Page 1
© 2020. Published by The Company of Biologists Ltd.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License
(http://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution and reproduction
in any medium provided that the original work is properly attributed.
Silencing of CCR4-NOT complex subunits affect heart structure and
function
Lisa Elmén1 PhD, Claudia B. Volpato2 PhD, Anaïs Kervadec1 PhD, Santiago Pineda1,# PhD,
Sreehari Kalvakuri1, PhD, Nakissa N. Alayari1+ MS, Luisa Foco2 PhD, Peter P. Pramstaller2
MD, Karen Ocorr1, PhD, Alessandra Rossini2 PhD, Anthony Cammarato3 PhD, Alexandre R.
Colas1 PhD, Andrew A. Hicks2* PhD, Rolf Bodmer1,* PhD
1 Development Aging and Regeneration Program, Sanford Burnham Prebys Medical
Discovery Institute, 10901 N Torrey Pines Rd, La Jolla, CA 92037, USA.
2 Institute for Biomedicine, Eurac Research, Affiliated Institute of the University of Lübeck,
Bolzano, Italy
3 Johns Hopkins University, Division of Cardiology, 720 Rutland Ave., Baltimore, MD 21205,
USA.
# Present address: University of California Los Angeles, Molecular, Cell, and Developmental
Biology, Terasaki Life Sciences Building, 610 Charles E. Young Dr. South, CA 90095, USA.
+ Present address: Illumina Inc., 5200 Illumina Way, San Diego CA 92122, USA.
* Corresponding authors: Rolf Bodmer, PhD, Development Aging and Regeneration
Program, Sanford Burnham Prebys Medical Discovery Institute, 10901 North Torrey Pines
Road, La Jolla, CA 92037. Telephone: (858) 795-5295. E-mail: [email protected] .
Andrew A. Hicks, PhD, Institute for Biomedicine, Eurac Research, Via Luigi Galvani 31,
Bolzano, 39100, Italy. Telephone +39 3347788721. E-mail: [email protected] .
Summary statement
This work demonstrates the successful approach of combining GWAS studies with in vitro
human cell assays and a suitable in vivo model organism, and clearly connects CNOT1,
CNOT7 and overall, the CCR4-NOT complex, to heart morbidity.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
http://dmm.biologists.org/lookup/doi/10.1242/dmm.044727Access the most recent version at First posted online on 29 May 2020 as 10.1242/dmm.044727
Page 2
ABSTRACT
Genome wide association studies (GWAS) have identified variants that associate with
QT-interval length. Three of the strongest associating variants (SNPs) are located in the
putative promotor region of CNOT1, a gene encoding the central subunit of CCR4-NOT, a
multi-functional, conserved complex regulating gene expression and mRNA stability and
turnover. We isolated the minimum fragment of the CNOT1 promoter containing all three
variants from individuals homozygous for the QT-risk alleles and demonstrated that the
haplotype associating with longer QT-interval caused reduced reporter expression in a cardiac
cell line, suggesting that reduced CNOT1 expression may contribute to abnormal QT-intervals.
Systematic siRNA-mediated knockdown of CCR4-NOT components in human induced
pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs) revealed that silencing CNOT1
and other CCR4-CNOT genes reduced their proliferative capacity. Silencing CNOT7 also
shortened action potential duration. Furthermore, cardiac-specific knockdown of Drosophila
orthologs of CCR4-NOT genes, CNOT1/not1 and CNOT7/8/pop2, in vivo, was either lethal or
resulted in dilated cardiomyopathy, reduced contractility, or a propensity for arrhythmia.
Silencing CNOT2/not2, CNOT4/not4 and CNOT6/6L/twin also affected cardiac chamber size
and contractility. Developmental studies suggested that CNOT1/not1 and CNOT7/8/pop2 are
required during cardiac remodeling from larval to adult stages. In sum, we have demonstrated
how disease associated genes identified by GWAS can be investigated, by combining human
cardiomyocyte cell-based and whole organism in vivo heart models. Our results also suggest
a potential link of CNOT1 and CNOT7/8 to QT alterations and further establish a critical role
of the CCR4-NOT complex in heart development and function.
Key words: CNOT1, GWAS, arrhythmia, long QT-syndrome, Drosophila heart, human iPSC,
cardiomyocytes.
D
isea
se M
ode
ls &
Mec
hani
sms
• D
MM
• A
ccep
ted
man
uscr
ipt
Page 3
INTRODUCTION
Despite the medical advances that have been made over the last decades,
cardiovascular disease remains the most common cause of mortality worldwide (WHO, 2018).
Understanding the mechanisms of heart morbidity is crucial for finding new therapies, and
determining which genetic variants predispose individuals to heart disease is necessary to
provide better preventative care. The challenge of connecting human genetic variants with
disease can be met by combining Genome Wide Association Studies (GWAS) with patient
sequencing and validation using disease-in-a-dish and in vivo cardiac model systems.
Drosophila melanogaster benefits from well-conserved genes and permits functional
assessment of genes of interest, which when manipulated may not be well-tolerated by the
vertebrate heart.
The QT interval on an electrocardiogram is a measure that reflects myocardial
repolarization. Short-QT syndrome (Rudic et al., 2014) and Long-QT syndrome (Amin et al.,
2013) are caused by different underlying mechanisms but are both risk factors for atrial and
ventricular arrhythmias and sudden cardiac death (Rudic et al., 2014, Amin et al., 2013,
Vacanti et al., 2017). Genome-wide association in up to 100,000 individuals has successfully
identified at least 35 common variant QT interval loci that collectively explain ∼8-10% of QT
variation (Arking et al., 2014). Some of the strongest QT-associating variants identified, center
around the CNOT1 gene which encodes the central scaffolding subunit (CNOT1) of the CCR4-
NOT complex. CCR4-NOT is conserved throughout the eukaryotic kingdoms and is involved
in the sequential processes of gene expression. Its activities can be divided in functional
modules involved in transcription (Kruk et al., 2011) (CNOT2, CNOT3), mRNA-degradation
(Bhandari et al., 2014, Yi et al., 2018, Temme et al., 2010, Webster et al., 2018), deadenylation
(CNOT6/6L, CNOT7/8) and protein quality control via ubiquitination (CNOT4) (Halter et al.,
2014, Collart, 2016, Collart and Panasenko, 2017).
The CCR4-NOT complex has previously been implicated in heart disease; we have
demonstrated that silencing of genes UBC4 and not3 cause cardiac dilation and dysfunction
in Drosophila (Neely et al., 2010). In addition, CNOT3 heterozygous knockout mouse hearts
displayed reduced contractility and increased susceptibility to failure following aortic
constriction (Neely et al., 2010). CNOT3 has also been found to interact with Atg7, which
affects cardiomyocyte (CM) survival and QT intervals in mice (Yamaguchi et al., 2018). In the
present study we investigated the individual role of additional CCR4-NOT complex subunits,
starting with positively QT-interval-associating variants in the CNOT1 putative promoter
region, to determine whether and in what direction they functionally influence reporter gene
expression. We further explored the effects of RNAi-mediated knockdown of CNOT1 and
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 4
complex subunit genes CNOT2, CNOT4, CNOT6/6L and CNOT7/8 in human induced
pluripotent stem cell-derived cardiomyocytes (hiPSC-CMs) (Cunningham et al., 2017, Yu et
al., 2018) on proliferation and electrophysiological properties, in particular action potential
duration (APD), (McKeithan et al., 2017), and in vivo, on cardiac structure and contractile
function using the Drosophila heart model (Ocorr et al., 2014). Overall, we find that silencing
CNOT1 and other CCR4-NOT components compromise cardiac development and function in
the two model systems, suggesting an important role of this complex in cardiac health and
disease.
RESULTS
Functional validation of CNOT1 promoter polymorphisms
Three of the strongest QT-interval associating variants (SNPs), in strong linkage
disequilibrium (LD) over CNOT1, are located in the putative promoter region of the gene (Fig.
1B), redrawn from Fig. S2 in Arking et al. (Arking et al., 2014)). All GWAS associated variants
over the whole length of this gene are in strong LD with the putative promoter variants, and
the GTEx data (https://doi.org/10.1089/bio.2015.0032) indicates strong tissue specific eQTL
variants over the whole gene, leading us to postulate that the promoter variants would
demonstrate functionally different expression levels. A fragment spanning all three of these
strongly QT-associated variants was confirmed by sequencing to contain four SNPs in total
(rs27097, rs37037, rs9941290 and rs863433, all associating strongly with QT interval, apart
from rs37037 which is not in strong linkage disequilibrium with the other three variants, but still
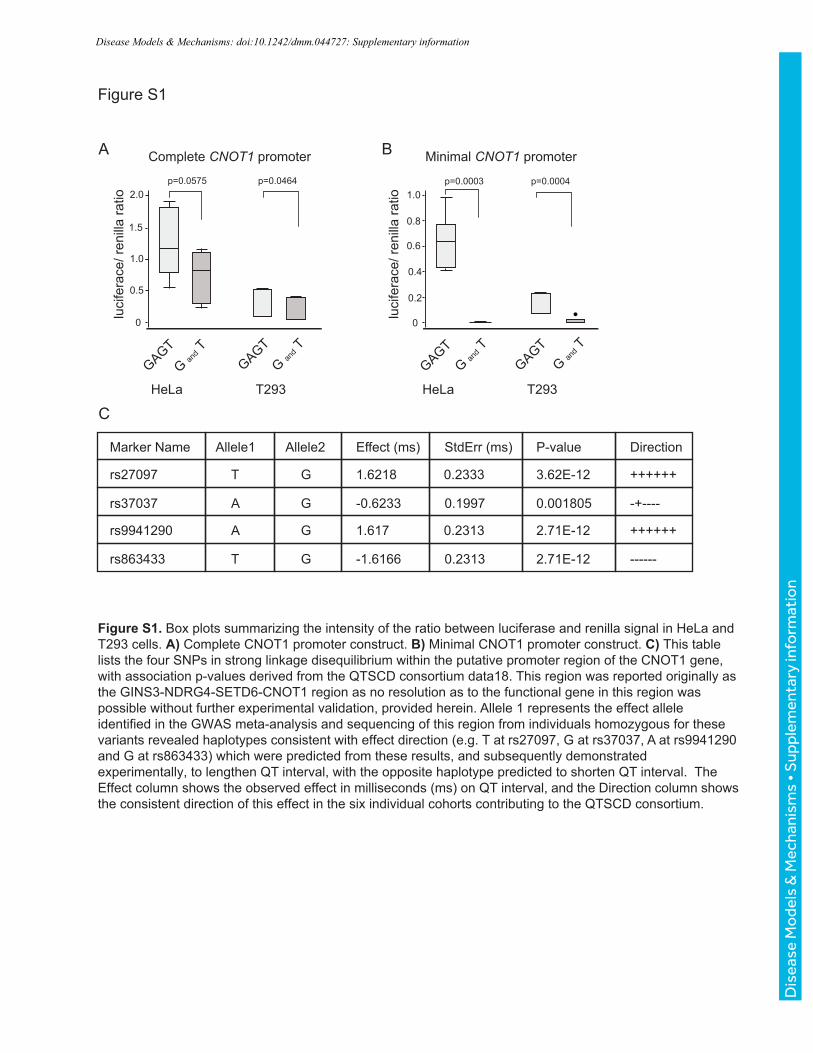
associated although with a less significant p value, Figure S1C). We identified two human
subjects homozygous for alleles at the four SNPs that fall within ~3.2 kb of the 5’ region of the
CNOT1 coding sequence (Fig. 1C). The two haplotypes (one with risk alleles and one with
alternate alleles) from the putative CNOT1 promoter region were isolated and cloned in two
forms into a plasmid vector (pGL4.1) to drive the firefly luciferase gene (Fig. 1D). The “minimal”
putative promoter region contained 657bp of the region around just 1 SNP (rs27097), located
closest to the start codon of the CNOT1 gene. The larger 3172bp fragment contained all of
the strongest QT-associating variants, potentially capturing the “complete” promoter region of
CNOT1 (with respect to the significantly associated variants in this region of the gene). Both
constructs were sequenced to confirm that the four variant positions under study were the only
ones differing between these natural promoter regions, and that the alleles were homozygous.
For both the “minimal” and “complete” constructs, one haplotype consisted of alleles that
significantly associate with increases in QT interval length (“TGAG” haplotype), while the other
haplotype consisted of the alternate alleles at these variants (“GAGT” haplotype) (see also
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 5
Fig. S1C) (Pfeufer et al., 2009). The “minimal” promoter constructs consisted of alleles T and
G respectively at rs27097 (Fig. 1C).
The “complete” and “minimal” promoter-luciferase constructs were transfected into HL-
1 cells, a cardiac muscle cell line that contracts and retains phenotypic characteristics of adult
CM, along with renilla luciferase in order to normalize signals. Both “minimal” and “complete”
promoter constructs were able to drive increased luciferase expression in the HL-1 cells with
significant differences between the two haplotypes. When considering the SNP closest to the
gene alone (“minimal” construct), expression was effectively silenced in the HL-1 cell line for
the haplotype that was associating with longer QT intervals (Fig. 1E). These differences were
also observed in both HeLa and T293 cells (Fig. S1A, B). With the larger promoter fragment,
expression was re-established in all lines, but was significantly reduced for the haplotype
associated with a longer QT compared with the haplotype carrying the alternate alleles. From
these experiments we conclude that the variants which significantly associate with QT interval
in human GWAS, are indeed functionally able to alter expression of the CNOT1 gene in
cardiac tissue, with the most significant differences for the “complete” haplotype being seen
in the cardiac cell line, and that the direction of the effect is such that reduced CNOT1 possibly
contributes to QT interval prolongation.
CCR4-NOT complex genes regulate proliferation and action potential duration in human
iPSC-derived cardiomyocytes (hiPSC-CMs)
CCR4-NOT is a multisubunit complex with different functional modules (including, but
not limited to, the subunits in this study) (Fig. 1A). It has been shown in HeLa and HEK293T
cells that siRNA depletion of CNOT1 decreases the amounts CCR4-NOT subunits and
reduces the complex’ deadenylase activity, and that simultaneous siRNA silencing of the
entire deadenylase module (CNOT6/6L, CNOT7/8) result in apoptosis similar of that of CNOT1
silencing alone (Ito et al., 2011). Therefore, we next decided to use hiPSC-CMs to investigate
the effect of silencing CNOT6/6L and CNOT7/8, genes encoding enzymes that affect
translation efficiency by removing mRNA poly(A) tails (Yi et al., 2018), and CNOT4/not4,
encoding a RING E3-ligase, important for assembly of the proteasome and proposed to be
involved in co-translational quality control (Halter et al., 2014). We also chose to examine
CNOT2, as it associates with CNOT3, for which we previously had identified a role in cardiac
function (Neely et al., 2010).
First, to evaluate a potential role of CCR4-NOT complex on human cardiac physiology,
we knocked down each of the CNOT genes and evaluated their effect on human iPSC-derived
CM (hiPSC-CM) proliferation. Individual knockdown of CNOT1, CNOT2, CNOT3, CNOT4 and
CNOT6 led to decreased EdU incorporation in day 25 hiPSC-CMs and reduced CM number
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 6
as compared to siControl (Fig. 2A-D), thereby suggesting a general role of the CCR4-NOT
complex in the regulation of proliferation in CMs.
A number of studies (Itzhaki et al., 2011, Matsa et al., 2011, Lahti et al., 2012) have
shown that hiPSC-CMs from patients with long QT syndromes consistently show prolonged
action potential duration (APD) phenotypes, thus suggesting that APD modulation in hiPSC-
CMs represent a reliable model system to evaluate the role of candidate genes for QT interval
modulation. Therefore, we asked whether the CCR4-NOT complex could also play such a role
and transfected day 25 hiPSC-CMs with siRNAs directed against each member of the complex
and APD parameters using a fluorescence-based single cell and high-throughput voltage
transient recording assay, based on (McKeithan et al., 2017) (Fig. 2E). Interestingly, we found
that CNOT7, but not CNOT1, knockdown led to a significant shortening (>20ms) of APD (Fig.
2F-I). Although it is possible that the level of CNOT1 knockdown was insufficient to cause a
change in APD, it did produce a proliferation deficit which indicates that the siRNAs were
transfected and active in hiPSC-CMs. This suggests a potential new role for CNOT7 and
deadenylation in the regulation of the QT interval in humans, an observation supported by the
suggestive association of a variant (rs183286310, p = 1.1x10-6) near the CNOT7 gene with
QT interval in ~5000 individuals within the CHRIS study (Pattaro et al., 2015).
Cardiac-specific in vivo knockdown of CNOT1/not1 and CNOT7/8/pop2 in Drosophila
results in dilated cardiomyopathy
Since experiments with hiPSC-CMs provided evidence that CNOT1 and CNOT7
regulate CM proliferation and APD respectively, we asked the question how the same
manipulations would affect the heart in vivo. The CNOT genes are conserved in Drosophila
and have the following orthologs to the human CNOT genes: CNOT2/not2, CNOT4/not4,
CNOT6 and CNOT6L/twin, CNOT7 and CNOT8/pop2 (Table 1).
Using the Drosophila UAS–Gal4 system (Brand and Perrimon, 1993) we silenced
CNOT1/not1 with Hand-Gal4, a driver specific for myocardial and pericardial cells of the heart,
acting throughout development (Han and Olson, 2005). At one-week post-eclosion (young
adult flies), we dissected the animals to expose the heart for video recording as previously
described (Ocorr et al., 2007a, Ocorr et al., 2014, Fink et al., 2009) (Fig. 3A). The Hand-Gal4-
driven CNOT1/not1 knockdown hearts exhibited normal beating frequency (shown as heart
period, Fig. 3B). However, these hearts were fragile and exhibited diastolic and systolic
diameters that were significantly larger compared to controls (Fig. 3C, D), which resulted in a
substantial decrease in fractional shortening (Fig. 3E), thus exhibiting reduced capacity for the
heart to contract. Fluorescent staining of actin revealed an abnormal myofibrillar structure with
large gaps and disarray in CNOT1/not1 knockdown fly hearts, as compared to control hearts
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 7
displaying typical tightly packed circumferential myofibrils (Fig. 3F, G). Knockdown with two
different CNOT1/not1 RNAi-lines (GD12571 and KK106587) resulted in similar phenotypes,
and knockdown with a third line, TRiP 28681 exhibited the same trend but was not statistically
significant from control fly hearts. The CNOT1/not1 RNAi-line VDRC GD12571 is shown in all
figures.
We had previously observed double beat EAD-associated arrhythmias in M-mode
traces from movies of CNOT3/not3 knockdown fly hearts (Neely et al., 2010), and therefore
an effort was made to record electrophysiology traces. However, this proved nearly impossible
due to the fragility of the CNOT1/not1 knockdown hearts. One successful recording did show
abnormal fibrillatory events, with increased event duration and number of peaks per burst (Fig.
S2A, B).
As the results of the hiPSC-CMs indicated the importance of the deadenylase CNOT7
for cardiac rhythm control, we next asked the question how knockdown of the Drosophila
ortholog CNOT7/8/pop2 would affect the fly heart. RNAi-mediated knockdown of
CNOT7/8/pop2, via the Hand-Gal4 driver line, resulted in pupal lethality at 25C. By lowering
the incubation temperature during development to 18C, and thereby reducing Gal4-
production, the flies did eclose. When one-week-old CNOT7/8/pop2 knockdown hearts were
functionally analyzed, they exhibited no change in heart period (Fig. 4A). However, significant
cardiac dilation, as measured by increased diastolic and systolic diameters (Fig. 4B, C), and
reduced contractility (Fig. 4D) were evident, consistent with the phenotype observed in
CNOT1/not1 knockdown fly hearts. Furthermore, fluorescent staining of CNOT7/8/pop2
knockdown hearts also revealed myofibrillar structure abnormalities (Fig. 4E, F), as seen in
CNOT1/not1 knockdown hearts (Fig. 3F, G). Electrophysiological recordings indicated that
cardiac-restricted silencing of CNOT7/pop2, as observed with the single CNOT1/not1
knockdown heart, triggered longer event durations and multiple peaks per burst (Fig. 4G-I)
compared to control hearts. While we cannot be certain that the single CNOT1/not1 recording
is representative, taken together with the CNOT7/8/pop2 electrophysiological recordings, we
find that this phenotype is consistent with a propensity for arrhythmia.
Hand-Gal4 driven RNAi-knockdown of CNOT2/not2, CNOT4/not4 and CNOT6/6L/twin
also caused cardiac dilation, as observed with the silencing of CNOT1/not1 and
CNOT7/8/pop2. Knockdown of CNOT2/not2, CNOT4/not4 and CNOT6/6L/twin induced a
significant increase in diastolic diameter (Fig. 5A), but only CNOT2/not2 and CNOT6/6L/twin
silencing also resulted in increased systolic diameters compared to controls (Fig. 5B). This
dilation did however not result in significantly diminished fractional shortening (Fig. 5C). The
results suggest that CNOT2/not2, CNOT4/not4 and CNOT6/6L/twin are needed for normal
heart dimensions in vivo, but their reduction did not significantly affect overall contractility.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 8
We repeated all RNAi-experiments with a second driver line, TinCΔ4-Gal4 (Lo and
Frasch, 2001), expressed in the myocardium during early development, during late pupal
stages of cardiac remodeling, and in the adult heart, but not during larval and early pupal
stages. Surprisingly, TinCΔ4-Gal4-mediated CNOT1/not1 knockdown did not cause a dilated
cardiac phenotype, except for a small increase in systolic heart diameter that resulted in
modestly decreased fractional shortening (Fig. S2C-E). TinCΔ4-Gal4 driven knockdown of
CNOT2/not2, CNOT4/not4 and CNOT6/6L /twin did not engender a cardiac phenotype
compared to controls, except for not4 knockdown, which resulted in substantially increased
diastolic and systolic diameters, but without an effect on fractional shortening (Fig. S3A-C).
Knockdown of CNOT1/not1 and CNOT7/8/pop2 during Drosophila development (larval
stages)
The finding that knockdown of CNOT1/not1 and CNOT7/8/pop2 with the pupal/adult
TinCΔ4-Gal4 myocardial driver (not expressed in larval hearts), did not recapitulate the results
obtained with the continuously expressed Hand-Gal4 heart driver, raised the question of
whether developmental expression in larvae/ early pupae was critical for normal adult heart
function. We therefore tested the hypothesis that the discrepancies observed between the
drivers were due to temporal expression differences. To test our hypothesis, we used the
driver NP1029-Gal4 that conferred larval/ early pupal-specific gene silencing (Monier et al.,
2005). CNOT1/not1 knockdown using NP1019-Gal4 was partially larval lethal, and completely
pupal lethal at 25C, while CNOT7/8/pop2-knockdown flies did eclose. Analysis of one-week-
old fly hearts upon larval/ early pupal CNOT7/8/pop2 knockdown revealed significant dilation
and reduction in fractional shortening and normal heart period (Fig. 6A-D), similar to the results
obtained with the Hand-Gal4 driver. Although we did not test for a cardiac phenotype at early
pupal stages, it is unlikely to be manifest similarly at adult stages, because most of the larval
heart will undergo histolysis and more anteriorly located portions of the larval aorta will
metamorphose during later pupal stages and become the adult heart (see Monier et al. 2005).
Thus, the observed adult heart phenotype is expected to be established during later pupal
stages but requiring CNOT7/8/pop2 function already during larval/early pupal stages, and
perhaps even in the embryo.
To test the latter, we further explored the developmental requirements by knocking
down CNOT7/8/pop2 in the cardiac mesoderm during early embryonal stages using the driver
line Tin-D-Gal4 (Reim and Frasch, 2005). This did not have any significant effects, except for
a prolonged heart period in CNOT7/8/pop2-knockdown flies compared to control (Fig. 6E-H).
The converse experiment was also performed, which was to knockdown of CNOT7/8/pop2 in
adult flies with the Hand-Gal4 Gene Switch system (Monnier et al., 2012) that activates RNAi-
mediated silencing in the myocardial and pericardial cells only when induced with RU-486.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 9
Adult flies were placed in food vials containing RU-486 at eclosion and were analyzed at one
week of age. Knockdown of CNOT7/8/pop2 in adult flies had no statistically significant effect
on any of the cardiac parameters measured (Fig. 6I-L).
Taken together, our data strongly suggest that the CCR4-NOT complex, in particular
CNOT1/not1 and CNOT7/8/pop2, along with CNOT3 (Neely et al., 2010, Yamaguchi et al.,
2018), are required during larval and/or early pupal stages of development, and thus during
the initial stages of cardiac remodeling from the larval to adult heart.
DISCUSSION
GWAS studies have successfully identified many genetic loci associated with multiple
disorders, including cardiovascular disease (Buniello et al., 2019). However, how to use
GWAS results, for recognizing the specific targets of these associations and for understanding
the biology of disease, is a major challenge in order to make progress towards diagnostics
improvement and personalized therapy. Here, we started with QT interval associating variants
in the CNOT1 gene, an integral component of the CCR4-NOT complex, and expanded on this
observation to include an investigation of additional complex subunits, with the combined
approach of using hiPSC-CMs and Drosophila, enabling a human and whole organ
assessment of cardiac physiology.
We examined the functionality of human QT associating variants in the CNOT1
promoter region and determined that the alleles of variants that significantly associate with
increases in QT interval are capable of lowering reporter gene expression, which may reflect
reduced transcription of CNOT1 as well. We do note, however, that the human SNPs tested
in vitro are located in the CNOT1 promoter region, and that mutation of the actual gene may
have different consequences in humans as compared with altered gene expression in cardiac
tissue alone. Since CNOT1 is essential for the function of the CCR4-NOT complex (Ito et al.,
2011), we expanded our study to include other subunits, to assess the range of potential
functional differences. We found that knockdown of not only CNOT1, but also CNOT2,
CNOT3, CNOT6, CNOT6L and CNOT7 decreased proliferation of hiPSC-CMs, and that
knockdown of CNOT7 also caused significant APD shortening. Knockdown of CNOT8 did not
change CM proliferation, which may be due to compensation by CNOT7 (Fig. 2).
Consistent with our observations in hiSPCs, in vivo findings also show that
CNOT1/not1 KD flies exhibit dilated hearts with reduced contractile ability, and severe
structural defects, similar to the myofibrillar reduction and cardiomyocyte death observed in
CNOT1 and CNOT3 muscle-specific knockout mice (Yamaguchi et al., 2018). Importantly,
silencing of CNOT7/8/pop2 resulted in cardiac damage similar to CNOT1/not1 knockdown,
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 10
and electrophysiological recordings demonstrated extended event duration and multiple peaks
per burst, which is indicative of a propensity for arrhythmias. Muscle tissue defects and
electrical activity have been linked in mouse and humans (Chinchilla and Franco, 2006).
Moreover, mutations in seizure, the Drosophila homolog of the human K+-channel hERG, both
important in cardiac repolarization, not only cause bradycardia and arrhythmia, but also
structural defects, such as myofibrillar disorganization (Ocorr et al., 2017). Silencing of
CNOT2/not2, CNOT4/not4 and CNOT6/6L/twin subunits led overall to similar, albeit weaker
phenotypes, limited to increased diastolic and systolic diameters. Importantly, the role of
CNOT subunits in the action potential (AP) repolarization phase was demonstrated not only
in the Drosophila heart model (Ocorr et al., 2017), but also in humans (Tse et al., 2017).
Notably, in our hands, dilated cardiomyopathy resulted as the main phenotype
produced by Hand-Gal4 driven knockdown of CNOT/not1 in the Drosophila heart.
Unfortunately, transthoracic echocardiography (TTE), the first-line imaging test in the
assessment of ventricular dilation (Mathew et al., 2017), was not performed in the individuals
from which we isolated the natural variants of the CNOT1 promoter and therefore we could
not evaluate structural alterations in those individuals. However, genetic based forms of long-
QT (LQT) have been associated with the development of dilated cardiomyopathy (DCM). An
overlap between DCM and LQT3 formed due to abnormalities of SCN5a gene, have been
described in multiple reports (Kwon et al., 2012, Shi et al., 2008) but there is also evidence for
the association between LQT1 and idiopathic DCM (Allen et al., 2016). In addition, it has been
reported that patients with both severe or mild forms of cardiomyopathies such as DCM or
hypertrophic cardiomyopathy (HCM), can show QT prolongation (Johnson et al., 2011, Jouven
et al., 2002, Ryerson and Giuffre, 2006). Of note, the role of the causative mutation in the
overlap between channelopathies and cardiomyopathies has not been fully understood and
the role of possible new players acting as phenotype modifiers, like the CCR4-NOT complex,
is yet to be determined.
When using a cardiac driver for gene knockdown that excluded the larval and early
pupal stages of fly heart development (TinCd4-Gal4), we failed to observe a strong
requirement for CNOT1/not1 and CNOT7/8/pop2. In contrast, when using a driver that was
restricted specifically to larval and early pupal stages (NP1029-Gal4), CNOT1/not1-
knockdown was lethal, but CNOT7/8/pop2 silencing at this stage of development resulted in
dilation and reduced contractility similar to knockdown exerted throughout life, whereas an
embryonic or adult-only driver had no effect. These findings suggest that CCR4-NOT function
is critical during cardiac remodeling from the larval to the adult heart. It is, however, also
possible that knockdown in adult flies would have an effect under stress conditions.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 11
Considering the effects on hiPSC-CM proliferation and the developmental defects
observed by silencing CNOT1/not1 and CNOT7/8/pop2 in Drosophila, we speculate that the
CCR4-NOT encoding genetic variants identified by GWAS in adult humans are those resulting
in less severe consequences, as the lack of proper mRNA regulation might be lethal at certain
stages of embryogenesis. Depletion of CNOT1 and the entire deadenylase module
(CNOT6/6L, CNOT7/8), respectively, has been demonstrated to promote ER-stress and
apoptosis in vitro (Ito et al., 2011). In turn, it has also been shown that activation of the
unfolded protein response impairs cardiac ion channel biogenesis leading to a prolongation of
the action potential duration (APD) (Liu et al., 2018). Taken together, these findings suggest
that disruption of CCR4-NOT complex function affect both structural (i.e. decreased CM
proliferation, myofibrillar structure abnormalities) and electrophysiological (i.e. shortened APD,
decreased contractility in flies) components of the heart. Whether knockdown of a specific
subunit produces one or both of those phenotypes may be influenced by silencing efficiency,
e.g. less ER-stress may lead to electrical remodeling, while more result in apoptosis. In
addition, specific RNA binding proteins that connect and guide CCR4-NOT to target specific
mRNAs are likely to have an influence. Collectively, our results show a prominent role of the
deadenylase module (CNOT7/8/pop2) both in vitro and in vivo.
This work demonstrates how the combinatorial use of GWAS studies and cardiac
model systems, enabled to connect CNOT1, CNOT7 and overall, the CCR4-NOT complex
function to cellular and whole heart phenotypes in the context human heart disease. However,
in this context, direct CCR4-NOT complex targets that influence heart rhythm and physiology
remain to be identified. Finally, strategies to modulate the expression of key components of
the CCR4-NOT complex, or stabilize its function, might be promising avenues for regulating
QT interval and preventing pro-arrhythmogenic substrates, especially targeted to those
individuals at increased risk due to their genetic background.
MATERIALS AND METHODS
Ethics statement
The DNA for promoter isolation was drawn from individual participants in the MICROS
study in South Tyrol (Pattaro et al., 2007). MICROS was approved by the Ethics Committee
of the Autonomous Province of Bolzano (Südtiroler Sanitätsbetrieb/Azienda Sanitaria dell’Alto
Adige). Each participant gave written informed consent.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 12
Generation of hiPSC-CMs
hiPSC were dissociated with 0.5 mM EDTA in PBS without CaCl2 and MgCl2 (Corning)
for 7min at RT, resuspended in mTeSR-1 media (StemCell Technologies) with 2 µM
Thiazovivin (StemCell Technologies) and 3 x 105 cells/well were plated in a Matrigel-coated
12-well plate. 24 hours after passage, cells were fed daily with mTeSR-1 media (without
Thiazovivin) for 3-5 days until ≥ 90% confluence. hiPSC-CMs were differentiated as previously
described (Burridge et al., 2015). Day 0, WNT signaling was activated by adding 6 µM
CHIR99021 (Selleck Chemicals) in S12 medium(Pei et al., 2017) for 48 hours. Day 2, cells
were treated with 2 µM Wnt-C59 (Selleck Chemicals) in S12 to inhibit WNT. Day 4, S12
medium was fully changed. Day 5, cells were dissociated with TrypLE Express (Gibco) for 4
minutes and blocked with RPMI (Gibco) +10% FBS (Omega Scientific). Cells were
resuspended in S12 supplemented with 4mg/L Recombinant Human Insulin (Gibco) (S12+
media) and 2µM Thiazovivin and 9 x 105 cells/well were plated in a Matrigel-coated 12-well
plate. S12+ media was changed Day 8 and replaced Day 10 by RPMI (Gibco) media + 213
µg/µL L-ascorbic acid (Sigma), 500 mg/L BSA-FV (Gibco), 0.5 mM L-carnitine (Sigma) and 8
g/L AlbuMAX Lipid-Rich BSA (Gibco) (CM medium). Under these conditions, hiPSC-CMs start
to beat around day 9-10. Day 15, cells were purified with lactate media, consisting of RPMI
without glucose, 213 µg/µL L-ascorbic acid, 500 mg/L BSA-FV and 8 mM Sodium-DL-Lactate
(Sigma) (Burridge et al., 2015, Tohyama et al., 2016), for 4-5 days and was replaced by CM
media until day 25.
Proliferation assay in hiPSC-CMs
At day 25 of differentiation, hiPSC-CMs were dissociated with TrypLE Select 10X
(Gibco), 12 min and neutralized with RPMI+10% FBS. Cells were resuspended in RPMI with
2% KOSR (Gibco) and 2% B27 50X with vitamin A (Life Technologies) supplemented with 2
µM Thiazovivin and plated at a density of 5000 cells/well in a Matrigel-coated 384-well plate.
hiPSC-CMs were transfected with siRNA (Dharmacon) targeting siCNOT1: L-015369-01,
siCNOT2: L020313-02, siCNOT3: L-020319-00, siCNOT4: L-020323-00, siNOT6: L-019101-
00, siNOT6L: L-016411-00, siCNOT7: L-012897-00 and siCNOT8: L-018791-00, using
lipofectamine RNAiMax (ThermoFisher). Each siRNA was tested in quadruplicate. 48 hours
post-transfection, cells were labelled with 10µM EdU (ThermoFisher). After 24h of EdU
incubation, cells were fixed with 4% paraformaldehyde for 30 min. EdU was detected
according to protocol and cells were stained with cardiac specific marker ACTN2 (Sigma,
dilution 1/800) and DAPI. Cells were imaged with ImageXpress Micro XLS microscope
(Molecular Devices) and custom algorithms were used to quantify EdU+ hiPSC-CMs.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 13
Voltage assay in hiPSC-CMs
Voltage assay was performed as described in McKeithan et al., 2017. Day 25 of
differentiation hiPSC-CMs were dissociated with TrypLE Select 10X for up to 12 min and
neutralized with RPMI+10% FBS. Cells were resuspended in RPMI with 2% KOSR (Gibco),
2% B27 50X with vitamin A (Life Technologies) and supplemented with 2µM Thiazovivin and
plated at 6.000 cells/well in a Matrigel-coated 384-well plate. hiPSC-CMs were transfected
with CCR4-NOT-NOT siRNAs as described above. Three days post-transfection, cells were
washed with pre-warmed Tyrode’s solution (Sigma) by removing 50 µL of media and adding
50 µL, five times. After the fifth wash, 50µL of 2x dye solution: voltage sensitive dye Vf2.1 Cl
(Fluovolt, 1:4000, ThermoFisher) diluted in Tyrode’s solution with 1µL of 10% Pluronic F127
(in water, ThermoFisher) and 20µg/mL Hoechst 33258 (in water, ThermoFisher) was added
to each well. The plate was returned to 37°C 5% CO2 incubator for 45 min. After incubation,
cells were washed four times with pre-warmed Tyrode’s. hiPSC-CMs were imaged with
ImageXpress Micro XLS microscope at 100 Hz for 5sec, with excitation wavelength at
485/20nm and emission filter 525/30nm. A single image of Hoechst was acquired before the
time series. Fluorescence over time quantification and trace analysis were automatically
quantified using custom software packages developed by Molecular Devices and Colas lab.
Three independent experiments were performed, each condition in quadruplicate.
Cell culture
HL-1 mouse atrial cardiomyocytes (Claycomb et al., 1998) were kindly donated by
William Claycomb (Louisiana State University, New Orleans) and cultured in Claycomb
medium (Sigma-Aldrich, USA) supplemented with 10% fetal bovine serum (FBS), 4 mM L-
glutamine, 100 U/mL Penicillin, 100 mg/mL Streptomycin, 0.3 mM Ascorbic Acid and 10 mM
Norepinephrine as previously described (Meraviglia et al., 2015).
Hela and 293T cells were cultured in Dulbecco's Modified Eagle Medium (DMEM),
GlutaMAX supplement (Thermo Fisher Scientific), supplemented with 10% Fetal bovine serum
(FBS) (Sigma) and 1% penicillin-streptomycin (Thermo Fisher Scientific). All cells were
maintained at 37°C in a saturated humidity atmosphere containing 5% CO2.
Luciferase assay
HL1, HeLa and T293 cells were seeded at 30,000, 60,000, and 150,000 cells/well,
respectively, into 24-well plates (Corning). 24 hours after seeding, 10ng of the reporter plasmid
pG4.74[hRluc/TK], was co-transfected with 10ng of either pGL4.10 vector (complete or
minimal for both haplotypes) or negative control vector (pG4.13 [luc2/SV40]). Transfection
was performed with the Lipofectamine plus reagent (Invitrogen), according to manufacturer’s
protocol. 48 hours post-transfection, cells were washed with PBS and lysed with 100 μl of
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 14
Passive Lysis Buffer (Promega) for 15 min at RT. Cell lysates were immediately used to
measure luciferase activity, using the Dual Luciferase Reporter Assay System kit (Promega).
Each lysate (20 μl) was incubated with 100 μl of Luciferase Assay Reagent II (LAR II). Firefly
luminescence was measured for 10 seconds using a luminometer (Victor X3-2030, Perkin
Elmer). After 2 seconds, 100μl of Dual-Glo Stop & Glo Reagent was added to each well.
Subsequently, renilla luminescence was measured for 10 seconds using the same
luminometer. Luciferase activity was calculated based on the ratio of the activities of firefly
and renilla luciferases. At least three independent experiments were performed in triplicate.
Fly stocks
All transgenic RNAi-fly lines were purchased from Vienna Drosophila RNAi Center
(VDRC) and from Bloomington Drosophila Stock Center at Indiana University (Transgenic
RNAi Project at Harvard Medical School, TRiP). VDRC IDs: not1 GD12571 and KK106587,
not2 GD20826, pop2 GD28396, twin GD13365. TRiP/ BDSC IDs: not1 28681, not4 JF03203,
pop2 HM05235, twin HMS00493. Control flies with corresponding genetic background: VDRC
w1118 (GD RNAi library), TRiP-fly line with attP2 docking site. Cardiac-specific drivers were
kind gifts from the following investigators: Manfred Frasch: TinCΔ4 12a-Gal4 (Lo and Frasch,
2001) and tin-D-Gal4 (Reim and Frasch, 2005), Eric Olsen: Hand-Gal4 (Han and Olson, 2005),
Laurent Perrin: NP1029-Gal4 (Monier et al., 2005) and Hand-Gal4 Gene Switch (Monnier et
al., 2012).
Fly medium
Ingredients: cornmeal (7.0%), malt (5.2%), molasses (5.2%), soy flour (1.7%), agar
(0.4%), autolyzed yeast (2.1%). All ingredients were mixed with water and cooked for 15
minutes without boiling. Preservatives, Tegosept in EtOH (1.8%) and propionic acid (2.1%),
were added once the batter cooled <65C. All percentages refer to final concentration. Dry
ingredients: weight/ total volume batter, liquid ingredients: volume/ total volume batter.
Fly crosses
Driver-line virgins were crossed to RNAi-males and corresponding isogenic control
males. Flies were raised on standard fly food and kept in 25C or 18C. Female progeny was
collected and aged to 1 week in 25C at which point they were imaged and analyzed.
Hand-Gal4 Gene-Switch knockdown
Eclosed female progeny was collected and placed in vials with fly food containing
100µg/ml RU-486, (40µl of 25mg/ml stock RU-486 dissolved in EtOH was added to 10ml fly
food). Control food contained an equal amount of EtOH. Flies were aged to 1 week in 25C at
which point they were dissected and analyzed.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 15
Fly heart dissection
Flies were anesthetized with FlyNap and dissected in artificial hemolymph according to a
previously described protocol (Vogler and Ocorr, 2009). The procedure includes removing the
fly head, intestines and some fat, resulting in a semi-intact preparation that visualizes the
beating heart. Artificial hemolymph was re-oxygenated for 20 minutes post dissection allowing
the hearts to stabilize before video recording.
High speed digital video imaging and analysis
All fly hearts were filmed with an EM-CDD Hamamatsu digital camera, using a Leica
DMFLSA microscope equipped with a 10x dipping lens. 30-second recordings of each fly heart
were made with a camera speed of 120-140 frames per second (Ocorr et al., 2007b). M-modes
describing fly heart contractions were created by Semi-automated Optical Heartbeat Analysis
(Fink et al., 2009), SOHA.
Fluorescent staining and imaging
According to a previously described protocol (Alayari et al., 2009), flies were dissected
in artificial hemolymph and hearts relaxed with 10mM EGTA before fixation with formaldehyde.
Flies were stained with Phalloidin, Alexa488 to visualize F-actin. Apotome images were taken
with a Zeiss Axio Imager.Z1 microscope at 10x and 25x. Images were processed with Adobe
Photoshop.
Statistical Analysis
To determine any statistical significance between experimental and control groups in
hiPSC-CMs and Drosophila experiments, we calculated two-sided p-values with Student’s t-
test, One-way- and Two-way-ANOVA with Tukey’s multiple comparisons test, using GraphPad
Prism software (2016). We analyzed CNOT1 expression data from HL-1, HeLa and T293 cell
lines with a two-sided non-parametric Wilcoxon rank-sum test, using Stata 13 (StataCorp.
2013. Stata Statistical Software: Release 13. College Station, TX: StataCorp LP). Population
distribution of siCtrl and siCNOT7-transfected hiPSC-CMs was generated with GraphPad
Prism using nonlinear regression. Unpaired nonparametric Kolmogorov-Smirnov test was
used to compare each treated condition to controls using APD75 of every measured cell.
Electrophysiology of Adult Hearts
Semi-intact heart preparations were incubated in artificial hemolymph containing 10µM
blebbistatin (Sigma Aldrich), left in the dark with oxygenation until the hearts stopped. Fresh
saline without blebbistatin was added and electrical potentials were recorded from the conical
chamber using glass electrodes (20-50 MΩ) filled with 3M KCl. Data were acquired using an
Axon-700B amplifier, signals were digitized using the DIGIDATA 1322A and data were
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 16
captured and analyzed using PClamp 9.0 and Clampfit 10.0 software from Molecular Devices.
Data was quantified from representative 30s recordings where the resting membrane potential
had remained stable.
Competing interests - No competing interests declared.
Funding
This study was supported by grants from NIH R01-HL054732 to RB and R01-
HL124091 to AC, the Wanek Foundation to RB and AC and by the Department of Innovation,
Research and Universities of the Autonomous Province of Bolzano-South Tyrol (Italy) to CV,
LF, AR and AH.
Author contributions
Lisa Elmén performed the Drosophila experiments and data analysis together with
Anthony Cammarato, Rolf Bodmer and Nakissa Alayari. Claudia Volpato, Andrew Hicks and
Luisa Foco analysed the genetics data, and created the resources to perform the cell culture
experiments, which were analyzed with Alessandra Rossini. Santiago Pineda performed the
Drosophila electrophysiology experiments and analyzed the recorded data with Karen Ocorr
and Rolf Bodmer. Anaïs Kervadec performed all hiPSC-experiments and analyzed the data
with Alexandre Colas. Lisa Elmén, Andrew Hicks and Rolf Bodmer wrote and edited the
manuscript with contributions from Alessandra Rossini, Alexandre Colas and Anthony
Cammarato.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 17
References
ALAYARI, N. N., VOGLER, G., TAGHLI-LAMALLEM, O., OCORR, K., BODMER, R. &
CAMMARATO, A. 2009. Fluorescent labeling of Drosophila heart structures. J Vis
Exp.
ALLEN, K. Y., VETTER, V. L., SHAH, M. J. & O'CONNOR, M. J. 2016. Familial long QT
syndrome and late development of dilated cardiomyopathy in a child with a KCNQ1
mutation: A case report. HeartRhythm Case Rep, 2, 128-131.
AMIN, A. S., PINTO, Y. M. & WILDE, A. A. 2013. Long QT syndrome: beyond the causal
mutation. J Physiol, 591, 4125-39.
ARKING, D. E., PULIT, S. L., CROTTI, L., VAN DER HARST, P., MUNROE, P. B.,
KOOPMANN, T. T., SOTOODEHNIA, N., ROSSIN, E. J., MORLEY, M., WANG,
X., JOHNSON, A. D., LUNDBY, A., GUDBJARTSSON, D. F., NOSEWORTHY, P.
A., EIJGELSHEIM, M., BRADFORD, Y., TARASOV, K. V., DORR, M., MULLER-
NURASYID, M., LAHTINEN, A. M., NOLTE, I. M., SMITH, A. V., BIS, J. C.,
ISAACS, A., NEWHOUSE, S. J., EVANS, D. S., POST, W. S., WAGGOTT, D.,
LYYTIKAINEN, L. P., HICKS, A. A., EISELE, L., ELLINGHAUS, D., HAYWARD,
C., NAVARRO, P., ULIVI, S., TANAKA, T., TESTER, D. J., CHATEL, S.,
GUSTAFSSON, S., KUMARI, M., MORRIS, R. W., NALUAI, A. T.,
PADMANABHAN, S., KLUTTIG, A., STROHMER, B., PANAYIOTOU, A. G.,
TORRES, M., KNOFLACH, M., HUBACEK, J. A., SLOWIKOWSKI, K.,
RAYCHAUDHURI, S., KUMAR, R. D., HARRIS, T. B., LAUNER, L. J.,
SHULDINER, A. R., ALONSO, A., BADER, J. S., EHRET, G., HUANG, H., KAO,
W. H., STRAIT, J. B., MACFARLANE, P. W., BROWN, M., CAULFIELD, M. J.,
SAMANI, N. J., KRONENBERG, F., WILLEIT, J., CONSORTIUM, C. A.,
CONSORTIUM, C., SMITH, J. G., GREISER, K. H., MEYER ZU
SCHWABEDISSEN, H., WERDAN, K., CARELLA, M., ZELANTE, L.,
HECKBERT, S. R., PSATY, B. M., ROTTER, J. I., KOLCIC, I., POLASEK, O.,
WRIGHT, A. F., GRIFFIN, M., DALY, M. J., DCCT/EDIC, ARNAR, D. O., HOLM,
H., THORSTEINSDOTTIR, U., E, M. C., DENNY, J. C., RODEN, D. M., ZUVICH,
R. L., EMILSSON, V., PLUMP, A. S., LARSON, M. G., O'DONNELL, C. J., YIN,
X., BOBBO, M., D'ADAMO, A. P., IORIO, A., SINAGRA, G., et al. 2014. Genetic
association study of QT interval highlights role for calcium signaling pathways in
myocardial repolarization. Nat Genet, 46, 826-36.
BHANDARI, D., RAISCH, T., WEICHENRIEDER, O., JONAS, S. & IZAURRALDE, E.
2014. Structural basis for the Nanos-mediated recruitment of the CCR4-NOT complex
and translational repression. Genes Dev, 28, 888-901.
BRAND, A. H. & PERRIMON, N. 1993. Targeted gene expression as a means of altering cell
fates and generating dominant phenotypes. Development, 118, 401-15.
BUNIELLO, A., MACARTHUR, J. A. L., CEREZO, M., HARRIS, L. W., HAYHURST, J.,
MALANGONE, C., MCMAHON, A., MORALES, J., MOUNTJOY, E., SOLLIS, E.,
SUVEGES, D., VROUSGOU, O., WHETZEL, P. L., AMODE, R., GUILLEN, J. A.,
RIAT, H. S., TREVANION, S. J., HALL, P., JUNKINS, H., FLICEK, P., BURDETT,
T., HINDORFF, L. A., CUNNINGHAM, F. & PARKINSON, H. 2019. The NHGRI-
EBI GWAS Catalog of published genome-wide association studies, targeted arrays and
summary statistics 2019. Nucleic Acids Res, 47, D1005-D1012.
BURRIDGE, P. W., HOLMSTROM, A. & WU, J. C. 2015. Chemically Defined Culture and
Cardiomyocyte Differentiation of Human Pluripotent Stem Cells. Curr Protoc Hum
Genet, 87, 21 3 1-15.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 18
CHINCHILLA, A. & FRANCO, D. 2006. Regulatory mechanisms of cardiac development and
repair. Cardiovasc Hematol Disord Drug Targets, 6, 101-12.
COLLART, M. A. 2016. The Ccr4-Not complex is a key regulator of eukaryotic gene
expression. Wiley Interdiscip Rev RNA, 7, 438-54.
COLLART, M. A. & PANASENKO, O. O. 2017. The Ccr4-Not Complex: Architecture and
Structural Insights. Subcell Biochem, 83, 349-379.
CUNNINGHAM, T. J., YU, M. S., MCKEITHAN, W. L., SPIERING, S., CARRETTE, F.,
HUANG, C. T., BUSHWAY, P. J., TIERNEY, M., ALBINI, S., GIACCA, M., MANO,
M., PURI, P. L., SACCO, A., RUIZ-LOZANO, P., RIOU, J. F., UMBHAUER, M.,
DUESTER, G., MERCOLA, M. & COLAS, A. R. 2017. Id genes are essential for early
heart formation. Genes Dev.
FINK, M., CALLOL-MASSOT, C., CHU, A., RUIZ-LOZANO, P., IZPISUA BELMONTE,
J. C., GILES, W., BODMER, R. & OCORR, K. 2009. A new method for detection and
quantification of heartbeat parameters in Drosophila, zebrafish, and embryonic mouse
hearts. Biotechniques, 46, 101-13.
HALTER, D., COLLART, M. A. & PANASENKO, O. O. 2014. The Not4 E3 ligase and CCR4
deadenylase play distinct roles in protein quality control. PLoS One, 9, e86218.
HAN, Z. & OLSON, E. N. 2005. Hand is a direct target of Tinman and GATA factors during
Drosophila cardiogenesis and hematopoiesis. Development, 132, 3525-36.
ITO, K., TAKAHASHI, A., MORITA, M., SUZUKI, T. & YAMAMOTO, T. 2011. The role
of the CNOT1 subunit of the CCR4-NOT complex in mRNA deadenylation and cell
viability. Protein Cell, 2, 755-63.
ITZHAKI, I., MAIZELS, L., HUBER, I., ZWI-DANTSIS, L., CASPI, O., WINTERSTERN,
A., FELDMAN, O., GEPSTEIN, A., ARBEL, G., HAMMERMAN, H., BOULOS, M.
& GEPSTEIN, L. 2011. Modelling the long QT syndrome with induced pluripotent
stem cells. Nature, 471, 225-9.
JOHNSON, J. N., GRIFONI, C., BOS, J. M., SABER-AYAD, M., OMMEN, S. R., NISTRI,
S., CECCHI, F., OLIVOTTO, I. & ACKERMAN, M. J. 2011. Prevalence and clinical
correlates of QT prolongation in patients with hypertrophic cardiomyopathy. Eur Heart
J, 32, 1114-20.
JOUVEN, X., HAGEGE, A., CHARRON, P., CARRIER, L., DUBOURG, O., LANGLARD,
J. M., ALIAGA, S., BOUHOUR, J. B., SCHWARTZ, K., DESNOS, M. & KOMAJDA,
M. 2002. Relation between QT duration and maximal wall thickness in familial
hypertrophic cardiomyopathy. Heart, 88, 153-7.
KRUK, J. A., DUTTA, A., FU, J., GILMOUR, D. S. & REESE, J. C. 2011. The multifunctional
Ccr4-Not complex directly promotes transcription elongation. Genes Dev, 25, 581-93.
KWON, H. W., LEE, S. Y., KWON, B. S., KIM, G. B., BAE, E. J., KIM, W. H., NOH, C. I.,
CHO, S. I. & PARK, S. S. 2012. Long QT syndrome and dilated cardiomyopathy with
SCN5A p.R1193Q polymorphism: cardioverter-defibrillator implantation at 27
months. Pacing Clin Electrophysiol, 35, e243-6.
LAHTI, A. L., KUJALA, V. J., CHAPMAN, H., KOIVISTO, A. P., PEKKANEN-MATTILA,
M., KERKELA, E., HYTTINEN, J., KONTULA, K., SWAN, H., CONKLIN, B. R.,
YAMANAKA, S., SILVENNOINEN, O. & AALTO-SETALA, K. 2012. Model for
long QT syndrome type 2 using human iPS cells demonstrates arrhythmogenic
characteristics in cell culture. Dis Model Mech, 5, 220-30.
LIU, M., SHI, G., ZHOU, A., RUPERT, C. E., COULOMBE, K. L. K. & DUDLEY, S. C., JR.
2018. Activation of the unfolded protein response downregulates cardiac ion channels
in human induced pluripotent stem cell-derived cardiomyocytes. J Mol Cell Cardiol,
117, 62-71.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 19
LO, P. C. & FRASCH, M. 2001. A role for the COUP-TF-related gene seven-up in the
diversification of cardioblast identities in the dorsal vessel of Drosophila. Mech Dev,
104, 49-60.
MATHEW, T., WILLIAMS, L., NAVARATNAM, G., RANA, B., WHEELER, R.,
COLLINS, K., HARKNESS, A., JONES, R., KNIGHT, D., O'GALLAGHER, K.,
OXBOROUGH, D., RING, L., SANDOVAL, J., STOUT, M., SHARMA, V.,
STEEDS, R. P. & BRITISH SOCIETY OF ECHOCARDIOGRAPHY EDUCATION,
C. 2017. Diagnosis and assessment of dilated cardiomyopathy: a guideline protocol
from the British Society of Echocardiography. Echo Res Pract, 4, G1-G13.
MATSA, E., RAJAMOHAN, D., DICK, E., YOUNG, L., MELLOR, I., STANIFORTH, A. &
DENNING, C. 2011. Drug evaluation in cardiomyocytes derived from human induced
pluripotent stem cells carrying a long QT syndrome type 2 mutation. Eur Heart J, 32,
952-62.
MCKEITHAN, W. L., SAVCHENKO, A., YU, M. S., CERIGNOLI, F., BRUYNEEL, A. A.
N., PRICE, J. H., COLAS, A. R., MILLER, E. W., CASHMAN, J. R. & MERCOLA,
M. 2017. An Automated Platform for Assessment of Congenital and Drug-Induced
Arrhythmia with hiPSC-Derived Cardiomyocytes. Front Physiol, 8, 766.
MONIER, B., ASTIER, M., SEMERIVA, M. & PERRIN, L. 2005. Steroid-dependent
modification of Hox function drives myocyte reprogramming in the Drosophila heart.
Development, 132, 5283-93.
MONNIER, V., ICHE-TORRES, M., RERA, M., CONTREMOULINS, V., GUICHARD, C.,
LALEVEE, N., TRICOIRE, H. & PERRIN, L. 2012. dJun and Vri/dNFIL3 are major
regulators of cardiac aging in Drosophila. PLoS Genet, 8, e1003081.
NEELY, G. G., KUBA, K., CAMMARATO, A., ISOBE, K., AMANN, S., ZHANG, L.,
MURATA, M., ELMEN, L., GUPTA, V., ARORA, S., SARANGI, R., DAN, D.,
FUJISAWA, S., USAMI, T., XIA, C. P., KEENE, A. C., ALAYARI, N. N.,
YAMAKAWA, H., ELLING, U., BERGER, C., NOVATCHKOVA, M.,
KOGLGRUBER, R., FUKUDA, K., NISHINA, H., ISOBE, M., POSPISILIK, J. A.,
IMAI, Y., PFEUFER, A., HICKS, A. A., PRAMSTALLER, P. P., SUBRAMANIAM,
S., KIMURA, A., OCORR, K., BODMER, R. & PENNINGER, J. M. 2010. A global
in vivo Drosophila RNAi screen identifies NOT3 as a conserved regulator of heart
function. Cell, 141, 142-53.
OCORR, K., PERRIN, L., LIM, H. Y., QIAN, L., WU, X. & BODMER, R. 2007a. Genetic
control of heart function and aging in Drosophila. Trends Cardiovasc Med, 17, 177-82.
OCORR, K., REEVES, N. L., WESSELLS, R. J., FINK, M., CHEN, H. S., AKASAKA, T.,
YASUDA, S., METZGER, J. M., GILES, W., POSAKONY, J. W. & BODMER, R.
2007b. KCNQ potassium channel mutations cause cardiac arrhythmias in Drosophila
that mimic the effects of aging. Proc Natl Acad Sci U S A, 104, 3943-8.
OCORR, K., VOGLER, G. & BODMER, R. 2014. Methods to assess Drosophila heart
development, function and aging. Methods, 68, 265-72.
OCORR, K., ZAMBON, A., NUDELL, Y., PINEDA, S., DIOP, S., TANG, M., AKASAKA,
T. & TAYLOR, E. 2017. Age-dependent electrical and morphological remodeling of
the Drosophila heart caused by hERG/seizure mutations. PLoS Genet, 13, e1006786.
PATTARO, C., GOGELE, M., MASCALZONI, D., MELOTTI, R., SCHWIENBACHER, C.,
DE GRANDI, A., FOCO, L., D'ELIA, Y., LINDER, B., FUCHSBERGER, C.,
MINELLI, C., EGGER, C., KOFINK, L. S., ZANIGNI, S., SCHAFER, T.,
FACHERIS, M. F., SMARASON, S. V., ROSSINI, A., HICKS, A. A., WEISS, H. &
PRAMSTALLER, P. P. 2015. The Cooperative Health Research in South Tyrol
(CHRIS) study: rationale, objectives, and preliminary results. J Transl Med, 13, 348.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 20
PATTARO, C., MARRONI, F., RIEGLER, A., MASCALZONI, D., PICHLER, I.,
VOLPATO, C. B., DAL CERO, U., DE GRANDI, A., EGGER, C., EISENDLE, A.,
FUCHSBERGER, C., GOGELE, M., PEDROTTI, S., PINGGERA, G. K.,
STEFANOV, S. A., VOGL, F. D., WIEDERMANN, C. J., MEITINGER, T. &
PRAMSTALLER, P. P. 2007. The genetic study of three population microisolates in
South Tyrol (MICROS): study design and epidemiological perspectives. BMC Med
Genet, 8, 29.
PEI, F., JIANG, J., BAI, S., CAO, H., TIAN, L., ZHAO, Y., YANG, C., DONG, H. & MA, Y.
2017. Chemical-defined and albumin-free generation of human atrial and ventricular
myocytes from human pluripotent stem cells. Stem Cell Res, 19, 94-103.
PFEUFER, A., SANNA, S., ARKING, D. E., MULLER, M., GATEVA, V., FUCHSBERGER,
C., EHRET, G. B., ORRU, M., PATTARO, C., KOTTGEN, A., PERZ, S., USALA,
G., BARBALIC, M., LI, M., PUTZ, B., SCUTERI, A., PRINEAS, R. J., SINNER, M.
F., GIEGER, C., NAJJAR, S. S., KAO, W. H. L., MUHLEISEN, T. W., DEI, M.,
HAPPLE, C., MOHLENKAMP, S., CRISPONI, L., ERBEL, R., JOCKEL, K. H.,
NAITZA, S., STEINBECK, G., MARRONI, F., HICKS, A. A., LAKATTA, E.,
MULLER-MYHSOK, B., PRAMSTALLER, P. P., WICHMANN, H. E.,
SCHLESSINGER, D., BOERWINKLE, E., MEITINGER, T., UDA, M., CORESH, J.,
KAAB, S., ABECASIS, G. R. & CHAKRAVARTI, A. 2009. Common variants at ten
loci modulate the QT interval duration in the QTSCD Study. Nature Genetics, 41, 407-
414.
REIM, I. & FRASCH, M. 2005. The Dorsocross T-box genes are key components of the
regulatory network controlling early cardiogenesis in Drosophila. Development, 132,
4911-25.
RUDIC, B., SCHIMPF, R. & BORGGREFE, M. 2014. Short QT Syndrome - Review of
Diagnosis and Treatment. Arrhythm Electrophysiol Rev, 3, 76-9.
RYERSON, L. M. & GIUFFRE, R. M. 2006. QT intervals in metabolic dilated
cardiomyopathy. Can J Cardiol, 22, 217-20.
SHI, R., ZHANG, Y., YANG, C., HUANG, C., ZHOU, X., QIANG, H., GRACE, A. A.,
HUANG, C. L. & MA, A. 2008. The cardiac sodium channel mutation delQKP 1507-
1509 is associated with the expanding phenotypic spectrum of LQT3, conduction
disorder, dilated cardiomyopathy, and high incidence of youth sudden death. Europace,
10, 1329-35.
TEMME, C., ZHANG, L., KREMMER, E., IHLING, C., CHARTIER, A., SINZ, A.,
SIMONELIG, M. & WAHLE, E. 2010. Subunits of the Drosophila CCR4-NOT
complex and their roles in mRNA deadenylation. RNA, 16, 1356-70.
TOHYAMA, S., FUJITA, J., HISHIKI, T., MATSUURA, T., HATTORI, F., OHNO, R.,
KANAZAWA, H., SEKI, T., NAKAJIMA, K., KISHINO, Y., OKADA, M., HIRANO,
A., KURODA, T., YASUDA, S., SATO, Y., YUASA, S., SANO, M., SUEMATSU,
M. & FUKUDA, K. 2016. Glutamine Oxidation Is Indispensable for Survival of Human
Pluripotent Stem Cells. Cell Metab, 23, 663-74.
TSE, G., CHAN, Y. W., KEUNG, W. & YAN, B. P. 2017. Electrophysiological mechanisms
of long and short QT syndromes. Int J Cardiol Heart Vasc, 14, 8-13.
VACANTI, G., MARAGNA, R., PRIORI, S. G. & MAZZANTI, A. 2017. Genetic causes of
sudden cardiac death in children: inherited arrhythmogenic diseases. Curr Opin
Pediatr, 29, 552-559.
VOGLER, G. & OCORR, K. 2009. Visualizing the beating heart in Drosophila. J Vis Exp.
WEBSTER, M. W., CHEN, Y. H., STOWELL, J. A. W., ALHUSAINI, N., SWEET, T.,
GRAVELEY, B. R., COLLER, J. & PASSMORE, L. A. 2018. mRNA Deadenylation
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 21
Is Coupled to Translation Rates by the Differential Activities of Ccr4-Not Nucleases.
Mol Cell, 70, 1089-1100 e8.
YAMAGUCHI, T., SUZUKI, T., SATO, T., TAKAHASHI, A., WATANABE, H.,
KADOWAKI, A., NATSUI, M., INAGAKI, H., ARAKAWA, S., NAKAOKA, S.,
KOIZUMI, Y., SEKI, S., ADACHI, S., FUKAO, A., FUJIWARA, T., NATSUME, T.,
KIMURA, A., KOMATSU, M., SHIMIZU, S., ITO, H., SUZUKI, Y., PENNINGER,
J. M., YAMAMOTO, T., IMAI, Y. & KUBA, K. 2018. The CCR4-NOT deadenylase
complex controls Atg7-dependent cell death and heart function. Sci Signal, 11.
YI, H., PARK, J., HA, M., LIM, J., CHANG, H. & KIM, V. N. 2018. PABP Cooperates with
the CCR4-NOT Complex to Promote mRNA Deadenylation and Block Precocious
Decay. Mol Cell, 70, 1081-1088 e5.
YU, M. S., SPIERING, S. & COLAS, A. R. 2018. Generation of First Heart Field-like Cardiac
Progenitors and Ventricular-like Cardiomyocytes from Human Pluripotent Stem Cells.
J Vis Exp.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 22
Tables
Table 1. CCR4-NOT genes in this study
Human Drosophila
CNOT1 not1
CNOT2 not2/rga
CNOT3 not3
CNOT4 not4
CNOT6 and CNOT6L twin
CNOT7 and CNOT8 pop2
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 23
Figures
Figure 1. A) Cartoon of the CCR4-Not complex with the subunits investigated in this study.
B) GWAS identifies CNOT1 SNPs associated with human QT syndrome. The four putative
promoter SNPs are highlighted by red box. C) Constructs generated for the CNOT1 complete
promoter region and the minimal promoter region, cloned from two subjects carrying haplotype
“GAGT” and haplotype “TGAG”. Numbers above each SNP denote how close the variant is to
the open reading frame (e.g. 1 is the closest). D) Schematic of experimental procedure. E)
Box plots summarizing the intensity of the ratio between luciferase and renilla signal in HL1
cells. Boxes: interquartile range, central line: median; whiskers: upper and lower adjacent
values as defined by Tukey (1977); dots: outside values. Two-sided P values were computed
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 24
using the Wilcoxon rank-sum test. For complete promoter, n independent experiments=3; for
each experiment, n of independent biological replicates per haplotype=3. Total n of
observations for each haplotype=9, total n=18. For minimal promoter: n independent
experiments=5; for each experiment, n of independent biological replicates per haplotype=3,
except one experiment (n=4). Total n of observations for each haplotype=16, total n=32.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 25
Figure 2. A) Schematic of proliferation assay in hiPSC-CMs. B) Representative
immunofluorescence images for EdU and ACTN1 in siCtrl, siCNOT1 and siCNOT3 conditions.
Scale bar, 25 µm. C-D) Histogram showing normalized % of EdU positive hiPSC-CMs and
normalized number of hiPSC-CMs. T-test was used to calculate p-values. E) Schematic
overview of single cell and high throughput voltage assay. F) Two-dimensional graph for
APD75 and Kolgomorov Smirnov distance (KS-D) representing screen results for CCR4-
CNOT components knockdown. G) Population distribution of APD75 measurements for
siCNOT7 vs siControl-transfected hiPSC-CMs. H) Median action potential traces for siCNOT7
and siControl transfected hiPSC-CMs. I) Table summarizing average and standard deviation
values for APD50, APD75 and APD90 for siCNOT7 and siControl transfected hiPSC-CMs.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 26
Figure 3. A) Schematic of heart analysis procedure in Drosophila. Dissection in artificial
hemolymph exposes the beating fly heart for video recording. Automated quantification
generates m-modes used to measure heart period, heart size and contractility. B-E) RNAi-
mediated knockdown of CNOT1/not1 (VDRC, GD12571) using cardiomyocyte and pericardial
cell specific driver Hand-Gal4. n=40 female flies per genotype. Reduced CNOT1/not1
expression increased diastolic and systolic diameters and reduced fractional shortening.
Student’s t-test was used to calculate two-sided p-values. Boxes: interquartile range, central
line: median; plus: mean; whiskers: upper and lower adjacent values as defined by Tukey
(1977); dots: outside values. F-G) Immunofluorescence staining with phalloidin visualized f-
actin of Drosophila hearts. Representative not1 knockdown heart show myofibrillar disarray
and gaps in muscle tissue compared to tightly packed circumferential myofibrils in control
heart. The scale bar is 200µm.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 27
Figure 4. A-D) RNAi-mediated knockdown of CNOT7/8/pop2 (TRiP HM05235) using
cardiomyocyte and pericardial cell specific driver Hand-Gal4. n=32 female flies per genotype.
Reduced pop2 expression increased diastolic and systolic diameters and reduced fractional
shortening. Student’s t-test was used to calculate two-sided p-values. Boxes: interquartile
range, central line: median; plus: mean; whiskers: upper and lower adjacent values as defined
by Tukey (1977); dots: outside values. E-F) Immunofluorescent staining with phalloidin
visualizes f-actin of Drosophila hearts. CNOT7/8/pop2 knockdown heart show dilation and
gaps in muscle tissue compared to tightly packed circumferential myofibrils in control heart.
The scale bar is 100µm. G) Table summarizing electrophysiology measurements of
CNOT7/8/pop2-knockdown fly hearts. H-I) Representative 10s m-modes show greater peaks
per burst and longer event duration of CNOT7/8/pop2-knockdown (Hand-
Gal4>CNOT7/8/pop2) fly hearts compared to control (w1118 x CNOT7/8/pop2).
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 28
Figure 5. RNAi-mediated knockdown of CNOT2/not2 (VDRC GD20826), CNOT4/not4 (TRiP
JF03203) and CNOT6/6L/twin (VDRC GD13365) using cardiomyocyte and pericardial cell
specific driver Hand-Gal4. n=15 female flies per genotype. A) Reduced expression of
CNOT2/not2, CNOT4/not4 and CNOT6/6L/twin significantly increased diastolic diameter. B)
Reduced expression of CNOT2/not2, and CNOT6/6L/twin significantly increased systolic
diameter. C) Fractional shortening was not changed by reduced expression of either
CNOT2/not2, CNOT4/not4 or CNOT6/6L/twin. One-way ANOVA with Tukey’s multiple
comparisons test was used to calculate two-sided p-values. Boxes: interquartile range, central
line: median; plus: mean; whiskers: upper and lower adjacent values as defined by Tukey
(1977); dots: outside values.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 29
Figure 6. RNAi-mediated knockdown of CNOT7/8/pop2 (TRiP HM05235) during
developmental stages and in adult fly hearts. A-D) RNAi-mediated knockdown of
CNOT7/8/pop2 using larval stage heart specific driver NP1029-Gal4. n=19 females flies per
genotype. Reduced CNOT7/8/pop2 expression increased diastolic and systolic diameters and
reduced fractional shortening. E-H) RNAi-mediated knockdown of CNOT7/8/pop2 using
embryonal stage heart specific driver TinD-Gal4 extended heart period but had no effect on
heart diameters and contractility. n=19 female flies per genotype. Student’s t-test was used to
calculate two-sided p-values. I-L) RNAi-mediated knockdown of CNOT7/8/pop2 using the RU-
486 inducible Hand-Gal4 Gene Switch driver. Reduced expression of CNOT7/8/pop2in adult
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 30
fly hearts had no effect on heart diameters or contractility. n=12 females flies per condition
and genotype. Two-way ANOVA was used to calculate p-values. Boxes: interquartile range,
central line: median; plus: mean; whiskers: upper and lower adjacent values as defined by
Tukey (1977); dots: outside values.
Dis
ease
Mo
dels
& M
echa
nism
s •
DM
M •
Acc
epte
d m
anus
crip
t
Page 31
Complete CNOT1 promoter
luci
fera
ce/ r
enill
a ra
tio 2.0
1.5
1.0
0.5
0
GAGTGAGT
p=0.0575
HeLa T293
p=0.0464
Figure S1
A Minimal CNOT1 promoterB
1.0
0.8
0.6
0.4
0
GAGTGAGT
p=0.0003 p=0.0004
0.2
luci
fera
ce/ r
enill
a ra
tioHeLa T293
C
Marker Name
rs27097
rs37037
rs9941290
rs863433
Allele1 Allele2
T
A
A
T
Effect (ms) StdErr (ms) P-value
G
G
G
G
Direction
1.6218
-0.6233
1.617
-1.6166
3.62E-12
0.1997
2.71E-12
0.2313
0.2313
2.71E-12
0.2333
0.001805
++++++
-+----
++++++
------
G and T
G and T
G and T
G and T
Figure S1. Box plots summarizing the intensity of the ratio between luciferase and renilla signal in HeLa and T293 cells. A) Complete CNOT1 promoter construct. B) Minimal CNOT1 promoter construct. C) This table lists the four SNPs in strong linkage disequilibrium within the putative promoter region of the CNOT1 gene, with association p-values derived from the QTSCD consortium data18. This region was reported originally as the GINS3-NDRG4-SETD6-CNOT1 region as no resolution as to the functional gene in this region was possible without further experimental validation, provided herein. Allele 1 represents the effect allele identified in the GWAS meta-analysis and sequencing of this region from individuals homozygous for these variants revealed haplotypes consistent with effect direction (e.g. T at rs27097, G at rs37037, A at rs9941290 and G at rs863433) which were predicted from these results, and subsequently demonstrated experimentally, to lengthen QT interval, with the opposite haplotype predicted to shorten QT interval. The Effect column shows the observed effect in milliseconds (ms) on QT interval, and the Direction column shows the consistent direction of this effect in the six individual cohorts contributing to the QTSCD consortium.
Disease Models & Mechanisms: doi:10.1242/dmm.044727: Supplementary information
Dis
ease
Mo
dels
& M
echa
nism
s •
Sup
plem
enta
ry in
form
atio
n
Page 32
w1118 x CNOT1/ not1
Time (s) 10 5
20
0
-20
-40
-60
IN 0
(mV
)
0
20
40
60
80
100
120
M
Diastolic Diameter
0
20
40
60
80
M
Systolic Diameter*
0.0
0.2
0.4
0.6
%
Fractional Shortening **
Hand-Gal4 > CNOT1/ not1
Time (s)
20
0
-20
-40
-60
IN 0
(mV
)
10 5
C D E
A B
Figure S2
w11
18 x C
NOT1/not1
TinC∆4
> CNOT1/n
ot1
Figure S2. A-B) Electrophysiological recordings of CNOT1/not1 (VDRC, GD12571) knockdown Drosophila hearts. One single electrophysiology trace obtained from fragile Hand-Gal4 driven CNOT1/not1-knockdown fly hearts show extended events and multiple peaks per burst com-pared to control fly. C-E) TinCΔ4-Gal4 driven CNOT1/not1-knockdown increased systolic diameter and reduced fractional shortening. n=40 female flies per genotype. Student’s t-test was used to calculate two-sided p-values. Boxes: interquartile range,central line: median; plus: mean; whiskers: upper and lower adjacent values as defined by Tukey (1977); dots: outside values.
w11
18 x C
NOT1/not1
TinC∆4
> CNOT1/n
ot1
w11
18 x C
NOT1/not1
TinC∆4
> CNOT1/n
ot1
Disease Models & Mechanisms: doi:10.1242/dmm.044727: Supplementary information
Dis
ease
Mo
dels
& M
echa
nism
s •
Sup
plem
enta
ry in
form
atio
n
Page 33
0
20
40
60
80
100
120
M
Diastolic Diameter ****
0
20
40
60
80
M
Systolic Diameter
***
0.0
0.1
0.2
0.3
0.4
0.5
Fractional ShorteningA B C
Figure S3
TinCΔ
4> C
NOT7/8/ p
op2
TinCΔ
4> C
NOT4/ no
t4
TinCΔ
4> C
NOT2/ no
t2
TinCΔ
4> C
NOT6/6L/
twin
w11
18 x T
inCΔ4
TinCΔ
4> C
NOT7/8/ p
op2
TinCΔ
4> C
NOT4/ no
t4
TinCΔ
4> C
NOT2/ no
t2
TinCΔ
4> C
NOT6/6L/
twin
w11
18 x T
inCΔ4
TinCΔ
4> C
NOT7/8/ p
op2
TinCΔ
4> C
NOT4/ no
t4
TinCΔ
4> C
NOT2/ no
t2
TinCΔ
4> C
NOT6/6L/
twin
w11
18 x T
inCΔ4
%
Figure S3. A-C) RNAi-mediated knockdown of CNOT7/8/pop2 (TRiP HM05235), CNOT2/not2 (VDRC GD20826), CNOT4/not4 (TRiP JF03203) and CNOT6/6L/twin (VDRC GD13365) using cardiomyocyte specific driver TinC∆4-Gal4 (n=15, n=20, n=19, n=20 of female flies per respective genotype). Reduced expression of CNOT7/8/pop2, CNOT2/not2 and CNOT6/6L/twin had no effect on diastolic diameter, systolic diameter or fractional shortening. Reduced expression of CNOT4/not4 increased both systolic and diastolic diameters but had no effect on contractility. One-way ANOVA with Tukey’s multiple comparisons test was used to calculate two-sided p-values. Boxes: interquartile range, central line: median; plus: mean; whiskers: upper and lower adjacent values as defined by Tukey (1977); dots: outside values.
Disease Models & Mechanisms: doi:10.1242/dmm.044727: Supplementary information
Dis
ease
Mo
dels
& M
echa
nism
s •
Sup
plem
enta
ry in
form
atio
n

































![Shukla, Lena and Ajram, Laura A. and Begg, Malcolm and ... · 1 2,8-Diazaspiro[4.5]decan-8-yl)pyrimidin-4-amine Potent CCR4 Antagonists Capable of Inducing Receptor Endocytosis. Lena](https://static.documents.pub/doc/80x56/607709dc4d4a0b4d06695c94/shukla-lena-and-ajram-laura-a-and-begg-malcolm-and-1-28-diazaspiro45decan-8-ylpyrimidin-4-amine.jpg)














