB), ha sido mejorada con ello se aumentó su precisión. Los objetivos de esta investigación fueron actualizar la clasificación de la cartografía del Instituto Nacional de Estadística y Geografía escala 1:50,000 de los suelos del Estado de México de la clasificación FAO 1970, a la versión de suelos de la WRB 2006; además de cuantificar la superficie de las Unidades y Subunidades de sus suelos y delimitar su distribución. Los suelos dominantes en el estado son los Andosoles con 4,799.08 km2, Feozems con 4,727.18, Regosoles con 2,656.83, Vertisoles con 2,414.85 y Cambisoles con 1,960.47 km2. Se agregaron los Leptosoles que agruparon a los Rankers, Rendzinas y Litosoles; estos suelos se denominan como Leptosoles Líticos (LPli), los cuales cubren 1,749.68 km2 y tienen una profundidad menor a 10 cm. Con ello se reduce el área para la producción de cultivos.
Palabras clave: Andosoles, cartografía de suelos, Leptosoles, planeación agropecuaria y forestal, recursos naturales, unidades de suelo.
ABSTRACT
Soil is one of the main natural resources, in which support forests and food production for an ever growing population. To have present-day information about this resource and its distribution help to plan and classify agricultural and game activities as well as forest management. The United Nations Food and Agriculture Organization (FAO) soil classification known as World Reference Base for Soil Resources (WRB) has been improved, and thus, it has increased its precision. The aim of this research was to update the FAO 1970 1:50,000 cartography of the National Statistics and Geography Institute of Mexico State to the WRB 2006 version, as well as to quantify the area of the Units and Subunits of its soils and to fix the boundaries of its distribution. The dominant soils in the State are Andosols with 4,799.08 km2, Feozems with 4,727.18, Regosols with 2,656.83, Vertisols with 2,414.85 and Cambisols with 1,960.47 km2. Leptosols were added, which included Rankers, Rendzinas and Litosols, and were named as Liptic Leptosols (LPli), which cover 1,749.68 km2 and are less than 10 cm deep. With this support, the area for crop production is reduced.
Key words: Andosols, soil cartography, Leptosols, agriculture and livestock planning, natural resources, soil units.
Fecha de recepción: 30 de abril de 2009.Fecha de aceptación: 23 de diciembre de 2011.
1Campo Experimental Toluca, CIR.- Centro, INIFAP. Correo-e: [email protected] Nacional de Investigación Disciplinaria en Conservación y Mejoramiento de Ecosistemas Forestales, INIFAP.3ExInvestigador del CENID-COMEF, INIFAP
4Génesis y Clasificación de Suelos, Colegio de Postgraduados.

72
Rev. Mex. Cien. For. Vol. 2 Núm. 8
Soil is one of the most important natural resources for man because of the direct relation with the development of forests, food production and fibers (Ortiz et al., 1990; Ortiz and Gutiérrez, 1999). Natural resources studies facilitate through storing, analysis and reproduction of computerized data. This system is good when the country or the organization that carries out the assessment have the right computation equipment to keep data (FAO, 1985).
There are different definitions of soils, according to their approach, based upon several properties: physical, chemical, nutritional, microbiological, of fertility, genesis and classification. In the actual work are taken into account genesis and classification of soils. Thus, among them can be quoted those that consider soil as the continuum of space and time that make up part of the terrestrial cortex, with the thickness of a few centimeters and where the roots of plants develop (FitzPatrick, 1987, 1996). For Boul et al. (2004), they are natural bodies that show their characteristics in a three-dimensional way, while for Brady and Weil (1999), soil is a natural three-dimensional body in the same sense as a mountain, a lake or a valley.
The following definitions are more complete and have the aim of classifying soils according to the Taxonomy of Soils and the United Nations Food and Agriculture Organization (FAO) or the World Reference Base for Soil Resources (WRB). The Soil Survey Staff (1992; 1994) states that soil is a collection of natural bodies, formed from mineral and organic materials, that cover great part of the terrestrial surface, have live mater and can support vegetation in a natural way and, in some places, have been transformed by human activities. The word “Soil”, as many others, has different meanings. In a traditional context, “it is the natural means for plant growth, and it may have or not, discernible horizons”.
Soil is a natural body formed by solids (minerals and organic matter), liquids and gases that occur over the surface of the Earth, that have a place in space and that have horizons or layer, that are distinguished from the original material as they result from additions, losses, transferences and transformation of energy and matter; also, they have the ability to support plant roots in a natural environment (Soil Survey Staff, 1998; Soil Survey Staff, 1999; Soil Survey Staff, 2003; Soil Survey Staff, 2006; Wilding, 2000).
FAO-WRB Classification defines soils the vertical combination of the present horizons in a specific depth and by the lateral organization (sequence) of horizons or by their absence at a scale that is at a scale that reflects in relief or that conceptualize it as a continuous natural body which has three special and temporal dimensions. The three most important properties of soil are: 1) they are formed by minerals and organic elements and include a solid, liquid and gaseous phase; 2) materials are organized into specific structures in the
El suelo es uno de los recursos naturales más significativos en la vida del hombre por su relación directa con el desarrollo de los bosques, la producción de alimentos y fibras (Ortiz et al . , 1990; Ortiz y Gutiérrez, 1999). Los estudios de los recursos naturales se facilitan mediante el almacenamiento, el análisis y la reproducción de datos computarizados. Este sistema ofrece ventajas cuando el país o la organización que realiza la evaluación poseen equipo de cómputo necesario para el almacenamiento de datos (FAO, 1985).
Existen diferentes definiciones de los suelos, en función del enfoque, basadas en diversas propiedades: físicas, químicas, de nutrición, microbiológicas, de fertilidad, de génesis y de clasificación. En el presente trabajo, se toman en cuenta la génesis y la clasificación de suelos. Así, entre ellas se pueden citar aquellas que considera al suelo como el continuo de espacio-tiempo que forma la parte superior de la corteza terrestre, con un espesor de pocos centímetros y donde se desarrollan las raíces de las plantas (Fitzpatrick, 1987; Fitzpatrick, 1996). Para Boul et al. (2004) son cuerpos naturales que exhiben sus características en forma tridimensional; en tanto que, para Brady y Weil (1999) el suelo es un cuerpo natural tridimensional en el mismo sentido que es una montaña, lago o valle.
Las siguientes definiciones son más completas y tienen el propósito de clasificar a los suelos según la Taxonomía de suelos y la Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO) o la Base Referencial Mundial del Recurso Suelo (WRB). El Soil Survey Staff (1992; 1994) establecen que el suelo es una colección de cuerpos naturales, formados a partir de materiales minerales y orgánicos, que cubren gran parte de la superficie terrestre, contienen materia viva y pueden soportar vegetación en forma natural y, en algunos lugares, han sido transformados por la actividad humana. La palabra Suelo, como otras, tienen diversos significados. En sentido tradicional “es el medio natural para el crecimiento de las plantas, puede tener o no horizontes discernibles”.
El suelo es un cuerpo natural integrado por sólidos (minerales y materia orgánica), líquidos y gases que ocurren sobre la superficie de la tierra, ocupan un lugar en el espacio y se caracterizan por tener horizontes o capas, que se distinguen del material inicial por ser el resultado de adiciones, pérdidas, transferencias y transformación de energía y materia; además poseen la habilidad de soportar raíces de plantas en un ambiente natural (Soil Survey Staff, 1998; Soil Survey Staff, 1999; Soil Survey Staff, 2003; Soil Survey Staff, 2006; Wilding, 2000).
La Clasificación de la FAO-WRB define a los suelos como la combinación vertical de horizontes presentes dentro de una profundidad específica y por la organización lateral

73
Sotelo et al., Los suelos del Estado de México y su actualización a la base referencial...
(secuencia) de horizontes o por su ausencia, a una escala que se refleja en el relieve o lo conceptualizan como un cuerpo natural continuo, el cual tiene tres dimensiones espaciales y una temporal. Las tres principales propiedades de un suelo son: 1) están formados por minerales y elementos orgánicos e incluye las fases sólida, líquida y gaseosa; 2) los materiales están organizados en estructuras específicas en el medio pedológico. Estas forman el aspecto morfológico de la cobertura del suelo, equivalente a la anatomía de un ser vivo. Su estudio facilita el entendimiento de las propiedades físicas, químicas y biológicas, que, a su vez, permite explicar el pasado, presente y futuro del suelo y 3) el suelo está en constante evolución, tiene una cuarta dimensión: el tiempo.
Por consiguiente, los objetivos de la clasificación WRB son identificar y clasificar cualquier material, hasta una profundidad de 2 m de la superficie de la tierra, que está en contacto con la atmósfera, a excepción de los organismos vivos, las áreas con hielos continuos no cubiertos por otro material y los cuerpos de agua más profundos de 2 m. La definición incluye roca continua, suelos con pavimento urbano, suelos de áreas industriales, de cuevas, así como los suelos subaquicos (IUSS Working Group WRB, 2006).
Clasificación de suelos
Los principales sistemas de clasificación que se utilizan en el mundo son la Taxonomía (Soil Survey Staff, 2006) y el Sistema de la FAO-WRB (IUSS Working Group WRB, 2006), cuyos criterios son:
1) Contenido de materia orgánica y arcilla2) Horizontes superficiales o epipedones3) Horizontes subsuperficiales4) Otras características de diagnóstico5) Régimen de humedad del suelo6) Régimen de temperatura del suelo.
Estos determinan el nombre de los suelos a nivel de Suborden, hacia las demás categorías inferiores en la Taxonomía y a Nivel de Subunidad en la WRB (IUSS Working Group WRB, 2006; Soil Survey Staff, 2006). Sin embargo, la WRB (2006) no contempla el clima para la clasificación de las unidades edáficas, esto es, no incluye los puntos cinco y seis.
Taxonomía de suelos. Es un sistema de clasificación morfogenético, que consiste en claves para clasificar a los suelos del mundo; su primera versión la publicó el Departamento de Agricultura de los Estados Unidos en 1960. Consta de seis categorías de mayor a menor jerarquía: Orden, Suborden, Gran grupo, Subgrupo, Familia y Serie (Soil Survey Staff, 1999; Soil Survey Staff, 2006).
Sistema FAO-WRB. Consta de dos categorías: Unidad y Subunidad. Este sistema se inicia en 1974, con la publicación
pedologic media. They form the morphology aspect of soil cover, which is equivalent to the anatomy of a living organism. Their study makes it easier to understand the physical, chemical and biological properties which lets to know the past, present and future of soil, and 3) soil is continuously evolving, as it has a fourth dimension: time.
Thus, the aims of the WRB classification are to identify and classify any material up to 2 m deep at the surface of earth, that is in contact with the atmosphere, except for living organisms, areas with permanent cover of ice not covered by other material and water bodies deeper than 2 m. The definition includes continuous rock, soils with urban pavement, soils of industrial areas, of caves, as well as underwater soils (IUSS Working Group WRB, 2006).
Soil Classification
The main classification systems that are used in the world are Taxonomy (Soil Survey Staff, 2006) and the FAO-WRB System (IUSS Working Group WRB, 2006), with the following criteria:
1) Organic matter and loam content 2) Superficial horizons or epipedons3) Subsuperficial horizons 4) Other diagnosis characteristics 5) Soil moisture regime6) Soil temperature regime
They determine the name of the soils at the Suborder level, towards the rest of the lower Taxonomy categories and at the Subunit Level of WRB (IUSS Working Group WRB, 2006; Soil Survey Staff, 2006). +However, WRB (2006) does not consider the weather for the classification of the edaphic units, that is, it does not include points 5 and 6.
Soil Taxonomy. It is a morphogenetic classification system, with keys to classify the soils of the world; the first edition was published in 1960 by the United States Department of Agriculture. It includes 6 categories from a major to a minor hierarchy: Order, Suborder, Great group, Subgroup, Family and Series (Soil Survey Staff, 1999; Soil Survey Staff, 2006).
FAO-WRB System. It has two categories: Unit and Subunit. This system was started in 1974, with the FAO/UNESCO (1974) Legend of the Soil Maps of the World and their units do not mach exactly with the categories of other systems, but in general, they are comparable in “great groups” with the soil taxonomy of the Soil Survey Staff.
WRB aim is to develop an international system good enough to describe soil, which can link and relate with national classifications by FAO Reviewed Legend as a basic structure; to provide a solid scientific base to this structure to be used

74
Rev. Mex. Cien. For. Vol. 2 Núm. 8
de la Leyenda del Mapa de suelos del Mundo (FAO/UNESCO, 1974) y sus unidades no corresponden exactamente con las categorías de otros sistemas, pero en general son comparables, en “grandes grupos” con la taxonomía de suelos del Soil Survey Staff.
La WRB tiene como objetivos desarrollar un sistema internacional aceptable para describir al suelo, el cual pueda vincularse y relacionarse con las clasificaciones nacionales, mediante la Leyenda Revisada de FAO como estructura básica; proporcionar una base científica sólida a esta estructura, para su uso en diferentes áreas: manejo de bosques, agricultura, geología, hidrología y ecología; reconocer dentro de la estructura importantes relaciones espaciales de suelos y horizontes edáficos, los cuales tienen topo y cronosecuencia; además de enfatizar la caracterización morfológica de los suelos, sobre la aproximación analítica basada en el análisis de laboratorio (FAO-ISRIC y SICS, 1999; ISSS-ISRIC-FAO, 1994; IUSS Working Group WRB, 2006).
La WRB está diseñada como un medio de comunicación sencillo entre científicos del suelo para identificar, caracterizar y nombrar tipos principales de suelos, finalmente es una herramienta para identificar estructuras pedológicas y su significado (FAO-ISRIC y SICS, 1999; IUSS Working Group WRB, 2006).
La cartografía de México se integra en seis cartas escala 1:1,000,000, todas ellas incluyen el aspecto edafológico; 122 cartas escala 1:250,000, de las cuales 49 son edafológicas y cubren 1,148,989 km2, que representa 59.4% del territorio nacional y 2,313 cartas a 1:50,000, 754 de las cuales son edafológicas, que comprenden 725, 725 km2 y que equivalen a 37.5% de la superficie total del país. Las cartas edáficas se elaboraron en 1970, por lo que es necesario actualizarlas a una escala 1:50,000 que permita conocer, ubicar y cuantificar los suelos y así definir el manejo adecuado para cada uno de ellos (INEGI, 2005).
Los objetivos de esta investigación consistieron en actualizar la clasificación de la cartografía de INEGI escala 1:50,000 de los suelos del Estado de México, de la clasificación FAO 1970 a la versión de suelos de la WRB 2006, cuantificar la superficie de las Unidades y Subunidades edafológicas y delimitar su distribución.
Localización de la zona de estudio
El Estado de México tiene 22,499.95 km2, que representan 1.1% del territorio nacional. Se localiza entre los paralelos 18° 21’ 15’’ y 20° 17’ 00’’ N y los meridianos 98° 35’ 30’’ y 100° 37’ 00’’ O. Colinda al norte con los estados de Querétaro e Hidalgo, al este con Puebla y Tlaxcala, al sur con Morelos y Guerrero,
in different areas: forest management, agriculture, geology, hydrology and ecology; to recognize within the structure, important space relations of soils and edaphic horizons, which have topo and chronosequence; in addition, to emphasize the morphologic composition of soils upon the analytic approach based on laboratory analysis (FAO-ISRIC and SICS, 1999; ISSS-ISRIC-FAO, 1994; IUSS Working Group WRB, 2006).
WRB is designed a simple communication media among scientists of soil to identify, describe and name main types of soils and at the end, it is a tool to identify pedologic structures and their meaning (FAO-ISRIC y SICS, 1999; IUSS Working Group WRB, 2006).
The cartography of Mexico is made up by 6 1:1,000,000 maps, which refer to edaphology; 122 1:250,000 maps, from which 49 are edaphologic and cover 1,148,989 km2 which is equivalent to 59.4% of the total national territory and 2,313 1:50,000 maps, 754 of which are edaphologic and cover 725, 725 km2, which is 37.5% of the total national territory. The edaphic maps were made in 1970, which makes it necessary to update them at a 1:50,000 scale making it possible to know, locate and quantify soils and thus, to define the right management for each one of them (INEGI, 2005).
The aims of this research study were to update the cartographic classification of INEGI at 1:50,000 of the soils of Mexico State, from FAO 1970 classification to the WRB (2006) version; to quantify the area of the edaphic Units and Subunits and to put the boundaries to their distribution.
The study area
Mexico State covers 22,499.95 km2, which are 1.1% of the national territory. It is located between 18° 21’ 15’’ and 20° 17’ 00’’ N and 98° 35’ 30’’ and 100° 37’ 00’’ W. To the north are Querétaro and Hidalgo States, to the al east, Puebla and Tlaxcala States, to the south, Morelos and Guerrero States, to the west, Michoacán State and it engulfs Distrito Federal (INEGI, 1999; INEGI, 2001) (Figure 1).
According to Köppen and adjusted by García (1988), the main climates that prevail in the high valleys of the north, center and east of the State are mild; semi-cold is the next one because of its importance and extent, which is present at the center and east, mainly over Valle de Toluca. In a smaller proportion, there are warm and semi-warm climates, which are at the extreme south of the State, where it neighbors Guerrero, Morelos and Michoacan States. Cold weather dominates small zones of the higher parts such as Nevado de Toluca and Popocatepetl volcano (INEGI, 1999; INEGI, 2001).

75
Sotelo et al., Los suelos del Estado de México y su actualización a la base referencial...
al oeste con Michoacán y envuelve al Distrito Federal (INEGI, 1999; INEGI, 2001) (Figura 1).
Fuente: INEGI (2005). Source: INEGI (2005).
Figura 1. Localización del Estado de México.Figure 1. Location of Mexico State
Los principales climas, de acuerdo con Köppen y modificados por García (1988), son el templado que predomina en los valles altos de la parte norte, centro y este de la entidad. Le sigue en importancia y extensión el clima semifrío, que se distribuye en regiones del centro y este, principalmente en el Valle de Toluca. En menor proporción están los climas cálidos y semicálidos, los cuales se ubican en el extremo sur del estado, en los límites con Guerrero, Morelos y Michoacán. El clima frío impera en pequeñas zonas de las partes más elevadas como el Nevado de Toluca y el volcán Popocatépetl (INEGI, 1999; INEGI, 2001).
Las temperaturas medias anuales van de 6 °C a 28 °C. La precipitación anual oscila entre 600 y 1,800 mm. La altura sobre el nivel del mar va de 340 a 5,100 m. Los suelos predominantes son: Feozem háplico (PHh), Andosol úmbrico (ANu), Vertisol eútrico (VRe), Regosol eútrico (RGe) y Arenosol háplico (ARh) (INEGI, 2001; FAO-ISRIC y SICS, 1999; SEMARNAP, 1999).
En primera instancia se realizó el acopio de la cartografía edafológica escala 1:50 000 del Instituto Nacional de Estadística, Geografía e Informática que cubre al Estado
from 340 to 5,100 m. The dominant soils are: haplic Feozem o (PHh), umbric Andosol (ANu), eutric Vertisol (VRe), eutric Regosol (RGe) and haplic Arenosol (ARh) (INEGI, 2001; FAO-ISRIC y SICS, 1999; SEMARNAP, 1999).The first instance was to gather the edaphologic cartography 1:50 000 of the National Institute of Statistics, Geography and Informatics that covers Mexico State (INEGI, 1974).
The edaphic inventory was achieved by the digitalization of 43 state maps of the 6.0 version of the ARC/INFO program (ESRI, 1992) (Table 1).
The cartographic information of soils belongs to the 1974-1985 period (FAO/UNESCO, 1974; SHR, 1974). The data base that was generated was classified and updated to FAO soil version (1988), FAO-ISRIC and SICS (1999) and IUSS Working Group WRB (2006).
Once they were digitalized, they were reviewed and inconsistencies between adjoining maps were corrected. Later, the data base of each map was made with the following entries: primary and secondary soil units, texture and physical
Annual mean temperatures vary from 6 °C to 28 °C. Annual precipitation, from 600 to 1,800 mm. Height over sea level,

76
Rev. Mex. Cien. For. Vol. 2 Núm. 8
de México (INEGI, 1974). El inventario edáfico se creó mediante la digitalización de las 43 cartas estatales en el programa ARC/INFO versión 6.0 (ESRI, 1992) (Cuadro 1).
and chemical phases. The digital archives of the 43 maps were imported to the geographic information system ARCGIS 9.0 version (ESRI, 2006) in shape format, which were put together into one single edaphologic map (Figure 2).
Cuadro 1. Cartas Edafológicas que cubren al Estado de México, escala 1:50,000.Table 1. 1:50,000 Soil maps for Mexico State.
Número Clave Nombre Número Clave Nombre
1 F14C77 San Juan del Río 23 E14A49 Milpa Alta2 F14C86 Amealco 24 E14A55 Bejucos
3 F14C87 Polotitlán 25 E14A56 Tejupilco
4 F14C88 Tula 26 E14A57 Ixtapán de la Sal
5 F14C89 Mixquihuala 27 E14A58 Tenancingo
6 F14D81 Pachuca 28 E14A59 Cuernavaca
7 E14A16 El oro 29 E14A65 Palmar Chico
8 E14A17 Atlacomulco 30 E14A66 Amatepec
9 E14A18 Tepeji 31 E14A67 Pilcaya
10 E14A19 Zumpango 32 E14A68 Taxco
11 E14A26 Agangueo 33 E14A75 Ciudad Altamirano
12 E14A27 Ixtlahuaca 34 E14A76 Arcelia
13 E14A28 Villa del Carbón 35 E14B11 Tizayuca
14 E14A29 Cuautitlán 36 E14B12 Ciudad Sahagún
15 E14A36 Villa de Allende 37 E14B21 Texcoco
16 E14A37 Zinacantepec 38 E14B22 Apan
17 E14A38 Toluca 39 E14B31 Chalco
18 E14A39 Ciudad de México 40 E14B32 Mariano Arista
19 E14A45 Tuzantla 41 E14B41 Amecameca
20 E14A46 Valle de Bravo 42 E14B42 Huejotzingo
21 E14A47 Nevado de Toluca 43 E14B51 Cuautla
22 E14A48 Tenango
La información cartográfica de los suelos corresponde al periodo 1974 a 1985 (FAO/UNESCO, 1974; SHR, 1974). La base de datos generada se clasificó y actualizó a la versión de suelos de la FAO (1988), FAO-ISRIC y SICS (1999) y IUSS Working Group WRB (2006).
Una vez digitalizadas las cartas, se revisaron y corrigieron las inconsistencias entre cartas contiguas. Posteriormente se elaboró la base de datos de cada carta con los campos unidad de suelo primario y secundario, textura y las fases físicas y químicas. Los archivos digitales de las 43 cartas se importaron al sistema de información geográfica ARCGIS versión 9.0 (ESRI, 2006) en formato shape, las cuales se unieron en un solo mapa edafológico (Figura 2).
En ARCGIS se cambió la base de datos de las unidades de suelo de la clasificación FAO-UNESCO 1970 a la clasificación FAO 1988 y WRB 2006. Se adicionaron tres nuevos campos para tener las tres clasificaciones (1988,1999 y 2006) en el mismo mapa. A continuación se procedió a recortar el Estado
By means of ARCGIS the data base of the soil units of FAO-UNESCO 1970 classification were changed to the FAO 1988 and WRB 2006 classification. Three new entries were added to have the three classifications 1988, 1999 and 2006) in the same map. Next, Mexico State was cut in ARCGIS and the area for each one of the Units was calculated. Finally, in ARGIS maps with Soil Units update to WRB (IUSS Working Group WRB, 2006) were designed.
Keys to classify and identify soils in FAO-WRB. To describe and define the Soil Units for FAO and WRB, characteristics, properties and horizons were used, which were combined to define soils and their relations (FAO/UNESCO, 1974; IUSS Working Group WRB, 2006; SRH, 1974) (Table 2).

77
Sotelo et al., Los suelos del Estado de México y su actualización a la base referencial...
Figura 2. Cartografía edafológica escala 1:50,000 que cubre al Estado de México.
Figure 2. 1:50,000 Soil cartography for Mexico State.
de México en ARCGIS, y se calculó la superficie para cada una de las Unidades. Finalmente, se diseñaron los mapas en ARGIS con las Unidades de suelos actualizadas a la WRB (IUSS Working Group WRB, 2006).
Claves para clasificar e identificar los suelos en la FAO-WRB. Para describir y definir las Unidades de suelos de la FAO y la WRB se usaron las características, propiedades y horizontes, que se combinaron para definir a los suelos y sus relaciones (FAO/UNESCO, 1974; IUSS Working Group WRB, 2006; SRH, 1974). (Cuadro 2).
Los resultados de las Unidades y Subunidades de suelos de la clasificación de la FAO/UNESCO de 1974 a la versión de la WRB de 2006 se presentan en cuadros y la distribución de cada Unidad se muestra en un mapa.
The results of the Units and Subunits of soils of the FAO/UNESCO (1974) classification to the WRB (2006) version are in tables and their distribution of each Unit in a map.
In Table 3 are shown the different Units and Subunits, while in Table 4, are the Units and the areas that they cover in the State (Figure 3). The dominant soils are Los suelos Andosols with 4,799.08 km2, Feozems with 4,727.18, Regosols with 2,656.83, Vertisols with 2,414.85 and Cambisols with 1,960.47 km2
In regard to the total area, there is a difference of 179.64 km2 compared to what is in the record for Mexico State which at the end sums up 22, 499.95 km2. It is worth-noticing that the boundaries that were used in this study were taken from INEGI (1999), which explains that difference.

78
Rev. Mex. Cien. For. Vol. 2 Núm. 8
Cuadro 2. Unidades de suelos y su simbología de las diferentes versiones de la clasificación FAO-WRB.Table 2. Soil units and symbols of the different versions of the FAO-WRB classification.
Unidades de sueloFAO 1970 FAO 1988 WRB 1999 WRB 2006
Acrisoles (A) Acrisoles (AC) Acrisoles (AC) Acrisoles (AC)Albeluvisoles (AB) Albeluvisoles (AB)
Alisoles (AL) Alisoles (AL) Alisoles(AL)Andosoles (T) Andosoles (AN) Andosoles (AN) Andosoles (AN)
Antrosoles (AT) Antrosoles (AT) Antrosoles (AT)Arenosoles (Q) Arenosoles (AR) Arenosoles (AR) Arenosoles (AR)
Calcisoles (CL) Calcisoles (CL) Calcisoles (CL)Cambisoles (B) Cambisoles (CM) Cambisoles (CM) Cambisoles (CM)Chernozems (C) Chernozem (CH) Chernozem (CH) Chernozem (CH)
Criosoles (CR) Criosoles (CR)Durisoles (DU) Durisoles (DU)
Feozems (H) Feozems (PH) Feozems (PH) Feozems (PH)Ferrasoles (F) Ferrasoles (FR) Ferrasoles (FR) Ferrasoles (FR)Fluvisioles (J) Fluvisioles (FL) Fluvisoles (FL) Fluvisoles (FL)Gleysoles (G) Gleysoles (GL) Gleysoles (GL) Gleysoles (GL)Greyzems (M) Greyzems (GR)
Gipsisoles (GY) Gipsisoles (GY) Gipsisoles (GY)Histosoles (O) Histosoles (HS) Histosoles (HS) Histosoles (HS)Kastañozems (K) Kastañozems (KS) Kastañozems (KS) Kastañozems (KS)Litosoles (I) Leptosoles (LP) Leptosoles (LP) Leptosoles (LP)Rendzinas (E)Rankers (U)
Lixisoles (LX) Lixisoles (LX) Lixisoles (LX)Luvisoles (L) Luvisoles (LV) Luvisoles (LV) Luvisoles (LV)Nitisoles (N) Nitisoles (NT) Nitisoles (NT) Nitisoles (NT)Planosoles (W) Planosoles (PL) Planosoles (PL) Planosoles (PL)
Plintosoles (PT) Plintosoles (PT) Plintosoles (PT)Podzoles (P) Podzoles (PZ) Podzoles (PZ) Podzoles (PZ)Podzoluvisoles (D) Podzoluvisoles (PD) Regosoles (R) Regosoles (RG) Regosoles (RG) Regosoles (RG)Solonchaks (Z) Solonchaks (SC) Solonchaks (SC) Solonchaks (SC)Solonetz (S) Solonetz (SN) Solonetz (SZ) Solonetz (SN)
Estagnosoles (ST)Tecnosoles (TC)
Umbrisoles (UM) Umbrisoles (UM)Vertisoles (V) Vertisoles (VR) Vertisoles (VR) Vertisoles (VR)Xerosoles (X)Yermosoles (Y)
En el Cuadro 3 se consignan las diferentes Unidades y Subunidades, mientras que en el Cuadro 4 las Unidades y la superficie que cubren del estado (Figura 3). Los suelos dominantes son los Andosoles con 4,799.08 km2, Feozems con 4,727.18, Regosoles con 2,656.83, Vertisoles con 2,414.85 y Cambisoles con 1,960.47 km2.
Con respecto a la superficie total, hay una diferencia en la que se registra para el Estado de México 179.64 km2, que al final da un total de 22, 499.95 km2. Cabe aclarar que los límites que se utilizaron para este trabajo fueron los que cita INEGI (1999), lo que justifica dicha diferencia.
In tables 5 and 6 are the areas of the phases and texture of soils. In figures 4 and 5 their distribution is shown.
The prevailing soils in Mexico State are Andosols, Feozems, Regosols, Vertisols, Cambisols and Leptosols (Figure 3). Andosols are at the center: center-west, center-east, and southeast. In these zones, height above sea level is 2,800 m.
Feozems are located at the north, with some small areas in the southeast section. Regosols dominate the south of the Tejupilco zone towards the limits of Guerrero and Michoacan State. Vertisols are in Ixtlahuaca-Atlacomulco valley (to the north), Jilotepec and in Texcoco zone. Cambisols prevail in the south, after Andosols, at altitudes beneath 1,800 m, but before Regosols.

79
Sotelo et al., Los suelos del Estado de México y su actualización a la base referencial...
Cuadro 3. Unidades y Subunidades de suelos en el Estado de México.Table 3. Units and subunits of Mexico State soils.
Tipo de suelos WRB 2006 Simbología Superficie (km2)
Acrisol Húmico AChu 45.08Acrisol Háplico ACha 424.59Andosol Úmbrico ANum 3,166.38Andosol Mólico ANmo 448.56Andosol Háplico ANha 1,184.14Cambisol Crómico CMcr 546.20Cambisol Dístrico CMdy 339.53Cambisol Eútrico CMeu 790.59Cambisol Ferrálico CMfl 66.58Cambisol Húmico CMhu 12.30Cambisol Háplico CMha 107.37Cambisol Calcárico CMca 58.39Cambisol Vértico CMvr 39.51Feozems Calcárico PHca 339.47Feozem Gléyico PHgl 48.02Feozem Háplico PHha 3,734.39Feozem Lúvico PHlv 605.30Fluvisol Dístrico FLdy 76.44Fluvisol Eútrico FLeu 174.88Fluvisol Calcárico FLca 0.84Gleysol Eútrico GLeu 0.29Gleysol Úmbrico GLum 1.08Gleysol Mólico GLmo 19.06Gleysol Háplico GLha 4.99Gleysol Vértico GLvr 2.24Histosol Eútrico HSeu 40.50Leptosol Lítico LPli 1,476.85Leptosol Réndzico LPrz 263.22Leptosol Úmbrico LPum 9.62Luvisol Cálcico LVcc 0.88Luvisol Crómico LVcr 1,346.94Luvisol Férrico LVfr 78.72Luvisol Háplico LVha 31.85Luvisol Vértico LVvr 10.66Planosol Eútrico PLeu 125.35Planosol Úmbrico PLum 71.38Planosol Mólico PLmo 563.45Regosol Calcárico RGca 27.50Regosol Dístrico RGdy 107.73Regosol Eútrico RGeu 2,520.04Regosol Gélico RGge 1.56Solonchak Gléyico SCgl 228.60Solonchak Mólico SCmo 104.61Solonchak Háplico SCha 59.69Vertisol Crómico VRcr 137.20Vertisol Pélico VRpe 2,277.64

80
Rev. Mex. Cien. For. Vol. 2 Núm. 8
Cuadro 4. Unidades de suelos presentes en el Estado de México.Table 4. Soil units of Mexico State.
Tipos de suelos WRB 2006 Simbología Superficie (km2)
Andosoles AN 4,799.08Feozems PH 4,727.18Regosoles RG 2,656.83Vertisoles VR 2,414.85Cambisoles CM 1,960.47Leptosoles LP 1,749.68Luvisoles LV 1,469.05Planosoles PL 760.18Acrisoles AC 469.68Solonchaks SC 392.90Fluvisoles FL 252.16Histosoles HS 40.50Gleysoles GL 27.65Superficie total 21,719.62
En los cuadros 5 y 6 se proporcionan la superficie de las fases y las texturas de los suelos. En las figuras 4 y 5 se observa su distribución.
Cuadro 5. Superficie de las diferentes texturas de los suelos en el Estado de México.
Table 5. Area of the different soil textures in Mexico State.Texturas Superficie (km2)
Fina 3,560.72Media 17,209.25Gruesa 951.31
Cuadro 6. Superficie de las fases físicas y químicas de suelos en el Estado de México.
Table 6. Area of the physical and chemical phases of the soils of Mexico State.
Fases físicas y químicas Superficie (km2)Dúrica 1,574.59Dúrica profunda 1,275.57Lítica 4,671.79Lítica profunda 2,936.39Pedregosa 605.25Gravosa 702.69Petrocálcica 9.80Ligeramente salino 65.19Moderadamente salino 5.04Sódica 464.01Sin fases 9,409.30
Los suelos que predominan en el Estado de México son los Andosoles, Feozems, Regosoles, Vertisoles, Cambisoles y Leptosoles (Figura 3). Los Andosoles se presentan en la parte central del estado: centro- oeste, centro- este y sureste. En estas zonas, la altura sobre el nivel del mar es superior a 2,800 m.
Los Feozems se sitúan en la parte norte, con unas pequeñas áreas en la porción suroeste. Los Regosoles dominan el sur de la zona de Tejupilco hacia los límites con Guerrero y Michoacán. Los Vertisoles se ubican en el valle de Ixtlahuaca-Atlacomulco (hacia el norte de la entidad), Jilotepec y en la zona de
Figura 3. Unidades de suelos FAO-WRB presentes en el Estado de México.
Figure 3. FAO-WRB soil units at Mexico State.
Leptosols are observed in the south in the limits with Morelos State, south of the Rural Development District of Texcoco in the boundaries of Guerrero State, in the Coatepec Harinas District and in the west adjacent with Michoacan State in Valle de

81
Sotelo et al., Los suelos del Estado de México y su actualización a la base referencial...
Figura 4. Texturas de suelos según FAO-WRB presentes en el Estado de México.
Figure 4. Soil textures in Mexico State according to FAO-WRB.
Texcoco. Los Cambisoles dominan en la parte sur, después de los Andosoles, en alturas inferiores a 1,800 msnm y están antes de los Regosoles.
Los Leptosoles se observan en el sur en los límtes con Morelos, sur del Distrito de Desarrollo Rural de Texcoco en los límites con Guerrero, en el Distrito de Coatepec Harinas y en el oeste en colindancia con Michoacán en el Distrito de Valle de Bravo. Son suelos que cambiaron de nombre, ya que antes se denominaban Litosoles (FAO/UNESCO, 1974). La mayor superficie agrícola del Estado de México se localiza en el Valle de Toluca, donde los suelos dominantes son los Feozems, el Valle de Ixtlahuaca-Atlacomulco y la zona de Jilotepec, donde dominan los Vertisoles y Feozems (SAGARPA, 2007).
Los suelos con texturas medias ocupan la mayor superficie con 17,209.25 km2, seguidos por los de textura fina con 3,560.72 km2 y por los de textura gruesa con 951.31 km2 (Figura 4). Las fases de los suelos, las cuales son una limitante para el desarrollo de los cultivos, son: lítica que cubren una superficie de 4,671.79 km2, lítica profunda con 2,936.39 km2,
Figura 5. Fases físicas y químicas de suelos según FAO-WRB en el Estado de México.
Figure 5. Physical and chemical phases of the soils of Mexico State according to FAO-WRB.
Bravo District. These soils changed their name, since they were known as Litosols (FAO/UNESCO, 1974). The largest agriculture area of Mexico State is located in Valle de Toluca, where the dominant soils are Feozems; Vertisols and Feozems are in the Ixtlahuaca-Atlacomulco valley and in Jilotepec (SAGARPA, 2007).
Soils with medium texture cover the greatest area, 17,209.25 km2, followed by those of fine texture, 3,560.72 km2 and by those of coarse texture, 951.31 km2 (Figure 4). The soil phases, which are a limitation for crops, are: lytic, which covers 4,671.79 km2, deep lytic, 2,936.39 km2, duric 1,574.59 km2 and deep duric 1,275.57 km2. The Lytic phase prevails in the southern part and in the highlands with a severe slope (Figure 5).
Finally, the studies that involve updating of FAO classification soil Units were made by Ortiz et al. (1994) and SEMARNAP (1996), in which they establish the foundations and they made an adjustment of Soil Units of the 1974 FAO/UNESCO version of 1988.

82
Rev. Mex. Cien. For. Vol. 2 Núm. 8
dúrica con 1,574.59 km2 y dúrica profunda con 1,275.57 km2. La fase lítica predomina en la parte sur y en las partes altas del estado con mucha pendiente (Figura 5).
Finalmente, los trabajos que se han realizado sobre actualizaciones de las Unidades de suelos de la clasificación FAO, los realizaron Ortiz et al. (1994) y la SEMARNAP (1996), en los cuales establecen los fundamentos y hacen una adecuación de las Unidades de suelos de la versión FAO/UNESCO de 1974 a la FAO/UNESCO de 1988.
CONCLUSIONES
Los suelos dominantes son los Andosoles con 4,799.08 km2, Feozems con 4,727.18 km2, Regosoles con 2,656.83 km2, Vertisoles con 2,414.85 km2 y Cambisoles con 1,960.47 km2. Los Leptosoles son suelos que cambiaron de nombre, antes eran los Litosoles, que ocupan 1,749.68 km2, lo que reduce la superficie para la producción de cultivos.
Los mejores suelos para las actividades agropecuarias son los Feozems y Verisoles que dominan en el Valle de Toluca, Ixtlahuaca-Atlacomulco, Jilotepec y Texcoco, que es donde existe la mayor superficie dedicada a la agricultura. Los Andosoles y Cambisoles dominan en toda la franja central del Estado que es la parte alta y en estos suelos la actividad forestal es el uso del suelo más importante.
Contar con cartografía digital de suelos con una clasificación actualizada facilita la aplicación de modelos edafológicos e hidrológicos recientes que la utilizan y son la base para la planeación e instrumentación de las acciones de manejo y conservación de los ecosistemas forestales, la producción agrícola y la ubicación de infraestructura humana.
REFERENCIAS
Brady, N. C. and R. R. Weil. 1999. The nature and properties of soils. 12th Ed. Prentice Hall. Upper Saddle River, NJ. USA. 881 p.
Boul, S. W., F. D. Hole and R. J. McCracken. 2004. Génesis y clasificación de suelos. Ed. Trillas, S. A. 4ª. reimpresión. México, D. F. México. 417 p.
Environmental Systems Research Institute (ESRI). 1992. ARC/INFO. User’s Guide. Vertion 6.0. New York, NY. USA. 125 p.
Environmental Systems Research Institute (ESRI). 2006. ArcGis 9.0. New York, NY. USA. 276 p.
Food and Agriculture Organization of the United Nations (FAO). 2000. Digital Soil Map of the World (www.fao.org/catalog/New/products). Rome, Italy. s/p.
Food and Agriculture Organization of the United Nations/United Nations Educational, Cientific and Cultural Organization (FAO/UNESCO). 1974. Soil Map of the World 1:1 000 000. Vol. I. Legend. Paris, France. 59 p.
Food and Agriculture Organization of the United Nations/United Nations Educational, Cientific and Cultural Organization (FAO/UNESCO). 1988. Soil Map of the World. Revised Legend. World Soil Resources Report 60. Rome, Italy. 136 p.
Fitzpatrick, E. A. 1987. Suelos. Su formación, clasificación y distribución. Compañía Editorial Contiental, S. A. de C. V. México, D. F. México. pp. 229-345.
Fitzpatrick, E. A. 1996. Introducción a la ciencia de los suelos. Editorial trillas. México, D. F. México. pp. 179-247.
CONCLUSIONS
Andosols with 4,799.08 km2, Feozems with 4,727.18 km2, Regosols with 2,656.83 km2, Vertisoles with 2,414.85 km2 and Cambisols with 1,960.47 km2 are the dominant types of soil. Leptosols are soils that changed their name, since they were the Litosols before, which cover 1,749.68 km2, a fact that reduces the area for crops.
The best soils for agriculture and livestock activities are Feozems and Vertisols in Valle de Toluca, Ixtlahuaca-Atlacomulco, Jilotepec and Texcoco, where the greatest area for agriculture is found. Andosols and Cambisols prevail in the whole central line of the State which is the high part, and in those soils forest activity is the most important land use.
To count with soil digital cartography with an updated class i fi cat ion favors the applicat ion of edapholog ic and hydrologic recent models that use them and is the basis for planning and implementation of management and conservation actions for forest ecosystems, agriculture production and location of human infrastructure.
End of the English version
García M., E. 1988. Modificaciones al Sistema de Clasificación Climática de Köppen para adaptarlo a las condiciones de la República Mexicana. México, D. F. México.185 p.
Instituto Nacional de Estadística Geografía e Informática (INEGI). 1999. Anuario Estadístico del Estado de México. Aguascalientes, Ags., México. 596 p.
Instituto Nacional de Estadística, Geografía e Informática (INEGI). 2001. Avance cartográfico edafológico 1:50, 000. Aguascalientes, Ags., México. s/p.
Instituto Nacional de Estadística, Geografía e Informática (INEGI). 2005. Marco Geoestadístico Municipal 2005 (MGM2005). Aguascalientes, Ags., México. s/p.
Instituto Nacional de Estadística, Geografía e Informática (INEGI). 2001. Síntesis de Información Geográfica del Estado de México. Nomenclatura y Anexo Cartográfico. Aguascalientes, Ags., México. 139 p.
Instituto Nacional de Estadística, Geografía e Informática (INEGI). Serie I. Carta temática edaológica escala 1:50 000. Estado de México, Edo. de Méx. México, s/p.
International Society of Soil Science - Society International of Sciences of Soil - Food and Agriculture Organization of the United Nations ( ISSS-ISRIC-FAO). 1994. World Reference Base for Soil Resources. Rome, Italy. 161 p.
International Union of Soil Science - International Soil Reference and Information Centre and Food and Agriculture Organization of the United Nations (IUSS Working Group WRB. IUSS-ISRIC and FAO). 2006. World reference base for soil resources. A framework for international classification, correlation and communication. 2nd edition. World Soil Resources Reports No. 103. Rome, Italy. 114 p
Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO). 1985. Directivas: evaluación de tierras para la agricultura en secano. Boletín de Suelos de la FAO No. 52. Roma, Italia. 228 p.
Organización de las Naciones Unidas para la Agricultura y la Alimentación - Centro Internacional de Referencia e Información en Suelos y Sociedad Internacional de las Ciencias del Suelo (FAO-ISRIC y SICS). 1999. Base Referencial Mundial del Recurso Suelo. Reporte No. 84. Roma, Italia. 93 p.
Ortiz S., C., D. Pájaro H. y V. Ordaz C. 1990. Manual para la cartografía de las clases de tierras campesinas. Centro de Edafología. Colegio de Postgraduados. Montecillo, Edo. de Méx., México. 62 p.

83
Sotelo et al., Los suelos del Estado de México y su actualización a la base referencial...
Ortiz S., C., D. Pájaro H. y Ma. del C. Gutiérrez C. 1994. Introducción a la leyenda del mapa mundial de suelos FAO/UNESCO, versión 1988. Instituto de Recursos Naturales. Programa de Edafología. Colegio de Postgraduados. Montecillo, Edo. de Méx., México. 40 p.
Ortiz S. C., y Ma. del C. Gutiérrez C. 1999. Fundamentos de pedología. Especialidad de Edafología. Instituto de Recursos Naturales. Colegio de Postgraduados. Montecillo, Texcoco, Edo., de Méx., México. 104 p.
Secretaria de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA). 2007. Estadísticas Estatales de la SAGARPA. Toluca, Edo. de México. s/p.
Secretaria de Medio Ambiente, Recursos Naturales y Pesca (SEMARNAP). 1999. Fundamentos para la actualización de la cartografía Edafológica de la Republica Mexicana, con el Sistema de clasificación FAO/UNESCO (1988). México, D. F. México. 22 p.
Soil Survey Staff. 1992. Keys to soil taxonomy. SMSS Technical Monograph No. 19, 5th ed. Pocahontas Press, Inc. Blacksburg, VI. USA. 541 p.
Soil Survey Staff. 1994. Keys to soil taxonomy. 6th ed. USDA-SCS. U. S. Government Priting Office. Washington, DC. USA. 306 p.
Soil Survey Staff. 1998. Soil taxonomy. A basic system of soil classification for making and interpreting soil surveys. Second edition. United States Department of Agriculture. Natural Resources Consrvation Service. Washington, DC. USA. 326 p.
Soil Survey Staff. 1999. Soil taxonomy. A basic system of soil classification for making and interpreting soil surveys. 2nd ed. United States Department of Agriculture. Natural Resources Consrvation Service. Washington, DC. USA. 865 p.
Soil Survey Staff. 2003. Keys to soil taxonomy. 9th ed. Department of Agriculture United States. Washington, DC. USA. 332 p.
Soil Survey Staff, 2006. Keys to soil taxonomy. 10th ed. United States Department of Agriculture. Natural Resources Conservation Service. Washington, DC. USA. 341 p.
Secretaría de Recursos Hidráulicos (SRH). 1974. Clave de unidades de suelos para el mapa de suelos del mundo. Dirección General de Estudios. Dirección de Agrología. México, D. F., México. 29 p.
Wilding, L. P. 2000. Classification of soil. In: M. E. Sumner (Ed.). Handbook of Soil Science. New York, NY. USA. pp. E175-E392.

84
Rev. Mex. Cien. For. Vol. 2 Núm. 8
Wangari Muta Maathai. Dominio público.

85
CONSEJO ARBITRAL INTERNACIONAL
ArgentinaInstituto Nacional de Tecnología Agropecuaria.- M.Sc. Leonel Harrand
Museo Argentino de Ciencias Naturales.- Dra. Ana María Faggi
Canadá Universitè Laval, Québec.- Ph. D. Roger Hernández.
Cuba Instituto de Investigaciones Fundamentales en Agricultura Tropical.- Dra. Amelia Capote Rodríguez
Unión Nacional de Escritores y Artistas de Cuba.- Dra. Raquel Carreras Rivery
ChileUniversidad del Bío Bío.- Dr. Rubén Andrés Ananias Abuter
EspañaCIFOR-INIA.- Dr. Eduardo López Senespleda, Dr. Gregorio Montero González, Dr. Sven Mutke Regneri
Fundación CEAM.- Dra. María José Sánz SánchezUniversidad de Oviedo.- Dr. Elías Afif Khouri
Universidad Politécnica de Madrid.- Dr. Alfredo Blanco Andray, Dr. Luis Gil Sánchez, Dr. Alfonso San Miguel-Ayanz,Dr. Eduardo Tolosana, Dr. Santiago Vignote Peña
Estados Unidos de AméricaNew Mexico State University.- Ph.D. John G. MexalNorthern Arizona University .- Ph.D. Peter Z. Fulé
University of Colorado at Denver.- Ph.D. Rafael Moreno SánchezUniversity of Florida.- Ph.D. Francisco Javier Escobedo Montoya
United States Department of Agriculture, Forest Service.- Dr. Mark E. Fenn, Dr. Carlos Rodriguez Franco
ItaliaInternational Plant Genetic Resources Institute.- Dra. Laura K. Snook
CONSEJO ARBITRAL NACIONAL
.Asociación Mexicana de Arboricultura.- Dr. Daniel Rivas Torres.Benemérita Universidad Autónoma de Puebla.- Dr. José F. Conrado Parraguirre Lezama.
Centro de Investigación y Docencia Económicas, A. C. (CIDE).- Dr. Alejandro José López-FeldmanCENTROGEO / CONACYT.- Dra. Alejandra López Caloca.
Colegio de Postgraduados.- Dr. Arnulfo Aldrete, Dr. Dionicio Alvarado Rosales, Dr. Víctor M. Cetina Alcalá,Dra. Ma. de Lourdes de la Isla de Bauer, Dr. Héctor M. de los Santos Posadas, Dr. Armando Equihua Martínez,
Dr. Ronald Ferrara-Cerrato, Dr. Edmundo García Moya, Dr. Jesús Jasso Mata, Dr. Lauro López Mata, Dr. Javier López Upton,Dr. Martín Alfonso Mendoza Briseño, Dr. Antonio Trinidad Santos, Dr. Juan Ignacio Valdés Hernández,Dr. José René Valdez Lazalde, Dr. J. Jesús Vargas Hernández, Dra. Heike Dora M. Vibrans Lindemann
El Colegio de la Frontera Sur.- Dr. Bernardus H. J. de Jong, Dr. Mario González Espinosa, Ph.D. Jorge E. Macías Sámano,Dr. Neptalí Ramírez Marcial, Dr. Cristian Tovilla Hernández, Dr. Henricus Franciscus M. Vester
El Colegio de México.- Dra. María PerevochtchikovaEl Colegio de Tlaxcala, A. C.- M.C. Noé Santacruz García

Rev. Mex. Cien. For. Vol. 2 Núm. 8
86
Instituto de Ecología, A. C..- Dr. Pedro Guillermo Ángeles Álvarez, Dr. Ismael Raúl López Moreno
Instituto Politécnico Nacional.- Dr. Alejandro Daniel Camacho Vera, Ph.D. José de Jesús Návar Cháidez,M.C. D. Leonor Quiroz García
PRONATURA.- Dr. José A. Benjamín Ordoñez DíazUniversidad Autónoma Agraria Antonio Narro.- Dr. Eladio Heriberto Cornejo Oviedo, M.C. Salvador Valencia Manzo
Universidad Autónoma Chapingo.- Dr. Leonardo Sánchez Rojas, Dr. Enrique Serrano Gálvez,Dra. Ernestina Valadez Moctezuma, M.C. Beatriz Cecilia Aguilar Valdez, M.C. Baldemar Arteaga Martínez,
M.C. Emma Estrada Martínez, M.C. Mario Fuentes Salinas, M.C. Enrique Guízar Nolazco,Dr. Hugo Ramírez Maldonado, Dr. Dante Arturo Rodríguez Trejo
Universidad Autónoma de Baja California Sur.- Dr. José Antonio Martínez de la Torre Universidad Autónoma de Chihuahua.- Ph.D. Concepción Luján Álvarez, Ph.D. Jesús Miguel Olivas García
Universidad Autónoma de Guadalajara.- Dr. Mauricio Alcocer RuthlingUniversidad Autónoma de Nuevo León .- Dr. Glafiro J. Alanís Flores, Dr. Enrique Jurado Ybarra,
Dr. José Guadalupe Marmolejo MonsiváisUniversidad Autónoma de Querétaro.- Dr. Luis Gerardo Hernández SandovalUniversidad Autónoma de San Luis Potosí.- M.C. Carlos Arturo Aguirre Salado
Universidad Autónoma del Estado de Hidalgo.- Dra. Ana Laura López Escamilla, Dr. Ángel Moreno FuentesUniversidad Autónoma del Estado de México.- Dr. Darío Ibarra Zavala
Universidad Autónoma Indígena de México.- Dra. Hilda Susana Azpiroz Rivero Universidad Autónoma Metropolitana.- Dr. Héctor Castillo Juárez, Dra. Carmen de la Paz Pérez Olvera
Dr. Luis Ramón Bravo García, Dr. Ezequiel Delgado Fourné, M.C. Francisco Javier Fuentes Talavera, Universidad de Guadalajara.- M.C. María Guadalupe Lomelí Ramírez, M.C. Roberto Novelo González,
Dr. Rubén Sanjuán DueñasUniversidad del Mar.- M.C. Verónica Ortega Baranda
Universidad Juárez Autónoma de Tabasco.- Dr. Miguel Alberto Magaña AlejandroDr. Javier Leonardo Bretado Velázquez, Dr. Hermes Alejandro Castellanos Bocaz, Dr. José Javier Corral Rivas,Universidad Juárez del Estado de Durango.- Ph.D. José Ciro Hernández Díaz, Dr. Marín Pompa García
Dr. José Cruz de León, M.C. Marco Antonio Herrera Ferreyra, Dr. Alejandro Martínez Palacios, Universidad Michoacana de San Nicolás de Hidalgo.- Dr. José Guadalupe Rutiaga Quiñones, Dr. David Zavala Zavala
Universidad Nacional Autónoma de México.- Dra. María del Consuelo Bonfil Sanders, Dr. Humberto Bravo Álvarez,Dra. Eliane Ceccón, Dr. Joaquín Cifuentes Blanco,
Dr. Abisaí Josué García Mendoza, Dr. Roberto Garibay Orijel, Dr. Julio Alberto Lemos Espinal, Dr. Daniel Piñero Dalmau, Dr. Américo Saldívar Valdés, Dra. Teresa Terrazas Salgado
Universidad Veracruzana.- Dr. Carlos H. Ávila Bello, Dr. Lázaro Rafael Sánchez VelásquezInstituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias.- Dr. Miguel Acosta Mireles,
Dr. Víctor Javier Arriola Padilla, Dr. Francisco Becerra Luna, Dr. Juan de Dios Benavides Solorio, Dr. Fernando Carrillo Anzures, Dr. Carlos Román Castillo Martínez, M.C. Alfonso de la Rosa Vázquez, Dr. José Germán Flores Garnica, M.C. Antonio González Hernández, Dr. Vidal Guerra de la Cruz,
Dr. José Amador Honorato Salazar, Dr. Fabián Islas Gutiérrez, M.C. Juan Islas Gutiérrez, Dr. Emiliano Loeza Kuk, M.C. José Francisco López Toledo, Dr. Martín Martínez Salvador, Dra. Aixchel Maya Martínez,
Dr. José Isidro Melchor Marroquín, M.C. Francisco Moreno Sánchez, Dr. Ramiro Pérez Miranda, Dr. José Ángel Prieto Ruíz,M.C. Fabiola Rojas García, Dr. Guillermo Sánchez Martínez, Dr. Erasto Domingo Sotelo Ruiz,
Dr. Arturo Gerardo Valles Gándara, Dr. José Villanueva Díaz, M.C. Eulalia Edith Villavicencio GutiérrezConsultores Privados.- M.Sc. Rosalía A. Cuevas Rangel , Dra. Teresita del Niño Jesús Marín Hernández, Dr. Gustavo Cruz Bello.
CONSEJO EDITORIAL
Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias.-Dr. Francisco Becerra Luna,M.C. Andrés Flores García, M.C. Georgel Moctezuma López, M.C. Francisco Moreno Sánchez,
M.C. Santa Ana Ríos Ruíz, M.C. Juan Carlos Tamarit Urias,Universidad Nacional Autónoma de México.- M.C. Verónica del Pilar Reyero Hernández

87
Carlos Galindo, (2011). Flor de Tulipan africano.

Wangari Muta Maathai. Dominio público.

A partir del Volumen 2, Número 3, enero-febrero 2011, se deberá cubrir una cuota de $350.00 (TRESCIENTOS CINCUENTA PESOS, 00/100 M.N.) por página completa publicada (traducción y gastos de edición).El pago de suscripciones y publicación de artículos se realizará por medio de un depósito a nombre del INIFAP/CENID-COMEF, en la cuenta No. 0657617851, Clabe Interbancaria 072 180 00657617851 2, del Grupo Financiero BANORTE, Sucursal No. 2037. En el caso de suscripciones internacionales, la Clave SWIFTT correspondiente es: MENOMXMT. Se deberá enviar copia del depósito por fax o correo electrónico. Si el pago es con cheque, se requiere expedirlo a nombre del INIFAP/CENID-COMEF.
Precios de suscripción (incluye envío)Nacional: $ 600.00 Institucional / IndividualExtranjero $ 90.00 USD Institucional / Individual
Toda correspondencia relacionada con la revista, favor de dirigirla a:
Editor en Jefe de la Revista Mexicana de Ciencias ForestalesAv. Progreso No. 5Barrio de Santa CatarinaDelegación CoyoacánC.P. 04010 México, D. F. México.Correo-e: [email protected]éfono y Fax: (+52-55) 3626-8697 Conmutador: (+52-55) 3626-8700 ext. 112
Producción: Carlos Mallén RiveraCuidado de la Edición: Marisela C. Zamora Martínez
Diseño y formación: Silvia Onodera HamanoGestión, seguimiento y galeras: Margarita Muñoz Morgado,Lourdes Velázquez Fragoso y Laura Gabriela Herrerías Mier
Impresión, encuadernación y terminado:Graphx, S.A. de C.V.
El
Vo lu m e n 2, Número 8
de la Revista Mexicana de Ciencias Forestales, se
terminó de imprimir en noviembre de 2011, “Año Internacional de
los Bosques”, por: Graphx, S.A. de C.V. , Tacuba 40-205, Col .
Centro Histórico, C.P. 06010 México, D.F . Tiraje: 1,000
ejemplares .

Rev. Mex. Cien. For. Vol. 2 Núm. 8
90

















