SOME OBSERVATIONS ON THE EFFECT OF COLCHICINE UPON SALMONELLA TYPHOSA' EDWARD J. MINSAVAGE AND EDWARD D. DELAMATER Section on Cytology and Genetics, Department of Physiology, School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania Received for publication February 25, 1955 Colchiciiie, a highly toxic plant alkaloid, is used by geneticists for the production of poly- ploidy in plants. While mitosis and cell division are suppressed, the multiplication of the chromo- somes may not be interrupted. Ostergren (1951) stated that the spindle is dis- organized by colchicine, and that the chromo- somes do not form a metaphase plate but are distributed at random over the cell or clustered in one or more irregular groups. The chromo- somes divide, but the daughter chromosomes remain lying side by side instead of separating to the two opposite poles of the spindle. They may form a single nucleus having double the number of chromosomes, but also they may form two or more aneuploid nuclei, depending on the random position of the chromosomes. The chromosomes may be excessively contracted while the cell is under the influence of the drug. The purpose of this paper is to describe the diverse nuclear and cellular effects produced by colchicine on Salmonella typhosa under controlled conditions. MATERIALS AND METHODS The culture (H901) of S. typhosa used is a non-lysogenic strain for which the mitotic cycle has been presented in AIinsavage and DeLamater (1955). The media employed were nutrient broth and agar (Difco), and MAorton and Engley (1945) trvptone-yeast extract-glucose broth (MI & E broth). The bacteria were kept in continuous culture by transferring 0.1 ml of a 24-hr culture in nutrient broth to a fresh tube of the same medium each day. Cultures on nutrient agar were also 1 This investigation was supported in part by, research grants (PHS # C-2189) from the Na- tional Institutes of Health, Public Health Service, and the Atomic Energy Commission, Contract § AT (30-1) -1341. maintained in a refrigerator; these served as the source of inoculum in some of the experiments. The bacteria were prepared for experimenta- tion as follows: 0.1 ml of a 24-hr culture of the bacteria wa3 introduced into a series of Was- sermann tubes containing 5 ml of M & E broth. The tubes were then incubated at 37 C for 3.5 hr. The cells in each tube were deposited by centrifugation for 10 minutes and the super- natant fluid was poured off; the cells were re- suspended immediately in 1 ml of M & E broth containing the proper concentrations of colchi- cine, or in 1 ml of M & E broth to serve as con- trols. The control cells in these experiments were used in the study of the mitotic cycle (1\Iinsavage and DeLamater, 1955). In experiments in which multiple successive doses of colchicine were employed, the cells were spun down at the end of each time interval of treatment; the supernatant fluid was poured off; the cells were then quickly resuspended in the next concentration of colchicine in M &E broth, or into fresh M & E broth to serve as controls. The volume of medium employed in the multiple successive dose experiments was 2 ml. In certain experiments a refrigerated culture on a nutrient agar slant served as the source of the inoculum in place of the continuous culture. The bacteria were suspended in 5 ml of nutrient broth, and 0.1 ml of this suspension was then introduced into the tubes containing M & E broth. Colchicine (Nutritional Biochemicals Corp., Cleveland, Ohio) was dissolved in M & E broth at a 1 per cent concentration, and was sterilized by sintered glass filtration. This stock solution was stored at 4 C, and was diluted in M & E broth to the required concentrations just prior to use. The solutions were shielded from light, to prevent photochemical degradation, and dis- carded after two weeks. The anaerobic method used was that of Morton 501 on March 27, 2020 by guest http://jb.asm.org/ Downloaded from
Transcript
SOME OBSERVATIONS ON THE EFFECT OF COLCHICINEUPON SALMONELLA TYPHOSA'
EDWARD J. MINSAVAGE AND EDWARD D. DELAMATER
Section on Cytology and Genetics, Department of Physiology, School of Medicine,University of Pennsylvania, Philadelphia, Pennsylvania
Received for publication February 25, 1955
Colchiciiie, a highly toxic plant alkaloid, isused by geneticists for the production of poly-ploidy in plants. While mitosis and cell divisionare suppressed, the multiplication of the chromo-somes may not be interrupted.
Ostergren (1951) stated that the spindle is dis-organized by colchicine, and that the chromo-somes do not form a metaphase plate but aredistributed at random over the cell or clusteredin one or more irregular groups. The chromo-somes divide, but the daughter chromosomesremain lying side by side instead of separating tothe two opposite poles of the spindle. They mayform a single nucleus having double the numberof chromosomes, but also they may form two ormore aneuploid nuclei, depending on the randomposition of the chromosomes. The chromosomesmay be excessively contracted while the cell isunder the influence of the drug.The purpose of this paper is to describe the
diverse nuclear and cellular effects produced bycolchicine on Salmonella typhosa under controlledconditions.
MATERIALS AND METHODS
The culture (H901) of S. typhosa used is anon-lysogenic strain for which the mitotic cyclehas been presented in AIinsavage and DeLamater(1955).The media employed were nutrient broth and
agar (Difco), and MAorton and Engley (1945)trvptone-yeast extract-glucose broth (MI & Ebroth).The bacteria were kept in continuous culture
by transferring 0.1 ml of a 24-hr culture innutrient broth to a fresh tube of the same mediumeach day. Cultures on nutrient agar were also
1 This investigation was supported in part by,research grants (PHS # C-2189) from the Na-tional Institutes of Health, Public Health Service,and the Atomic Energy Commission, Contract §AT (30-1) -1341.
maintained in a refrigerator; these served as thesource of inoculum in some of the experiments.The bacteria were prepared for experimenta-
tion as follows: 0.1 ml of a 24-hr culture of thebacteria wa3 introduced into a series of Was-sermann tubes containing 5 ml of M & E broth.The tubes were then incubated at 37 C for 3.5hr. The cells in each tube were deposited bycentrifugation for 10 minutes and the super-natant fluid was poured off; the cells were re-suspended immediately in 1 ml of M & E brothcontaining the proper concentrations of colchi-cine, or in 1 ml of M & E broth to serve as con-trols. The control cells in these experiments wereused in the study of the mitotic cycle (1\Iinsavageand DeLamater, 1955).
In experiments in which multiple successivedoses of colchicine were employed, the cells werespun down at the end of each time interval oftreatment; the supernatant fluid was poured off;the cells were then quickly resuspended in thenext concentration of colchicine in M & E broth,or into fresh M & E broth to serve as controls.The volume of medium employed in the multiplesuccessive dose experiments was 2 ml.
In certain experiments a refrigerated cultureon a nutrient agar slant served as the source ofthe inoculum in place of the continuous culture.The bacteria were suspended in 5 ml of nutrientbroth, and 0.1 ml of this suspension was thenintroduced into the tubes containing M & Ebroth.
Colchicine (Nutritional Biochemicals Corp.,Cleveland, Ohio) was dissolved in M & E brothat a 1 per cent concentration, and was sterilizedby sintered glass filtration. This stock solutionwas stored at 4 C, and was diluted in M & Ebroth to the required concentrations just priorto use. The solutions were shielded from light,to prevent photochemical degradation, and dis-carded after two weeks.The anaerobic method used was that of Morton
(1943). The air in the tubes was replaced, afterevacuation, by a nitrogen-carbon dioxide mixturewith a small volume of hydrogen to combinewith any remaining traces of oxygen.Maximum unforced aeration was obtained in
certain experiments by layering 2 ml of inoculatedcolchicine solution on the bottom of a 50-mlErlenmeyer flask.To prepare the slides a drop of a broth culture
was placed on a partially dried block of agar toremove the excess moisture. The cells on theagar block were fixed in the vapors of a 2 percent aqueous solution of osmium tetroxide for 1minute. An impression smear was then made ona coverslip. The cells on the coverslip werehydrolyzed in 1 N HCl at 60 C for a period of 7to 16 min. After rinsing in distilled water, thecoverslip was immersed in 10 ml of a 0.25 percent aqueous solution of azure A (Cat. No. 136,Harleco) to which 2 drops of thionyl chloride hadbeen added about 15 minutes prior to use. Thesmears were immersed in the nuclear dye for 18hours. The coverslip was then rinsed in distilledwater and placed in fresh 10 per cent tannic acidfor 30 min, rinsed, and dipped briefly into a 0.1per cent aqueous solution of basic fuchsin, ac-cording to Hunter's ( DeLamater, 1953) modi-fication of Cassel's (1951) cell wall stain. Thecoverslip was then thoroughly rinsed in distilledwater, blotted, allowed to dry in air until thefilm of water on the glass had just evaporated, andthen mounted according to the method of Min-savage (1955). The mounting medium consists of75 per cent heavy mineral oil and 25 per centa-bromonaphthalene. The edges of the coverslipwere sealed with a colorless cement, such asDuPont "Duco" cement.The endpoint for optimal hydrolysis was de-
termined by making a series of slides of the samepopulation of cells with varying lengths of timeof exposure to the acid. The slide showing thegreatest clarity of nuclear configurations wasconsidered optimally hydrolyzed.The hydrogen ion concentrations of the media
containing colchicine were determined withHydrion pH paper; colchicine had no detectableeffect upon pH of the media.Growth curves were determined both tur-
bidimetrically and by plating. One-tenth ml of a24-hr culture of S. typhosa in nutrient broth wassubcultured in 9.9 ml M & E broth. After incuba-tion at 37 C for 3.5 hr, 1 ml of this culture was
serially diluted in normal saline and plated innutrient agar. One ml was also introduced intoKlett tubes containing 9 ml M & E broth (con-trol) or the proper concentration of colchicine in9 ml M & E broth. Klett readings were taken athourly intervals, at which time 1 ml of mediumfrom each tube was serially diluted in normalsaline and plated in nutrient agar.The slides were examined with a Bausch and
Lomb research microscope employing a 12.5 Xeyepiece and a 97X achromatic oil immersionobjective of NA 1.25. Upon occasion a 61 X apo-chromatic oil immersion objective of NA 1.40was used in conjunction with 20X and 28Xeyepieces. Photomicrographs were taken with aBausch and Lomb L type camera so as to givea 2X further magnification on 5 x 7 sheets ofKodak Panatomic X or Kodagraph microfilefilm. The latter is especially good for photo-graphing cells with a low density. The negativeswere developed in Kodak microdol developer at20 C for 17 minutes. The prints were furtherenlarged 2X, giving a final magnification of4850X. The light source employed was a 100-watt zirconium arc lamp used with Bausch andLomb interference filters (570 and 590 mu). Thislight source, for practical purposes, renders anessentially monochromatic system. Exposuretimes were determined with a Photovolt exposurephotometer and by test.
OBSERVATIONS
Optimal hydrolysis times. The purpose of acidhydrolysis is either to remove or to changeribonucleic acid; thus it no longer stains with thedye used to demonstrate the nuclei. Aldehydegroups in desoxyribonucleic acid are also liber-ated.The optimal hydrolysis time for normal bac-
teria used in these experiments has usually been7 to 8 minutes in 1 N HCl at 60 C. One effect ofcolchicine is to increase the optimal hydrolysistime in S. typhosa. In some instances the hy-drolysis time was doubled.
Growth curves. In the normal growth curve ofS. typhosa the logarithmic growth phase beginsabout 1.5 hr after inoculation and persists untilabout the sixth hour. The bacteria used in mostof these experiments were grown for 3.5 hr beforethey were exposed to colchicine. At this timethere was an average of 136 million cells per ml,and the population was in the lower third of the
logarithmic growth phase. Thus, 2.5 hr of thelogarithmic period remained for experimentalobservation.When turbidity readings and plate counts were
used as methods of measurement, it was foundthat colchicine in concentrations of 1 and 7.5mg/ml had no effect upon the growth rate ofS. typhosa. Cytologic analysis, however, indicatedthat approximately 10 per cent of the cells wereaffected by the drug.The bacteria exposed to 1 and 7.5 mg/ml
colchicine in MI & E broth for 3 hr showedcolonies similar to the controls when plated onnutrient agar. These cultures were unable toutilize sucrose and lactose, which were incorpor-ated in Difco phenol-red nutrient agar base.
Cytology. Cells 3jg hr old were exposed to 1mg/ml colchicine in M & E broth for 1 hr an-aerobically, according to the method of Morton(1943). The optimal hydrolysis time for thecontiol cells was 7 to 9 min, and 7 to 10 minfor the cells treated with colchicine in this con-centration. No difference in the cytology of thesetwo groups of bacteria was detected, and in eachcase the cells were small.
Cells 3ki hr old were exposed to 1 mg/mlcolchicine in M & E broth which was layered in2-ml quantities on the bottoms of 50-ml Erlen-meyer flasks so as to secure maximum unforcedaeration. Cells exposed for 1 hr had an optimalhydrolysis time of 11 to 13 min as compared to7 to 8 min for their control. Cells exposed for 2hr had an optimal hydrolysis time of 14 min ascompared to 7 to 8 min for their controls. Thecolchicine-treated cells were longer than thecontrols, but no observable difference in nuclearconfigurations was detected in the aerobic ex-periments.
TABLE 1Nuclear changes, due to single doses of colchicinefor various periods of time, expressed in per cent
found in 1000 or more cells per slide
La rge Axial LargeTreatment-Continuous Culture Round "Poly- is-
TABLE 2Nuclear changes, due to multiple successive ex-
posures to high and low doses of colchicine for1-hour periods in cells derived from refrigeratedand continuous culture seed inocula, expressed inper cent found in 1000 or more cells per slide
Large Axial LargeRound "Poly- Dis-
Treatment Dense ploid" BacdeNucleus Nuclei teBac
Continuous culture seedinoculum:
Control ................... 1.3 0.3 0
1.5 mg/ml/l hr............ 1.0 0.3 0To:
2 mg/ml/i hr.............. 2.2 1.8 0.2To:
2.5 mg/ml/i hr............ 1.1 3.6 0
7.5 mg/ml/i hr............ 2.3 0.3 0To:
8 mg/ml/i hr.............. 1.0 2.3 0To:
8.5 mg/ml/i hr............ 2.7 2.8 0
Refrigerated seed inoculum:Control... 1.4 0.08 0
1.5 mg/ml/i hr ............ 0.8 0.2 0To:
2 mg/ml/i hr.. 1 .5 68.1 0.2To:
2.5 mg/ml/i hr............ 1 .9 0.2 0.2
7.5 mg/ml/i hr............ 7.9 0.7 0To:
8 mg/ml/i hr.............. 7.9 0.4 0To:
8.5 mg/ml/I hr............ 6.5 3.6 0
The nuclear changes induced by colchicine intypical experiments in Wassermann tubes areshown in tables 1 and 2. Two types of nuclearconfigurations and a cellular change are tabulatedwith the percentage of incidence. An incidence ofless than 0.1 per cent was scored as 0. Large denseround nuclei and what are interpreted as "tran-sient polyploid nuclei" (DeLamater, 1953 and1954; DeLamater et al., 1955) were also found inthe control preparations in small numbers. Inthe control preparations the bacteria containingthese axial nuclei were usually of the same di-mensions as the neighboring cells. In the bacteria
£Figures 1-6. All photographs are of Salmonella typhosa at a magnification of 4850 X. All cells were
treated with colchicine for designated time. 1: Three lower nuclei condensed. Upper nucleus extended.1 mg/ml for 1 hr. 2: Two nuclei condensed. Others extended. Cell division inhibited. 1 mg/ml for 1 hr.3: Axial nucleus segmenting. 1 mg/ml for 1 hr. 4: Axial nucleus segmenting into individual nuclei whichare undergoing mitosis. 7.5 mg/ml for 1 hr. 5: Individual nuclei undergoing division while axial nucleuscontinues to segment. Telophase seen at bottom of cell. Anaphase at top. 1 mg/ml for 75 min. 6: Nucleiappear to have returned to normal haploid condition. 1 mg/ml for 1 hr.
exposed to colchicine the cells containing the axialnuclei were usually greatly increased in size ascompared to the presumably unaffected cellswhich surrounded them. Data in the tablesemphasize the variability in the response of S.typhosa to colchicine. Repetition of the experi-ments, however, demonstrated that the resultsfor each dose of the drug fell within a range ofvalues.
Figures 1 to 6 represent a reconstructed se-quence of steps in the production of long axialnuclei ("transient polyploid nuclei") and whatappears to be their subsequent reorganization tonormal nuclei. Cells of similar size have beenobserved in unfixed, unstained, wet-mountpreparations in which they appeared as activelymotile bacteria during the period of time coveredby the experiments. Figure 1 shows an elongatedcell containing three dense nuclei and a fourthwhich appears to be elongating. As growth con-tinues in the medium containing colchicine, thecells which are thought to have been arrested inmitosis continue to elongate, and the chromo-somal mass in the center elongates with it, asseen in figures 2 and 3. These cells were exposedto 1 mg/ml colchicine. The continuing reduplica-tion of the chromosomes within these axialchromatinic masses is demonstrated as recoveryfrom the drug proceeds. The nuclei are fre-quently too dense to permit clear observation ofthis internal structure. Recovery stages, observedeven in the presence of the drug, are seen infigures 4 to 6. The recovering nuclei ultimatelybecome separated, each of which appears to con-tain three chromosomes. This reorganizationbecomes evident between the first and the thirdhour of exposure to colchicine. The reorganizationof the "transient polyploid" nuclei in S. typhosaappears to be similar to that seen in Bacillusmegaterium exposed to oxytetracycline (De-Lamater, 1953).What appears to be inhibition of cellular
division is seen in figures 7 and 8. The develop-ment of aberrant branches resulting in "Y"-shaped configurations is shown in figure 7.Similar cells have been shown by Zapf (1953),Bergersen (1953), Stempen and Hutchinson(1951), Duguid (1948), Mellon (1925), andGardner (1925) in other bacteria under otherconditions. Fusiform cells and "large bodies"were also observed. A "large body" is shown infigure 8. These three configurations were occa-
sionally and inconsistently observed in controlpreparations in smaller numbers than in colchi-cine-treated cultures.
Figures 9 to 11 depict stages suggesting thosereported by Mellon (1925) as representing asexual process in Escherichia coli and S. typhosa.In figure 9 two dense nuclei are shown in a largebud. In figure 10 a bud containing a dense massof nuclear material is seen between two cells,while in figure 11 is seen a detached bud inter-preted as containing six chromosomes.The fourth type of response to colchicine is
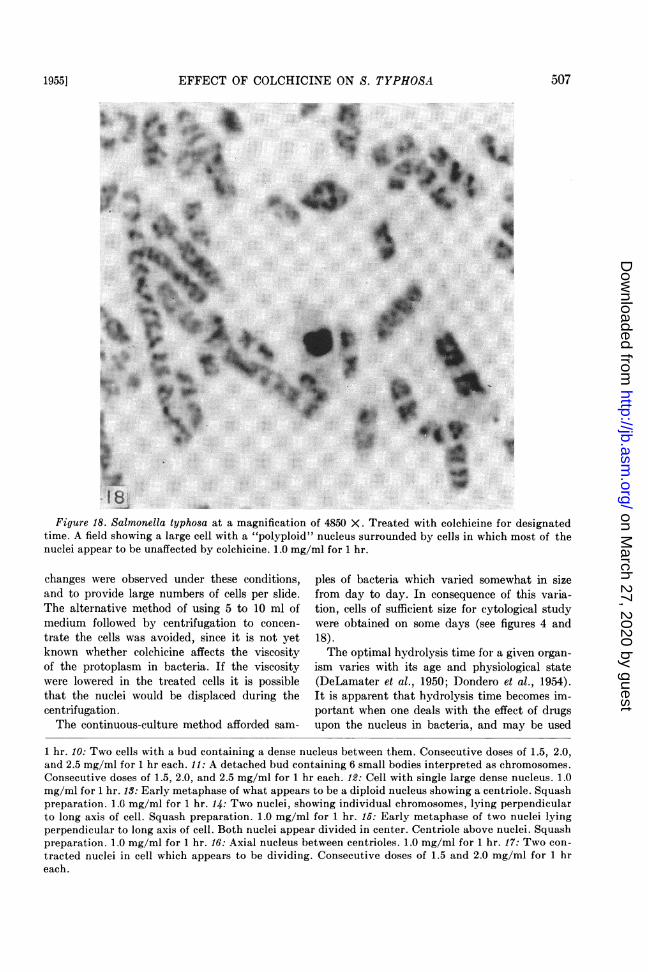
shown by figures 12 to 17. One or two large densenuclei are produced in some cells. These con-figurations were seen when the cells were sub-jected to high and low concentrations of thedrug, but were more prevalent at high concen-trations. Figure 18 shows a cell containing a largedense nucleus which appears to be "polyploid."The nuclei in the surrounding cells, for the mostpart, appear to be unaffected by colchicine.
DISCUSSION
The term polyploidy is used here with reserva-tion. The condition is distinguished from thatinduced in higher forms by c-mitotic substances,such as colchicine, in that a true polyploid nucleusis not constituted and perpetuated. For thisreason it is referred to as "transient polyploidy"(DeLamater et al., 1955).The practice in bacteriology of placing a small
inoculum in the presence of a drug, and observingthe turbidity of the medium at 24 and 48 hrto ascertain whether or not the drug had an ef-fect upon the organisms, has been modified inthis work. It was considered desirable to have asthe test organisms large populations of more orless homogenous, actively multiplying bacteria.This would closely approach the conditions underwhich the action of colchicine has been observedin plants and animals (Levine, 1951). Experi-mentation was confined to the logarithmic phaseof growth, since this is the period of most rapidmultiplication and, hence, the period in whichthe mitotic cycle would be most in evidence.Since the bacteria in most of these experimentswere 3.5 hr old, 2.5 hr of the logarithmic periodremained for observation of nuclear changes in-duced by action of the drug.
Small quantities of medium were employed toavoid, so far as possible, anaerobic conditionsat the bottom of the tube, since no nuclear
14 15 16 17Figures 7-17. All photographs are of Salmonella typhosa at a magnification of 4850 X. All cells were
treated with colchicine for designated time. 7: An asteroid cell. Lower branch appears to be separatingfrom main body of cell. 1 mg/ml for 2 hr. 8: "Large body" containing large dense nucleus bounded bywhat appears to be a "nuclear membrane." 1 mg/ml for 2 hr. 9: Two cells, with axial nuclei, whichappear to have merged at their tips to produce a large bud containing two dense nuclei. 1 mg/ml for
Figure 18. Salmonella typhosa at a magnification of 4850 X. Treated with colchicine for designatedtime. A field showing a large cell with a "polyploid" nucleus surrounded by cells in which most of thenuclei appear to be unaffected by colchicine. 1.0 mg/ml for 1 hr.
changes were observed under these conditions,and to provide large numbers of cells per slide.The alternative method of using 5 to 10 ml ofmedium followed by centrifugation to concen-trate the cells was avoided, since it is not yetknown whether colchicine affects the viscosityof the protoplasm in bacteria. If the viscositywere lowered in the treated cells it is possiblethat the nuclei would be displaced during thecentrifugation.The continuous-culture method afforded sam-
ples of bacteria which varied somewhat in sizefrom day to day. In consequence of this varia-tion, cells of sufficient size for cytological studywere obtained on some days (see figures 4 and18).The optimal hydrolysis time for a given organ-
ism varies with its age and physiological state(DeLamater et al., 1950; Dondero et al., 1954).It is apparent that hydrolysis time becomes im-portant when one deals with the effect of drugsupon the nucleus in bacteria, and may be used
1 hr. 10: Two cells with a bud containing a dense nucleus between them. Consecutive doses of 1.5, 2.0,and 2.5 mg/ml for 1 hr each. 11: A detached bud containing 6 small bodies interpreted as chromosomes.Consecutive doses of 1.5, 2.0, and 2.5 mg/ml for 1 hr each. 12: Cell with single large dense nucleus. 1.0mg/ml for 1 hr. 13: Early metaphase of what appears to be a diploid nucleus showing a centriole. Squashpreparation. 1.0 mg/ml for 1 hr. 14: Two nuclei, showing individual chromosomes, lying perpendicularto long axis of cell. Squash preparation. 1.0 mg/ml for 1 hr. 15: Early metaphase of two nuclei lyingperpendicular to long axis of cell. Both nuclei appear divided in center. Centriole above nuclei. Squashpreparation. 1.0 mg/ml for 1 hr. 16: Axial nucleus between centrioles. 1.0 mg/ml for 1 hr. 17: Two con-tracted nuclei in cell which appears to be dividing. Consecutive doses of 1.5 and 2.0 mg/ml for 1 hreach.
as an indicator of an effect of the drug. In someinstances the optimal hydrolysis time was doubledas compared to untreated populations of bacteria.The question of whether ribonucleic acid is in-creased in amount due to colchicine action, or hasbecome more highly polymerized, or both, is atpresent unresolved.
All the effects of colchicine presented in thispaper were observed in 1- and 2-ml quantitiesof medium in Wassermann tubes under normalatmospheric conditions at 37 C. When S. typhosawas exposed to colchicine under anaerobic con-ditions and in aerated cultures, no significantdifferences in the nuclei, as compared to un-treated populations, were observed. Optimalhydrolysis time was unchanged in the anaerobiccultures, but was doubled in the aerobic cultures.
Cytologically, S. typhosa displays at least fourdifferent and definable responses to colchicine:(a) very long cells in which the chromosomes ap-pear to have multiplied within the "nuclear mem-brane" so as to produce axial nuclei; (b) "largebodies," fusiform cells, or "Y"-shaped cellscontaining either one dense nucleus or whatappear to be "polyploid" nuclei; (c) round cells,containing nuclear material, either lying free orattached to one or two bacteria; (d) cells whichcontain one or two dense nuclei which appear tobe "polyploid." Responses (a) and (d) are sug-gestive of the results obtained by subjectingB. megaterium to oxytetracycline (DeLamater,1953). Similar results were reported by Sterzl(1948, 1949) and Johnson and Gray (1949) withother bacteria under different conditions. Theseindividual effects can be controlled to some ex-tent in their inducement by specific concentra-tions of, and duration of exposure to, the drug.
Figures 4 to 6 are interpreted as stages in therecovery of the nuclei from a transient polyploidstate. The configurations may also be interpretedas cells which were belatedly or irreversibly af-fected by the drug. The former interpretation ispreferred since these stages resemble recoverystages seen in B. megaterium which were exposedto oxytetracycline (DeLamater, 1953). In thelatter organism a 100 per cent response was ob-tained. The exact time at which recovery beginsin S. typhosa is not known, since living cells werenot studied as with B. megaterium.
Levine (1951) observed that in the root tipsof Allium cepa 3.6 peI cent of the cells were inmetaphase. After 24 hr exposure to 0.01 per cent
colchicine, 14.88 per cent metaphases were seen.The 11.2 per cent increase in metaphases over thenumber counted in untreated roots appeared tobe the maximum effect. Giant cells with ab-normally shaped nuclei were seen. Levine con-cluded that colchicine, in a given concentrationfor a given period up to and including 24 hr, in-creases the number of metaphases by arrestingthe nucleus in that phase of division. Levinefurther states: "The lack of an adequate spindlemechanism, however, does not prevent thechromosomes from dividing, but apparentlyits failure to function prevents cell division andgives rise to giant cells with multiploid chromo-some numbers." The extreme variation in thenumber of metaphases counted at the 24th hourby Levine indicated the beginning of the toxicinfluence exerted by colchicine or the inabilityof the colchicine to hold the dividing nucleus inmetaphase any longer.
It is interesting to note the similarity of theeffects of colchicine upon S. typhosa and Levine'sresults with A. cepa. In S. typhosa approximately10 per cent of the cells were affected. Among theeffects noted were the inhibition of division, theconsequent production of giant cells, the enlarge-ment of the nuclei due to the reduplication of thechromosomes, and the configurations which wereinterpreted as recovery stages.
If we are to accept Levan's (1951) premise thatpolyploidization under the action of colchicine isdirect evidence for the presence of a spindle anda true mitotic process, the evidence presentedhere would substantiate the concept that thesebacteria possess a mitotic process.Ludford (1953) has stated that some mitotic
poisons, such as colchicine, produce the sameaction on all cells which normally develop aspindle during mitosis. In these cases the chem-icals presumably act on a physico-chemicalmechanism common to all cells. The colchicineresponse in S. typhosa appeared, in general, to besimilar to those shown in numerous plant andanimal experiments.
SUMMARY
The effect of colchicine on actively multiply-ing Salmonella typhosa was investigated by meansof growth curves and cytologic procedures.The drug was effective in a wide range of con-
centrations and exerted its influence on the bac-teria for only a few hours. Four different de-
finable responses were noted. These individualeffects could be controlled to some extent in theirinducement by specific concentrations of, andduration of exposure to, the mitotic inhibitor.The cellular division and the nuclear mechanismwere inhibited in approximately 10 per cent ofthe cells. This resulted in what is interpreted as"polyploidy." In many instances the volume ofcells was greatly increased. Exposure to colchicineincreased the optimal hydrolysis time of the popu-lations of cells.
REFERENCESBERGERSEN, F. J. 1953 Cytological changes
induced in Bacterium coli by chloramphenicol.J. Gen. Microbiol., 9, 353-356.
CASSEL, W. A. 1951 A procedure for the simul-taneous demonstration of the cell walls andthe chromatinic bodies of bacteria. J. Bac-teriol., 62, 239.
DELAMATER, E. D. 1953 Structure and divisionof the bacterial nucleus. Symposium onBacterial Cytology. Intern. Congr. Micro-biol., Rome, Suppl. Rend., ist. super. sanita.,108-135.
DELAMATER, E. D. 1954 A demonstration ofthe antimitotic nature of the inhibition pro-duced in bacteria by antibiotics and otherdrugs. Am. J. Syphilis, Gonorrhea, VenerealDiseases, 38, 593-598.
DELAMATER, E. D., HUNTER, M. E., SZYBALSKI,W., AND BRYSON, V. 1955 Chemically in-duced aberrations of mitosis in bacteria. J.Gen. Microbiol., 12, 203-212.
DELAMATER, E. D., MESCON, H., AND BARGER,J. D. 1950 The chemistry of the Feulgenreaction and related histo- and cytochemicalmethods. J. Invest. Dermatol., 14, 133-151.
DONDERO, N. C., ADLER, H. I., AND ZELLE, M. R.1954 Quantification of the Feulgen reactionin bacteria. J. Bacteriol., 68, 483-492.
DUGUID, J. P. 1948 The influence of culturalconditions on the morphology of Bacteriumaerogenes with reference to nuclear bodies andcapsule size. J. Pathol. Bacteriol., 60, 265-274.
GARDNER, A. D. 1925 The growth of branchingforms of bacilli ("3 point multiplication").J. Pathol. Bacteriol., 28, 189-194.
JOHNSON, F. H., AND GRLAY, D. H. 1949 Nucleiand large bodies of luminous bacteria in rela-
tion to salt concentration, osmotic pressure,temperature, and urethane. J. Bacteriol.,58, 675-688.
LEVAN, A. 1951 Discussion to DeLamater'spaper: A new cytological basis for bacterialgenetics. Cold Spring Harbor SymposiaQuant. Biol., 16, 409-410.
LEVINE, M. 1951 The action of colchicine oncell division in human cancer, animal, andplant tissues. Ann. N. Y. Acad. Sci., 51,1365-1408.
LUDFORD, R. J. 1953 Chemically induced de-rangements of cell division. J. Roy. Mi-croscop. Soc., 73, 1-23.
MELLON, R. R. 1925 Studies in microbic hered-ity. I. Observations on a primitive form ofsexuality (zygospore formation) in the colon-typhoid group. J. Bacteriol., 10, 481-501.
MINSAVAGE, E. J. 1955 A permanent mountingprocedure for bacteria. Stain Technol., 30,231-233.
MINSAVAGE, E. J., AND DELAMATER, E. D. 1955The nuclear cytology of Salmonella typho8a.J. Bacteriol., 70, 482-483.
MORTON, H. E. 1943 An improved technic forgrowing microorganisms under anaerobicconditions. J. Bacteriol., 46, 373-376.
MORTON, H. E., AND ENGLEY, F. B., JR. 1945The protective action of dysentery bacteri-ophage in experimental infections in mice.J. Bacteriol., 49, 245-255.
O5TERGREN, G. 1951 Narcotized mitosis andthe precipitation hypothesis of narcosis.Colloq. Intern. Centre Nat. recherche Sci.(Paris), 26, 77-88.
STEMPEN, H., AND HuTcMNSON, W. G. 1951The formation and development of largebodies in Proteus vulgaris OX-19. J. Bac-teriol., 61, 337-344.
STERZL, J. 1948 Tvarova menlivost jardernehmoty a zmeny geneticke u Escherichia colivyvolane kolchicinem. Biol. Listy, 29, 198-206.
STERZL, J. 1949 Morphological variability ofthe nuclear substance and genetic changesinduced by colchicine in Escherichia coli.Nature, 163, 28.
ZAPF, K. 1953 Licht-und elektronenoptischeUntersuchungen uber die Ultraschallwirkungauf Bacterium coli unter Penicillineinfluss.Zentr. Bakteriol. Parasitenk., Abt. I, Orig.,159, 436-453.