Original article The effect of Saccharomyces cerevisiae and Aspergillus oryzae on the digestion of the cell wall fraction of a mixed diet in defaunated and refaunated sheep rumen Jean-Pierre Jouany a Frédérique Mathieu a Jean Senaud b Jacques Bohatier Gérard Bertin c Mariette Mercier d a Inra, SRNH, Centre de Clermont-Theix, 63122 Saint-Genès-Champanelle, France b Université Blaise-Pascal, URA CNRS 1944, Les Cézeaux, 63177 Aubière cedex, France c Santel, 22-24, rue du Président-Wilson, 92300 Levallois-Perret, France d Laboratoire de biostatistiques, Université de Franche-Comté, 4, place Saint-Jacques, 25030 Besançon, France (Received 16 September 1997; accepted 20 June 1998) Abstract - The objective of this study was to determine the effect of two probiotics, Saccharo- myces cerevisiae (SC) and Aspergillus oryzae (AO), without their culture medium, on the digestion of plant cell wall components in sheep that had been successively defaunated and refaunated. Six sheep fitted with large rumen cannulae were used to study 1) defaunated sheep with no probiotic, 2) defau- nated sheep with SC or AO, 3) refaunated sheep with no probiotic, 4) refaunated sheep with SC or AO. The apparent digestibility of the plant cell walls was not altered (P > 0.05) by the probiotics in defau- nated sheep but was increased (P < 0.05) with SC (+ 16 %) in refaunated sheep. Simultaneously, SC stimulated the growth of the protozoal population in the rumen. As noted in most previous experiments, the positive effect of the presence of protozoa on plant cell wall digestion (P < 0.001) was confirmed here. The effect (P > 0.05) of SC or AO on in situ ADF digestion was either not significant or nega- tive in defaunated rumens, whereas it became positive in refaunated rumen after a residence time of 12 h. The improvement of in situ ADF digestion due to the presence of protozoa was significant (P < 0.05 for NDF; P < 0.001 for ADF). However, we could not determine whether this was a direct effect of protozoa or an indirect effect operating via bacteria. The limits of the nylon bag technique for evaluating the microbial activity are discussed in relation to the ability of protozoa to enter and grow in the bags, and the pH regulation inside the bags. The pH values below 5.5 noted inside the bags can significantly alter the bacterial and protozoal populations and limit the validity of the technique. SC increased (P < 0.05) the specific activity of CMCase and xylanase of the solid-adherent bacteria (SAB) isolated from the rumen digesta of refaunated sheep. Stimulation of both the bacterial activity and protozoa numbers could explain the positive effect of SC on cell wall digestion in the rumen since the modelling of marker excretion in faeces showed that the ruminal mean retention time of hay measured from the model of Danhoa et al. was not modified (P > 0.05) by either refaunation or the * Correspondence and reprints E-mail: [email protected]
Transcript
Original article
The effect of Saccharomyces cerevisiae and Aspergillusoryzae on the digestion of the cell wall fraction of
a mixed diet in defaunated and refaunated sheep rumen
Jean-Pierre Jouanya Frédérique Mathieua Jean Senaudb
Jacques Bohatier Gérard Bertinc Mariette Mercierd
a Inra, SRNH, Centre de Clermont-Theix, 63122 Saint-Genès-Champanelle, Franceb Université Blaise-Pascal, URA CNRS 1944, Les Cézeaux, 63177 Aubière cedex, France
c Santel, 22-24, rue du Président-Wilson, 92300 Levallois-Perret, Franced Laboratoire de biostatistiques, Université de Franche-Comté, 4, place Saint-Jacques,
25030 Besançon, France
(Received 16 September 1997; accepted 20 June 1998)
Abstract - The objective of this study was to determine the effect of two probiotics, Saccharo-myces cerevisiae (SC) and Aspergillus oryzae (AO), without their culture medium, on the digestionof plant cell wall components in sheep that had been successively defaunated and refaunated. Six sheepfitted with large rumen cannulae were used to study 1) defaunated sheep with no probiotic, 2) defau-nated sheep with SC or AO, 3) refaunated sheep with no probiotic, 4) refaunated sheep with SC or AO.The apparent digestibility of the plant cell walls was not altered (P > 0.05) by the probiotics in defau-nated sheep but was increased (P < 0.05) with SC (+ 16 %) in refaunated sheep. Simultaneously, SCstimulated the growth of the protozoal population in the rumen. As noted in most previous experiments,the positive effect of the presence of protozoa on plant cell wall digestion (P < 0.001) was confirmedhere. The effect (P > 0.05) of SC or AO on in situ ADF digestion was either not significant or nega-tive in defaunated rumens, whereas it became positive in refaunated rumen after a residence time of12 h. The improvement of in situ ADF digestion due to the presence of protozoa was significant(P < 0.05 for NDF; P < 0.001 for ADF). However, we could not determine whether this was a directeffect of protozoa or an indirect effect operating via bacteria. The limits of the nylon bag techniquefor evaluating the microbial activity are discussed in relation to the ability of protozoa to enter andgrow in the bags, and the pH regulation inside the bags. The pH values below 5.5 noted inside the bagscan significantly alter the bacterial and protozoal populations and limit the validity of the technique.SC increased (P < 0.05) the specific activity of CMCase and xylanase of the solid-adherent bacteria(SAB) isolated from the rumen digesta of refaunated sheep. Stimulation of both the bacterial activityand protozoa numbers could explain the positive effect of SC on cell wall digestion in the rumen sincethe modelling of marker excretion in faeces showed that the ruminal mean retention time of haymeasured from the model of Danhoa et al. was not modified (P > 0.05) by either refaunation or the
The intensive production of milk and meatthat has developed over the last 40 yearsrequires high-energy diets. Feed additiveshave been proposed to stabilize and stimulate
the digestive efficiency of the ruminal eco-system, thereby improving the nutrient sup-ply to ruminants. Among these additives,direct-fed microbials (DFM) are viewed asbiologically harmless agents, whereas chem-ical additives such as antibiotics are consi-
dered risky. Also, European Union regula-tions restrict these to veterinary use.
Live yeasts and fungi have been extensi-vely used as feed additives over the lastdecade, but their real efficiency for rumi-nants is still being debated [45]. Early stu-dies [1, 42] indicated that DFMs increasedfeed intake. According to Wallace and New-bold [45], this could result from a stimulatedrate of fibre degradation. However, a fullsearch of the literature shows that no consis-tent effect of DFM has ever been obtainedon ruminal plant cell wall degradation [35].
The objective of this study was to exa-mine the effect of supplemental Saccharo-myces cerevisiae (SC) and Aspergillus ory-zae (AO) on several parameters involved inthe digestion of cell wall carbohydrates. Theeffects of the two probiotics were measuredfirst in defaunated sheep and then in thesame refaunated sheep, because recent workhas indicated that these probiotics canchange the ruminal protozoa population [3,10, 34]. This experimental design enabled usto analyse the interactions between proto-zoa and probiotics for different factorscontrolling the digestion of plant cell walls.
2. MATERIALS AND METHODS
A total of nine castrated male adult Texel
sheep (72.5 ± 5.0 kg LW) fitted with rumen can-nulae and labelled from A to I were used. Theywere defaunated by emptying and washing therumen according to Jouany and Senaud [21]. Theexperiment began following a 6-week period ofadaptation to the defaunated state. Rumencontents were examined each week to verify theabsence of protozoa within the rumen. AnimalB died at the end of the third period followingnecrosis of the liver aggravated by heparin injec-tions during blood sampling. Animals E and Fdied after undergoing two closely spaced suc-cessive defaunation treatments following an acci-dental contamination with small entodinia at the
end of the second period.
The two probiotics were a strain of Saccha-romyces cerevisiae (SC, registered as CNCM I-1096, Institut Pasteur, France), and a strain ofAspergillus oryzae (AO) of industrial origin. Thetwo additives were composed of living cells(20 x 109 cells/g for SC; 6.5 x 108 spores/g forAO) without their culture medium, and were sup-plied by Santel*. They were administered throughthe rumen cannulae, just before the morningmeal, so the exact daily dose of the two addi-tives was known.
The experimental design has been describedby Mathieu et al. [26]. During the first period,the six sheep used (A-F) were defaunated andreceived no probiotics. During the second per-iod, three of them (A-C) received SC (50 mg/day),while the other three were fed AO (3 g/day). Thesedoses are commonly used in current practice.During the third period, six sheep (A-D, G, H)were each refaunated with 200 mL of rumencontents containing Isotricha (103/mL), Epidi-nium (104/mL), Eudiplodinium (104/mL) andEntodinium (105/mL). Measurements started 4weeks after the ciliate population had stabilized.During the fourth period, three sheep (A, I, C)received SC (50 mg/day); three others (D, G, H)were fed AO (3 g/day). Periods 2 and 4 began2 weeks after the addition of probiotics althoughthe recommended level of live cells was reachedin the rumen digesta on the day following thefirst administration (Chaucheyras, pers. comm.). ).
The sheep were housed in individual stalls ina building specifically designed to avoid all directcontact between animals and so prevent conta-mination. They received 1 350 g/day of a mixeddiet (table n fed twice daily in equal meals, at0900 and 1600 hours. Because of the high pro-portion of barley, the sheep were adapted to thediet gradually for 1 month before measurementsbegan. The measurements were carried outconcurrently on the six sheep using the sameexperimental design for each period (figure 1 ).
Rumen fluid samples were taken for pH deter-mination from the bottom of the ventral sac bysuction through a tube ( cm in internal diameter)connected to a large rubber teat. Total rumendigesta were collected by completely emptyingthe rumen 5 h after the morning meal. Thevolume and weight of total digesta were deter-mined. A representative sample of digesta wastaken for analyses of DM, OM, NDF, ADF, ADLand determination of their ruminal pools.
* SANTEL, 22-24, rue du Pr6sident-Wilson, 92300 Levallois-Perret, France.
Nylon bags** (5 x 10 cm; 53 microns poresize) filled with 3 g (DM) of hay from the dietwere incubated (0, 1, 3, 6, 12, 24, 48, 72 h) usingthe in situ technique [27]. Hay was ground usinga 4-mm screen before being introduced into thebags using the standard method developed atInra. Kinetic studies were repeated six times persheep. Bags removed from the rumen were gentlyhand-pressed before washing in cold tap wateruntil the rinse water was clear. Because therewas no plateau after a 72-h retention time in therumen, the kinetic data could not be fitted to themodel described by 0rskov and Mc Donald [32].The pH was determined in the liquid extracted
from the bags by hand-pressure and the proto-zoa contained in this liquid were counted.
A single dose of 20 g chopped hay previouslylabelled with ytterbium acetate (12.5 mg Yb/gDM) according to Bernard [4] was introducedinto the rumen of each animal via the cannula toevaluate the mean retention time of forage par-ticles. Following this addition, the faeces werecollected every 6 h for the first 24 h, then every3 h up to 60 h, then every 6 h up to 96 h, thenevery 12 h up to 168 h. Yb was assayed by ato-mic absorption spectrophotometry (Perkin-ElmerSpectrophotometer 2380) as described by Sid-
** ANKOM, 140 Turk Hill Park Fairport, NY 14450, USA.
dons et al. [37]. Solutions for calibration plotswere prepared with unlabelled faeces from thesheep. The mean retention times of hay particlesin the whole digestive tract and its compartmentswere calculated from the models applied to thekinetic excretion of Yb in faeces and established,respectively, by Thielemans et al. [39], Danhoaet al. [9] and Faichney and Boston [11]. I -
The turnover of the liquid phase (k) in therumen was measured from the kinetic decrease
(2, 4, 6, 8, 10, 12, 18, 24, 32 h) of the solublemarker PEG 4000 introduced through the rumencannulae in a single dose (100 mL of PEG solu-tion 20 % w/v ) just before the morning meal.The dilution rate was calculated from the equa-tion: Ct = Co x e(-kt). This determination wasrepeated twice for each animal. PEG was assayedaccording to Hyden [18] using an autoanalyser.
Enzymatic activities were determined onliquid associated bacteria (LAB) and on solidadherent bacteria (SAB) only during periods 3and 4, when the sheep were refaunated. Liquidsamples taken from the rumen just before themorning meal were filtered on two gauze layersunder anaerobic conditions. Filtrates were cen-
trifuged at 1 000 g for 10 min to remove protozoaand feed particles. The supernatants were cen-trifuged at 20 000 g for 20 min to isolate theLAB fraction. All centrifuging was carried outunder anaerobiosis. Fifteen grams of ground hay(4-mm screen) from the diet was placed in nylonbags (8 x 18 cm, 95 microns pore size), whichwere then left in the rumen for 24 h. Removed
bags were preserved in airtight bottles. Ten-gramwet samples from the bags were suspended in10 mL of buffer MES (4.88 g/L, pH 6.5) in poly-ethylene bags and treated by a stomacher for4 min under C02 to detach the SAB fraction.The suspensions of SAB were then treated likethe LAB. Samples were ultrasonicated for 2 min(four periods of 30 s) to release the enzymes.Liquids were centrifuged at 3 000 g for 20 minand the enzymes were analysed in the superna-tants according to Williams and Withers [48].Protein contents in the enzymatic preparationswere determined according to Bradford [5].
NDF, ADF and ADL were determined byprocedures outlined by Goering and Van Soest[14]. Protozoa were counted in rumen fluid andin the liquid extracted by hand-pressure fromnylon bags with a Dollfuss cell as described byJouany [19]. Dry matter (DM) was determined byoven drying at 80 °C for 48 h. Organic matter(OM) was measured after ashing in a muffle fur-nace (550 °C for 6 h).
The experimental design was chosen to pre-vent protozoal contamination of the sheep during gthe defaunated state. The study of the effect ofprobiotics was thus carried out first on defauna-ted sheep. We subsequently refaunated all theanimals and ran the two experiments with andwithout probiotics. Data were processed byvariance analysis using the GLM procedure ofSAS [36] using the following model: Y!.k = m+ Ai + F + F.(Pk) + Eij’k, where Y = measuredparameter, m = overall mean, A = animal effect,F = fauna effect, F. (Pk) = probiotic effect nestedwithin fauna effect, and E = residual error. Thismodel, in which the probiotic effect is ’nestedwithin the fauna effect’, was well-adapted to ourexperimental design, especially where there is asignificant interaction between fauna and pro-biotics. Interactions between protozoa and pro-biotics were assessed with a second model: Y ii -k= m + Ai + F. + Pk + FPjk + Eijk’ where Y = mea-sured parameter, m = overall mean, A = animaleffect, F = fauna effect, P = probiotic effect, FP= interaction between fauna and probiotic, and E= residual error. The option ’repeated measuresfor variance analyses’ was used to take intoaccount the non-independence of the data in allthe kinetic studies performed here. Because dataexpressed in percentages do not have a normaldistribution, they were converted into their ’arc-sinus’ values before the statistical analysis. Meanswere compared in pairs using the Duncan mul-tiple rank test. The significance threshold wasset at 5 %.
3. RESULTS AND DISCUSSION
The apparent total tract digestion of thecomponents of the diet was not modified bythe addition of AO or SC (P > 0.05) in thedefaunated rumens (table In. In contrast,both DFMs stimulated the digestibility ofthe dietary OM (P < 0.05) in refaunatedsheep. The hemicellulose and cellulose frac-tions of the diet estimated as (NDF - ADF)and (ADF - lignin), respectively, were alsosignificantly (P < 0.05) better digested afterthe addition of SC in refaunated animals.We confirmed here that protozoa have astrong stimulating effect (P < 0.001 for the
global effect of protozoa) on the apparenttotal tract digestion of OM and cell wallcomponents, mainly accounted for by a
powerful stimulation of the ruminal diges-tion as discussed by Jouany [20].
in situ hay ADF disappearance was onlysignificantly (P < 0.05) stimulated by theaddition of protozoa after a 12- or 24-h incu-bation time in the rumen when DFMs wereadded or a 48-h incubation time in the non-treated animals (table 111). The same effectof protozoa on NDF degradation was obser-ved in the rumen of the DFM-treated ani-mals while no effect or a significantdecrease was noted during the first 12 h ofthe kinetics. SC and AO had either noeffect or decreased (P < 0.05) the in situdegradation of NDF and ADF fractions indefaunated animals. This could indicatethat SC operates in cell wall digestion viathe rumen protozoa. Our results concer-
ning the global positive effect of protozoaon the in situ degradation of hay cell wallsshown by variance analysis (P < 0.05 for
NDF; P < 0.001 for ADF) confirmed theabundant data currently available in the lit-erature (see Jouany and Martin [22]).
The modifications reported in the litera-ture on the in situ degradation of forage cellwalls induced by probiotics range widelyand are generally non-significant [13, 15,16, 25, 33, 43, 44]. For example, Wallaceet al. [46] indicated that the rate of degra-dation was stimulated by SC while the maxi-mum level of degradation was not influen-ced. In agreement with our results, Roa et al.[35] and Miranda et al. [29] observed thatthe degradation rate of alfalfa hay cell wallswas not influenced by SC during short incu-bations of bags in the rumen, whereas theamount of degraded cell walls can increase(P < 0.05) after 24 h incubation. These dis-crepancies are probably due to a strain effectof the yeasts as shown by Jouany et al. [23]and Newbold et al. [30], and to experimen-tal conditions such as diet and level of feedintake. Not enough results are available inthe literature for a precise explanation ofthe differences in the mode of action ofDFMs on plant cell wall degradation. Alongwith most authors we consider that DFMs
are mainly active at the rumen level, prin-cipally when the ruminal ecosystem is dis-turbed by extreme dietary conditions such ashigh-starch diets. They have been shown tostabilize the pH and prevent lactate accu-mulation in the rumen [26, 28]. Our resultsconfirm that such a stabilizing effect ofDFMs is probably involved in the impro-vement of the in situ ADF degradationbeyond the 12-h residence time in normallyfaunated rumen [26]. We can also supposethat the presence of large amounts of bar-ley (45 % of dietary DM in our experiment),as a cereal rich in rapidly degradable starch,depressed the ruminal fibre degradation [38].As a consequence, the intestinal digestionof plant cell wall components increased at
the expense of the ruminal digestion.According to Archimède et al. [2], who ana-lysed 348 results from 113 recent digestivetrials, the intestinal digestion of plant cellwall components increases with the dietarylevel of concentrate but remains less than20 % of the total digestible cell walls.Because Newbold et al. [30] showed thatsurvival live cells of SC are found at theileal level at the same concentration as in
the rumen, they considered that probioticscould also act at the intestinal level, essen-
tially at the large intestine level for the diges-tion of cell walls. In the absence of intesti-nal cannula on our sheep it was impossibleto allow for the partition between the rumi-nal and intestinal digestion of dietary plant
cell walls. However, 1) the close agreementbetween the positive effect of SC on thedigestion of cellulosic and hemicellulosicfractions, and 2) the stimulating effect ofSC on the specific activity of carboxyme-thycellulase in the rumen SAB and LABand on the xylanase activity in the rumenSAB (table IV), indicates that SC acts pri-marily at the rumen level. Likewise, theabsence of any effect of AO on either thetotal cell wall digestion or the rumen bacte-rial enzyme activities supports this hypo-thesis. The reason why SC decreased the(3D-glucosidase activity in the SAB and bothprobiotics had a negative effect on the (3D-xylosidase in LAB is difficult to explain.
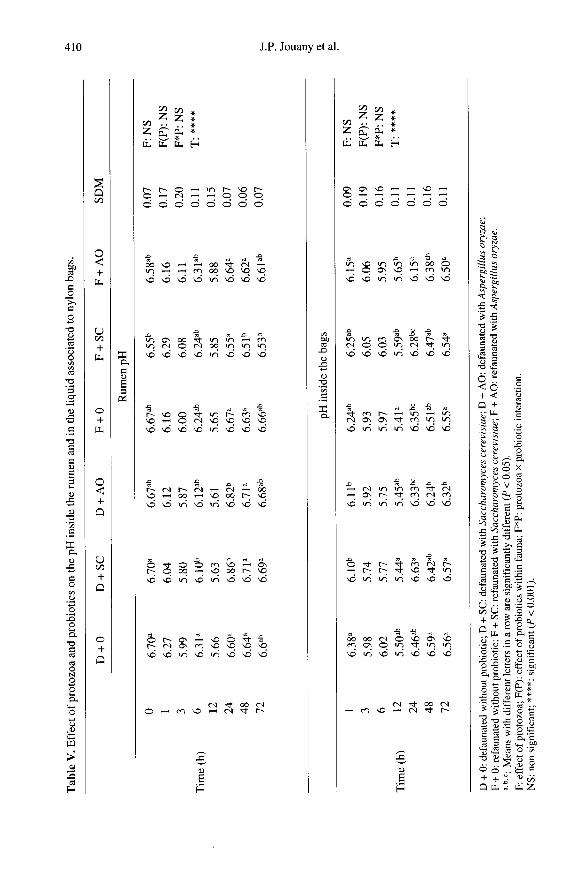
During the part of our study carried outwith the nylon bags, we measured the pHand determined the protozoa populationsinside the bags to compare the values withthose obtained in the rumen. Due to a defi-cit of exchange with the rumen liquid asindicated by Trabalza-Marinucci [40], thepH values in the bags were 0.1 to 0.6 unitslower than the values obtained in the rumen(table V) and the protozoa were less nume-rous (table Vn. Compared with the rumen,large-size protozoa were three times lessnumerous in nylon bags, small entodiniadecreased by 30 %, and holotrichs showedinconsistent modifications. The protozoalpopulation became established after 1 h inthe bags and then remained stable for 72 h.This may indicate that the growth and acti-vity of protozoa were limited by unfavour-able environmental conditions inside the
bags. If, as we assume, the positive effectof probiotics mainly observed with SC isalso explained by a stimulation of the pro-tozoal population in the rumen as observedin this experiment (table Vn and in otherswith SC [3, 34] and AO [12]. This effectdid not fully take place in the nylon bagswhere the concentration of protozoa wasless strongly influenced by the two probio-tics than in the rumen, which can explainwhy no effect of probiotics (P > 0.05 forthe global effect of probiotics) appeared inour study on in situ degradation of cell walls
(table III). Of course, the stimulation notedon protozoa at the rumen level induceschanges in the bacterial population [22]. It isgenerally considered that protozoa have adetrimental effect on amylolytic bacteriaand a positive effect on the bacterial cellu-lolytic activity in the rumen [41, 48]. Howe-ver, the latter cannot be demonstrated inside
nylon bags, as reported by Nozi6re andMichalet-Doreau [31], who showed that theactivities of SAB were lower in bag resi-dues than in rumen digesta. This limitation,added to the reserves expressed on the per-meability of the nylon tissue to protozoaand pH regulation, means the nylon bagtechnique is not sensitive enough for thetesting of feed additives in such dietaryconditions.
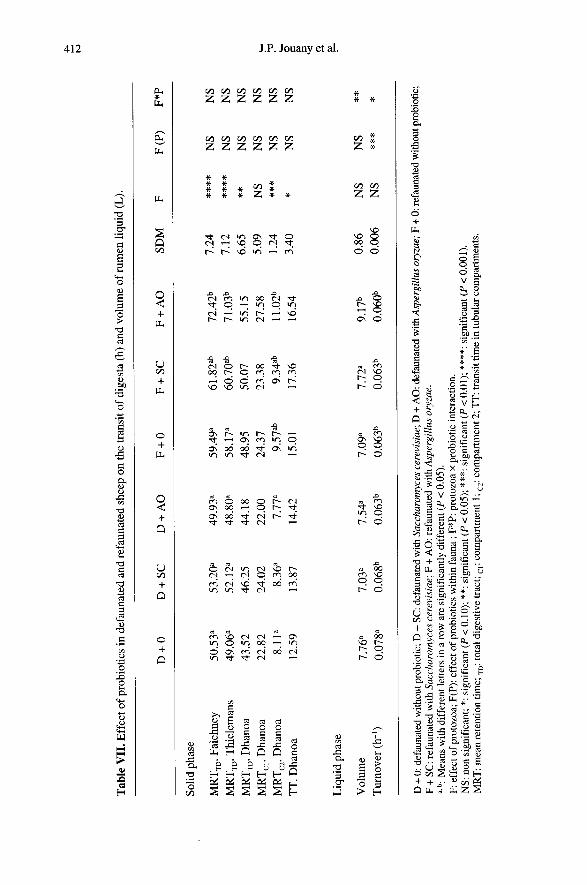
Addition of probiotics in the rumen ofdefaunated sheep decreased (P < 0.05) theturnover of the ruminal liquid phase (tableVII). Unlike Yoon and Stern [49], whoobserved a significant decrease (P > 0.05) inthe liquid turnover following the additionof SC in the rumen of cows, we noted noeffect of probiotics on this parameter mea-sured in refaunated sheep. Comparisonsmade on defaunated and refaunated sheepwithout probiotics indicated that protozoadecreased the turnover of the ruminal liquidphase. Differences between defaunated andrefaunated sheep disappeared when probio-tics were given.
In agreement with Caton et al. [6], West-wig et al. [47], Plata et al. [34], there wasno significant overall effect of the probioticson the retention time of solid particles inthe digestive tract whatever the model used(table VII). The retention time of digesta inthe total digestive tract was significantlylengthened with AO when the faecal excre-tion of marker was analysed according toThielemans et al. [39] and Faichney andBoston [11]. The increase was not signifi-cant (P > 0.05) when the model of Dhanoaet al. [9] was used. These models gave noinformation on the retention of digesta inthe caecum and colon which could support
any shift of cell wall digestion from therumen to the large intestine.
From comparisons between defaunatedanimals without probiotics and refaunatedanimals without probiotics, we observedthat the presence of protozoa significantly(P < 0.001) lengthened the mean retentiontime in the whole digestive tract by 5 to 10h, whereas no significant effect of protozoawas noted on the retention time of particlesin ’compartment 1’, which is often identifiedas the rumen. However, as indicated above,it is impossible to assert from these tooimprecise models that digesta are retainedlonger in the intestinal portion of the digest-ive tract. Literature data on the effect of pro-tozoa on the transit time of solid particlesare conflicting (see [7]). De Smet et al. [8]suggest that protozoa increase the retentionof particles in sheep fed diets rich in concen-trate but decrease it in sheep fed roughagediets. Our results fit this hypothesis. Howe-ver, recent experiments carried out byHegarty et al. [17] and Jouany et al. [24]confirm the wide range of individual ani-mal response to the addition of protozoa intheir previously defaunated rumens on thetransit time of rumen digesta.
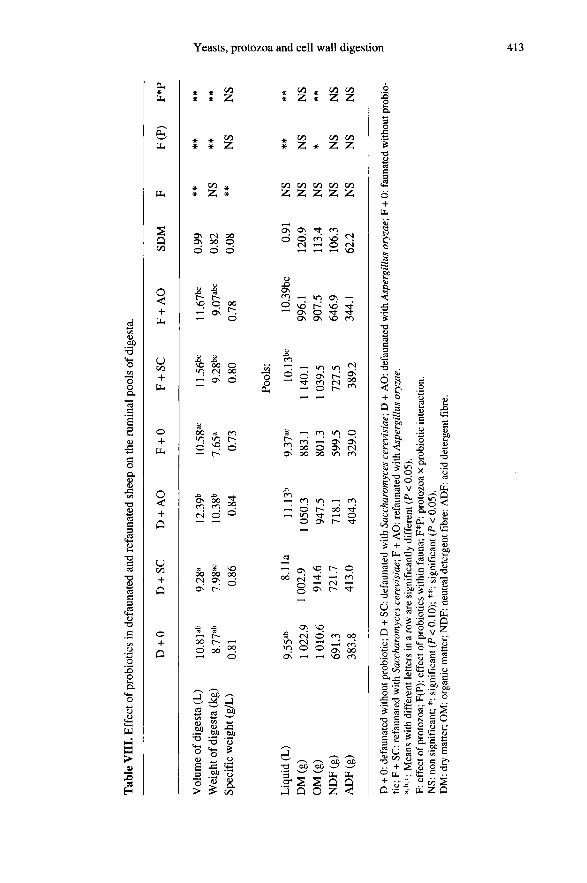
The volume and the ruminal pool of totaldigesta measured 5 h after feeding in the defau-nated animals were significantly (P < 0.05)
higher in the presence of AO compared withSC in defaunated sheep. Differences be-tween the two probiotics disappeared inrefaunated sheep (table VIII). AO increa-sed the total pool of liquid only in the defau-nated rumen (P < 0.05). Neither SC nor AOaltered the pools of NDF or ADF in therumen (P > 0.05). Because the animals weregiven constant amounts of feed, this resultmeans that the rate of cell wall digestionwas not changed for the first 5 h by the addi-tion of SC or AO, which agrees with theresults from our kinetic in situ study. Thelower specific weight (P < 0.05) of therumen contents in the faunated comparedwith the defaunated state may be due to the
higher production of fermentation gases inthe presence of protozoa resulting from lar-ger amounts of fermented OM [20], smallgas bubbles being in that case trapped in therumen digesta and decreasing their specificweight.
4. CONCLUSION
SC without its culture medium stimulatedthe apparent digestibility of dietary OM inthe whole digestive tract of refaunated ani-mals. This result is explained by the positiveeffect of SC on the digestibility of plant cell
walls. AO had no effect on plant cell walldigestion. SC increased the population ofprotozoa and stabilized the rumen pH ondiets rich in rapidly fermentable starch,which explains the stimulation of the bac-terial specific activity of polysaccharidedepolymerases involved in the degradationof hemicellulose and cellulose, thus impro-ving the total cell wall digestion observed inrefaunated animals. Protozoa increased the
digestive retention time of solid particleswhich could explain the positive effect ofSC on cell wall digestion in these animals.The results obtained with nylon bags are cri-tically assessed. Probiotics had no effect onthe ruminal retention time of solid particles.
ACKNOWLEDGEMENTS
The authors are indebted to A.G. Williamsfor his advice and help for the determination ofthe enzymatic activities of rumen microbes. Theythank Sylvie Toillon and M. Fabre for their tech-nical assistance, and J. Lefaivre for surgery.
REFERENCES
[1] ] Adams D.C., Galyean M.L., Kiesling H.E., Wal-lace J.D., Finker M.D., Influence of viable yeastculture, sodium bicarbonate and monensin onliquid dilution rate, rumen fermentation andfeedlot performance of growing lambs anddigestibility in lambs, J. Anim. Sci., 53 (1981)780-789.
[2 Archim6de H., Sauvant D., Schmidely P., Quan-titative review of mineral and total tract digestionof mixed diet organic matter and carbohydrates,Reprod. Nutr. Dev. 37 (1997) 173-189.
[3] Ayala O.J., Gonzales S.S., Herrera R., Barcena R.,Mendoza G.D., Effect of a probiotic and amolasses-urea supplement on fiber digestibilityof sesame straw, J. Anim. Sci. (1992) 70 (suppl.1 ) 307.
[4] Bernard L., Etude de la dynamique des parti-cules et des liquides dans le r6ticulo-rumen, chezle mouton recevant une ration de foin de dac-
tyle sous forme hachde et broy6e en différentesproportions, These d’Universit6, UniversiteBlaise Pascal de Clermont-Ferrand II, n° d’ordre466, 1992, 180 pp.
[5] Bradford M., A rapid and sensitive method forthe quantitation of microgram quantities of pro-tein utilizing the principle of protein-dye brin-ding, Anal. Biochem. 72 (1996) 248-254.
[6] Caton J.S., Erickson D.O., Carey D.A., Ulmer D.L.,Influence of Aspergillus oryzae fermentationextract on forage intake, site of digestion, in situdegradability, and duodenal amino acid flow insteers grazing cool-season pasture, J. Anim. Sci.71 (1993) 779-787.
[7] Demeyer D.I., Effect of defaunation on rumenfibre digestion and digesta kinetics, in: Nolan J.V.,Leng R.A., Demeyer D.I. (Eds.), The Roles ofProtozoa and Fungi in Ruminant Digestion,Penambul Books, Armidale, Australia, 1989,pp.171-179.
[8] De Smet S., Demeyer D.I., Van Nevel C.J.,Effect of defaunation and hay: concentrate ratioon fermentation, fiber digestion and passage inthe rumen of sheep, Anim. Feed Sci. Technol. 37(1992) 333-344.
[9] Dhanoa N.S., Siddons R.C., France J., Gale D.L.,A multicompartmental model to describe markerexcretion patterns in ruminant faeces, Br. J. Nutr.53 (1985) 663-371. 1 .
[10] Edwards I.E., Practical uses of yeast culture inbeef production: insight into its mode of action,in: Lyons T.P. (Ed.), Biotechnology in the FeedIndustry, Alltech Techn. Publ., Nicholasville,USA, 1991, pp. 51-63.
[11] Faichney G.J., Boston R.C., Interpretation ofthe faecal excretion patterns of solute and par-ticle markers introduced into the rumen of sheep,J. Agric. Sci. (Cambridge) 101 (1983) 575-581. 1 .
[12] Fondevila M., Newbold C.J., Hotten P.M.,0rskov E.R., A note on the effect ofAspergillusoryzae fermentation extract on the rumen fer-mentation of sheep given straw, Anim. Prod. 51 1(1990) 422!25.
( 13] Frumholz P.P., Manipulation of the rumen fer-mentation and its effects on digestive physio-logy, Ph.D. dissertation, University of Aber-deen, Scotland, 1991, 240 pp.
[ l4] Goering H.K., Van Soest P.J., Forage fiber ana-lysis (apparatus, reagents, procedures and someapplications), Agric. Handbook N° 379, ARS-USDA, Washington, DC, USA, 1970.
[15] Gomez-Alarcon R.A., Effects of Aspergillusoryzae on milk production, feed utilization andrumen fermentation in lactating dairy cows,Ph.D. dissertation, University of Arizona, Tuc-son, USA, 1988, 102 pp.
[16] Guilford R.J., Olson K.C., Caton J.S., KirbyD.R., Influence of yeast culture supplementa-tion and advancing season on in situ NDF degra-dability in beef cattle grazing native range inthe Northern great plains, J. Anim. Sci. 71 1(suppl. 1) (1993) 83.
[ 17] Hegarty R.S., Nolan J.V., leng R.A., The effectsof protozoa and of supplementation with nitro-gen and sulfur on digestion and microbial meta-bolism in the rumen of sheep, Austr. J. Agric.Res. 45 (1994) 1215-1227.
[18] Hyden S., A turbidimetric method for the deter-mination of higher polyethylene glycols in bio-logical materials, Ann. Lantlrhghs. 22 (1955)139-145.
[19] Jouany J.P., Contribution a l’étude des proto-zoaires cili6s du rumen: leur dynamique, leur roledans la digestion et leur interet pour le ruminant,These de Doctorat, Université Blaise-Pascal deClermont-Ferrand, Vol. 1 (text), 1978, 195 pp.
[20] Jouany J.P., Defaunation of the rumen, in:Jouany J.P. (Ed.), Rumen Microbial Metabo-lism and Ruminant Digestion, Inra Editions,Science Update, Versailles, France, 1991, pp.239-261.
[21 Jouany J.P., Senaud J., Defaunation du rumen demouton, Ann. Biol. Anim. Bioch. Biophys. 19(1979) 619-624.
[22] Jouany J.P., Martin C., Effect of protozoa inplant cell wall and starch digestion in the rumen,in: Onodera R. et al. (Eds.), Japan Sci. Soc. Press,Tokyo/S. Karger, Basel, Japan, 1997, pp. 11-24.
[23] Jouany J.P., Fonty G., Lassalas B., Dore J.,Gouet Ph., Bertin G., Effect of live yeast cul-tures on feed degradation in the rumen as asses-sed by in vitro measurements, 21st BiennalConference on Rumen Function, Chicago, Illi-nois, USA, Abstract 6, 1991.
[24] Jouany J.P., Senaud J., Toillon S., Ben SalahM., Bohatier J., Prensier G., Effect of ruminalinoculation of Isotricha alone or a mixed (3-typefauna in a defaunated rumen on the digestionof a hay-maize diet (70/30) in sheep, Reprod.Nutr. Dev. 35 (1995) 11-25.
[25] Kumar U., Sareen V.K., Singh S., Effect of Sac-charomyces cerevisiae yeast culture supplementon ruminal metabolism in buffalo calves givena high concentrate diet, Anim. Prod. 59 (1994)209-215.
[26] Mathieu F., Jouany J.P., Senaud J., Bohatier J.,Bertin G., Mercier M., The effect of Saccharo-myces cerevisiae and Aspergillus oryzae on fer-mentations in the rumen of faunated and defau-nated sheep; protozoal and probiotic interactions,Reprod. Nutr. Dev. 36 (1996) 271-287.
[27] Mehrez A.Z., 0rskov E.R., A study of artificialbag technique for determining the digestibility offeed in the rumen, J. Agric. Sci. (Cambridge)88 (1997) 645-660.
[28] Michalet Doreau B., Morand D., Martin C.,Effect of the microbial additive Levuce110O SCon microbial activity in the rumen during thestepwise adaptation of sheep to high concen-trate diet, Reprod. Nutr. Dev. 5 (suppl.) (1997)81-82.
[29] Miranda R.L.A., Mendoza M.G.D., Barcena-Gama J.R., Gonzales M.S.S., Ferrara R., OrtegaC.M.E., Cobos P.M.A., Effect of Saccharomycescerevisiae or Aspergillus oryzae cultures andNDF level on parameters of ruminal fermenta-tion, Anim. Feed Sci. Technol. 63 (1996)289-296.
[30] Newbold C.J., Wallace R.J., Chen X.B., McIntosh F.M., Different strains of Saccharomycescerevisiae differ in their effects on ruminal bac-teria numbers in vitro and in sheep, J. Anim.Sci. 73 (1995) 1811-1818.
[31 ] Noziere P., Michalet Doreau B., Validation of insacco method: influence of sampling site, nylonbag or rumen contents on fibrolytic activity ofsolid-associated microorganisms, Anim. FeedSci. Technol. 57 (1996) 203-210.
[32] 0rskov E.R., Mc Donald LW., The estimation ofprotein degradability from incubation measure-ments weighted according to rate of passage, J.Agric. Sci. (Camb.) 92 (1979) 499-503.
[33] Plata P.F., Gonzales S.S., Mendoza G., BarcenaR., Effect of a yeast culture (Saccharomycescerevisiae) on nutritive value of oat straw baseddiets fed to Holstein cows, J. Anim. Sci. 71 1
(suppl. 1) (1993) 307.
[34] Plata P.F., Mendoza M.G.D., Barcena-GamaJ.R., Gonzales M.S., Effect of yeast culture (Sac-charomyces cerevisiae) on neutral detergentfiber digestion in steers fed oat straw based diets,Anim. Feed Sci. Technol. 49 (1994) 203-210.
[35] Roa M.L., Barcena-Gama J.R., Gonzales M.S.,Mendoza G.M., Ortega M.E., Garcia B.C.,Effect of fiber source and a yeast culture (Sac-charomyces cerevisiae 1026) on digestion andthe environment in the rumen of cattle, Anim.Feed Sci. Technol. 64 (1997) 327-336.
[36] SAS Institute Inc., SAS-STAT, Guide for Per-sonal Computers, version 6, Cary, NC, USA,1987.
[37] Siddons R.C., Paradine J., Beever D.E., Comell P.R.,Ytterbium acetate as a particulate phase digesta-flow marker, Br. J. Nutr. 54 (1985) 509-519.
[38] Tamminga S., Influence of feeding managementon ruminant fiber digestibility, in: Jung H.G.,Buxton D., Hatfield R.D., Ralph J. (Eds.), ForageCell Wall Structure and Digestibility, ASACSSA SSA, Madison, 1993, pp. 571-602.
[39] Thielemans R.F., Francois E., Bodart C., The-wis A., Mesure du transit gastro-intestinal chezle porc a I’aide de radiolanthanides. Comparai-son avec le mouton, Ann. Biol. Anim. Bioch.
Biophys. 18 (1978) 237-247.
[40] Trabalza-Marinucci M., Dehority B.A., Loech S.C.,in vitro and in vivo studies of factors affectingdigestion of feeds in synthetic fiber bags, J.Anim. Sci. 70 (1992) 296-307.
[41 ] Ushida K., Jouany J.P., Demeyer D.E., Effectsof presence or absence of rumen protozoa onthe efficiency of utilization of concentrate onfibrous feeds, in: Tsuda T., Sasaki Y., Kawa-shima R. (Eds.), Physiological Aspects of Diges-tion and Metabolism in Ruminants, AcademicPress, Tokyo, 1991, pp. 625-654.
[42] Van Horn H.H., Harris B., Taylor M.J., Bach-man K.C., Wilcox C.J., By-products feeds forlactating dairy cows: effects of cottonseed hulls,sunflower hulls, corrugated paper, peanut hulls,sugarcane bagasse and whole cottonseed withadditives of fat, sodium bicarbonate and Asper-gillus oryzae product on milk production, J.Dairy Sci. 67 (1984) 2922-2938.
[43] Varel V.H., Krieikmeir K.K., Influence of fee-ding Aspergillus oryzae fermentation extract(Amaferm) on in situ fiber degradation, rumi-nal fermentation, and microbial protein synthe-sis in non lactating cows fed alfalfa or brome-grass hay, J. Anim. Sci. 72 (1994) 1814-1822.
[44] Varel V.H., Krieikmeir K.K., Response tovarious amounts of Aspergillus oryzae fermen-tation extract on ruminal metabolism in cattle, J.Dairy Sci. 77 (1994) 3081-3086.
[45] Wallace R.J., Newbold C.J., Probiotics for rumi-nants, in: Fuller R. (Fd.), Probiotics, The Scien-tific Basis, Chapman and Hall, London, 1992,pp. 317-353.
[46] Wallace R.J., Newbold C.J., Mc Intosh F.M.,Influence of Saccharomyces cerevisiaeNCYC240 and malic acid on bacterial numbersand fiber breakdown in the sheep rumen, J.Anim. Sci. 71 (suppl. 1) (1993) 287.
[47] Westig R.M., Caton J.S., Erickson D.O.,Influence of barley and Aspergillus oryzae fer-mentation extract supplementation on micro-bial efficiency, duodenal crude protein andamino acid flows, and digesta kinetics in steersfed prairie hay, J. Anim. Sci. 71 (suppl. 1 ) ( 1993)81. 1 .
[48] Williams A.G., Withers S.E., Changes in therumen microbial population and its activitiesduring the refaunation period after the reintro-duction of ciliate protozoa into the rumen ofdefaunated sheep, Can. J. Microbiol. 39 (1993)61-69.
[49] Yoon I.K., Stern M.D., Effects of Saccharo-myces cerevisiae and Aspergillus oryzae cul-tures on ruminal fermentation in dairy cows, J.Dairy Sci. 79 (1996) 411-417.