Segmentation and specification of the Drosophila mesoderm Natalia Azpiazu, 1,3 Peter A. Lawrence, 2 Jean-Paul Vincent, 2 and Manfred Frasch 1'4 1Brookdale Center for Molecular Biology, Mount Sinai School of Medicine, New York, New York 10029 USA; 2Medical Research Council Laboratory of Molecular Biology, Cambridge CB2 2QH, UK Patterning of the developing mesoderm establishes primordia of the visceral, somatic, and cardiac tissues at defined anteroposterior and dorsoventral positions in each segment. Here we examine the mechanisms that locate and determine these primordia. We focus on the regulation of two mesodermal genes: bagpipe (hap), which defines the anlagen of the visceral musculature of the midgut, and serpent (srp), which marks the anlagen of the fat body. These two genes are activated in specific groups of mesodermal cells in the anterior portions of each parasegment. Other genes mark the anlagen of the cardiac and somatic mesoderm and these are expressed mainly in cells derived from posterior portions of each parasegment. Thus the parasegments appear to be subdivided, at least with respect to these genes, a subdivision that depends on pair-rule genes such as even-skipped (eve). We show with genetic mosaics that eve acts autonomously within the mesoderm. We also show that hedgehog (hh) and wingless (wg) mediate pair-rule gene functions in the mesoderm, probably partly by acting within the mesoderm and partly by inductive signaling from the ectoderm, hh is required for the normal activation of hap and srp in anterior portions of each parasegment, whereas wg is required to suppress bap and srp expression in posterior portions. Hence, hh and wg play opposing roles in mesoderm segmentation. [Key Words: Mesoderm; segmentation; visceral musculature; fat body; heart; even-skipped; wingless; hedgehog] Received September 4, 1996; revised version accepted November 1, 1996. How does the Drosophila mesoderm become subdivided into different anlagen? The mesoderm is partitioned along the dorsoventral axis: dorsally, cells found the pri- mordia of the cardiac and visceral mesoderm and this subdivision is triggered by inductive inputs from the ec- toderm. Dpp is a TGF~ protein that is secreted from dorsal ectodermal cells, and has been identified as the inductive signal (Staehling-Hampton et al. 1994; Frasch 1995). Dpp signaling is required for the formation of the anlagen of the heart and the midgut visceral mesoderm, and the dorsoventral extent of the visceral mesoderm is directly determined by the dorsoventral limits of dpp expression in the ectoderm (Frasch 1995; Maggert et al. 1995). An early response to the Dpp signal in mesoder- mal cells is the spatial restriction of expression of the homeo-box gene tinman (tin) to dorsal portions of the mesoderm, tin provides the dorsal mesoderm with the competence to form midgut visceral mesoderm, dorsal muscles, and the heart (Azpiazu and Frasch 1993; Bod- mer 1993). In combination with additional regulators that may include Dpp, tin activates the expression of a second homeo-box gene, bagpipe (bap), in dorsal meso- 3Present address: Centro de Biologia Molecular"Severo Ochoa," Facul- tad de Ciencias, Universita Autonoma de Madrid,Madrid28034, Spain. 4Corresponding author. dermal areas (Azpiazu and Frasch 1993; Staehling-Hamp- ton et al. 1994). bap is involved in the determination of the anlagen of the midgut visceral mesoderm. Analogous events could activate tin target genes that would specify the anlagen of the heart and dorsal body wall muscula- ture. We now study patterning of the mesoderm along the anteroposterior axis of the embryo. The morphology of the early mesoderm and the spatial domains of expres- sion of homeotic genes indicate that the mesoderm is organized into parasegmental units (for review, see Lawrence 1992). Previous attempts to identify distinct subdivisions within each mesodermal parasegment that might be homologous to the anterior and posterior com- partments of the ectoderm were negative (Lawrence and Johnston 1984). However, more recently, the analysis of expression patterns of genes that control mesoderm de- velopment has suggested that mesodermal parasegments are indeed subdivided into anterior and posterior por- tions with different developmental fates. A clear exam- ple is the expression of the homeo-box gene bap which is restricted to metameric clusters of cells in the dorsal mesoderm (Azpiazu and Frasch 1993). Under the control of bap, these cell clusters develop into midgut visceral mesoderm, whereas cells in segmental portions lacking bap expression form other mesodermal derivatives. The expression of several marker genes in the anlagen of the GENES & DEVELOPMENT10:3183-3194 9 1996 by Cold SpringHarborLaboratoryPress ISSN 0890-9369/96 $5.00 3183 Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.org Downloaded from
Transcript
Segmentation and specification of the Drosophila mesoderm Natal ia Azpiazu , 1,3 Peter A. Lawrence, 2 Jean-Paul Vincent , 2 and Manfred Frasch 1'4
1Brookdale Center for Molecular Biology, Mount Sinai School of Medicine, New York, New York 10029 USA; 2Medical Research Council Laboratory of Molecular Biology, Cambridge CB2 2QH, UK
Patterning of the developing mesoderm establishes primordia of the visceral, somatic, and cardiac tissues at defined anteroposterior and dorsoventral positions in each segment. Here we examine the mechanisms that locate and determine these primordia. We focus on the regulation of two mesodermal genes: bagpipe (hap), which defines the anlagen of the visceral musculature of the midgut, and serpent (srp), which marks the anlagen of the fat body. These two genes are activated in specific groups of mesodermal cells in the anterior portions of each parasegment. Other genes mark the anlagen of the cardiac and somatic mesoderm and these are expressed mainly in cells derived from posterior portions of each parasegment. Thus the parasegments appear to be subdivided, at least with respect to these genes, a subdivision that depends on pair-rule genes such as even-skipped (eve). We show with genetic mosaics that eve acts autonomously within the mesoderm. We also show that hedgehog (hh) and wingless (wg) mediate pair-rule gene functions in the mesoderm, probably partly by acting within the mesoderm and partly by inductive signaling from the ectoderm, hh is required for the normal activation of hap and srp in anterior portions of each parasegment, whereas wg is required to suppress bap and srp expression in posterior portions. Hence, hh and wg play opposing roles in mesoderm segmentation.
Received September 4, 1996; revised version accepted November 1, 1996.
How does the Drosophila mesoderm become subdivided into different anlagen? The mesoderm is partitioned along the dorsoventral axis: dorsally, cells found the pri- mordia of the cardiac and visceral mesoderm and this subdivision is triggered by inductive inputs from the ec- toderm. Dpp is a TGF~ protein that is secreted from dorsal ectodermal cells, and has been identified as the inductive signal (Staehling-Hampton et al. 1994; Frasch 1995). Dpp signaling is required for the formation of the anlagen of the heart and the midgut visceral mesoderm, and the dorsoventral extent of the visceral mesoderm is directly determined by the dorsoventral limits of dpp expression in the ectoderm (Frasch 1995; Maggert et al. 1995). An early response to the Dpp signal in mesoder- mal cells is the spatial restriction of expression of the homeo-box gene tinman (tin) to dorsal portions of the mesoderm, tin provides the dorsal mesoderm with the competence to form midgut visceral mesoderm, dorsal muscles, and the heart (Azpiazu and Frasch 1993; Bod- mer 1993). In combination with additional regulators that may include Dpp, tin activates the expression of a second homeo-box gene, bagpipe (bap), in dorsal meso-
3Present address: Centro de Biologia Molecular "Severo Ochoa," Facul- tad de Ciencias, Universita Autonoma de Madrid, Madrid 28034, Spain. 4Corresponding author.
dermal areas (Azpiazu and Frasch 1993; Staehling-Hamp- ton et al. 1994). bap is involved in the determination of the anlagen of the midgut visceral mesoderm. Analogous events could activate tin target genes that would specify the anlagen of the heart and dorsal body wall muscula- ture.
We now study patterning of the mesoderm along the anteroposterior axis of the embryo. The morphology of the early mesoderm and the spatial domains of expres- sion of homeotic genes indicate that the mesoderm is organized into parasegmental units (for review, see Lawrence 1992). Previous attempts to identify distinct subdivisions within each mesodermal parasegment that might be homologous to the anterior and posterior com- partments of the ectoderm were negative (Lawrence and Johnston 1984). However, more recently, the analysis of expression patterns of genes that control mesoderm de- velopment has suggested that mesodermal parasegments are indeed subdivided into anterior and posterior por- tions with different developmental fates. A clear exam- ple is the expression of the homeo-box gene bap which is restricted to metameric clusters of cells in the dorsal mesoderm (Azpiazu and Frasch 1993). Under the control of bap, these cell clusters develop into midgut visceral mesoderm, whereas cells in segmental portions lacking bap expression form other mesodermal derivatives. The expression of several marker genes in the anlagen of the
GENES & DEVELOPMENT 10:3183-3194 �9 1996 by Cold Spring Harbor Laboratory Press ISSN 0890-9369/96 $5.00 3183
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from
fat body and studies on the regulatory gene twist also indicate that cells at different anteroposterior positions within each mesodermal parasegment give rise to differ- ent mesodermal derivatives (Abel et al. 1993; Hoshizaki et al. 1994; Dunin-Borkowski et al. 1995; Baylies and Bate 1996).
By analogy to dorsoventral patterning, inductive sig- nals from the segmented ectoderm could help segment the mesoderm. Alternatively, the mesoderm could seg- ment itself autonomously. For example, wingless (wg) acts in heart and muscle development and can function either from within the ectoderm or from the mesoderm (Baylies et al. 1995; Lawrence et al. 1995; Wu et al. 1995; Ranganayakulu et al. 1996). However, there must be ad- ditional segmental regulators, because both in the ab- sence of wg and in the presence of uniformly distributed Wg protein the mesoderm is still segmentally patterned.
Our experiments show that the mesoderm forms seg- ments autonomously, although induction from the ecto- derm also contributes. We show that wg and hedgehog (hh), which are candidates for inductive signals from the ectoderm, play opposite and antagonistic roles in the subdivision of mesodermal parasegments, wg is mainly involved in the formation of derivatives from posterior portions of each parasegment, including somatic muscu- lature and the heart, whereas hh functions mainly in the formation of derivatives in anterior parasegmental por- tions, in particular the visceral mesoderm of the midgut musculature and the fat body.
R e s u l t s
Segmental organization of mesodermal primordia
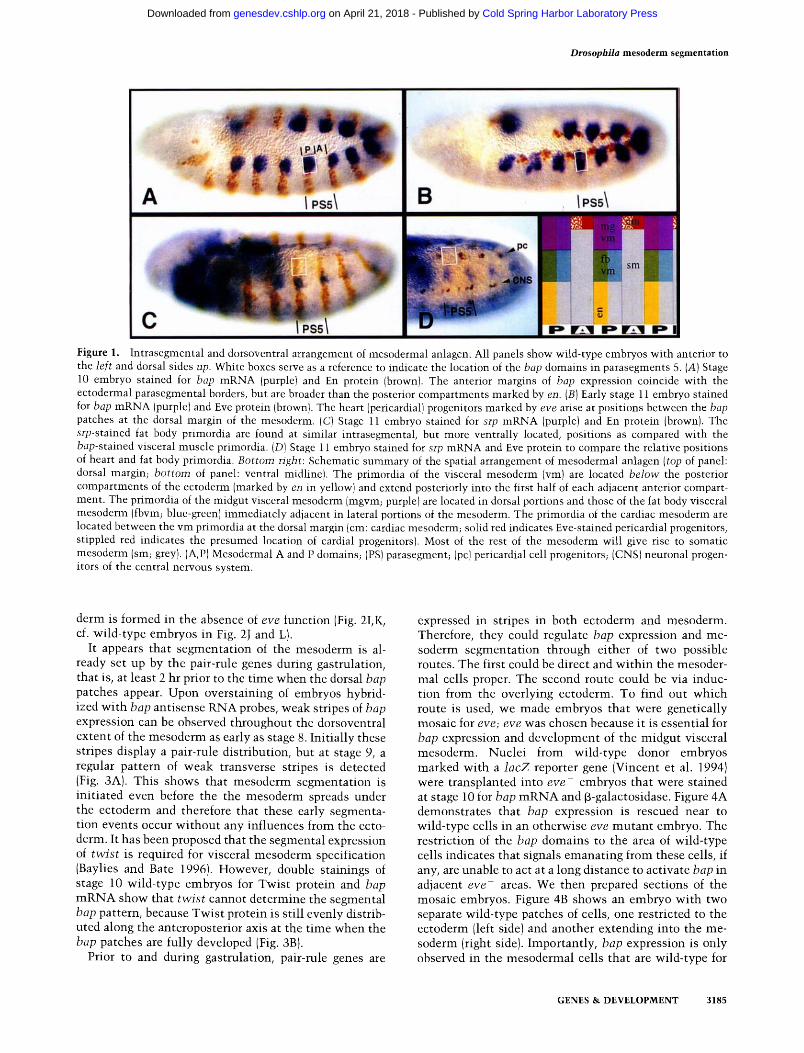
The expression patterns of several genes reveal that the early mesoderm is segmented. Notably, the homeo-box gene bap is expressed in 11 metameric clusters of dorsal mesodermal cells that develop into visceral musculature of the midgut (Azpiazu and Frasch 1993). The anterior borders of each of the bap patches coincide with the parasegmental borders of the ectoderm--thus the pri- mordia of the midgut visceral mesoderm are largely po- sitioned below the posterior compartments of the ecto- derm (Fig. 1 A; Azpiazu and Frasch 1993). The homeo-box gene even-skipped (eve) marks heart progenitors that will form pericardial cells (Frasch et al. 1987). Double stainings for bap and eve demonstrate that these heart progenitors form between the bap patches at the dorsal crest of the mesoderm (Fig. 1B). In addition, the progen- itors of the cardioblasts are formed in close juxtaposi- tion, just anterior to the pericardial progenitors (K. Jagla, pets. comm.). Thus, it appears that the anlagen of the cardiac mesoderm alternate with the anlagen of the mid- gut visceral mesoderm. There are also primordia of the fat body in each segment; these are marked by the ex- pression of the serpent (srp) gene that is required for fat body formation (srp encodes the GATA transcription fac- tor ABF; Abel et al. 1993; Rehorn et al. 1996). Double stainings for srp and engrailed (en; Fig. 1C) or srp and eve
(Fig. 1D) show that these cells are at the same antero- posterior positions and lie just ventrolateral to the pri- mordia of the midgut visceral mesoderm.
In summary, the mesoderm of stage 10-11 embryos appears to be organized into parasegmental repeats that are in exact register with the ectodermal parasegments. Each parasegment is subdivided into two domains along the anteroposterior axis. In keeping with the nomencla- ture used for the ectoderm (Lawrence 1992), we term the mesodermal domains below the anterior compartments "A domains" and those below the posterior compart- ments "P domains." As shown schematically in Figure 1E, the P domains express bap and srp and give rise to visceral mesoderm derivatives that include midgut mus- culature and the fat body. The A domains include the primordia of the cardiac mesoderm and the bulk of the somatic mesoderm and give rise to the heart and most of the body wall muscles.
A mesoderm-autonomous role of pair rule genes in segmentation and specification of the visceral mesoderm
The close alignment between mesodermal and ectoder- mal parasegments suggests that the two germ layers might be segmented by the same genes. Indeed, muta- tions in pair rule genes have similar effects in both the ectoderm and the mesoderm. For example, fushi tarazu (ftz) mutant embryos retain only six out of normally 13 en stripes between parasegments 2 and 14 in wild-type embryos, and the number of bap patches is similarly reduced from normally 11 to 5 (Fig. 2A; note that para- segments 13 and 14 lack bap expression even in wild- type embryos; see Fig. 1A). srp patches are also reduced in ftz mutant embryos (Fig. 2B; wild-type expression only occurs between parasegments 4 and 12; see Fig. 1C). In spite of their aberrant patterns, the normal spatial relationships between the bap or srp domains and the en stripes are maintained in ftz mutants, indicating that ftz mutations cause identical alterations in the mesoderm and ectoderm. Concomitant alterations of bap and en stripes are observed in mutants for all other pair-rule genes tested, including hairy (h; Fig. 2C), runt (run; Fig. 2D), paired (prd; Fig. 2E), odd-paired (opa; Fig. 2F), and odd-skipped (odd; data not shown). Pair-rule mutations that cause mirror-image arrangements in the ectoderm, such as runt (Nhsslein-Volhard and Wieschaus 1980), re- sult in mirror-image arrangements of the bap patches in the mesoderm (Fig. 2D).
The importance of pair-rule genes to segmentation of the mesoderm is particularly obvious in embryos that are mutant for eve. In embryos lacking eve function, the ectoderm is unsegmented and no en stripes are formed in the trunk region (Nhsslein-Volhard et al. 1985; Harding et al. 1986; Macdonald et al. 1986). Similarly, bap and srp patches are completely lost in the trunk region of eve mutant embryos, indicating that the mesoderm is unseg- mented as well (Fig. 2G, H). As a consequence, neither midgut visceral mesoderm nor fat body visceral meso-
3184 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from
Figure 1. Intrasegmental and dorsoventral arrangement of mesodermal anlagen. All panels show wild-type embryos with anterior to the left and dorsal sides up. White boxes serve as a reference to indicate the location of the bap domains in parasegments 5. CA) Stage 10 embryo stained for bap mRNA (purple) and En protein (brown). The anterior margins of bap expression coincide with the ectodermal parasegmental borders, but are broader than the posterior compartments marked by en. (B) Early stage 11 embryo stained for bap mRNA (purple) and Eve protein (brown). The heart (pericardial} progenitors marked by eve arise at positions between the bap patches at the dorsal margin of the mesoderm. (C) Stage ll embryo stained for srp mRNA (purple) and En protein (brown). The srp-stained fat body primordia are found at similar intrasegmental, but more ventrally located, positions as compared with the bap-stained visceral muscle primordia. (D) Stage 11 embryo stained for srp mRNA and Eve protein to compare the relative positions of heart and fat body primordia. Bottom right: Schematic summary of the spatial arrangement of mesodermal anlagen (top of panel: dorsal margin; bottom of panel: ventral midline). The primordia of the visceral mesoderm {vm) are located below the posterior compartments of the ectoderm (marked by en in yellow) and extend posteriorly into the first half of each adjacent anterior compart- ment. The primordia of the midgut visceral mesoderm (mgvm; purple) are located in dorsal portions and those of the fat body visceral mesoderm (fbvm; blue-green) immediately adjacent in lateral portions of the mesoderm. The primordia of the cardiac mesoderm are located between the vm primordia at the dorsal margin (cm: cardiac mesoderm; solid red indicates Eve-stained pericardial progenitors, stippled red indicates the presumed location of cardial progenitors). Most of the rest of the mesoderm will give rise to somatic mesoderm (sm; grey). (A,P) Mesodermal A and P domains; (PS) parasegment; (pc) pericardial cell progenitors; (CNS) neuronal progen- itors of the central nervous system.
derm is formed in the absence of eve function (Fig. 2I,K, cf. wild-type embryos in Fig. 2J and L}.
It appears that segmentat ion of the mesoderm is al- ready set up by the pair-rule genes during gastrulation, that is, at least 2 hr prior to the t ime when the dorsal bap patches appear. Upon overstaining of embryos hybrid- ized with bap antisense RNA probes, weak stripes of bap expression can be observed throughout the dorsoventral extent of the mesoderm as early as stage 8. Initially these stripes display a pair-rule distribution, but at stage 9, a regular pattern of weak transverse stripes is detected (Fig. 3A). This shows that mesoderm segmentat ion is initiated even before the the mesoderm spreads under the ectoderm and therefore that these early segmenta- tion events occur wi thout any influences from the ecto- derm. It has been proposed that the segmental expression of twist is required for visceral mesoderm specification (Baylies and Bate 1996). However, double stainings of stage 10 wild-type embryos for Twist protein and bap m R N A show that twist cannot determine the segmental bap pattern, because Twist protein is still evenly distrib- uted along the anteroposterior axis at the t ime when the bap patches are fully developed (Fig. 3B).
Prior to and during gastrulation, pair-rule genes are
expressed in stripes in both ectoderm and mesoderm. Therefore, they could regulate bap expression and me- soderm segmentat ion through either of two possible routes. The first could be direct and wi th in the mesoder- real cells proper. The second route could be via induc- tion from the overlying ectoderm. To find out which route is used, we made embryos that were genetically mosaic for eve; eve was chosen because it is essential for bap expression and development of the midgut visceral mesoderm. Nuclei from wild-type donor embryos marked with a lacZ reporter gene (Vincent et al. 1994) were transplanted into eve embryos that were stained at stage 10 for bap m R N A and f~-galactosidase. Figure 4A demonstrates that bap expression is rescued near to wild-type cells in an otherwise eve mutan t embryo. The restriction of the bap domains to the area of wild-type cells indicates that signals emanat ing from these cells, if any, are unable to act at a long distance to activate bap in adjacent eve- areas. We then prepared sections of the mosaic embryos. Figure 4B shows an embryo with two separate wild-type patches of cells, one restricted to the ectoderm (left side) and another extending into the me- soderm (right side). Importantly, bap expression is only observed in the mesodermal cells that are wild-type for
GENES & DEVELOPMENT 3185
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from
Figure 2. Regulation of mesoderm segmentation and specification by pair-rule genes, bap and srp mRNAs are stained purple; En, fasciclin III, and [3-galactosidase proteins are stained brown. Em- bryos shown in A - H are in stage 10. {A)ftz mutant embryo stained for hap and En. (B) ftz mutant em- bryo stained for srp and En. (C) hairy mutant stained for bap and En. {D) runt embryo stained for bap and En. The wedges demarcate examples for segments arranged in a mirror image fashion. (E) paired mutant stained for bap and En. Although the number of fully developed bap patches is re- duced from normally 11 to 6, weak bap signals are detected in intervening areas that lack detectable en stripes. (F) opa mutant stained for bap and En. Both the en and bap stripes are reduced in number and irregularly spaced. {G) bap expression in a stage 10 eve ~ mutant embryo. No bap expression in the midgut visceral mesoderm primordia is de- tected. (H) Stage 11 eve ~ embryo stained for srp (purple) and En (brown). srp is not expressed in the trunk region. (I) eve ~ mutant, and {J) wild-type em- bryo at early stage 12, both stained for fasciclin III. No midgut visceral mesoderm is formed in eve mutants. (K) eve ~ and {L) wild-type embryo at stage 13, both containing an enhancer trap inser- tion expressing ~-galactosidase in the fat body. The fat body is missing in the eve mutant embryo. (mgvm) Midgut visceral mesoderm; (fb) fat body.
eve, but not in eve mutan t cells that underly and contact wild-type cells of the dorsal ectoderm (Fig. 4B). The fail- ure of wild-type ectoderm to induce bap in the underly- ing eve mutan t mesoderm is not attributable to the small size of the wild-type patch in this particular em- bryo, because embryos with more extensive areas of wild-type cells in the ectoderm also fail to activate bap (Fig. 4C). These results demonstrate that at least one function of eve is required within the mesoderm itself for activating bap and for mesoderm segmentation.
Opposing roles of hh and wg in es tabl ishing m e s o d e r m a l A and P d o m a i n s
Because the pair-rule gene products disappear prior to the stage when bap and srp are fully activated, their function
in regulating these two genes mus t be indirect and in- volve intermediates. Here we examine whether segment polarity genes, which mediate pair-rule gene functions in the ectoderm, are also required for the segmentat ion of the mesoderm. The spatial overlap between the stripes of en or hh expression and the patches of bap or srp sug- gests that these two segment polarity genes are candi- dates (Figs. 1A, C and 5A; note that segment polarity gene expression is predominant ly ectodermal at this stage).
Both hh and en are indeed required for full activation of bap. In hh mutan t embryos, the bap domains are sig- nificantly reduced in size (Fig. 5B). There is also some reduction in embryos deficient for en (Fig. 5C). In Df(en);hh double mutants , there is an even stronger re- duction of bap expression than in mutan t s for either hh
3186 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from
Figure 3. Dynamics of bap expression in wild-type embryos. (A) Wild-type embryo at stage 9, overstained with a bap RNA probe. Weak transverse stripes are detected transiently even in ventral portions of the mesoderm. (This ventral expression ceases at stage 10 when dorsal mesodermal expression reaches high levels.) (B) Stage 10 wild-type embryo, stained for bap and Twist. Twist protein is still evenly distributed at the time when the mature bap pattern appears.
or en alone (Fig. 5D), therefore the function of en in al- lowing full bap activation is not mediated soley through hh. The lack of either hh or en function alone has only minor consequences for visceral mesoderm formation (data not shown), whereas the s imultaneous loss of func- tion of both genes results in a clear reduction of visceral mesoderm derivatives. As shown in Figure 5E, the nor- mal ly continuous band of the midgut visceral mesoderm (see Fig. 2J) is interrupted in Df(en);hh double mutants, and resembles that of mutan t embryos with strongly re- duced bap function (Azpiazu and Frasch 1993). The num- bers and sizes of cell clusters of the fat body visceral mesoderm are both reduced in the double mutants (Fig. 5E).
The requirement for hh and en for normal bap activa- tion could partially explain the absence of bap expres- sion in eve mutan t embryos, because these embryos lack hh and en expression in their t runk region. To test whether hh or en could rescue bap expression in the absence of eve, we ectopically expressed each of these two genes in the mesoderm of eve mutant embryos (see Materials and Methods). bap is indeed activated upon mesodermal expression of hh in eve mutant embryos, although the number of rescued bap patches is less than in the wild-type (Fig. 5F). The striped expression of bap in these embryos indicates that the mesoderm of eve mutan t embryos retains some periodic properties, which could reflect the contributions of other pair-rule genes to mesoderm segmentation. There is also a partial rescue of midgut visceral mesoderm formation. Ubiquitous ex- pression of en in the mesoderm of eve mutant embryos activates bap expression as well, albeit less efficiently than hh (data not shown). Taken together, these results
Figure 4. Mesoderm-autonomous requirement for eve func- tion to activate bap. Shown are mosaic embryos where host tissues are eve ~ Donor-derived tissues are wild-type and carry an arm-lacZ reporter gene insertion. Embryos are stained for bap mRNA {purplel and ~-galactosidase (brown). (A) bap expres- sion is strictly confined to a wild-type patch of cells {brown) in the otherwise eve mutant embryo. (B,C) Cross-sections of stained mosaic embryos, bap expression is seen only seen when wild-type patches encompass dorsal mesoderm (B, right), but not when eve function is provided in the dorsal ectoderm only (B, lefL, and C).
suggest that hh and en participate in the es tabl ishment of the mesodermal P domains that express both bap and srp and form visceral mesoderm derivatives.
bap is expressed in the mesodermal P domains, whereas the secreted protein Wg is synthesized adjacent in the A domains and, more prominently, in the A com- partments of the ectoderm (Fig. 6A). The alternating stripes of expression abut at the parasegmental borders, but small areas posterior to each bap patch lack detect- able levels of Wg protein. Wg appears to act negatively on bap, because bap expression is expanded in wg mutant embryos (this interaction may not be direct, however; see Discussion). In posterior regions of these embryos, a continuous rather than periodic pattern of bap expres- sion is observed in the dorsal mesoderm (Fig. 6B). More anteriorly, some periodic bap expression is still ob-
GENES & DEVELOPMENT 3187
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from
Figure 5. Positive regulation of bap by hh and en. Embryo in E is at stage 12, all others shown are at stage 10. mRNA detection appears purple and protein detection appears brown. CA) Wild-type embryo stained for bap mRNA and Hh protein. The bap and hh stripes are largely congruent. (B) hh mutant stained for bap mRNA and Wg protein. The bap patches are reduced in size. (C) Df(en) embryo stained for bap mRNA (and En protein). A minor reduction of bap expression is observed. (D) Df(en);hh double mutant embryo, stained as in C. bap expression is strongly reduced. (E) Df(en);hh double mutant embryo, stained for fasciclin III protein and srp mRNA. The visceral mesoderm of the midgut musculature and fatbody are substantially reduced, iF) eve ~ mutant embryo with ectopic mesodermal hh expression (using a twist-GAL4 driver) and stained for bap mRNA and Hh protein. Partial rescue of bap expression is observed.
served. Mutations in genes downstream of wg such as armadillo (arm; Fig. 6C) and dishevelled (dsh; Fig. 6D) have a similar phenotype. The expression of srp is also continuous in wg mutant embryos (Fig. 6E).
The subdivision of the mesoderm into A and P domains involves antagonistic interactions between hh and wg
Because the results described above indicate a positive function of hh and a negative function of wg in bap reg- ulation, we tested whether ectopic expression of hh or wg would affect bap expression in opposite directions. Ectopic expression of hh in the Krfippel domain of the ectoderm (Fig. 7A; Frasch 1995), in the whole ectoderm (data not shown), or in the whole mesoderm (Fig. 7B) of wild-type embryos results in a minor expansion of the bap domains only. By contrast, ectopic hh expression in either the ectoderm (Fig. 7C) or mesoderm (Fig. 7D) of wg mutant embryos causes massive overexpression of bap, which comes to resemble the expression pattern of tin at this stage (Bodmer et al. 1990; Azpiazu and Frasch 1993). If hh is ectopically expressed together with dpp in the ectoderm of wg mutant embryos, bap expression ex- pands also ventrally and is observed in the whole meso- derm between parasegments 2 and 12 (Fig. 7F; cf. ectopic expression of dpp in wild-type embryos, Fig. 7E). This clearly illustrates the combinatorial roles of anteropos- terior and dorsoventral cues in mesoderm patterning.
Ectopic expression of wg has opposite effects to those observed for ectopic hh. Whereas wg expression either in the ectoderm (within the Krfippel domain, Fig. 7G, or in the whole ectoderm, data not shown) or in the mesoderm of wild-type embryos (Fig. 7H) results only in a slight and partially penetrant reduction of bap expression, similar ectopic expression of wg in hh mutant embryos almost
entirely blocks bap expression in the midgut visceral mesoderm primordia (Fig. 7I, J). These results indicate that overexpression of hh in wg- , and of wg in h h - , produce unsegmented mesoderm. The patterns of bap expression suggest that the mesoderm has opposite iden- tities in these two experimental situations. Ectopic hh in wg- appears to result in a uniform P character, whereas the lack of bap expression upon ectopic wg expression in h h - signifies a uniform A character of the resulting me- soderm. Ectopic hh and wg can produce these alterations only if the other gene is not functional, but are unable to do so in wild-type embryos. Thus, we conclude that wg and hh normally antagonize each other's function. This antagonism is likely to be important for the proper sub- division of the mesoderm into A and P domains.
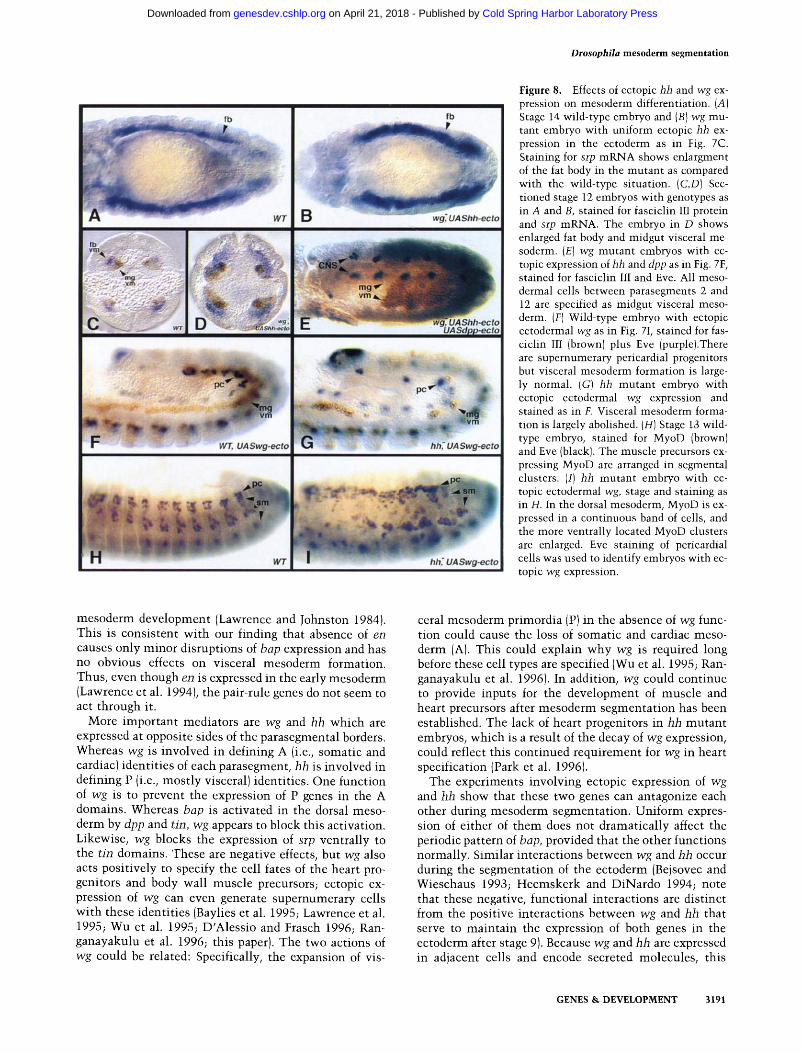
Analysis of wg- embryos with ectopic hh, and of h h - embryos with ectopic wg, confirmed that the mesoder- mal fate maps are shifted in opposite directions in these two situations. For example, the fat body in wg-/ec topic hh embryos is expanded as compared with wild-type and wg- embryos (Fig. 8A-D). The midgut visceral meso- derm appears also to be enlarged (Fig. 8C, D) at the ex- pense of heart precursors and somatic musculature (data not shown). The latter phenotypes are also observed in wg- embryos (Baylies et al. 1995; Wu et al. 1995), and it is difficult to judge whether the reduction of somatic muscles is enhanced upon ectopic hh expression. In wg mutant embryos with ectopic ectodermal expression of both hh and dpp, the visceral midgut mesoderm is even more expanded. It occupies the whole mesoderm be- tween parasegment 2 and 12, and no other mesodermal derivatives are formed in this region (Fig. 8E).
In stark contrast to wg-/ec topic hh embryos, hh-/ec- topic wg embryos show a massive reduction of visceral mesodermal tissues (Fig. 8G). Ectopic wg expression in wild-type embryos causes a minor reduction of visceral
3188 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from
Figure 6. Negative regulation of bap and srp by the wg path- way. (A) Wild-type embryo stained for bap mRNA and Wg pro- tein. The bap patches abut the wg stripes at their posterior margins but do not appear to fill the entire wg interstripe re- gions. (B) wg mutant embryo stained for bap mRNA and En protein, bap expression expands and becomes continuous, par- ticularly in the posterior segments. (C) Embryo lacking both the maternal and zygotic contributions of arm, stained as in B. (D) Embryo lacking both the maternal and zygotic contributions of dsh, stained as in B. bap expression in arm and dsh mutants is altered in a similar fashion as in wg-. (E) wg mutant embryo stained for srp mRNA expression, which becomes almost con- tinuous (cf. Fig. 1C).
mesoderm formation only, but the number of heart pro- genitors is clearly increased (Fig. 8F; Lawrence et al. 1995). The reduction of visceral mesoderm in hh mu- tants wi th ectopic wg is accompanied by an increased
number of cells with somatic mesodermal cell fates, as shown by the larger number of MyoD expressing cells. Whereas, in stage 13 wild-type embryos, there are seg- mental clusters of MyoD-expressing cells in the somatic mesoderm (Fig. 8H; Michelson et al. 1990; Paterson et al. 1991 ), hh- /ec top ic wg embryos have uniform MyoD ex- pression in the dorsal mesoderm, and the more ventral clusters appear expanded (Fig. 8I). Taken together, these results show that expansion of the mesodermal P do- mains at the expense of the A domains, as exemplified by wg /ectopic hh embryos, causes the mesoderm to form mostly visceral organs. By contrast, fate map shifts in the opposite direction in hh- /ec top ic wg embryos result in the exclusive formation of somatic muscles and heart.
D i s c u s s i o n
Anteroposterior subdivisions of mesodermal parasegments and the mesodermal fate map
Our analysis of mesodermal gene expression patterns shows that the early mesoderm is organized into para- segments that are in exact register wi th the ectodermal parasegments. The existence of stable parasegmental mesoderm boundaries is supported by our observation that the stripes of eve-lacZ reporter genes perdure in the mesoderm with sharp anterior borders that coincide with the anterior borders of the bap patches (M. Frasch, unpubl.). Each mesodermal parasegment is subdivided along the anteroposterior axis into a P and an A domain that express different control genes, have different devel- opmental fates, and respond differently to mutat ions in segmentation genes. The P domains comprise the ex- pression domains of bap plus those of srp, genes that mark the anlagen of the transverse midgut muscles and of the fat body, respectively. The A domains contain the anlagen of the heart and the bulk of the body wall mus- cles. This picture conforms wi th the morphological anal- ysis of Dunin-Borkowski et al. (1995), except we observe that the anlagen of the fat body are located ventrally rather than dorsally to those of the midgut musculature. Our assignment is further supported by an analysis of bap reporter genes showing that all bap expressing cells from the dorsal mesoderm will form midgut visceral me- soderm (Z. Yin and M. Frasch, unpubl.). We are not sure of the fate of those mesodermal cells located between the fat body anlagen and the ventral mid l ine in the P do- mains of each parasegment (Fig. 1). Although it is possi- ble that some of them give rise to mesodermal glial cells (Gorczyka et al. 1994), others probably generate specific body wall muscles. Thus, al though most of the somatic mesoderm is derived from the A domains, some comes from the P domains and therefore could be subject to different control. This could explain the presence of some residual somatic mesodermal cells and muscles in embryos with genetic backgrounds that appear to pro- duce a uniform "P" character in the mesoderm.
The subdivision of the mesodermal parasegments into P and A domains is reminiscent of the subdivision of ectodermal parasegments into posterior and anterior compartments. At present, we do not know whether
GENES & DEVELOPMENT 3189
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from
Figure 7. Effects of ectopic hh and wg ex- pression on bap expression. (A) Wild-type embryo with ectopic ectodermal hh expres- sion (ZKrGAL driver, see Frasch 1995) and stained for bap mRNA and Hh protein. (B) Wild-type embryo with ectopic mesoder- mal hh expression (twist-GAL4 driver) and stained as in A. In both situations, the bap patches are only slightly expanded. (C) wg mutant embryo with ectopic ectodermal hh expression (E22cGAL4 driver), stained as in A. bap expression is strongly ex- panded and becomes continuous along the anteroposterior axis. (D) wg mutant em- bryo with ectopic mesodermal hh expres- sion (twist-GAL4 driver), stained for bap mRNA and En protein. The bap pattern is similar to that seen with ectodermal over- expression of hh. (E) Wild-type embryo with ectopic ectodermal expression of dpp (E22cGAL4 driver), stained for bap mRNA and Eve protein. The bap stripes extend from the dorsal margin (d) to the ventral midline (v). (F) wg mutant embryo with ec- topic expression of hh and dpp (driver as in E), showing uniform bap expression be- tween parasegments 2 and 12. (G) Wild- type embryo with ectopic ectodermal ex- pression of wg (ZKrGAL driver as in Fig. 7A), stained for bap mRNA and Wg protein. A weak suppression of bap expression is observed, mostly in the patches #4 and #5. (H) Wild-type embryo with ectopic meso- dermal expression of wg (twist-GAL4 driver), stained as in A. Some suppression of bap expression is observed, particularly in more anterior parts of the mesoderm. Note that the examples shown in E and F are among the most strongly affected em- bryos found, and usually the reduction of bap expression is less conspicuous. (I) hh mutant embryo with ectopic ectodermal expression of wg (E22cGAL4 driver as in C) and {1) hh mutant embryo with ectopic me- sodermal expression of wg, stained as in E. In both situations, bap expression in the midgut visceral mesoderm primordia is al- most completely abolished.
there are lineage restrictions in the mesoderm prior to or during the stage when this subdivision becomes visible through the expression of genes such as bap and srp. However, it is l ikely that any such restrictions would occur before the onset of the third wave of mitot ic divi- sions in the mesoderm. By that t ime the outer borders of the bap patches form sharp boundaries and all bap-ex- pressing cells start segregating into the interior to form midgut visceral mesoderm (Azpiazu and Frasch 1993). Our observation that most body wall muscles are derived from the A domains may explain why it has not been possible to detect A and P compartments wi th in somatic musculature (Lawrence 1982). If there are lineage bound- aries between A and P domains in the gastrulating me- soderm, they would be expected to demarcate visceral
(gut muscle and fat body) P compar tments from somatic / cardiac A compartments.
Regulation of mesoderm patterning
The segmentat ion of the mesoderm and the anteropos- terior subdivision of the parasegments are controlled by gene systems that are similar, but not identical, to those that segment the ectoderm. In particular, the pair-rule genes and their upstream regulators have similar roles in both ectoderm and mesoderm and their muta t ions cause identical alterations of the number and polarity of seg- ments in the two germ layers. However, these genes may not act through an identical set of downstream genes. Indeed, previous experiments wi th genetic mosaics for en suggested that this gene is not required for normal
3190 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from
Figure 8. Effects of ectopic hh and wg ex- pression on mesoderm differentiation. (A) Stage 14 wild-type embryo and (B) wg mu- tant embryo with uniform ectopic hh ex- pression in the ectoderm as in Fig. 7C. Staining for srp mRNA shows enlargment of the fat body in the mutant as compared with the wild-type situation. (C,D) Sec- tioned stage 12 embryos with genotypes as in A and B, stained for fasciclin III protein and srp mRNA. The embryo in D shows enlarged fat body and midgut visceral me- soderm. (E) wg mutant embryos with ec- topic expression of hh and dpp as in Fig. 7F, stained for fasciclin III and Eve. All meso- dermal cells between parasegments 2 and 12 are specified as midgut visceral meso- derm. (F) Wild-type embryo with ectopic ectodermal wg as in Fig. 7I, stained for fas- ciclin III (brown) plus Eve (purple).There are supernumerary pericardiat progenitors but visceral mesoderm formation is large- ly normal. (G) hh mutant embryo with ectopic ectodermal wg expression and stained as in F. Visceral mesoderm forma- tion is largely abolished. (H) Stage 13 wild- type embryo, stained for MyoD (brown) and Eve (black). The muscle precursors ex- pressing MyoD are arranged in segmental clusters. (I) hh mutant embryo with ec- topic ectodermal wg, stage and staining as in H. In the dorsal mesoderm, MyoD is ex- pressed in a continuous band of cells, and the more ventrally located MyoD clusters are enlarged. Eve staining of pericardial cells was used to identify embryos with ec- topic wg expression.
mesoderm development (Lawrence and Johnston 1984). This is consistent wi th our finding that absence of en causes only minor disruptions of bap expression and has no obvious effects on visceral mesoderm formation. Thus, even though en is expressed in the early mesoderm (Lawrence et al. 1994), the pair-rule genes do not seem to act through it.
More important mediators are wg and hh which are expressed at opposite sides of the parasegmental borders. Whereas wg is involved in defining A (i.e., somatic and cardiac) identities of each parasegment, hh is involved in defining P (i.e., most ly visceral) identities. One function of wg is to prevent the expression of P genes in the A domains. Whereas bap is activated in the dorsal meso- derm by dpp and tin, wg appears to block this activation. Likewise, wg blocks the expression of srp ventrally to the tin domains. These are negative effects, but wg also acts positively to specify the cell fates of the heart pro- genitors and body wall muscle precursors; ectopic ex- pression of wg can even generate supernumerary cells wi th these identities (Baylies et al. 1995; Lawrence et al. 1995; Wu et al. 1995; D'Alessio and Frasch 1996; Ran- ganayakulu et al. 1996; this paper). The two actions of wg could be related: Specifically, the expansion of vis-
ceral mesoderm primordia (P) in the absence of wg func- tion could cause the loss of somatic and cardiac meso- derm (A). This could explain why wg is required long before these cell types are specified (Wu et al. 1995; Ran- ganayakulu et al. 1996). In addition, wg could continue to provide inputs for the development of muscle and heart precursors after mesoderm segmentat ion has been established. The lack of heart progenitors in hh mutan t embryos, which is a result of the decay of wg expression, could reflect this continued requirement for wg in heart specification (Park et al. 1996).
The experiments involving ectopic expression of wg and hh show that these two genes can antagonize each other during mesoderm segmentation. Uniform expres- sion of either of them does not dramatical ly affect the periodic pattern of bap, provided that the other functions normally. Similar interactions between wg and hh occur during the segmentat ion of the ectoderm (Bejsovec and Wieschaus 1993; Heemskerk and DiNardo 1994; note that these negative, functional interactions are distinct from the positive interactions between wg and hh that serve to maintain the expression of both genes in the ectoderm after stage 9). Because wg and hh are expressed in adjacent cells and encode secreted molecules, this
GENES & DEVELOPMENT 3191
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from
functional antagonism could help ensure that wg func- tion is limited to the A domains and hh function to the P domains of the mesoderm.
Mesoderm-autonomous vs. inductive regulation of mesoderm segmentation
The close contact between ectoderm and mesoderm after gastrulation could allow indirect regulation of meso- derm segmentation by pair-rule genes, through their role in the segmental subdivision of the ectoderm. Inductive signals from ectodermal cells could transmit this posi- tional information to the mesoderm and instruct meso- dermal cells to assume either A or P properties. Such a mechanism would be analogous to the dorsoventral sub- division of the mesoderm, which is induced by dorsal ectodermal cells through the secreted signal molecule Dpp (Staehling-Hampton et al. 1994; Frasch 1995). Both wg and hh act in the mesoderm and their ectopic expres- sion in the ectoderm can indeed alter the pattern of me- sodermal bap expression and induce changes in visceral mesoderm development. Similarly, ectopic expression of wg in the ectoderm affects the development of somatic muscle precursors (Baylies et al. 1995; Ranganayakulu et al. 1996; this paper). However, embryos that are geneti- cally mosaic for wg show that the function of wg in heart formation can be provided either from the ectoderm or from within the mesoderm (Lawrence et al. 1995). We believe that this function of wg is, at least in part, a consequence of its general role in determining the A do- mains of each parasegment. Therefore, it is likely that its function to promote somatic mesoderm development and to suppress visceral mesoderm in the A domains is also provided from both ectoderm and mesoderm. Note that the gradual decrease of wg and hh expression in the mesoderm during gastrulation suggests that the ectoder- mal contributions of both genes, which persist at high levels, are most significant for normal mesoderm devel- opment (Baker 1988; Lee et al. 1992; Mohler and Vani 1992; Tabata et al. 1992; Baylies et al. 1995).
Importantly, our results with eve mosaic embryos demonstrate that pair-rule genes act within the meso- derm to segment it. Thus, wg and hh signals provided by ectodermal patches of wild-type cells are not sufficient to mediate normal mesoderm segmentation and bap ac- tivation. Probably, the pair-rule genes establish first a prepattern of gene expression within the mesoderm and later inductive inputs from the ectoderm ensure that the mesodermal and ectodermal parasegments remain in ex- act register.
What are the candidates for genes that are regulated by the pair-rule genes in the mesoderm? We postulate that there exist as yet unidentified downstream genes in ad- dition to hh and wg that are expressed in striped patterns in the early mesoderm. Such genes could be responsible for the residual segmental expression of bap in wg;hh double or wg, en;hh triple mutant embryos (M. Frasch, unpubl.) and the faint stripes of bap expression in the early wild-type mesoderm. A potential candidate is the paired-domain gene pox meso, which is expressed in seg-
mental stripes in the early mesoderm (Bopp et al. 1989). The expression of twist becomes striped, with high-pro- tein levels present in the A domains, and this periodic modulation is required to allow visceral mesoderm de- velopment (Baylies and Bate 1996). However, twist can- not be involved in organizing the mesodermal A and P domains, because its periodic expression appears only after the segmental expression of bap has been estab- lished. It is more probable that twist could be controlled at a similar level of the regulatory hierarchy as bap and its disappearance in the A domains could be required for the function, rather than the regulation, of bap.
In conclusion, our current view of mesodermal seg- mentation and patterning can be summarized as follows (see Fig. 9): (1) During gastrulation, pair-rule genes con- trol the expression of target genes in segmentally re- peated stripes in the mesoderm. This process is autono- mous to the mesoderm and establishes the A and P do- mains. Mesodermal targets of the pair-rule genes include hh and en (in P domains) and wg (in A domains), en plays only a marginal role, and therefore we believe that there are as yet unidentified pair-rule target genes, M and N, that are important for the allocation of mesodermal cells to the A and P domains {Fig. 9, left). Gene M could be required to activate bap and srp in P domains, whereas N could activate target genes in A domains that are in- volved in somatic and cardiac mesoderm formation. Nei- ther M nor N is sufficient to activate their target genes; they both would later require localized coregulators that are not active in the early mesoderm.
(2) After gastrulation, the pair-rule gene products fade away and the maintenance of the striped expression of M and N requires hh and wg function. The mesodermally expressed hh and wg products can only maintain normal A and the P domains (i.e., normal M and N expression) transiently as they are not expressed beyond stage 8 or 9. In the ectoderm, hh and wg maintain each other's ex- pression after this stage. Therefore, the contribution of hh and wg products secreted from the ectoderm is likely
Figure 9. Model of mesoderm segmentation and the specifica- tion of mesodermal tissues. We propose that mesoderm seg- mentation occurs in two phases; an early, mesoderm-autono- mous phase (left) and a subsequent phase that involves meso- derm-autonomous as well as inductive regulation (right). During the second phase, developmental control genes are ac- tivated in quadrants at the intersections of transverse and lon- gitudinal domains of patterning genes (see text for details).
3192 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from
to become s igni f icant later. Funct iona l an tagon i sm be- tween hh and wg cont r ibutes to the format ion of sharp borders be tween A and P domains (Fig. 9, right).
(3) After the m e s o d e r m has spread below the ectoderm, the cells in the dorsal por t ions of each t ransverse stripe receive Dpp signals from the dorsal ectoderm and there- fore express h igh levels of tin. This combina t ion of trans- verse and long i tud ina l ac t iv i t ies (gene M plus T i n / D p p 1 is suff icient to ac t ivate h igh levels of bap in a dorsal quadrant of each P domain. Similarly, different combi- na t ions could act ivate deve lopmcnta l control genes in other quadrants and thereby define other anlagen.
(4) bap specifies the midgu t visceral mcsoderm, whereas genes such as srp de te rmine the fat body vis- ceral mesoderm. Both types of visceral mesoderm segre- gate into the in ter ior and their deve lopment becomes independen t from the ectoderm. However, the mesodcr- real cells of the A domains remain in contact w i t h the ec toderm and wg could therefore con t inue to inf luence the deve lopment and pa t t e rn ing of the somat ic and car- diac mesoderm.
Mater ia l s and m e t h o d s
Drosophila mutant strains
The following segmentation mutants were used in this study: eve L27, ftz 9H34, h 7A94, prd 2.45, odd 1.36.1~, o[,Ta Ilp32, rlln LBF',
armHS 6; Df(2R)en c Ideleting both en and invected (inv); Gustav- son et al. 19961, Df(2R)gsb and Cl I)ga-mlA (both of which had normal hap expression, data not shown), dsh 477, hh l'<:>), and Wc~ cx4. Mosaic germlines for arm and dsh were generated by using the yeast recombinase-based FLP-DTS system (Chou and Perrimon 1992). Recombination was induced in femalcs ho- mozygous for an FRT insertion on the X chromosome (FRT mr) and heterozygous for the dominant female sterile mutation ovo TM, as well as the mutation to be studied. The source of FLP recombinase was an autosomal insertion of FLP recombinase under the control of the heat shock promoter (FLp~S). The FLP/ FRT stocks were provided by E. Siegfried (Pennsylvania State University, State College). Homozygous pair-rule mutant em- bryos were identified through altered e , patterns, hh mutant embryos through the use of a TM3,lacZ balancer or their de- creased wg expression, and wg mutants through their decreased en or hh expression.
UAS/GAL4 strains
Generation of UASwg: wg cDNA c14 (provided by N. Baker, Albert Einstein College of Medicine, Bronx, NY) was cloned as a BamHI-XbaI fragment into BglII-Xbal of pUAST (Brand and Perrimon 1993). An insertion of this P element on the third chromosome (UASwqg2) was used throughout the experiments.
Generation of UAShh: hh cDNA 11 (provided by Phil Beachy, Johns Hopkins University, Baltimore, MD) was made by first cloning a ftindIII-EcoRI insert from hhl l -pNB40 into pBlue- script KS+ and then a KpnI-Xbal insert was recloned into pUAST. The line UAShhl with an insertion on the third chro- mosome was used in all experiments described.
For ectopic expression in the mesoderm, we used the strains GALSG24 (twist-GAL4 insertion on the second chromosome) or GALSG30 (twist-GALd + 2dB enhancer trap insertion on the third chromosome)(Brand and Perrimon 1993; Greig and Akam 1993), both provided to us by A. Michelson (Brigham and Women's Hospital, Boston, MA). Ectopic expression in the ec-
toderm was achieved with the driver lines ZKrGAL8 (in the Kr0ppel domain; Frasch 1995} or E22c-GAL4 (made by K. Yoffe, Harvard Medical School, Boston, MA). The expression pattern of E22c-GAL4 was tested by crossing it with Bg4-i-2 (Brand and Perrimon 1993). Araldite sections of embryos stained for f3-galactosidase demonstrated that the expression from this driver is restricted to the embryonic ectoderm (M. Frasch, un- publ.). The line UASdpp-5 (third chromosome insertion) was used for dpp misexpression (Frasch 19951. Embryos from UAS- GAL4 crosses were collected at 27~ The enhancer trap line E7-3-63 (provided by V. Hartenstein, University of California, Los Angeles) was used to analyze fat body formation.
Nuclear transplantation and analysis of mosaic embryos
Nuclear transplantations were done as described previously (Lawrence et al. 1994; Vincent and Lawrence 1994), using do- nors marked with arm-lacZ. Hosts were an eve 127 mutant strain maintained over a CyO,hb-lacZ balancer. Mosaic em- bryos were processed for staining with ~-gal antibodies, fol- lowed by hybridization with digoxygenin-labeled bap cDNA probes as described in Azpiazu and Frasch (1993). Nine of the obtained mosaic embryos were analyzed in serial sections. Of these, three had mesodermally restricted wild-type cells show- ing hap expression, four had ectodermally restricted wild type cells and lacked bap expression, and in two embryos both ec- toderm and the underlying mesoderm was wild-type for eve.
Antibody staining and in situ hybridization of embryos
The following antibodies were used for the phenotypic analysis: Monoclonal mouse-anti En (4D9, provided by C. Doe, Univer- sity of illinois, Urbana), rabbit-anti Eve (Frasch et al. 1987), rabbit-anti Hh (rabbit 1255, provided by J. Knight and T. Korn- berg, University of California, San Francisco), rabbit-anti Wg (provided by R. Nusse, Stanford University, CA), polyclonal mouse-anti 6-galactosidase (Sigma), monoclonal mouse-anti fasciclm III (7G 10, provided Developmental Studies Hybridoma Bank), rabbit-anti Twist (provided by S. Roth, Max Planck In- stitut for Entwicklungsbiologie, Tfibingen, Germany), rabbit- anti Dmyd (1184, provided by B. Paterson, National Institute of Health, Bethesda, MD}. Antibody double stainings and anti- body-in situ hybridization double stainings were performed as described in Azpiazu and Frasch (1993).
A c k n o w l e d g m e n t s
We thank Zhizhang Yin for technical assistance and Patrick Lo for bap RNA in situ hybridizations. We thank Esther Siegfried, Andrea Brand, Tom Kornberg, Volker Hartenstein, Alan Mich- elson, Leslie Pick, Jym Mohler, and Ken Yoffe for fly strains, and Chris Doe, Jonathan Knight, Tom Kornberg, Roel Nusse, Bruce Paterson, Siegfried Roth, and the Developmental Studies Hy- bridoma Bank for antibodies. We appreciate the helpful com- ments of Hanh Nguyen on the manuscript. This work was sup- ported by National Institutes of Health grant HD30832 and a Pew Scholarship in the Biomedical Sciences to M.F. and by a fellowship from the Basque Country to N.A.
The publication costs of this article were defrayed in part by payment of page charges. This article must therefore be hereby marked "advertisement" in accordance with 18 USC section 1734 solely to indicate this fact.
References
Abel, T., A. Michelson, and T. Maniatis. 1993. A Drosophila GATA family member that binds to Adh regulatory se- quences is expressed in the developing fat body. Develop- ment 119: 623-633.
GENES & DEVELOPMENT 3193
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from
Azpiazu, N. and M. Frasch. 1993. tinman and bagpipe: Two homeo box genes that determine cell fates in the dorsal me- soderm of Drosophila. Genes & Dev. 7" 1325-1340.
Baker, N.E. 1988. Localization of transcripts from the wingless gene in whole Drosophila embryos. Development 103: 289- 298.
Baylies, M., A. Martinez Arias, and M. Bate. 1995. wingless is required for the formation of a subset of muscle founder cells during Drosophila embryogenesis. Development 121- 3829- 3837.
Baylies, M.K. and M. Bate. 1996. twist: A myogenic switch in Drosophila. Science 272: 1481-1484.
Bejsovec, A. and E. Wieschaus. 1993. Segment polarity gene interactions modulate epidermal patterning in Drosophila embryos. Development 119:501-517.
Bodmer, R. 1993. The gene tinman is required for specification of the heart and visceral muscles in Drosophila. Develop- ment 118: 719-729.
Bodmer, R., L.Y. Jan, and Y.N. Jan. 1990. A new homeobox- containing gene, rash-2, is transiently expressed early during mesoderm formation of Drosophila. Development 110: 661- 669.
Bopp, D., E. Jamet, S. Baumgartner, M. Burri, and M. Noll. 1989. Isolation of two tissue-specific Drosophila paired box genes, Pox meso and Pox neuro. EMBO J�9 8: 3447-3457.
Brand, A.H. and N. Perrimon. 1993. Targeted gene expression as a means of altering cell fates and generating dominant phe- notypes. Development 118: 401-415.
Chou, T.-B. and N. Perrimon. 1992. Use of a yeast site specific recombinase to produce female germline chimeras in Droso- phila. Genetics 131: 643-653.
D'Alessio, M. and M. Frasch. 1996. rash may play a conserved role in dorsoventral patterning of the neuroectoderm and mesoderm. Mech. Dev. 58: 217-231.
Dunin-Borkowski, O., N. Brown, and M. Bate. 1995. Anterior- posterior subdivision and the diversification of the meso- derm in Drosophila. Developm en t 121" 4183-4193.
Frasch, M. 1995. Induction of visceral and cardiac mesoderm by ectodermal Dpp in the early Drosophila embryo. Nature 374: 464--467.
Frasch, M., T. Hoey, C. Rushlow, H.J. Doyle, and M. Levine. 1987. Characterization and localization of the even-skipped protein of Drosophila. EMBO J. 6: 749-759.
Gorczyka, M.G., R.W. Phyllis, and V. Budnik. 1994. The role of tinman, a mesodermal cell fate gene, in axon pathfinding during the development of the transverse nerve in Droso- phila. Development 120: 2143-2152.
Greig, S. and M. Akam. 1993. Homeotic genes autonomously specify one aspect of pattern in the Drosophila mesoderm. Nature 362: 630-632.
Gustavson, E., A.S. Goldsborough, Z. Ali, and T.B. Komberg. 1996. The Drosophila engrailed and invected genes: Part- ners in regulation, expression and function. Genetics 142: 893-906.
Harding, K., C. Rushlow, H.J. Doyle, T. Hoey, and M. Levine. 1986. Cross-regulatory interactions among pair-rule genes in Drosophila. Science 233: 953-959.
Heemskerk, J. and S. DiNardo. 1994. Drosophila hedgehog acts as a morphogen in cellular patterning. Cell 76: 449-460.
Hoshizaki, D.K., T. Blackburn, C. Price, M. Ghosh, K. Miles, M. Ragucci, and R. Sweis. 1994. Embryonic fat-cell lineage in Drosophila melanogaster. Development 120: 2489-2499.
Lawrence, P.A. 1982. Cell lineage of the thoracic muscles of Drosophila. Cell 29: 493-503.
�9 1992. The making of a fly: The genetics of animal de- sign. Blackwell Scientific Publications, Oxford.
Lawrence, P.A. and P. Johnston. 1984. On the role of the en- grailed + gene in the internal organs of Drosophila. EMBO J. 3: 2839-2844.
Lawrence, P.A., P. Johnston, and J.-P. Vincent. 1994. Wingless can bring about a mesoderm-to-ectoderm induction in Drosophila. Development 120" 3355-3359.
Lawrence, P.A., R. Bodmer, and J.-P. Vincent. 1995. Segmental patterning of heart precursors in Drosophila. Development 121: 4303-4308.
Lee, J.J., D.P. yon Kessler, S. Parks, and P.A. Beachy. 1992. Se- cretion and localized transcription suggest a role in posi- tional signalling for products of the segmentation gene hedgehog. Cell 71: 33-50.
Macdonald, P.M., P. Ingham, and G. Struhl. 1986. Isolation, structure, and expression of even-skipped: A second pair- rule gene of Drosophila containing a homeo box. Cell 47: 721-734.
Maggert, K., M. Levine, and M. Frasch. 1995. The somatic-vis- ceral subdivision of the embryonic mesoderm is initiated by dorsal gradient thresholds in Drosophila. Development 121: 2107-2116.
Michelson, A.M., S.M. Abmayr, M. Bate, A. Mart/nez-Arias, and T. Maniatis. 1990. Expression of a MyoD family member prefigures muscle pattern in Drosophila embryos. Genes & Dev. 4: 2086-2097.
Mohler, J. and K. Vani. 1992. Molecular organization and em- bryonic expression of the hedgehog gene involved in cell- cell communication in segmental patterning of Drosophila. Development 115:957-971.
Nfisslein-Volhard, C. and E. Wieschaus. 1980. Mutations affect- ing segment number and polarity in Drosophila. Nature 287: 795-801.
Nfisslein-Volhard, C., H. Kluding, and G. Jiirgens. 1985. Genes affecting the segmental subdivision of the Drosophila em- bryo. Cold Spring Harbor Symp. Quant. Biol. 50: 145-154.
Park, M., X. Wu, K. Golden, J.D. Axelrod, and R. Bodmer. 1996. The Wingless signaling pathway is directly involve in Droso- phila heart development. Dev. Biol. 177: 104--116.
Paterson, B.M., V. Walldorf, J. Eldrigde, A. Dfibendorfer, M. Frasch, and W. Gehring. 1991. The Drosophila homologue of vertebrate myogenic-determination genes encodes a tran- siently expressed nuclear protein marking primary myo- genic cells. Proc. Natl. Acad. Sci. 88: 3782-3786.
Ranganayakulu, G., R.A. Schulz, and E.N. Olson. I996. wing- less signaling induces nautilus expression in the ventral me- soderm of the Drosophila embryo. Dev. Biol. 176: 143-148.
Rehorn, K.-P., H. Thelen, A. Michelson, and R. Reuter. 1996. A molecular aspect of hematopoiesis and endoderm develop- ment common to vertebrates and Drosophila. Development 122:4023-4031.
Staehling-Hampton, K., F.M. Hoffmann, M.K. Baylies, E. Rush- ton, and M. Bate. 1994. dpp induces mesodermal gene ex- pression in Drosophila. Nature 372: 783-786�9
Tabata, T., S. Eaton, and T.B. Kornberg. 1992. The Drosophila hedgehog gene is expressed specifically in posterior compart- ment cells and is a target of engrailed regulation. Genes & Dev. 6: 2635-2645.
Vincent, J.-P. and P.A. Lawrence. 1994. Drosophila wingless sustains engrailed expression only in adjoining cells: Evi- dence from mosaic embryos. Cell 77: 909-915.
Vincent, J.-P., C. Girdham, and P. O'Farrell. 1994. A cell-auton- omous, ubiquitous marker for the analysis of Drosophila genetic mosaics. Dev. Biol. 164: 328-331.
Wu, X., K. Golden, and R. Bodmer. 1995. Heart development in Drosophila requires the segment polarity gene wingless. Dev. Biol. 169: 619-628.
3194 GENES & DEVELOPMENT
Cold Spring Harbor Laboratory Press on April 21, 2018 - Published by genesdev.cshlp.orgDownloaded from