1
Role of FimW, FimY, and FimZ in regulating the expression of type I 1
fimbriae in Salmonella enterica serovar Typhimurium 2
3
Supreet Saini§, Jeffrey A. Pearl§, and Christopher V. Rao* 4
5
Department of Chemical and Biomolecular Engineering 6
University of Illinois at Urbana-Champaign 7
Urbana, Illinois, United States, 61801 8
9
10
Running Title: Regulation of type I fimbriation in S. typhimurium 11
Key words: Type I fimbriae, FimW, FimY, FimZ, Gene regulation 12
13
§ Authors contributed equally to this work 14
15
* Corresponding author. Department of Chemical and Biomolecular Engineering, 16
University of Illinois at Urbana-Champaign, 600 S. Mathews Ave, Urbana, IL, United 17
States, 61810. Phone: (217) 244-2247. Fax: (217) 333-5052. Email: 18
[email protected]. 19
Copyright © 2009, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.J. Bacteriol. doi:10.1128/JB.01694-08 JB Accepts, published online ahead of print on 13 February 2009
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
2
Abstract 20
Type I fimbriae in Salmonella enterica serovar Typhimurium are surface appendages 21
that facilitate binding to eukaryotic cells. Expression of the fim gene cluster is known to 22
be regulated by three proteins – FimW, FimY, and FimZ – and a tRNA encoded by 23
fimU. In this work, we investigated how these proteins and tRNA coordinately regulate 24
fim gene expression. Our results indicate that FimY and FimZ independently activate 25
the PfimA promoter which controls the expression of the fim structural genes. FimY and 26
FimZ were also found to strongly activate each other’s expression and weakly activate 27
their own expression. FimW was found to negatively regulate fim gene expression by 28
repressing transcription from the PfimY promoter, independent of FimY or FimZ. 29
Moreover, FimW and FimY interact within a negative feedback loop as FimY was found 30
to activate the PfimW promoter. In the case of fimU, expression of this gene was not 31
found to be regulated by FimW, FimY, or FimZ. We also explored the effect of fim gene 32
expression on Salmonella Pathogenecity Island 1 (SPI1). Our results indicate that FimZ 33
alone is able to enhance the expression of hilE, a known repressor of SPI1 gene 34
expression. Based on our results, we were able to propose an integrated model for the 35
fim gene circuit. As this model involves a combination of positive and negative 36
feedback, we hypothesized that the response of this circuit may be bistable and thus a 37
possible mechanism for phase variation. However, we found that the response was 38
continuous and not bistable. 39
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
3
Introduction 40
Type I fimbriae in Salmonella enterica serovar Typhimurium (S. typhimurium) are 41
proteinaceous surface appendages that carry adhesions specific for mannosylated 42
glycoproteins (9). Type I fimbriae are involved in S. typhimurium pathogenicity by 43
facilitating the binding to and invasion of intestinal epithelial cells (44). In orally 44
inoculated mice, a wild-type strain has been shown to cause more infections and deaths 45
than a fim- mutant strain (19). A fim- mutant has also been shown to exhibit several-fold 46
weaker binding to HEp-2 and HeLa cells, and the defect in binding could be restored by 47
complementing the fim system on a plasmid (4). Apart from type I fimbriae, mutations in 48
different Salmonella fimbrial systems - lpf, pef, and agf - have all been also shown to 49
greatly reduce virulence in mice (48). These systems appear to work synergistically in 50
order to facilitate colonization of the ileum (5). In S. typhimurium, the fim gene cluster 51
possesses all of the genes necessary for type I fimbrial production. This gene cluster is 52
composed of six structural genes, three regulators, and a tRNA specific for rare arginine 53
codons (AGA and AGG). The structural genes - fimA, fimI, fimC, fimD, fimH, and fimF - 54
are all expressed in one transcript from the PfimA promoter (27, 37-39). The regulators - 55
fimZ, fimY, and fimW - are all expressed from independent promoters (45, 47, 49). The 56
tRNA encoded by fimU is located at one end of the cluster and is required for the 57
effective translation of the regulatory genes that all carry rare arginine codons (43). 58
Type I fimbriation is environmentally regulated with fim gene expression favored in 59
static liquid media, whereas growth on solid media inhibits expression (17). Moreover, 60
S. typhimurium cultures in fimbriae-inducing conditions contain cells in both fimbriated 61
and non-fimbriated states (36). While the regulation of fim gene expression has been 62
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
4
studied extensively in Escherichia coli, far less is known about the regulation in S. 63
typhimurium (1, 28). In particular, despite homology between the structural genes for 64
type I fimbriae in E. coli and S. typhimurium, their expression is regulated in completely 65
different manners. No homologs of E. coli regulators, FimB and FimE, are present in S. 66
typhimurium (25, 29). Also, the S. typhimurium PfimA promoter is inactive in E. coli, 67
indicating that the PfimA promoter is regulated by different factors in these two organisms 68
(49). In S. typhimurium, the expression of the structural genes is regulated by three 69
transcription factors: FimY, FimZ, and FimW (45, 47, 49). Both FimZ and FimY are 70
essential for expression of the structural genes from the PfimA promoter (49). In 71
particular, deletion of either the fimY or fimZ gene reduces expression from the PfimA 72
promoter and prevents S. typhimurium from making type I fimbriae. FimZ has been 73
shown to bind the PfimA promoter and promote transcription (13, 49). FimY, on the other 74
hand, is thought to facilitate activation of the PfimA promoter as direct binding has not 75
been observed (45). FimW is a negative regulator of fim gene expression (46). FimW 76
has also been suggested to auto regulate its expression as enhanced PfimW activity has 77
been observed in ∆fimW mutant. In DNA-binding assays, FimW was not observed to 78
bind any of the fim promoters. However, FimW was found to interact with FimZ in a 79
LexA-based two-hybrid system in E. coli (46). Thus, a possible mechanism for FimW-80
mediated repression may be that it binds FimZ and prevents it from activating 81
transcription. However, analysis for FimW’s amino acid sequence predicts that it has a 82
DNA-binding domain. Moreover, it is related to a broad range of prokaryotic 83
transcription factors, with its closest relatives being BpdT from Rhodococcus and an 84
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
5
uncharacterized response regulator, TodD, from Pseudomonas putida (30, 31). Thus, 85
FimW may also act by an alternate mechanism involving DNA binding. 86
In addition to these transcription factors, the fimU tRNA also plays a role in fim gene 87
expression (43). All three regulators - FimZ, FimY, and FimW - contain a number of the 88
rare arginine codons, AGA and AGG, recognized by the fimU tRNA. In the case of 89
FimY, ∆fimU mutants have been shown to be non-fimbriated due to inefficient 90
translation of fimY mRNA. This translational regulation results from FimY having three 91
rare arginine codons within its first 14 amino acids. The phenotypic effect of ∆fimU 92
mutation could, however, be overcome by expressing fimU from a plasmid or by 93
changing these three rare arginine codons in fimY to ones more efficiently translated. 94
As a pathogen, S. typhimurium invades host cells by a process in which effector 95
proteins are injected into the target cells with the help of the Salmonella Pathogenicity 96
Island 1 (SPI1) type III secretion system (T3SS) (12, 14). SPI1 gene expression is 97
regulated by a number of proteins, with the critical activator being HilA (2). Expression 98
of hilA, in turn, is regulated by three AraC-like transcriptional activators: hilC, hilD, and 99
rtsA (20, 22, 23, 33, 41, 42). HilD activity is controlled by HilE; this protein binds HilD 100
and is thought to prevent it from activating the PhilA promoter (6, 8). FimY and FimZ 101
have been previously shown to regulate SPI1 gene expression by repressing hilA 102
expression through their activation of the PhilE promoter (7). 103
In this work, we investigated the gene circuit regulating fim expression. Using 104
genetic approaches, we found that FimZ and FimY activate each other’s expression, 105
and that each protein can independently activate the PfimA promoter. Moreover, FimZ 106
and FimY were found to be weak auto-activators. Our data also suggests that FimW-107
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
6
mediated repression occurs at the level of fimY transcription. With regards to fimU, we 108
found that none of the fim regulatory genes had any effect on its transcription. As the 109
fim gene circuit involves a combination of positive and negative feedback, we tested 110
whether induction was bistable. However, we found the cell population responded 111
homogeneously when induced. Finally, we looked at the link between the fim and SPI1 112
gene circuits and found that the PhilE promoter is activated solely by FimZ. Collectively, 113
these results allow us to propose an integrated model for the regulation of the fim gene 114
circuit in S. typhimurium. 115
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
7
Materials and Methods 116
General Techniques and Growth Conditions. All culture experiments were 117
performed in Luria-Bertani (LB) broth (tryptone: 10 g/l, yeast extract: 5 g/l, and NaCl: 10 118
g/l) at 37˚C unless otherwise noted. Antibiotics were used at the following 119
concentrations: ampicillin at 100 µg/ml, chloramphenicol at 20 µg/ml, and kanamycin at 120
40 µg/ml. All experiments involving growth of cells carrying pKD46 were performed at 121
30˚C as previously described (15). Loss of the helper plasmid pKD46 was achieved by 122
growth in non-selective conditions on LB agar at 42˚C. Removal of the antibiotic 123
cassette from the FRT-Cm/Kan-FRT insert was obtained by transformation of pCP20 124
into the respective strain and selection on ampicillin at 30˚C. Loss of the helper plasmid 125
pCP20 was obtained by growth at 42˚C under non-selective conditions on LB agar (10). 126
Integrations into the λattB sites of the S. typhimurium and E. coli genomes were done 127
using the helper plasmid, pInt-ts, as described previously (26). Loss of the helper 128
plasmid, pInt-ts, was obtained by growth at 42˚C under non-selective conditions. 129
Primers were purchased from IDT Inc. Enzymes were purchased from New England 130
Biolabs and Fermentas and used according to the manufacturer’s recommendations. 131
Strain and Plasmid Construction. All bacterial strains and plasmids used in this 132
study are described in Tables 1 and 2, respectively. All S. enterica serovar Typhimurium 133
are isogenic derivatives of strain 14028 (American Type Culture Collection, ATCC). The 134
generalized transducing phage of S. typhimurium P22 HT105/1int-201 was used in all 135
transductional crosses (16). 136
The plasmids pKD3 or pKD4 were used as templates to generate scarred FLP 137
recombinant target (FRT) mutants as described previously (14). The ∆fimYZ mutant 138
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
8
was made using primers SS105F and SS105R. The ∆fimZ mutant was made using 139
primers SS105FII and SS105F. The ∆fimY mutant was made using the primers 140
SS105RII and SS105R. The ∆fimW mutant was made using primers SS152F and 141
SS152R. The ∆fimU mutant was made using the primers SS165F and SS165R. All 142
mutations were checked by PCR using primers that bound outside the deleted region. 143
Prior to removal of the antibiotic resistance marker, the constructs resulting from this 144
procedure were moved into a clean wild-type background (14028) by P22 transduction. 145
In order to construct the fluorescent Venus reporter plasmid (35), PCR was used to 146
amplify Venus from pBS7 using primers LC294F and LC296R. The resulting PCR 147
product was used as a template with primers LC295F and LC296R to add three out of 148
frame stop codons and a synthetic Shine-Dalgarno sequence before the Venus start 149
codon. The resultant PCR product was then digested with EcoRI and HindIII and sub-150
cloned into the EcoRI and HindIII cut-sites of pQE80L (Qiagen), yielding pQE80L-151
Venus. The plasmid pQE80L was digested with EcoRI and NheI, and the fragment was 152
cloned into the conditional-replication, integration, and modular (CRIM) plasmid pAH125 153
digested with EcoRI and NheI (26). The resulting CRIM plasmid was called pVenus. 154
Venus transcriptional fusions were made by amplifying the promoter of interest and then 155
cloning these PCR fragments into the multiple cloning site of pVenus. The fimA 156
transcriptional fusion was made using primers SS104F and SS104R. The fimY 157
transcriptional fusion was made using primers SS037F and SS037R. The fimZ 158
transcriptional fusion was made using primers SS103F and SS103R. The fimW 159
transcriptional fusion was made using primers SS154F and SS154R. The fimU 160
transcriptional fusion was made using primers SS162F and SS162R. The hilE 161
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
9
transcriptional fusion was made using primers SS024F and SS024R. The PCR 162
fragments were then digested with KpnI and EcoRI (sequences underlined) and cloned 163
into the multiple cloning site of the pVenus vector. The resulting transcriptional fusions 164
were integrated into the S. typhimurium and E. coli chromosomes at the λattB site using 165
λInt produced from the CRIM helper plasmid pInt-ts, thus creating single-copy 166
transcriptional fusions. In the case of S. typhimurium, the integrated plasmid was moved 167
into different mutant strains by P22 transduction. 168
Expression plasmids for fimY, fimZ, and fimW were made by cloning the respective 169
gene into the multiple cloning site of pPROTet.E (Clontech) under the control of a strong 170
promoter PLTetO1, resulting in plasmids pFimY, pFimZ, and pFimW (34). The plasmid 171
pFimZ was made first by amplifying the fimZ gene using the primers SS106F and 172
SS106R. The PCR product was then digested with EcoRI and KpnI (sequence 173
underlined) and cloned into pPROTet.E. The plasmid pFimY was made by amplifying 174
the fimY gene using the primers SS107F and SS107R. The PCR product was then 175
digested with SalI and BamHI (sequence underlined) and cloned into pPROTet.E. The 176
plasmid pFimW was made by amplifying the fimW gene using the primers SS160F and 177
SS160R. The PCR product was then digested with EcoRI and HindIII (sequence 178
underlined) and cloned into pPROTet.E. In order to mutate the first three arginine rare 179
codons at position 7, 9, and 14 in fimY, primers SS162F and SS107R were used to 180
amplify fimY with the rare arginine codons mutated to consensus arginine codons. The 181
resulting PCR product was used as a template with primers SS167F and SS107R. The 182
amplified product was digested with EcoRI and BamHI and cloned into the multiple 183
cloning site of pPROTet.E. The plasmid is called pFimY*. 184
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
10
In our expression plasmids, in the absence of TetR, the PLTetO-1 promoter is 185
constitutively active. To regulate the expression levels from the PLTetO-1 promoter, the 186
tetR gene was also cloned downstream of the gene target into the plasmids as 187
previously described (40). In this arrangement, in absence of the inducer 188
anhydrotetracycline (aTc), expression from the promoter is inhibited due to TetR. The 189
inhibition, however, is relieved upon addition of 100 ng/ml of aTc, and expression from 190
the PLTetO-1 promoter then takes place. All constructs were sequenced prior to 191
transforming into wild type and mutant strains. The sequences for all the primers used 192
in this study are given in Table 3. 193
Fluorescence Assays. As an indirect measure of gene expression, end-point and 194
dynamic measurement of the fluorescent reporter system were made using a Tecan 195
Safire2 microplate reader. For fluorescence end-point measurements, 1 ml culture was 196
grown at 37˚C overnight and then sub-cultured 1:1000 into fresh media and grown in 197
static conditions for 24 hours at 37˚C. 100 µL of the culture was then transferred into a 198
96 well microplate, and the relative fluorescence and optical density at 600 nm (OD600) 199
measured. The fluorescence readings were normalized with the OD600 absorbance to 200
account for cell density. For time-course measurements, overnight cultures at 37˚C 201
were sub-cultured to an OD of 0.05 into fresh medium and allowed to grow to an OD 202
0.15. 100 µL of the culture was then transferred into a 96 well microplate and overlaid 203
with 25 µL of oil to prevent evaporation. The temperature was maintained at 37˚C, and 204
fluorescence and optical density readings were taken every 5 minutes. All experiments 205
were done in triplicate and average values with the standard deviations reported. 206
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
11
Single-cell measurements were done similarly by growing the cells in non-inducing 207
conditions with vigorous shaking at 37˚C. Overnight cultures were sub-cultured to an 208
OD of 0.05 into fresh media (LB) and grown in inducing conditions of high oxygen and 209
no shaking at 37˚C. Samples were collected at different time points by spinning the cells 210
down, resuspending in phosphate buffered saline (PBS) supplemented with 211
chloramphenicol (34 µg/ml) to stop all translation and arrest cells in their respective 212
state, and finally storing on ice. All flow cytometry experiments were performed on a BD 213
LRS II system (BD Biosciences). Data extraction and analysis for the flow cytometry 214
experiments was done using FCS Express Version 3 (De Novo Software). 215
216
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
12
Results 217
FimZ and FimY are activators and FimW is a repressor of fim gene expression 218
FimZ and FimY have previously been reported as activators of fim gene expression 219
in S. typhimurium (45, 49). Both have also been reported as essential for fimbriation as 220
deletion of either one results in loss of expression from the PfimA promoter (50). To 221
understand the roles of FimZ and FimY in the fim gene circuit, we measured expression 222
from the PfimA, PfimZ, PfimY, and PfimW promoters in wild type and ∆fimZ, ∆fimY, ∆fimYZ, 223
and ∆fimW mutants (Figure 1). Chromosomally-integrated Venus transcriptional 224
reporters were employed as indirect measures of promoter activities (35). In the cases 225
of all four promoters, activity levels were found to be about two times less active in 226
∆fimZ, ∆fimY, and ∆fimYZ mutants relative to wild type. For all four promoters, note that 227
no further reduction in promoter activity was observed in the double mutant. In a ∆fimW 228
mutant, the activities of all four promoters were approximately two times higher than 229
wild-type levels. While these results agree with previously published data regarding the 230
fim system in S. typhimurium, they still do not tell us how FimW, FimY, and FimZ 231
individually contribute to PfimA activation. 232
233
FimY and FimZ are strong activators of each other’s expression and weak 234
activators of their own expression. 235
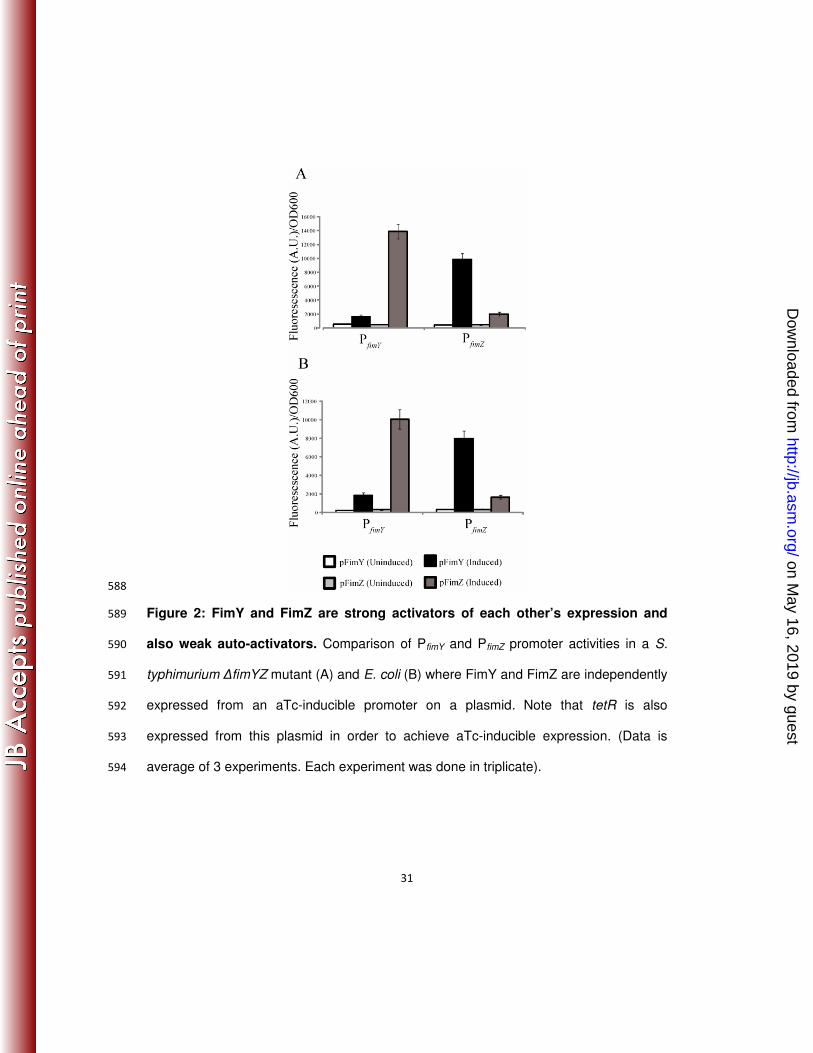
To determine the relative effect of FimY and FimZ on fim gene expression, PfimY and 236
PfimZ promoter activities were measured in a ∆fimYZ mutant where either FimZ or FimY 237
was expressed from a strong, aTc-inducible promoter on a plasmid (see Materials and 238
Methods). Using this system, we found that expressing FimZ in the ∆fimYZ mutant led 239
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
13
to a more than ten-fold increase in PfimY activity (Figure 2a). Likewise, expressing FimY 240
in the ∆fimYZ mutant led to about a ten-fold increase in PfimZ levels. In addition to their 241
ability to activate each other’s promoters, FimY and FimZ were found to increase 242
expression from their own promoters roughly three fold. 243
Even though E. coli makes type I fimbriae, the S. typhimurium fim promoters by 244
themselves are inactive in this organism. Therefore, we performed an identical set of 245
experiments in E. coli using the S. typhimurium proteins and promoters. Overall, the 246
results were identical to those in E. coli (Figure 2b). In particular, FimZ expression led 247
to a more than ten-fold increase in PfimY promoter activity and FimY expression led to a 248
ten-fold increase in PfimZ activity. Both FimZ and FimY were again found to weakly 249
activate expression from their own promoters. The goal of these experiments was to 250
remove the affect of any S. typhimurium specific regulatory mechanisms, thus allowing 251
us to more confidently conclude that the observed results are due to direct interactions. 252
Collectively, these results show that FimY and FimZ strongly activate each other’s 253
expression and weakly activate their own expression. This cross regulation also 254
explains why both FimY and FimZ are required for strong PfimA promoter activity, as the 255
expression of each is dependent on the other. 256
257
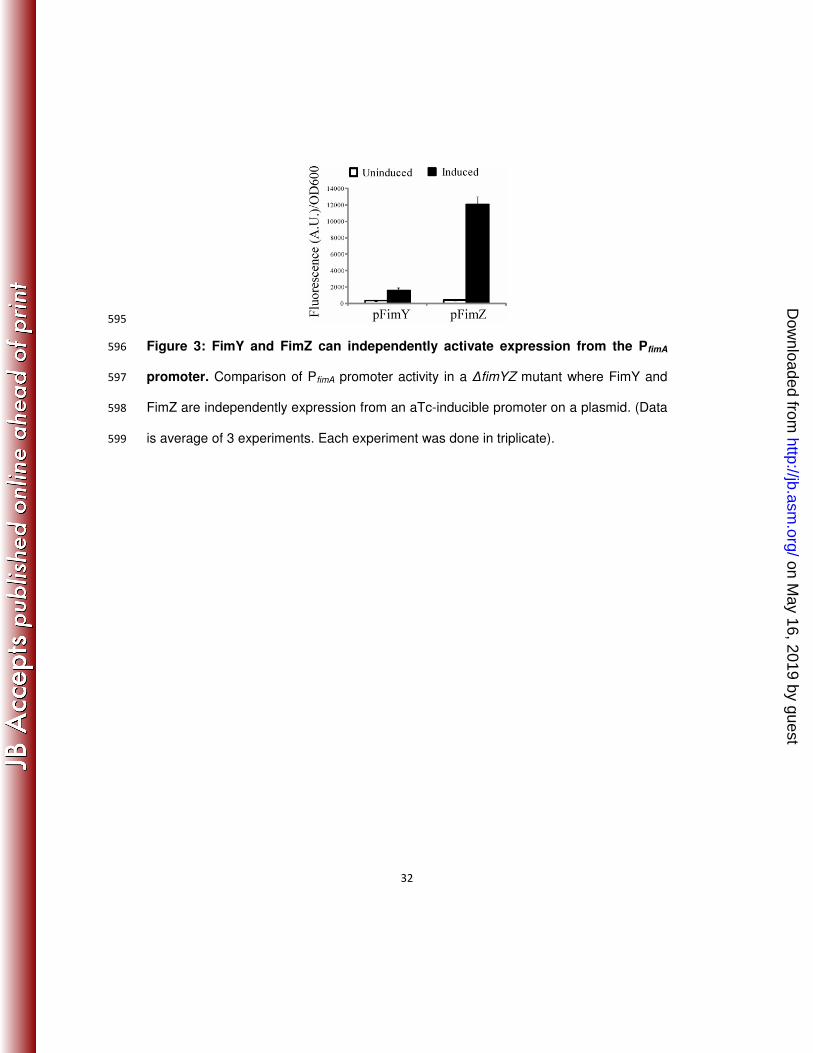
FimZ and FimY can independently activate expression from PfimA promoter 258
Next, we looked at how FimZ and FimY independently effected PfimA expression. To 259
investigate this problem, we measured PfimA promoter activity in a ∆fimYZ mutant where 260
either FimY or FimZ was expressed using the aTc-inducible system. FimZ expression 261
was found to strongly activate (>15 fold) the PfimA promoter whereas FimY could only 262
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
14
weakly activate (>2 fold) it (Figure 3). We also performed these experiments in E. coli 263
with similar results (data not shown). Based on these results, we conclude that FimZ 264
and FimY can both independently activate the PfimA promoter. In the case of FimY, weak 265
activation of the PfimA promoter is likely due to its strong dependence on the fimU tRNA 266
(see below) (43). 267
268
FimY activates the PfimW promoter and FimW represses the PfimY promoter 269
FimW has previously been observed to repress fim gene expression (46). Consistent 270
with these results, we observed that PfimA, PfimW, PfimY, and PfimZ promoter activities were 271
all elevated in a ∆fimW mutant (Figure 1). To understand the mechanism of FimW-272
mediated repression, we first sought to identify the proteins that regulate expression 273
from the PfimW promoter. To answer this question, we measured expression from the 274
PfimW promoter in a ∆fimYZ mutant where FimW, FimY, and FimZ were independently 275
expressed using the aTc-inducible system. In the case of FimW and FimZ, expression 276
had no effect on PfimW promoter activity (data not shown). However, in the case of FimY, 277
we observed a significant increase in PfimW promoter activity (1052 +/- 381 RFU/OD 278
[uninduced] versus 14718 +/- 1032 RFU/OD [induced]). Similar results were also 279
obtained when these experiments were performed in E. coli (data not shown). To 280
identify the regulatory targets of FimW, we measured the expression of the PfimA, PfimZ, 281
and PfimY promoters in a ∆fimW ∆fimYZ mutant where FimW was expressed using the 282
aTc-inducible system. In the case of the PfimA and PfimZ promoters, we found that FimW 283
expression had no effect. However, in the case of the PfimY promoter, FimW expression 284
led to about a three-fold decrease in PfimY activity (PfimY: 7462 +/- 319 RFU/OD 285
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
15
[uninduced] versus 2781 +/- 188 RFU/OD [induced]). Based on these results, we 286
conclude that FimY activates expression from the PfimW promoter and that FimW 287
represses expression from the PfimY promoter. 288
289
The PfimU promoter is not regulated by FimW, FimY, or FimZ 290
Both fimY and fimZ contain rare arginine codons (AGA and AGG) and need fimU, a 291
tRNA specific for rare arginine codons, for effective translation. In a ∆fimU mutant, PfimA 292
activity was less than ten-fold as compared to the wild type levels (wild type: 16723 +/- 293
1173 RFU/OD; ∆fimU: 1389 +/- 261 RFU/OD). Expression of FimY in the ∆fimU mutant 294
using the aTc-inducible system, however, did not increase PfimA activity (988 +/- 319 295
[uninduced] versus 1343 +/- 166 [induced]). Replacing the rare arginine codons in the 296
fimY gene with consensus ones did restore PfimA activity to wild-type levels (817 +/- 73 297
RFU/OD [uninduced] versus 11294 +/- 462 RFU/OD [induced]). These experiments are 298
consistent with previously published results (46), and indicate that fimU is essential for 299
effective fimY translation. 300
As fimU has a strong effect on PfimA promoter activity, we hypothesized that it may 301
be subject to regulation by the other proteins within the circuit. To test this hypothesis, 302
we measured PfimU promoter activity in different regulatory mutants. Contrary to our 303
hypothesis, we did not observe any change in PfimU promoter activity in any mutant (wild 304
type: 26717 +/- 1381 RFU/OD; ∆fimZ: 28991 +/- 2164 RFU/OD; ∆fimY: 25884 +/- 1983 305
RFU/OD; ∆fimYZ: 26516 +/- 1772 RFU/OD; and ∆fimW: 24829 +/- 2073 RFU/OD). 306
Likewise, we did not observe any change in PfimU promoter activity when FimW, FimY, 307
and FimZ were expressed using the aTc-inducible system in wild-type S. typhimurium or 308
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
16
E. coli (data not shown). Based on these results, we conclude that the PfimU promoter is 309
not regulated by any fim protein. 310
311
FimZ alone is able to regulate SPI1 gene expression 312
Previous studies have shown that both FimY and FimZ regulate SPI1 expression 313
through their activation of the PhilE promoter (7). HilE, in turn, is known to bind HilD and 314
repress HilD-mediated activation of PhilA, PhilC, PrtsA, and PhilD promoters (6, 21). To test 315
which protein activates the PhilE promoter, we independently expressed FimY and FimZ 316
in a ∆fimYZ mutant using the aTc-inducible system and then measured expression from 317
the PhilE promoter. Of the two, only FimZ was found to affect PhilE expression: 1089 +/- 318
421 RFU/OD [uninduced] versus 17654 +/- 2234 RFU/OD [induced]. Similar results 319
were also observed in E. coli (data not shown). 320
We note that these results are contrary to those previously reported, where it was 321
shown that both FimY and FimZ were necessary for activation of the PhilE promoter (7). 322
One possible explanation for the discrepancy involves how the two gene were 323
selectively expressed. In the original study, a DNA fragment containing the fimYZ gene 324
cluster was cloned onto a plasmid and expressed using the tetracycline promoter. To 325
study their relative effect, each gene was selectively inactivated using a universal 326
promoter. As part of the PfimY and the whole PfimZ promoter were left intact in their 327
construct, transcriptional inference may have occurred between the various promoters. 328
In our design, we selectively cloned each gene and then expressed it from an inducible 329
promoter, eliminating any potential interfering effects from having the native promoters 330
still present. 331
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
17
Dynamics of fim gene expression. 332
Finally, we wished to investigate the dynamics of fim gene expression. We first 333
measured PfimA promoter activity using a microplate reader in wild type and ∆fimY, 334
∆fimZ, ∆fimYZ, and ∆fimW mutants (Figure 4a). Consistent with our end-point 335
measurements, we found that the PfimA promoter was weakly expressed in the ∆fimY, 336
∆fimZ, and ∆fimYZ mutants. Likewise, expression was enhanced in a ∆fimW mutant. 337
Note that the microplate experiments tell us only about the average response of the 338
population and nothing about how individual cells are behaving. To test whether the 339
cells were responding homogenously, we also performed single-cell measurements of 340
PfimA promoter activity at select times in wild type and a ∆fimW mutant using flow 341
cytometry (Figure 4b). Our results indicate that individual wild-type and ∆fimW mutant 342
cells are responding homogenously with respect to PfimA promoter activity at all times 343
tested. In other words, we did not observe any phase variation or heterogeneity with 344
regards to PfimA promoter activity in our kinetic experiments. 345
346
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
18
Discussion 347
In this work, we investigated the regulatory gene circuit controlling the expression of 348
type I fimbriae in S. typhimurium. Using genetic approaches, we demonstrated that 349
FimY and FimZ independently activate the PfimA promoter. Of the two, FimZ was found 350
to be the dominant activator. We also found that FimY and FimZ strongly activate each 351
other’s expression and weakly activate their own expression. In addition to these two 352
positive regulators, a third regulator, FimW, is known to repress fim gene expression. 353
We found that FimW negatively regulates fim gene expression by repressing expression 354
from the PfimY promoter. Furthermore, FimW participates in a negative feedback loop as 355
FimY was found to enhance PfimW expression. Interestingly, these results suggest that 356
FimY is both an activator and repressor of fim gene expression, as it can directly 357
activate the PfimZ, PfimY, and PfimA promoters and indirectly repress them by enhancing 358
FimW expression. In addition to these regulators, type I fimbriation is also dependent on 359
expression of rare arginine codon tRNA, fimU. However, our results showed that the 360
PfimU promoter is not regulated by FimY, FimZ, or FimW. The results suggest that fimU 361
does not play a role in the internal regulation of the circuit. Finally, we demonstrated that 362
the previously observed coordinate regulation of SPI1 gene expression by the fim gene 363
circuit (7) occurs through the activation of hilE expression by FimZ. Based on these 364
results, we are able to propose the following model for the fim gene circuit in S. 365
typhimurium (Figure 5). 366
According to our model, induction of the fim circuit begins with activation of the PfimY 367
and PfimZ promoters, resulting in small amounts of fimY and fimZ being expressed. FimY 368
and FimZ then rapidly accumulate in the cell due to the positive feedback loop formed 369
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
19
by the cross-activation of PfimY and PfimZ promoters by these two proteins. Expression of 370
the type I fimbrial structural genes from the PfimA promoter commences when the 371
concentration of FimY and FimZ accumulates within the cell beyond a critical level. 372
These two regulators can independently activate the PfimA promoter; however, their 373
expression is correlated as each activates the others expression. Moreover, FimY and 374
FimZ protein expression levels are controlled by a negative feedback loop involving 375
FimW. In this loop, FimY activates expression of the PfimW promoter, and FimW 376
represses expression from the PfimY promoter. We hypothesize that this negative 377
feedback loop involving FimW prevents runaway expression of FimY and FimZ arising 378
from their participation in an interacting positive feedback. Specifically, we hypothesize 379
that when FimY and FimZ reach their optimum expression levels, the FimW negative 380
feedback loop is activated and halts expression from the PfimY and PfimZ promoters. 381
While our model for the fim circuit explains internal regulation, it still does not explain 382
how the circuit is activated. In particular, we do not know which factors induce the PfimY 383
and PfimZ promoters. We suspect that these factors activate both promoters as each 384
alone exhibits some activity in a ∆fimYZ mutant (Figure 1). In addition to these factors, 385
another open question concerns whether fimU plays a role in regulating circuit 386
dynamics. While it is tempting to speculate that fimU expression is tuned in response to 387
environmental signals and thus affects circuit dynamics, more likely this gene is 388
constitutively expressed like other tRNAs. 389
Our results also indicate that FimW-mediated inhibition of fim gene expression is 390
through repression of the PfimY promoter. Earlier reports suggested that FimW binds 391
FimZ and somehow inhibits FimZ-dependent activation of fim promoters (46). Moreover, 392
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
20
FimW was not found to bind to the PfimW promoter. Based on these results, FimW would 393
appear to repress the PfimY promoter by preventing FimZ from activating it. However, we 394
found that FimW is able to repress the PfimY promoter in the absence of FimZ. Our 395
results would suggest that FimW directly binds the PfimY promoter and represses 396
transcription, irrespective of FimZ. Consistent with our model, FimW has a C-terminal 397
LuxR-type helix-turn-helix DNA domain (SM00421) (32). However, at this time we have 398
no direct experimental support for such a mechanism. Moreover, an equally likely 399
hypothesis is that repression by FimW is indirect. Further experiments are clearly 400
required to determine the mechanism of FimW-mediated repression and distinguish 401
between these different putative models. 402
A final unanswered question concerns the role of the positive and negative feedback 403
loops in the fim gene circuit. Our initial hypothesis was that these feedback loops would 404
result in bistability. In particular, interacting positive and negative feedback loops are 405
known to be sufficient ingredients for bistability (24). This bistability could potentially 406
explain the phase variation observed in type I fimbriation during growth in inducing 407
conditions (36). To test whether the fim circuit exhibited bistability, we measured PfimA 408
activity at single-cell resolution as a function of time. Contrary to our initial hypothesis, 409
we did not observe a heterogeneous or switch-like response in induction, the tell tale 410
indicator of bistability. Rather, we observed a continuous or rheostatic-like response in 411
both wild type and a ∆fimW mutant (3). One possibility is that there is a lack of 412
correlation between fim gene expression and the production of type I fimbriae in S. 413
typhimurium (11). Another is that the environment may dictate the response 414
characteristics. For example, under conditions different from those used in our study, 415
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
21
Duguid and coworkers observed subpopulations of cells expressing type I fimbriae, 416
indicative of phase variation (18). With these in mind, we hypothesize that the bacteria 417
exhibit type I fimbriae phase variation under specific environmental conditions and that 418
the regulation of this process involves post-transcriptional mechanisms as well. 419
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
22
Acknowledgements 420
We thank James Slauch and Philip Aldridge for strains and technical advice. This work 421
was partially supported by grants from the National Science Foundation and the 422
National Institutes of Health. 423
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
23
References 424
1. Abraham, J. M., C. S. Freitag, J. R. Clements, and B. I. Eisenstein. 1985. An 425
invertible element of DNA controls phase variation of type 1 fimbriae of 426
Escherichia coli. Proc Natl Acad Sci U S A 82:5724-7. 427
2. Bajaj, V., C. Hwang, and C. A. Lee. 1995. hilA is a novel ompR/toxR family 428
member that activates the expression of Salmonella typhimurium invasion genes. 429
Mol Microbiol 18:715-27. 430
3. Batchelor, E., T. J. Silhavy, and M. Goulian. 2004. Continuous control in 431
bacterial regulatory circuits. J Bacteriol 186:7618-7625. 432
4. Baumler, A. J., R. M. Tsolis, and F. Heffron. 1996. Contribution of fimbrial 433
operons to attachment to and invasion of epithelial cell lines by Salmonella 434
typhimurium. Infect Immun 64:1862-5. 435
5. Baumler, A. J., R. M. Tsolis, and F. Heffron. 1997. Fimbrial adhesins of 436
Salmonella typhimurium. Role in bacterial interactions with epithelial cells. Adv 437
Exp Med Biol 412:149-58. 438
6. Baxter, M. A., T. F. Fahlen, R. L. Wilson, and B. D. Jones. 2003. HilE interacts 439
with HilD and negatively regulates hilA transcription and expression of the 440
Salmonella enterica serovar Typhimurium invasive phenotype. Infect Immun 441
71:1295-305. 442
7. Baxter, M. A., and B. D. Jones. 2005. The fimYZ genes regulate Salmonella 443
enterica Serovar Typhimurium invasion in addition to type 1 fimbrial expression 444
and bacterial motility. Infect Immun 73:1377-85. 445
8. Boddicker, J. D., B. M. Knosp, and B. D. Jones. 2003. Transcription of the 446
Salmonella invasion gene activator, hilA, requires HilD activation in the absence 447
of negative regulators. J Bacteriol 185:525-33. 448
9. Buchanan, K., S. Falkow, R. A. Hull, and S. I. Hull. 1985. Frequency among 449
Enterobacteriaceae of the DNA sequences encoding type 1 pili. J Bacteriol 450
162:799-803. 451
10. Cherepanov, P. P., and W. Wackernagel. 1995. Gene disruption in Escherichia 452
coli: TcR and KmR cassettes with the option of Flp-catalyzed excision of the 453
antibiotic-resistance determinant. Gene 158:9-14. 454
11. Clegg, S., L. S. Hancox, and K. S. Yeh. 1996. Salmonella typhimurium fimbrial 455
phase variation and FimA expression. J Bacteriol 178:542-545. 456
12. Collazo, C. M., and J. E. Galan. 1997. The invasion-associated type-III protein 457
secretion system in Salmonella--a review. Gene 192:51-9. 458
13. Cotter, P. A., and J. F. Miller. 1994. BvgAS-mediated signal transduction: 459
analysis of phase-locked regulatory mutants of Bordetella bronchiseptica in a 460
rabbit model. Infect Immun 62:3381-90. 461
14. Darwin, K. H., and V. L. Miller. 1999. Molecular basis of the interaction of 462
Salmonella with the intestinal mucosa. Clin Microbiol Rev 12:405-28. 463
15. Datsenko, K. A., and B. L. Wanner. 2000. One-step inactivation of 464
chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl 465
Acad Sci U S A 97:6640-5. 466
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
24
16. Davis, R. W., Botsein, D., Roth, J.R. 1980. Advanced bacterial genetics: a 467
manual for genetic engineering. Cold Spring Harbor Laboratory Press, Cold 468
Spring Harbor, NY. 469
17. Duguid, J. P., E. S. Anderson, and I. Campbell. 1966. Fimbriae and adhesive 470
properties in Salmonellae. J Pathol Bacteriol 92:107-38. 471
18. Duguid, J. P., E. S. Anderson, and I. Campbell. 1966. Fimbriae and adhesive 472
properties in Salmonellae. J Pathol Bacteriol 92:107-138. 473
19. Duguid, J. P., M. R. Darekar, and D. W. Wheater. 1976. Fimbriae and infectivity 474
in Salmonella typhimurium. J Med Microbiol 9:459-73. 475
20. Eichelberg, K., W. D. Hardt, and J. E. Galan. 1999. Characterization of SprA, 476
an AraC-like transcriptional regulator encoded within the Salmonella typhimurium 477
pathogenicity island 1. Mol Microbiol 33:139-52. 478
21. Ellermeier, C. D., J. R. Ellermeier, and J. M. Slauch. 2005. HilD, HilC and RtsA 479
constitute a feed forward loop that controls expression of the SPI1 type three 480
secretion system regulator hilA in Salmonella enterica serovar Typhimurium. Mol 481
Microbiol 57:691-705. 482
22. Ellermeier, C. D., and J. M. Slauch. 2003. RtsA and RtsB coordinately regulate 483
expression of the invasion and flagellar genes in Salmonella enterica serovar 484
Typhimurium. J Bacteriol 185:5096-108. 485
23. Fahlen, T. F., N. Mathur, and B. D. Jones. 2000. Identification and 486
characterization of mutants with increased expression of hilA, the invasion gene 487
transcriptional activator of Salmonella typhimurium. FEMS Immunol Med 488
Microbiol 28:25-35. 489
24. Ferrell, J. E. 2002. Self-perpetuating states in signal transduction: positive 490
feedback, double-negative feedback and bistability. Curr Opin Cell Biol 14:140-491
148. 492
25. Gally, D. L., J. Leathart, and I. C. Blomfield. 1996. Interaction of FimB and 493
FimE with the fim switch that controls the phase variation of type 1 fimbriae in 494
Escherichia coli K-12. Mol Microbiol 21:725-38. 495
26. Haldimann, A., and B. L. Wanner. 2001. Conditional-replication, integration, 496
excision, and retrieval plasmid-host systems for gene structure-function studies 497
of bacteria. J Bacteriol 183:6384-6393. 498
27. Hancox, L. S., K. S. Yeh, and S. Clegg. 1997. Construction and 499
characterization of type 1 non-fimbriate and non-adhesive mutants of Salmonella 500
typhimurium. FEMS Immunol Med Microbiol 19:289-96. 501
28. Klemm, P. 1984. The fimA gene encoding the type-1 fimbrial subunit of 502
Escherichia coli. Nucleotide sequence and primary structure of the protein. Eur J 503
Biochem 143:395-9. 504
29. Klemm, P. 1986. Two regulatory fim genes, fimB and fimE, control the phase 505
variation of type 1 fimbriae in Escherichia coli. Embo J 5:1389-93. 506
30. Labbe, D., J. Garnon, and P. C. Lau. 1997. Characterization of the genes 507
encoding a receptor-like histidine kinase and a cognate response regulator from 508
a biphenyl/polychlorobiphenyl-degrading bacterium, Rhodococcus sp. strain M5. 509
J Bacteriol 179:2772-6. 510
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
25
31. Lau, P. C., Y. Wang, A. Patel, D. Labbe, H. Bergeron, R. Brousseau, Y. 511
Konishi, and M. Rawlings. 1997. A bacterial basic region leucine zipper 512
histidine kinase regulating toluene degradation. Proc Natl Acad Sci U S A 513
94:1453-8. 514
32. Letunic, I., R. R. Copley, B. Pils, S. Pinkert, J. Schultz, and P. Bork. 2006. 515
SMART 5: domains in the context of genomes and networks. Nucleic Acids Res 516
34:257-260. 517
33. Lucas, R. L., and C. A. Lee. 2001. Roles of hilC and hilD in regulation of hilA 518
expression in Salmonella enterica serovar Typhimurium. J Bacteriol 183:2733-519
45. 520
34. Lutz, R., and H. Bujard. 1997. Independent and tight regulation of transcriptional 521
units in Escherichia coli via the LacR/O, the TetR/O and AraC/I1-I2 regulatory 522
elements. Nucleic Acids Res 25:1203-10. 523
35. Nagai, T., K. Ibata, E. S. Park, M. Kubota, K. Mikoshiba, and A. Miyawaki. 524
2002. A variant of yellow fluorescent protein with fast and efficient maturation for 525
cell-biological applications. Nat Biotechnol 20:87-90. 526
36. Old, D. C., and J. P. Duguid. 1970. Selective outgrowth of fimbriate bacteria in 527
static liquid medium. J Bacteriol 103:447-56. 528
37. Piknova, L., E. Kaclikova, D. Pangallo, B. Polek, and T. Kuchta. 2005. 529
Quantification of Salmonella by 5'-nuclease real-time polymerase chain reaction 530
targeted to fimC gene. Curr Microbiol 50:38-42. 531
38. Purcell, B. K., J. Pruckler, and S. Clegg. 1987. Nucleotide sequences of the 532
genes encoding type 1 fimbrial subunits of Klebsiella pneumoniae and 533
Salmonella typhimurium. J Bacteriol 169:5831-4. 534
39. Rossolini, G. M., P. Muscas, A. Chiesurin, and G. Satta. 1993. Analysis of the 535
Salmonella fim gene cluster: identification of a new gene (fimI) encoding a 536
fimbrin-like protein and located downstream from the fimA gene. FEMS Microbiol 537
Lett 114:259-65. 538
40. Saini, S., J. D. Brown, P. D. Aldridge, and C. V. Rao. 2008. FliZ Is a 539
posttranslational activator of FlhD4C2-dependent flagellar gene expression. J 540
Bacteriol 190:4979-88. 541
41. Schechter, L. M., S. M. Damrauer, and C. A. Lee. 1999. Two AraC/XylS family 542
members can independently counteract the effect of repressing sequences 543
upstream of the hilA promoter. Mol Microbiol 32:629-42. 544
42. Schechter, L. M., and C. A. Lee. 2001. AraC/XylS family members, HilC and 545
HilD, directly bind and derepress the Salmonella typhimurium hilA promoter. Mol 546
Microbiol 40:1289-99. 547
43. Swenson, D. L., K. J. Kim, E. W. Six, and S. Clegg. 1994. The gene fimU 548
affects expression of Salmonella typhimurium type 1 fimbriae and is related to 549
the Escherichia coli tRNA gene argU. Mol Gen Genet 244:216-8. 550
44. Tavendale, A., C. K. Jardine, D. C. Old, and J. P. Duguid. 1983. 551
Haemagglutinins and adhesion of Salmonella typhimurium to HEp2 and HeLa 552
cells. J Med Microbiol 16:371-80. 553
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
26
45. Tinker, J. K., and S. Clegg. 2000. Characterization of FimY as a coactivator of 554
type 1 fimbrial expression in Salmonella enterica serovar Typhimurium. Infect 555
Immun 68:3305-13. 556
46. Tinker, J. K., and S. Clegg. 2001. Control of FimY translation and type 1 fimbrial 557
production by the arginine tRNA encoded by fimU in Salmonella enterica serovar 558
Typhimurium. Mol Microbiol 40:757-68. 559
47. Tinker, J. K., L. S. Hancox, and S. Clegg. 2001. FimW is a negative regulator 560
affecting type 1 fimbrial expression in Salmonella enterica serovar typhimurium. J 561
Bacteriol 183:435-42. 562
48. van der Velden, A. W., A. J. Baumler, R. M. Tsolis, and F. Heffron. 1998. 563
Multiple fimbrial adhesins are required for full virulence of Salmonella 564
typhimurium in mice. Infect Immun 66:2803-8. 565
49. Yeh, K. S., L. S. Hancox, and S. Clegg. 1995. Construction and 566
characterization of a fimZ mutant of Salmonella typhimurium. J Bacteriol 567
177:6861-5. 568
50. Yeh, K. S., J. K. Tinker, and S. Clegg. 2002. FimZ binds the Salmonella 569
typhimurium fimA promoter region and may regulate its own expression with 570
FimY. Microbiol Immunol 46:1-10. 571
572
573
574
575
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
27
576
Table 1: Strains used during this study. 577
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
28
578
Table 2: Plasmids used during this study. 579
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
29
580
Table 3: List of Primers used in the study 581
582
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
30
583
Figure 1. FimY and FimZ are activators and FimW is repressor of fim gene 584
expression. Comparison of PfimA, PfimY, PfimZ, and PfimW promoter activites in wild type 585
(WT) and ∆fimY, ∆fimZ, ∆fimYZ, and ∆fimW mutants. (Data is average of 3 586
experiments. Each experiment was done in triplicate). 587
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
31
588
Figure 2: FimY and FimZ are strong activators of each other’s expression and 589
also weak auto-activators. Comparison of PfimY and PfimZ promoter activities in a S. 590
typhimurium ∆fimYZ mutant (A) and E. coli (B) where FimY and FimZ are independently 591
expressed from an aTc-inducible promoter on a plasmid. Note that tetR is also 592
expressed from this plasmid in order to achieve aTc-inducible expression. (Data is 593
average of 3 experiments. Each experiment was done in triplicate). 594
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
32
595
Figure 3: FimY and FimZ can independently activate expression from the PfimA 596
promoter. Comparison of PfimA promoter activity in a ∆fimYZ mutant where FimY and 597
FimZ are independently expression from an aTc-inducible promoter on a plasmid. (Data 598
is average of 3 experiments. Each experiment was done in triplicate). 599
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
33
600
Figure 4: Dynamics of PfimA promoter activity. (A) Population-average PfimA activity 601
as a function of time in wild type (WT) and ∆fimY, ∆fimZ, ∆fimYZ, and ∆fimW mutants. 602
(B) Histogram of single-cell PfimA promoter activity at select times in wild type and a 603
∆fimW mutant. Single-cell measurements of promoter activity were obtained using flow 604
cytometry. (Figure 4A: Data is average of single experiment with average of 6 605
independent cultures. The experiment was repeated thrice and identical results 606
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
34
observed. Figure 4B: Population distribution data is from a single experiment. The 607
experiment was repeated thrice and identical results were observed (data not shown)). 608
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
35
609
610
Figure 5: Model for the type I fimbriae gene circuit in S. typhimurium. 611
612
613
614
615
616
617
618
on May 16, 2019 by guest
http://jb.asm.org/
Dow
nloaded from














































![Pork Contaminated with Salmonella enterica Serovar …aem.asm.org/content/76/14/4601.full.pdfstudy indicates that in Germany S. enterica serovar 4,[5],12:i: strains isolated from pig,](https://static.documents.pub/doc/80x56/5b30ee7e7f8b9a81728b54ae/pork-contaminated-with-salmonella-enterica-serovar-aemasmorgcontent76144601fullpdfstudy.jpg)



