THE JOURNAL OF Bro~oouxr. CHEMISTRY Vol. 245, No. 20, Issue of October 25, PP. 5228-5233, 1970 Printed in U.S.A. Ribonucleotide Reductase and Cell Proliferation I. VARIATIONS OF RIBONUCLEOTIDE REDUCTASE ACTIVITY WITH TUMOR GROWTH RATE IN A SERIES OF RAT HEPATOMAS* (Received for publication, May 5, 1970) HOWARD L. ELFORD,$ MICHAEL FREESE, EUGENE PASSAMANI, AND HAROLD P. MORRIS From the Department of Biological Chemistry, University of Michigan, Ann Arbor, Michigan 48104, and De- partment of Biochemistry, Howard University Medical Center, Washington, D. C. 2OOOi SUMMARY The relationship between ribonucleotide reductase and tumor growth rate was measured in a series of rat hepatomas of different growth rates. An excellent correlation was found between tumor growth rate and the specific activity of ribo- nucleotide reductase. Differences of ZOO-fold in enzyme specific activities exist between the very fast and the slow growing hepatomas. This represents the largest enhance- ment of any enzyme activity in these tumors. No activity was detected in either normal rat liver or liver from a tumor- bearing animal although l/500 of the enzyme level in the rapid tumor could have been detected. The thioredoxin- thioredoxin reductase complex which serves as the hydrogen donor for this reaction was eliminated as a contributory factor for these differences in activities because this complex is found at comparable levels in both fast growing tumor and normal liver extracts. Two other enzymes involved in deoxyribonucleotide syn- thesis, thymidylate synthetase and thymidine kinase, did not exhibit such a close degree of correlation with tumor growth rate. Thymidylate synthetase activity was found to be less markedly enhanced with only a 15- to 20-fold difference between the slow and fast growing tumors. Thymidine kinase exhibited differences of activity of approximately 40-fold. These data, as well as other evidence, indicate that the ribonucleotide reductase reaction is a rate-limiting step in DNA synthesis and cell division and that enzyme synthesis and degradation may be involved in control of this reaction. DNA synthesis requires the availability of a balanced supply of deoxyribonucleotides. Since deoxyribonucleotides in con- trast to ribonucleotides are found at extremely low levels in animal cells, the reductive conversion of ribonucleotides to deoxyribonucleotides is believed to be a crucial and rate-con- trolling step in the pathway leading to the biosynthesis of DNA. * This work was supported by American Cancer Society Grant P-407 and by United States Public Health Service Grants CA- 10441 and 10729 from the National Cancer Institute. $ To whom reprint requests should be directed. Present ad- dress, Department of, Medicine, Duke University Medical Center, Durham, North Carolina 27706. Ribonucleotide reductase has been well studied in Escherichia coli and Lactobacillus leichmannii but less information is known about the mammalian ribonucleotide reductase (1, 2). However, in all systems studied so far, reduction occurs without breakage of the glycosidic bond, one enzyme complex reduces all four ribonucleotides and reduced disulfhydryl protein serves as the reducing agent. In addition, this reductive event has been shown to be under stringent control by a complex regulatory mechanism involving ATP and deoxyribonucleotide tri- phosphates which serve as both positive and negative allosteric effecters. The present investigation was undertaken to determine whether or not there is a correlation between ribonucleotide reductase activity and tumor growth rate. Such a correlation would provide additional evidence that this reaction plays a critical role in DNA synthesis and cell division and that ribo- nucleotide reductase activity fluctuates greatly, depending on the requirement of the cell for proliferation. The degree of change of the apparent activity of ribonucleotide reductase in relation to other enzymes in deoxyribonucleotide synthesis, thymidylate synthetase and thymidine kinase, was also studied. A preliminary report of this work has been presented (3). EXPERIMENTAL PROCEDURES Materials The unlabeled nucleotides were purchased from Calbiochem and P-L Biochemicals. The dithiothreitol, N-2-hydroxyethyl- piperazine-N-2-ethanesulfonic acid were products of Calbiochem. The c&L-tetrahydrofolic acid was obtained from Sigma. All other chemicals were of reagent grade. Radioactive Biochemicals-The aH-CDP was purchased from Schwarz BioResearch, thymidine-methyl-3H from Schwarz and New England Nuclear, and dUMP-5-aH from Calbiochem. The dUMP-5-aH was purified before use by adsorption onto activated charcoal and elution with 0.1 M NHQH in 60% eth- anol. The eluate was concentrated by vacuum distillation. The concentration and specific activity were adjusted with unlabeled dUMP to the desired value. Tumor-bearing and Control Animals-The Morris solid hepa- tomas were obtained from Dr. H. P. Morris of the National Cancer Institute and Howard University. They were shipped by Air Express sometime after implantation from the Washington, D.C. area to the University of Michigan. The transplantable 5228 by guest on February 16, 2020 http://www.jbc.org/ Downloaded from
Transcript
THE JOURNAL OF Bro~oouxr. CHEMISTRY Vol. 245, No. 20, Issue of October 25, PP. 5228-5233, 1970
Printed in U.S.A.
Ribonucleotide Reductase and Cell Proliferation
I. VARIATIONS OF RIBONUCLEOTIDE REDUCTASE ACTIVITY WITH TUMOR GROWTH RATE IN A SERIES OF RAT HEPATOMAS*
(Received for publication, May 5, 1970)
HOWARD L. ELFORD,$ MICHAEL FREESE, EUGENE PASSAMANI, AND HAROLD P. MORRIS
From the Department of Biological Chemistry, University of Michigan, Ann Arbor, Michigan 48104, and De- partment of Biochemistry, Howard University Medical Center, Washington, D. C. 2OOOi
SUMMARY
The relationship between ribonucleotide reductase and tumor growth rate was measured in a series of rat hepatomas of different growth rates. An excellent correlation was found between tumor growth rate and the specific activity of ribo- nucleotide reductase. Differences of ZOO-fold in enzyme specific activities exist between the very fast and the slow growing hepatomas. This represents the largest enhance- ment of any enzyme activity in these tumors. No activity was detected in either normal rat liver or liver from a tumor- bearing animal although l/500 of the enzyme level in the rapid tumor could have been detected. The thioredoxin- thioredoxin reductase complex which serves as the hydrogen donor for this reaction was eliminated as a contributory factor for these differences in activities because this complex is found at comparable levels in both fast growing tumor and normal liver extracts.
Two other enzymes involved in deoxyribonucleotide syn- thesis, thymidylate synthetase and thymidine kinase, did not exhibit such a close degree of correlation with tumor growth rate. Thymidylate synthetase activity was found to be less markedly enhanced with only a 15- to 20-fold difference between the slow and fast growing tumors. Thymidine kinase exhibited differences of activity of approximately 40-fold. These data, as well as other evidence, indicate that the ribonucleotide reductase reaction is a rate-limiting step in DNA synthesis and cell division and that enzyme synthesis and degradation may be involved in control of this reaction.
DNA synthesis requires the availability of a balanced supply of deoxyribonucleotides. Since deoxyribonucleotides in con- trast to ribonucleotides are found at extremely low levels in animal cells, the reductive conversion of ribonucleotides to deoxyribonucleotides is believed to be a crucial and rate-con- trolling step in the pathway leading to the biosynthesis of DNA.
* This work was supported by American Cancer Society Grant P-407 and by United States Public Health Service Grants CA- 10441 and 10729 from the National Cancer Institute.
$ To whom reprint requests should be directed. Present ad- dress, Department of, Medicine, Duke University Medical Center, Durham, North Carolina 27706.
Ribonucleotide reductase has been well studied in Escherichia coli and Lactobacillus leichmannii but less information is known about the mammalian ribonucleotide reductase (1, 2). However, in all systems studied so far, reduction occurs without breakage of the glycosidic bond, one enzyme complex reduces all four ribonucleotides and reduced disulfhydryl protein serves as the reducing agent. In addition, this reductive event has been shown to be under stringent control by a complex regulatory mechanism involving ATP and deoxyribonucleotide tri- phosphates which serve as both positive and negative allosteric effecters.
The present investigation was undertaken to determine whether or not there is a correlation between ribonucleotide reductase activity and tumor growth rate. Such a correlation would provide additional evidence that this reaction plays a critical role in DNA synthesis and cell division and that ribo- nucleotide reductase activity fluctuates greatly, depending on the requirement of the cell for proliferation. The degree of change of the apparent activity of ribonucleotide reductase in relation to other enzymes in deoxyribonucleotide synthesis, thymidylate synthetase and thymidine kinase, was also studied. A preliminary report of this work has been presented (3).
EXPERIMENTAL PROCEDURES
Materials
The unlabeled nucleotides were purchased from Calbiochem and P-L Biochemicals. The dithiothreitol, N-2-hydroxyethyl- piperazine-N-2-ethanesulfonic acid were products of Calbiochem. The c&L-tetrahydrofolic acid was obtained from Sigma. All other chemicals were of reagent grade.
Radioactive Biochemicals-The aH-CDP was purchased from Schwarz BioResearch, thymidine-methyl-3H from Schwarz and New England Nuclear, and dUMP-5-aH from Calbiochem. The dUMP-5-aH was purified before use by adsorption onto activated charcoal and elution with 0.1 M NHQH in 60% eth- anol. The eluate was concentrated by vacuum distillation. The concentration and specific activity were adjusted with unlabeled dUMP to the desired value.
Tumor-bearing and Control Animals-The Morris solid hepa- tomas were obtained from Dr. H. P. Morris of the National Cancer Institute and Howard University. They were shipped by Air Express sometime after implantation from the Washington, D.C. area to the University of Michigan. The transplantable
Issue of October 25, 1970 Elford, Freese, Passamani, and Morris 5229
Morris hepatomas were carried in Buffalo strain rats except for 3683F and 3924A which were carried in ACI/N rats. The tumors were transplanted bilaterally in the femoral muscle of the host rats, except 3683F and 39248 which were grown by transplantation subcutaneously. The Novikoff tumor was obtained from Dr. C. Wu of the University of Michigan and Dr. S. Weinhouse, Temple University. Dr. Wu also supplied the Walker 256 hepatoma. The Walker and Novikoff tumors were maintained at Michigan by transplantation subcutaneously into male Sprague-Dawley rats over a number of generations. The biology and growth properties of the Morris hepatomas have been described previously (4). The animals were fed ad libitum on Ralston Purina rat chow and housed in air conditioned quarters with 12 hours of light daily.
Methods
Preparation of Cell-free Extracts-Tumor- and nontumor- bearing animals were killed by decapitation at approximately the same time each day to avoid diurnal effects. The tumor was excised from the muscle or skin and washed with cold 0.9% sodium chloride solution. The vital tumor tissue was dis- sected from the necrotic or hemorrhagic tissue and washed again with 0.9% sodium chloride solution, then cut into small pieces and placed in a Potter-Elvehjem homogenizer with an equal volume of potassium phosphate buffer, pH 7.4,0.025 M. Homog- enization was accomplished by the use of a motor driven Potter- Elvehjem homogenizer with 10 strokes with a wait of at least 2 min between sets of 5 strokes. The homogenate was cen- trifuged at 25,000 x g for 30 min. In certain cases, the super- natant solution was recentrifuged in a Beckman model L2 ultra- centrifuge at 104,000 x g for 60 min. The clear supernatant fluids were obtained by withdrawing with a disposable pipette, avoiding a lipid layer floating at the top of the centrifuged tube. The liver extracts from tumor-bearing and control animals were obtained in a similar manner.
Separation of Ribonucleotide Reductase from Thioredoxin- Thioredoxin Reductase Complex-Both tumor and liver extracts were treated in the same manner. All operations were carried out at close to 4”. To the crude extracts was added a 1% solu- tion of protamine sulfate until the ratio of protein to protamine sulfate was 6: 1. The mixture was stirred for 30 min and then centrifuged at 25,000 x g for 30 min. The precipitate was discarded and solid ammonium sulfate was added to 40% satu- ration. The mixture was stirred for 30 min and then centri- fuged for 30 min at 25,000 x g. The precipitate was dissolved in a minimal amount of potassium phosphate buffer, 0.05 M, pH 7.4, usually 10% of the original volume, and dialyzed for 3 hours against the same buffer. This fraction contains the ribonu- cleotide reductase. The supernatant fluid was then brought to 70% saturation with solid ammonium sulfate and stirred slowly for 30 min. The resulting precipitate was collected by centrifugation at 25,000 x g for 30 min, and dissolved in a mini- mum amount of 0.05 M phosphate buffer, pH 7.4. The material precipitated by ammonium sulfate between 40 and 70% of satu- ration was dialyzed for 6 hours against 0.05 M phosphate buffer with one change of buffer. The dialyzed material was dis- tributed in 2.5-ml portions to several tubes and placed in a 70” water bath for 6 min. The denatured material was removed by centrifugation at 25,000 x g for 20 min. This preparation is the souree of the physiological reducing complex.
Ribonucleotide Reductase Assay-The reduction of ribonucleo-
tides to deoxyribonucleotides was measured by monitoring the conversion of cytosine ribonucleotide to the deoxyribonucleotide compound utilizing a slightly modified assay procedure originally developed by Reichard, Baldesten, and Rutberg (5). The incubation mixture contained 0.12 pmole of 3H-CDP (specific activity 3.0 X lo6 to 5.0 x lo7 cpm per pmole), 1.125 pmoles of ATP, 0.50 pmole of MgC&, and 3.0 pmoles of potassium phosphate buffer, pH 7.4, 5.0 pmoles of dithiothreitol, and en- zyme protein between 1 and 4 mg in a total volume of 0.345 ml. Incubation was at 37” for 1 hour. The reaction was terminated by the addition of 1 ml of 1.25 M perchloric acid; the denatured protein was removed by centrifugation and carrier dCMP was added. The deproteinized mixture was hydrolyzed in a boiling water bath for 20 min, cooled and neutralized with 4 N KOH. The KC104 precipitate was removed by centrifugation and the neutralized mixture was applied to a Dowex 50W[H+] column (13.5 x 0.5 cm) and eluted with 0.2 N acetic acid. Four-mil- liliter fractions were collected and each fraction was concentrated to about 0.4 ml by heating. Ten milliliters of a liquid scintil- lation solution (5 g of 2,5-diphenyloxazole and 100 g of recrystal- lized naphthalene dissolved in 1 liter of p-dioxane) were added and the fractions were counted in a Nuclear-Chicago or Picker- Nuclear liquid scintillation spectrometer. Only data obtained from columns exhibiting two distinct radioactive peaks, which cochromatographed with CMP and dCMP were utilized. All the radioactive determinations were corrected for a small amount of radioactivity that was apparent with a no enzyme or zero time control. One unit of activity is defined as that amount of enzyme that catalyzes the formation of 1 ppmole of deoxycyti- dylate in 1 hour per mg of protein.
Thymidylate Synthetase Assay-Thymidylate synthetase ac- tivities were measured essentially by the method of Lomax and Greenberg (6) which involves exchange of aH of dUMP-5-OH into water during the course of the reaction. The modification of Kammen (7) which utilizes the charcoal adsorption and Milli- pore filtration to separate the water from the reaction was em- ployed. The assay mixture contained 25 PM dUMP-5-3H (70,000 to 90,000 cpm per mpmole), 42 mM Tris-HCl (pH 7.4), 26 mM MgC&, 15.8 m&r formaldehyde, 106 m&r 2-mercapto- ethanol, 1.06 mM EDTA, 0.30 mM dl-n-tetrahydrofolate, and enzyme preparation (1 to 2 mg) in a final volume of 0.2 ml. The reaction was initiated by the addition of dUMP-5-aH to 2-min preliminary incubation reaction mixtures. After 10 min of incubation at 25”, the reaction was terminated by the addition of 0.5 ml of a charcoal suspension of 100 mg per ml. Specific activity is defined as the release of 1 ppatom of tritium per mg of protein per hour.
Thymidine Kinase-The capacity of the extracts to phos- phorylate thymidine was determined by the method of Bollum and Potter (8) as modified by Breitman (9). This method was further modified in that we eluted the adsorbed thymidylate from the Whatman DE 20, diethylaminoethyl cellulose paper, with 4 ml of 1 N ammonium hydroxide for 40 min. The paper square was removed, and the ammonium hydroxide eluate con- centrated by heating to a volume of 0.5 ml or less. The radio- activity was determined as described earlier. The incubation mixture contained thymidine-3H, 60 mpmoles (14,750 cpm per mpmole) ; ATP, 400 mpmoles; MgC&, 100 mpmoles; MnClz, 50 mpmoles; N-2-hydroxyethylpiperazine-N-2Tethanesulfonic acid buffer, pH 7.4, 2.4 pmoles; and enzyme in a total volume of 0.07 ml. Incubation was carried out at 37” and 0.02-ml aliquots
5230 Ribmucleotide Reductase and Cell Proliferation. I Vol. 245, No. 20
TABLE I Relationship between ribonucleotide reductase activity
and tumor growth rate The assay is described under “Experimental Procedure.” At
least three enzyme concentration levels over a $-fold range were tested for each tumor extract (25,000 X g supernatant fluid). Only data within the linear portion of the curve were utilized. The mean and range for the different animal tumors are listed.
(1 Based on data from Morris and Wagner (4), H. P. Morris (unpublished), and growth of the tumors at the University of Michigan. Killing was at the transplantation time.
b Tumor 3683F was derived from a frozen tissue bank and yielded a tumor with much less hemorrhage and necrosis than previously seen in this tumor.
c Undetectable 5 3 pprnoles per mg per hour.
were removed at 123 and 25 min and spotted on a Whatman DEAE paper square 1.5 cm2 and treated as described by Breit- man (9) with the modification as previously noted. Specific activity is defined as 1 mpmole nucleotide formed per mg of protein per hour.
Protein concentration in the various preparations was deter- mined by the method of Lowry et al. (10) with the use of crystal- line bovine serum albumin as a standard.
RESULTS
Ribonucleotide Reductase-The relationship between ribonu- cleotide reductase activity and tumor growth rate, as measured by transplantation time, was determined in extracts of a series of rat hepatomas including several hepatomas of the Morris series and livers from normal and tumor-bearing animals. The results shown in Table I indicate a correlation between tumor growth rate and enzymatic activity. No activity could be detected in crude extracts of livers from normal or tumor-bearing rats, where the sensitivity of the assay was such that one one- hundredth of the specific activity of the fast growing tumors could have been detected. Therefore, Novikoff tumor extracts had at least 100 times more ribonucleotide reductase activity than did extracts of rat livers. The activity of extracts of tumors with growth rates of 3 months or longer was very low and could not be detected in every instance. Changing the rnncmtrat,ion of the substrate CDP-3H several-fold higher or
, , l v . , I I ” I I
/
3924 A
GROWTH RATE IN WEEKS
FIG. 1. Relationship between tumor growth rate and ribo- nucleotide reductase specific activity. The experimental condi- tions for the assay is described under “Experimental Procedure” except 20 #Si of CDP-3H of 13.1 Ci per mmole was used in place of CDP-aH of 5 X lo3 or 5 X lo4 cpm per mpmole used in Table I. The assays were performed on crude enzyme preparations (104,000 X g) described under “Methods.” Incubation was 1 hour. The growth rate of rat liver is set at zero. The values are means. The standard deviation is approximately 15% of the mean for extracts prepared from tumors of growth rate of 2 months or less. The standard deviation increases to 3Ooj, for extracts of tumors of growth rates of 3 months or longer.
lower than the optimal concentration determined for the Novi- koff extract did not enhance the activity of the slow growing tumor or liver extracts. The activity was hardly enhanced by varying the concentration of ATP, MgClz, or dithiothreitol, or by treatment of the crude extracts by dialysis, molecular ex- clusion, ion exchange chromatography, or charcoal. In some instances, a 10 to 20% increment above the crude activities were obtained after dialysis.
A second assay was employed in which the sensitivity was increased about 40 times over that of the previous assay. How- ever, suboptimal amounts of substrate were used. The crude extracts were subjected to ultracentrifugation at 104,000 x g for 1 hour in addition to 25,000 x g centrifugation. In this series of experiments with this very sensitive assay, a fairly smooth curve can be drawn through the various points when the log of the specific activity of ribonucleotide reductase is plotted against the log of the tumor growth rate (Fig. 1). Again no activity could be detected in the normal or host rat liver extracts or in the very slow growing tumors, such as 38A. Differences of 500-fold between the rapid growing hepatomas and the meas- urably slower growing hepatoma were seen with this assay; therefore, the activity in liver is at least lOOO-fold less than that of Novikoff or Walker extracts. Fetal liver (17-day fetus) had the highest activity observed, 300,000 cpm per mg or about 3 times higher than that of the ranidlv growing tumors. Results
Issue of October 25, 1970 Elford, Freese, Passamani, and Morris 5231
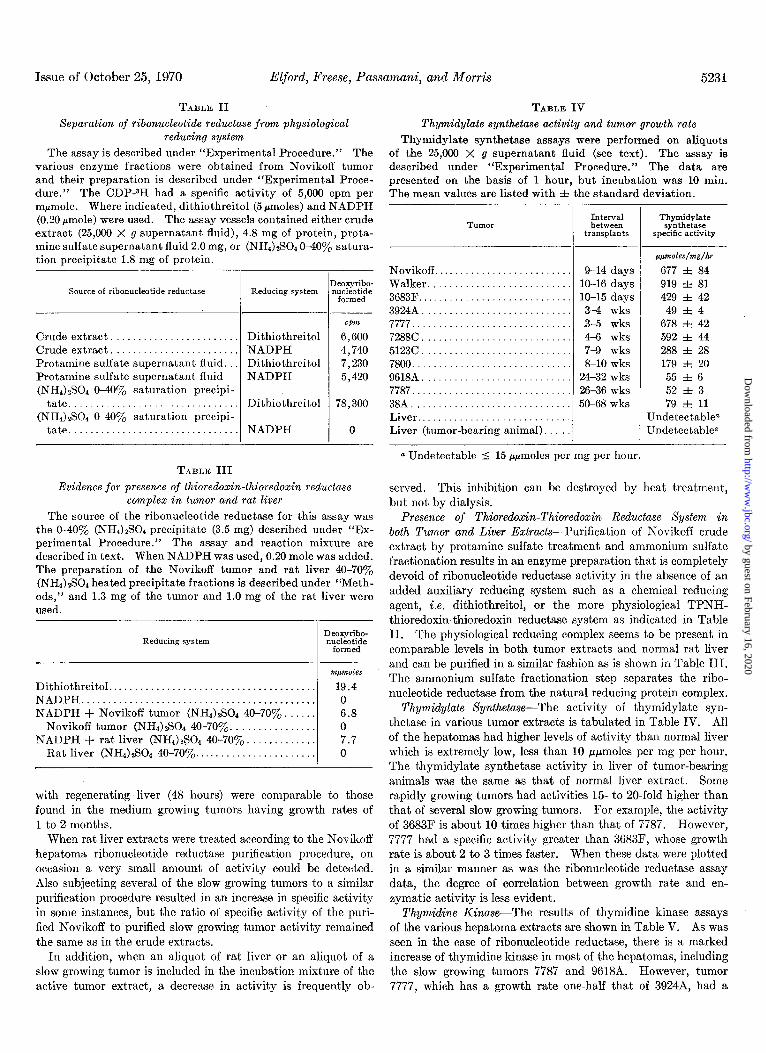
TABLE II TABLE IV
Separation of ribonucleotide reductase from physiological Thymidylate synthetase activity and tumor growth rate reducing system Thymidylate synthetase assays were performed on aliquots
The assay is described under “Experimental Procedure.” The of the 25,000 X g supernatant fluid (see text). The assay is various enzyme fractions were obtained from Novikoff tumor described under “Experimental Procedure.” The data are and their preparation is described under “Experimental Proce- presented on the basis of 1 hour, but incubation was 10 min. dure.” The CDP-3H had a specific activity of 5,000 cpm per The mean values are listed with ZIZ the standard deviation. mpmole. Where indicated, dithiothreitol (5 rmoles) and NADPH (0.20 pmole) were used. The assay vessels contained either crude extract (25,000 X g supernatant fluid), 4.8 mg of protein, prota- mine sulfate suuernatant fluid 2.0 mg. or (NH4)2S04 040% satura- -. tion precipitate 1.8 mg of protein.
9-14 days 10-16 days lo-15 days 3-4 wks 3-5 wks 4-6 wks 7-9 wks 8-10 wks
24-32 wks 26-36 wks 50-68 wks
a Undetectable 4 15 ppmoles per mg per hour.
,qmoles/mg/hr
677 i 84 919 i 81 429 i 42 49 32 4
678 r!z 42 592 i 44 288 3~ 28 179 i 20 55 f 6 52 f 3 79 i 11
Undetectablea Undetectable”
Evidence for presence of thioredoxin-thioredoxin reductase served. This inhibition can be destroyed by heat treatment, complex in tumor and rat liver but not by dialysis.
The source of the ribonucleotide reductase for this assay was Presence of Thioredoxin-Thioredoxin Reductase Xysfem in the 040% (NHI)zS04 precipitate (3.5 mg) described under “I&- both Tumor and Liver Extracts-Purification of Novikoff crude perimental Procedure.” The assay and reaction mixture are described in text. When NADPH was used, 0.20 mole was added.
extract by protamine sulfate treatment and ammonium sulfate
The preparation of the Novikoff tumor and rat liver 40-70yo fractionation results in an enzyme preparation that is completely
(NH4).#04 heated precipitate fractions is described under “Meth- devoid of ribonucleotide reductase activity in the absence of an
ods,” and 1.3 mg of the tumor and 1.0 mg of the rat liver were added auxiliary reducing system such as a chemical reducing
used. agent, i.e. dithiothreitol, or the more physiological TPNH-
with regenerating liver (48 hours) were comparable to those found in the medium growing tumors having growth rates of 1 to 2 months.
When rat liver extracts were treated according to the Novikoff hepatoma ribonucleotide reductase purification procedure, on occasion a very small amount of activity could be detected. Also subjecting several of the slow growing tumors to a similar purification procedure resulted in an increase in specific activity in some instances, but the ratio of specific activity of the puri- fied Novikoff to purified slow growing tumor activity remained the same as in the crude extracts.
In addition, when an aliquot of rat liver or an aliquot of a slow growing tumor is included in the incubation mixture of the active tumor extract, a decrease in activity is frequently ob-
thioredoxin-thioredoxin reductase system as indicated in Table II. The physiological reducing complex seems to be present in comparable levels in both tumor extracts and normal rat liver and can be purified in a similar fashion as is shown in Table III. The ammonium sulfate fractionation step separates the ribo- nucleotide reductase from the natural reducing protein complex.
Thymidylate Xynfhetase-The activity of thymidylate syn- thetase in various tumor extracts is tabulated in Table IV. All of the hepatomas had higher levels of activity than normal liver which is extremely low, less than 10 ppmoles per mg per hour. The thymidylate synthetase activity in liver of tumor-bearing animals was the same as that of normal liver extract. Some rapidly growing tumors had activities 15- to 20.fold higher than that of several slow growing tumors. For example, the activity of 3683F is about 10 times higher than that of 7787. However, 7777 had a specific activity greater than 3683F, whose growth rate is about 2 to 3 times faster. When these data were plotted in a similar manner as was the ribonucleotide reductase assay data, the degree of correlation between growth rate and en- zymatic activity is less evident.
Thymidine Kinase-The results of thymidine kinaee assays of the various hepatoma extracts are shown in Table V. As was seen in the case of ribonucleotide reductase, there is a marked increase of thymidine kinase in most of the hepatomas, including the slow growing tumors 7787 and 9618A. However, tumor 7777, which has a growth rate one-half that of 39248, had a
5232 Ribonucleotide Reductase and Cell Proliferation. I Vol. 245, No. 20
TABLE V Tumor growth rate and thymidine kinase activity
The assay is described in “Experimental Procedure” and was performed on the 25,000 X g supernatant aliquots. The values are presented on the basis of 1 hour although the assay was only for 123 min and 25 min, respectively. The mean values are given with i the standard deviation.
9-14 days 10-16 days 10-15 days 3-4 wks 3-5 wks 4-6 wks 7-9 wks S-10 wks
13-16 wks 24-32 wks 26-36 wks
mpmoles/mg/lir
14.0 z.t 1.6 12.3 i 1.2 10.2 i 1.1 0.8 f 0.1 4.0 f 0.7 2.8 f 0.8 1.2 f 0.2 1.9 f 0.3 2.2 zk 0.4 2.0 f 0.4 0.6 31 0.1
Undetectablea Undetectable”
a Undetectable 5 0.10 mpmole per mg per hour.
specific activity about &fold higher than 39248. Several slow growing tumors had activities only one-twentieth that of the fast growing hepatomas Novikoff, Walker 256, and 3683F. Under the assay conditions employed, no activity was detected in livers from adult or tumor-bearing animals.
DISCUSSION
The results reported in this paper indicate that ribonucleotide reductase activity in crude extracts had a high degree of cor- relation with growth rate in a series of hepatomas, including several of the Morris series. Growth rate, as used in this report, is defined as the average time required for the develop- ment of a viable transplantable tumor (4). An excellent re- lationship between this parameter of tumor growth and develop- ment, and the rate of increase in tumor size has been reported (4, 11).
The data presented here are in accord with the previous findings that DNA synthesis, as measured by thymidine in- corporation into DNA (12) or by crucial enzyme activities in- volved in thymidylate or DNA synthesis, or both, thymidine kinase (13-15), thymidylate synthetase (15, 16), deoxycytidylate deaminase (15-l@, and DNA polymerase (19-21) are markedly elevated in several rat hepatomas. The observations reported herein support the “molecular correlation concept” of Weber (22) that certain key enzymes and metabolic pathways are cor- related with tumor growth rates. The data also fit the hypo- thetical enzyme pattern proposed by Potter (23) for the enzymes involved in cell replication in which he relates normal liver development with “minimal deviation” hepatomas and the more “progressed” hepatomas.
Since DNA synthesis is an obligatory requirement for cellular replication, we planned to determine whether ribonucleotide reductase was regulated to a greater or lesser degree than other enzymes involved in deoxyribonucleotide synthesis, and to ascertain if any of these enzymes were coordinately regulated.
Thymidylate synthetase and thymidine kinase were chosen in addition to ribonucleotide reductase because of the unique role of thymidylate in DNA synthesis.
Thymidine kinase has been examined in several Morris hepa- tomas by Bukovsky and Roth (13)) Bresnick et al. (14), and Sneider, Potter, and Morris (15). Their data, as well as the present study, indicate that this enzyme activity is markedly elevated in the hepatomas and that the increases are related to growth rate. However, several discrepancies were observed, the most notable being the relatively low level of activity found in 39248 in the present study and that 96188 activity is almost as high as 7800, although its growth rate is several-fold faster. It should be noted that thymidine kinase is a salvage pathway enzyme and not a truly synthetic one de novo.
On the other hand, thymidylate synthetase, although studied previously in Morris hepatomas by Sneider et al. (15) and Maley and Maley (16), had not been examined in Morris tumors with growth rates faster than 2 months. It appears that although thymidylate synthetase activity in hepatomas is elevated over the activity found in adult liver, the activity is not as markedly increased as either ribonucleotide reductase or thymidine kinase. The largest observed difference in activities between hepatomas is only about 20-fold, while for ribonucleotide reductase vari- ations of 500-fold have been observed. Also the degree of correlation between growth rate of the tumors and thymidylate synthetase activity is quite limited, especially in the rapidly growing tumors. For example, tumor 5123C has a specific activity 6- to &fold higher than 39248 which has a growth rate approximately 3 times faster than 5123C.
Ribonucleotide reductase, thymidylate synthetase, and thy- midine kinase are all minimally active in adult liver and elevated in tumor tissue; therefore the mechanisms that commit the cells to replication release the restraining influence on all three enzymes. However, as ribonucleotide reductase activity is more closely correlated with tumor growth than the others, these three enzymes do not appear to be coordinately regulated.
One possible variable in ribonucleotide reductase reaction is the reducing component which appears to be similar to the thioredoxin-thioredoxin reductase of E. coli and L. leichmannii (1,2). This reductive complex was eliminated as a possible controlling factor in our results by including a chemical reducing agent, dithiothreitol, in the incubation mixture which would substitute for the natural hydrogen donor. In addition, the experimental evidence reported in Table IV indicates that the reductive complex is found at comparable levels in adult rat liver and Novikoff tumor. This hydrogen donor complex can be also shown in slow growing tumors. The presence of the thioredoxin-like protein portion of this complex has been pre- viously shown in rat liver (24). Thus it seems that the thio- redoxin-thioredoxin reductase complex is not the limiting factor which could account for the large variation of ribonucleotide reductase activity between the various hepatomas. It also suggests that there are other functions for the thioredoxin- thioredoxin reductase complex.
The question arises whether the differences in ribonucleotide reductase activity we observed between the various tumors are the consequence of an alteration in the enzyme from an in- active to active state as mediated by allosteric agents, or whether they represent an increment in enzyme synthesis. The work of Turner, Abrams, and Lieberman (25) provide some informa- tion relating to this question. They found that in synchronized
Issue of October 25, 1970 Elford, Freese, Passamani, and ibforris 5233
cultures of mouse L-cells, ribonucleotide reductase activity is in rapidly proliferating tumors represents the most striking detected only in the S-phase of the cell cycle and that the enzyme elevation of activity of any enzyme studied so far in the Morris activity decreases rapidly with a half-life of 2 hours when cyclo- hepatomas. This increase is also greater than that shown for heximide, a protein synthesis inhibitor, is added to the cultures. incorporation of radioactive thymidine into DNA. Therefore, In addition, Elford has found’ that cycloheximide prevents the ribonucleotide reductase could have considerable interest as a large increase in ribonucleotide reductase activity observed in barometer of malignancy, and the reaction is a logical target for the first few days of neonatal development of rat spleen and chemotherapy. thymus. Therefore it is our belief that the gross changes in enzyme activity observed in the present series of tumors represent different rates of enzyme synthesis and degradation in response to the cell requirement for DNA synthesis rather than to an
REFERENCES 1. REICHARD, P., Eur. J. Biochem., 3, 259 (1968). 2. LARSSON, A., AND REICHARD, P., in J. N. DAVIDSON AND W.
E. COHN (Editors), Progress in nucleic acid research. Vol. 7.
alteration in the state of the enzymes from an inactive to an Academic Press, New York, 1967, p. 303.
active state. 3. ELFORD, H. L., Fed. Proc., 27, 300 (1968).
The allosteric control of this reaction exerted by the nucleo- 4. MORRIS, H. P., AND WAGNER, B. P., in H. BUSCH (Editor),
tide effecters, ATP, and deoxyribonucleotide triphosphates, Methods in cancer research, Vol. 4, Academic Press, New York, 1968. D. 125.
which alter the specificity of the enzyme, probably exert a fine control on the reaction by assuring that a balanced supply of deoxyribonucleotides is synthesized and preventing an excess supply of free deoxyribonucleotides. However, to affect a rapid increment in deoxyribonucleotide synthesis, additional mech- anisms must be involved. Evidence obtained in this laboratory suggests that synthesis of ribonucleotide reductase enzyme is a major contributor to the ability to synthesize deoxyribonu- cleotides at an adequate rate to allow sufficient DNA synthesis to occur in rapidly proliferating tissue. An additional factor could be the existence of a specific inhibitor of the enzyme in- dicated by the inhibitory material seen in adult liver extracts and slow growing tumors.
The assumption has been made that mammalian ribonucleotide reductase systems are similar to E. co& and L. Zeichmannii re- ductase systems in that a single enzyme complex catalyzes the reduction of all four ribonucleotides, and the specificity for a particular ribonucleotide is governed by nucleotide allosteric effecters (26). However, this premise requires further experi- mental examination.
The large differences in ribonucleotide reductase activity that we observed do not appear to be the consequence of nucleotides or other low molecular weight substances in the extracts acting as effecters because treatments such as dialysis, molecular exclusion, and ion exchange chromatography did not result in a significant change in activity. However, it is possible that a negative effector could have been too tightly bound to be re- moved by these treatments. It is difficult by enzyme assay to differentiate between quantity and state of the enzyme. Studies to distinguish between these alternatives are being conducted.
The marked increase in activity of ribonucleotide reductase
1 H. L. Elford, manuscript in preparation,
5. REICHA~D, P.; BALDESTEN, A., AND RUTBERG, L., J. Biol. Chem., 236, 1150 (1961).
6. LOMAX, M. I. S., AND GREENBERG, G. R., J. Biol. Chem., 242, 109 (1967).
7. KAMMEN, H. O., Anal. Biochem., 17, 553 (1966). 8. BOLLIJM, F. J., AND POTTER, V. R., Cancer Res., 19, 561 (1959). 9. BREITMAN, T. R., Biochim. Biophys. Acta, 67, 153 (1963).
10. LOWRY, 0. H., ROSEBROUGH, N. J., FARR, A. L., AND RANDALL, R. J., J. Biol. Chem., 193, 265 (1951).
11. LINDER-HOROWITZ, M., KNOX, W. E., AND MORRIS, H. P., Cancer Res., 29, 1195 (1969).
12. LEA, M. A., MORRIS, H. P., AND WEBER, G., Cancer Res., 26, 465 (1966).
13. BUKOVSKY, J., AND ROTH, J. S., Cancer Res., 26, 358 (1965). 14. BRESNICK, E., THOMPSON, U. B., MORRIS, H. P., AND LIEBELT,
A. G., Biochem. Biophys. Res. Commun., 16, 278 (1964). 15. SNEIDER, T. W., POTTER, V. R., AND MORRIS, H. P., Cancer
Res., 29, 40 (1969). 16. NIALEY, F., AND MALEY, G. F., Cancer Res., 21, 1421 (1961). 17. POTTER, V. R., PITOT, H. C., ONO, T., AND MORRIS, H. P.,
Cancer Res., 20,1255 (1960). 18. ROTH, J. S., SHEID, B., AND MORRIS, H. P., Cancer Res., 23,
454 (1963‘). 19. WHEELER, G. P., ALEXANDER, J. A., HILL, D. D., AND MORRIS,
H. P.. Cancer Res., 26, 2470 (1966). 20. IWAMU~A, Y., ONO, T., AND GORR&, H. P., Cancer Res., 23,
2466 (1968). 21. OVE, P., LASZLO, J., JENKINS, M. D., AND MORRIS. H. P..
Cancer Res., 29; 1557 (1969). ’ 22. WEBER, G., Gann Monoar.. 1. 151 (1966). 23. POTTER, V.‘R., AND WATANABE, Ml in~b. J. D. ZARAFONETIS
(Editor), Proceedings of International Conference on Leu- kemia-Lymphoma, Lea and Febiger Publishing Company, Philadelphia, 1968, p. 33.
24. MOORE, E. C., AND HURLBERT, R. B., Biochim. Biophys. Acta, 66, 651 (1962).
25. TURNER, M. K., ABRAMS, R., AND LIEBERMAN, I., J. Biol. Chem., 243, 3725 (1968).
26. MOORE, E. C., AND HURLBERT, R. B., J. Biol. Chem., 241.4802 (1966).