COMMUNICATION Role of the N and C-terminal Strands of Beta 2-Microglobulin in Amyloid Formation at Neutral pH Susan Jones, David P. Smith and Sheena E. Radford* School of Biochemistry and Molecular Biology, University of Leeds, Leeds LS2 9JT, UK Beta 2-microglobulin (b 2 m) is known to form amyloid fibrils de novo in vitro under acidic conditions (below pH 4.8). Fibril formation at neutral pH, however, has only been observed by deletion of the N-terminal six residues; by the addition of pre-assembled seeds; or in the presence of Cu 2þ . Based on these observations, and other structural data, models for fibril formation of b 2 m have been proposed that involve the fraying of the N and C-terminal b-strands and the consequent loss of edge strand protective features. Here, we examine the role of the N and C-terminal strands in the initiation of fibrillogenesis of b 2 m by creating point mutations in strands A and G and comparing the properties of the result- ing proteins with variants containing similar mutations elsewhere in the protein. We show that truncation of buried hydrophobic side-chains in strands A and G promotes rapid fibril formation at neutral pH, even in unseeded reactions, and increases the rate of fibril formation under acidic conditions. By contrast, similar mutations created in the remaining seven b-strands of the native protein have little effect on the rate or pH depen- dence of fibril formation. The data are consistent with the view that per- turbation of the N and C-terminal edge strands is an important feature in the generation of assembly-competent states of b 2 m. q 2003 Elsevier Ltd. All rights reserved Keywords: b2-microglobulin; amyloid fibril; edge strands; cooperativity; aggregation *Corresponding author Amyloid and amyloid-related diseases result from the deposition of normally soluble proteins into insoluble fibrillar plaques. Nature has selected against this process using various approaches, including stabilisation of the native state 1,2 or by the intervention of molecular chaperones that pre- vent partially folded species accumulating in high concentrations. 3,4 Analysis of naturally occurring b-sheet proteins has also highlighted common structural features employed by these proteins to prevent edge strand associations. These “negative design” features include the covering of b-strand edges with loops, the incorporation of short edge strands, b-bulges, prolines or the inclusion of inwardly pointing charged side-chains. 5 Repeating patterns of polar and non-polar amino acids (binary patterning) are also disfavoured in b-sheet edge strands as such sequences promote lateral association. 6–8 All of these arrangements have evolved with the common purpose of preventing edge-to-edge aggregation of b-sheet proteins. The observation that many proteins can assem- ble into a common amyloid-like state, regardless of their initial structure, suggests that amyloid may form by a common mechanism or from pre- cursor states with common structural properties. Indeed, a number of proteins, including some that are not known to be involved in human disease, have been shown to form amyloid-like fibrils in vitro via the population of partially folded states. 9,10 These states are often formed by changes in solu- tion conditions, such as reducing the pH, 11 – 13 increasing the temperature, 14 by adding organic solvents 15 or metal ions. 16 Environmental changes such as these decrease the stability of the native state relative to that of the partially unfolded ensemble, promoting amyloid formation. 4,9 Native b 2 -microglobulin (b 2 m) is a 99-residue protein that forms the non-covalently bound light chain of the class I major histocompatability 0022-2836/$ - see front matter q 2003 Elsevier Ltd. All rights reserved E-mail address of the corresponding author: [email protected]Abbreviations used: b 2 m, human b 2 -microglobulin; ThT, thioflavin-T; DRA, dialysis-related amyloidosis; EM, electron microscopy. doi:10.1016/S0022-2836(03)00688-0 J. Mol. Biol. (2003) 330, 935–941
Transcript
COMMUNICATION
Role of the N and C-terminal Strands of Beta2-Microglobulin in Amyloid Formation at Neutral pH
Susan Jones, David P. Smith and Sheena E. Radford*
School of Biochemistry andMolecular Biology, Universityof Leeds, Leeds LS2 9JT, UK
Beta 2-microglobulin (b2m) is known to form amyloid fibrils de novo invitro under acidic conditions (below pH 4.8). Fibril formation at neutralpH, however, has only been observed by deletion of the N-terminal sixresidues; by the addition of pre-assembled seeds; or in the presence ofCu2þ. Based on these observations, and other structural data, models forfibril formation of b2m have been proposed that involve the fraying ofthe N and C-terminal b-strands and the consequent loss of edge strandprotective features. Here, we examine the role of the N and C-terminalstrands in the initiation of fibrillogenesis of b2m by creating pointmutations in strands A and G and comparing the properties of the result-ing proteins with variants containing similar mutations elsewhere in theprotein. We show that truncation of buried hydrophobic side-chains instrands A and G promotes rapid fibril formation at neutral pH, even inunseeded reactions, and increases the rate of fibril formation under acidicconditions. By contrast, similar mutations created in the remaining sevenb-strands of the native protein have little effect on the rate or pH depen-dence of fibril formation. The data are consistent with the view that per-turbation of the N and C-terminal edge strands is an important feature inthe generation of assembly-competent states of b2m.
Amyloid and amyloid-related diseases resultfrom the deposition of normally soluble proteinsinto insoluble fibrillar plaques. Nature has selectedagainst this process using various approaches,including stabilisation of the native state1,2 or bythe intervention of molecular chaperones that pre-vent partially folded species accumulating in highconcentrations.3,4 Analysis of naturally occurringb-sheet proteins has also highlighted commonstructural features employed by these proteins toprevent edge strand associations. These “negativedesign” features include the covering of b-strandedges with loops, the incorporation of short edgestrands, b-bulges, prolines or the inclusion ofinwardly pointing charged side-chains.5 Repeatingpatterns of polar and non-polar amino acids(binary patterning) are also disfavoured in b-sheet
edge strands as such sequences promote lateralassociation.6 – 8 All of these arrangements haveevolved with the common purpose of preventingedge-to-edge aggregation of b-sheet proteins.
The observation that many proteins can assem-ble into a common amyloid-like state, regardlessof their initial structure, suggests that amyloidmay form by a common mechanism or from pre-cursor states with common structural properties.Indeed, a number of proteins, including some thatare not known to be involved in human disease,have been shown to form amyloid-like fibrils invitro via the population of partially folded states.9,10
These states are often formed by changes in solu-tion conditions, such as reducing the pH,11 – 13
increasing the temperature,14 by adding organicsolvents15 or metal ions.16 Environmental changessuch as these decrease the stability of the nativestate relative to that of the partially unfoldedensemble, promoting amyloid formation.4,9
Native b2-microglobulin (b2m) is a 99-residueprotein that forms the non-covalently bound lightchain of the class I major histocompatability
0022-2836/$ - see front matter q 2003 Elsevier Ltd. All rights reserved
Abbreviations used: b2m, human b2-microglobulin;ThT, thioflavin-T; DRA, dialysis-related amyloidosis;EM, electron microscopy.
doi:10.1016/S0022-2836(03)00688-0 J. Mol. Biol. (2003) 330, 935–941
complex (MHC-I). It has a seven-stranded b-sand-wich fold with the two b-sheets comprisingstrands A, B, D, E and C, F, G, respectively (Figure1). These sheets are held together by a single disul-phide bond between Cys25 and Cys80 in strands Band F.17,18 Full length wild-type b2m is the key pro-tein component of dialysis-related amyloidosis(DRA).19,20 Renal failure disrupts the clearance ofb2m from the serum, resulting in a significantincrease in its concentration (by up to 60-fold).20
As a consequence, b2m self-associates into amyloidfibrils that are deposited in the musculoskeletalsystem.21 Recent studies have shown that partialunfolding of native b2m is a pre-requisite of fibrilformation in vitro12 (Smith et al., accompanyingmanuscript).22 NMR analysis of the partiallyunfolded states of b2m formed under acidic con-ditions in vitro,23,24 together with a number ofother studies,16,18,25,26 have provided importantclues into the structural rearrangements requiredfor the normally highly soluble native protein toform amyloid-like fibrils in vitro. These studieshave shown that partially unfolded b2m formedby acidification to pH 3.6 displays significantdestabilisation of both the N and C-terminalstrands (A and G, respectively), but retains astable, compact core involving strands B, C, D, Eand F.23 This species assembles spontaneously denovo in vitro into short (,600 nm) curved amy-loid-like fibrils.12,27 Further acidification of b2m to
pH 2.5 (at low ionic strength) results in the popu-lation of a more highly denatured state, in whichthe N and C-terminal strands are unstructured24
(V. J. McParland, A. P. Kalverda, S. W. Homans &S.E.R., unpublished results). Incubation of thisspecies results in the assembly of long (,1 mm),straight amyloid-like fibrils de novo in vitro by lagdependent kinetics.27,28 Interestingly, hydrogenexchange experiments on b2m fibrils formed at pH2.5 in vitro have shown that the A and G strandsare also relatively weakly protected from exchangein amyloid fibrils,29 and analysis of ex vivo fibrilsfrom DRA patients has shown that up to 20% ofthe b2m molecules display truncations in the N-terminal strand,20,30 suggesting that these strandsare not involved in the core of the assembled fibril.Together with the observation that b2m can beinduced to form fibrils at neutral pH by theremoval of the N-terminal six amino acids,25 theseresults suggest that strands A and G play animportant role in protecting monomeric b2m fromfibril formation. Here, we test this hypothesis bymutating buried hydrophobic residues in strandsA and G such that hydrophobic packing betweenthese strands and the core of native b2m isreduced. We show that substitution of isoleucineor valine residues in these strands with alanineresults in the rapid formation of amyloid-likefibrils at neutral pH and increases the rate of amy-loid formation at acidic pH. By contrast, similarmutations created elsewhere in the protein haveno such effects. The data are consistent with theview that destabilisation of strands A and/or G isan important step in the initiation of b2m fibrillo-genesis and point the way towards new routes ofpreventing amyloid formation in vitro.
Point mutations in the N and C-terminalstrands destabilise native b2m
To examine the role of global stability in deter-mining the amyloid-forming properties of b2m, 13point mutations were made throughout the nativestructure and the stability of the resulting variantswas correlated with their ability to form amyloid-like fibrils under acidic conditions (accompanyingmanuscript).22 Here, we extend these studies byfocusing on three point mutants that were createdspecifically to examine the role of strands A and Gin the generation of amyloid-like fibrils at neutralpH. Two mutations (I7A and V9A) were intro-duced in strand A and one (V93A) in strand G(Figure 1). These residues are hydrophobic andburied in the core of native b2m.
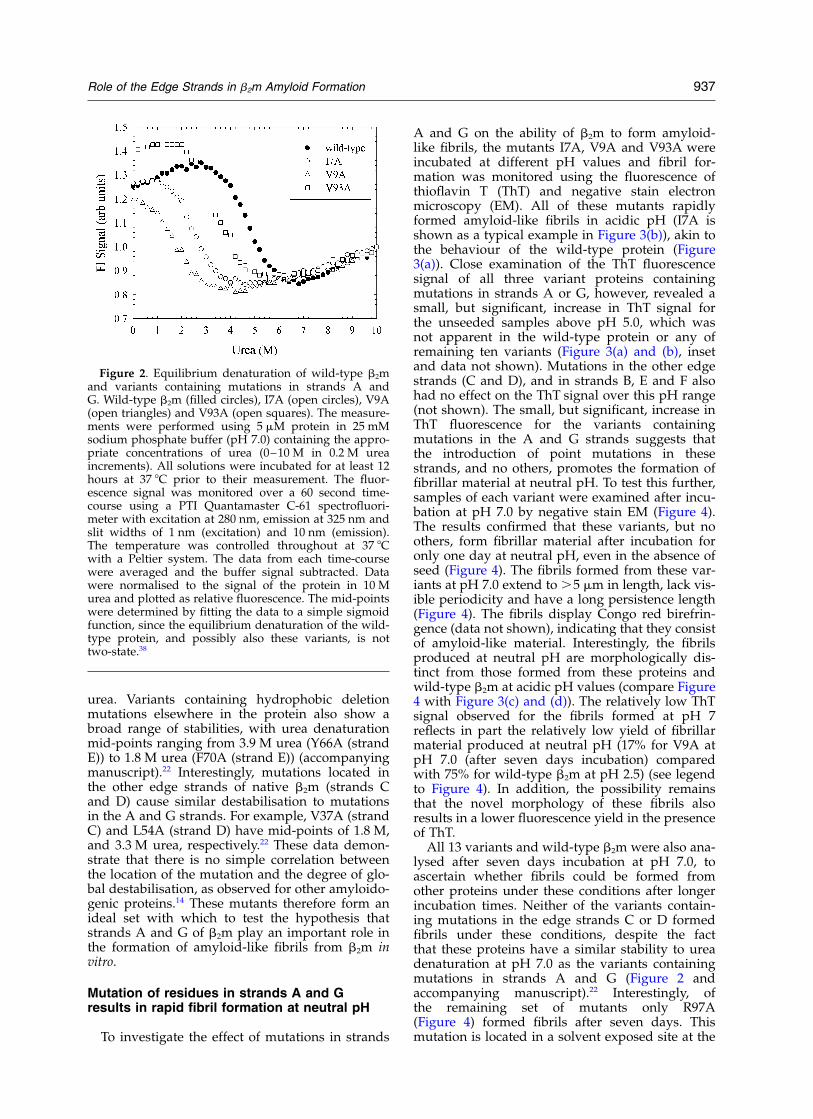
The effect of mutations in strands A and G onthe stability of native b2m at pH 7.0, 37 8C isshown in Figure 2. V9A (strand A) is the mostdestabilised protein, with a mid-point of ureadenaturation at 1.6 M (the midpoint of wild-typeprotein is 4.7 M urea). I7A (strand A) is also signifi-cantly destabilised, with a mid-point of 2.7 M urea,whilst V93A (strand G) is the least destabilised ofthese three variants with a mid-point of 3.4 M
Figure 1. Ribbon diagram of the X-ray structure ofhuman b2m (PDB 1DUZ).17 Strands A and G are shownin red and residues I7, V9 and V93, which were mutatedto alanine and characterised in this paper, are shown inyellow. The disulphide bond is shown in grey. TheFigure was drawn using SPOCK.37 Mutagenesis was car-ried out using the QuikChange site-directed mutagenesiskit (Stratagene) with the plasmid pINKwt
27 as a template.All mutant proteins were expressed and purified as pre-viously described.27 An average yield of 10–20 mg pureb2m/l culture was obtained. The proteins were shownto be .95 % pure by SDS-PAGE and of the expectedmolecular weight by electrospray ionisation massspectrometry.
936 Role of the Edge Strands in b2m Amyloid Formation
urea. Variants containing hydrophobic deletionmutations elsewhere in the protein also show abroad range of stabilities, with urea denaturationmid-points ranging from 3.9 M urea (Y66A (strandE)) to 1.8 M urea (F70A (strand E)) (accompanyingmanuscript).22 Interestingly, mutations located inthe other edge strands of native b2m (strands Cand D) cause similar destabilisation to mutationsin the A and G strands. For example, V37A (strandC) and L54A (strand D) have mid-points of 1.8 M,and 3.3 M urea, respectively.22 These data demon-strate that there is no simple correlation betweenthe location of the mutation and the degree of glo-bal destabilisation, as observed for other amyloido-genic proteins.14 These mutants therefore form anideal set with which to test the hypothesis thatstrands A and G of b2m play an important role inthe formation of amyloid-like fibrils from b2m invitro.
Mutation of residues in strands A and Gresults in rapid fibril formation at neutral pH
To investigate the effect of mutations in strands
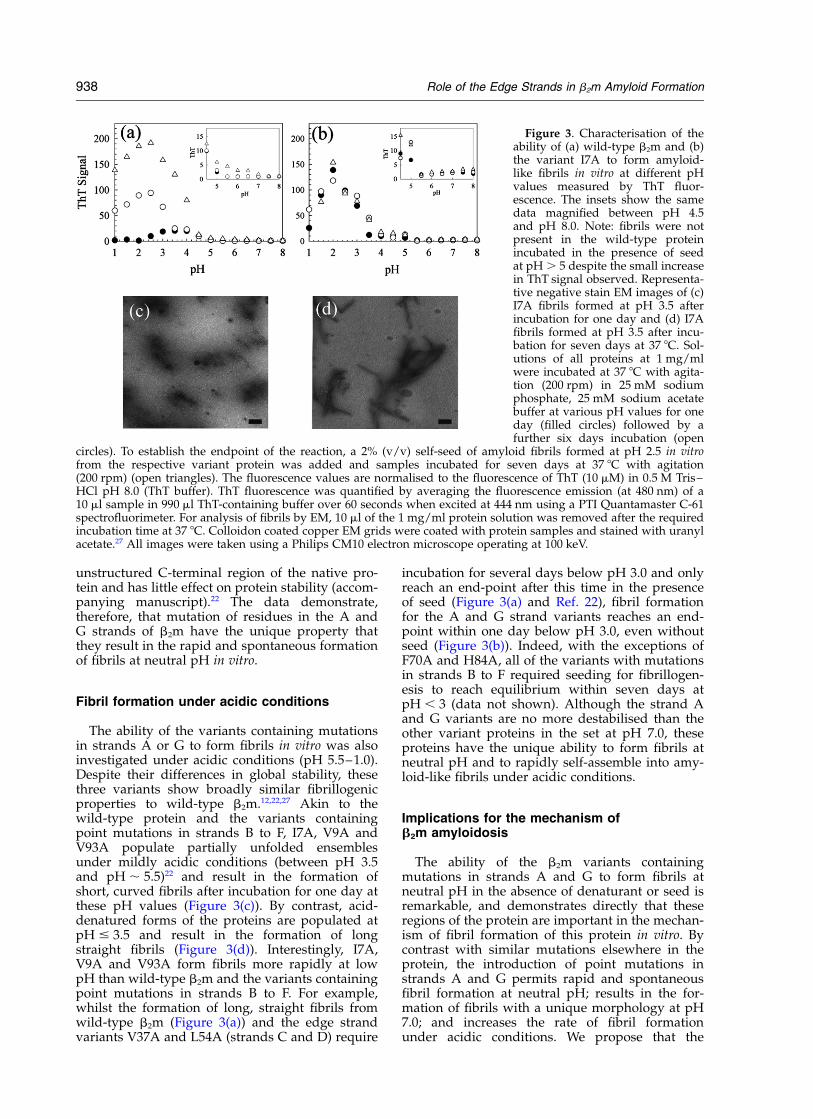
A and G on the ability of b2m to form amyloid-like fibrils, the mutants I7A, V9A and V93A wereincubated at different pH values and fibril for-mation was monitored using the fluorescence ofthioflavin T (ThT) and negative stain electronmicroscopy (EM). All of these mutants rapidlyformed amyloid-like fibrils in acidic pH (I7A isshown as a typical example in Figure 3(b)), akin tothe behaviour of the wild-type protein (Figure3(a)). Close examination of the ThT fluorescencesignal of all three variant proteins containingmutations in strands A or G, however, revealed asmall, but significant, increase in ThT signal forthe unseeded samples above pH 5.0, which wasnot apparent in the wild-type protein or any ofremaining ten variants (Figure 3(a) and (b), insetand data not shown). Mutations in the other edgestrands (C and D), and in strands B, E and F alsohad no effect on the ThT signal over this pH range(not shown). The small, but significant, increase inThT fluorescence for the variants containingmutations in the A and G strands suggests thatthe introduction of point mutations in thesestrands, and no others, promotes the formation offibrillar material at neutral pH. To test this further,samples of each variant were examined after incu-bation at pH 7.0 by negative stain EM (Figure 4).The results confirmed that these variants, but noothers, form fibrillar material after incubation foronly one day at neutral pH, even in the absence ofseed (Figure 4). The fibrils formed from these var-iants at pH 7.0 extend to .5 mm in length, lack vis-ible periodicity and have a long persistence length(Figure 4). The fibrils display Congo red birefrin-gence (data not shown), indicating that they consistof amyloid-like material. Interestingly, the fibrilsproduced at neutral pH are morphologically dis-tinct from those formed from these proteins andwild-type b2m at acidic pH values (compare Figure4 with Figure 3(c) and (d)). The relatively low ThTsignal observed for the fibrils formed at pH 7reflects in part the relatively low yield of fibrillarmaterial produced at neutral pH (17% for V9A atpH 7.0 (after seven days incubation) comparedwith 75% for wild-type b2m at pH 2.5) (see legendto Figure 4). In addition, the possibility remainsthat the novel morphology of these fibrils alsoresults in a lower fluorescence yield in the presenceof ThT.
All 13 variants and wild-type b2m were also ana-lysed after seven days incubation at pH 7.0, toascertain whether fibrils could be formed fromother proteins under these conditions after longerincubation times. Neither of the variants contain-ing mutations in the edge strands C or D formedfibrils under these conditions, despite the factthat these proteins have a similar stability to ureadenaturation at pH 7.0 as the variants containingmutations in strands A and G (Figure 2 andaccompanying manuscript).22 Interestingly, ofthe remaining set of mutants only R97A(Figure 4) formed fibrils after seven days. Thismutation is located in a solvent exposed site at the
Figure 2. Equilibrium denaturation of wild-type b2mand variants containing mutations in strands A andG. Wild-type b2m (filled circles), I7A (open circles), V9A(open triangles) and V93A (open squares). The measure-ments were performed using 5 mM protein in 25 mMsodium phosphate buffer (pH 7.0) containing the appro-priate concentrations of urea (0–10 M in 0.2 M ureaincrements). All solutions were incubated for at least 12hours at 37 8C prior to their measurement. The fluor-escence signal was monitored over a 60 second time-course using a PTI Quantamaster C-61 spectrofluori-meter with excitation at 280 nm, emission at 325 nm andslit widths of 1 nm (excitation) and 10 nm (emission).The temperature was controlled throughout at 37 8Cwith a Peltier system. The data from each time-coursewere averaged and the buffer signal subtracted. Datawere normalised to the signal of the protein in 10 Murea and plotted as relative fluorescence. The mid-pointswere determined by fitting the data to a simple sigmoidfunction, since the equilibrium denaturation of the wild-type protein, and possibly also these variants, is nottwo-state.38
Role of the Edge Strands in b2m Amyloid Formation 937
unstructured C-terminal region of the native pro-tein and has little effect on protein stability (accom-panying manuscript).22 The data demonstrate,therefore, that mutation of residues in the A andG strands of b2m have the unique property thatthey result in the rapid and spontaneous formationof fibrils at neutral pH in vitro.
Fibril formation under acidic conditions
The ability of the variants containing mutationsin strands A or G to form fibrils in vitro was alsoinvestigated under acidic conditions (pH 5.5–1.0).Despite their differences in global stability, thesethree variants show broadly similar fibrillogenicproperties to wild-type b2m.12,22,27 Akin to thewild-type protein and the variants containingpoint mutations in strands B to F, I7A, V9A andV93A populate partially unfolded ensemblesunder mildly acidic conditions (between pH 3.5and pH , 5.5)22 and result in the formation ofshort, curved fibrils after incubation for one day atthese pH values (Figure 3(c)). By contrast, acid-denatured forms of the proteins are populated atpH # 3.5 and result in the formation of longstraight fibrils (Figure 3(d)). Interestingly, I7A,V9A and V93A form fibrils more rapidly at lowpH than wild-type b2m and the variants containingpoint mutations in strands B to F. For example,whilst the formation of long, straight fibrils fromwild-type b2m (Figure 3(a)) and the edge strandvariants V37A and L54A (strands C and D) require
incubation for several days below pH 3.0 and onlyreach an end-point after this time in the presenceof seed (Figure 3(a) and Ref. 22), fibril formationfor the A and G strand variants reaches an end-point within one day below pH 3.0, even withoutseed (Figure 3(b)). Indeed, with the exceptions ofF70A and H84A, all of the variants with mutationsin strands B to F required seeding for fibrillogen-esis to reach equilibrium within seven days atpH , 3 (data not shown). Although the strand Aand G variants are no more destabilised than theother variant proteins in the set at pH 7.0, theseproteins have the unique ability to form fibrils atneutral pH and to rapidly self-assemble into amy-loid-like fibrils under acidic conditions.
Implications for the mechanism ofb2m amyloidosis
The ability of the b2m variants containingmutations in strands A and G to form fibrils atneutral pH in the absence of denaturant or seed isremarkable, and demonstrates directly that theseregions of the protein are important in the mechan-ism of fibril formation of this protein in vitro. Bycontrast with similar mutations elsewhere in theprotein, the introduction of point mutations instrands A and G permits rapid and spontaneousfibril formation at neutral pH; results in the for-mation of fibrils with a unique morphology at pH7.0; and increases the rate of fibril formationunder acidic conditions. We propose that the
Figure 3. Characterisation of theability of (a) wild-type b2m and (b)the variant I7A to form amyloid-like fibrils in vitro at different pHvalues measured by ThT fluor-escence. The insets show the samedata magnified between pH 4.5and pH 8.0. Note: fibrils were notpresent in the wild-type proteinincubated in the presence of seedat pH . 5 despite the small increasein ThT signal observed. Representa-tive negative stain EM images of (c)I7A fibrils formed at pH 3.5 afterincubation for one day and (d) I7Afibrils formed at pH 3.5 after incu-bation for seven days at 37 8C. Sol-utions of all proteins at 1 mg/mlwere incubated at 37 8C with agita-tion (200 rpm) in 25 mM sodiumphosphate, 25 mM sodium acetatebuffer at various pH values for oneday (filled circles) followed by afurther six days incubation (open
circles). To establish the endpoint of the reaction, a 2% (v/v) self-seed of amyloid fibrils formed at pH 2.5 in vitrofrom the respective variant protein was added and samples incubated for seven days at 37 8C with agitation(200 rpm) (open triangles). The fluorescence values are normalised to the fluorescence of ThT (10 mM) in 0.5 M Tris–HCl pH 8.0 (ThT buffer). ThT fluorescence was quantified by averaging the fluorescence emission (at 480 nm) of a10 ml sample in 990 ml ThT-containing buffer over 60 seconds when excited at 444 nm using a PTI Quantamaster C-61spectrofluorimeter. For analysis of fibrils by EM, 10 ml of the 1 mg/ml protein solution was removed after the requiredincubation time at 37 8C. Colloidon coated copper EM grids were coated with protein samples and stained with uranylacetate.27 All images were taken using a Philips CM10 electron microscope operating at 100 keV.
938 Role of the Edge Strands in b2m Amyloid Formation
enhanced amyloidogenicity of these variantsresults from a specific decrease in the local stabilityof the A and G strands, caused by truncation ofhydrophobic residues that dock these strands intothe core of the native protein. Increased localdynamics of these strands and the resultingdecrease in cooperativity of the variant proteinsmay then result in the significant, albeit transient,population of precursor state(s) that expose newassembly-competent surfaces and ablate the needfor acidification or seeding to initiate fibrillogen-esis. These data are consistent with a number ofother studies that have suggested that amyloid for-mation from wild-type b2m is initiated by the fray-ing of the N and/or C-terminal strands23,25 and thepopulation of specific precursor states that are notdirectly related to the global stability of theprotein.22 For example, deletion of the N-terminalsix amino acids (in DN6b2m) results in the popu-lation of a partially unfolded ensemble of non-native b2m molecules that are unstructured inresidues corresponding to the native strand A.25
This species is able to extend pre-formed fibrils atpH 7 and displays increased rates of fibrillogenesisat acidic pH, similar to the results presented herefor I7A, V9A and V93A. DN6b2m contains struc-tural features similar to those observed in the amy-
loidogenic precursors of wild-type b2m formed atpH 3.623 and 2.524 and proposed here for the var-iants I7A, V9A and V93A. Destabilisation ofstrands A and G in molecules that retain significantstable structure in the remainder of the b-sandwichis thus a common feature of the different amyloidprecursor states of b2m identified to date.
Negative design features have evolved in manyb-sheet proteins to prevent inter-molecular associ-ation between edge strands.5 In this regard b2m isno exception: the native protein incorporates aninwardly pointing positively charged residue instrand G (K91) that could protect the A and Gedge strands. A number of protective features arealso found in the C and D edge strands, includinga b-bulge and inwardly pointing His in strand Dand a short two residue b-strand (C0) that hydro-gen bonds to strand C.5,17,18 Transient unfolding ofone or more of these strands would result in theexposure of new edge strands, which may lackprotective features and facilitate assembly. Forexample, transient unfolding of the A and Gstrands would expose the B and F strands tosolvent. These strands are linked by a disulphidebond in native b2m and lack protective features.Interestingly, a peptide encompassing strands Band C has been shown to be capable of forming
Figure 4. Characterisation of amyloid-like fibrils formed from different b2m variants at neutral pH in vitro. Negativestain EM images of fibrils formed from (left to right) I7A, V9A, V93A and R97A. Samples of 1 mg/ml of each of theb2m variants were incubated in 25 mM sodium phosphate, 25 mM sodium acetate buffer at pH 7.0, 37 8C for one daywith agitation at 200 rpm. R97A was incubated for seven days with agitation at 200 rpm. A 10 ml sample was thenremoved and added to a colloidon coated copper EM grid and stained with uranyl acetate.27 All images were takenusing a Philips CM10 electron microscope operating at 100 keV. The yield of fibrils was quantified using a procedurebased on depolymerisation of the fibrils and denaturation of the resulting monomers in guanidinium chloride. Briefly,fibrils were grown for six days (in triplicate) using 1 mg/ml b2m as described above. Each fibril suspension was thencentrifuged at 13,000 rpm for five minutes. The supernatants were removed immediately and transferred to Eppendorftubes. The pellet was resuspended to the original volume in 25 mM sodium phosphate, 25 mM sodium acetate bufferat the appropriate pH. Solutions of freshly dissolved monomeric protein at 1 mg/ml in the same buffer were used ascontrols. Each solution of 100 ml was then added to 900 ml of 6 M guanidinium chloride (pH 7.0) and the sampleswere incubated at 25 8C for two hours. EM and ThT fluorescence showed that this procedure resulted in the completedissociation of fibrillar material. The A280 of the samples was then taken and the amount of protein in the supernatantand fibril pellet was calculated by comparison of their A280 values with that of the monomeric control.
Role of the Edge Strands in b2m Amyloid Formation 939
amyloid-like fibrils in isolation that promotes amy-loid formation from the intact protein.31 Moreover,maintenance of the disulphide bond is requiredfor fibril extension, suggesting that the B and Fstrand pair is important for fibril growth in vitro.32
We have recently shown that the b-bulge in strandD, that is an ideal protective feature,5 is replacedby a single, long b-strand in the crystal structureof the protein at pH 5.7.18 As a consequence, anassembly competent surface is created on thisedge strand of the protein that could promote fibrilformation. Alternatively, more dramatic changes instructure may be necessary for the initiation ofamyloid formation, such that new assembly com-petent surfaces are created, possibly involvingstrand E, which has been shown to be inherentlyfibrillogenic in isolation.33 The regions of b2m thatcould be involved in promoting fibril formationare therefore diverse and reinforce the likely het-erogeneity of the assembly mechanism. Intrigu-ingly, strands A, C, D and G are all involved incontacts with the heavy chain in the intact MHC-Icomplex,17 emphasising the pivotal role of theheavy chain in stabilising b2m and preventing itsself-assembly.
The precise conformational changes required foramyloid formation of b2m remain unknown andmore experiments will be needed to determine themechanisms of unfolding and assembly of thedifferent monomeric precursor states. Possibilitiesfor the assembly mechanism include head-to-tailassociation, stacking of head-to-head dimers,domain swapping,34 – 36 or assembly following lar-ger conformational changes. Which, if any of thesemechanisms occurs in the assembly of amyloidfibrils from b2m under different conditions willrequire further investigation. Nonetheless, thedata presented here suggest that an increase in thelocal dynamics of the N and C-terminal strandsand the consequent decrease in cooperativity ofthe native structure are important features in initi-ating fibrillogenesis of b2m. Decreasing the localdynamics of the edge strands, rather than increas-ing the global stability of the native state, could bea potential mechanism of preventing self-assemblyof this protein. The behaviour of the variant pro-teins studied here predict specific regions of theprotein that could be important targets for thedevelopment of potential therapies to prevent theassociation of b2m monomers into amyloid-likefibrils in vitro and possibly ultimately in vivo.
Acknowledgements
We thank Victoria McParland and all othermembers of the S.E.R. group for helpful dis-cussions and Chi Trinh for help with Figure 1. Wealso thank the BBSRC, the Wellcome Trust and theUniversity of Leeds for financial support. Theauthors are members of the Astbury Centre for
Structural Molecular Biology, which is part ofthe North of England Structural Biology Centre,and is funded by the BBSRC. S.E.R. is a BBSRCProfessorial Fellow.
References
1. Hammarstrom, P., Wiseman, R. L., Powers, E. T. &Kelly, J. W. (2003). Prevention of transthyretin amy-loid disease by changing protein misfolding ener-getics. Science, 299, 713–716.
2. Sacchettini, J. C. & Kelly, J. W. (2002). Therapeuticstrategies for human amyloid diseases. Nature Rev.Drug Discov. 1, 267–275.
3. Sittler, A., Lurz, R., Lueder, G., Priller, J., Lehrach, H.,Hayer-Hartl, M. K., Hartl, F. U. & Wanker, E. E.(2001). Geldanamycin activates a heat shockresponse and inhibits huntingtin aggregation in acell culture model of Huntington’s disease. Hum.Mol. Genet. 10, 1307–1315.
4. Fink, A. L. (1998). Protein aggregation: folding aggre-gates, inclusion bodies and amyloid. Fold. Des. 3,R9–R23.
5. Richardson, J. S. & Richardson, D. C. (2002). Naturalbeta-sheet proteins use negative design to avoidedge-to-edge aggregation. Proc. Natl Acad. Sci. USA,99, 2754–2759.
6. Broome, B. M. & Hecht, M. H. (2000). Nature disfa-vors sequences of alternating polar and non-polaramino acids: implications for amyloidogenesis.J. Mol. Biol. 296, 961–968.
7. Wang, W. & Hecht, M. H. (2002). Rationally designedmutations convert de novo amyloid-like fibrils intosoluble monomeric b-sheet proteins. Proc. Natl Acad.Sci. USA, 99, 2760–2765.
8. Mandel-Gutfreund, Y. & Gregoret, L. M. (2002). Onthe significance of alternating patterns of polar andnon-polar residues in beta-strands. J. Mol. Biol. 323,453–461.
9. Dobson, C. M. (1999). Protein misfolding, evolutionand disease. Trends Biochem. Sci. 24, 329–332.
10. Rochet, J. C. & Lansbury, P. T. (2000). Amyloid fibril-logenesis: themes and variations. Curr. Opin. Struct.Biol. 10, 60–68.
11. Lai, Z. H., Colon, W. & Kelly, J. W. (1996). The acid-mediated denaturation pathway of transthyretinyields a conformational intermediate that can self-assemble into amyloid. Biochemistry, 35, 6470–6482.
12. McParland, V. J., Kad, N. M., Kalverda, A. P., Brown,A., Kirwin-Jones, P., Hunter, M. G. et al. (2000). Par-tially unfolded states of beta-2-microglobulin andamyloid formation in vitro. Biochemistry, 39,8735–8746.
13. Khurana, R., Gillespie, J. R., Talapatra, A., Minert,L. J., Ionescu-Zanetti, C., Millett, I. & Fink, A. L.(2001). Partially folded intermediates as critical pre-cursors of light chain amyloid fibrils and amorphousaggregates. Biochemistry, 40, 3525–3535.
14. Ramirez-Alvarado, M. & Regan, L. (2002). Does thelocation of mutations determine the ability to formamyloid fibrils. J. Mol. Biol. 323, 17–22.
15. Chiti, F., Webster, P., Taddei, N., Clark, A., Stefani, M.,Ramponi, G. & Dobson, C. M. (1999). Designingconditions for in vitro formation of amyloid protofila-ments and fibrils. Proc. Natl Acad. Sci. USA, 96,3590–3594.
16. Morgan, C. J., Gelfand, M., Atreya, C. & Miranker,A. D. (2001). Kidney dialysis-associated amyloidosis:
940 Role of the Edge Strands in b2m Amyloid Formation
a molecular role for copper in fibre formation. J. Mol.Biol. 309, 339–345.
17. Khan, A. R., Baker, B. M., Ghosh, P., Biddison, W. E.& Wiley, D. C. (2000). The structure and stability ofan HLA-Ap201/octameric tax peptide complex withan empty conserved peptide-N-terminal bindingsite. J. Immunol. 164, 6398–6405.
18. Trinh, C. H., Smith, D. P., Kalverda, A. P., Phillips,S. E. & Radford, S. E. (2002). Crystal structure ofmonomeric human beta-2-microglobulin revealsclues to its amyloidogenic properties. Proc. NatlAcad. Sci. USA, 99, 9771–9776.
19. Gejyo, F., Yamada, T., Odani, S., Nakagawa, Y.,Arakawa, M., Kunitomo, T. et al. (1985). A new formof amyloid protein associated with chronic hemodia-lysis was identified as beta-2-microglobulin. Biochem.Biophys. Res. Commun. 129, 701–706.
20. Floege, J. & Ketteler, M. (2001). Beta-2-microglobulin-derived amyloidosis: an update. Kidney Int. 59,164–171.
21. Homma, N., Gejyo, F., Isemura, M. & Arakawa, M.(1989). Collagen-binding affinity of beta-2-microglo-bulin, a preprotein of hemodialysis-associated amy-loidosis. Nephron, 53, 37–40.
22. Smith, D. P., Jones, S., Serpell, L. C., Sunde, M. &Radford, S. E. (2003). A systematic investigation intothe effect of protein destabilisation on beta 2-micro-globulin amyloid formation. J. Mol. Biol. 330, 943–954.
23. McParland, V. J., Kalverda, A. P., Homans, S. W. &Radford, S. E. (2002). Structural properties of anamyloid precursor of beta-2-microglobulin. NatureStruct. Biol. 9, 326–331.
24. Katou, H., Kanno, T., Hoshino, M., Hagihara, Y.,Tanaka, H., Kawai, T. et al. (2002). The role of disul-fide bond in the amyloidogenic state of beta 2-micro-globulin studied by heteronuclear NMR. Protein Sci.11, 2218–2229.
25. Esposito, G., Michelutti, R., Verdone, G., Viglino, P.,Hernandez, H., Robinson, C. V. et al. (2000). Removalof the N-terminal hexapeptide from human beta-2-microglobulin facilitates protein aggregation andfibril formation. Protein Sci. 9, 831–845.
26. Eakin, C. M., Knight, J. D., Morgan, C. J., Gelfand,M. A. & Miranker, A. D. (2002). Formation of a cop-per specific binding site in non-native states of beta-2-microglobulin. Biochemistry, 41, 10646–10656.
27. Kad, N. M., Thomson, N. H., Smith, D. P., Smith,D. A. & Radford, S. E. (2001). Beta-2-microglobulinand its deamidated variant, N17D form amyloidfibrils with a range of morphologies in vitro. J. Mol.Biol. 313, 559–571.
28. Naiki, H., Hashimoto, N., Suzuki, S., Kimura, H.,Nakakuki, K. & Gejyo, F. (1997). Establishment of akinetic model of dialysis-related amyloid fibril exten-sion in vitro. Amyloid Int. J. Expt. Clin. Invest. 4,223–232.
29. Hoshino, M., Katou, H., Nakasima, Y., Hagihama, Y.,Hasegawa, K., Naiki, H. & Goto, Y. (2002). Mappingof the beta-2-microglobulin core by H/D exchangemonitored by NMR. Nature Struct. Biol. 9, 323–325.
30. Bellotti, V., Stoppini, M., Mangione, P., Sunde, M.,Robinson, C., Asti, L. et al. (1998). Beta-2-microglobu-lin can be refolded into a native state from ex vivoamyloid fibrils. Eur. J. Biochem. 258, 61–67.
31. Kozhukh, G. V., Hagihara, Y., Kawakami, T., Hase-gawa, K., Naiki, H. & Goto, Y. (2002). Investigationof a peptide responsible for amyloid fibril formationof beta 2-microglobulin by achromobacter proteaseI. J. Biol. Chem. 277, 1310–1315.
32. Ohashi, Y., Hagihara, Y., Kozhukh, G., Hoshino, M.,Hasegawa, K., Yamaguchi, I. et al. (2002). The intra-chain disulfide bond of beta-2-microglobulin is notessential for the immunoglobulin fold at neutral pH,but is essential for amyloid fibril formation at acidicpH. J. Biochem. 131, 45–52.
33. Jones, S., Manning, J., Kad, N. M. & Radford, S. E.(2003). Amyloid-forming peptides from beta 2-micro-globulin—insights into the mechanism of fibril for-mation in vitro. J. Mol. Biol. 325, 249–257.
34. Sinha, N., Tsai, C. J. & Nussinov, R. (2001). A pro-posed structural model for amyloid fibril elongation:domain swapping forms an interdigitating beta-structure polymer. Protein Eng. 14, 93–103.
35. Liu, Y., Gotte, G., Libonati, M. & Eisenberg, D. (2001).A domain-swapped RNase A dimer with impli-cations for amyloid formation. Nature Struct. Biol. 8,211–214.
36. Staniforth, R. A., Giannini, S., Higgins, L. D., Conroy,M. J., Hounslow, A. M., Jerala, R. et al. (2001). Three-dimensional domain swapping in the folded andmolten-globule states of cystatins, an amyloid-form-ing structural superfamily. EMBO J. 20, 4774–4781.
37. Christopher, J. A. (1998). SPOCK: the structural proper-ties observation and calculation kit. (The Center forMacromolecular Design, Texas A & M University,College Station).
38. Chiti, F., Mangione, P., Andreola, A., Giorgetti, S.,Stefani, M., Dobson, C. M. et al. (2001). Detection oftwo partially structured species in the folding pro-cess of the amyloidogenic protein beta 2-microglobu-lin. J. Mol. Biol. 307, 379–391.
Edited by J. Thornton
(Received 28 February 2003; received in revised form 20 May 2003; accepted 22 May 2003)
Role of the Edge Strands in b2m Amyloid Formation 941