JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY 69(4) suppl., 1996, pp. 85-97 Social Behavior and Communication in the Neotropical Cicada Fidicina mannifera (Fabricius) (Homoptera: Cicadidae) Reginald B. Cocroft1 Department of Zoology, University of Texas, Austin, Texas 78712 Michael Pogue Department of Entomology, National Museum of Natural History, Smithsonian Institution, Washington, D.C. 20560 abstract: We investigated signalling behavior in relation to social context in the cicada Fidicina mannifera. Four types of acoustic signals were recorded from males: songs, calls, low-amplitude songs, and disturbance sounds. Signal types were similar in frequency and fine temporal structure, but differed markedly in gross temporal structure. Songs and calls were the most frequently observed signals, and these differed in the social context in which they were used. When inter-male distances were large, males produced songs, overlapping them with songs of neighboring males. As nearest-neighbor distance decreased, males switched to calls, which were typically alternated back and forth between males. Bouts of calls often were followed by a parallel walk display, which apparently was involved in settling disputes over calling sites. Singing cicadas are a prominent feature of the acoustic environment in many temperate and tropical areas. Cicada songs are diverse and often have provided a basis for taxonomic decisions and species identification (Jiang, 1985; Joermann and Schneider, 1987; Villet, 1988; Fonseca, 1991; Daniel et al., 1993). Further more, the mechanisms of sound production and sound reception have been well studied (Pringle, 1954; Moore and Sawyer, 1966; Simmons and Young, 1978; Josephson and Young, 1979, 1981; Huber, 1983; Young and Josephson, 1983; Popov and Sergeeva, 1987; Young, 1990; Popov, 1990; Bennet-Clark and Young, 1992; Fonseca and Popov, 1994; Henning et al., 1994a, b). In contrast, there are relatively few studies of the social behavior of cicadas in the field (Alexander and Moore, 1958, 1962; Claridge et al., 1979; Doolan, 1981; Gwynne, 1987; Doolan and Young, 1989; Villet, 1992), in part because many species sing from the forest canopy and are difficult to observe. As a result, the functional aspects of most cicada communication systems remain poorly known. In this study we describe the social behavior of a neotropical cicada, Fidicina mannifera (Fabricius). This species provides a rare opportunity to study cicada behavior in the field, because most of its activity takes place on tree trunks well below the canopy. In addition to describing the signals used by males, we examine the social context in which two of the most commonly-used signals are given. We also present previously unreported aspects of their social behavior. Materials and Methods The study was conducted at the BIOLAT Biological Station, located at Puesto de Vigilancia Pakitza in the Zona Reservada del Manu, about 10 km south of the 1 Present address: Section of Neurobiology and Behavior Mudd Hall, Cornell University, Ithaca, New York 14853. This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PM All use subject to JSTOR Terms and Conditions
Transcript
JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY 69(4) suppl., 1996, pp. 85-97
Social Behavior and Communication in the Neotropical Cicada Fidicina mannifera (Fabricius) (Homoptera: Cicadidae)
Reginald B. Cocroft1
Department of Zoology, University of Texas, Austin, Texas 78712
Michael Pogue
Department of Entomology, National Museum of Natural History, Smithsonian Institution, Washington, D.C. 20560
abstract: We investigated signalling behavior in relation to social context in the cicada Fidicina mannifera. Four types of acoustic signals were recorded from males: songs, calls,
low-amplitude songs, and disturbance sounds. Signal types were similar in frequency and fine temporal structure, but differed markedly in gross temporal structure. Songs and calls were the most frequently observed signals, and these differed in the social context in which
they were used. When inter-male distances were large, males produced songs, overlapping them with songs of neighboring males. As nearest-neighbor distance decreased, males switched to calls, which were typically alternated back and forth between males. Bouts of calls often were followed by a parallel walk display, which apparently was involved in
settling disputes over calling sites.
Singing cicadas are a prominent feature of the acoustic environment in many temperate and tropical areas. Cicada songs are diverse and often have provided a
basis for taxonomic decisions and species identification (Jiang, 1985; Joermann and Schneider, 1987; Villet, 1988; Fonseca, 1991; Daniel et al., 1993). Further
more, the mechanisms of sound production and sound reception have been well studied (Pringle, 1954; Moore and Sawyer, 1966; Simmons and Young, 1978; Josephson and Young, 1979, 1981; Huber, 1983; Young and Josephson, 1983; Popov and Sergeeva, 1987; Young, 1990; Popov, 1990; Bennet-Clark and Young, 1992; Fonseca and Popov, 1994; Henning et al., 1994a, b). In contrast, there are
relatively few studies of the social behavior of cicadas in the field (Alexander and Moore, 1958, 1962; Claridge et al., 1979; Doolan, 1981; Gwynne, 1987;
Doolan and Young, 1989; Villet, 1992), in part because many species sing from the forest canopy and are difficult to observe. As a result, the functional aspects
of most cicada communication systems remain poorly known.
In this study we describe the social behavior of a neotropical cicada, Fidicina mannifera (Fabricius). This species provides a rare opportunity to study cicada behavior in the field, because most of its activity takes place on tree trunks well below the canopy. In addition to describing the signals used by males, we examine the social context in which two of the most commonly-used signals are given.
We also present previously unreported aspects of their social behavior.
Materials and Methods
The study was conducted at the BIOLAT Biological Station, located at Puesto de Vigilancia Pakitza in the Zona Reservada del Manu, about 10 km south of the
1 Present address: Section of Neurobiology and Behavior Mudd Hall, Cornell University, Ithaca, New York 14853.
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
86 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
Parque Nacional del Manu (11?55'48"S, 71?15'18"W; elevation 350 m). This area is in the department of Madre de Dios in southeastern Peru. The study was con
ducted from 26 September to 25 October, 1990, during the transition from the
dry to the wet season.
Fidicina mannifera is widely distributed, ranging from Costa Rica to Argentina (Distant, 1883; Jacobi, 1907; Goding, 1925; Torres, 1953; Young, 1976). In the
moist tropical forests of southeastern Peru, adults of F. mannifera have been collected from 12 September to 25 November, which includes the late dry season and early rainy season. At Pakitza, adults occur on trees in non-flooded forest
(especially old alluvial terrace forest) and in the clearing surrounding the build
ings. Males call on tree trunks during sunny periods in small aggregations on one or several nearby trees. Males also form a pronounced evening chorus.
Sounds were recorded using either a Sony TCD-5M or a Marantz PMD 420 cassette recorder with a Sennheiser ME-80 microphone and K3U power module. Both recorder-microphone combinations provided a flat frequency response up to 14 kHz. Air temperature at or near the recorded male's calling site was measured to the nearest 0.2?C with a Miller & Weber quick-reading thermometer. Sound pressure level was measured with a Realistic Sound Pressure Level meter, using the Fast response setting and A weighting.
Recordings were analyzed using a DATA 6000A Universal Waveform Analyzer for frequency and fine-temporal measurements and a Multigon Uniscan II Real Time Analyzer for gross temporal measurements. The sampling rate for waveform
analysis was 40 kHz. Fourier transforms were performed on samples of 8192
points, and thus the frequency resolution (sampling rate/number of points) was about 5 Hz. In practice, factors such as small variations in tape recorder speed are likely to lessen the actual level of resolution.
Five-minute observational samples of calling behavior were made on focal males and their nearest calling neighbors. Signals emitted during the sample pe
riod were counted and scored as songs or calls (these signal types were differ
entiated easily on the basis of temporal differences; see below). Distance between males was measured with a tape measure or, for pairs of males on the same tree
but too high to measure, estimated to the nearest 0.5 m. The proportion of calls vs. songs was arcsine-transformed and regressed on the log of nearest-neighbor distance.
Signals also were scored as overlapping or alternating with signals of the neigh boring male. For each observational sample we calculated the average signal duty cycle, and, using an average signal length, we calculated the proportion of signals expected to overlap due to chance (i.e., if males were signalling randomly with
respect to each other). This expectation was then compared with the overlap observed during each sample. Each focal male was scored as overlapping more or less than would be expected due to chance, then used as a single data point in a binomial probability test. Preliminary observations had suggested the hypoth eses that songs were actively overlapped with songs of neighboring males, while calls were actively alternated, and thus the binomial tests used were one-tailed.
Results
Sizes of male and female F. mannifera are reported in Table 1. Measurements were made from specimens collected at the study site and deposited in the col lection of the Department of Entomology, Smithsonian Institution.
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
VOLUME 69, NUMBER 4 87
Table 1. Dimensions of male and female Fidicina mannifera from Pakitza, Peru (x ? SD). All measurements are in millimeters.
Body Head Pronotum Mesonotum Tegmen Sex Length Width Width Width Length N
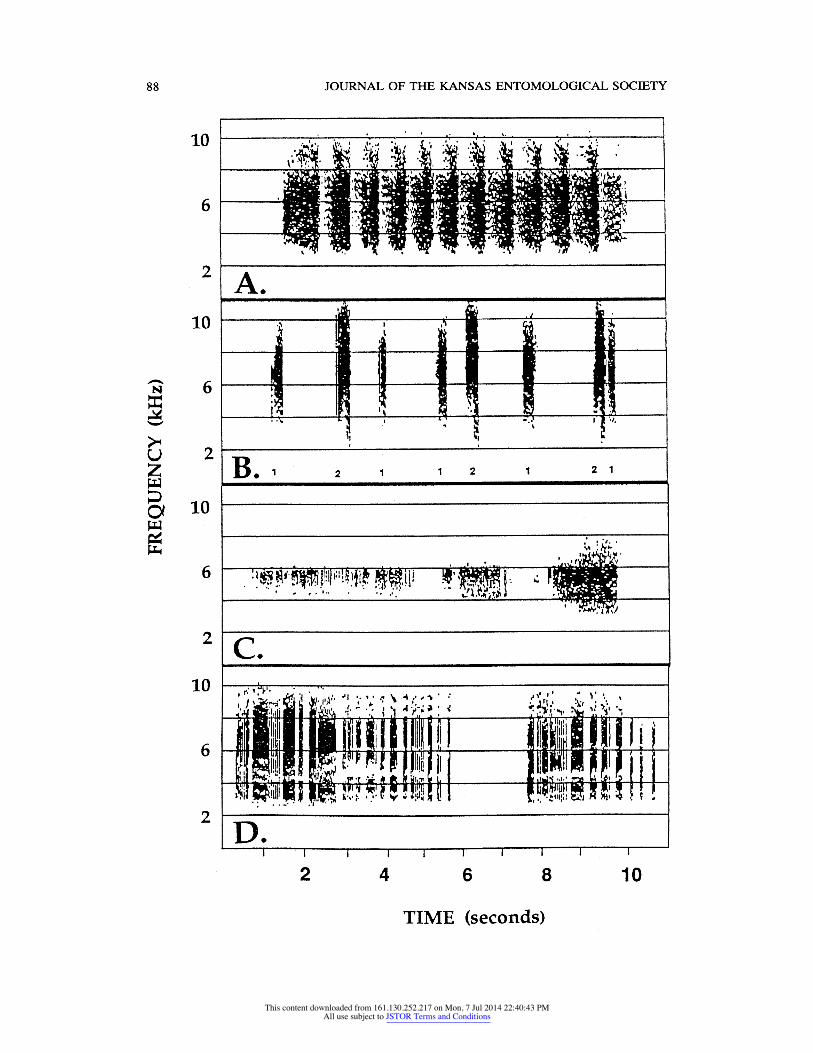
We recorded four distinct types of acoustic signals from male F. mannifera. Signal types were similar in spectral characteristics: males produced relatively broadband sounds with energy concentrated between 3 and 9 kHz, with a mean dominant frequency between 4.6 and 5.6 kHz (Table 2).
Gross temporal features, in contrast, differed markedly between signals. Two
signals were given much more frequently than the others. The first of these was the song (or "calling song"; Fig. IA, Table 2). This was the longest signal, with a mean length of 8.6 sec. Song length varied considerably between males and between different songs of the same male, spanning an overall range of 2.4-13.6 seconds. Songs were characterized by an initial, gradual rise in amplitude, fol lowed by pronounced, regular amplitude modulation. Each rise in amplitude was
accompanied by a raising of the abdomen, and the rate of amplitude modulation (and of corresponding raising and lowering of the abdomen) sometimes increased or decreased slightly within the song. The length of the initial buildup in ampli tude was variable within and between males, and songs occasionally began with
out it.
Songs were produced at a mean rate of 1.35 per minute during periods of active
singing. Song rate within males was variable, with consecutive intervals between songs of an individual male varying by up to 51%. The maximum sound pressure level of song had a mean of 91.9 dB at 1 m (ref. 0.0002 dynes/cm2). Because the
meter used for this measurement provided A weighting rather than flat weighting, this value should be considered only as a measure of relative intensity with respect to other signals of similar frequency content (see below).
The second of the two commonly heard signals was the call (Figs. IB, 2A; Table 2). Calls consisted of a short burst of sound with a mean length of 0.34 sec. In most cases, amplitude was low at the start and rapidly increased to a peak
Table 2. Characteristics of songs and calls (jc ? SE) of male Fidicina mannifera from Pakitza, Peru. Sample sizes are reported as # males/# calls. *Note: for SPL measurements, N = 1 males (song),
N = 4 males (call). SE values are not symmetrical as measurements were converted into pressure, averaged, and re-converted into dB.
Amplitude Dominant Pulse Rate Modu- Signal Sound
Rise Fre- _ lation Repetition Pressure Af Signal Duration Time quency Min Rate Rate Level* (males/ Type (ms) (ms) (Hz) (/s) Max. (/s) (/min) (dB) calls)
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
88 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
-|n -_:?.-1,-_:-1-*;-!*-.-??-^-^
MM J-iJJJJ J JJU 1
u 2.r=-1-! -
Z I 1J? 1 2 1_1 2_1_21_
a io ?:?
w ~s -_
r^ ? : J'
2{c. I j . ' . ? .mmt???<~4
10 ,< >. .-.,'-.?-,, ? . t,, -
lllllfe*sd|jl?fa Sid!!
21 p."";' ... i -1-1-1-1-1-r-1-1-1?-1
2 4 6 8 10
TIME (seconds)
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
VOLUME 69, NUMBER 4 89
A.
B.
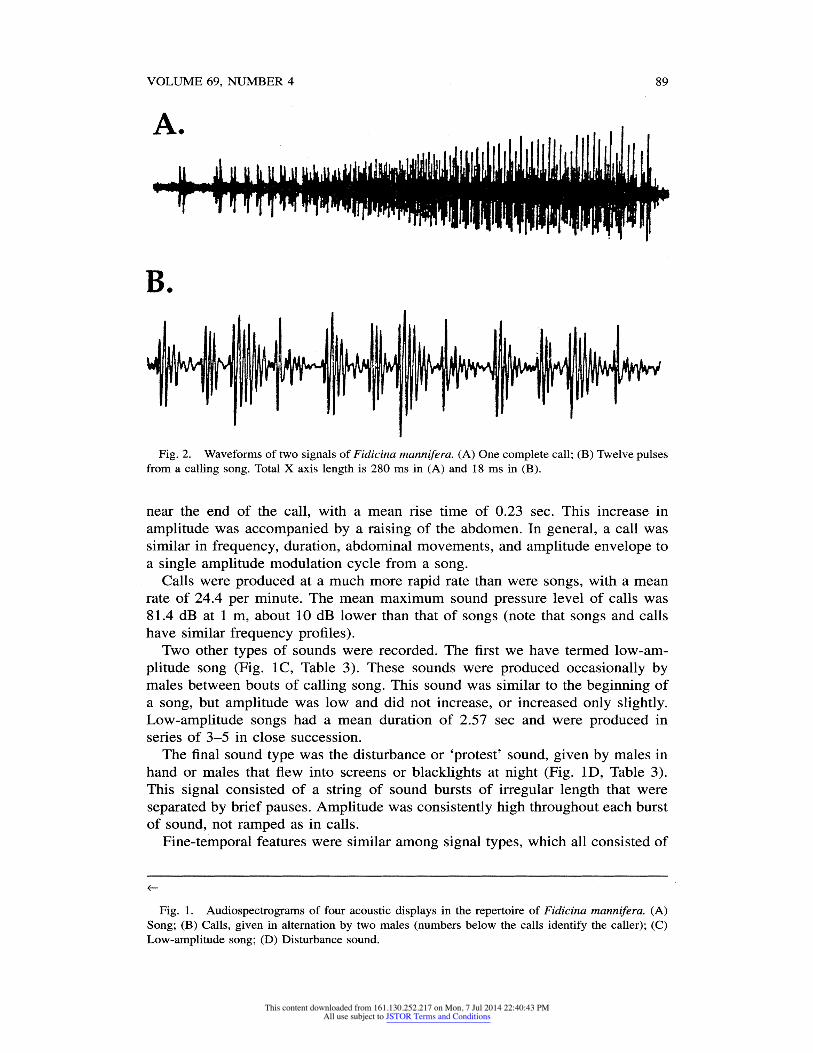
Fig. 2. Waveforms of two signals of Fidicina mannifera. (A) One complete call; (B) Twelve pulses from a calling song. Total X axis length is 280 ms in (A) and 18 ms in (B).
near the end of the call, with a mean rise time of 0.23 sec. This increase in
amplitude was accompanied by a raising of the abdomen. In general, a call was similar in frequency, duration, abdominal movements, and amplitude envelope to a single amplitude modulation cycle from a song.
Calls were produced at a much more rapid rate than were songs, with a mean rate of 24.4 per minute. The mean maximum sound pressure level of calls was 81.4 dB at 1 m, about 10 dB lower than that of songs (note that songs and calls have similar frequency profiles).
Two other types of sounds were recorded. The first we have termed low-am
plitude song (Fig. 1C, Table 3). These sounds were produced occasionally by males between bouts of calling song. This sound was similar to the beginning of a song, but amplitude was low and did not increase, or increased only slightly. Low-amplitude songs had a mean duration of 2.57 sec and were produced in series of 3-5 in close succession.
The final sound type was the disturbance or 'protest' sound, given by males in hand or males that flew into screens or blacklights at night (Fig. ID, Table 3). This signal consisted of a string of sound bursts of irregular length that were
separated by brief pauses. Amplitude was consistently high throughout each burst of sound, not ramped as in calls.
Fine-temporal features were similar among signal types, which all consisted of
<
Fig. 1. Audiospectrograms of four acoustic displays in the repertoire of Fidicina mannifera. (A)
Song; (B) Calls, given in alternation by two males (numbers below the calls identify the caller); (C)
Low-amplitude song; (D) Disturbance sound.
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
90 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
Table 3. Characteristics of low-amplitude songs and disturbance sounds of male Fidicina mannifera from Pakitza, Peru. Sample sizes and values are reported as in Table 2.
Dominant Pulse Duration Frequency Rate N
Signal TVpe (ms) (Hz) (/s) (males/calls)
Low-Amplitude 2567 4686 499.0 3/11
Song (?738) (?1271) (?35.5) Protest 211 4633 499.5 1/5
Sound ? ? ?
trains of pulses (Fig. 2B). Individual pulses had a rapid onset and exponential decay, with a mean total duration of 1.3 msec. Pulse repetition rate varied with signal amplitude. In calls, pulse rate changed slightly, with a mean initial rate of 602.5 per sec increasing to a maximum of 613.8 per sec, coinciding approximately with the peak amplitude of the call. In songs, pulse rate underwent regular in creases and decreases, with mean pulse rates of 479.4 per sec during amplitude
minima and 639.2 per sec during amplitude maxima. ambient temperature and song structure: In this study, pulse rate was not
correlated with air temperature near the calling site (r = 0.00, d.f. = 8, NS). Although this sample size is small, the similarity of maximum pulse rates over a 7?C range in ambient temperature suggests that different males achieved a similar
tymbal muscle temperature during singing. Josephson and Young (1979) showed that tymbal muscle temperature in the bladder cicada {Cystosoma saundersii) in creased by about 12? over ambient temperature during continuous singing. Fur thermore, some species of cicadas thermoregulate by basking (Heath and Wilkin, 1970) and/or evaporative cooling (Toolson and Hadley, 1987; Hastings and Tool son, 1991; Heinrich, 1993), although it is unknown whether these mechanisms occur in F. mannifera (Bartholomew and Barnhart, 1984).
variation in signalling behavior: Male signalling behavior varied markedly with social context. As the distance between neighboring sound-producing males
increased, males gave proportionally more songs and fewer calls (r =
0.88, d.f. = 17, P < 0.001; Fig. 3). At distances of one meter or less, males gave calls almost exclusively. Between 2 and 5 m, males began giving songs as well as calls, while at greater distances males primarily produced songs. A threshold effect is suggested by the relatively sharp drop in the proportion of calls after 3 m.
Signalling strategy also differed between songs and calls (Table 4). Males over
lapped songs with those of neighboring males significantly more than expected if
they were singing randomly with respect to one another. In fact, almost all songs were overlapped with songs of neighbors. Even though the intervals between successive songs were variable (with a mean intersong interval of 44 sec), the
average delay time between the beginnings of songs of two neighboring males was only 1.3 sec. Each male essentially tuned its singing rate to that of its neigh bors. The chorusing pattern of a group of males consisted of a wave of singing as one male started and successively more distant neighbors responded, followed
by a period of silence, then another wave of singing, etc.
Calls, in contrast, were primarily alternated rather than overlapped between members of pairs of calling males (Table 4; cf. Fig. IB). Evidence from this study
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
VOLUME 69, NUMBER 4 91
? -\- . s v ? 0.8- \ \ CD \
< 0.6 -i \ U \ fe 1 \
I X o 02" ^^
o ^^^ g o.o
-1? ? ? ? ? ? ? ? ?' ?^ ? ? ?
???'? ? ? ?
0 5 10 15 20
DISTANCE BETWEEN MALES (m) Fig. 3. Proportion of calls vs. songs given by male Fidicina mannifera in relation to nearest
neighbor distance. Note that the relationship with distance is not linear (fitted curve is logarithmic).
is inconclusive about whether this involved active avoidance of call overlap by neighboring males. However, even though calls were given at much faster rates than were songs, the short duration of calls makes it probable that most would not overlap even if males did not actively avoid giving their calls while another
male is calling. In order for significant overlap to occur, males would have had to actively overlap calls, which they clearly did not.
At least during daytime singing, males typically gave series of songs while
remaining at the same position on a tree trunk. For example, during 5-min ob servation periods, males gave from 3 to 12 (mean = 6.5) songs without changing
position. Counts were not made beyond the 5-min period, so the maximum num
ber of successive songs given by stationary males was not determined.
In addition to daytime singing, males also formed evening choruses during the half-hour before dusk. During these evening activity periods, males changed po sition more frequently, seldom giving more than 2 or 3 calls before flying to a new position. On two occasions, most of the individuals in an area were seen to
gradually converge onto a single tree (a behavior seen in Fidicina pronoe by
Table 4. Signal timing strategies of male Fidicina mannifera. Value represents number of focal males overlapping or alternating signals more than would be expected if males placed signals randomly with respect to signalling neighbors.
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
92 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
Moore (1962)). By the end of the evening activity period, these focal trees were
occupied by 19 individuals on one occasion and 26 on the other; nearby trees (5? 7 m away) that previously had served as calling sites were unoccupied. Many of these individuals were singing (and, especially, calling) males; the proportion of females in these aggregations is unknown, but we did not observe courtship or
copulation. This density of individuals on a single tree was far higher than that seen during the day.
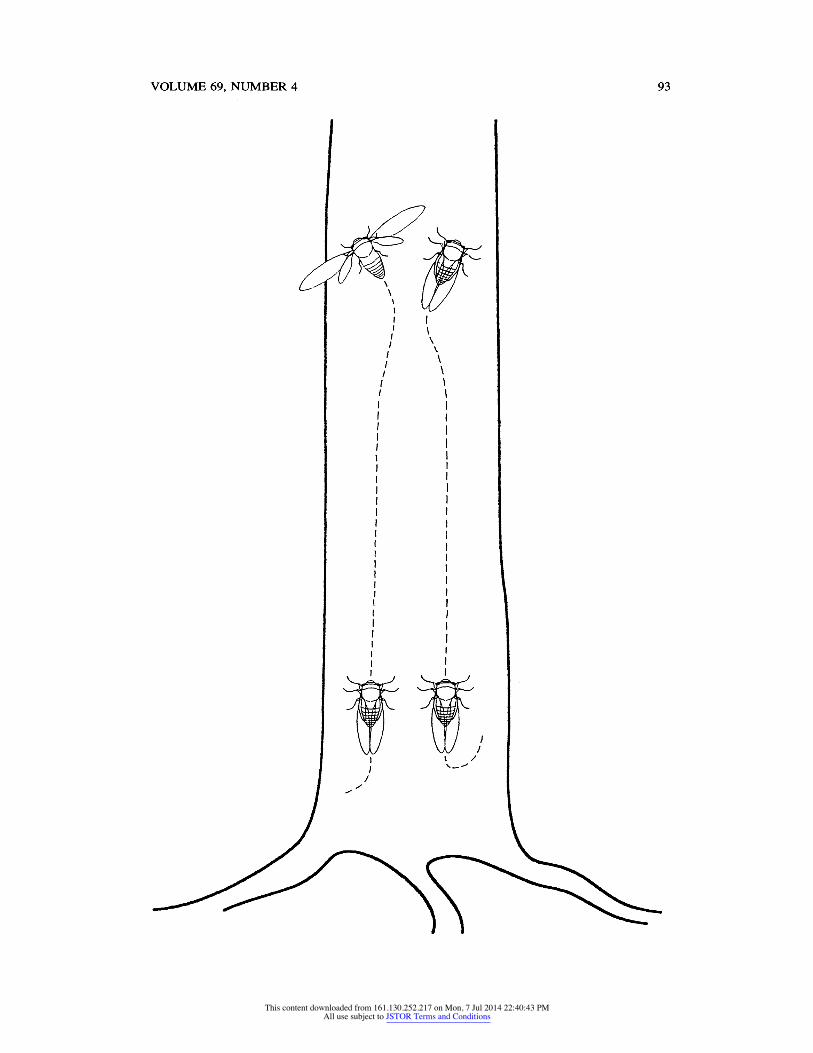
visual display: Males engaged in a stereotyped visual display that apparently was involved in settling disputes over calling sites (Af = 20 observed). This dis
play, which we have termed a parallel walk display, was performed in a coordi nated manner between two males (Fig. 4).
Signalling males less than 1 m apart did not produce songs, but instead called back and forth. After a bout of calling, one male often flew to another tree, moved to a different area of the same tree, or became silent. However, another frequent outcome was for the two males to approach each other until they were side by side, facing vertically up the tree trunk. They then walked up the trunk side by side, one occasionally jostling the other, for about 25 cm. In each of the 20 observed occasions, one of the males then flew away, either to a different tree or to elsewhere on the same tree. In each of 5 cases in which prior residency of one of the males was known, the male that arrived second was the one that left the site. The entire display lasted 3 to 5 seconds. We observed no other obvious
components to this display; for example, the males' wings appeared to be held at rest during the vertical walk. Males did not call during this display.
Discussion
Repertoire composition in Fidicina mannifera is similar to that of other cicadas whose signals have been reported. In particular, calling songs given by males in the field and disturbance sounds are widespread (Alexander and Moore, 1958;
Alexander, 1960; Moore, 1962; Simmons and Young, 1978; Smith and Langley, 1978; Josephson and Young, 1981; Young and Josephson, 1983; Villet, 1988;
Young, 1990; Fonseca, 1991; Villet, 1992; Daniel et al., 1993). Distinct courtship songs also have been reported in some species and may be typical of cicadas (Alexander and Moore, 1958; Alexander, 1960, 1967, 1968; Simmons and Young, 1978; Gwynne, 1987; Fonseca, 1991), but we did not witness pairing in F. man
nifera and thus have no information about sound production in this context by either males or females.
Other call types have been less thoroughly investigated. Aidley (1969; see also
Enger et al., 1969) describes a "zeep" call of F. mannifera (=rana) from Brazil that is similar to the call reported here for F. mannifera from Peru (Note: although these taxa have been synonymized, Aidley [1969] describes the calling song as "an uninterrupted train of clicks lasting from 2 to 20 sec, of a constant and very loud sound intensity." Given the apparent differences between the songs of F.
mannifera from Peru and Brazil, distinct taxa may be involved). Fonseca (1991) describes a male-male interaction signal of Cicada barbara lusitanica that appears
Fig. 4. Parallel walk display between two male Fidicina mannifera on the same tree trunk.
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
VOLUME 69, NUMBER 4 93
r I
/ \
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
94 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
similar in structure and function to the F. mannifera call. We also observed a
similar call of an undescribed Fidicina from Pakitza, Peru. We are unaware of reports of other cicada species with signals similar to the low-amplitude songs reported here in F. mannifera, and the role of these sounds in communication (if any) remains unclear.
Songs and calls are the two signals most frequently given by males in the field, and these differ in the social context in which they are produced. Songs in cicadas have been demonstrated to function in mate attraction (Alexander and Moore, 1958; Claridge et al., 1979; Doolan, 1981; Doolan and Young, 1989; Villet, 1992).
We have no data on female phonotactic behavior in F. mannifera, but the song in this species appears to function as a long-distance advertisement signal, as in other cicadas. First, it apparently is a long-distance signal, since it is primarily given when males are distant from other individuals. Second, most of a male's song is overlapped by songs of neighboring males. It is unlikely that a male can
perceive another male's signal while singing: much of a male's sound is radiated
through its tympana (Young, 1990; Fonseca and Popov, 1994), and the hearing sensitivity of male cicadas is reduced during singing by folding of the tympanal
membranes (Hennig et al., 1994a). Thus, although males coordinate the timing of their songs using the beginning of a neighbor's song, most of the signal will not be perceived by other males, suggesting that male-male communication is not its
primary function. The function of calls, as opposed to songs, has been less well documented in
other cicada species. In F. mannifera, calls are produced at close range by inter
acting males. Pairs of males that begin producing songs within a few meters of each other switch to calls; after a bout of calling, one or both males may become silent, one male may fly to another tree, or the two males may engage in a parallel
walk display, after which one of the males leaves the site. Calls thus appear to be aggressive signals involved in competitive interactions between males. A male
male interaction signal also has been reported for Cicada barbara lusitanica (Fon
seca, 1991); as in F. mannifera, this signal is given in close-range interactions, as when a second male begins producing calling song near a singing male. An alternative hypothesis for explaining the switch to a different signal type when distances between males are small is suggested by Ryan (1985) for frogs: when vocal competition increases, males may be forced to give an otherwise costly call that is more attractive to females. This function is unlikely in F. mannifera, both because bouts of calling regularly precede active, aggressive interactions between
males and because males primarily produce songs, regardless of chorus size, un
less two males begin singing within a few meters of each other. In the tick-tock cicada {Cicadetta quadricincta), males employ a "call-fly"
strategy, remaining at a given calling site for less than a minute before flying to a new site (Gwynne, 1987). This strategy is characteristic of at least some other cicadas (Alexander, 1960; Moore, 1962) as well as of many Auchenorrhyncha that communicate using substrate-borne vibrations (Hunt and Nault, 1991; Tish
echkin, 1994; Cocroft, unpub. data). This behavior has been interpreted as a mech anism for increasing the active space of a male's signalling effort (Gwynne, 1987;
Hunt and Nault, 1991). In contrast, singing male F. mannifera remain at a single site for extended periods, at least during daytime singing. Furthermore, males
apparently defend these sites against other males by means of calls and visual
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
VOLUME 69, NUMBER 4 95
displays. Male bladder cicadas (Cystosoma saundersii) also remain at one singing site for extended periods (Doolan, 1981). These differences in calling strategy among species remain to be explained, and presumably depend on a number of inter-related factors including the active space of the song, the nature and intensity of predation, and the distribution and behavior of females. Doolan (1981) sug gested that the sedentary singing behavior of C. saundersii may be related to
possession of an enlarged abdomen (used in sound radiation) that limits their
flying ability. However, the direction of evolutionary causality between sedentary singing and an enlarged radiating structure that hampers flight is not clear. Fidi cina mannifera appear to change singing sites more frequently during the evening than during the day, but the social and/or ecological correlates of this difference are unknown.
Songs and calls differ not only in the social context in which they are used, but also in structure. The song, a long-range signal, is higher in intensity (about 10 dB SPL) than the call. Given that the two signals are similar in frequency
composition, songs will have a larger active space. This difference between the two signal types is not unexpected, given that songs are used over longer distances than are calls.
Songs and calls also differ in the timing strategy used by signalling males. A number of studies have established differences among calling species in whether
signals are overlapped or alternated with signals of neighboring males (e.g., Greenfield and Shaw, 1983; Sullivan, 1985). In F. mannifera, calling strategy changes depending on the kind of signal a male is producing; calling songs are
actively overlapped by neighboring males, while calls are alternated (or at least not actively overlapped).
The parallel walk display performed by pairs of male F. mannifera recalls
displays performed in a similar context in other taxa, including red deer, giraffes, and thrips (Clutton-Brock and Albon, 1979; Walther, 1984; Crespi, 1986). We are unaware of reports of similar parallel walk displays in other cicadas. If this be havior is in fact analogous in these examples, it would represent a convergent visual display across a strikingly broad range of taxa.
Understanding the function of communication signals requires a two-part pro cess. One line of evidence is provided by description of the context in which these signals are used in the field, as was done in this study for signals of F.
mannifera. Complementary evidence is provided by experimental studies of the response of receivers to these signals?a line of evidence that has proved even
more difficult to obtain than observational evidence for most cicadas. In any case, our observational study has narrowed the range of hypotheses of function for the communication signals of this cicada species, and revealed a rich repertoire of social signals that may turn out to be characteristic of cicadas as the behavior of
more species becomes known.
Acknowledgments This study was supported by BIOLAT research grants to the authors for work
at Pakitza. We thank the Direccion General Forestal y de Fauna, Ministerio de
Agricultura, Lima, Peru for permission to work at Pakitza. We also thank R. Burnham, M. Ryan, two anonymous reviewers and G. C. Eickwort for providing comments on the manuscript, and G. Venable for producing Fig. 4. This is con
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
96 JOURNAL OF THE KANSAS ENTOMOLOGICAL SOCIETY
tribution No. 25, Biological Diversity in Latin America (BIOLAT) Project, Smith sonian Institution.
Literature Cited
Aidley, J. D. 1969. Sound production in a Brazilian cicada. J. Exp. Biol. 51:325-337.
Alexander, R. D. 1960. Sound communication in Orthoptera and Cicadidae. In W. E. Lanyon and W. N. Tavolga (eds.), Animal Sounds and Communication, pp. 38-92. American Institute of Bio
logical Sciences, Washington, D.C.
Alexander, R. D. 1967. Acoustical communication in arthropods. Ann. Rev. Entomol., 12:495-526.
Alexander, R. D. 1968. Arthropods. In T A. Sebeok (ed.), Animal Communication: Techniques of
Study and Results of Research, pp. 167-216. Indiana University Press, Bloomington. Alexander, R. D., and T. E. Moore. 1958. Studies on the acoustical behavior of seventeen-year cicadas
(Homoptera: Cicadidae: Magicicada). Ohio J. Sci. 58(2): 107-127.
Alexander, R. D., and T. E. Moore. 1962. The evolutionary relationships of 17-year and 13-year cicadas and three new species (Homoptera: Cicadidae: Magicicada). Univ. Mich. Mus. Zool.
Misc. Publ. 121:5-59.
Bartholomew, G. A., and M. C. Barnhart. 1984. Tracheal gases, respiratory gas exchange, body temperature and flight in some tropical cicadas. J. Exp. Biol. 111:131-144.
Bennet-Clark, H. C, and D. Young. 1992. A model of the mechanism of sound production in cicadas.
J. Exp. Biol. 173:123-153.
Claridge, M. E, M. R. Wilson, and J. S. Singhrao. 1979. The songs and calling sites of two European cicadas. Ecol. Entomol. 4:225-229.
Clutton-Brock, T. H. and S. D. Albon. 1979. The roaring of red deer and the evolution of honest advertisement. Behaviour 69:145-169.
Crespi, B. J. 1986. Size assessment and alternative fighting tactics in Elaphrothrips tuberculatus
Doolan, J. M., and D. Young. 1989. Relative importance of song parameters during flight phonotaxis and courtship in the bladder cicada Cystosoma saundersii. J. Exp. Biol. 141:113-131.
Enger, P. S., D. J. Aidley, and T Szabo. 1969. Sound reception in the Brazilian cicada Fidicina rana
Walk. J. Exp. Biol. 51:339-345.
Fonseca, P. J. 1991. Characteristics of the acoustic signals in nine species of cicadas (Homoptera, Cicadidae). Bioacoustics 3:173-182.
Fonseca, P. J., and A. V. Popov. 1994. Sound radiation in a cicada: the role of different structures. J.
Comp. Physiol. A 175:349-361.
Goding, F. W. 1925. Synopsis of the Cicadidae of Ecuador. Rev. Col. Nac. Vicente Rocafuerte 7:1-34.
Greenfield, M. D., and K. C. Shaw. 1983. Adaptive significance of chorusing with special reference
to the Orthoptera. In D. T. Gwynne, and G. K. Morris (eds.), Orthopteran Mating Systems, pp. 1-27. Westview Press, Boulder.
Gwynne, D. T 1987. Sex-biased predation and the risky mate-locating behaviour of male tick-tock
Hastings, J. M., and E. C. Toolson. 1991. Thermoregulation and activity patterns of two syntopic cicadas, Tibicen chiricahua and T. duryi (Homoptera: Cicadidae), in central New Mexico.
Oecologia 85:513-520.
Heath, J. E., and P. J. Wilkin. 1970. Temperature responses of the desert cicada, Diceroprocta apache
Heinrich, B. 1993. The Hot-Blooded Insects: Strategies and Mechanisms of Thermoregulation. Har
vard University Press, Cambridge.
Hennig, R. M., T. Weber, F. Huber, H. U. Kleindienst, T. E. Moore, and A. V. Popov. 1994a. Auditory threshold change in singing cicadas. J. Exp. Biol. 187:45-55.
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions
VOLUME 69, NUMBER 4 97
Hennig, R. M., T. Weber, T. E. Moore, F. Huber, H. U. Kleindienst, and A. V Popov. 1994b. Function of the tensor muscle in the cicada Tibicen linnet J. Exp. Biol. 187:33-44.
Huber, F. 1983. Neural correlates of orthopteran and cicada phonotaxis. In F. Huber and H. Markl
(eds.), Neuroethology and Behavioral Physiology: Roots and Growing Points, pp. 108-135.
Springer-Verlag, Berlin.
Hunt, R. E., and L. R. Nault. 1991. Roles of interplant movement, acoustic communication, and
phototaxis in mate-location behavior of the leafhopper Graminella nigrifrons. Behav. Ecol. Sociobiol. 28:315-320.
Jacobi, A. 1907. Homoptera Andina. Die Zikaden des Kordillerengebietes von Sudamerika nach
Systematik und Verbreitung. 1. Cicadidae. (Dresden) K. Zool. u. Antrhop.-Ethnog. Mus. Ab handl. u. Ber. 11:1-28.
Jiang, J. C. 1985. A study of the song characteristics in cicadas at Jinghong in Yunnan province, China. Acta Entomol. Sinica 28:257-265.
Joermann, G., and H. Schneider. 1987. The songs of four species of cicada in Yugoslavia (Homoptera: Cicadidae). Zoologischer Anzeiger 218:283-296.
Josephson, R. K., and D. Young. 1979. Body temperature and singing in the bladder cicada, Cysto soma saundersii. J. Exp. Biol. 80:69-81.
Josephson, R. K., and D. Young. 1981. Synchronous and asynchronous muscles in cicadas. J. Exp. Biol. 91:219-237.
Moore, T. E. 1962. Acoustical behavior of the cicada Fidicina pronoe (Walker) (Homoptera: Cicad
idae). Ohio J. Sci. 62(3):113-119. Moore, T. E., and R. T. Sawyer. 1966. The mechanism of cicada timbal action (Insecta: Homoptera:
Cicadidae). Am. Zool. 6:509.
Popov, A. V. 1990. Co-evolution of sound-production and hearing in insects. In F. G. Gribalkin, K.
Wiese, and A. V Popov (eds.), Sensory Systems and Communication in Arthropods. Birkhauser
Verlag, Basel.
Popov, A. V, and M. V Sergeeva. 1987. Sound signalization and hearing in the Baikal cicada Cicadetta yezoensis (Homoptera: Cicadidae). Zoologischeskii Zhurnal 66:681-691.
Pringle, J. W S. 1954. A physiological analysis of cicada song. J. Exp. Biol. 32:525-560.
Ryan, M. J. 1985. The Tungara Frog: A Study in Sexual Selection and Communication. Univ. of
Chicago Press, Chicago. Simmons, P., and D. Young. 1978. The tymbal mechanism and song patterns of the bladder cicada,
Cystosoma saundersii. J. Exp. Biol. 76:27-45.
Smith, R. L., and W M. Langley. 1978. Cicada stress sound: an assay of its effectiveness as a predator defense mechanism. Southwest. Nat. 23:187-196.
Sullivan, B. K. 1985. Male calling behavior in response to playback of conspecific advertisement calls in two bufonids. J. Herpetol. 19(l):78-83.
Tishechkin, D. 1994. Acoustic communication system in Macropsinae leafhoppers (Homoptera: Ci
cadellidae): adaptive features and trend of evolution. Entomologisheckoe Obozrenie 73:3-15.
Toolson, E. C, and N. E. Hadley. 1987. Energy-dependent facilitation of transcuticular water flux contributes to evaporative cooling in the Sonoran Desert cicada, Diceroprocta apache (Ho
moptera: Cicadidae). J. Exp. Biol. 131:439-444.
Torres, B. A. 1953. Estudio biologico sobre Fidicina mannifera (Fab., 1803) y su importancia econ omica en la Argentina. Rev. Museo Ciudad Eva Peron (N. S.) 6(Zool.):285-295.
Villet, M. 1988. Calling songs of some South African cicadas (Homoptera: Cicadidae). S. Afr. J. Zool. 23(2):71-77.
Villet, M. 1992. Responses of free-living cicadas (Homoptera: Cicadidae) to broadcasts of cicada
songs. J. Ent. Soc. South Africa 55:93-97.
Walther, F. R. 1984. Communication and Expression in Hoofed Mammals. Bloomington: Indiana
University Press.
Young, A. M. 1976. Notes on the faunistic complexity of cicadas (Homoptera; Cicadidae) in northern Costa Rica. Rev. Biol. Trop. 24:267-279.
Young, D. 1990. Do cicadas radiate sound through their ear-drums? J. Exp. Biol. 151:41-56.
Young, D., and R. K. Josephson. 1983. Mechanisms of sound-production and muscle contraction kinetics in cicadas. J. Comp. Physiol. 152:183-195.
This content downloaded from 161.130.252.217 on Mon, 7 Jul 2014 22:40:43 PMAll use subject to JSTOR Terms and Conditions