Page 1
REVIEW ARTICLE
Role of peroxynitrite-modified biomoleculesin the etiopathogenesis of systemic lupus erythematosus
Rizwan Ahmad • Haseeb Ahsan
Received: 24 July 2012 / Accepted: 6 November 2012 / Published online: 23 November 2012
� Springer-Verlag Italia 2012
Abstract Systemic lupus erythematosus (SLE) is a
chronic inflammatory autoimmune disease characterized
by autoantibodies directed against various biomolecules.
The initial immunogens that drive the development of SLE
are unknown, but characteristics of the immune response in
SLE suggest that it is an antigen-driven response, and a
chromatin antigen could be one of the immunogens for the
production of antinuclear antibodies (ANA) in SLE. Other
factors implicated in the pathogenesis of SLE include
nitrogen-free radicals such as nitric oxide and peroxyni-
trite. The free radical-mediated damage to proteins results
in the modification of amino acid residues, cross-linking of
side chains and fragmentation. The tyrosine residues in
proteins are susceptible to attack by various reactive
nitrogen intermediates, including peroxynitrite to form
3-nitrotyrosine (3-NT). The presence of nitrated proteins
in vivo indicates that peptides derived from the proteolytic
degradation of modified proteins could serve as neoanti-
gens. Histones are highly conserved proteins that are rich in
basic amino acids lysine and arginine. Autoantibodies
against histones and anti-DNA antibodies are present in
SLE. The anti-DNA autoantibodies coexist with anti-his-
tone autoantibodies and may react with chromatin-associ-
ated histones and histone complexes. Elevated levels of
reactive nitrogen species (RNS) in SLE patients suggest a
possible role in the pathogenesis of the disease. The
alteration of proteins resulting from photomodification or
peroxynitrite could lead to the development of antibodies.
Therefore, the modified proteins or photoadducts could
have important implications in autoimmunity, and under-
standing the pathophysiology of peroxynitrite-modified
biomolecules could lead to a better understanding of
autoimmune phenomenon in SLE.
Keywords Peroxynitrite � Nitric oxide � Nitrotyrosine �SLE � Histone proteins � Lysine � Arginine � Photoadduct
Abbreviations
SLE Systemic lupus erythematosus
3-NT 3-nitrotyrosine
NO Nitric oxide
O��2 Superoxide anion
RNS Reactive nitrogen species
nDNA Native DNA
RNOS Reactive nitrogen oxide species
Introduction
Systemic lupus erythematosus (SLE) is a chronic multi-
system autoimmune disease with a wide range of clinical
manifestations and pathogenesis. Its pathology is still not
fully understood, and its etiopathogenesis has remained
relatively elusive. Clinical symptoms commonly appear in
different organs such as the skin, kidney, musculoskeletal
and hematologic systems, but can also affect the lungs,
central nervous system and serous membranes [1, 2]. The
disease is responsible for significant morbidity and mor-
tality [3, 4]. Both genetic and environmental factors have
been linked to SLE [5, 6]. Environmental triggers include
R. Ahmad
Department of Biochemistry, Oman
Medical College, Sohar, Oman
H. Ahsan (&)
Department of Biochemistry, Faculty of Dentistry, Jamia Millia
Islamia (A Central University), Okhla, New Delhi 110025, India
e-mail: [email protected]
123
Clin Exp Med (2014) 14:1–11
DOI 10.1007/s10238-012-0222-5
Page 2
smoking [7], sunlight [8], drugs [9] and certain viruses [5].
SLE involves the innate and adaptive immunopathologies
with both endogenous and exogenous triggers contributing
in the initiation and progression of the disease such as viral
infections, ultraviolet (UV) light exposure and certain
drugs [10].
The nuclear antigens are generated and released in vivo
during apoptosis. The defective clearance of apoptotic
cellular debris in SLE patients causes a loss of self-toler-
ance, autoantibody generation and the formation of
immune complexes [11–14]. Several clinical manifesta-
tions of SLE are thought to be the result of autoantibody
and immune complex deposition in tissues leading to a
secondary inflammatory response [15]. The inflammatory
responses may involve the reaction of nitric oxide (NO)
and superoxide anions (O��2 ) leading to the formation of a
reactive nitrogen species such as peroxynitrite.
It has been established that not only oxygen but nitro-
gen-free radicals play an important role in the pathogenesis
of several human diseases. Nitric oxide radicals participate
in some pathological conditions such as arthritis, vasculitis,
asthma, hypertension, etc. It is also an unstable molecule-
like oxygen-free radical but less reactive and can react with
proteins and other biomolecules [16].
Peroxynitrite is a reactive nitrogen species, making it a
potent oxidizing and nitrating species. In vivo, it is pro-
duced by the reaction between NO and O��2 . It is formed in
the macrophages, endothelial cells, platelets, leukocytes,
neurons, etc. by the reaction between NO and O��2 . Tissue
inflammation and chronic infection lead to the overpro-
duction of NO and O��2 , which rapidly combine to yield
peroxynitrite. The studies suggest that most of the cyto-
toxicity attributed to NO is due to peroxynitrite [17].
Evidence suggests a role of reactive nitrogen species in
the initiation and progression of autoimmune phenomenon.
In chronic inflammatory diseases, peroxynitrite generation
by the phagocytic cells may cause damage to DNA and
proteins, generating neoantigenic epitopes that lead to the
production of antibodies cross-reacting with native mac-
romolecular structures. Therefore, the generation of neo-
epitopes on the molecules, may be one of the factors for the
induction of an immune response as seen in autoimmune
diseases such as SLE. Hence, understanding the contribu-
tion of peroxynitrite and other nitrogen-free radicals in the
etiopathogenesis of SLE could lead to better perception of
these systemic autoimmune disorders and the design of
new therapies directed against the disease process.
Since a lot of literature is available on the contribution
of nitric oxide in the pathogenesis of various diseases, this
review article will focus on the effects of peroxynitrite on
biomolecules (particularly amino acids, proteins and
nucleosomes) in autoimmune phenomenon especially SLE.
Peroxynitrite, a reactive nitrogen species
Peroxynitrite is a relatively long-lived oxidant that may
serve as an important cytotoxic agent. Its biological effects
are due to its reactivity toward a large number of molecules
including lipids, amino acids and nucleic acids. It is
involved in tissue damage in a number of pathophysio-
logical conditions such as neurodegenerative diseases,
cardiovascular disorders, etc. [18–20]. Peroxynitrite inter-
acts with lipids, DNA and proteins causing oxidative
damage and other free radical-induced chain reactions.
These reactions trigger cellular responses such as cellular
signaling, oxidative injury, committing cells to necrosis or
apoptosis. In vivo, peroxynitrite generation represents a
crucial pathological mechanism in conditions such as
stroke, myocardial infarction, chronic heart failure, diabe-
tes, inflammation, neurodegenerative disorders and cancer.
The cell signaling cascade that depends on reversible
phosphorylation of tyrosine is disrupted by tyrosine nitra-
tion. Thus, the occurrence of nitrotyrosine containing
proteins in vivo may be regarded as a general indicator of
tissue damage induced by peroxynitrite [21, 22]. The
nitrosative stress under certain in vivo conditions can result
in the formation of carcinogenic nitrosoamines [23–26].
Both mammalian and bacterial cells when exposed to NO
lead to deamination of guanine, cytosine and adenine via
nitrosative reactions [27].
Peroxynitrite exhibits unique chemical reactivities such
as protein nitration, DNA strand breakage, base modifica-
tion, etc., which may have cytotoxic effects and also lead to
mutagenesis. It is thought to be involved in both cell death
and an increased cancer risk [28–30]. The reaction of
peroxynitrite with lipids leads to peroxidation (malondial-
dehyde and conjugated diene formation) and formation of
nitrito-, nitro-, nitrosoperoxo- and nitrated lipid oxidation
derivatives [31–33]. Peroxynitrite is a particularly effective
oxidant of aromatic molecules, thioethers and organosulfur
compounds that include free amino acids and polypeptide
residues.
Peroxynitrite can influence protein and enzyme function
by nitration of tyrosine residues in tissues leading to
pathological malfunction [34]. The formation of 3-ni-
trotyrosine (3-NT) is contributed by many reactive nitrogen
species such as peroxynitrite, nitrogen dioxide, nitrous
acid, nitronium ion. NO alone is not capable of nitrating
tyrosine [35]. In addition, another way of tyrosine nitration
involves myeloperoxidase, secreted by monocytes and
polymorphonuclear neutrophils under inflammatory con-
ditions [36]. The nitrated product, 3-NT, is found in many
pathophysiological conditions such as chronic inflamma-
tion, atherosclerosis, acute lung injury, etc. [37, 38].
The formation of nitrotyrosine represents a specific
2 Clin Exp Med (2014) 14:1–11
123
Page 3
peroxynitrite-mediated protein modification; thus, detec-
tion of nitrotyrosine in proteins is considered as a bio-
marker for endogenous peroxynitrite activity [31, 39–41].
Even though nucleic acid antigens are by themselves
poorly immunogenic, their antigenicity can be enhanced by
modification through different free radicals [20]. DNA
damage also occurs by RNOS formed under oxidative
stress. This damage is primarily caused due to formation of
peroxynitrite [42].
Autoimmunity
It has been postulated that the immune response against
host antigens could be due to genetic predisposition,
exaggerated B cell activity or cross-reactivity between
foreign and host antigens. The foreign antigens arise as a
consequence of infection, inflammation, drugs, environ-
mental factors, free radicals, etc. [43, 44].
The mechanism of autoantibody production in diseases
such as SLE has not yet been clearly identified. If antigen
selection is an important aspect of differentiation, the
nature of the stimulating antigen also remains to be
determined. The origin of antibodies remains an enigma,
although modified DNA appears to be a causative factor in
RA and SLE. It is possible that the production of autoan-
tibodies may be the result of free radical attack on chro-
matin or histone proteins causing changes at the
macromolecular level. It is, therefore, postulated that in
chronic inflammatory diseases, free radicals generated by
phagocytic cells may cause damage to DNA and proteins
and antibodies to self-antigen are produced. Also, a defect
in the control of apoptosis and delayed clearance of
apoptotic debris provides sustained interaction between
free radicals and macromolecules, generating neoepitopes
which subsequently result in autoimmunity and generating
poly-specific autoantibodies [45].
The origin of autoantibody remains ambiguous, and the
production of anti-DNA antibodies is even more intricate.
Even though nucleic acid antigens are themselves poorly
immunogenic, their antigenicity can be enhanced by
modification with agents such as free radicals. The auto-
antibodies produced against modified biomolecules are the
characteristic feature of systemic human disease, SLE. B
cell hyperactivity and the production of pathogenic auto-
antibodies are the main immunological event in the path-
ogenesis of this disease. The amino acid and nucleotide
sequence of autoantibodies derived from human lupus
subjects with active disease show features of diversification
with a high rate of replacement or silent mutations and the
clustering of mutations in the hypervariable region. This
distinctive feature implies that a pure polyclonal activation
cannot be the only mechanism responsible for autoantibody
production. An antigen-driven process is more likely to
play a role in their generation. It has been suggested that
the antibodies may be stimulated by nucleic acid antigens
or pathogens. B cells whose paratopes have complementary
determining regions (CDR) which are formed by amino
acids that can promote DNA binding may be selectively
stimulated by nucleic acid-related structures [16, 17].
Systemic lupus erythematosus
Two diseases that are considered as a prototype for sys-
temic autoimmunity are SLE and rheumatoid arthritis
(RA). SLE is a multi-systemic disorder characterized by a
variety of autoantibodies and abnormal lymphocyte func-
tion that are responsible for many of the clinical manifes-
tations important in diagnosis. A hallmark of SLE is the
presence of antinuclear antibodies (ANA). The ANA are
prototype autoantibodies that distinctly mark the course of
rheumatic diseases [46]. Because of the close association
between ANA and clinical diagnosis, these antibodies have
become a key component in the evaluation of patients.
These antibodies target a diverse range of macromolecules
including DNA, RNA, proteins and protein–nucleic acid
(PNA) complexes.
SLE is a chronic multisystem and multifactorial
inflammatory autoimmune disease, characterized by a
range of autoantibodies directed against the native nucle-
osome; its DNA and/or its histone components. Nuclear
antigens are generated and released in vivo during pro-
grammed cell death or apoptosis. A hallmark of apoptosis
is the cleavage of chromatin by caspase-activated deoxy-
ribonuclease (DNase) (CAD) [47]. CAD is an endonucle-
ase that is activated by active caspase 3 during apoptosis
and is responsible for degradation of chromatin into
nucleosomal units. The presence of apoptotic bodies is
significant for the generation of autoantigen in autoimmune
diseases, such as SLE, which is characterized by the
presence of antinuclear antibodies [48]. The initial immu-
nogen(s) that drive the development of SLE are unknown,
but characteristics of the immune response in SLE suggest
that it is an antigen-driven response, and a modified
chromatin antigen(s) could be one of the immunogens for
the production of antinuclear antibodies of various types in
SLE [49].
Other factors implicated in the development or pro-
gression of SLE include altered cytokine levels, altered sex
hormone metabolism, increased apoptosis and oxidative
stress. In oxidative stress conditions, there is an evidence of
increased levels of phospholipid peroxidation products and
plasma DNA oxidative products, 8-oxo-20-deoxyguanosine
[50].
Antibodies to DNA have been particularly associated
with SLE which is considered to be a prototype
Clin Exp Med (2014) 14:1–11 3
123
Page 4
autoimmune disease. Native DNA is no longer regarded as
the antigen initiating the disease due to the fact that
immunization of experimental animals with nDNA does
not produce SLE-like symptoms. A few of the possible
candidates could be polynucleotides, denatured DNA,
RNA or modified DNA. While antibodies to single-stran-
ded DNA are formed in several inflammatory complexes
including RA; antibodies to double-stranded DNA serve as
an immunological marker in the diagnosis of SLE [51].
Serum obtained from SLE individuals has been shown to
possess anti-DNA antibodies of diverse antigenic speci-
ficity. These anti-DNA autoantibodies have been used to
evaluate therapeutic effect and clinical features of SLE
patients [52, 53].
Among protein modifications, protein tyrosine nitration
is widely recognized as a hallmark of inflammation that is
associated with the upregulation of inducible nitric oxide
synthase [54]. Both free and protein-associated 3-NT are
commonly found in mammalian tissues and are dramati-
cally increased in pathological conditions previously
associated with the production of reactive oxygen and
nitrogen species [55]. Elevated levels of RNS in SLE
patients suggest a possible role in the pathogenesis of the
disease. Murine models of SLE demonstrate abnormally
high levels of RNS compared with normal mice, whereas
systemic blockade of RNS production reduces disease
activity [56]. A correlation between serum nitrate/nitrite
(NO3/NO2) levels and serum nitrotyrosine levels with
lupus disease activity has been observed. Nitration of
tyrosine is irreversible and can be detected as long as the
nitrated protein is in circulation [57]. Subjects with active
lupus nephritis have higher levels of serum nitrotyrosine
than those without renal disease, suggesting that overpro-
duction of NO and its derived species may have a patho-
logical role in SLE and lupus nephritis [57, 58].
Elevated levels of NO in SLE patients suggest a role of
NO in the pathogenesis of the disease. Murine models of SLE
demonstrate abnormally high levels of NO compared with
normal mice, whereas systemic blockade of NO production
reduces disease activity. Elevated serum nitrate levels cor-
relate with disease activity and along with serum titers of
anti-(dsDNA) antibodies serve as an indicators of SLE [59,
60]. Auto-antibody production in SLE has been attributed to
either selective stimulation of autoreactive B cells by self-
antigens or antigens cross-reactive with self. The persistence
of anti-DNA antibodies in SLE patients, despite systems to
suppress self-recognition, suggests that the response is dri-
ven by an antigen-resembling nDNA. The DNA damage by
peroxynitrite is far more lethal than that caused by NO alone,
leading to the perturbations in nDNA that renders it immu-
nogenic. The modified DNA might, therefore, play a role in
the induction of circulating anti-DNA autoantibodies in
various autoimmune disorders including SLE [61, 62].
Anti-nDNA autoantibodies are considered a diagnostic
marker of SLE. They are expressed in association with
multiple ANA specificities, suggesting a role for both
generalized and antigen-specific immune abnormalities in
their etiology [63, 64]. Histones are small cationic proteins
that bind DNA and remain confined to the nucleus. They
are weak immunogens, probably because of their con-
served nature. However, after apoptosis, they may appear
in the circulation as nucleosomes. Histones show strong
immunogenicity after acetylation and nitration [65]. The
occurrence of autoantibodies against histone proteins has
been reported in sera of SLE patients [66]. In addition, anti-
nDNA autoantibodies are generally present with anti-his-
tone autoantibodies and may react with each of the 5
chromatin-associated histones, as well as with the H3–H4
complex [66]. It has been suggested that alteration in the
amino acid structure or sequence may generate neoepitopes
on self-proteins, leading to an autoaggressive immune
attack. Furthermore, autologous proteins may also become
immunogenic if they are structurally modified. The modi-
fications may generate or mask antigenic epitopes and
stimulate relevant B cells and/or T cells, leading to a
breakdown or bypass of tolerance.
Role of peroxynitrite in autoimmunity
A number of studies support the role of free radicals in the
initiation and progression of autoimmune response. There-
fore, in chronic inflammatory diseases, peroxynitrite gener-
ated by phagocytic cells may cause damage to DNA and
proteins, generating neoantigens that lead to the production
of antibodies cross-reactive with nDNA or proteins. Modi-
fication of native DNA or proteins by peroxynitrite might
also lead to the generation of neoepitopes on the molecule
and may be one of the factors for the induction of the immune
responses as seen in the autoimmune disease-like SLE [66,
67]. The peroxynitrite-modified human DNA was found to
be highly immunogenic in rabbits inducing high titer
immunogen-specific antibodies. The data demonstrate that
the antibodies, though cross-reactive with various nucleic
acids and polynucleotides, preferentially bind peroxynitrite-
modified epitopes on DNA [67, 68].
Immunopathology of peroxynitrite: effect on DNA
DNA is a non-immunogenic entity, but any significant
unrepaired alteration in its basic structure could turn it
‘‘foreign,’’ leading to the activation of immune pathways.
A change in the structure of DNA could either be due to
radiation or interaction with different free radicals. NO and
its derivatives are among the radicals known to interact
with DNA and are primarily involved in deamination of
4 Clin Exp Med (2014) 14:1–11
123
Page 5
DNA bases. Peroxynitrite, on the other hand, leads to more
extensive damage than that caused by an equivalent dose of
NO. The formation of peroxynitrite occurs both intracel-
lularly inside macrophages and extracellularly causing
DNA strand breaks and modification of guanine [69]. The
two main products identified from the reaction of deoxy-
guanosine with peroxynitrite are 8-oxodeoxyguanine and
8-nitroguanine. The former is regarded as a reliable bio-
marker for monitoring DNA damage in with various oxi-
dizing agents. The peroxynitrite-modified DNA has been
shown to acquire immunogenicity and was suspected to be
one of the causes for generation of autoantibodies in cancer
and autoimmune disorders [61, 70].
The peroxynitrite-modified DNA is a potent immunizing
stimulus, inducing high-titer immunogen-specific antibod-
ies in rabbits. Peroxynitrite modification may generate
potential neoepitopes against which antibodies are raised.
The analysis of cross-reactivity indicates that anti-perox-
ynitrite-DNA IgG is immunogen-specific, showing differ-
ent amount of cross-reactivity attributable to sharing of
common antigenic determinants. The common antigenic
determinants between peroxynitrite-DNA and nDNA could
possibly be the sugar-phosphate backbone, since perox-
ynitrite attacks DNA and causes single-strand breaks
through sugar fragmentation. Induced antibodies also rec-
ognized synthetic polynucleotides, representing A/B con-
formations, with a preference for the B-form [61].
The studies in our laboratory have investigated the
photochemical addition of lysine and arginine to native
DNA in view of its potential importance in the photo-cross-
linking of histones to DNA in chromatin [62, 71]. The C-2
carbon atom of thymine in DNA undergoes a covalent
photoaddition reaction with the e-amino group of lysine on
UV irradiation to form a DNA-lysine photoconjugate or
photoadduct [43]. The UV spectroscopic analysis of the
DNA-lysine photoadduct showed hyperchromism, indicat-
ing either the formation of single-stranded breaks in DNA
or ‘‘breathing’’ of a double-stranded polymer at the site of
lysine conjugation. Peroxynitrite caused substantial dam-
age to the DNA-lysine adduct as evident from the hyper-
chromicity of the spectral curve, which could be attributed
to the generation of strand breaks (Fig. 1a). On peroxyni-
trite modification, the hypochromicity increased, which
may be due to the shielding effect of lysine, limiting the
sites for peroxynitrite action. Hypochromicity may also be
attributed to the extensive cross-linking between perox-
ynitrite and the DNA-lysine adduct [62]. The UV spectral
analysis of the DNA-arginine photoadduct showed changes
in DNA spectra as a result of photomodification. A
hyperchromic effect was observed due to increase in
absorbance for the DNA-arginine photoadduct when
compared with that of native DNA and the unirradi-
ated DNA-arginine complex as control (Fig. 1b).
Hyperchromicity represents the formation of single-stran-
ded regions due to adduct formation. The UV absorbance
ratios (A260/A280) of DNA-arginine adduct decreased from
that of native DNA [71]. The results of these studies have
also been summarized in Table 1 [62, 71].
Immunopathology of peroxynitrite: effect on amino
acids and proteins
The free radical-mediated damage to proteins results in the
modification of amino acid residues, cross-linking of side
chains and fragmentation. The aromatic amino acids
(tyrosine, tryptophan and phenylalanine) and cysteine are
more susceptible to modification affecting the general
properties of protein, such as change in ionic charge,
hydrophobicity and conformation [72–74]. L-tyrosine and
1 – DNA-arginine adduct 2 –native DNA3 –peroxynitrite-modified adduct4 –arginine
1
2
3
4
Wavelength (nm)
Ab
sorb
ance
0.8
0.6
0.4
0.2
0200 250 300 350 400
A
B
Fig. 1 a Effect of peroxynitrite on human DNA-lysine photoadduct.
UV absorption spectra of lysine (times symbol), native DNA
(triangle), DNA-lysine adduct (square) and peroxynitrite-modified
adduct (diamond) (Ahmad et al. [62]). b Effect of peroxynitrite on
human DNA–arginine photoadduct. UV absorption spectra of argi-
nine (times symbol), native DNA (diamond), DNA-arginine adduct
(square) and peroxynitrite-modified DNA-arginine adduct (triangle)
(Ahmad et al. [71])
Clin Exp Med (2014) 14:1–11 5
123
Page 6
protein-bound tyrosine are vulnerable to attack by various
reactive nitrogen intermediates, including peroxynitrite to
form 3-NT and protein-associated 3-NT [75]. Tyrosine
nitration is a post-translational modification in proteins that
occurs under conditions of oxidative stress and plays a role
in the pathogenesis of diseases such as asthma [76], Alz-
heimer’s disease [77], atherosclerosis, endotoxic shock,
human acute lung injury [78], SLE [58] and rheumatoid
arthritis [37]. The presence of nitrated proteins in vivo
indicates that nitrated polypeptides derived from the pro-
teolytic degradation of these proteins could serve as anti-
gens. Hence, antibodies that recognize 3-NT have been
successfully raised in rabbits and mice [79, 80].
Alterations in the amino acid sequence or structure can
generate neoantigenic epitopes from self-proteins, causing
aggressive autoimmune attack. There is a range of possible
post-translational modifications of proteins (transglutami-
nation, glycosylation, oxidation, nitration and proteolytic
cleavage) that allow recognition of these self neoepitopes
activating B cells and/or T cells, thus leading to the
breakdown or evasion of immune tolerance [81, 82]. These
alterations can lead to acute and chronic disease states and
may be predictors of disease risk.
The reaction of various amino acids with peroxynitrite
leads to the following products: (a) cysteine and glutathione
are converted to disulfides; (b) methionine is converted to
sulfoxide or is fragmented to ethylene and dimethyl disulfides.
Dimethyl sulfoxide is oxidized to formaldehyde; and
(c) tyrosine and tryptophan undergo one electron oxidation to
radical cations, which are hydroxylated, nitrated and dimer-
ized [83–85]. Exposure of amino acids, peptides and proteins
to ionizing radiation such as gamma radiation and peroxyni-
trite in the presence of O2 gives rise to hydroperoxides. These
hydroperoxides decompose to oxygen- and carbon-centered
radicals on exposure to copper (Cu?) and other transition
metal ions. Hydroperoxide formation on nuclear proteins
results in oxidative damage to associated DNA. These
hydroperoxide-derived radicals react readily with nucleosides
and pyrimidine bases in DNA to form adduct, for example,
8-oxo-dG. This adduct is highly mutagenic and induces G:C to
T:A transversions in human DNA after replication [86].
Both chromatin and histone proteins are components of
the nucleosome [87]. Each nucleosome is composed of 2
molecules each of H3 and H4 histones, flanked by a dimer
of histones-H2A and H2B and surrounded by about
140–200 bp of DNA. Histone H1 is located on the outside
of the nucleosome core which helps in keeping the struc-
ture locked or ‘‘stapled’’. Antinuclear antibodies in human
and murine lupus can distinguish complex epitopes that
result from the ordered interactions between histones and
DNA [88, 89]. Monoclonal antibodies to H2A-H2B-DNA
or other more complex nucleosome epitopes have been
isolated from spontaneously autoimmune mice [90, 91].
Histones are small highly conserved cationic proteins
which bind DNA and are major components of chromatin
and remain confined to nucleus. They are poorly immu-
nogenic because of their conserved nature. However, after
apoptosis, they may appear in circulation as nucleosomes.
The incidence of autoantibodies against various histones
(H1, H2A, H2B, H3 and H4) has been reported to be
between 30 and 60 % in the sera of SLE patients [66].
Histones also act as autoantigens in humoral factors-med-
iated neoplastic diseases [92]. Furthermore, anti-histone
antibodies have also been reported in conditions such as
polymyositis/dermatomyositis [93].
As peroxynitrite reaction involves free radical interme-
diates, it may favor cross-linking and aggregation during
nitration. Formation of tyrosyl radical by peroxynitrite and
its reaction with another tyrosyl radical may generate O,O0-dityrosine covalent cross-links. Peroxynitrite induces an
array of modifications in H2A structure, for example, tyro-
sine nitration, protein carbonyl, dityrosine and crosslinking.
Such gross structural changes might favor polymerization of
native epitopes of H2A histone into potent immunogenic
neoepitopes. The histone proteins are conserved proteins and
act as weak immunogens. However, they show enhanced
immunogenicity after acetylation and alterations in amino
acid structure or sequence and can generate neoantigens on
self-proteins causing and immune attack. The oxidative and
nitrative action of peroxynitrite confers additional immu-
nogenicity on H2A histone indicating a direct correlation
between nitration and immunogenicity. Hence, immuniza-
tion with peroxynityrite-modified H2A may produce poly-
specific antibodies which can recognize and bind to both old
and newly generated epitopes [45].
Contribution of peroxynitrite in SLE
SLE is a complex autoimmune inflammatory disease, and
people are affected by this disorder worldwide. Despite
Table 1 UV absorption and
thermal melting characteristics
of native and modified lysine
and arginine photoadducts
(Ahmad et al. [62, 71])
Parameter/property DNA–lysine
photoadduct
DNA–arginine
photoadduct
Peroxynitrite-modified
DNA-lysine adduct
Peroxynitrite-modified
DNA-arginine adduct
A260/A280 1.2 1.1 1.0 1.1
Tm (oC) 70 61 85 69.5
% Hyperchromicity 52 66 84 69
6 Clin Exp Med (2014) 14:1–11
123
Page 7
decades of intensive research, the origin of SLE has not
been traced. Although the presence of anti-native DNA
autoantibodies characterizes SLE, but native DNA has
been rejected as an antigen-initiating SLE. A previous
report on binding of SLE anti-dsDNA antibodies to a
peptide DNA surrogate tilts the support in favor of protein
antigens as likely initiators of SLE [94]. Cells of the
immunoregulatory network produce both NO and O��2during oxidative burst triggered by inflammation which
may combine to generate peroxynitrite. The peroxynitrite-
driven oxidation and nitration of biomolecules (protein,
lipid, DNA) may lead to SLE-like autoimmunity and age-
related diseases [95, 96].
Furthermore, nitration of tyrosine residues can pro-
foundly alter protein function, suggesting that protein
nitration may be fundamentally related to, and be predic-
tive of, oxidative cell injury. The subsequent release of
altered proteins may enable them to act as antigen-inducing
antibodies against self-proteins. The biological significance
of tyrosine nitration is of interest, because evidence sup-
ports the formation of 3-NT in vivo in diverse pathologic
conditions and 3-NT is thought to be a relatively specific
marker of oxidative damage mediated by peroxynitrite.
Free/protein-bound tyrosine are attacked by various RNS,
including peroxynitrite, to form free/protein-bound 3-NT,
which may provide insight into the mechanism of severe
disease activity seen in lupus patients and among those
developing nephritis [97]. In addition, numerous other
disease states using non-human models have been shown to
involve the formation of 3-NT [55]. At the very least, the
presence of 3-NT in biological samples indicates that
reactive NO-derived species were produced in vivo,
although the exact nature of these species remains to be
determined. The biological markers that have been impli-
cated in the etiopathogenesis of autoimmune phenomenon
and/or produced as a result of reactive nitrogen species
modified-molecules have been identified and include his-
tone proteins, collagen, various types of DNA structures
and modified amino acids. Table 2 lists the various bio-
logical molecules modified by nitric oxide (2a) and per-
oxynitrite (2b) involved in the pathogenesis of autoimmune
conditions such as SLE, RA and OA [38, 56, 58, 98–104].
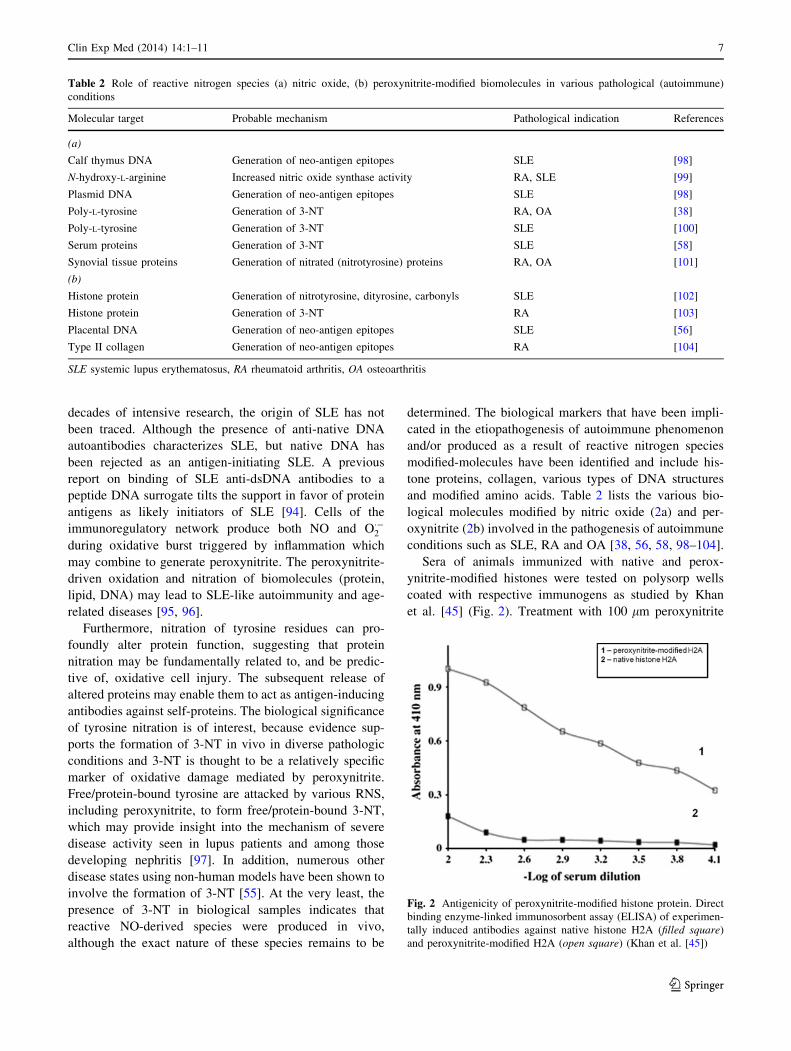
Sera of animals immunized with native and perox-
ynitrite-modified histones were tested on polysorp wells
coated with respective immunogens as studied by Khan
et al. [45] (Fig. 2). Treatment with 100 lm peroxynitrite
Table 2 Role of reactive nitrogen species (a) nitric oxide, (b) peroxynitrite-modified biomolecules in various pathological (autoimmune)
conditions
Molecular target Probable mechanism Pathological indication References
(a)
Calf thymus DNA Generation of neo-antigen epitopes SLE [98]
N-hydroxy-L-arginine Increased nitric oxide synthase activity RA, SLE [99]
Plasmid DNA Generation of neo-antigen epitopes SLE [98]
Poly-L-tyrosine Generation of 3-NT RA, OA [38]
Poly-L-tyrosine Generation of 3-NT SLE [100]
Serum proteins Generation of 3-NT SLE [58]
Synovial tissue proteins Generation of nitrated (nitrotyrosine) proteins RA, OA [101]
(b)
Histone protein Generation of nitrotyrosine, dityrosine, carbonyls SLE [102]
Histone protein Generation of 3-NT RA [103]
Placental DNA Generation of neo-antigen epitopes SLE [56]
Type II collagen Generation of neo-antigen epitopes RA [104]
SLE systemic lupus erythematosus, RA rheumatoid arthritis, OA osteoarthritis
Fig. 2 Antigenicity of peroxynitrite-modified histone protein. Direct
binding enzyme-linked immunosorbent assay (ELISA) of experimen-
tally induced antibodies against native histone H2A (filled square)
and peroxynitrite-modified H2A (open square) (Khan et al. [45])
Clin Exp Med (2014) 14:1–11 7
123
Page 8
conferred a higher immunogenicity on H2A histone than
other lower concentration of peroxynitrite tested [45]. It
is speculated that anti-histone antibodies observed in
autoimmune conditions such as SLE patients might
originate from immunological activity of peroxynitrite-
modified histones. Peroxynitrite-modified H2A histone
could act as an autoantigen leading to generation of anti-
H2A histone antibodies. It is anticipated that anti-histone
antibodies seen in a sub-group of SLE patients might
originate from immunological activity of peroxynitrite-
modified histones due to their protection from digestion
by normal proteolytic machinery [44]. In the context of
anti-histone antibodies in drug-induced lupus erythema-
tosus, it is quite possible that the drug itself might mimic
reactions or pathways leading to abnormal synthesis of
peroxynitrite. The peroxynitrite may then modify the
structure of histone making it immunogenic [17].
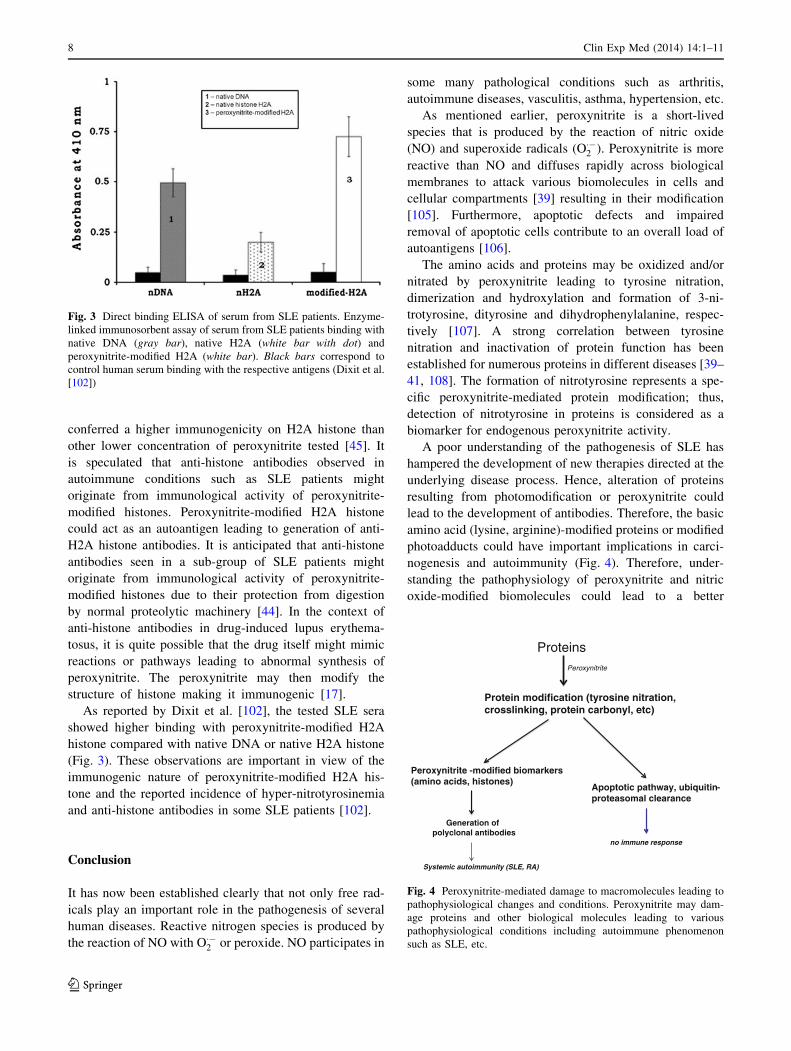
As reported by Dixit et al. [102], the tested SLE sera
showed higher binding with peroxynitrite-modified H2A
histone compared with native DNA or native H2A histone
(Fig. 3). These observations are important in view of the
immunogenic nature of peroxynitrite-modified H2A his-
tone and the reported incidence of hyper-nitrotyrosinemia
and anti-histone antibodies in some SLE patients [102].
Conclusion
It has now been established clearly that not only free rad-
icals play an important role in the pathogenesis of several
human diseases. Reactive nitrogen species is produced by
the reaction of NO with O��2 or peroxide. NO participates in
some many pathological conditions such as arthritis,
autoimmune diseases, vasculitis, asthma, hypertension, etc.
As mentioned earlier, peroxynitrite is a short-lived
species that is produced by the reaction of nitric oxide
(NO) and superoxide radicals (O��2 ). Peroxynitrite is more
reactive than NO and diffuses rapidly across biological
membranes to attack various biomolecules in cells and
cellular compartments [39] resulting in their modification
[105]. Furthermore, apoptotic defects and impaired
removal of apoptotic cells contribute to an overall load of
autoantigens [106].
The amino acids and proteins may be oxidized and/or
nitrated by peroxynitrite leading to tyrosine nitration,
dimerization and hydroxylation and formation of 3-ni-
trotyrosine, dityrosine and dihydrophenylalanine, respec-
tively [107]. A strong correlation between tyrosine
nitration and inactivation of protein function has been
established for numerous proteins in different diseases [39–
41, 108]. The formation of nitrotyrosine represents a spe-
cific peroxynitrite-mediated protein modification; thus,
detection of nitrotyrosine in proteins is considered as a
biomarker for endogenous peroxynitrite activity.
A poor understanding of the pathogenesis of SLE has
hampered the development of new therapies directed at the
underlying disease process. Hence, alteration of proteins
resulting from photomodification or peroxynitrite could
lead to the development of antibodies. Therefore, the basic
amino acid (lysine, arginine)-modified proteins or modified
photoadducts could have important implications in carci-
nogenesis and autoimmunity (Fig. 4). Therefore, under-
standing the pathophysiology of peroxynitrite and nitric
oxide-modified biomolecules could lead to a better
Fig. 3 Direct binding ELISA of serum from SLE patients. Enzyme-
linked immunosorbent assay of serum from SLE patients binding with
native DNA (gray bar), native H2A (white bar with dot) and
peroxynitrite-modified H2A (white bar). Black bars correspond to
control human serum binding with the respective antigens (Dixit et al.
[102])
ProteinsPeroxynitrite
Protein modification (tyrosine nitration, crosslinking, protein carbonyl, etc)
Peroxynitrite -modified biomarkers (amino acids, histones)
Apoptotic pathway, ubiquitin-proteasomal clearance
Generation of polyclonal antibodies
no immune response
Systemic autoimmunity (SLE, RA)
Fig. 4 Peroxynitrite-mediated damage to macromolecules leading to
pathophysiological changes and conditions. Peroxynitrite may dam-
age proteins and other biological molecules leading to various
pathophysiological conditions including autoimmune phenomenon
such as SLE, etc.
8 Clin Exp Med (2014) 14:1–11
123
Page 9
understanding of the autoimmune phenomenon in diseases
such as SLE.
Acknowledgments RA is grateful to the administration of Oman
Medical College, Sohar Campus, Oman for the continued support
during the preparation of this manuscript. We express our sincere
regrets if citations of some authors may have been inadvertently
missed in the review article.
Conflict of interest None.
References
1. Lahita RG (1999) Systemic lupus erythematosus, 3rd edn.
Academic Press, San Diego
2. Cervera R, Khamashta MA, Font J, Sebastiani GD, Gil A, La-
villa P, Mejia JC, Aydintug AO, Chwalinska-Sadowska H, de
Ramon E, Fernandez-Nebro A, Galeazzi M, Valen M, Mathieu
A, Houssiau F, Caro N, Alba P, Ramos-Casals M, Ingelmo M,
Hughes GR (2003) Morbidity and mortality in systemic lupus
erythematosus during a 10-year period: a comparison of early
and late manifestations in a cohort of 1,000 patients. Medicine
82:299–308
3. Pons-Estel GJ, Alarcon GS, Scofield L, Reinlib L, Cooper GS
(2010) Understanding the epidemiology and progression of sys-
temic lupus erythematosus. Semin Arthritis Rheum 39:257–268
4. Trager J, Ward MM (2001) Mortality and causes of death in
systemic lupus erythematosus. Curr Opin Rheumatol 13:345–351
5. Tsokos GC (2011) Systemic lupus erythematosus. N Engl J Med
365:2110–2121
6. Jonsen A, Bengtsson AA, Nived O, Truedsson L, Sturfelt G
(2007) Gene–environment interactions in the aetiology of sys-
temic lupus erythematosus. Autoimmunity 40:613–617
7. Simard JF, Costenbader KH, Liang MH, Karlson EW, Mittle-
man MA (2009) Exposure to maternal smoking and incident
SLE in a prospective cohort study. Lupus 18:431–435
8. Bijl M, Kallenberg CG (2006) Ultraviolet light and cutaneous
lupus. Lupus 15:724–727
9. Ballestar E, Esteller M, Richardson BC (2006) The epigenetic face
of systemic lupus erythematosus. J Immunol 176:7143–7147
10. Lichtman EI, Helfgott SM, Kriegel MA (2012) Emerging ther-
apies for systemic lupus erythematosus–focus on targeting
interferon-alpha. Clin Immunol 143(3):210–221
11. Munoz LE, Gaipl US, Franz S, Sheriff A, Voll RE, Kalden JR,
Herrmann M (2005) SLE-disease of clearance deficiency?
Rheumatology 44:1101–1107
12. Herrmann RE Voll, Zoller OM, Hagenhofer M, Ponner BB,
Kalden JR (1998) Impaired phagocytosis of apoptotic cell
material by monocyte-derived macrophages from patients with
systemic lupus erythematosus. Arthritis Rheum 41:1241–1250
13. Janko C, Schorn C, Grossmayer GE, Frey B, Herrmann M, Gaipl
US, Munoz LE (2008) Inflammatory clearance of apoptotic
remnants in systemic lupus erythematosus (SLE). Autoimmun
Rev 8:9–12
14. Rahman A, Isenberg DA (2008) Systemic lupus erythematosus.
N Engl J Med 358:929–939
15. Koffler D, Schur PH, Kunkel HG (1967) Immunological studies
concerning the nephritis of systemic lupus erythematosus. J Exp
Med 126:607–624
16. Ahsan H, Ali A, Ali R (2003) Oxygen free radicals and systemic
autoimmunity. Clin Exp Immunol 131(3):398–404
17. Ahmad R, Ahsan H (2011) Contribution of peroxynitrite, a
reactive nitrogen species, in the pathogenesis of autoimmunity.
In: Mavragani CP (ed) Autoimmune disorders-pathogenetic
aspects. Intech Open Access Publishers, Croatia, pp 141–156
18. Szabo C (2003) Multiple pathways of peroxynitrite cytotoxicity.
Toxicol Lett 140–141:105–112
19. Pacher P, Beckman JS, Liaudet L (2007) Nitric oxide and per-
oxynitrite in health and disease. Physiol Rev 87(1):315–424
20. Szabo C, Ischiropoulos H, Radi R (2007) Peroxynitrite: bio-
chemistry, pathophysiology and development of therapeutics.
Nat Rev Drug Discov 6(8):662–680
21. Beckman JS, Koppenol WH (1996) Nitric oxide, superoxide and
peroxynitrite: the good the bad and the ugly. Am J Physiol
271:C1424–C1437
22. Sampson JB, Ye Y, Rosen H, Beckman JS (1998) Myeloper-
oxidase and horseradish peroxidase catalyze tyrosine nitration in
proteins from nitric oxide and hydrogen peroxide. Arch Bio-
chem Biophys 356:207–213
23. Marletta MA (1988) Mammalian synthesis of nitrite, nitrate, nitric
oxide and N-nitrosating agents. Chem Res Toxicol 1:249–257
24. Miles AM, Gibson M, Krishna M, Cook JC, Pacelli R, Wink
DA, Grisham MB (1995) Effects of superoxide on nitric oxide
dependent N-nitrosation reactions. Free Radic Res 233:379–390
25. Liu RH, Baldwin B, Tennant BC, Hotchkiss JH (1991) Elevated
formation of nitrate and N-nitrosodimethylamine is wood
chucks (Marmota monax) associated with chronic woodchuck
hepatitis virus infection. Cancer Res 51:3925–3929
26. Liu RH, Jacob JR, Tennant BD, Hotchkiss JH (1992) Nitrite and
nitrosoamine synthesis by hepatocytes isolated from normal
woodchucks, (Marmota Monax) and woodchucks chronically
infected with woodchuck hepatitis virus. Cancer Res
52:3925–3929
27. Merchant K, Chen H, Gonzalez TC, Keefer LK, Shaw BR
(1996) Deamination of single stranded DNA cytosine residues in
aerobic nitric oxide solution at micromolar total NO exposures.
Chem Res Toxicol 9:891–896
28. Ahmad R, Rasheed Z, Ahsan H (2009) Biochemical and cellular
toxicology of peroxynitrite: implications in cell death and
autoimmune phenomenon. Immunopharmacol Immunotoxicol
18(7):388–396
29. Maeda H, Akaike T (1998) Nitric oxide and oxygen radicals in
infection, inflammation, and cancer. Biochemistry (Moscow)
63(7):854–865
30. Ohshima H, Bartsch H (1994) Chronic infections and inflam-
matory processes as cancer risk factors: possible role of nitric
oxide in carcinogenesis. Mutat Res 305(2):253–264
31. Rubbo H (1998) Nitric oxide and peroxynitrite in lipid peroxi-
dation. Medicina B Aires 58(4):361–366
32. Rubbo H, Freeman BA (1996) Nitric oxide regulation of lipid
oxidation reactions: formation and analysis of nitrogen-contain-
ing oxidized lipid derivatives. Methods Enzymol 269:385–394
33. Rubbo H, Radi R, Trujillo M, Telleri R, Kalyanaraman B,
Barnes S, Kirk M, Freeman BA (1994) Nitric oxide regulation of
superoxide and peroxynitrite-dependent lipid peroxidation.
Formation of novel nitrogen-containing oxidized lipid deriva-
tives. J Biol Chem 269(42):26066–26075
34. Pfeiffer S, Mayer B, Hemmens B (1999) Nitric oxide: chemical
puzzles posed by a biological messenger. Angew Chem Int Ed
8:1714–1731
35. Beckman JS (1996) Oxidative damage and tyrosine nitration by
peroxynitrite. Chem Res Toxicol 9:836–844
36. Eiserich JP, Hristova M, Goss CE, Jones AD, Freeman BA,
Halliwell B, Vander Vliet A (1998) Formation of nitric oxide
derived inflammatory oxidants by myeloperoxidase in neutro-
phils. Nature 391:393–397
37. Kaur H, Halliwell B (1994) Evidence for nitric oxide mediated
oxidative damage in chronic inflammation, nitrotyrosine in serum
and synovial fluid from rheumatoid patients. FEBS Lett 350:9–12
Clin Exp Med (2014) 14:1–11 9
123
Page 10
38. Khan F, Siddiqui AA (2006) Prevalence of anti-3-nitrotyrosine
antibodies in the joint synovial fluid of patients with rheumatoid
arthritis, osteoarthritis and systemic lupus erythematosus. Clin
Chim Acta 370:100–107
39. Radi R, Peluffo G, Alvarez MN, Naviliat M, Cayota A (2001)
Unraveling peroxynitrite formation in biological systems. Free
Radic Biol Med 30:463–488
40. Greenacre SAB, Ischiropoulos H (2001) Tyrosine nitration:
localization, quantification, consequences for protein function
and signal transduction. Free Radic Biol Med 34:541–581
41. Turko IV, Murad F (2002) Protein nitration in cardiovascular
diseases. Pharmacol Rev 54:619–634
42. Wink DA, Look JA, Kim S, Vodovotz Y, Pacelli R, Krishna
MC, Russo A, Mitchel JB, Jourdheuil D, Miles AM, Grisham
MB (1997) Superoxide modulates the oxidation and nitrosation
of thiols by nitric oxide derived reactive intermediates. J Biol
Chem 272:11147–11151
43. Islam N, Ali R (1998) Immunological studies on DNA-lysine
photoadduct. Biochem Mol Biol Int 45(3):453–464
44. Ahmad R, Alam K, Ali R (2000) Antigen binding characteristics
of antibodies against hydroxyl radical modified thymidine
monophosphate. Immunol Lett 71:111–115
45. Khan MA, Dixit K, Jabeen S, Moinuddin, Alam K (2009)
Impact of peroxynitrite modification on structure and immuno-
genicity of H2A histone. Scan J Immunol 69:99–109
46. Pisetsky DS (1994) Antinuclear antibodies. Diagn Lab Immunol
14:371–385
47. Gabler C, Kalden JR, Lorenz HM (2003) The putative role of
apoptosis-modified histones for the induction of autoimmunity in
systemic lupus erythematosus. Biochem Pharmacol 66:1441–1446
48. Jog NR, Frisoni L, Shi Q, Monestier M, Hernandez S, Craft J,
Prak ET, Caricchio R (2012) Caspase-activated DNase is
required for maintenance of tolerance to lupus nuclear autoan-
tigens. Arthritis Rheum 64(4):1247–1256
49. Morino N, Sakurai H, Yamada A, Yazaki Y, Minota S (1995)
Rabbit anti-chromatin antibodies recognize similar epitopes on a
histone H1 molecule as lupus autoantibodies. Clin Immunol
Immunopathol 77:52–58
50. Khan F, Khan F, Siddiqui AA, Ali R (2006) Plasmid DNA
acquires immunogenicity on exposure to singlet oxygen. Bio-
chemistry (Moscow) 71:871–878
51. Isenberg D, Shenfeld Y (1997) The origin and significance of
anti-DNA antibodies. Immunol Today 8:279–281
52. Tan EM (1982) Autoantibody to nuclear antigens (ANA): their
immunobiology and medicine. Adv Immunol 33:167–208
53. Pollard KM, Jones JE, Tan EM, Theofilopoulos AN, Dixon FJ,
Rubin RL (1986) Polynucleotide specificity of murine monoclonal
anti-DNA antibodies. Clin Immunol Immunopathol 40:197–208
54. Aulak KS, Miyagi M, Yan L (2001) Proteomic method identifies
proteins nitrated in vivo during inflammatory challenge. Proc
Natl Acad Sci USA 98:12056–12061
55. Crow JP (1999) Measurement and significance of free and
protein-bound 3-nitrotyrosine, 3-chlorotyrosine, and free
3-nitro-4-hydroxyphenylacetic acid in biologic samples: a high-
performance liquid chromatography method using electro-
chemical detection. Methods Enzymol 301:151–160
56. Habib S, Moinuddin, Ali R (2006) Peroxynitrite-modified DNA:
a better antigen for systemic lupus erythematosus anti-DNA
autoantibodies. Biotechnol Appl Biochem 43:65–70
57. Gilkeson G, Cannon C, Oates J, Reilly C, Goldman D, Petri M
(1999) Correlation of serum measures of nitric oxide production
with lupus disease activity. J Rheumatol 26:318–324
58. Oates JC, Christensen EF, Reilly CM, Self SE, Gilkeson GS
(1999) Prospective measure of serum 3-nitrotyrosine levels in
systemic lupus erythematosus: correlation with disease activity.
Proc Assoc Am Physicians 111:611–621
59. Wanchu A, Khullar M, Deodhar SD, Bambery P, Sud A (1998)
Nitric oxide synthesis is increased in patients with systemic
lupus erythmatosus. Rheumatol Int 18(2):41–43
60. Oates JC, Ruiz P, Alexander A, Pippen AM, Gilkeson GS
(1997) Effect of late modulation of nitric oxide production on
murine lupus. Clin Immunol Immunopathol 83(1):86–92
61. Browne SE, Ferrante RJ, Beal MF (1999) Oxidative stress in
Huntington’s disease. Brain Pathol 9(1):147–163
62. Ahmad R, Rasheed Z, Kaushal E, Singh D, Ahsan H (2008)
Biochemical evaluation of human DNA-lysine photoadduct
treated with peroxynitrite. Toxicol Mech Method 18:589–595
63. Pisetsky DS (1992) Anti-DNA antibodies in systemic lupus
erythematosus. Rheum Dis Clin North Am 18:437–454
64. Herrmann M, Winkler TH, Fehr H, Kalden JR (1995) Prefer-
ential recognition of specific DNA motifs by anti-double-
stranded DNA autoantibodies. Eur J Immunol 25:1897–1904
65. Muller S, Isabey A, Cooppez M, Plaue S, Sommermeyer G, Van
Regenmortel MH (1987) Specificity of antibodies raised against
triacetylated histone H4. Mol Immunol 24:779–789
66. Gheidira I, Andolsi H, Mankai A, Fabien N, Jeddi M (2006)
Anti-histone antibodies in systemic lupus erythmatosus, com-
parison of three assays: ELISA, dot blot and immunoblot. Pathol
Biol 54:148–154
67. Ahsan H, Abdi S, Ali A (2002) Recognition of DNA-arginine
photoadduct by anti-DNA auto-antibodies in systemic lupus
erythematosus. Ind J Med Res 115:201–211
68. Dixit K, Moinuddin, Ali A (2005) Immunological studies on
peroxynitrite modified DNA. Life Sci 77(21):2626–2642
69. Burney S, Caulfield JL, Niles JC, Wishnok JS, Tannenbaum SR
(1999) The chemistry of DNA damage from nitric oxide and
peroxynitrite. Mutat Res 424(1–2):37–49
70. Habib S, Moinuddin, Ali R (2005) Acquired antigenicity of
DNA after modification with peroxynitrite. Int J Biol Macromol
35(3–4):221–225
71. Ahmad R, Yadav N, Chaudhary K, Heming T, Ahsan H (2012)
Analysis of human DNA-arginine photoadduct modified with
peroxynitrite. Nucleosides Nucleotides Nucleic Acids 31:
377–387
72. Davies KJ (1987) Protein damage and degradation by oxygen
radicals. I. General aspects. J Biol Chem 262:9895–9901
73. Davies KJ, Delsignore ME, Lin SW (1987) Protein damage and
degradation by oxygen radicals. II. Modification of amino acids.
J Biol Chem 262:9902–9907
74. Davies KJ, Delsignore ME (1987) Protein damage and degra-
dation by oxygen radicals. III. Modification of secondary and
tertiary structure. J Biol Chem 262:9908–9913
75. Tsikas D, Caidahl K (2005) Recent methodological advances in
the mass spectrometric analysis of free and protein-associated
3-nitrotyrosine in human plasma. J Chromatogr B Anal Technol
Biomed Life Sci 814:1–9
76. Xiao GG, Nel AE, Loo JA (2005) Nitrotyrosine-modified pro-
teins and oxidative stress induced by diesel exhaust particles.
Electrophoresis 26:280–292
77. Reynolds MR, Berry RW, Binder LI (2005) Site-specific nitra-
tion and oxidative dityrosine bridging of the tau protein by
peroxynitrite: implications for Alzheimer’s disease. Biochem-
istry 44:1690–1700
78. Brito C, Naviliat M, Tiscornia AC, Vuillier F, Gualco G, Dig-
hiero G, Radi R, Cayota AM (1999) Peroxynitrite inhibits T
lymphocyte activation and proliferation by promoting impair-
ment of tyrosine phosphorylation and peroxynitrite-driven
apoptotic death. J Immunol 162:3356–3366
79. Heijnen HF, van Donselaar E, Slot JW, Fries DM, Blachard-
Fillion B, Hodara R, Lightfoot R, Polydoro M, Spielberg D,
Thomson L, Regan EA, Crapo J, Ischiropoulos H (2006) Sub-
cellular localization of tyrosine-nitrated proteins is dictated by
10 Clin Exp Med (2014) 14:1–11
123
Page 11
reactive oxygen species generating enzymes and by proximity to
nitric oxide synthase. Free Radic Biol Med 40:1903–1913
80. Fries DM, Paxinou E, Themistocleous M, Swanberg E, Grien-
dling KK, Salvemini D, Slot JW, Heijnen HF, Hazen SL, Is-
chiropoulos H (2003) Expression of inducible nitric-oxide
synthase and intracellular protein tyrosine nitration in vascular
smooth muscle cells: role of reactive oxygen species. J Biol
Chem 278:22901–22907
81. Anderton SM (2004) Post-translational modifications of self
antigens: implications for autoimmunity. Curr Opin Immunol
16:753–758
82. Ohmori H, Oka M, Nishikawa Y (2005) Immunogenicity of
autologous IgG bearing the inflammation-associated marker
3-nitrotyrosine. Immunol Lett 96:47–54
83. Ramezanian MS, Padmaja S, Koppenol WH (1996) Nitration
and hydroxylation of phenolic compounds by Peroxynitrite.
Chem Res Toxicol 9(1):232–240
84. Ramezanian MS, Padmaja S, Koppenol WH (1996) Nitration
and hydroxylation of phenolic compounds by peroxynitrite.
Methods Enzymol 269:195–201
85. Alvarez B, Radi R (2003) Peroxynitrite reactivity with amino
acids and proteins. Amino Acids 25(3–4):295–311
86. Luxford C, Morin B, Dean RT, Davies MJ (1999) Histone H1
and other protein- and amino acid-hydroperoxides can give rise
to free radicals which oxidize DNA. Biochem J 344(1):125–134
87. van Holde KE (1989) Chromatin. In: Rich A (ed) Springer series
in molecular biology. Springer, New York
88. Burlingame RW, Rubin RL, Balderas RS, Theofilopoulos AN
(1993) Genesis and evolution of antichromatin autoantibodies in
murine lupus implicates T-dependent immunization with self
antigen. J Clin Invest 91:1687–1696
89. Burlingame RW, Boey ML, Starkebaum G (1994) The central
role of chromatin in autoimmune responses to histones and DNA
in systemic lupus erythematosus. J Clin Invest 94:184–192
90. Losman MJ, Fasy TM, Novick KE, Monestier M (1992)
Monoclonal autoantibodies to subnucleosomes from a MRL/
MP- ?/? mouse. Oligoclonality of the antibody response and
recognition of a determinant composed of histones H2A, H2B
and DNA. J Immunol 48:1561–1569
91. Losman MJ, Fasy TM, Novick KE (1993) Relationships among
antinuclear antibodies from autoimmune MRL mice reacting with
histone H2A–H2B dimers and DNA. Int Immunol 5:513–523
92. Marc M, Thomas M, Lothar B, Frank SL (1991) Anti-histone
antibodies in subacute sensory neuropathy. J Neurooncol 11:71–75
93. Masahidi K, Hironobu I, Norhito Y, Shinichi S, Kanako K,
Kunihiko T (1999) Prevalence and antigen specificity of anti-
histone antibodies in patients with polymyositis/dermatomyosi-
tis. J Invest Dermatol 112:1523–1747
94. Zhang W, Reichlin M (2005) A peptide DNA surrogate that binds
and inhibits anti-dsDNA antibodies. Clin Immunol 117:214–220
95. Drew B, Leeuwenburg C (2002) Aging and the role of reactive
nitrogen species. Ann N Y Acad Sci 959:66–81
96. Oates JC, Gilkeson GS (2006) The biology of nitric oxide and
other reactive intermediates in systemic lupus erythematosus.
Clin Immunol 121:243–250
97. Ohshima H, Friesen M, Brouet I, Bartsch H (1999) Nitrotyrosine
as new marker for endogenous nitrosation and nitration of pro-
teins. Food Chem Toxicol 28:647–652
98. Dixit K, Ali R (2004) Role of nitric oxide modified DNA in the
etiopathogenesis of systemic lupus erythematosus. Lupus 13(2):
95–100
99. Wigand R, Meyer J, Busse R, Hecker M (1997) Increased serum
NG-hydroxy-L-arginine in patients with rheumatoid arthritis and
systemic lupus erythematosus as an index of an increased nitric
oxide synthase activity. Ann Rheum Dis 56:330–332
100. Khan F, Ali R (2006) Antibodies against nitric oxide damaged
poly-L-tyrosine and 3-nitrotyrosine levels in systemic lupus
erythematosus. J Biochem Mol Biol 39(2):189–196
101. Sandhu JK, Robertson S, Birnboim HC, Goldstein R (2003)
Distribution of protein nitrotyrosine in synovial tissues of
patients with rheumatoid arthritis and osteoarthritis. J Rheuma-
tol 30(6):1173–1181
102. Dixit K, Khan MA, Sharma YD, Moinuddin, Alam K (2011)
Peroxynitrite-induced modification of H2A histone presents
epitopes which are strongly bound by human anti-DNA auto-
antibodies: role of peroxynitrite-modified-H2A in SLE induction
and progression. Hum Immunol 72(3):219–225
103. Khan M, Dixit K, Moinuddin, Malik A, Alam K (2012) Role of
peroxynitrite-modified H2A histone in the induction and pro-
gression of rheumatoid arthritis. Scan J Rheumatol 1–8 (in press)
104. Nissim A, Winyard PG, Corrigall V, Fatah R, Perrett D, Pinayi G,
Chernajovsky Y (2005) Generation of neoantigenic epitopes after
posttranslational modification of type II collagen by factors
present within the inflamed joint. Arthritis Rheum 52(12):3829–3838
105. Lowenstein CJ, Dinerman JL, Snyder SH (1994) Nitric oxide: a
physiologic messenger. Ann Intern Med 120:227–237
106. Ballestar E, Esteller M, Richardson BC (2006) The epigenetic face
of systemic lupus erythematosus. J Immunol 176:7143–7147
107. Radi R (2004) Nitric oxide, oxidants, and protein tyrosine
nitration. Proc Natl Acad Sci USA 101:4003–4008
108. Ischiropoulos H (1998) Biological tyrosine nitration: a patho-
physiological function of nitric oxide and reactive oxygen spe-
cies. Arch Biochem Biophys 356:1–11
Clin Exp Med (2014) 14:1–11 11
123