REGULAR ARTICLE Septin9 is involved in T-cell development and CD8 + T-cell homeostasis Louise Berkhoudt Lassen & Annette Füchtbauer & Alexander Schmitz & Annette Balle Sørensen & Finn Skou Pedersen & Ernst-Martin Füchtbauer Received: 29 August 2012 / Accepted: 11 March 2013 / Published online: 4 May 2013 # Springer-Verlag Berlin Heidelberg 2013 Abstract SEPTIN9 (SEPT9) is a filament-forming protein involved in numerous cellular processes. We have used a conditional knock out allele of Sept9 to specifically delete Sept9 in T-cells. As shown by fluorescence-activated cell sorting, loss of Sept9 at an early thymocyte stage in the thymus results in increased numbers of double-negative cells indicating that SEPT9 is involved in the transition from the double-negative stage during T-cell development. Accordingly, the relative numbers of mature T-cells in the periphery are decreased in mice with a T-cell-specific dele- tion of Sept9. Proliferation of Sept9-deleted CD8 + T-cells from the spleen is decreased upon stimulation in culture. The altered T-cell homeostasis caused by the loss of Sept9 results in an increase of CD8 + central memory T-cells. Keywords Sept9 . T-celldevelopment . CD8 + T-cells . T-cell homeostasis . T-cell proliferation . Mice (Sept9 cond :Mx-Cre) Introduction Septins are a group of filament-forming proteins which have been called the fourth component of the cytoskeleton (Mostowy and Cossart 2012). They were originally found in cell-cycle mutants of yeast (Hartwell 1971). In budding yeast, they serve as a platform formed as an hourglass- shaped collar at which components needed for septation are recruited. At later stages of the septation process, the septin collar transforms into a pair of rings functioning as diffusion barriers at the neck between the mother and daugh- ter cells (McMurray and Thorner 2009). In mammals, septins are associated with cytokinesis and other membrane-associated events (Hall and Russell 2012), e.g., as diffusion barriers at the base of cilia (Hu et al. 2010). The alteration of normal septin expression might cause diseases such as cancer, whereas a point mutation results in the rare disorder of hereditary neuralgic amyothrophy (Kuhlenbaumer et al. 2005; Hall and Russell 2012). Furthermore, septins play a role in bacterial infection as they form cages around certain bacteria (Mostowy et al. 2010). Thirteen different septins are known in mammals (SEPT1-12 and SEPT14; Russell and Hall 2011), which, on the basis of sequence similarity, are divided into four subgroups (Hall and Russell 2004). SEPT9 belongs to the SEPT3 subgroup together with SEPT3 and SEPT12. Septins associate with each other to form heteromeric complexes. The best-described mammalian septin complex is the six- subunit heteromer built up in the following order SEPT7, SEPT6, SEPT2, SEPT2, SEPT6, and SEPT7 (Sirajuddin et Finn Skou Pedersen and Ernst-Martin Füchtbauer equally contributed to this work. This study was supported in part by grants from the Danish Cancer Society, the Danish Medical Research Council, the Novo Nordisk Foundation, and the Danish Genetically Modified Animal Resource (DAGMAR) funded by the Danish Agency for Science, Technology, and Innovation. The authors declare no conflict of interest. L. B. Lassen : A. Füchtbauer : A. Schmitz : F. S. Pedersen : E.-M. Füchtbauer (*) Department of Molecular Biology and Genetics, Aarhus University, Bldg. 1130, C.F. Mollers Allé 3, 8000 Aarhus, Denmark e-mail: [email protected]A. Schmitz Department of Haematology, Aalborg University Hospital, Aalborg University, Aarhus, Denmark A. B. Sørensen The State and University Library, Aarhus, Denmark Cell Tissue Res (2013) 352:695–705 DOI 10.1007/s00441-013-1618-6
Transcript
REGULAR ARTICLE
Septin9 is involved in T-cell development and CD8+ T-cellhomeostasis
Louise Berkhoudt Lassen & Annette Füchtbauer &
Alexander Schmitz & Annette Balle Sørensen &
Finn Skou Pedersen & Ernst-Martin Füchtbauer
Received: 29 August 2012 /Accepted: 11 March 2013 /Published online: 4 May 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract SEPTIN9 (SEPT9) is a filament-forming proteininvolved in numerous cellular processes. We have used aconditional knock out allele of Sept9 to specifically deleteSept9 in T-cells. As shown by fluorescence-activated cellsorting, loss of Sept9 at an early thymocyte stage in thethymus results in increased numbers of double-negativecells indicating that SEPT9 is involved in the transition fromthe double-negative stage during T-cell development.Accordingly, the relative numbers of mature T-cells in theperiphery are decreased in mice with a T-cell-specific dele-tion of Sept9. Proliferation of Sept9-deleted CD8+ T-cellsfrom the spleen is decreased upon stimulation in culture.The altered T-cell homeostasis caused by the loss of Sept9results in an increase of CD8+ central memory T-cells.
Septins are a group of filament-forming proteins which havebeen called the fourth component of the cytoskeleton(Mostowy and Cossart 2012). They were originally foundin cell-cycle mutants of yeast (Hartwell 1971). In buddingyeast, they serve as a platform formed as an hourglass-shaped collar at which components needed for septationare recruited. At later stages of the septation process, theseptin collar transforms into a pair of rings functioning asdiffusion barriers at the neck between the mother and daugh-ter cells (McMurray and Thorner 2009).
In mammals, septins are associated with cytokinesis andother membrane-associated events (Hall and Russell 2012),e.g., as diffusion barriers at the base of cilia (Hu et al. 2010).The alteration of normal septin expressionmight cause diseasessuch as cancer, whereas a point mutation results in the raredisorder of hereditary neuralgic amyothrophy (Kuhlenbaumeret al. 2005; Hall and Russell 2012). Furthermore, septins play arole in bacterial infection as they form cages around certainbacteria (Mostowy et al. 2010).
Thirteen different septins are known in mammals(SEPT1-12 and SEPT14; Russell and Hall 2011), which,on the basis of sequence similarity, are divided into foursubgroups (Hall and Russell 2004). SEPT9 belongs to theSEPT3 subgroup together with SEPT3 and SEPT12. Septinsassociate with each other to form heteromeric complexes.The best-described mammalian septin complex is the six-subunit heteromer built up in the following order SEPT7,SEPT6, SEPT2, SEPT2, SEPT6, and SEPT7 (Sirajuddin et
Finn Skou Pedersen and Ernst-Martin Füchtbauer equally contributedto this work.
This study was supported in part by grants from the Danish CancerSociety, the Danish Medical Research Council, the Novo NordiskFoundation, and the Danish Genetically Modified Animal Resource(DAGMAR) funded by the Danish Agency for Science, Technology,and Innovation.
The authors declare no conflict of interest.
L. B. Lassen :A. Füchtbauer :A. Schmitz : F. S. Pedersen :E.-M. Füchtbauer (*)Department of Molecular Biology and Genetics, AarhusUniversity, Bldg. 1130, C.F. Mollers Allé 3,8000 Aarhus, Denmarke-mail: [email protected]
A. SchmitzDepartment of Haematology, Aalborg University Hospital,Aalborg University, Aarhus, Denmark
A. B. SørensenThe State and University Library, Aarhus, Denmark
Cell Tissue Res (2013) 352:695–705DOI 10.1007/s00441-013-1618-6
al. 2007). SEPT2 and SEPT6 can be substituted with othermembers of their subgroup, possibly explaining the largedegree of redundancy seen among septins. Recently, SEPT9has been localized to each end of the non-polar heteromerfilament creating an eight-unit heteromer, but whether thefunction of SEPT9 can be substituted by subgroup membersremains unknown (Kim et al. 2011; Sellin et al. 2011). Inembryonic fibroblasts, the deletion of Sept9 leads to changesin the subcellular localization and filament appearance ofSEPT7 thus showing a dependency on SEPT9 for the cor-rect function of SEPT7 (Füchtbauer et al. 2011).
SEPT9/Sept9 has a wide-spread expression (Sørensen etal. 2002), which is especially high in lymphoid tissue (Hallet al. 2005). The importance for lymphoid development isfurther highlighted by murine Sept9 being identified as acommon integration site in retroviral-induced T-cell lym-phomas (Sørensen et al. 2000).
In addition, SEPT1, SEPT6, SEPT7, and SEPT9 havebeen found to co-immunoprecipitate in the murine D10.G4CD4+ T-cell line. In motile D10.G4 cells, septins have beenfound at the uropod cortex, possibly serving as a rigidifyingcorset. SEPT7 depletion in these cells leads to disruptedmorphology and motility, whereas the depletion of SEPT9has no obvious effect (Tooley et al. 2009).
In order to study the function of SEPT9 in T-cell develop-ment in vivo, we have deleted Sept9 at the double-negative(DN) stage of T-cell development by crossing our conditionalSept9 knock out mouse with the T-cell-specific Lck-Cremouse (Orban et al. 1992). We report here that SEPT9 isimportant for T-cell development and proliferation.
Materials and methods
Mice
The Sept9TM2-EMFU (in this article termed Sept9cond) and theSept9TM1-EMFU (in this article termed Sept9neo) alleles aredescribed in Füchtbauer et al. (2011). Sept9cond mice werecrossed with Lck-Cre mice (B6-Cg-TgN (Lck-Cre)548Jxm;Orban et al. 1992) in order to generate Sept9cond:Lck-Cremice deleted for Sept9 in T-cells, and with Mx-Cre mice(Kuhn et al. 1995) in order to generate Sept9cond:Mx-Cremice deleted for Sept9 in bone marrow, liver, and lymphatictissue upon polyI polyC treatment. Sept9cond:Mx-Cre micewere treated with five doses of 300 μg polyI polyC over17 days. All experiments were performed with permissionfrom the responsible Danish authorities.
RNA purification
Total RNAwas purified from whole organs or leukocytes byusing TRIzol reagent (Invitrogen). Total RNA from cells
after fluorescence-activated cell sorting (FACS) analysiswas purified with a high performance isolation kit (Roche)and was reverse-transcribed by using random hexamerprimers with M-MLV reverse transcriptase (Invitrogen).
Determination of mRNA levels was carried out by quan-titative real-time polymerase chain reaction studies withTaqman hydrolysis probes (Applied Biosystems) in aStratagene Mx3000 apparatus. Data were analyzed withMxPro Software. The [Mm00450491_g1] Taqman probewas utilized for expression analysis of Sept9. B2m[Mm00437762_m1] was used as an internal standard.
FACS analysis
Fluorochrome-conjugated antibodies were used in a singletube for analysis of T-cells (all derived from BDBioscience). Anti-CD8-PerCP-Cy5.5 and anti-CD4-FITCwere used in combination with anti-CD3-PE as a pan T-cellmarker to distinguish between the various T-cell differentia-tion stages. Anti-CD25-PE-Cy7, anti-CD44-V450, and anti-CD62L-APC were included in the panel to distinguish sub-populations of DN subsets in thymus and central memory T-cells of spleen. After lysis of erythrocytes, suspensions of onemillion single cells from either spleen or thymus were incu-bated with 1 μg blocking antibody (FcγIII/II receptor)followed by 30-min incubation with surface marker monoclo-nal antibody combinations for 30 min at 4°C, washed exten-sively, and analyzed in phosphate-buffered saline plus 2%fetal bovine serum. Non-stained samples were run as negativecontrols. Propidium iodide (PI) was used to recognize deadcells. Anti-CD19-PerCP-Cy5.5 and anti-CD11b-FITC wereused to distinguish B-cells and myeloid cells, respectively.
FACS analysis was primarily performed on a FACSAriaIII at the FACS Core Facility, The Faculty of HealthSciences, Aarhus University, Denmark.
Unstained samples and samples including single-stainsamples served as controls for subsequent software compen-sation by using the “FlowJo” analysis software compensa-tion module (Treestar).
Blood stains
Blood smears were stained with Wright stain (Sigma Aldrich)for 15 s according to the manufacturer’s recommendations.
Cell proliferation assay
Single-cell suspensions were purified from spleen and thy-mus. Cells were stained with 5 μM carboxyfluoresceindiacetate succinimidyl ester (CFSE; Invitrogen, C34554) inRPMI glutaMAX and subsequently washed three times withmedium containing 20 mMHEPES. Cells (106 per well) wereseeded in a round-bottom 96-well plate. Anti-CD3e (BD
696 Cell Tissue Res (2013) 352:695–705
553238) and anti-CD28 (BD 557393) antibodies were addedto a final concentration of 0.5 μg/ml. Unstimulated cells weregrown as controls. After 5 days, cells were analyzed by flowcytometry. The following antibody panel was used: anti-CD8-PerCP-Cy5.5, anti-CD4-APC-Cy7, anti-CD44-v450, anti-CD25-PE-Cy7, and anti-CD62L-APC. PI was used to distin-guish dead and live cells but was excluded from the finalanalysis to avoid spectral overlap with CFSE.
Statistical analysis
We used a standard logistic model and a likelihood ratio testto test equality of the proportion of mononuclear cells. Aone-sided test was used to test the proportion of CFSE-positive cells within the CD8+ T-cell population. A pairedStudent’s t-test was used to test for statistical significance inall other experiments. P-values less than 0.05 were consid-ered to be statistically significant.
Results
Sept9 expression in lymphatic tissues
In a screen for putative proto-oncogenes in murine T-celllymphomas, we earlier detected the murine Sept9. We foundSept9 to be a frequent retroviral insertion site (Sørensen et al.2000), and subsequent expression analysis showed that theexpression of the ubiquitously expressed gene was increasedin thymus compared with other tissues. This encouraged us tocross mice carrying the conditional Sept9cond allele with trans-genic mice expressing CRE recombinase under the control ofthe T-cell-specific Lck promoter. All mice were homozygousfor the conditional allele Sept9cond and differed only in thepresence or absence of the Lck-Cre transgene. In this article,mice homozygously deleted for Sept9 in T-cells will thereforebe referred to as Sept9:LckCre, whereas control littermates arereferred to as wild-type (Wt).
In whole thymus and spleen, Sept9:LckCre mice showed14% and 70% of Sept9 expression, respectively, comparedwith Wt littermates, consistent with the T-cell contributionto the two tissues.
In isolated leukocytes from thymus, Sept9:LckCre miceshowed only 8% of the Sept9 expression observed in Wt(Fig. 1a). Consistent with the lower contribution of T-cells tospleen leukocytes, the level of Sept9 expression was here onlyreduced to 68% of theWt level in Sept9:LckCremice (Fig. 1b).
To determine at what stage of T-cell developmentSept9:LckCremice lose Sept9 expression, we purified variousstages of pre-T-cells from the thymus by FACS analysis anddetermined Sept9 expression. Sept9 expression started de-creasing in DN cells (CD4-CD8-). In later stages of T-celldevelopment (double-positive [DP] CD4+CD8+ and single-
positive CD4+ and CD8+) less than 10% of Wt Sept9 expres-sion, representing the level of non-deleted alleles, was foundin Sept9:LckCre mice. Interestingly, we observed a 5- to 10-fold increase in Sept9 expression in Wt mice with the shiftfrom DN to DP T-cells, suggesting a functional requirementfor SEPT9 (Fig. 1c). T-cells of the DN fraction can besubdivided into stages DN1 (CD4-CD8-CD25-CD44+), DN2(CD4-CD8-CD44+CD25+), DN3 (CD4-CD8-CD25+CD44-),and DN4 (CD4-CD8-CD25-CD44-). To reveal the time pointof Sept9 deletion, we analyzed Sept9 expression levels inDN1-DN4 cells. Sept9 expression decreased as early as theDN1 stage in Sept9:LckCre mice (Fig. 1d).
In order to analyze Sept9 expression in mature peripheralCD4+ and CD8+ T-cells, we isolated cells from the spleenand found a high efficiency of the Sept9 deletion, althoughsome Sept9:LckCre mice had a lower level of the Sept9deletion indicating an inefficiency of CRE recombinase(Fig. 1e). This explanation was supported by the findingthat individual mice that had a low level of Sept9 deletion inthe CD4+ cells also had a low level of Sept9 deletion in theCD8+ cells (exemplified in Fig. 1e in which the dotted linesconnect the data from identical mice).
DN pre-T-cells are increased in Sept9:LckCre mice
In order to identify whether the loss of Sept9 has functionalconsequences for T-cell development, we compared the size(measured in percentage of the total leukocyte cell population)of the various T-cell populations between Sept9:LckCre andWt mice in the thymus (Fig. 2a, b). The results showed arelative increase of DN T-cells in Sept9:LckCremice to 171%compared with Wt mice, whereas the relative numbers of DPand single-positive T-cells (CD4+ and CD8+) were unalteredsuggesting a requirement for SEPT9 at the transition from theDN to the DP stage (Fig. 2c).
To further analyze the requirement for SEPT9 at the vari-ous DN sub-stages (DN1–DN4), leukocytes were stained withanti-CD25 and anti-CD44 in combination with anti-CD4 andanti-CD8 (Fig. 2d, e). Because the size of the DN populationvaries between Wt and Sept9:LckCre mice, we analyzed thecells of the DN1-4 stages within the DN cell gate (percentageof gate) in order to reveal whether the distribution of the cellsin different substages of the DN stage changes irrespectivelyof the variation in the DN population. This revealed not onlyan increase of cells in DN3, but also a decrease of the DN4population in Sept9:LckCre mice compared with Wt micesuggesting a functional requirement for SEPT9 at the DN3-DN4 transition (Fig. 2f).
Mature T-cells are downregulated in Sept9:LckCre mice
To investigate whether the reduced development from DN3through DN4 to the DP stage has consequences for peripheral
Cell Tissue Res (2013) 352:695–705 697
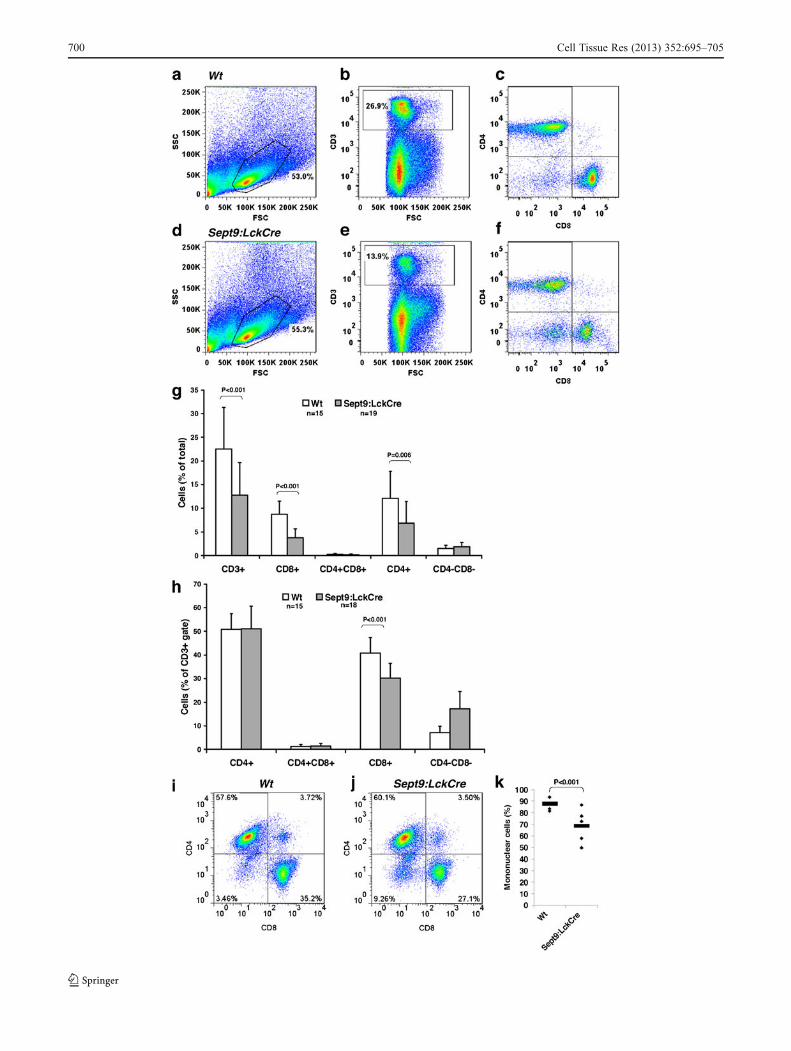
T-cells, we compared various mature T-cell populations of thespleen from adult mice (Fig. 3a-f). This revealed a generalreduction to 57% of T-cells (CD3+) in Sept9:LckCre micecompared with Wt. Likewise, we found a reduction of bothmature regulatory CD4+ T-cells and of cytotoxic CD8+ T-cellsin the spleen of Sept9:LckCre mice compared with Wt (bothpopulations also CD3+). The reduction was more prominent inCD8+ T-cells (down to 44%) compared with CD4+ T-cells(down to 57%) (Fig. 3g). Consistent with this, only CD8+
T-cells were reduced within the CD3+ T-cell gate (Fig. 3h).Compared with Wt mice, the reduction of single-positive
CD4+ and CD8+ T-cells were most prominent in youngSept9:LckCre mice (40 days) in which mature single-positive CD4+ and CD8+ T-cells in the spleen were reducedto 29.6% (from 5.4% of the total leukocyte population[±2.0, n=3] to 1.6% [±0.4, n=3]) and 21.8% (from 6.4%[±2.6, n=3] to 1.4% [±0.1, n=3]), respectively. In olderSept9:LckCre mice (60 days), this reduction compared with
Wt was only down to 66.3% (from 9.5% [±4.4, n=4] to 6.3%[±2.2, n=3]) and 50.0% (from 6.8% [±2.2, n=4]) to 3.4%[±1.4, n=3]) in CD4+ and CD8+ populations, respectively,indicating that the partial developmental block at the DN stageis mostly visible at the peak of T-cell development.
In lymph node tissue pooled from three Sept9:LckCreand three Wt mice (40 days old), respectively, the numberof CD3+ T-cells was reduced to a level of 80% comparedwith Wt. Again, this effect was stronger in CD8+ T-cells(reduced to 62%) compared with CD4+ T-cells (reduced to84%). Interestingly, in the lymph node, the population ofDN cells (CD3+CD4-CD8-), which in Wt mice constituted2.5% of the total cell population, was also increased to 5.4%in Sept9:LckCre mice (Fig. 3i, j). Whether this increase ofDN cells in lymph nodes is related to the increase of DNcells in the thymus of Sept9:LckCre mice remains to beinvestigated. However, the decrease of mature T-cells (bothCD8+ and CD4+) in Sept9:LckCre mice and the shift in the
Fig. 1 Relative Sept9 expression measured by quantitative real-timepolymerase chain reaction. Beta-2-microglobulin (b2m) is used as aninternal reference. Individual mice are shown as dots to indicate thevariability in the deletion efficiency between mice (bar mean). aThymus leukocytes. b Spleen leukocytes. Sept9 is highly expressedin wild-type (Wt) thymus compared with Wt spleen (note the differentscales). In thymus, Sept9 expression is on average decreased to 8% inSept9:LckCre mice (n=3) compared with Wt mice (n=6). In spleen,Sept9 expression is on average decreased to 68% in Sept9:LckCre mice(n=5) compared with Wt mice (n=6). c T-cell populations of thethymus. Sept9 deletion starts at the double-negative (DN) stage (Sept9expression decreased to 26% of Wt [n=6] in Sept9:LckCre mice [n=5])
but is more efficient at the double-positive stage (Sept9 expressiondown to 9% of Wt [n=6] in Sept9:LckCre mice [n=5]) and the single-positive stages (Sept9 expression down to 8% of Wt [n=6] in bothCD4+ cells [n=6] and CD8+ cells [n=4]) of Sept9:LckCre mice. Sept9expression is increased in the Wt double-positive cells (n=6) comparedwith Wt DN and single-positive cells. d DN T-cell populations ofthymus. Sept9 expression is decreased as early as the DN1 stage. Fourmice were analyzed within each group. e Mature T-cells of the spleen.Dotted lines connect data from identical mice; these data suggest thatthe high values of Sept9 expression found in the two mice are attributableto inefficient CRE recombinase activity. Six and five mice of eachgenotype were analyzed for, respectively, CD4+ and CD8+ cells
698 Cell Tissue Res (2013) 352:695–705
distribution of CD8+ versus CD4+ T-cells demonstrate ageneral alteration of mature T-cells in the periphery as aconsequence of Sept9 deletion in early T-cells.
In order to test whether the reduction of T-cells is alsofound in the blood, we performed staining of blood smearsfrom 10- to 12-month-old mice. Such staining showed, inSept9:LckCremice, a decrease (down to 78%) of the relativenumber of mononuclear cells (which included T-cells) com-pared with granulocytic cells, which were used as an inter-nal standard (Fig. 3k). Even though these counts do notreveal the nature of the missing cells, the data are consistentwith those that we obtained by FACS analysis in otherperipheral tissues.
In conclusion, our data show that Sept9 deletion in T-cellsleads to a reduction of mature T-cells in the periphery.
To investigate further whether SEPT9 is only transientlyrequired for T-cell development in young mice, we crossedSept9cond mice with the transgenic mice expressing CRErecombinase under the control of the interferon-responsiveMx promoter, which can be induced in bone marrow andlymphoid tissue upon injection of polyI polyC (here denotedSept9:MxCre). Analysis of mice, injected at the age of6 weeks and exhibiting a Sept9 deletion efficiency of99.4% in thymus, 96% in spleen, and 98.9% in bone mar-row, showed no difference between Sept9:MxCre and Wtmice in the number of T-cells in the spleen (CD3+:Wt 12.7%
Fig. 2 Increased numbers of DN cells in the thymus of adultSept9:LckCre mice. T-cell populations in the thymus measured byfluorescence-activated cell sorting (FACS) analysis. a, b Representa-tive FACS data plots of thymus from a Wt mouse (a) and aSept9:LckCre mouse (b) showing gates used to detect lymphocytes(within the forward scatter channel [FSC]—side scatter channel [SSC]plot), which were analyzed for the expression of CD4 and CD8. cHistogram showing the average sizes of T-cell populations of 18 Wtand 15 Sept9:LckCre mice. DN cells (CD4-CD8-) are increased inSept9:LckCre mice compared with Wt. Note that the percentage of
cells refers to the entire leukocyte population and therefore does notadd up to 100%. d, e Percentage of cells within the subpopulations ofDN early T-cells (DN1, DN2, DN3, and DN4). Representative FACSdata plots of thymus from a Wt mouse (d) and a Sept9:LckCre mouse(e). DN cells (shown in a) are gated for the expression of CD25 andCD44. f Histogram showing the average sizes of the DN subpopula-tions within the DN gate for seven Wt and six Sept9:LckCre mice.Compared with Wt, DN3 (CD4-CD8-CD25+CD44-) cells are in-creased, whereas DN4 (CD4-CD8-CD25-CD44-) cells are decreasedwithin the CD4-CD8- gate in Sept9:LckCre mice
Cell Tissue Res (2013) 352:695–705 699
700 Cell Tissue Res (2013) 352:695–705
of the total leukocyte population [±4.2, n=2], Sept9:MxCre12.3% [±0.2, n=3]; CD4+:Wt 7.1% [±1.7, n=2], Sept9:MxCre7.9% [±1.3, n=3]; CD8+:Wt 4.6% [±2.2, n=2], Sept9:MxCre3.6% [±0.4, n=3]) or thymus (DN:Wt 2.5% [±1.3, n=2],Sept9:MxCre 2.5% [±0.2, n=3]). In bone marrow, the CD3+
population (Wt 3.4% [±0.7, n=2], Sept9:MxCre 2.8% [±1.0,n=3]) and the CD4+ subpopulation (Wt 0.6% [±0.2, n=2],Sept9:MxCre 0.5% [±0.1, n=3]) were unaffected by the lossof Sept9, but the CD8+ subpopulation was decreased to 61.5%(from Wt 1.3% [±0.1, n=2], Sept9:MxCre 0.8% [±0.04, n=3],P=0.04) emphasizing the importance of Sept9 for CD8+
T-cells (Fig. 4). We likewise checked the level of B-cells (bystaining with CD19) and the level of myeloid cells (bystaining with CD11b) and found no differences betweenSept9:MxCre and Wt mice. The mice were analyzed 100 daysafter the induction of CRE recombinase expression. Thissuggests that, if T-cells have developed normally during thefirst 6 weeks of life, the normal T-cell development in thymusand turn-over of T-cells in spleen is Sept9-independent,whereas the minor CD8+ population in bone marrow seemsto be affected by the deletion of Sept9. Whether the lower rateof T-cell development at this age can explain the Sept9-inde-pendent development remains unanswered.
To analyze whether the gene dose of Sept9 is critical forproper T-cell development, we analyzed thymus and spleenfrom Sept9neo/+ mice (n=3), which have only one functionalSept9 allele, at age 32 days. We found Sept9 expression levelsto be approximately 50% compared with Wt (n=3). Flowanalysis revealed no phenotypic differences in the number ofT-cells in thymus, spleen, or lymph node indicating that 50% ofSept9 expression is sufficient for proper T-cell development.
Upregulation of central memory CD8+ T-cells
In order to analyze the capacity of Sept9-deleted T-cells todevelop into other subtypes of T-cells, we performed FACS
�Fig. 3 Decreased mature T-cell populations in the periphery of adultSept9:LckCre mice. a-f T-cell populations in the spleen measured byFACS analysis. Representative FACS data plots of spleen from a Wtmouse (a-c) and a Sept9:LckCre mouse (d–f). Lymphocytes are gatedwithin the FSC—SSC—plot (a, d) and then CD3-positive cells areselected (b, e). These cells are then analyzed for the expression of CD4and CD8 (c, f). The lymphocyte populations are not significantlydifferent between Wt and Sept9:LckCre mice, but the CD3+ populationis smaller in Sept9:LckCre mice compared with Wt mice. Within theCD3+ population, only the CD8+ population is significantly smaller inSept9:LckCre mice compared with Wt mice. g T-cell populations in thespleen measured by FACS analysis. The average percentage of cells inthe total leukocyte population within the CD3+, the CD3+CD4+, theCD3+CD8+, the CD3+CD4+CD8+, and the CD3+CD4-CD8-
populations from 15 Wt mice and 19 Sept9:LckCre mice measuredby FACS analysis as shown in a–f. CD3+, CD3+CD4+, and CD3+CD8+
cells are decreased in Sept9:LckCre compared with Wt. h T-cellpopulations in the spleen measured by FACS analysis. Average per-centage of CD4+, CD8+, CD4+CD8+, and CD4-CD8- T-cells within theCD3+ population from 15Wt and 18 Sept9:LckCremice. Compared withWt, CD8+ T-cells are decreased among CD3+ T-cells in Sept9:LckCremice. i, j CD4/CD8 T-cell distribution within the CD3+ gate of lymphnode cells measured by FACS analysis. Lymph nodes from, respectively,three Wt mice (i) and three Sept9:LckCre mice (j) were pooled. CD8+ T-cells are decreased, and CD4-CD8- T-cells are increased in Sept9:LckCremice compared with Wt mice. k Analysis of blood smears fromSept9:LckCre and Wt mice. Mononuclear and granular cells were count-ed, and the percentages of mononuclear cells were calculated. Fewermononuclear cells were found in Sept9:LckCre mice compared with Wtindicating a reduced number of T-cells (dots individual mice). FiveSept9:LckCre and four Wt mice were analyzed
Fig. 4 Most T-cell populations were unaffected by Sept9 loss at theage of 6 weeks, but the CD8+ T-cell population from the bone marrowwas decreased. a, b Representative FACS data plots of a Wt mouse (a)and a Sept9:MxCremouse (b). Bone marrow cells were gated as shownfor splenocytes in Fig. 3a. In the Wt mouse, the CD8+ T-cell proportionconstitutes 1.3% of the total leukocyte population, whereas only 0.9%
CD8+ T-cells were found in the Sept9:MxCre mouse. c Average per-centage of CD8+ T-cells in the total leukocyte population from bonemarrow of two Wt and three Sept9:MxCre mice. The numbers of CD8+
T-cells are statistically significantly decreased in Sept9:MxCre micecompared with Wt mice
Cell Tissue Res (2013) 352:695–705 701
analysis of spleen from adult Sept9:LckCre mice with anti-CD44 and anti-CD62L antibodies in combination with anti-CD3, anti-CD4, and anti-CD8 . Within the CD8+ T-cellpopulation, we observed an increase in the number of cellspositive for CD44 and CD62L, which have been describedas central memory cells (Ammirati et al. 2012; Laouar et al.2008; Fig. 5a, b).
In accordance with the reduced number of CD3+ T-cellsin Sept9:LckCre mice, we observed reduced absolute num-bers of CD4+ and CD8+ central memory cells (Fig. 5c).However, when analyzed within the CD4+ or CD8+ cellgate, respectively, the number of CD8+ (but not CD4+)central memory cells was significantly increased to 183%(P<0.001) of that found in Wt (Fig. 5d).
To exclude the possibility that the long-living centralmemory T-cells originate from an expansion of a few T-cellsthat might have escaped the CRE-recombinase-mediateddeletion, we analyzed Sept9 expression in these cells andfound Sept9 expression decreased to 0.4% and 3.10% in,respectively, CD4+ and CD8+ central memory T-cells ofSept9:LckCre mice.
Proliferation capacity is decreased in Sept9-depleted CD8+
T-cells
The decrease in mature T-cells in the periphery ofSept9:LckCre mice compared with Wt mice and the shiftin distribution of T-cells (fewer CD8+ compared with CD4+
T-cells), together with the finding that SEPT9 is importantfor cytokinesis (Füchtbauer et al. 2011; Estey et al. 2010)led us to test the proliferative capacity of Sept9-deleted T-cells compared to Wt T-cells. We cultured CFSE-labeledspleen leukocytes for 5 days. The loss of the otherwisestable membrane label is a direct measure of cell division.After stimulation with anti-CD3 and anti-CD28 antibodies,subsequent FACS analysis revealed a general decrease (downto 41%) of living lymphocytes in cells isolated fromSept9:LckCre mice (based on PI staining and the size andgranularity of the cells) compared with cells isolated from Wtmice (Fig. 6a, d, g). Again, CD8+ T-cells were most affectedby the loss of Sept9, as these cells were decreased to 20% ofWt (Fig. 6b, e, g). Within the lymphocyte gate, Sept9-deletedCD8+ T-cells were on average decreased to a level of 42%compared with Wt (Fig. 6b, e). As in vivo, Sept9-deletedCD4+ T-cells were decreased compared with Wt (Fig. 6g),and Sept9-deleted CD8+ central memory T-cells were in-creased within the CD8+ gate compared with Wt (not shown),although neither effect was statistically significant.
To standardize the relative loss of the CFSE label, weanalyzed the CD8+ T-cells by FACS analysis and determinedthe percentage of cells that were labeled more strongly thanthe maximum of the CFSE histogram (gate represented inFig. 6c, f). CFSE staining was found to decrease in allpopulations of both Sept9:LckCre and Wt mice comparedwith non-stimulated cells, but we found Sept9:LckCre miceto have a higher level of CD8+ CFSE-positive cells comparedwith Wt upon stimulation as a result of reduced proliferationcapacity (Fig. 6h).
Discussion
We have analyzed the development of T-cells in the absenceof Sept9. We have found that the Sept9 deletion causes T-cell development to be partially inhibited at the DN stage,more specifically at the DN3 stage, when we see an accu-mulation of cells.
Fig. 5 Increased CD8+ central memory T-cell population inSept9:LckCre mice. CD4+ and CD8+ central memory T-cells withinspleen of Sept9:LckCre and Wt mice measured by FACS analysis. a, bRepresentative FACS data plot showing the CD8+ gate from Fig. 3c, fanalyzed for the expression of CD44 and CD62L from a Wt mouse (a)and a Sept9:LckCre mouse (b). Percentages of central memory cellswithin the CD8+ population are indicated. c Percentage of CD4+ andCD8+ central memory T-cells in the total leukocyte population. CD4+
central memory cells were analyzed as shown for CD8+ central memorycells in a, b. The numbers of central memory cells are not significantlydifferent between Sept9:LckCre and Wt mice. d Average percentage ofCD4+ and CD8+ central memory T-cells within the CD4+ and the CD8+
gate, respectively, from fiveWt and six Sept9:LckCremice. CD8+ centralmemory T-cells (CD3+CD8+CD44+CD62L+) are increased inSept9:LckCre mice compared with Wt as also seen in a, b
702 Cell Tissue Res (2013) 352:695–705
We have also found the Sept9:LckCre transgene is activeas early as the DN1 stage, a finding that raises the possibilitythat the change in population size at the transition from DN3to DN4 represents a functional requirement for SEPT9 rath-er than the onset of Sept9 deletion. However, we know littleabout the stability of the SEPT9 protein in DN T-cells, as
this protein is difficult to quantify in the minute amounts ofmaterial available after FACS analysis. The massiveupregulation of Sept9 at the DP stage might also reflect afunctional role for SEPT9, and although not statisticallysignificant, Sept9 expression increases in Wt mice as earlyas the DN3 and DN4 stages (compared with DN1 and DN2),
Fig. 6 Altered CD8+ T-cell proliferation rate in Sept9:LckCre mice.Distribution of T-cells from Sept9:LckCre and Wt mice stimulated withanti-CD3 and anti-CD28 and grown in culture for 5 days measured byFACS analysis. a–f FACS data plots from a representative culture froma Wt mouse (a-c) and a Sept9:LckCre mouse (d–f). The lymphocytegate (a, d) is set on the basis of a separate run including propidiumiodide to exclude dead cells. Fewer live lymphocytes are found in theSept9:LckCre culture compared with Wt. Within the lymphocyte gate,the expression of CD4 and CD8 were analyzed (b, e). Fewer CD8+
cells were found within the lymphocyte gate of the Sept9:LckCreculture compared with Wt. Within the CD8 gate shown in b, e, thecells were analyzed for the intensity of carboxyfluorescein diacetatesuccinimidyl ester (CFSE; c, f). The CFSE-positive cells are defined as
cells with higher CFSE labeling than the maximum of the CFSEhistogram. A higher proportion of CFSE-positive cells were found inthe Sept9:LckCre culture compared with Wt indicating a lower rate ofproliferation. g Average percentage of live cells of the total leucocytepopulation within the lymphocyte, CD8+ T-cell, CD4+ T-cell, CD8+
central memory T-cell, and CD4+ central memory T-cell gate fromeight Wt and five Sept9:LckCre cultures analyzed as shown in a, b,d, e and Fig. 5 a, b. Lymphocytes and CD8+ T-cells are decreased inSept9:LckCre cultures compared with Wt. h Average of CFSE-positivecells defined by the gates shown in c, f from five Wt and eightSept9:LckCre mice. Sept9-deleted cells have a significantly higherlevel of CFSE-positive cells, because they proliferate to a lower extentthan Wt cells
Cell Tissue Res (2013) 352:695–705 703
in agreement with a role for Sept9 in the DN3-DN4-DPtransition.
After T-cell-specific deletion of Sept9, the number of ma-ture T-cells markedly decreases, and in particular, the CD8+
T-cell population is affected causing a shift in the CD4+/CD8+
T-cell distribution. These changes are less pronounced in olderanimals indicating that T-cells deleted for Sept9 do not have adecreased survival potential. This interpretation is supportedby the finding that the deletion of Sept9 by using aMx-drivenCre transgene at the age of 6 weeks when the majority of themature T-cells have developed, does not change the size of theT-cell population in the spleen.
Lymphopenia-induced proliferation has been shown toincrease the number of central memory T-cells (Takadaand Jameson 2009). We expect this phenomenon to explainthe increase in CD8+ central memory T-cells, which thuswould be a secondary event arising from the reduced CD8+
T-cell population in Sept9:LckCre mice. The reduced pro-liferation capacity of Sept9-deleted CD8+ T-cells might bethe reason that these cells are more affected by the reducedtransition from DN3 to DN4 compared with CD4+ T-cells.Assuming that SEPT9 is important for the proliferation in T-cells in general, the stronger effect on CD8+ T-cells com-pared with CD4+ T-cells upon Sept9 deletion might be theconsequence of the higher proliferation level of CD8+ T-cells. The proliferative capacity of CD8+ T-cells is reportedto be greater than that of CD4+ T-cells, both upon anti-CD3and anti-CD28 stimulation in culture and in the response toseveral kinds of pathogens (Foulds et al. 2002). We confirmthis in our studies, as we find higher percentages of prolif-erative cells (CFSE-negative) in the Wt CD8+ T-cell popu-lation compared with the Wt CD4+ T-cell population (datanot shown). In Sept9:LckCre mice, the growth of CD4+ T-cells upon stimulation in culture is decreased compared withthat in Wt mice, although not at a statistically significantlevel (Fig. 6g), a result that is possibly explained by thedecreased proliferation capacity of CD4+ T-cells comparedwith CD8+ T-cells in general.
Earlier studies of septin function in T-cells have beenmainly performed in the CD4+-derived D10.G4 cell line(Tooley et al. 2009); this might explain the minor effect seenupon deletion of Sept9. However, the need for SEPT9 specif-ically during T-cell development or redundancy from otherseptins might compensate for the loss of SEPT9 in these cells.
Several functional effects of SEPT9 could be importantin T-cell development. SEPT7 is involved in T-cell migra-tion, which is crucial for T-cell development in the thymus(Tooley et al. 2009), and SEPT9 is vital for SEPT7 filamentformation, at least in embryonic fibroblasts (Füchtbauer etal. 2011). Furthermore, SEPT9 and other septins are in-volved in autophagy (Mostowy et al. 2010), a process highlyrelevant in T-cells (Crotzer and Blum 2009). Moreover, actinand septin filaments interact in a still not fully understood way
(Kinoshita et al. 2002), and the loss of Sept9 in embryonicfibroblasts reduces actin filaments and actin-associated focalcontacts (Füchtbauer et al. 2011). Actin filaments are impor-tant for the activation of the T-cell receptor (Billadeau et al.2007), which is an important step in T-cell development; thus,we are tempted to speculate that the septin-actin interactioninfluences T-cell development.
In conclusion, we show here, for the first time, a phenotypicdefect upon Sept9 deletion in T-cell development. Despite theeffects seen in T-cell numbers, the mice are viable and do notsuffer from disease probably because T-cell development is notcompletely blocked. The lack of a complete inhibition of T-celldevelopment might be caused by redundancy from the tworemaining members of the SEPT3 group or from other septinsthat might functionally compensate for the loss of Sept9. Septinresearch is, in general, made difficult by complex and not yetfully understood interactions. However, specific functions ofindividual septins are established, for example, by the death ofembryos deprived of Sept7, Sept9, or Sept11 (Ware et al. 2008;Hall et al. 2008; Füchtbauer et al. 2011; Roseler et al. 2011),although the reason for the embryonic lethality is still elusive.Other knock out mouse models of septins are viable but sufferfrom sterility (Ihara et al. 2005) and abnormalities in the brain(Ihara et al. 2007; Sept4-/-) or elevated platelet sensitivity (Dentet al. 2002) and affective behavior (Suzuki et al. 2009;Sept5−/−) indicating a widespread function of the septins.
Acknowledgments The authors thank Lone Højgaard Nielsen fortechnical assistance, and Charlotte Christie Petersen, Rodrigo Labouriauand Bo Porse for advice regarding the set-up and analysis of experiments.Flow cytometry/cell sorting was performed at the FACS Core Facility,The Faculty of Health Sciences, Aarhus University, Denmark.
References
Ammirati E, Cianflone D, Vecchio V, Banfi M, Vermi AC, De MetrioM, Grigore L, Pellegatta F, Pirillo A, Garlaschelli K, ManfrediAA, Catapano AL, Maseri A, Palini AG, Norata GD (2012)Effector memory T cells are associated with atherosclerosis inhumans and animal models. J Am Heart Assoc 1:27–41
Billadeau DD, Nolz JC, Gomez TS (2007) Regulation of T-cell acti-vation by the cytoskeleton. Nat Rev Immunol 7:131–143
Crotzer VL, Blum JS (2009) Autophagy and its role in MHC-mediatedantigen presentation. J Immunol 182:3335–3341
Dent J, Kato K, Peng XR, Martinez C, Cattaneo M, Poujol C, NurdenP, Nurden A, Trimble WS, Ware J (2002) A prototypic plateletseptin and its participation in secretion. Proc Natl Acad Sci USA99:3064–3069
Estey MP, Di Ciano-Oliveira C, Froese CD, Bejide MT, Trimble WS(2010) Distinct roles of septins in cytokinesis: SEPT9 mediatesmidbody abscission. J Cell Biol 191:741–749
Foulds KE, Zenewicz LA, Shedlock DJ, Jiang J, Troy AE, Shen H(2002) Cutting edge: CD4 and CD8 T cells are intrinsicallydifferent in their proliferative responses. J Immunol 168:1528–1532
704 Cell Tissue Res (2013) 352:695–705
Füchtbauer A, Lassen LB, Jensen AB, Howard J, Quiroga Ade S,Warming S, Sørensen AB, Pedersen FS, Füchtbauer EM (2011)Septin9 is involved in septin filament formation and cellularstability. Biol Chem 392:769–777
Hall PA, Russell SE (2004) The pathobiology of the septin gene family.J Pathol 204:489–505
Hall PA, Russell SE (2012) Mammalian septins: dynamic heteromerswith roles in cellular morphogenesis and compartmentalization. JPathol 226:287–299
Hall PA, Jung K, Hillan KJ, Russell SE (2005) Expression profiling thehuman septin gene family. J Pathol 206:269–278
Hall PA, Russell SEH, Pringle JR (2008) The septins. Wiley, New YorkHartwell LH (1971) Genetic control of the cell division cycle in yeast.
IV. Genes controlling bud emergence and cytokinesis. Exp CellRes 69:265–276
Hu Q, Milenkovic L, Jin H, Scott MP, Nachury MV, Spiliotis ET,Nelson WJ (2010) A septin diffusion barrier at the base of theprimary cilium maintains ciliary membrane protein distribution.Science 329:436–439
Ihara M, Kinoshita A, Yamada S, Tanaka H, Tanigaki A, Kitano A, GotoM, Okubo K, Nishiyama H, Ogawa O, Takahashi C, Itohara S,Nishimune Y, Noda M, Kinoshita M (2005) Cortical organizationby the septin cytoskeleton is essential for structural and mechanicalintegrity of mammalian spermatozoa. Dev Cell 8:343–352
Ihara M, Yamasaki N, Hagiwara A, Tanigaki A, Kitano A, Hikawa R,Tomimoto H, Noda M, Takanashi M, Mori H, Hattori N,Miyakawa T, Kinoshita M (2007) Sept4, a component of presyn-aptic scaffold and Lewy bodies, is required for the suppression ofalpha-synuclein neurotoxicity. Neuron 53:519–533
KimMS, Froese CD, Estey MP, Trimble WS (2011) SEPT9 occupies theterminal positions in septin octamers and mediates polymerization-dependent functions in abscission. J Cell Biol 195:815–826
Kinoshita M, Field CM, Coughlin ML, Straight AF, Mitchison TJ(2002) Self- and actin-templated assembly of mammalian septins.Dev Cell 3:791–802
Kuhlenbaumer G, Hannibal MC, Nelis E, Schirmacher A, VerpoortenN, Meuleman J, Watts GD, De Vriendt E, Young P, Stogbauer F,Halfter H, Irobi J, Goossens D, Del-Favero J, Betz BG, Hor H,Kurlemann G, Bird TD, Airaksinen E, Mononen T, Serradell AP,Prats JM, Van Broeckhoven C, De Jonghe P, Timmerman V,Ringelstein EB, Chance PF (2005) Mutations in SEPT9 causehereditary neuralgic amyotrophy. Nat Genet 37:1044–1046
Kuhn R, Schwenk F, Aguet M, Rajewsky K (1995) Inducible genetargeting in mice. Science 269:1427–1429
Laouar A, Manocha M, Haridas V, Manjunath N (2008) Concurrentgeneration of effector and central memory CD8 T cells duringvaccinia virus infection. PLoS One 3:e4089
McMurray MA, Thorner J (2009) Reuse, replace, recycle. Specificityin subunit inheritance and assembly of higher-order septin struc-tures during mitotic and meiotic division in budding yeast. CellCycle 8:195–203
Mostowy S, Cossart P (2012) Septins: the fourth component of thecytoskeleton. Nat Rev Mol Cell Biol 13:183–194
Mostowy S, Bonazzi M, Hamon MA, Tham TN, Mallet A, Lelek M,Gouin E, Demangel C, Brosch R, Zimmer C, Sartori A, KinoshitaM, Lecuit M, Cossart P (2010) Entrapment of intracytosolicbacteria by septin cage-like structures. Cell Host Microbe8:433–444
Orban PC, Chui D, Marth JD (1992) Tissue- and site-specific DNArecombination in transgenic mice. Proc Natl Acad Sci USA89:6861–6865
Roseler S, Sandrock K, Bartsch I, Busse A, Omran H, Loges NT,Zieger B (2011) Lethal phenotype of mice carrying a Sept11 nullmutation. Biol Chem 392:779–781
Russell SE, Hall PA (2011) Septin genomics: a road less travelled. BiolChem 392:763–767
Sellin ME, Holmfeldt P, Stenmark S, Gullberg M (2011) Microtubulessupport a disk-like septin arrangement at the plasma membrane ofmammalian cells. Mol Biol Cell 22:4588–4601
Sirajuddin M, Farkasovsky M, Hauer F, Kuhlmann D, Macara IG,Weyand M, Stark H, Wittinghofer A (2007) Structural insightinto filament formation by mammalian septins. Nature 449:311–315
Sørensen AB, Lund AH, Ethelberg S, Copeland NG, Jenkins NA,Pedersen FS (2000) Sint1, a common integration site in SL3-3-induced T-cell lymphomas, harbors a putative proto-oncogenewith homology to the septin gene family. J Virol 74:2161–2168
Suzuki G, Harper KM, Hiramoto T, Sawamura T, Lee M, Kang G,Tanigaki K, Buell M, Geyer MA, Trimble WS, Agatsuma S, HiroiN (2009) Sept5 deficiency exerts pleiotropic influence on affec-tive behaviors and cognitive functions in mice. Hum Mol Genet18:1652–1660
Takada K, Jameson SC (2009) Naive T cell homeostasis: from aware-ness of space to a sense of place. Nat Rev Immunol 9:823–832
Tooley AJ, Gilden J, Jacobelli J, Beemiller P, Trimble WS, KinoshitaM, Krummel MF (2009) Amoeboid T lymphocytes require theseptin cytoskeleton for cortical integrity and persistent motility.Nat Cell Biol 11:17–26
Ware J, Martínez C, Zieger B (2008) Septins and platelets. In: Hall PA,Russell SEH, Pringle JR (eds)The septins. Wiley, New York,pp 269–280