Page 1
Indian Journal of Experimental Biology
Vol. 51, September 2013, pp. 739-745
Somatic embryo-like structures of strawberry regenerated in vitro on media
supplemented with 2,4-D and BAP
Genesia F Omar1, Fouad H Mohamed
1, Klaus-Thomas Haensch
2*, Sawsan H Sarg
1 & Mohamed M Morsey
1
1Department of Horticulture, Faculty of Agriculture, Suez Canal University, Ismailia, Egypt 2Department Plant Propagation, Leibniz-Institute of Vegetable and Ornamental Crops,
Kühnhäuser Straße 101, D-99090 Erfurt, Germany
Received 18 January 2013; revised 5 June 2013
Somatic embryo-like structures (SELS) were produced in vitro from leaf disk and petiole explants of two cultivars of
strawberry (Fragaria × ananassa Duch) on Murashige and Skoog medium with different concentrations and combinations
of 2,4-dichlorophenoxyacetic acid (2,4-D), 6-benzylaminopurine (BAP) and sucrose to check the embryonic nature of these
structures histologically. A large number of SELS could be regenerated in both cultivars on media with 2 - 4 mg L-1 2,4-D in
combination with 0.5 - 1 mg L-1 BAP and 50 g L-1 sucrose. Histological examination of SELS revealed the absence of a root
pole. Therefore these structures cannot be strictly classified as somatic embryos. The SELS formed under the tested culture
conditions represent malformed shoot-like and leaf-like structures. The importance of these results for the propagation of
strawberries via somatic embryogenesis is discussed.
Keywords: Auxin, Cytokinin, Fragaria × ananassa, Somatic embryogenesis
Strawberries (Fragaria × ananassa Duch) are one of
the most important fruits. In 2010 the worldwide
production of strawberries has been reported to be
around 4.3 million tonnes1. Such a huge production
is dependent upon efficient nursery production of
certified transplants. This in turn requires enough
in vitro disease-indexed plants from nuclear stocks
for further propagation in the nursery fields.
Micropropagation is a tool for the multiplication
of disease-free strawberry plants2,3
. Potentially
micropropagation of strawberry plants can be
carried out via somatic embryogenesis4-12
. Somatic
embryogenesis is a better alternative because of its
high propagation rates, the presence of both root
and shoot meristems in the same propagation unit and
the easy manageability for scale up as well as
other manipulations since embryos can be grown
freely floating in liquid cultures13
.
Plant growth regulators play a key role in the
induction of somatic embryogenesis14
. Raemakers
et al.15
estimated that in dicotyledonous species
among the different growth regulators applied for
somatic embryo induction, 2,4-dichlorophenoxyacetic
acid (2,4-D) has been the most commonly applied
auxin (49%) and 6-benzylaminopurine (BAP) the
most frequently used cytokinin (57%).
Auxins and cytokinins have also been used
to induce somatic embryogenesis in strawberry.
The auxins 2,4-D4,6,9-12
, 4-amino-3,5,6-trichloro-2-
pyridinecarboxylic acid (picloram)9, indole-3-butyric
acid (IBA)5,9
and α–naphthalene acetic acid
(NAA)4,6,9-11
alone or in combination with the
cytokinins BAP4-6,10,11
or thidiazuron (TDZ)12
have
been reported to induce embryogenesis.
All of the previous research dealing with
strawberry somatic embryogenesis included only
limited histological examination of the obtained
structures which were classified as somatic embryos.
However somatic embryos are defined as bipolar
structures, which do not have a vascular connection
with the mother tissue at any time of their life16
and develop through characteristic embryological
stages17
. None of the above mentioned reports
dealing with somatic embryogenesis in strawberry
presented unambiguous histological proof of somatic
embryogenesis. Pallavi et al.12
presented squash
preparations of callus with structures designated as
pro-embryos but the applied method was unsuitable
to show the undisturbed anatomy of these
early formations. Kordestani and Karami9 published
——————
*Correspondent author
Telephone: +49 (0) 36201-785 224
Fax: +49 (0) 36201-785 250
E-mail: [email protected]
Page 2
INDIAN J EXP BIOL, SEPTEMBER 2013
740
histological sections of structures which they
classified as globular somatic embryos but this
classification is not convincing because these
structures are broadly connected to the explant and
no information was given concerning the anatomy of
the more developed structures. The only histological
examination of a putative advanced stage of a somatic
embryo in strawberry is presented by Biswas et al.6
which shows anatomy of the structures classified as
early torpedo shaped embryos. Also this examination
represents no proof of somatic embryogenesis in
strawberry because the longitudinal section shows no
cotyledons and a root meristem consisting of small
cells with a dense cytoplasm and large nuclei.
The aim of the present study has been therefore to
check the embryonic nature of the somatic embryo-
like structures (SELS) that are regenerated in in vitro
cultures of strawberry on media supplemented with
different combinations of 2,4-D and BAP.
Materials and Methods Regeneration in vitro—Young leaves and petioles
from two strawberry cultivars namely Sweet Charlie
and Chandler (Fragaria × ananassa Duch), were
obtained from greenhouse-grown plants during
November. These were rinsed several times with
normal tap-water, surface sterilized in sodium
hypochlorite (14% active chlorine; VWR
International GmbH Deutschland) for 10 min,
followed by a treatment with 70% ethanol (VWR
International GmbH Deutschland) for 1 min and
rinsed three times in sterilized distilled water.
Leaf disks (5 × 5 mm) and petiole segments
(10 mm) were cultured on the surface of media
solidified with agar under sterile conditions in a
50 mL glass jar containing 10 mL medium. The basal
medium was MS medium18
with additional 500 mg L-1
casein hydrolysate. This medium was supplemented
with different concentrations and combinations of
2,4-D, BAP and sucrose. For each cultivar all possible
combinations of five concentrations (1.0, 2.0, 3.0,
4.0 and 5.0 mg L-1
) of 2,4-D, three concentrations of
BAP (0.5, 1.0 and 2.0 mg L-1
) and two levels of
sucrose (30 and 50 g L-1
) were prepared. Media were
solidified with 7.0 g L-1
agar and the pH was adjusted
to 5.7 before autoclaving (121 °C, 15 min).
Cultures were incubated at 25 °C in dark for two
weeks and then transferred under light (16/8 h
photoperiod at 69 µmol m-2
s-1
white fluorescent light)
at 18 °C for three weeks. After this time cultures were
transferred to MS medium without any plant growth
regulators with 30 g L-1
sucrose and grown for three
weeks (same light conditions as above and 25 °C
temperature). Three replicates were used per
treatment and each replicate comprised 10 glass jars.
Callus formation and regenerating SELS were
observed after the total time of eight weeks using
a stereo microscope.
Histological examination—Histological examination
was carried out following Haensch19
. Altogether eight
representative explants with regenerating SELS were
examined. The material was fixed using FAA, a
solution of 5.4 mL formalin (37%), 65.6 mL ethanol
(96%), 5 mL glacial acetic acid and 24 mL distilled
water20
. Parts with representative structures
were embedded in hydroxyethylmethacrylate (Histo-
technique-set Technovit 7100; Kulzer, Wehrheim,
Germany). The specimens were dehydrated in steps of
2 h through a graded series of ethanol (70, 90, 96 and
100%), then pre-infiltrated overnight with a mixture
of equal parts of 100% ethanol and Technovit
7100 base liquid and subsequently transferred into an
infiltration solution of 100 mL Technovit 7100 base
liquid and 1 g hardener I for 1 day. For proper
infiltration a vacuum was applied for 30 min at the
start of the last two processes. Explants were put in
Teflon molds with a mixture consisting of 15 parts
infiltration solution and one part hardener II and
polymerized for 1 h at room temperature and
afterwards for 6 h at 37 °C. The explants were then
mounted on block holders using Technovit 3040.
Afterwards they were cut into 6 µm slices at room
temperature. For this purpose a Jung CM1800
microtome with type 818 disposable microtome
blades was used (both from Leica Instruments,
Nussloch, Germany). Sections were stretched on the
surface of distilled water, then mounted on slides
and stained with 0.05% toluidine blue O (Serva,
Heidelberg, Germany) which was dissolved in
1% sodium tetraborate decahydrate buffer following
Hutchinson et al21
. The slices were then rinsed with
distilled water, dried and covered with Entellan
(Merck, Darmstadt, Germany) and a cover slip. This
technique stains the cytoplasm and unlignified cell
walls red and the lignified cell walls and DNA-
containing structures blue20
. Microscopic analysis was
performed using an Axio Imager A1 microscope
(Zeiss, Jena, Germany).
Statistical analysis—Quantitative data were analyzed
using analysis of variance (software STATISTICA 8,
Page 3
OMAR et al.: SOMATIC EMBRYO LIKE STRUCTURES IN STRAWBERRY
741
StatSoft 2009), and the means were compared by
Least Significant Difference Test (LSD) at 5% level
of probability. Ordinal data were analyzed in the
same way following Thöni22
.
Results Regeneration in vitro—The culture of the leaf and
petiole segments of both the cultivars in all the
treatments resulted in formation and proliferation of
callus (Fig. 1 A-F). However, the callus was creamy,
yellowish or brownish and granular depending
on the hormone combination. The degree of callus
formation varied between slight and massive
(Table 1). There were many treatments reaching the
highest possible rating for callus production without
showing significant differences to each other. A
center within this experimental scheme with many of
such very suitable treatments was observed in both
explant types of cv. Sweet Charlie in the range of 3 –
4 mg L-1
2,4-D in combination with 0.5 – 1 mg L-1
BAP and 50 g L-1
sucrose. In cv. Chandler most
treatments with this highest possible rating were
found in leaf disk cultures in a range of treatments
with 1 - 2 mg L-1
2,4-D and 2 mg L-1
BAP or with 2 -
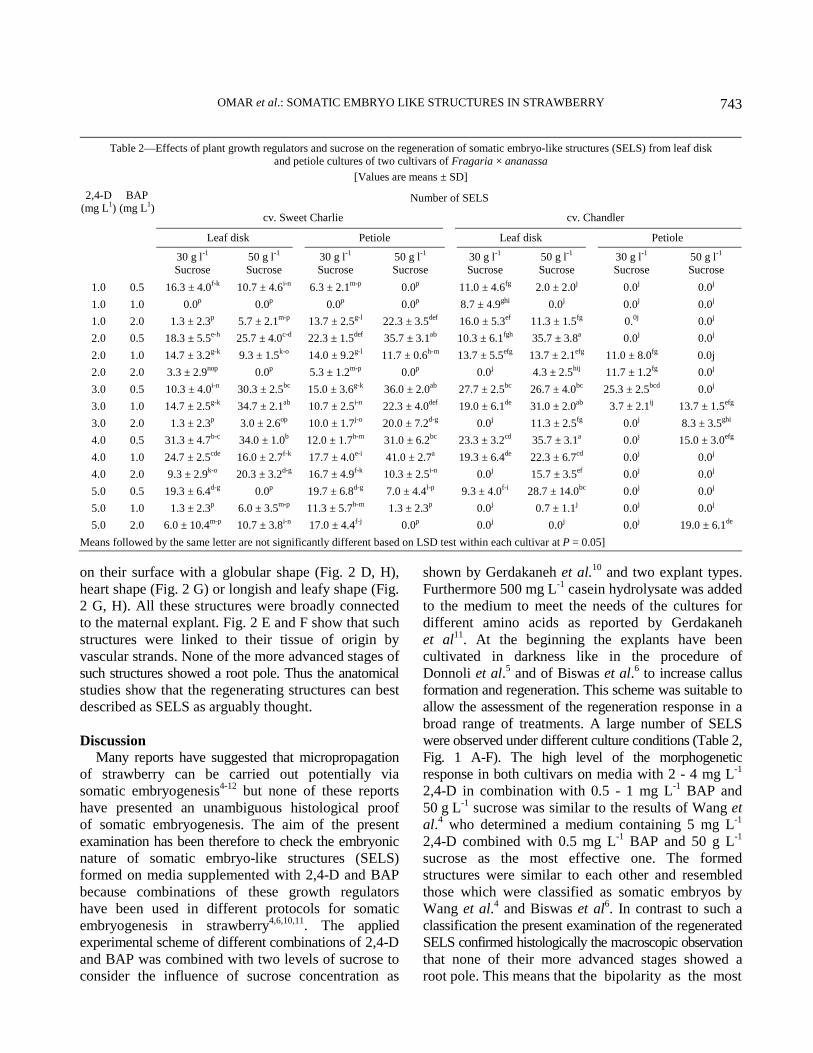
Fig. 1—Representative examples of callus formation and regeneration of somatic embryo-like structures (SELS) showing a high
variability in shape. Bar = 1 mm (A-F)
Page 4
INDIAN J EXP BIOL, SEPTEMBER 2013
742
3 mg L-1
2,4-D and 0.5 mg L-1
BAP each combined
with 50 g L-1
sucrose.
Regenerating structures were observed on both the
explant types of both the cultivars. These structures
resembled somatic embryos (Fig. 1 A-F). They
were globular- to leaf-shaped or similar to shoots,
yellowish or greenish and sometimes covered with
hairs. However, a root pole was never visible. A clear
classification of these structures was not possible
without histological examination. These structures
were designated as “somatic embryo-like structures”
(SELS). This regeneration response was obtained with
a broad spectrum of different combinations of the
applied growth regulators and sucrose but, in contrast
to callus formation, regeneration of SELS did not
occur in all the treatments (Table 2). The SELS
induced on different combinations were similar in
external characters. The best response in terms of
regeneration was observed with petioles of cv. Sweet
Charlie and leaf disks of cv. Chandler on media with
2 - 4 mg L-1
2,4-D in combination with 0.5 - 1 mg L-1
BAP and 50 g L-1
sucrose and with leaf disks of cv.
Sweet Charlie on a medium with 3 mg L-1
2,4-D in
combination with 1 mg L-1
BAP and 50 g L-1
sucrose.
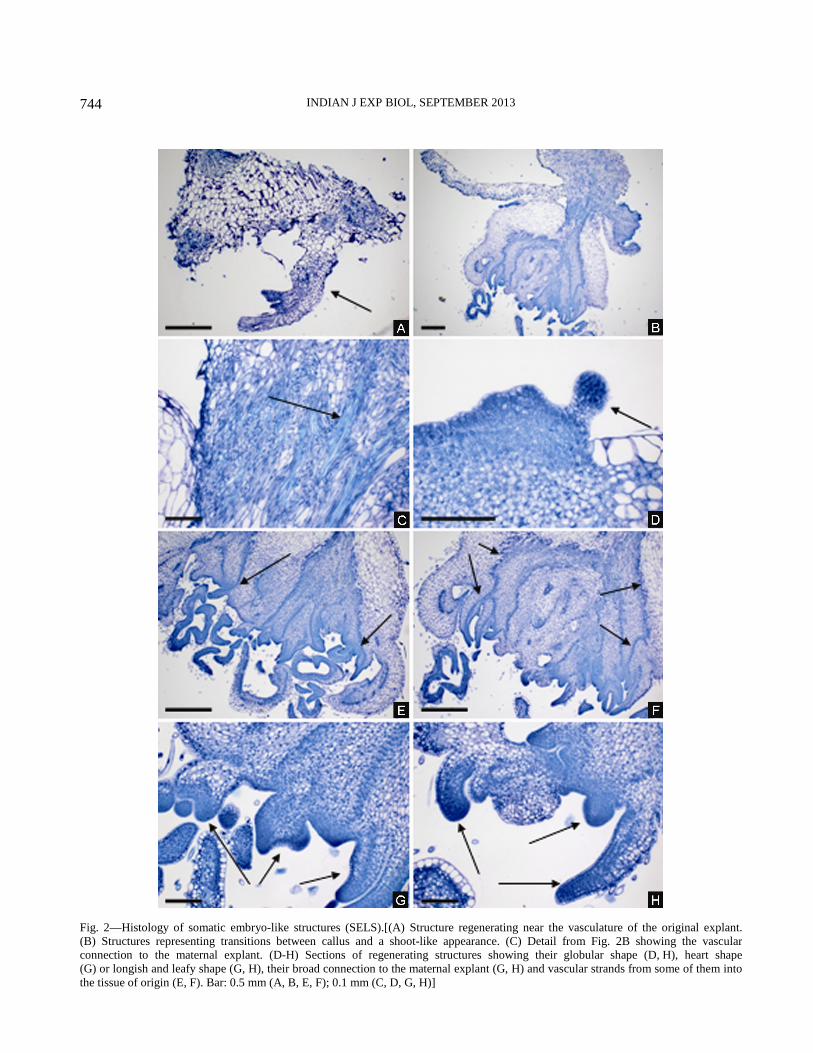
Histological examination—The light microscopic
examination of the histological sections revealed that
the regeneration of SELS occurred both when the
callus was slight as well as when callus formation was
much more extensive (Fig. 2 A, B). In case of slight
callus formation the regenerating structures originated
near the vasculature of the original explant (Fig. 2 A).
In case of extensive callus formation the origin of the
regenerating structures was difficult to determine
because some of the observed structures represent
transitions between callus and shoot like structures.
For example Fig. 2 B shows a remaining leaf disc
explant and on its right margin three structures. The
upper one is a callus clump showing irregular tissues.
The structure beneath looks on the whole like a very
malformed shoot, which is connected to the maternal
explant through vasculature (see this detail in Fig. 2 C)
and has meristem-like tissues and very callus-like
leafy substructures. The structure to the right hand
side with its cauliflower-like appearance and small
round formations on its surface represents a transition
between the two structures described above. Sections
of such malformed structures at different positions of
the explant show the regeneration of further structures
Table 1—Effects of plant growth regulators and sucrose on callus formation in leaf disk and petiole cultures of two cultivars of
Fragaria × ananassa
[Values are means]
Degree of callus formation
cv. Sweet Charlie cv. Chandler
Leaf disk Petiole Leaf disk Petiole
2,4-D
(mg L-1)
BAP
(mg L-1)
30 g l-1
Sucrose
50 g l-1
Sucrose
30 g l-1
Sucrose
50 g l-1
Sucrose
30 g l-1
Sucrose
50 g l-1
Sucrose
30 g l-1
Sucrose
50 g l-1
Sucrose
1.0 0.5 2.3ef 3.0cd 3.0cd 2.7de 3.0cd 1.3gh 1.0h 2.0efg
1.0 1.0 3.3bc 3.0cd 2.3ef 2.0f 2.7cde 3.0cd 2.0efg 2.0efg
1.0 2.0 3.0cd 2.7de 3.7ab 4.0a 3.3abc 4.0a 3.3abc 1.0h
2.0 0.5 3.0cd 3.0cd 3.0cd 3.7ab 3.0cd 4.0a 2.3def 3.0cd
2.0 1.0 3.0cd 2.0f 2.0f 3.3bc 3.3abc 3.0cd 3.0cd 3.3abc
2.0 2.0 3.0cd 2.0f 3.3bc 3.3bc 2.0efg 4.0a 2.3def 3.0cd
3.0 0.5 3.0cd 4.0a 3.0cd 4.0a 3.3abc 4.0a 3.0cd 2.3def
3.0 1.0 3.3bc 4.0a 2.0f 4.0a 3.7ab 3.0cd 1.0h 4.0a
3.0 2.0 2.0f 3.0cd 2.0f 3.7ab 4.0a 2.3def 2.0efg 3.0cd
4.0 0.5 4.0a 4.0a 3.7ab 4.0a 3.3abc 3.7ab 1.3gh 3.0cd
4.0 1.0 3.7ab 4.0a 4.0a 4.0a 3.0cd 3.0cd 2.0efg 3.0bcd
4.0 2.0 3.0cd 3.0cd 3.0cd 2.0f 3.3abc 3.0cd 2.0efg 3.0bcd
5.0 0.5 3.3bc 2.0f 4.0a 4.0a 2.0efg 4.0a 3.0bcd 1.0h
5.0 1.0 2.3ef 3.0cd 3.0cd 3.0cd 1.0h 4.0a 3.0bcd 1.0h
5.0 2.0 2.0f 3.0cd 2.0f 4.0a 1.0h 1.0h 2.0efg 1.7fgh
Means followed by the same letter are not significantly different within each cultivar based on LSD test at P=0.05. Degree of callus
formation: 1 = slight, 2 = minor moderate, 3 = moderate, 4 = massive]
Page 5
OMAR et al.: SOMATIC EMBRYO LIKE STRUCTURES IN STRAWBERRY
743
on their surface with a globular shape (Fig. 2 D, H),
heart shape (Fig. 2 G) or longish and leafy shape (Fig.
2 G, H). All these structures were broadly connected
to the maternal explant. Fig. 2 E and F show that such
structures were linked to their tissue of origin by
vascular strands. None of the more advanced stages of
such structures showed a root pole. Thus the anatomical
studies show that the regenerating structures can best
described as SELS as arguably thought.
Discussion
Many reports have suggested that micropropagation
of strawberry can be carried out potentially via
somatic embryogenesis4-12
but none of these reports
have presented an unambiguous histological proof
of somatic embryogenesis. The aim of the present
examination has been therefore to check the embryonic
nature of somatic embryo-like structures (SELS)
formed on media supplemented with 2,4-D and BAP
because combinations of these growth regulators
have been used in different protocols for somatic
embryogenesis in strawberry4,6,10,11
. The applied
experimental scheme of different combinations of 2,4-D
and BAP was combined with two levels of sucrose to
consider the influence of sucrose concentration as
shown by Gerdakaneh et al.10
and two explant types.
Furthermore 500 mg L-1
casein hydrolysate was added
to the medium to meet the needs of the cultures for
different amino acids as reported by Gerdakaneh
et al11
. At the beginning the explants have been
cultivated in darkness like in the procedure of
Donnoli et al.5 and of Biswas et al.
6 to increase callus
formation and regeneration. This scheme was suitable to
allow the assessment of the regeneration response in a
broad range of treatments. A large number of SELS
were observed under different culture conditions (Table 2,
Fig. 1 A-F). The high level of the morphogenetic
response in both cultivars on media with 2 - 4 mg L-1
2,4-D in combination with 0.5 - 1 mg L-1
BAP and
50 g L-1
sucrose was similar to the results of Wang et
al.4 who determined a medium containing 5 mg L
-1
2,4-D combined with 0.5 mg L-1
BAP and 50 g L-1
sucrose as the most effective one. The formed
structures were similar to each other and resembled
those which were classified as somatic embryos by
Wang et al.4 and Biswas et al
6. In contrast to such a
classification the present examination of the regenerated
SELS confirmed histologically the macroscopic observation
that none of their more advanced stages showed a
root pole. This means that the bipolarity as the most
Table 2—Effects of plant growth regulators and sucrose on the regeneration of somatic embryo-like structures (SELS) from leaf disk
and petiole cultures of two cultivars of Fragaria × ananassa
[Values are means ± SD]
Number of SELS
cv. Sweet Charlie cv. Chandler
Leaf disk Petiole Leaf disk Petiole
2,4-D
(mg L1)
BAP
(mg L1)
30 g l-1
Sucrose
50 g l-1
Sucrose
30 g l-1
Sucrose
50 g l-1
Sucrose
30 g l-1
Sucrose
50 g l-1
Sucrose
30 g l-1
Sucrose
50 g l-1
Sucrose
1.0 0.5 16.3 ± 4.0f-k
10.7 ± 4.6i-n
6.3 ± 2.1m-p
0.0p 11.0 ± 4.6
fg 2.0 ± 2.0
j 0.0
j 0.0
j
1.0 1.0 0.0p 0.0
p 0.0
p 0.0
p 8.7 ± 4.9
ghi 0.0
j 0.0
j 0.0
j
1.0 2.0 1.3 ± 2.3p 5.7 ± 2.1
m-p 13.7 ± 2.5
g-l 22.3 ± 3.5
def 16.0 ± 5.3
ef 11.3 ± 1.5
fg 0.
0j 0.0
j
2.0 0.5 18.3 ± 5.5e-h
25.7 ± 4.0c-d
22.3 ± 1.5def
35.7 ± 3.1ab
10.3 ± 6.1fgh
35.7 ± 3.8a 0.0
j 0.0
j
2.0 1.0 14.7 ± 3.2g-k
9.3 ± 1.5k-o
14.0 ± 9.2g-l
11.7 ± 0.6h-m
13.7 ± 5.5efg
13.7 ± 2.1efg
11.0 ± 8.0fg
0.0j
2.0 2.0 3.3 ± 2.9nop
0.0p 5.3 ± 1.2
m-p 0.0
p 0.0
j 4.3 ± 2.5
hij 11.7 ± 1.2
fg 0.0
j
3.0 0.5 10.3 ± 4.0i-n
30.3 ± 2.5bc
15.0 ± 3.6g-k
36.0 ± 2.0ab
27.7 ± 2.5bc
26.7 ± 4.0bc
25.3 ± 2.5bcd
0.0j
3.0 1.0 14.7 ± 2.5g-k
34.7 ± 2.1ab
10.7 ± 2.5i-n
22.3 ± 4.0def
19.0 ± 6.1de
31.0 ± 2.0ab
3.7 ± 2.1ij 13.7 ± 1.5
efg
3.0 2.0 1.3 ± 2.3p 3.0 ± 2.6
op 10.0 ± 1.7
j-o 20.0 ± 7.2
d-g 0.0
j 11.3 ± 2.5
fg 0.0
j 8.3 ± 3.5
ghi
4.0 0.5 31.3 ± 4.7b-c
34.0 ± 1.0b 12.0 ± 1.7
h-m 31.0 ± 6.2
bc 23.3 ± 3.2
cd 35.7 ± 3.1
a 0.0
j 15.0 ± 3.0
efg
4.0 1.0 24.7 ± 2.5cde
16.0 ± 2.7f-k
17.7 ± 4.0e-i
41.0 ± 2.7a 19.3 ± 6.4
de 22.3 ± 6.7
cd 0.0
j 0.0
j
4.0 2.0 9.3 ± 2.9k-o
20.3 ± 3.2d-g
16.7 ± 4.9f-k
10.3 ± 2.5i-n
0.0j 15.7 ± 3.5
ef 0.0
j 0.0
j
5.0 0.5 19.3 ± 6.4d-g
0.0p 19.7 ± 6.8
d-g 7.0 ± 4.4
l-p 9.3 ± 4.0
f-i 28.7 ± 14.0
bc 0.0
j 0.0
j
5.0 1.0 1.3 ± 2.3p 6.0 ± 3.5
m-p 11.3 ± 5.7
h-m 1.3 ± 2.3
p 0.0
j 0.7 ± 1.1
j 0.0
j 0.0
j
5.0 2.0 6.0 ± 10.4m-p
10.7 ± 3.8i-n
17.0 ± 4.4f-j
0.0p 0.0
j 0.0
j 0.0
j 19.0 ± 6.1
de
Means followed by the same letter are not significantly different based on LSD test within each cultivar at P = 0.05]
Page 6
INDIAN J EXP BIOL, SEPTEMBER 2013
744
Fig. 2—Histology of somatic embryo-like structures (SELS).[(A) Structure regenerating near the vasculature of the original explant.
(B) Structures representing transitions between callus and a shoot-like appearance. (C) Detail from Fig. 2B showing the vascular
connection to the maternal explant. (D-H) Sections of regenerating structures showing their globular shape (D, H), heart shape
(G) or longish and leafy shape (G, H), their broad connection to the maternal explant (G, H) and vascular strands from some of them into
the tissue of origin (E, F). Bar: 0.5 mm (A, B, E, F); 0.1 mm (C, D, G, H)]
Page 7
OMAR et al.: SOMATIC EMBRYO LIKE STRUCTURES IN STRAWBERRY
745
important characteristic of an embryo could not be
proved and that therefore these SELS cannot be
classified as somatic embryos. These SELS have to be
regarded partially as malformed shoots because the
histology revealed a shoot-like structure (Fig. 2 A, B,
E, F) with meristematic areas (Fig. 2 G, H) and
partially very thick callus-like leaves (Fig. 2 B, E, F).
These shoot-like structures originate near the
vasculature of the original explant (Fig. 2 A) or
develop their own vasculature connection with the
tissue from which they originate (Fig. 2 C). Other
SELS have to be considered as malformed leaf-like
structures having a broad connection to the tissue of
origin but showing only restricted vascular strands
(Fig. 2 H). All these structures resemble somatic
embryos only externally. They are the result of the
two overlapping processes of callus formation and
regeneration as it can be seen from Fig. 2 B. This is
the reason for the malformations and the great
variability of structure shapes which can be observed.
Considering the results from this study as
well as those of previously published examinations
concerning somatic embryogenesis in strawberry
leads to the conclusion that until now there is no proof
for the regeneration of somatic embryos from media
which have been supplemented with combinations of
2,4-D and BAP in this species.
Acknowledgment This investigation was undertaken with the support
of the Channel System of the Arab Republic of Egypt,
the State of Brandenburg, the Free State of Thuringia
and the Federal Republic of Germany. Thanks are due
to Mrs. Barbara Weinlich for technical assistance.
References 1 FAOSTAT 2012, Worldwide Strawberry production 2010,
State on 22nd November 2012, http://faostat.fao.org/site/567/
DesktopDefault.aspx?PageID=567#ancor.
2 Boxus P, The production of strawberry plants by in vitro
micropropagation, J Hortic Sci Biotechnol, 49 (1974) 209.
3 Swartz H J, Galletta G J & Zimmermann R H, Field
performance and phenotypic stability of tissue culture-
propagated strawberries, J Am Soc Hortic Sci, 106 (1981) 667.
4 Wang D, Wergin W P & Zimmerman R Z, Somatic
embryogenesis and plant regeneration from immature
embryos of strawberry, Hort Science, 19 (1984) 71.
5 Donnoli R, Sunseri F, Martelli G & Greco I, Somatic
embryogenesis, plant regeneration and genetic
transformation in Fragaria spp. Acta Hortic, 560 (2001) 235.
6 Biswas M K, Islam R & Hossain M, Somatic embryogenesis
in strawberry (Fragaria sp.) through callus culture, Plant
Cell Tissue Organ Cult, 90 (2007) 49.
7 Husaini A M & Abdin M Z, Interactive effect of light,
temperature and TDZ on the regeneration potential of leaf
discs of Fragaria × ananassa Duch. In Vitro Cell Dev Biol:
Plant, 43 (2007) 576.
8 Husaini A M, Aquil S, Bhat M, Qadri T, Kamaluddin &
Abdin M Z, A high-efficiency direct somatic embryogenesis
system for strawberry (Fragaria × ananassa Duch.) cultivar
Chandler, J Crop Sci Biotechnol, 11 (2008) 107.
9 Kordestani G K & Karami O, Picloram-induced somatic
embryogenesis in leaves of strawberry (Fragaria ananassa
L.), Acta Biol Crac Ser Bot, 50 (2008) 69.
10 Gerdakaneh M, Mozafari A A, Khalighi A & Sioseh-mardah
A, The effects of carbohydrate source and concentration on
somatic embryogenesis of strawberry (Fragaria × ananassa
Duch.), Am-Eurasian J Agric Environ Sci, 6 (2009) 76.
11 Gerdakaneh M, Mozafari A A, Sioseh-mardah A & Sarabi B,
Effects of different amino acids on somatic embryogenesis of
strawberry (Fragaria × ananassa Duch.). Acta Physiol Plant,
33 (2011) 1847.
12 Pallavi C M; Rekha R & Neelambika T M, Indirect somatic
embryogenesis from petiole segment in strawberry cv. Sweet
Charlie, Indian J. Hortic, 68 (2011) 24.
13 Ammirato P V, Organizational events during somatic
embryogenesis, in Plant Tissue and Cell Culture, edited by
C E Green, D A Sommers, W P Hacket, & D D Biersboer
(Alan R Liss Inc, New York) 1987, 57.
14 Jimenez V M & Bangerth F, Endogenous hormone levels in
explants and in embryogenic and non-embryogenic cultures
of carrot, Physiol Plant, 111 (2001) 389.
15 Raemakers C J J M, Jacobsen E & Visser R G F, Secondary
somatic embryogenesis and applications in plant breeding,
Euphytica, 81 (1995) 93.
16 Haccius B, Question of unicellular origin of non-zygotic
embryos in callus cultures, Phytomorphology, 28 (1978) 74.
17 Williams EG & Maheswaran G, Somatic embryogenesis:
factors influencing coordinated behavior of cells as an
embryogenic group, Ann Bot, 57 (1986) 443.
18 Murashige T & Skoog F, A revised medium for rapid growth
and bio assays with tobacco tissue cultures, Physiol Plant,
15 (1962) 473.
19 Haensch K T, Morpho-histological study of somatic embryo-
like structures in hypocotyl cultures of Pelargonium x
hortorum Bailey, Plant Cell Rep, 22 (2004) 376.
20 Gerlach D, Botanische mikrotechnik (Georg Thieme,
Stuttgart New York) 1984, 32.
21 Hutchinson M J, KrishnaRaj S & Saxena P K, Morphological
and physiological changes during thidiazuron-induced
somatic embryogenesis in geranium (Pelargonium ×
hortorum Bailey) hypocotyl cultures, Int J Plant Sci, 157
(1996) 440.
22 Thöni H, Auswertung von Bonituren: ein empirischer
Methodenvergleich, EDV in Medizin und Biologie, 16
(1985) 108.