Multilayer structures are known to produce vivid ir-idescent coloration in certain butterflies.1–3 The na-ture of these structures shows several variations ontwo central design themes.4 The first, termed class
or Morpho type,2 comprises layering within discreteridged structures on the surface of scales that coverthe wing. The second, referred to as class II or Ura-nia type,2 comprises continuous multilayering withinthe body of the iridescent scales.
One particular variation in class II multilayer de-sign is brought about by modulations in the profile ofthe multilayering. These modulations, in orthogo-nal directions across the scale surface, have the effectof imposing concave structures into the scale. Theseconcavities are characteristic of the iridescent scalesof many gloss-Papilio butterflies and, in certain spe-cies, produce specific optical effects. Recent re-search5 shows that the design of the multilayering in
apilio palinurus iridescent scales leads to bicolorroduction ~with subsequent color stimulus synthe-is6 for human vision!, retroreflection, and strong
monocolor polarization conversion.In this paper we present detailed evidence to ac-
P. Vukusic [email protected]! and R. Sambles are with Thinilm Photonics, School of Physics, Exeter University, Exeter EX4QL, UK. C. Lawrence is with the Mechanical Sciences Sector,efence Evaluation and Research Agency, Farnborough, GU14LX, UK. G. Wakely is with the School of Biological Sciences,xeter University, Exeter EX4 4QL, UK.Received 5 July 2000; revised manuscript received 27 November
1116 APPLIED OPTICS y Vol. 40, No. 7 y 1 March 2001
count for the mechanism of coloration of P. palinurusand P. ulysses butterflies. Both species exhibit theoncavity-variation of class II iridescent scales. Ac-ordingly, despite their different coloration, oneight expect them to employ the same mechanism
or the production of analogous optical effects. Wehow here that this is not the case and that concavityepth and profile are the key factors responsible forhis difference.
2. Experimental Procedures
Scanning and transmission electron microscopy areimportant techniques for analysis of scale surfaces,cross sections, and concavity profiles. For scanningelectron microscopy a Hitachi Model S-3200N elec-tron microscope was used, with samples first sput-tered with 4 nm of gold. Transmission electronmicroscope ~TEM! images were taken after fixing ofamples in 3% glutaraldehyde at 21 °C for 2 h fol-owed by rinsing in sodium cacodylate buffer. Theyere then fixed in 1% osmic acid in buffer for 1 h
ollowed by block staining in 2% aqueous uranyl ac-tate for 1 h, dehydration through an acetone seriesending with 100% acetone!, and embedding in Spurresin. Postmicrotomed sample sections weretained with lead citrate and examined with a JEOLodel 100S TEM.Reflection spectra from wing samples were col-
ected with a Perkin-Elmer Model Lambda 900 UVyisyNIR spectrometer. Optical images of iridescentcales were taken through a Zeiss Jenalab polarizingicroscope with a JVC Model TK-1280E color video
amera.
acadim
sli
l
itc
tstsobcfdpaslw
bWdcbiatfl
FrlStn
ptoismeospf
tasifl
3. Results and Analysis
To the human observer, P. palinurus, and P. ulyssesre distinct by their bright green and bright blueoloration, respectively ~Fig. 1!. The former exhibitsband of green iridescence across both fore and hindorsal wings and the latter, bright blue across most ofts fore and hind dorsal wings. Scanning electron
icroscope ~SEM! images of the superficial layer ofscales within these colored regions show their sur-faces to consist of a fairly regular array of concavities
(a)
(b)
Fig. 1. Full-color image of ~a! P. palinurus and ~b! P. ulyssesbutterflies showing their iridescent green and iridescent blue col-oration @scale bars: ~a! 1.5 cm and ~b! 1.5 cm#.
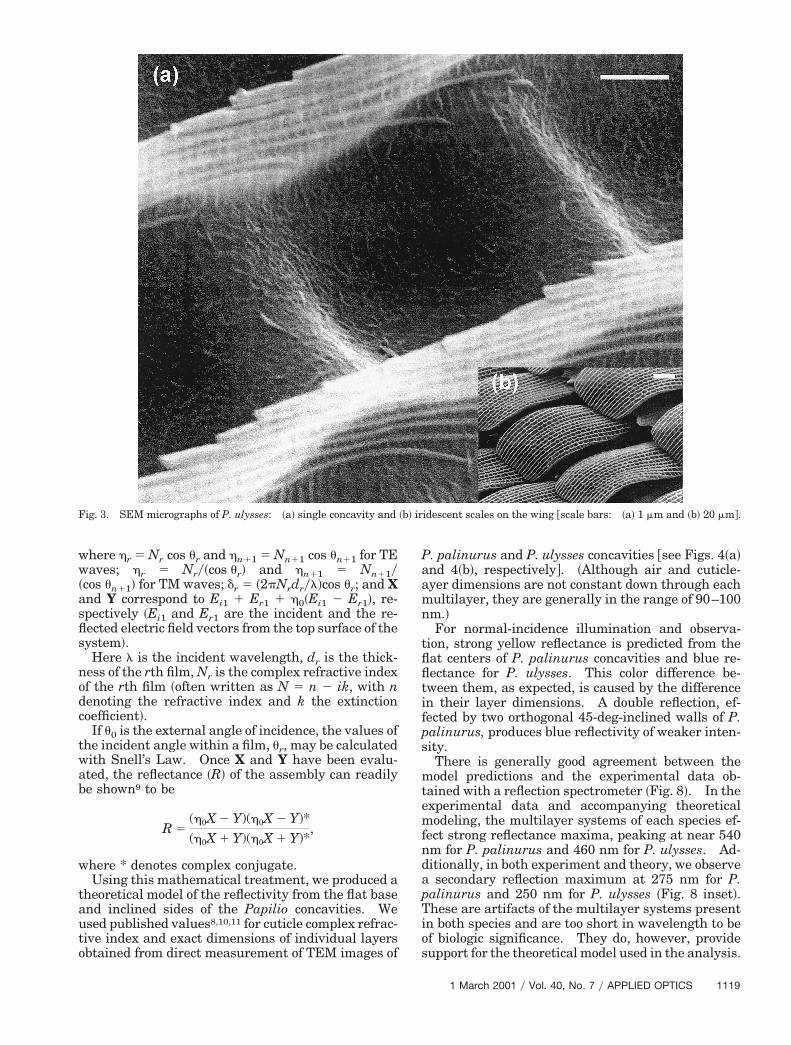
~Figs. 2 and 3!. Ridging, a characteristic of the ma-jority of lepidopteran wing scales, runs the full lengthof each scale with periodicity 4–6 mm and abruptlyeparates these depressions in one direction. Paral-el to the ridging, the transition between concavitiess less abrupt.
TEM images of the samples in cross section high-ight the unique form of gloss-Papilio multilayering
~Figs. 4 and 5!. Sections taken through the concav-ties, perpendicular and parallel to the ridging, showhe modulation in multilayering that causes the con-avity profile.
Close inspection highlights three important fea-ures. First, in the scales of both species, the dimen-ions of each layer in the direction perpendicular tohe local layer surface remain approximately con-tant regardless of position around a concavity. Sec-nd, the multilayer dimensions are smaller for thelue-colored P. ulysses than for the apparently green-olored P. palinurus. Finally, there is a distinct dif-erence between the scales of the two species in theepths and the profiles of their concavities. In P.alinurus, the concavities are deeper and their sidesre steeply inclined with respect to the plane to thecale. For P. ulysses, the scale concavities are shal-ower and their profiles effect much less-inclined sidealls.The optical effect of the different concavity profiles
ecomes evident in optical micrographs ~Fig. 6!.hen illuminated and observed at near-normal inci-
ence, the flat central regions in and between theoncavities strongly reflect yellow in P. palinurus andlue in P. ulysses. Furthermore, in P. palinurus, thenclined sides of each concavity appear as blue annuliround the yellow concavity centers. The sides ofhe concavities in P. ulysses exhibit no analogous re-ection.It is interesting at this stage to ask two questions.
irst, for P. palinurus, why is normally incident lighteflected back along the incident direction, as blueight, by the 45-deg-inclined sides of the concavities?econd, why is an equivalent effect not observed withhe inclined sides of P. ulysses concavities even atear-UV wavelengths?The key to answering these questions lies in ap-
reciation of the contrast in concavity profiles be-ween the two species. Whereas a substantial partf P. palinurus concavity sides are inclined at approx-mately 45 deg relative to the plane of the scale, theides of P. ulysses concavities have inclinations of notore than 30 deg. Consequently, opposite sides of
ach P. palinurus concavity are perpendicular to eachther, whereas in P. ulysses they are not. Notwith-tanding actual multilayer dimensions, it is this thatroduces the fundamental differences in optical ef-ects between the two species.
Clearly, for P. palinurus, single inclined sides ofhe concavities of the scales cannot reflect light backlong the incident direction. In reality, each singleide combines with the surface that is orthogonal totself on the opposite side of the concavity. Retrore-ection is effected in this way; i.e., normally incident
1 March 2001 y Vol. 40, No. 7 y APPLIED OPTICS 1117
pww
mf
1
blue light, reflected from one 45-deg-inclined surface,is directed across the concavity to the opposite orthog-onal surface from where it returns parallel to theoriginal incident direction. These pairs of inclinedsurfaces comprise near-identical multilayering andare both inclined at approximately 45 deg to the di-rection of normally incident light on the scale surface.Accordingly, their spectral reflectivity characteristicsare closely matched. For P. ulysses, in which thissurface orthogonality does not exist, such retroreflec-tion is not possible.
Polarization conversion of blue light, through thisdouble reflection, confirms this mechanism in P.
alinurus. Upon crossing an input linear polarizerith an exit analyzer ~while the sample is viewedith normally incident light!, all yellow reflected
light ~reflected directly from the bottom of the con-cavities! is extinguished while a substantial portionof blue reflected light remains observable @Fig. 7~a!#.This necessarily implies that only the blue reflectedlight has undergone polarization conversion.
Fig. 2. SEM micrographs of P. palinurus: ~a! whole iridescentconcavity @scale bars: ~a! 10 mm, ~b! 5 mm, and ~c! 1 mm#.
118 APPLIED OPTICS y Vol. 40, No. 7 y 1 March 2001
Such retroreflected polarization conversion is pre-dicted from orthogonal surfaces only when the polar-ization vector of the incident light is at 45 deg to theplane of incidence. It does not occur when the inci-dent polarization is perpendicular or parallel to theplane of incidence. Rotation of the wing scale,through 45 deg in the plane of the wing, changes theregions of the inclined sides of each concavity thatexhibit this strong polarization conversion in the ex-pected manner @Fig. 7~b!#.
Optical theory can be used to predict reflectivityfrom a nonuniform system of flat multilayers.7–9
The characteristic matrix of an assembly of n filmsay be calculated from the product of the matrices
or the individual films taken in the correct order,
SXYD 5 5)r51
n 3 cos dr
i sin dr
hr
ihr sin dr cos dr46 S 1
hn11D,
on the wing, ~b! a small region of iridescent scale, and ~c! single
scale
t
ab
amn
tflfltifps
mtemfndapTios
where hr 5 Nr cos ur and hn11 5 Nn11 cos un11 for TEwaves; hr 5 Nry~cos ur! and hn11 5 Nn11y~cos un11! for TM waves; dr 5 ~2pNrdryl!cos ur; and Xand Y correspond to Ei1 1 Er1 1 h0~Ei1 2 Er1!, re-spectively ~Ei1 and Er1 are the incident and the re-flected electric field vectors from the top surface of thesystem!.
Here l is the incident wavelength, dr is the thick-ness of the rth film, Nr is the complex refractive indexof the rth film ~often written as N 5 n 2 ik, with ndenoting the refractive index and k the extinctioncoefficient!.
If u0 is the external angle of incidence, the values ofhe incident angle within a film, ur, may be calculated
with Snell’s Law. Once X and Y have been evalu-ted, the reflectance ~R! of the assembly can readilye shown9 to be
R 5(h0X 2 Y)(h0X 2 Y)*(h0X 1 Y)(h0X 1 Y)*
,
where * denotes complex conjugate.Using this mathematical treatment, we produced a
theoretical model of the reflectivity from the flat baseand inclined sides of the Papilio concavities. Weused published values8,10,11 for cuticle complex refrac-tive index and exact dimensions of individual layersobtained from direct measurement of TEM images of
Fig. 3. SEM micrographs of P. ulysses: ~a! single concavity and
P. palinurus and P. ulysses concavities @see Figs. 4~a!and 4~b!, respectively#. ~Although air and cuticle-yer dimensions are not constant down through eachultilayer, they are generally in the range of 90–100m.!For normal-incidence illumination and observa-
ion, strong yellow reflectance is predicted from theat centers of P. palinurus concavities and blue re-ectance for P. ulysses. This color difference be-ween them, as expected, is caused by the differencen their layer dimensions. A double reflection, ef-ected by two orthogonal 45-deg-inclined walls of P.alinurus, produces blue reflectivity of weaker inten-ity.There is generally good agreement between theodel predictions and the experimental data ob-
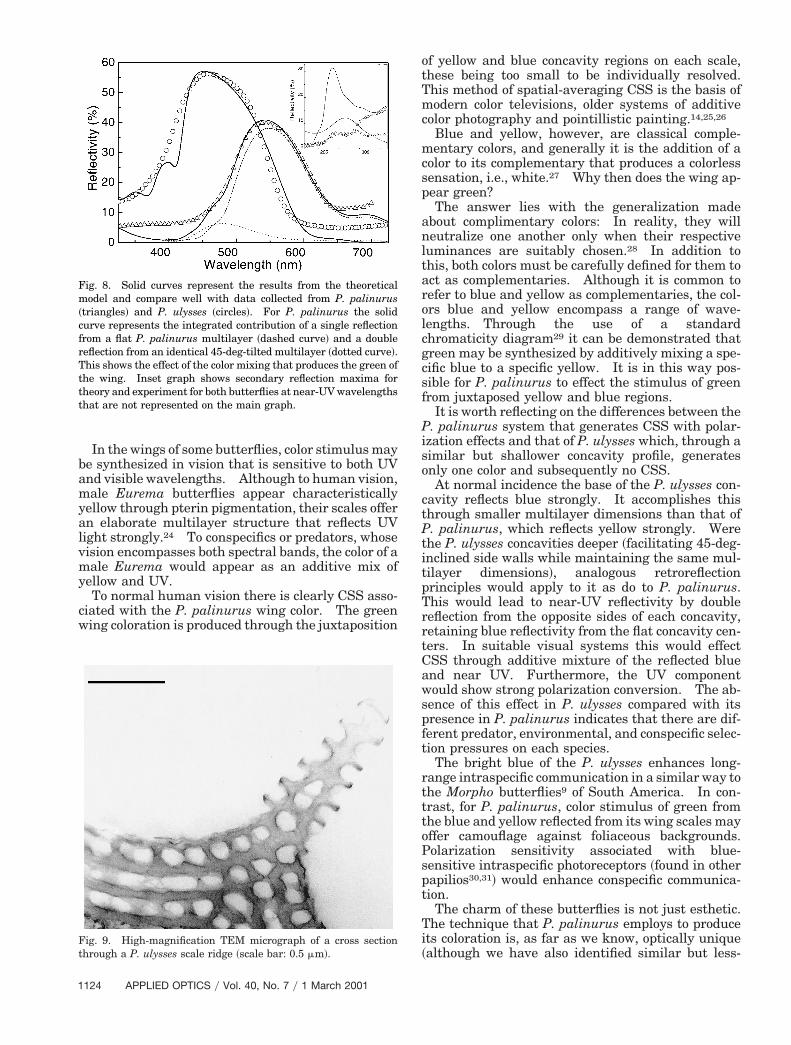
ained with a reflection spectrometer ~Fig. 8!. In thexperimental data and accompanying theoreticalodeling, the multilayer systems of each species ef-
ect strong reflectance maxima, peaking at near 540m for P. palinurus and 460 nm for P. ulysses. Ad-itionally, in both experiment and theory, we observesecondary reflection maximum at 275 nm for P.
alinurus and 250 nm for P. ulysses ~Fig. 8 inset!.hese are artifacts of the multilayer systems present
n both species and are too short in wavelength to bef biologic significance. They do, however, provideupport for the theoretical model used in the analysis.
idescent scales on the wing @scale bars: ~a! 1 mm and ~b! 20 mm#.
~b! ir
1 March 2001 y Vol. 40, No. 7 y APPLIED OPTICS 1119
Hcmiirittvou
dm
trgmtrydta
1
However, it is worth making several additionalpoints. The discrepancies in reflectivity levels be-tween model and data, either side of the main peaks,are attributed to scatter from scale ridges and under-lying noniridescent ground scales that cannot be ac-counted for in the modeling. Additionally, furtherdiscrepancy is brought about by uncertainty in dis-persion of cuticle complex refractive index across theexperimental wavelength range.
The TEM images of both species show evidence ofstrong differential staining ~Figs. 4 and 5!. This haspreviously been observed in TEM’s of iridescent but-terfly scales12 but with less-extreme differential con-trast. It is not considered to be an artifact of thepreparation procedure: Therefore we should con-sider its significance. The purpose of block-and-section staining of the sample during the preparationprocess is well known.13 Block staining with uranylacetate renders fine structure visible by causing cer-tain components to attract heavy metals ions andconsequently scatter electrons differentially. Fur-ther staining with lead citrate, once the sample is insectional form, enhances contrast between fine struc-ture in the image. The dark contrast of the lowercuticle layers is indicative of the presence of differentmolecular species compared with the light-contrastedupper layers. We conjecture that this darker con-trast is evidence of higher concentrations of melanin
Fig. 4. TEM micrographs showing cross sections through iridesshowing the arrangement of several of the concavities across a sin
120 APPLIED OPTICS y Vol. 40, No. 7 y 1 March 2001
in the lower layers ~melanin is the optically absorbingspecies present in the majority of butterfly scales14!.
igh levels of melanin in the lower layers would in-rease optical absorption toward the bottom of theultilayer system while permitting strong reflectiv-
ty without absorption in the upper layers. Thisdea has been included in the theoretical modelingepresented in Fig. 8. Within the model, the imag-nary component of refractive index used to representhe optical absorption in the upper layers is less thanhat in the lower layers; this difference and theiralues are in line with measurements of k coefficientsf single iridescent scales in other species10 ~i.e., forpper layers, k 5 0.06 and for lower layers, k 5 0.25,
both at l 5 550 nm!. Poorer fits between theory andata are produced if this differential absorbanceodel is not used.Both species also exhibit some even finer surface
exture across their iridescent scales. In P. palinu-us this texture is in the form of a small zero-orderrating of approximately 170-nm pitch and approxi-ately 50-nm depth. It follows the surface profile in
he area between the ridges running at 45 deg to theidges. Similar surface texture is present in P. ul-sses but in a much less-ordered arrangement. Theimensions of both these textures are believed to beoo small to contribute any significant optical effectst visible and near UV. However, it is possible that
scales of P. palinurus ~perpendicular to ridges, with inset imagecale!. @scale bars: 1 mm ~inset 3 mm!#.
centgle s
q
this provides some level of impedance matching, re-ducing broadband reflectivity from the topmost sur-face of the scale multilayer system.
It is generally believed that iridescent species exhib-iting the continuous body-lamellae of class II scales donot posses ridge-lamellae associated with class Iscales. Such lamellae, however, are evident in theridges of P. ulysses ~Fig. 9!. One would assume thatsuch complex structure would serve an optical pur-pose. Reflectivity calculations using layer dimen-sions suggest that the layered structure of the ridgingis designed to reflect a wavelength band centered atapproximately 380 nm. However, after we accountfor the limited number of layers and ridge occupancy,the maximum absolute level at which this band isreflected is less than 10%. This appears to makeridge-lamellae reflectivity insignificant in comparisonwith the reflectivity from the body-lamellae, even at380 nm. The function, therefore, of the lamellar na-ture of the ridges on P. ulysses remains a puzzle.
When P. palinurus is illuminated with diffuse sun-light, green coloration can be observed in a limited
Fig. 5. TEM micrographs showing cross sections through iridesceshowing cross section taken parallel to ridges and top inset image1 mm ~top inset 3 mm and bottom inset 1 mm!#.
region above its wings. Outside this perspective thewing coloration changes predictably; i.e., shorterwavelengths are reflected more strongly as observa-tion angle approaches grazing incidence. It is forthese conditions that the reflectivity characteristicsof each orthogonal surface become mismatched, caus-ing the retroreflective effect of each concavity to func-tion less effectively. However, observation atincreasingly nonnormal incidence is effected throughlarge-angle reflections from the center and from sin-gle sides of each concavity.
P. ulysses visibility is more straightforward. Itappears blue from directly above the wing; this isentirely due to single reflections from either the bot-tom or shallow sides of the concavities. Towardgrazing incidence the wing color approaches deep vi-olet, owing to larger angle reflections from the bottomand single sides of the shallow concavities.
4. Discussion
Color stimulus synthesis6 ~CSS! is a phenomenon fre-uently found in nature, especially appearing in the
les of P. ulysses ~perpendicular to ridges, with bottom inset imageing arrangement of several neighboring concavities!. @scale bars:
nt scashow
1 March 2001 y Vol. 40, No. 7 y APPLIED OPTICS 1121
1
production of green colors. It is an effect whereby anadditive mixture of two or more colors synthesizes thestimulus of a different color in the visual system of anobserver.
122 APPLIED OPTICS y Vol. 40, No. 7 y 1 March 2001
In the animal kingdom, CSS of green is usuallyachieved through an additive mixture of structurallyeffected blue and pigmentary yellow15 ~or occasion-ally, both pigmentary blue and yellow16!. The green
Fig. 6. Optical microscopy images of a region of ~a! P. palinurus iridescent scale and ~b! P. ulysses iridescent scale @scale bars: ~a! and~b!, 10 mm#.
17–20 18,19
of many species of birds’ feathers and the integ-ument of some amphibians15,19,21 and reptiles19 re-sults from color mixing of yellow pigmentation withblue structural scattering effects. Purple, another
synthesized color found in some feathers, butter-flies,22 and in port-wine birthmarks,23 is produced bythe combination of red pigmentation with blue fromstructural scattering.
Fig. 7. Optical microscopy images of a region of a P. palinurus iridescent scale: ~a! using input and output polarizers crossed with respectto each other and ~b! input and output polarizers crossed with respect to each other but the sample rotated azimuthally through 45 degfrom position in ~a! @scale bars: ~a! and ~b!, 10 mm#. C1 and C2 represent concavities 1 and 2 to clarify the sample rotation.
1 March 2001 y Vol. 40, No. 7 y APPLIED OPTICS 1123
yal
so
ctPtitpTrrtCawspft
rt
toPsp
rTttt
1
In the wings of some butterflies, color stimulus maybe synthesized in vision that is sensitive to both UVand visible wavelengths. Although to human vision,male Eurema butterflies appear characteristicallyellow through pterin pigmentation, their scales offern elaborate multilayer structure that reflects UVight strongly.24 To conspecifics or predators, whose
vision encompasses both spectral bands, the color of amale Eurema would appear as an additive mix ofyellow and UV.
To normal human vision there is clearly CSS asso-ciated with the P. palinurus wing color. The greenwing coloration is produced through the juxtaposition
Fig. 8. Solid curves represent the results from the theoreticalmodel and compare well with data collected from P. palinurus~triangles! and P. ulysses ~circles!. For P. palinurus the solidcurve represents the integrated contribution of a single reflectionfrom a flat P. palinurus multilayer ~dashed curve! and a doubleeflection from an identical 45-deg-tilted multilayer ~dotted curve!.his shows the effect of the color mixing that produces the green ofhe wing. Inset graph shows secondary reflection maxima forheory and experiment for both butterflies at near-UV wavelengthshat are not represented on the main graph.
Fig. 9. High-magnification TEM micrograph of a cross sectionthrough a P. ulysses scale ridge ~scale bar: 0.5 mm!.
124 APPLIED OPTICS y Vol. 40, No. 7 y 1 March 2001
of yellow and blue concavity regions on each scale,these being too small to be individually resolved.This method of spatial-averaging CSS is the basis ofmodern color televisions, older systems of additivecolor photography and pointillistic painting.14,25,26
Blue and yellow, however, are classical comple-mentary colors, and generally it is the addition of acolor to its complementary that produces a colorlesssensation, i.e., white.27 Why then does the wing ap-pear green?
The answer lies with the generalization madeabout complimentary colors: In reality, they willneutralize one another only when their respectiveluminances are suitably chosen.28 In addition tothis, both colors must be carefully defined for them toact as complementaries. Although it is common torefer to blue and yellow as complementaries, the col-ors blue and yellow encompass a range of wave-lengths. Through the use of a standardchromaticity diagram29 it can be demonstrated thatgreen may be synthesized by additively mixing a spe-cific blue to a specific yellow. It is in this way pos-sible for P. palinurus to effect the stimulus of greenfrom juxtaposed yellow and blue regions.
It is worth reflecting on the differences between theP. palinurus system that generates CSS with polar-ization effects and that of P. ulysses which, through aimilar but shallower concavity profile, generatesnly one color and subsequently no CSS.At normal incidence the base of the P. ulysses con-
avity reflects blue strongly. It accomplishes thishrough smaller multilayer dimensions than that of. palinurus, which reflects yellow strongly. Werehe P. ulysses concavities deeper ~facilitating 45-deg-nclined side walls while maintaining the same mul-ilayer dimensions!, analogous retroreflectionrinciples would apply to it as do to P. palinurus.his would lead to near-UV reflectivity by doubleeflection from the opposite sides of each concavity,etaining blue reflectivity from the flat concavity cen-ers. In suitable visual systems this would effectSS through additive mixture of the reflected bluend near UV. Furthermore, the UV componentould show strong polarization conversion. The ab-
ence of this effect in P. ulysses compared with itsresence in P. palinurus indicates that there are dif-erent predator, environmental, and conspecific selec-ion pressures on each species.
The bright blue of the P. ulysses enhances long-ange intraspecific communication in a similar way tohe Morpho butterflies9 of South America. In con-
trast, for P. palinurus, color stimulus of green fromhe blue and yellow reflected from its wing scales mayffer camouflage against foliaceous backgrounds.olarization sensitivity associated with blue-ensitive intraspecific photoreceptors ~found in otherapilios30,31! would enhance conspecific communica-
tion.The charm of these butterflies is not just esthetic.
The technique that P. palinurus employs to produceits coloration is, as far as we know, optically unique~although we have also identified similar but less-
ldqtcotttc
es0s
11. M. Land, School of Biological Sciences, Sussex University,
pronounced effects in the related species of P. crino,P. buddha, and P. blumei!. Through simple modu-ation in an otherwise uniform multilayer system, itisplays high bicolor reflectivity that synthesizes auite different color stimulus in certain visual sys-ems. The structure shows strong local polarizationonversion of one of the colors through a mechanismf orthogonal-surface retroreflection.5 P. ulysses,hrough a shallower concavity profile, does not createhis effect. It exhibits strong blue iridescence by vir-ue of appropriate layer dimensions in a 10–12uticle-layer system.
This research was funded by the University of Ex-ter, the Biotechnology and Biological Sciences Re-earch Council ~BBSRC!, and the Technology Group8 of the Ministry of Defence ~MoD! Corporate Re-earch Program.
References1. H. Ghiradella, “Light and color on the wing: structural col-
ors in butterflies and moths,” Appl. Opt. 30, 3492–3500~1991!.
2. W. Lippert and K. Gentil, “Uber lamellare Feinstrukturen beiden Schillerschuppen der Schmetterlinge vom Urania- undMorpho-typ,” Z. Morphol. Oekol. Tiere 48, 115–122 ~1959!.
3. T. F. Anderson and A. G. Richards, “An electron microscopestudy of some structural colors of insects,” J. Appl. Phys. 13,748–758 ~1942!.
4. P. Vukusic, J. R. Sambles, and H. Ghiradella, “Optical classi-fication of microstructure in butterfly wing scales,” PhotonicsSci. News 6, 61–66 ~2000!.
5. P. Vukusic, J. R. Sambles, and C. R. Lawrence, “Structuralcolour: colour mixing in wing scales of a butterfly,” Nature404, 457 ~2000!.
6. R. W. Burnham, R. M. Hanes, and C. J. Bartleson, Color~Wiley, New York, 1963!.
7. A. Vasicek, Optics of Thins Films ~North-Holland, Amsterdam,1960!.
8. M. F. Land, “The physics and biology of animal reflectors,”Prog. Biophys. Mol. Biol. 24, 75–106 ~1972!.
9. H. A. Macleod, Thin-Film Optical Filters ~Adam Hilger, Lon-don, 1969!.
10. P. Vukusic, J. R. Sambles, C. R. Lawrence, and R. J. Wootton,“Quantified interference and diffraction in single Morpho but-terfly scales,” Proc. R. Soc. London Ser. B 266, 1403–1411~1999!.
Brighton BN1 9QG, UK ~personal communication, 1999!.12. J. Huxley, “The basis of structural colour variation in two
species of Papilio,” J. Entomol. Ser. A 50, 9–22 ~1975!.13. P. R. Lewis and D. P. Knight, Staining Methods for Sectioned
Material ~North-Holland, Oxford, 1977!.14. H. F. Nijhout, The Development and Evolution of Butterfly
15. D. L. Fox, Animal Biochromes and Structural Colours ~Uni-versity of California Press, Berkeley, Calif., 1976!.
16. R. I. Vane-Wright, “The coloration, identification and phylog-eny of Nessaea butterflies ~Lepidoptera: Nymphalidae!,” Bull.Br. Mus. ~Nat. Hist.! Entomol. 38, 2, 29–56 ~1979!.
17. R. Lewis, “The optics of feather colour,” Biophotonics Int.~April, 1999!, pp. 38–39.
18. C. W. Mason, “Structural colours in feathers I,” J. Phys. Chem.27, 205–251 ~1923!.
19. H. M. Fox and G. Vevers, The Nature of Animal Colours ~Sidg-wick and Jackson, London, 1960!.
20. F. Frank, “Die farbung der vogelfeder durch pigment undstruktur,” J. Orn. Lpz. 3, 426–523 ~1939!.
21. C. E. Geldern von, “Color changes and structure of the skin ofAnolis carolinensis,” Proc. Calif. Acad. Sci. 10, 77–117 ~1921!.
22. R. I. Vane-Wright, Keeper of Entomology, Natural History Mu-seum, London SW7 5BD, UK ~personal communication, 1999!.
23. J. Verne and F. Leyani, “Les dyschromies,” Traite de derma-tologie ~Paris! 2, 745–811 ~1938!.
24. H. Ghiradella, D. Aneshansley, T. Eisner, R. E. Silbergleid,and H. E. Hinton, “Ultra-violet reflection of a male butterfly:interference colour caused by thin layer elaboration of wingscales,” Science 178, 1214–1217 ~1972!.
25. R. M. Evans, “Visual processes and color photography,” J. Opt.Soc. Am. 33, 579–614 ~1943!.
26. D. L. MacAdam, “Perceptions of colour in projected and tele-vised pictures,” J. Soc. Motion Pict. Tel. Eng. 65, 455–4669~1956!.
27. F. W. Billmeyer and M. Saltzman, Principles of Color Tech-nology ~Wiley, New York, 1981!.
28. M. I. Sobel, Light ~U. of Chicago Press, Chicago, 1987!.29. G. A. Agoston, Color Theory and its Applications in Art and
Design ~Springer-Verlag, New-York, 1987!.30. A. Kelber, “Why ‘false’ colours are seen by butterflies,” Nature
402, 251 ~1999!.31. K. Bandai, K. Arikawa, and E. Eguchi, “Localisation of spec-
tral receptors in the ommatidium of butterfly compound eyedetermined by polarisation sensitivity,” J. Comp. Physiol. A171, 289–297 ~1992!.
1 March 2001 y Vol. 40, No. 7 y APPLIED OPTICS 1125