Page 1
RESEARCH PAPER
Spatiotemporal distribution of bacteriochlorophyllsin the meromictic Lake Suigetsu, Japan
Ryuji Kondo • Misa Kodera • Yumi Mori •
Takahiko Okamura • Shinya Yoshikawa •
Kaori Ohki
Received: 15 November 2012 / Accepted: 15 July 2013
� The Japanese Society of Limnology 2013
Abstract The spatiotemporal distribution of chlorophyll
pigments (chloropigments) in the water column of a mer-
omictic lake, Lake Suigetsu (Fukui, Japan), was investi-
gated. Water samples were collected from the central basin
of Lake Suigetsu bimonthly between May 2008 and March
2010 at appropriate depths, including the oxic surface,
oxic–anoxic interface, and anoxic bottom layers. Chloro-
phyll a, related to cyanobacteria and eukaryotic phyto-
plankton, was detected throughout the water column during
the years of the study, whereas bacteriochlorophyll e,
related to brown-colored green sulfur bacteria, was detec-
ted in the anoxic layers below the chemocline at a maxi-
mum concentration of 825 lg L-1. The concentration of
bacteriochlorophyll e was generally maximal at or just
below the chemocline of the lake. The cellular content of
bacteriochlorophyll e was estimated to be low in the upper
part of the chemocline and tended to increase with
increasing water depth. Bacteriochlorophyll a, which was
presumably related to purple sulfur bacteria, was only
detected at the chemocline during summer and autumn at
concentrations of 5.4–16.3 lg L-1. Our analysis of the
chloropigment distribution for the two years of the study
suggested that brown-colored green sulfur bacteria are the
predominant phototroph in the anoxic layers of Lake Sui-
getsu, and that these play a significant role in the carbon
and sulfur cycling of the lake, especially from spring to
summer.
Keywords Bacteriochlorophyll � Chlorophyll �HPLC � Meromictic lake
Introduction
Lake Suigetsu is a typical meromictic lake located on the
coast at Wakasa Bay (which faces the Sea of Japan) in
Fukui, Japan. This lake is part of a chain of five lakes with
a maximum depth of 34 m. Two different types of water
enter the lake—freshwater (through Lake Mikata) and
saltwater (through Lakes Kugushi and Hiruga), so Lake
Suigetsu is a meromictic lake characterized by a permanent
chemocline at a depth of 5–6 m that separates the aerobic
freshwater epilimnion from the anaerobic, saline, sulfido-
genic hypolimnion (Kondo et al. 2000; Matsuyama and
Saijo 1971; Takahashi and Ichimura 1968). Thus, the
deeper layers of the lake stagnate (Matsuyama 1973). With
prolonged meromixis, the anoxic water chemistry of Lake
Suigetsu is dominated by high levels of sulfides (Kondo
et al. 2000; Matsuyama and Saijo 1971).
Lake Suigetsu has attracted interest concerning micro-
bial sulfate reduction and sulfur oxidation in relation to
both the carbon and sulfur cycles because of the charac-
teristics of its water, including the chemocline in the water
column and high concentrations of sulfide in the monim-
olimnion. Thus, we are currently using molecular methods
to investigate the diversity and distribution of sulfate-
reducing bacteria in the water column of Lake Suigetsu
(Kondo et al. 2006; Kondo and Butani 2007). Lake Sui-
getsu is highly sulfidic and the chemocline water is colored
pale red or pink, so it is assumed that this colored water is a
dense bloom of phototrophic sulfur bacteria that is
responsible for microbial sulfur oxidation and carbon fix-
ation. Matsuyama and Saijo (1971) and Takahashi and
Handling Editor: Hideaki Miyashita
R. Kondo (&) � M. Kodera � Y. Mori � T. Okamura �S. Yoshikawa � K. Ohki
Department of Marine Bioscience, Fukui Prefectural University,
Obama, Fukui 917-0003, Japan
e-mail: [email protected]
123
Limnology
DOI 10.1007/s10201-013-0415-y
Page 2
Ichimura (1968) reported that maximum photosynthetic
bicarbonate fixation took place at the chemocline, and
suggested that sulfur bacteria were involved in the photo-
synthetic carbon assimilation that occurred in that region.
A few studies were carried out to demonstrate the existence
of phototrophic sulfur bacteria in the water column of Lake
Suigetsu, utilizing denaturing gradient gel electrophoresis
(DGGE) of the 16S rRNA gene fragment (Kondo et al.
2009), chlorophyll pigment (chloropigment) analyses
(Okada et al. 2007; Takahashi and Ichimura 1968), and a
functional gene sequence analysis (Mori et al. 2010). More
recently, we demonstrated that brown-colored green sulfur
bacterial counts, as estimated using a new quantitative PCR
method targeting a portion of the a-subunit of the dissim-
ilatory sulfite reductase gene (dsrA), account for up to
35 % of the total bacterial count in the chemocline of Lake
Suigetsu (Mori et al. 2013). However, this molecular
technique for counting populations can detect dead and/or
metabolically inactive microbial cells. On the other hand,
oxygenic pico-sized cyanobacteria were detected at a
maximum density of 4.8 9 105 cell mL-1 even in the
anoxic chemocline of Lake Suigetsu (Okada et al. 2007).
So, it is still unclear which phototrophic microorganisms
are physiologically active in the anoxic layer of Lake
Suigetsu.
Chloropigment analysis potentially provides quantita-
tive estimates of phototrophic bacteria in environments
without the biases attributed to the efficiencies of PCR
amplification and DNA extraction (Nakajima et al. 2003).
Chloropigment analysis has been used for the in situ
characterization of phototrophic sulfur bacterial assem-
blages in stratified water columns (Manske et al. 2005;
Nakajima et al. 2003; Okada et al. 2007). In some cases,
the abundance of phototrophic sulfur bacteria has been
reported as the amount of bacteriochlorophylls (Van
Gemerden and Mas 1995). Such abundances, as reported in
previous studies, vary by several orders of magnitude in the
water column. In the work described in the present paper,
we examined the distribution of phototrophic microbes
using a diagnostic chloropigment analysis to determine the
type of phototrophic population that plays a potential role
in photosynthesis in the anoxic layers of Lake Suigetsu and
to identify spatiotemporal shifts in the phototrophic
populations.
Materials and methods
Samples
Water samples were collected from the central basin of
Lake Suigetsu (35�350 N, 135�530 E) bimonthly between
May 2008 and March 2010 using a Kitahara’s water
sampler (Rigo Co., Ltd., Saitama, Japan) at appropriate
depths, including the oxic surface, oxic–anoxic interface,
and anoxic bottom layers. The samples were immediately
poured into brown bottles to prevent light exposure. All
samples were kept in an ice-cooled box and transported to
the laboratory within a few hours of sampling. Tempera-
ture, salinity, and dissolved oxygen (DO) concentration
were measured using oxygen meters (Model 85; YSI,
Yellow Springs, OH, USA and HQ30d; HACH, Loveland,
CO, USA). The vertical turbidity profile (as kaolin
mg L-1; ppm) was obtained using a turbidity meter (model
PT-1; JFE Advantec Co., Ltd., Kobe, Japan). Vertical
profiles of photosynthetically active radiation (PAR,
400–700 nm) were measured using an LI-192SA under-
water quantum sensor and an LI-1400 datalogger (LI-COR,
Lincoln, NE, USA).
Chemicals and chloropigment preparation
All chemicals used in this study were of analytical grade or
HPLC grade. Chlorophyll a (Chl a) and bacteriochloro-
phyll a (BChl a) were purchased from Wako Pure Chem-
ical Industries (Osaka, Japan) and Sigma–Aldrich Fine
Chemicals (St. Louis, MO, USA), respectively. The bac-
teriochlorophyll e (BChl e) used in this study was extracted
from Chlorobium phaeovibrioides DSM 269T, which was
obtained from Deutsche Sammlung von Mikroorganismen
und Zellkulturen GmbH (DSMZ) and cultured in our
laboratory using DSMZ medium 29 (http://www.dsmz.de/
microorganisms/medium/pdf/DSMZ_Medium29.pdf). These
chloropigments were dissolved in diethyl ether and
used for calibration. The chloropigment concentrations
in the standard solutions were calculated using molar
extinction coefficients of 8.98 9 104 M-1 cm-1 for Chl
a at 661 nm, 9.70 9 104 M-1 cm-1 for BChl a at 771 nm,
and 3.4 9 104 M-1 cm-1 for BChl e at 647 nm (Takamiya
et al. 2003), after performing absorption spectrum mea-
surements using a PharmaSpec UV-1700 spectrophotom-
eter (Shimadzu, Kyoto, Japan).
Pigment extraction
A 100-mL water aliquot was filtered through a 25-mm GF-
75 glass fiber filter (nominal pore size, 0.3 lm; Advantec,
Tokyo, Japan) to collect suspended particulate matter for
subsequent chloropigment extraction. The filters were
stored in a 2-mL screw-cap plastic tube at -135 �C in the
dark until processed. The filter samples were suspended in
1 mL of acetone and sonicated (10 W) for 1 min on ice.
The samples were then centrifuged at 20,0009g for 15 min
at 4 �C and the supernatant was filtered through a 0.45-lm
Cosmonice Filter S (Nacalai Tesque, Kyoto, Japan). A
portion (500 lL) of the filtrate was evaporated in vacuo
Limnology
123
Page 3
using a centrifugal vacuum evaporator (Iwaki VEC-100;
AGC Techno Glass, Chiba, Japan). After evaporation, the
chloropigment was redissolved in 50 ll of acetone. All
processes were performed under dim light conditions.
High-performance liquid chromatography
and quantification
The acetone extract was analyzed via high-performance
liquid chromatography (HPLC), as described previously
(Okada et al. 2007) with some modification. Briefly, the
HPLC system consisted of a Class LC-VP/SPD-M10AVP
system (Shimadzu, Kyoto, Japan). Chloropigments were
separated with a Wakosil-II 5C18HG column
(250 9 4.6 mm i.d.; Wako Pure Chemical Industries,
Osaka, Japan) and a guard column, and detected with a
photodiode array detector (SPD-M10AVP; Shimadzu). All
analyses were performed at 40 �C with a flow rate of
1 mL min-1 for 60 min. Methanol was used as the solvent.
Aliquots of 20-ll chloropigment extracts or standard
solutions were injected into the HPLC system. Chromato-
grams were digitally recorded on a computer using the
CLASS-VP chromatography data system (Shimadzu). The
chloropigment concentrations were quantified at wave-
lengths of 431 nm for Chl a, 770 nm for BChl a, and
473 nm for BChl e. Three BChl e homologs can be
detected by our HPLC procedure (Okada et al. 2007).
Concentrations of BChl e were calculated by summing the
three HPLC peaks.
Results
Water column profiles
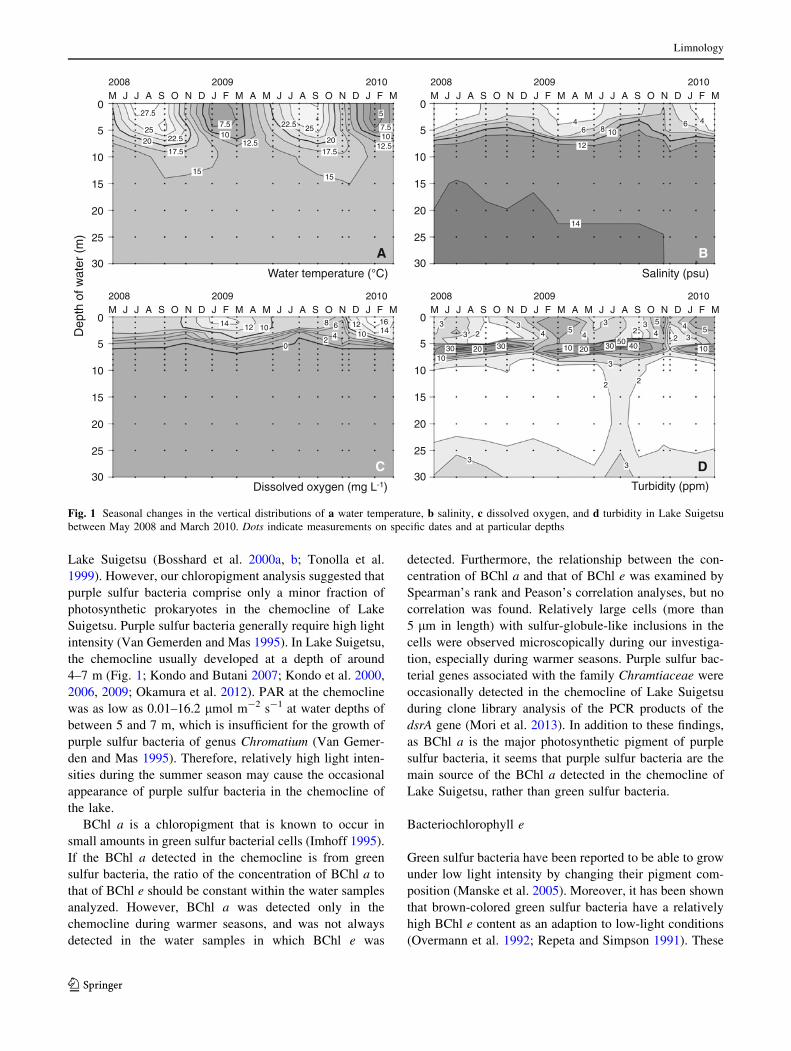
Figure 1 shows seasonal changes in the depth distributions
of the physicochemical properties in the central basin of
Lake Suigetsu during the sample collection times. The
epilimnion temperature in Lake Suigetsu ranged from 3 �C
in the winter to 30 �C in the summer. The hypolimnion
temperature was, however, constant at ca. 14 �C through-
out the year. The salinity was 2–4 practical salinity units
(psu) for the epilimnion water and 12–15 psu for the
hypolimnion water. Surface water was saturated with DO,
and the DO concentration decreased rapidly below 3–5 m
to below the detection limit at 5–8 m and down to the
bottom at 34 m. This observation shows the stagnation of
the anoxic saline water in deeper layers from approxi-
mately 6 m to the bottom at 34 m. Turbidity was 2–8 ppm
in the surface layer and increased to its maximum at the
oxic–anoxic interface zone. Turbidity decreased with depth
between 9 and 25 m, remained constant from 25 m to
30 m, and below that increased slightly with depth.
Distribution of chloropigments in the water column
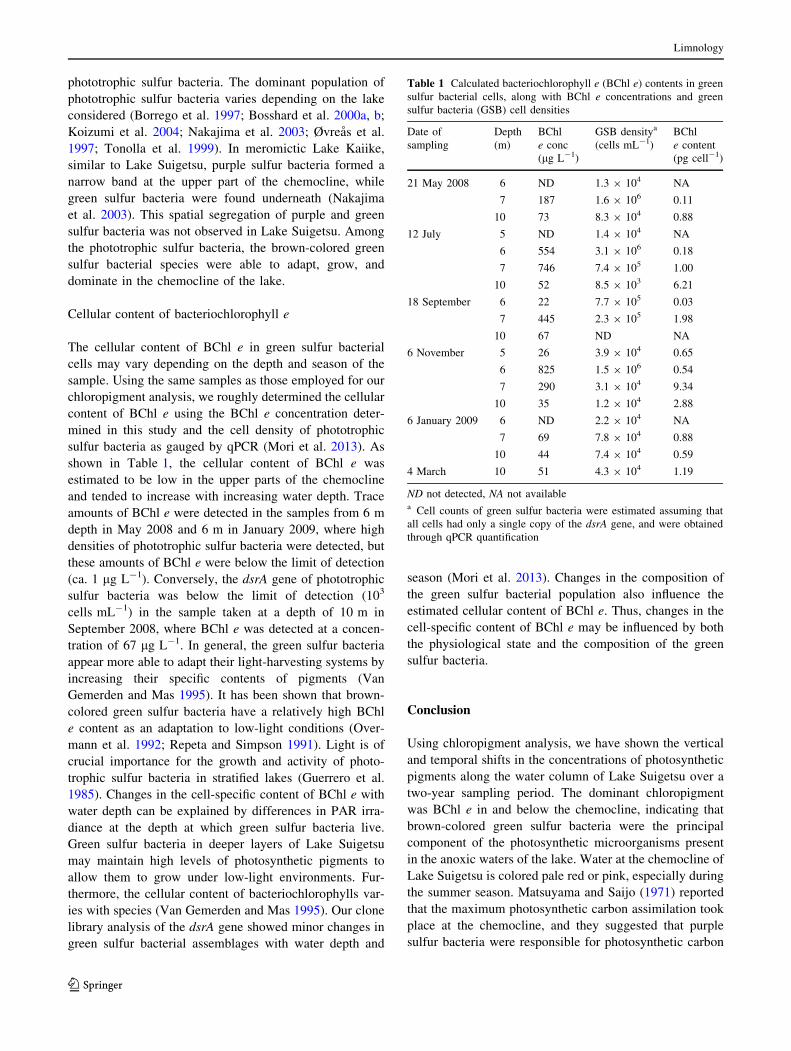
Chlorophyll a (Chl a) was detected in all samples,
including those from the oxic surface, oxic–anoxic inter-
face, and anoxic bottom layers, at concentrations ranging
from 3.5 to 80.6 lg L-1 (Fig. 2). High concentrations of
Chl a were observed in the surface water in January and
March in both years. Apart from the high concentrations
observed in the epilimnion in winter, Chl a concentrations
ranged from 3.6 to 16.8 lg L-1 in the surface water and
increased to maxima of 5.6–11.6 lg L-1 at the chemo-
cline, and then gradually decreased with water depth
between the vernal and autumnal seasons (Fig. 2).
Bacteriochlorophyll a (BChl a) was detected in the
redox boundary waters between July and November at low
concentrations ranging from 5.4 to 16.3 lg L-1 (Fig. 2).
Bacteriochlorophyll e (BChl e) was detected in and
below the chemocline during all sampling occasions at
high concentrations ranging from 21.6 to 825 lg L-1,
whereas BChl e was not detected in the oxic surface layer
(Fig. 2). The concentrations of BChl e changed with
depth and/or season. A high concentration of BChl
e ([100 lg L-1) was generally detected in metalimnetic
water during the warm season (from July to November),
whereas the concentration decreased from November to
March during the next year.
Discussion
Chlorophyll a
In the surface water of Lake Suigetsu, according to our
microscopic observations, dinoflagellate blooms of Het-
erocapsa occur during the winter season, while cyano-
bacteria such as Microcystis and Anabaena and/or diatoms
dominate during the summer season. High concentrations
of Chl a in the winter season appear to correspond with the
Heterocapsa bloom. The vertical profiles of Chl a concen-
tration with a broad peak at the chemocline correspond to
those of pico-sized cyanobacterioplankton within the Syn-
echococcus/Cyanobium clade (Okada et al. 2007; Ohki
et al. 2012). Ohki et al. (2012) isolated sulfide-tolerating
pico-sized cyanobacteria from the chemocline of Lake
Suigetsu. However, considering the low concentration of
Chl a, the cyanobacteria seem to be less abundant and to
have lower photosynthetic activities than anoxygenic
phototrophs in the chemocline of Lake Suigetsu.
Bacteriochlorophyll a
Purple sulfur bacteria have been reported to form dense
blooms in the chemoclines of meromictic lakes similar to
Limnology
123
Page 4
Lake Suigetsu (Bosshard et al. 2000a, b; Tonolla et al.
1999). However, our chloropigment analysis suggested that
purple sulfur bacteria comprise only a minor fraction of
photosynthetic prokaryotes in the chemocline of Lake
Suigetsu. Purple sulfur bacteria generally require high light
intensity (Van Gemerden and Mas 1995). In Lake Suigetsu,
the chemocline usually developed at a depth of around
4–7 m (Fig. 1; Kondo and Butani 2007; Kondo et al. 2000,
2006, 2009; Okamura et al. 2012). PAR at the chemocline
was as low as 0.01–16.2 lmol m-2 s-1 at water depths of
between 5 and 7 m, which is insufficient for the growth of
purple sulfur bacteria of genus Chromatium (Van Gemer-
den and Mas 1995). Therefore, relatively high light inten-
sities during the summer season may cause the occasional
appearance of purple sulfur bacteria in the chemocline of
the lake.
BChl a is a chloropigment that is known to occur in
small amounts in green sulfur bacterial cells (Imhoff 1995).
If the BChl a detected in the chemocline is from green
sulfur bacteria, the ratio of the concentration of BChl a to
that of BChl e should be constant within the water samples
analyzed. However, BChl a was detected only in the
chemocline during warmer seasons, and was not always
detected in the water samples in which BChl e was
detected. Furthermore, the relationship between the con-
centration of BChl a and that of BChl e was examined by
Spearman’s rank and Peason’s correlation analyses, but no
correlation was found. Relatively large cells (more than
5 lm in length) with sulfur-globule-like inclusions in the
cells were observed microscopically during our investiga-
tion, especially during warmer seasons. Purple sulfur bac-
terial genes associated with the family Chramtiaceae were
occasionally detected in the chemocline of Lake Suigetsu
during clone library analysis of the PCR products of the
dsrA gene (Mori et al. 2013). In addition to these findings,
as BChl a is the major photosynthetic pigment of purple
sulfur bacteria, it seems that purple sulfur bacteria are the
main source of the BChl a detected in the chemocline of
Lake Suigetsu, rather than green sulfur bacteria.
Bacteriochlorophyll e
Green sulfur bacteria have been reported to be able to grow
under low light intensity by changing their pigment com-
position (Manske et al. 2005). Moreover, it has been shown
that brown-colored green sulfur bacteria have a relatively
high BChl e content as an adaption to low-light conditions
(Overmann et al. 1992; Repeta and Simpson 1991). These
Dep
th o
f wat
er (
m)
Water temperature (°C)
A
27.5
2522.52017.5
1515
12.5107.5
17.5
20
22.5 25
12.5107.5
5
Salinity (psu)
B
4 46
68 10
12
14
Dissolved oxygen (mg L-1)
C
14 1012468
141612
102
0
Turbidity (ppm)
D
2
3
4
5
10 20 30 4050
2
3
45
2
3
45
10203010
30
223
3 3 3 3
3
4
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
0
5
10
15
20
25
30
010290028002M J J A S O N D J F M A M J J A S O N D J F M
010290028002M J J A S O N D J F M A M J J A S O N D J F M
010290028002M J J A S O N D J F M A M J J A S O N D J F M
010290028002M J J A S O N D J F M A M J J A S O N D J F M
Fig. 1 Seasonal changes in the vertical distributions of a water temperature, b salinity, c dissolved oxygen, and d turbidity in Lake Suigetsu
between May 2008 and March 2010. Dots indicate measurements on specific dates and at particular depths
Limnology
123
Page 5
observations indicate that chloropigment concentrations in
natural environments do not necessarily represent the cell
density of phototrophic sulfur bacteria. The relationship
between the BChl e concentration and the phototrophic
sulfur bacterial density as determined by a quantitative
real-time PCR (qPCR) targeting the dsrA gene (Mori et al.
2013) was examined using Spearman and Pearson corre-
lation analyses, but only a weak correlation was found
(q = 0.516, p = 0.061 for the Spearman test and
r = 0.576, p = 0.031 for the Pearson test). This weak
correlation suggests that the BChl e detected in the
chemocline of Lake Suigetsu originates from green sulfur
bacteria.
The high concentrations of BChl e are within the range
of, or higher than, values previously reported for other
meromictic lakes (Borrego et al. 1997; Nakajima et al.
2003; Van Gemerden and Mas 1995 and references
therein). Green sulfur bacteria of Chlorobium, Prosthec-
ochloris, and Pelodictyon have been detected mainly in or
just below the chemocline of Lake Suigetsu by DGGE
analysis of 16S rRNA gene fragments (Kondo et al. 2009)
and clone library analysis of PCR products of the dsrA
gene (Mori et al. 2010; 2013). These results, together with
our chloropigment determination, indicate that brown-col-
ored green sulfur bacteria form the dense bacterial layer in
the chemocline of Lake Suigetsu. In general, the dominant
species of green sulfur bacteria is Chlorobium phaeobac-
teroides in meromictic freshwater lakes or Chlorobium
phaeovibrioides and Pelodictyon phaeum in meromictic
brackish waters (Borrego et al. 1997). Phototrophic sulfur
bacterial assemblages in the saline meromictic Lake Sui-
gesu coincide with this general trend in species habitat
segregation.
Two groups of phototrophic sulfur bacteria, purple sul-
fur bacteria and green sulfur bacteria, inhabit similar
environments, where anoxic layers containing reduced
sulfur compounds are exposed to light. However, due to
differences in the physiological characteristics (such as
optimum temperature, light illumination/wavelength, and
electron donor concentrations for growth) between the two
groups (Van Gemerden and Mas 1995), environmental
conditions may determine the dominant group of
1
5
6
7
10
1
5
6
7
10
1
5
6
7
10
1
5
6
7
10
1
5
6
7
10
1
5
6
7
10
100 101 102 103 100 101 102 103 100 101 102 103 100 101 102 103 100 101 102 103 100 101 102 103
100 101 102 103 100 101 102 103 100 101 102 103 100 101 102 103 100 101 102 103 100 101 102 103
1
5
6
7
10
1
4
5
6
10
1
5
6
7
10
1
5
6
7
10
1
5
6
7
10
1
5
6
7
10
Dep
th o
f wat
er (
m)
Chloropigment concentration (µg L-1)
Chloropigment concentration (µg L-1)
May 2008 July 2008 September 2008 November 2008 January 2009 March 2009
May 2009 July 2009 September 2009 November 2009 January 2010 March 2010
Fig. 2 Seasonal changes in the vertical distributions of chlorophyll
a (open bar), bacteriochlorophyll a (stippled bar), and bacteriochlo-
rophyll e (cross-hatched bar) in the water column of Lake Suigetsu
between May 2008 and March 2010. Error bars represent the
standard error of the mean (n = 3)
Limnology
123
Page 6
phototrophic sulfur bacteria. The dominant population of
phototrophic sulfur bacteria varies depending on the lake
considered (Borrego et al. 1997; Bosshard et al. 2000a, b;
Koizumi et al. 2004; Nakajima et al. 2003; Øvreas et al.
1997; Tonolla et al. 1999). In meromictic Lake Kaiike,
similar to Lake Suigetsu, purple sulfur bacteria formed a
narrow band at the upper part of the chemocline, while
green sulfur bacteria were found underneath (Nakajima
et al. 2003). This spatial segregation of purple and green
sulfur bacteria was not observed in Lake Suigetsu. Among
the phototrophic sulfur bacteria, the brown-colored green
sulfur bacterial species were able to adapt, grow, and
dominate in the chemocline of the lake.
Cellular content of bacteriochlorophyll e
The cellular content of BChl e in green sulfur bacterial
cells may vary depending on the depth and season of the
sample. Using the same samples as those employed for our
chloropigment analysis, we roughly determined the cellular
content of BChl e using the BChl e concentration deter-
mined in this study and the cell density of phototrophic
sulfur bacteria as gauged by qPCR (Mori et al. 2013). As
shown in Table 1, the cellular content of BChl e was
estimated to be low in the upper parts of the chemocline
and tended to increase with increasing water depth. Trace
amounts of BChl e were detected in the samples from 6 m
depth in May 2008 and 6 m in January 2009, where high
densities of phototrophic sulfur bacteria were detected, but
these amounts of BChl e were below the limit of detection
(ca. 1 lg L-1). Conversely, the dsrA gene of phototrophic
sulfur bacteria was below the limit of detection (103
cells mL-1) in the sample taken at a depth of 10 m in
September 2008, where BChl e was detected at a concen-
tration of 67 lg L-1. In general, the green sulfur bacteria
appear more able to adapt their light-harvesting systems by
increasing their specific contents of pigments (Van
Gemerden and Mas 1995). It has been shown that brown-
colored green sulfur bacteria have a relatively high BChl
e content as an adaptation to low-light conditions (Over-
mann et al. 1992; Repeta and Simpson 1991). Light is of
crucial importance for the growth and activity of photo-
trophic sulfur bacteria in stratified lakes (Guerrero et al.
1985). Changes in the cell-specific content of BChl e with
water depth can be explained by differences in PAR irra-
diance at the depth at which green sulfur bacteria live.
Green sulfur bacteria in deeper layers of Lake Suigetsu
may maintain high levels of photosynthetic pigments to
allow them to grow under low-light environments. Fur-
thermore, the cellular content of bacteriochlorophylls var-
ies with species (Van Gemerden and Mas 1995). Our clone
library analysis of the dsrA gene showed minor changes in
green sulfur bacterial assemblages with water depth and
season (Mori et al. 2013). Changes in the composition of
the green sulfur bacterial population also influence the
estimated cellular content of BChl e. Thus, changes in the
cell-specific content of BChl e may be influenced by both
the physiological state and the composition of the green
sulfur bacteria.
Conclusion
Using chloropigment analysis, we have shown the vertical
and temporal shifts in the concentrations of photosynthetic
pigments along the water column of Lake Suigetsu over a
two-year sampling period. The dominant chloropigment
was BChl e in and below the chemocline, indicating that
brown-colored green sulfur bacteria were the principal
component of the photosynthetic microorganisms present
in the anoxic waters of the lake. Water at the chemocline of
Lake Suigetsu is colored pale red or pink, especially during
the summer season. Matsuyama and Saijo (1971) reported
that the maximum photosynthetic carbon assimilation took
place at the chemocline, and they suggested that purple
sulfur bacteria were responsible for photosynthetic carbon
Table 1 Calculated bacteriochlorophyll e (BChl e) contents in green
sulfur bacterial cells, along with BChl e concentrations and green
sulfur bacteria (GSB) cell densities
Date of
sampling
Depth
(m)
BChl
e conc
(lg L-1)
GSB densitya
(cells mL-1)
BChl
e content
(pg cell-1)
21 May 2008 6 ND 1.3 9 104 NA
7 187 1.6 9 106 0.11
10 73 8.3 9 104 0.88
12 July 5 ND 1.4 9 104 NA
6 554 3.1 9 106 0.18
7 746 7.4 9 105 1.00
10 52 8.5 9 103 6.21
18 September 6 22 7.7 9 105 0.03
7 445 2.3 9 105 1.98
10 67 ND NA
6 November 5 26 3.9 9 104 0.65
6 825 1.5 9 106 0.54
7 290 3.1 9 104 9.34
10 35 1.2 9 104 2.88
6 January 2009 6 ND 2.2 9 104 NA
7 69 7.8 9 104 0.88
10 44 7.4 9 104 0.59
4 March 10 51 4.3 9 104 1.19
ND not detected, NA not availablea Cell counts of green sulfur bacteria were estimated assuming that
all cells had only a single copy of the dsrA gene, and were obtained
through qPCR quantification
Limnology
123
Page 7
assimilation in this zone. However, brown-colored green
sulfur bacteria species are responsible for the pale-red
colored water of the chemocline; they appear to be active
and play an important role in carbon assimilation and
sulfide oxidation in Lake Suigesu. Although chloropigment
analysis is useful for the in situ characterization of photo-
trophic sulfur bacterial assemblages in nature, it remains to
be determined whether the green sulfur bacteria contribute
to carbon assimilation in Lake Suigetsu as well as the
oxidation of reduced sulfur compounds. Molecular analy-
ses such as stable-isotope probing and mRNA quantifica-
tion of functional genes could be useful in this respect.
Acknowledgments We are grateful to N. Kawahara and J. Butani
from our laboratory for their assistance with the field sampling. We
also gratefully acknowledge the constructive comments of anony-
mous reviewers and the handling editor and their suggestions for
improving this paper. This study was supported in part by a Fukui
Prefectural Fund for the Promotion of Science to R.K.
References
Borrego CM, Gracia-Gil LJ, Vila X, Cristina XP, Figueras JB, Abella
CA (1997) Distribution of bacteriochlorophyll homologs in
natural populations of brown-colored phototrophic sulfur bacte-
ria. FEMS Microbiol Ecol 24:301–309
Bosshard PP, Santini Y, Gruter D, Stettler R, Bachofen R (2000a)
Bacterial diversity and community composition in the chemo-
cline of the meromictic alpine Lake Cadagno as revealed by 16S
rDNA analysis. FEMS Microbiol Ecol 31:173–182
Bosshard PP, Stettler R, Bachofen R (2000b) Seasonal and spatial
community dynamics in the meromictic Lake Cadagno. Arch
Microbiol 174:168–174
Guerrero R, Mantesinos E, Pedros-Alio C, Esteve I, Mas J, Van
Gemerden H, Hofman PAG, Bakker JF (1985) Phototrophic
sulfur bacteria in two Spanish lakes: vertical distribution and
limiting factors. Limnol Oceanogr 30:919–931
Imhoff JF (1995) Taxonomy and physiology of phototrophic purple
bacteria and green sulfur bacteria. In: Blankenship RE, Madigan
MT, Bauer CE (eds) Anoxygenic photosynthetic bacteria.
Kluwer, Dordrecht, pp 1–15
Koizumi Y, Kojima H, Fukui M (2004) Dominant microbial
composition and its vertical distribution in saline meromictic
Lake Kaiike (Japan) as revealed by quantitative oligonucleotide
probe membrane hybridization. Appl Environ Microbiol
70:4930–4940
Kondo R, Butani J (2007) Comparison of the diversity of sulfate-
reducing bacterial communities in the water column and the
surface sediments of a Japanese meromictic lake. Limnology
8:131–141
Kondo R, Kasashima N, Matsuda H, Hata Y (2000) Determination of
thiosulfate in a meromictic lake. Fish Sci 66:1076–1081
Kondo R, Osawa K, Mochizuki L, Fujioka Y, Butani J (2006)
Abundance and diversity of sulphate-reducing bacterioplankton
in Lake Suigetsu, a meromictic lake in Fukui, Japan. Plankton
Benthos Res 1:165–177
Kondo R, Nakagawa A, Mochizkui L, Osawa K, Fujioka Y, Butani J
(2009) Dominant bacterioplankton populations in the meromictic
Lake Suigetsu as determined by denaturing gradient gel
electrophoresis of 16S rRNA gene fragments. Limnology
10:63–69
Manske AK, Glaeser J, Kuyper MMM, Overmann J (2005) Physi-
ology and phylogeny of green sulfur bacteria forming a
monospecific phototrophic assemblage at a depth of 100 meters
in the Black Sea. Appl Environ Microbiol 71:8049–8060
Matsuyama M (1973) Changes in the limnological features of a
meromictic Lake Suigetsu during the years, 1926–1967. J Oce-
anogr Soc Japan 29:131–139
Matsuyama M, Saijo Y (1971) Studies on biological metabolism in a
meromictic Lake Suigetsu. J Oceanogr Soc Japan 27:197–206
Mori Y, Purdy KJ, Oakley BB, Kondo R (2010) Comprehensive
detection of phototrophic sulfur bacteria using PCR primers that
target reverse dissimilatory sulfite reductase gene. Microbes
Environ 25:190–196
Mori Y, Kataoka T, Okamura T, Kondo R (2013) Dominance of green
sulfur bacteria in the chemocline of the meromictic Lake
Suigetsu, Japan, as revealed by dissimilatory sulfite reductase
gene analysis. Arch Micorbiol 195:303–312
Nakajima Y, Okada H, Oguri K, Suga H, Kitazato H, Koizumi Y,
Fukui M, Ohkouchi N (2003) Distribution of chloropigments in
suspended particulate matter and benthic microbial mat of a
meromictic lake, Lake Kaiike, Japan. Environ Microbiol
5:1103–1110
Ohki K, Yamada K, Kamiya M, Yoshikawa S (2012) Morphological,
phylogenetic and physiological studies of pico-cyanobacteria
isolated from the halocline of a saline meromictic lake, Lake
Suigetsu, Japan. Microbes Environ 27:171–178
Okada M, Taniuchi Y, Murakami A, Takeuchi S, Ohtake S, Ohki K
(2007) Abundance of picophytoplankton in the halocline of a
meromictic lake, Lake Suigetsu, Japan. Limnology 8:271–280
Okamura T, Mori Y, Nakano S, Kondo R (2012) Abundance and
bacterivory of heterotrophic nanoflagellates in the meromictic
Lake Suigetsu, Japan. Aquat Microb Ecol 66:149–158
Overmann J, Cypionka H, Pfennig N (1992) An extremely low-light
adapted phototrophic sulfur bacterium from the Black Sea.
Limnol Oceanogr 37:150–155
Øvreas L, Forney L, Daae FL, Torsvik V (1997) Distribution of
bacterioplankton in meromictic Lake Salenvannet, as determined
by denaturing gradient gel electrophoresis of PCR-amplified
gene fragments coding for 16S rRNA. Appl Environ Microbiol
63:3367–3375
Repeta DJ, Simpson DJ (1991) The distribution and recycling of
chlorophyll, bacteriochlorophyll and carotenoids in the Black
Sea. Deep-Sea Res 38:969–984
Takahashi M, Ichimura S (1968) Vertical distribution and organic
matter production of photosynthetic sulfur bacteria in Japanese
lakes. Limnol Oceanogr 13:644–655
Takamiya K, Ikegami I, Ohta H, Ono T, Tanaka A, Terashima I et al
(2003) Encyclopedia of photosynthesis. Japan Scientific Socie-
ties Press, Tokyo (in Japanese)
Tonolla M, Demarta A, Peduzzi R, Hahn D (1999) In situ analysis of
phototrophic sulfur bacteria in the chemocline of meromictic
Lake Cadagno (Switzerland). Appl Environ Microbiol
65:1325–1330
Van Gemerden H, Mas J (1995) Ecology of phototrophic sulfur
bacteria. In: Blankenship RE, Madigan MT, Bauer CE (eds)
Anoxygenic photosynthetic bacteria. Kluwer, Dordrecht,
pp 49–85
Limnology
123