14
Science for Defra: excellence in the application of evidence Defra Science Showcase Session 6 Animal health and food and farming Chair: Professor James Wood

Science for Defra: excellence in the application of evidence
Defra Science Showcase Session 6
Animal health and food and farming
Chair: Professor James Wood

New paradigms to solve the aquaculture disease crisis
Prof. Grant D. Stentiford, Pathology and Microbial Systematics Theme, Cefas
@grantstent
Source: Worldfish.org
Now: 50%
1975: <10%
2050: >65%+

Disease is the #1 issue in limiting yield, reducing profit and preventing investment
Emergence rate is high Deficit in trained professionals/AH investment Dispersed industry. 90% in Asia
Extensive losses Infamous pathogens Pandemic spread
@grantstent

Decentralise diagnostics
Centralise data
Close the gap between farmer, scientist, policy maker
Poverty alleviation. Food. Trade
Simple and quick
Pond-side testing
Remote locations
Smartphone app
Centralised data
@grantstent

Next generation thinking
Reduce impact of disease to promote insurability of industry
Digital pathology for networking
and training
Pathogen genomics for diagnostics/therapy
eDNA/microbiomes for early warning
Host genetics for resilience
@grantstent

When you have 5 mins….
@grantstent

Molecular characterisation of colistin resistance in bacteria isolated from food animals.
Dr. Muna Anjum
Molecular Lead: Antimicrobial Resistance and Enteric Pathogens,Department of Bacteriology

The colistin outbreak…
• Colistin is a last resort antibiotic with limited usage in humans due to its toxicity.
• It is authorised for usage in animals for treatment of gastrointestinal infections, and was first authorised in the UK in 2004 for veterinary medicine.
• Until recently all reported polymyxin resistance mechanisms have been chromosomally mediated, involving for example SNPs in genes encoding two component regulatory systems (e.g. pmrAB, phoPQ), resulting in modification of lipid A.
• Liu et al 2015 reported the first case of a plasmid-mediated colistinresistance mechanism, designated mcr-1, in E. coli and Klebsiellapneumoniae.
8

Usage levels reported in 2011:

Occurrence of mcr-1 in animals 2015-16 in GB:
• The mcr-1 E. coli was detected on two pig farms in GB through anonymised surveillance of 387 caecal samples collected in 2015 from pigs at slaughter from 313 different herds, thus 0.6% of those pig herds sampled were positive.**
• mcr-1 E. coli was also detected on 2/105 (1.9%) of pig farms from which archived E. coli isolates from veterinary diagnostic investigations in 2015/16 were available.**
*(Anjum et al, 2016, Journal of Antimicrobial Chemotherapy)
*(Duggett et al, 2016, Journal of Antimicrobial Chemotherapy)

11
Characterising of mcr-1 isolates by WGS using SeqFinder(Duggett et al, JAC 2016).
Isolate and origin Inc-types present
Estimated
size of
mcr-1
containing
plasmid
(kb)**
Similarity to
pHNSHP45
(%)
Maximum
similarity to
publicly
available
mcr-1
plasmids
(%)
Colistin
resistance
genes
Other AMR genes Virulence genes
E4 #
O139:K82
2015 Veterinary
submission 4
IncX4, IncIFIB(K),
repB, pO111, IncI1,
IncX1,
IncFIA(HI1),
IncA/C2
78.5 16 89
pmrA*,
pmrB*,
phoP*,
phoQ*, etk*
and mcr-1
aadA2, aac3-Iva,
aph4-Ia, aph3-Ib,
aph6-Id, blaTEM-1,
blaLAT-1, cml,
dfrA12, sul1, sul2,
tet(A), gyrA*
cfaA-D, ecpA-E, ecpR, fimF-I,
hlyE, iss
Clinical isolate ^
O149:H10
2015 Veterinary
submission
Inc1, IncX4,
IncFII(pCoo),
IncFIB(AP001918),
IncFIC(FII), IncY
32.7 28 99
phoP*,
phoQ*,
pmrA*, etk*
and mcr-1
aadA1b, ant3-Ia,
dfrA1, folP*, sul2
aec15-19, aec22-27, aec29-32,
astA9, cah, eaeH, ecpA-E, ecpR,
ehaB, eltA-B, espL4, espR1, faeC-
E, faeG-J, fimF-G, hlyA-E, ibeB-C,
ItcA, stb1, shf
PO155 ^
-:H56
2015 surveillance
study
Incl1, Col8282,
pO111, IncX1 91.2 19 90 mcr-1
aac3-IVa, aadA2,
ant3-Ia, aph3-Ib,
aph4-Ia, aph6-Id,
blaTEM-1, cml,
dfrA12, inuF, sul2,
tetA
aec19, aec32, astA ecpA-B, ecpD-
E, ecpR, espL1, espL4, espX5,
fimB-C, fimF-G, fimI, hlyE, iss,
iucA, sitA-C
PO169 ^
-:H2
2015 surveillance
IncX1, IncI2,
IncFII(pCoo),
IncB/O/K/Z
59.2 90 97
acrR*,
phoP* and
mcr-1
blaTEM-1, gyrA*,
qnrS1, tetA
aec31-32, ecpA-E, ecpR, espR1,
fimA-C, fimE-I, hlyE, ibeC, iss,
mchF, tia
12
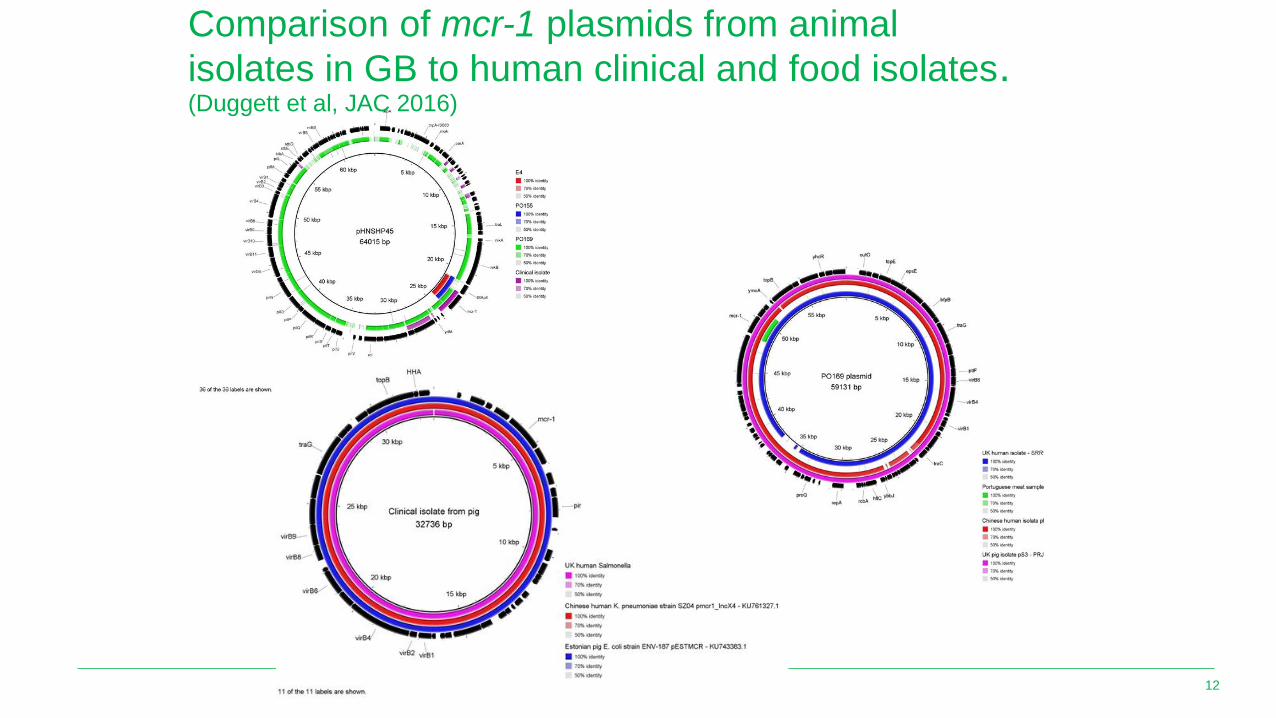
Comparison of mcr-1 plasmids from animal
isolates in GB to human clinical and food isolates.(Duggett et al, JAC 2016)

Conclusion
• First reporting of mcr-1 in enteric bacteria from animals in GB, indicating surveillance was a good mechanism for detection.
• The isolates are MDR but mcr-1 harbours no other AMR gene, so spread of mcr-1 plasmid is not leading to MDR.
• Scanning of archived isolates indicated prevalence of mcr-1 was low (0.6%) in healthy pigs in GB.
• Plasmid comparison showed GB mcr-1 plasmids are quite diverse with high similarity to mcr-1 plasmid in isolates from human, food, animals from various countries, indicating that this is a global epidemic with a worldwide spread.
13

Acknowledgement
APHA colleagues:
Nick Duggett Camilla Brena
Manal AbuOun Richard Ellis
Miranda Kirchner Francesca Martelli
Ellie Saywers Javier Nunez-Garcia
Luke Randall Robert Horton
Sarah Evans Chris Teale
Fabrizio Lemma Jon Rogers
Richard Smith
Rob Davies
Susanna Williamson
14

















