Species delimitation of the white-tailed rubythroat Calliope pectoralis complex (Aves, Muscicapidae) using an integrative taxonomic approach
Yang Liu, Guoling Chen, Qin Huang, Chenxi Jia, Geoff Carey, Paul Leader, Yun Li, Fasheng Zou, Xiaojun Yang, Urban Olsson and Per Alström
Y. Liu ([email protected]), G. Chen, Q. Huang and Y. Li, State Key Laboratory of Biocontrol, College of Ecology and Evolution/School of Life Sciences, Sun Yat-sen Univ., Guangzhou, China. – C. Jia, The National Zoological Museum of China, Inst. of Zoology, Chinese Academy of Sciences, Chaoyang District, Beijing, China. – G. Carey and P. Leader, c/o AEC, Palm Springs, Yuen Long, Hong Kong SAR, China. – F. Zou, Guangdong Entomological Inst./South China Inst. of Endangered Animals, Guangzhou, China. – X. Yang, State Key Laboratory of Genetic Resources and Evolution, Kunming Inst. of Zoology, Chinese Academy of Sciences, Kunming, China. – U. Olsson, Dept of Biology and Environ-mental Sciences, Systematics and Biodiversity, Univ. of Gothenburg, Göteborg, Sweden. – P. Alström, Dept of Animal Ecology, Evolutionary Biol-ogy Centre, Uppsala Univ., Uppsala, Sweden, and Key Laboratory of Zoological Systematics and Evolution, Inst. of Zoology, Chinese Academy of Sciences, Chaoyang District, Beijing, China, and Swedish Species Information Centre, Swedish Univ. of Agricultural Sciences, Uppsala, Sweden.
Our knowledge of the systematics and taxonomy of Asian birds has improved much in the last two decades, and the num-ber of recognised species has increased significantly as a result of in-depth studies using an integrative taxonomic approach. The Sino-Himalayan mountains harbor a high level of passerine diversity. Several allopatric or parapatric taxa that are currently treated as subspecies of polytypic species within that region are likely to deserve full species status, and thus their taxonomic status needs to be revisited. Based on analyses of multilocus data, vocalizations and morphology, we propose that the white-tailed rubythroat Calliope pectoralis should be treated as two species, the Himalayan rubythroat C. pectoralis sensu stricto in the Tian Shan and Himalayan mountains, and the Chinese rubythroat C. tschebaiewi in the mountains of southwestern and north-central China. According to our dating analyses based on mitochondrial loci, these two species diverged approximately 2.2 million yr ago. We further found that C. tschebaiewi was paraphyletic to C. pectoralis sensu stricto in nuclear data, which demonstrates a state of mitonuclear discordance that warrants further work. Our results sug-gest that geographic changes and glacial cycles in the Pleistocene may have caused allopatric divergence in the C. pectoralis complex. Our study stresses the importance of applying an integrative taxonomy approach to fully unravel the true avian diversity in the Sino-Himalayan Mountains.
Species is a fundamental concept in biology, and is central for biodiversity estimates and conservation management. Species delimitation and estimates of number of spe-cies represent crucial efforts towards understanding global biological diversity (de Queiroz 2007, Tobias et al. 2010, Fujita et al. 2012). Ongoing progress in coalescent theory and molecular phylogenetics provide methodological tools to assess evolutionary relationships and infer taxonomic affinities among members of species complexes (Drummond et al. 2012, Fujita et al. 2012). Although there is continual debate over species criteria and species concepts (de Queiroz 2007, Tobias et al. 2010), it is now widely agreed that species should be defined based on multiple independent lines of evidence, such as multiple, functionally independent char-acters, e.g. morphological, bioacoustic and behavioural data (Helbig et al. 2002), and/or multilocus DNA data (Yang and Rannala 2010). This modern approach is now generally referred to as integrative taxonomy (Padial et al. 2010).
The understanding of avian taxonomy and systematics in Asia is far from complete due to its high bird diversity, complex biogeography and the many taxa that have not been investigated using modern integrative approaches. However, continual efforts in molecular phylogenetics have steadily improved our knowledge of higher-level systematics of conti-nental Asian birds in the last two decades (Olsson et al. 2004, Nylander et al. 2008, Gelang et al. 2009, Päckert et al. 2010, Sangster et al. 2010, Alström et al. 2011a, b, 2013a, 2014, Yeung et al. 2011, Moyle et al. 2012, Zuccon et al. 2012, Drovetski et al. 2013, Tietze et al. 2013, Price et al. 2014). In addition, an increasing number of studies using integra-tive approaches have recently been carried out to address taxonomic questions in closely related species (Alström et al. 2007, 2008, 2015, 2016, Martens et al. 2008, Päckert et al. 2009, Mahood et al. 2013, Hung et al. 2014, Mckay et al. 2014, Dong et al. 2015). Apart from sequence data, also morphology, egg coloration, bioacoustics, distributional
Journal of Avian Biology 47: 899–910, 2016 doi: 10.1111/jav.01015
900
patterns and ecological niche profiles have been used in the aforementioned species delimitation studies as evidence of lineage separation.
The genus Calliope is endemic to the Palearctic, and was previously synonymized with Luscinia (Sibley and Monroe 1990, Dickinson 2004, Collar 2005, Gill and Donsker 2015) or Erithacus (Ripley 1964). However, it was recently suggested that the genus Luscinia sensu lato (s.l.) is non-monophyletic, and that it should be split into four genera: Calliope, Larvivora, Luscinia (sensu stricto, s.s.) and Tarsiger (Sangster et al. 2010, Clement and Rose 2015). The genus Calliope consists of four members, Siberian rubythroat Calliope calliope and its sister species white-tailed rubythroat C. pectoralis, and the sister species firethroat C. pectardens and blackthroat C. obscura (Alström et al. 2013b). The latter two species are monotypic and have restricted ranges in the mountains in central China (Collar 2005, Song et al. 2014, Clement and Rose 2015).
Calliope pectoralis has the second largest breeding range after C. calliope, and breeds in semi-open alpine or subalpine meadows with scattered bushes above the tree-line, at approx-imately 2600–4800 m a.s.l. (Collar 2005) (Fig. 1). Four sub-species are usually recognized: C. p. ballioni in southern Tian Shan mountains, Dzungarian Alatau, China and Kazakh-stan to southern Tajikistan (Pamir range) and northeastern Afghanistan; C. p. pectoralis in southeastern Afghanistan and northwestern Himalayas to central Nepal; C. p. confusa in eastern Nepal and Sikkim; and C. p. tschebaiewi in northern
Kashmir, on the eastern edge of the Qinghai-Tibetan Plateau (hereafter ‘QTP’), including northeastern India and extreme northern Myanmar to north-central China (Collar 2005, Clement and Rose 2015). The non-breeding ranges of the subspecies are poorly known, and all are believed to winter in the lowlands and foothills of their breeding ranges (Collar 2005, Rasmussen and Anderton 2005, Galewski et al. 2006, Clement and Rose 2015). The subspecies C. p. tschebaiewi is easily told from the others in adult male plumage by its white submoustachial stripe (Fig. 1), whereas the other subspecies are more subtly different from each other (Collar 2005, Rasmussen and Anderton 2005, Clement and Rose 2015).
The role of mountains as cradles of avian diversity has been widely recognized (Fjeldså et al. 2012). As a high ele-vation bird species, C. pectoralis inhabits highland habitats spanning several mountain ranges encompassing the core QTP (e.g. Tian Shan, the Himalayas and the mountains surrounding the Sichuan Basin in south central China, hereafter the Sino-Himalayan mountains). Because of the heterogeneity of alpine climatic conditions and vegetation in each of these mountain ranges (Favre et al. 2015), allo-patric or parapatric subspecies of C. pectoralis may have experienced different climatic and ecological conditions. Therefore intraspecific divergences in genetic and generic traits are expected due to processes such as local adaptation and genetic drift. In C. pectoralis, apparent plumage differ-ences and songs were already noted between C. p. tschebaiewi
Figure 1. Breeding distribution and sampling localities of the white-tailed rubythroat Calliope pectoralis complex. Red dotted lines show boundaries among subspecies. Triangles represent sampling localities of Siberian rubythroat Calliope calliope beicki and circles represent sampling localities of the Calliope pectoralis complex. Unfilled symbols indicate localities with DNA samples; filled ones indicate localities with sound recordings; and half filled circles indicate localities with DNA and sound recordings. The photo captions present the new taxonomy proposed in this paper. Photos by Menxiu Tong (Chinese rubythroat) and Andrew Lassey (Himalayan rubythroat).
901
and the other subspecies (Collar 2005). This indicated a potential subdivision among the subspecies of C. pectoralis, and its species limits merit further study using an integra-tive approach. Moreover, because the vocalizations of C. pectoralis and C. calliope are considered similar, and as C. p. tschebaiewi is somewhat intermediate between the other subspecies of C. pectoralis and C. calliope, it has been sug-gested that C. pectoralis and C. calliope may be better treated as a single species, although the possibility of three species has also been postulated (Rasmussen and Anderton 2005). The isolated breeding population of Siberian rubythroat in central China is sometimes treated as the subspecies C. c. beicki (Spiridonova et al. 2014). Its breeding range is sym-patric with C. p. tschebaiewi in Gansu, Qinghai and northern Sichuan (Collar 2005, Clement and Rose 2015) (Fig. 1).
In this study, we examined species delimitation of the C. pectoralis complex based on multiple lines of evidence: 1) mitochondrial DNA (mtDNA) and nuclear DNA (nucDNA); 2) song; and 3) plumage and morphometrics.
Methods
Sample collection and DNA extraction
For molecular genetic analysis, twenty-two DNA samples of C. pectoralis were included in this study, from China, Kazakhstan, Tajikistan and Sikkim (India), representing the subspecies C. p. ballioni, C. p. confusa and C. p. tschebaiewi (Fig. 1, Supplementary material Appendix 1, Table A1). Among them, we collected mostly feathers from live-trapped birds in a noninvasive manner, but also some muscle and blood samples. Three toepad samples were obtained from a museum specimen of C. p. confusa. Unfortunately, we did not obtain any samples representing the subspecies of C. p. pectoralis. In addition, we also collected samples of the closely related species C. calliope and C. pectardens, and homologous sequences of C. obscura and collared bush robin Tarsiger johnstoniae were downloaded from GenBank (Supplementary material Appendix 1, Table A1).
All tissue samples were stored at –20°C or in 95% etha-nol. Total genomic DNA was extracted using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol and quantified on a NanoDrop ND-1000 (Thermo Fisher Scientific, Waltham, MA, USA).
Gene amplification and sequencing
We amplified a fragment of the mitochondrial Cytochrome Oxidase C subunit 1 (COI) gene using the primer pairs BirdF1 and BirdR1 (Hebert et al. 2004), and mitochondrial cyto-chrome b (cytb) gene using the primer L14995 and H16065 (Groth 1998). Because the DNA from three museum samples of C. p. confusa was degraded and fragmented, we were only able to amplify and sequence the cytb gene from these using our own designed internal primers (Supplementary material Appendix 1, Table A2). We also amplified two introns of nuclear DNA (nucDNA), myoglobin intron 2 (myo) and ornithine decarboxylase (ODC). Primer pair myo2 and myo3F (Kimball et al. 2009) were used for myo, primer pair OD6 and OD8R, and primer pair ODC2-F and ODC2-R
for ODC (Allen et al. 2003). PCR amplifications were per-formed in a reaction volume of 25 ml, containing 15.3 ml ddH2O, 2.5 ml 10 buffer (containing 1.5 mM MgCl2), 2 ml dNTPs (2.5 mM), 1 ml MgCl2 (25 mM), 1 ml of each primer (10 pmol ml–1), and 0.2 ml (5 unit ml–1) Taq poly-merase (Qiagen) and 2 ml template DNA (20–100 ng ml–1). The PCR condition for each gene is provided as Supplemen-tary material Appendix 1, Table A2. The purified products were sequenced with both forward and reverse primers using BigDye Terminator ver. 3.1 Cycle Sequencing Kit (Applied Biosystems, Carlsbad, CA, USA) according to the guide-lines of the manufacturer. The products were sequenced on an ABI Prism 3730 automated DNA sequencers (service by Shanghai Majorbio Bio-pharm Technology).
Phylogenetic analyses
Sequences were aligned using the Clustal W algorithm (Thompson et al. 1994) in MEGA ver. 6.06 (Tamura et al. 2013), with default parameters, and the alignment was checked and adjusted manually if necessary. The protein-coding fragments were checked for the presence of stop codons and correctness of the reading frames to identify potential pseudogenes. All loci were analyzed sepa-rately, as well as concatenated. In the multi-locus analyses, the data were partitioned by locus, using rate multipliers to allow different rates for the different partitions (Ronquist and Huelsenbeck 2003, Nylander et al. 2004). We applied nucleotide substitution model selection using the Akaike information criterion, AIC (Akaike 1974) in jModelTest ver. 1.0 (Posada 2008). The Hasegawa–Kishino–Yano (HKY) model (Hasegawa et al. 1985) assuming a gamma-shaped (G) distribution across sites was selected for the COI, cytb and myo genes, and the general time reversible (GTR) model (Waddell and Steel 1997) assuming a gamma-shaped distribution across sites was chosen for ODC.
The mitochondrial gene tree was estimated by maxi-mum likelihood (ML) implemented in RaxML ver. 7.3.1 (Stamatakis 2006). Bootstrap with 1000 replicates was used to estimate nodal support in the ML analyses. Branches corresponding to partitions reproduced in 50% of the bootstrap replicates were collapsed. We also co-estimated mtDNA gene trees and divergence times using Bayes-ian inference (BI) implemented in BEAST ver. 1.7.3 (Drummond et al. 2012). We applied a cytb substitution rate of 2.1% per million yr (my) (0.0105 substitution/site/my) (Weir and Schluter 2008). All analyses were per-formed using HKY I G substitution model. Because the molecular clock hypothesis was rejected for mtDNA loci, we employed an uncorrelated lognormal relaxed and a birth–death tree prior for each mtDNA gene. We ran the BEAST analysis for 60 million generations with trees being sampled every 1000 generations. We generated trace plots and checked parameters using Tracer ver. 1.6 (Rambaut et al. 2014). Sufficient effective sample sizes (ESS 200) indicated convergence and mixing of chains. The first 10% of trees were discarded as burn-in.
We further estimated phylogenetic trees based on con-catenated nuclear loci using ML implemented in RaxML ver. 7.3.1 and BI by MrBayes ver. 3.2.1 (Ronquist and Huelsenbeck 2003). In the BI analyses, two independent
902
in Fig. 2. For each individual, five strophes were selected randomly, and the following measurements were taken for each individual: delta time (s) (the difference between begin time and end time for the selection); duration 90% (s) (the difference between the time 5 and time 95%; time 5% is the point in time that divides the selection into two time intervals containing 5 and 95% of the energy in the selec-tion, while the time 95% is the point in time that divides the selection into two time intervals containing 95 and 5% of the energy in the selection); low frequency (Hz) (the lower frequency limit of the selection); high frequency (Hz) (the upper frequency limit of the selection); delta frequency (Hz) (the difference between the upper and lower frequency limits of the selection); center frequency (Hz) (the frequency that divides the selection into two frequency intervals of equal energy); max frequency (Hz) (the frequency at which max power occurs within the selection); frequency 5% (Hz) and frequency 95% (Hz) (the frequency that divides the selection into two frequency intervals containing 5 and 95% of the energy in the selection); and bandwidth 90% (Hz) (the difference between the 5 and 95% frequencies). Moreover, the number of phrases/syllables/notes in each strophe was counted (Fig. 2). In total, we measured 13 song variables for each male and calculated their mean values and standard deviations (Table 2).
Before advanced statistical analysis, we performed the Kolmogorov–Smirnov analyses to test whether the vari-ables were normally distributed or not. We then separately ran Pearson or Spearman correlation analysis for normally distributed and non-normally distributed variables. After these pre-procedures, we removed seven variables that were significantly correlated with the six others (Supplementary material Appendix 1, Table A4). The remaining variables, ‘delta time’, ‘low frequency’, ‘high frequency’, ‘max fre-quency’, ‘bandwidth 90%’ and ‘number of syllables in a strophe’, were retained and further used to quantify acoustic differentiation using multivariate analyses including prin-cipal components analysis (PCA) and discriminant factor analysis (DFA). All statistical analyses were performed using SPSS ver. 18.0 for Windows.
Morphometric analyses
Because male plumage differences between C. p. tschebaiewi and the remaining subspecies of C. p. pectoralis are very obvious (Fig. 1), we only measured six morphometric characteristics including bill length (from skull to bill tip),
runs, each with four Markov Chain Monte Carlo (MCMC) simulations, were performed for 20 million iterations and sampled every 1000th step. The first 25% of the samples were discarded as burn-in. Convergence of the MCMC was assessed by checking the average standard deviation of split frequencies between two runs in MrBayes, and by examining the parameter values and ESS in Tracer ver. 1.6 (Rambaut et al. 2014).
To infer a multilocus species tree, we applied the mul-tispecies coalescent algorithm in *BEAST as implemented in BEAST ver. 1.7.3 (Drummond et al. 2012). For these analyses, only samples with sequence data for all four loci were used. The two linked mtDNA loci, myo and ODC, were treated as three separate partitions (mtDNA and two introns). Substitution models were assigned for each locus based on the results of jModelTest. Because the molecular clock hypothesis was not rejected for nucDNA genes, we then used a strict molecular clock model for each nucDNA partition and an uncorrelated lognormal relaxed model for mtDNA. We further employed a Yule speciation process as tree prior, and a piecewise linear and constant root popu-lation size model. We ran MCMCs for 90 million genera-tions, sampling trees every 1000 generations, and this was repeated three times. We discarded the first 10% of trees as burn-in. Convergence among the independent runs was determined by observing the ESS of the posterior distribu-tion and monitoring likelihoods and posterior probabilities of all resulting parameters in Tracer ver. 1.6 (Rambaut et al. 2014). The resulting tree was visualized using FigTree ver. 1.4.2 (Rambaut 2006).
We further estimated genetic divergences among subspe-cies of C. pectoralis and its congeners. Because only partial cytb of C. p. confusa was sequenced, this taxon was excluded. We calculated uncorrected p-distances and Kimura-2 param-eter genetic distances for cytb in MEGA, following the other recommendations on calculating genetic distance by Fregin et al. (2012). We also calculated average evolutionary diver-gence (D) over multiple sequence pairs within each taxon to represent interspecific genetic variation.
Sonogram analyses
We obtained song recordings of 37 singing males from a large part of the range of C. pectoralis (Supplementary mate-rial Appendix 1, Table A3 and Fig. 1). We also obtained four recordings of song of C. calliope beicki from central China, where its distribution range overlaps with C. p. tschebaiewi, and one recording of song of C. calliope from Kazakhstan, where it breeds close to C. p. ballioni, to examine the differ-ences among all the taxa. All song recordings were obtained during the breeding season and only full songs were included for further analyses. For C. p. confusa, the only two avail-able song recordings were taken during the winter season. These were visualized in sonograms but not used for statisti-cal analyses.
Before measurements were taken, all sound recordings were transformed into 48000 Hz, 24 bit in a .wav format. This process was performed in Sound Forge Audio Studio 9.0 (Sony Creative Software). Sonograms were created and analyzed in Raven Pro ver. 1.4 (Cornell Laboratory of Ornithology 2014). The sound terminology is explained Figure 2. Song terminology used in this study.
903
Sonogram analyses revealed song division within the Calliope pectoralis complex
The songs were basically similar in C. p. ballioni and C. p. pectoralis, whereas the song of C. p. tschebaiewi was more divergent (Fig. 4, Table 2). The strophes were on average considerably longer in C. p. ballioni and C. p. pectoralis than in C. p. tschebaiewi, with more variation and changes in a strophe (as in operatic coloratura). The song of C. p. tschebaiewi had more regular rhythm with more repetitions, usually including three or four similar phrases in succession. C. p. tschebaiewi also had relatively lower high-frequency and center-frequency. The song of C. calliope was more similar to the song of C. p. tschebaiewi, with closely similar strophe duration, but lower average center frequency (Table 2) and different timbre.
Principal component analysis (PCA) was performed with six principal components (PC) and the first three explained 78.6% of the total variation. PC1 mainly represented ‘delta time’ and ‘number of syllables in each strophe’; PC2 rep-resented ‘low frequency’; PC3 represented ‘bandwidth 90%’ and ‘max frequency’. Scatterplots were made with PC 1–3. In the plot PC1 vs PC2 (Fig. 5A) and PC1 vs PC3 (Fig. 5B), C. p. tschebaiewi, C. p. ballioni/C. p. pectoralis and C. calliope were separated with only marginal overlap, and C. p. tschebaiewi and C. calliope were only separable on PC2. In contrast C. p. ballioni and C. p. pectoralis were almost indistiguishable.
Based on the PCA results, we assigned three groups: 1) C. p. ballioni/pectoralis, 2) C. p. tschebaiewi and 3) C. calliope for discriminant function analyses (DFA), on the same six variables used in the PCA. The DFA resulted in 91.7% correct classification of the assigned groups. The group specific cor-rect rate was 88.1, 97 and 80% for C. p. ballioni/pectoralis, C. p. tschebaiewi and C. calliope, respectively, suggesting distinctive song of, especially, C. p. tschebaiewi. If we assigned C. p. ballioni and C. p. pectoralis as independent groups, the overall rate of correct classification was 84.4%. The correct rate of identification for C. p. tschebaiewi and C. calliope was not changed irrespective of groupings.
Morphometric analyses revealed slight differences
Calliope p. tschebaiewi differed only slightly in morphomet-rics from C. p. pectoralis and C. p. ballioni (Table 3). All measured variables were non-significant, except for the depth of the black breast-band, which was marginally broader in C. p. ballioni than in C. p. tschebaiewi (Mann–Whitney U test, z –1.93, p 0.046 [based on 9999 Monte Carlo simulations]; Table 3, Supplementary material Appendix 1, Fig. A3). Due to the limited sample size for both taxa, this result should be interpreted with caution.
Discussion
Species delimitation in the Calliope pectoralis complex
Plumages, songs and genetic analysis supported three main groups in the Calliope pectoralis complex, namely C. p. pectoralis,
wing length (flattened), tarsal length, tail length and depth of black breast band (from throat towards belly) in the specimen collection at The National Zoological Museum of China, Inst. of Zoology, Chinese Academy of Sciences, Beijing. All measurements were taken by YL, and Mann– Whitney U tests were performed to test whether any mor-phometric characteristics show statistical differentiation between C. p. tschebaiewi and the remaining subspecies of C. pectoralis (no samples of C. p. confusa available).
Results
Molecular phylogenetic analyses support the split of the polytypic Calliope pectoralis
Overall, 32 individuals of four Calliope species were studied. For each individual, we obtained a sequence of 693 bp of COI, 1018 bp of cytb, 690 bp of myo and 663 bp of ODC, giving up to 3064 bp of combined DNA sequence data. We did not find any heterozygous sites in the two nucDNA gene sequences. However, not all individuals were genotyped at four loci due to the low quality of DNA template or because sequences were not available in GenBank (Supplementary material Appendix 1, Table A1). From the three toepad sam-ples of C. p. confusa, we were only able to obtain 600-bp partial sequences of cytb.
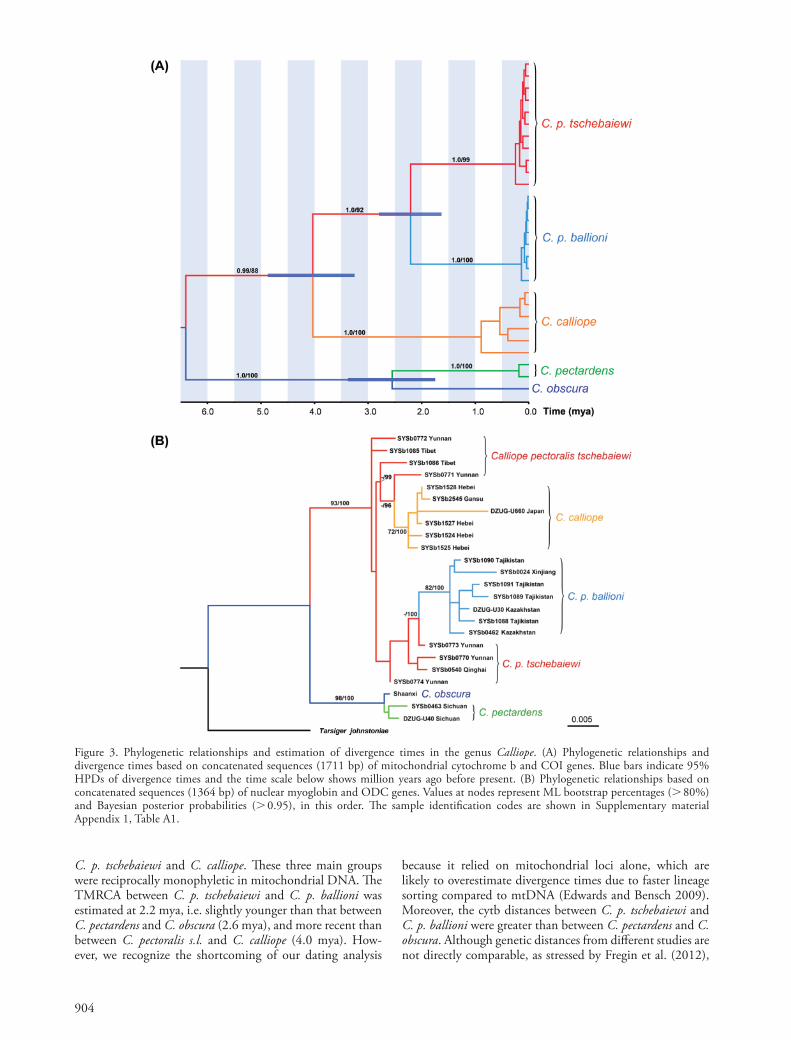
The ML and BI mtDNA gene trees resulted in virtually identical topologies, and all terminal clades had relatively high support values (Fig. 3A). In these trees, C. pectoralis ballioni (Tajikistan, Kazakhstan, and Xinjiang Uygur region, China) and C. pectoralis tschebaiewi (Sichuan, Tibet, Qinghai and Yunnan provinces, China) were reciprocally monophyl-etic. The gene trees based on partial cytb sequences suggested further subdivision between C. p. ballioni and C. p. confusa (Supplementary material Appendix 1, Fig. A1).
Due to a low level of polymorphism in the two nuclear loci, we carried out divergence analysis using mtDNA loci only. Our analysis revealed that the two taxa of C. pectoralis were sister to Calliope calliope, and these three taxa diverged around 4.0 mya (95% highest posterior density [HPD] 3.2–4.8 mya) from their most recent common ancestor (Fig. 3A). The time to most recent common ancestor (TMRCA) between C. p. tschebaiewi and C. p. ballioni was estimated to be 2.2 mya (95% HPD 1.6–2.8 mya). This time was close to the TMRCA between the two sister species C. pectardens and C. obscura (2.6 mya; 95% HPD 1.8–3.4 mya). Pair-wise genetic divergence based on uncorrected or Kimura 2-parameter cytb distances were 6.5% between C. p. tschebaiewi and C. p. ballioni, which is larger than those between C. pectardens and C. obscura (4.5–4.7%) (Table 1). In the nucDNA concatenated gene tree, C. p. ballioni and C. calliope were reciprocally monophyletic, with fairly high statistical support, although both these clades were nested within C. p. tschebaiewi (Fig. 3B).
The *BEAST multilocus species tree topology was con-cordant with the mtDNA gene tree (Supplementary material Appendix 1, Fig. A2) and further supported the split of C. p. tschebaiewi and C. p. ballioni, although it should be borne in mind that this analysis was heavily influenced by the mtDNA due to low levels of polymorphism in nucDNA.
904
because it relied on mitochondrial loci alone, which are likely to overestimate divergence times due to faster lineage sorting compared to mtDNA (Edwards and Bensch 2009). Moreover, the cytb distances between C. p. tschebaiewi and C. p. ballioni were greater than between C. pectardens and C. obscura. Although genetic distances from different studies are not directly comparable, as stressed by Fregin et al. (2012),
C. p. tschebaiewi and C. calliope. These three main groups were reciprocally monophyletic in mitochondrial DNA. The TMRCA between C. p. tschebaiewi and C. p. ballioni was estimated at 2.2 mya, i.e. slightly younger than that between C. pectardens and C. obscura (2.6 mya), and more recent than between C. pectoralis s.l. and C. calliope (4.0 mya). How-ever, we recognize the shortcoming of our dating analysis
Figure 3. Phylogenetic relationships and estimation of divergence times in the genus Calliope. (A) Phylogenetic relationships and divergence times based on concatenated sequences (1711 bp) of mitochondrial cytochrome b and COI genes. Blue bars indicate 95% HPDs of divergence times and the time scale below shows million years ago before present. (B) Phylogenetic relationships based on concatenated sequences (1364 bp) of nuclear myoglobin and ODC genes. Values at nodes represent ML bootstrap percentages ( 80%) and Bayesian posterior probabilities ( 0.95), in this order. The sample identification codes are shown in Supplementary material Appendix 1, Table A1.
905
C. calliope and C. p. ballioni, but not in C. p. tschebaiewi (Fig. 3B). This very likely resulted from a larger and sta-ble population of C. p. tschebaiewi within stable montane habitats, given its larger extant geographic range compared to the other subspecies. Apart from demographic dispari-ties, alternative scenarios including adaptive introgression and sex-biased gene flow are often invoked to explain mito-nuclear discordances (Toews and Brelsford 2012). It seems more promising to test these alternative hypotheses using a coalescent model with a better coverage of population sampling and a reasonable number of independent nuclear loci (Papakostas et al. 2016).
the pairwise comparisons among the Calliope taxa are gener-ally considerably higher than normal intraspecific variation (Aliabadian et al. 2009).
In contrast to mtDNA genealogical patterns, our nuclear gene trees show paraphyly and lower genetic divergences among the members of the C. pectoralis complex. This is expected due to lower substitution rate and larger effec-tive population size of nuclear genes compared to mtDNA, and can be interpreted as recent divergence and retention of ancestral polymorphisms, as shown in previous studies of other birds (reviewed by Zink and Barrowclough 2008). However, lineage sorting of nucDNA was completed in
Table 1. Pairwise genetic distances (uncorrected p-distance at lower diagonal and Kimura 2-parameter distance at upper diagnoal) based on 1018 bp of cytochrome b between, and average evolutionary divergence (D) over sequence pairs within each taxonomic groups of Calliope taxa.
Figure 4. Sonograms of songs in the Calliope pectoralis complex and C. calliope. Calliope p. ballioni: b1) Kalon, Sughd Region, Tajikistan, 20 July 2014 (C. Jia); b2) Almaty, Kazakhstan, 14 May 2011(A. Wassink); b3) Almaty, Kazakhstan, 11 May 2009 (G. Carey); b4) Xinyuan, Xinjiang, China, 16 June 2014 (C. Jia); b5) Wusu, Xinjiang, China, 31 May 2014 (C. Jia); b6) Bole, Xinjiang, China, 19 June 2014 (C. Jia); C. p. pectoralis: p1) and p2) Gilgit, Pakistan, 3 June 1998 (P. Alström); C. p. confusa: c1) Assam, India, 23 November 2012 (M. Nel-son); c2) Assam, India, 3 January 1998 (D. Allen); C. p. tschebaiewi : t1) Deqin, Yunan, China, 9 June 2008 (P. Alström); t2), t3) and t4) Kangding, Sichuan, China, from 30–31 May 2013 (P. Alström); t5) Balangshan, Sichuan, China, 2 June 2013 (P. Alström); t6) Balangshan, Sichuan, China, 8 June 2012 (G. Carey); t7) Balangshan, Sichuan, China, 1 June 2012 (F. Lambert); t8) Yajiang, Sichuan, China, 7 May 2012 (C. Jia); t9) Heimahe, Qinghai, China, 1 June 1987 (P. Alström); t10) Yushu, Qinghai, China, 10 June 2005 (F. Lambert); t11) Zhada, Xizang, China, 7 June 2014 (C. Jia); t12) Linzhi, Xizang, China, 28 May 2014 (C. Jia); C. calliope: ca1) Huzhu, Qinghai, China, 18 June 2013 (T. Carlberg); ca2) Jiuzhaigou, Sichuan, China, 4 June 2013 (F. Lambert).
906
It has been argued that the songs of C. pectoralis and C. calliope are very similar, and that the song of C. p. tschebaiewi is somewhat intermediate between the other subspecies of C. pectoralis and C. calliope (Collar 2005, Rasmussen and Anderton 2005, Clement and Rose 2015). Our results show that the songs are indeed closely similar in all taxa, although C. calliope, C. p. pectoralis/C. p. ballioni and C. p. tschebaiewi are separable both audibly and in the PCA/DFA.
The plumage differences between C. calliope and the C. pectoralis complex are striking in adult male, and diagnostic also in adult female plumage (Collar 2005, Rasmussen and Anderton 2005, Clement and Rose 2015). In adult male plumage, C. p. tschebaiewi is diagnosably different from C. p. ballioni and C. p. pectoralis by qualitative characters, whereas females are only subtly different (Collar 2005, Rasmussen and Anderton 2005, Clement and Rose 2015). In contrast, C. p. pectoralis, C. p. ballioni and C. p. confusa are much more similar to each other, and differ mainly in color hues of the upperparts in adult male plumage (Collar 2005, Rasmussen and Anderton 2005, Clement and Rose 2015).
The breeding distributions of C. calliope and C. p. tschebaiewi are marginally overlapping, with no evidence of interbreeding. Field observations at one sympatric area, Gonghe, Qinghai, indicate that C. calliope occurs in decidu-ous and mixed forests at low to mid-elevation, whereas C. p. tschebaiewi breeds in alpine and subalpine bushes (M. Ma pers. comm.). The subspecies of the C. pectoralis complex are allo-/parapatrically distributed in the breeding season, with-out any known contact, although C. p. tschebaiewi is at least geographically very close to C. p. pectoralis and C. p. confusa (Rasmussen and Anderton 2005, Clement and Rose 2015); cf. Fig. 1).
Due to the congruent differences in adult male plum-age, song and mitochondrial DNA and a substantial time of separation between C. p. tschebaiewi and C. p. ballioni, we suggest that these should be treated as separate species. Although C. p. pectoralis, C. p. ballioni and C. p. confusa dif-fer somewhat, especially in adult male plumage, the differ-ences are comparatively slight and concern colour hues only. Our acoustic analyses also suggest almost no difference in song types between C. p. pectoralis and C. p. ballioni (Fig. 5), but the sample size for C. p. pectoralis was small. These three subspecies are parapatric along the Tian Shan–Himalayan mountain chains, and whether they show disjunct or clinal variation in morphology, songs and genetics is not resolved in this study.
Calliope calliope should continue to be recognized as a distinct species. Although significant genetic divergence at a mitochondrial level, indicating genetic clusters between western and eastern Siberian C. calliope has been revealed (Spiridonova et al. 2014), we found no evidence of differ-entiation between central Chinese and Siberian/Japanese C. calliope, and this question needs further study.
Speciation in the Calliope pectoralis complex
Calliope pectoralis sensu lato has a geographical range that extends from the Tian Shan mountains in central Asia, through the Himalayas to the mountainous areas in central China (Collar 2005, Clement and Rose 2015). The lat-ter area is also called the Sino-Himalayan region, which is Ta
ble
2. T
he m
ean
valu
e an
d st
anda
rd d
evia
tion
of th
e m
easu
rem
ents
of s
elec
ted
stro
phes
of s
ong
in e
ach
taxo
nom
ic g
roup
of C
allio
pe ta
xa.
Tax
onN
o. o
f in
divi
dual
sN
o. o
f st
roph
esV
alue
Del
ta
time
(s)
Dur
atio
n 90
% (s
)
Low
fr
eque
ncy
(Hz)
Hig
h fr
eque
ncy
(Hz)
Del
ta
freq
uenc
y (H
z)
Cen
ter
freq
uenc
y (H
z)
Max
fr
eque
ncy
(Hz)
Freq
uenc
y 5%
(Hz)
Freq
uenc
y 95
% (H
z)B
andw
idth
90
% (H
z)
No.
of
phra
ses
in e
ach
stro
phe
No.
of
sylla
bles
in
eac
h st
roph
e
No.
of
note
s in
ea
ch
stro
phe
C. p
. bal
lioni
1252
Mea
n4.
033.
0613
42.8
680
94.3
267
51.4
544
71.1
543
66.5
931
80.2
958
99.0
428
70.1
918
.226
.942
.4SD
1.18
0.96
325.
7447
8.75
582.
6326
0.93
500
411.
7371
7.55
717.
636.
36.
310
.9C
. p. p
ecto
ralis
315
Mea
n4.
964.
0513
09.4
982
30.1
669
20.6
744
23.0
543
25.0
832
63.9
857
30.3
924
66.4
128
.935
.456
.9SD
1.85
1.71
183.
8138
7.75
451.
0520
7.62
290.
0115
5.24
196.
1129
1.35
7.9
9.3
12.4
C. p
. tsc
heba
iew
i20
100
Mea
n1.
611.
1915
34.2
7458
.77
5924
.57
4126
.88
4385
.63
2821
.88
5765
.63
2943
.75
8.4
13.0
21.6
SD0.
390.
3526
2.11
396.
7850
7.09
423.
4773
1.74
200.
151
1.5
563.
761.
92.
76.
1C
. cal
liope
525
Mea
n2.
491.
9689
6.17
8412
.05
7515
.88
3937
.541
2527
0057
9030
9010
.815
.025
.5SD
1.75
1.71
237.
4576
0.66
818.
151
3.49
924.
9235
4.93
581.
2562
6.84
4.0
6.2
9.8
907
feasible that the divergence between C. p. tschebaiewi and the remaining taxa of C. pectoralis was associated with allopat-ric divergence due to long-term geographical fragmentation between Himalayan and south-central Chinese mountain chains. In the east, populations of C. p. tschebaiewi may have been maintained in a single and stable refuge along the eastern margin of the QTP, which has been suggested for other bird species (Qu et al. 2011, Gu et al. 2012). In the west, the lineage diversification of three Tian Shan–Himalayan subspecies of C. pectoralis might be a consequence of in situ diversification within the Himalayas (Päckert et al. 2012), or range expansion to the Western Himalayas and Tian Shan mountains from a source population in a single refuge in the eastern Himalayas (Srinivasan et al. 2014). More data and analysis are needed to evaluate this.
Taxonomic implications
We propose that C. pectoralis s.l. should be treated as two species: C. pectoralis, with subspecies C. p. pectoralis, C. p. ballioni and C. p. confusa; and C. tschebaiewi monotypic. We suggest the use of Himalayan rubythroat for C. pectoralis (already adopted in Rasmussen and Anderton 2005) and Chinese rubythroat for C. tschebaiewi. The English name of the latter species reflects its geographical range.
Some passerine birds in the region that have been upgraded from subspecies to full species based on previ-ous integrative studies, e.g. Horornis and Locustella warblers
a hotspot for bird diversity (Stattersfield et al. 1998). The biogeographical patterns by phylogeography of several bird species from this area have been studied in recent years (Qu and Lei 2009, Martens et al. 2011, Qu et al. 2011, Gu et al. 2012, Lei et al. 2014, Liu et al. 2016). It has been suggested that, in general, immigration of pre-adapted lineages fol-lowed by geographical separation and subsequent speciation has been an important driver of bird diversification in this region (Päckert et al. 2012, Lei et al. 2014, Price et al. 2014). For example, a comprehensive analysis of the complete song-bird community of the east Himalayas suggested that the richness of passerines in the Himalayas was largely caused by immigration followed by allopatric divergence, especially from central China and southeastern Asia (Price et al. 2014). According to our analyses, the Calliope pectoralis complex diverged from its more northerly distributed closest relative, C. calliope, around the Early Pliocene (3.2–4.8 mya), which might correspond with the early uplift of QTP (ca 3.6 mya, Lei et al. 2014).
The lineage divergence within the Calliope pectoralis com-plex was estimated at around 2.2 mya (95% HPD 1.6–2.8 mya), coinciding with recent violent uplifts of the QTP (ca 3.4–1.7 mya, Li et al. 1996) and associated climatic change (Qu et al. 2011, Wang et al. 2014). Because the Calliope pectoralis complex breeds in subalpine vegetated habitats, the glacial cycles since the early Pleistocene in the QTP (ca 1.17 mya, Zheng et al. 2002) may also have affected lineage diver-sification as in other subalpine species (Luo et al. 2016). It is
Table 3. Morphological comparisons between Calliope pectoralis ballioni and C. p. tschebaiewi. Measurements included are wing length (WL), tail length (TL), bill length (BL), tarsal length (TarL) and breast-band depth (BBD). Average values and associated standard deviation are given in millimeters (mm).
Taxon (sample size) WL TL BL TarL BBD
C. p. ballioni (3) 73.08 2.68 62.50 2.75 13.05 0.38 30.41 1.10 15.87 2.22C. p. tschebaiewi (6) 70.17 3.27 60.33 2.64 12.50 0.57 30.13 0.52 23.33 4.98
Figure 5. Scatterplot based on the results of principal component analyses of song divergence among four studied taxa in the genus Calliope.
908
mandelli complex reveals a new species from central China. –Avian Res. 6: 1–33.
Alström, P., Rasmussen, P. C., Zhao, C., Xu, J., Dalvi, S., Cai, T., Guan, Y., Zhang, R., Kalyakin, M. V., Lei, F. M. and Olsson, U. 2016. Integrative taxonomy of the plain-backed thrush (Zoothera mollissima) complex (Aves, Turdidae) reveals cryptic species, including a new species. – Avian Res. 7: 1.
Clement, P. and Rose, C. 2015. Robins and chats: Helm identifica-tion guide series. – Christopher Helm.
Collar, N. 2005. Family Turdidae (Thrushes). – In: Del Hoyo, J., Elliott, A. and Christie, D. (eds), Handbook of the birds of the World, vol. 10. Cuckoo-shrikes to thrushes. Lynx Edicions, pp. 514–807.
Cornell Laboratory of Ornithology 2014. Raven Pro: interactive sound analysis software. – Cornell Lab of Ornithology, Ithaca, NY.
de Queiroz, K. 2007. Species concepts and species delimitation. – Syst. Biol. 56: 879–886.
Dickinson, E. C. (ed.) 2004. The Howard and Moore complete checklist of birds of the World, 3nd ed. – Christopher Helm.
Dong, L., Wei, M., Alström, P., Huang, X., Olsson, U., Shigeta, Y., Zhang, Y. and Zheng, G. 2015. Taxonomy of the narcissus flycatcher Ficedula narcissina complex: an integrative approach using morphological, bioacoustic and multilocus DNA data. – Ibis 157: 312–325.
Drovetski, S. V., Semenov, G., Drovetskaya, S. S., Fadeev, I. V., Red’kin, Y. A. and Voelker, G. 2013. Geographic mode of speciation in a mountain specialist Avian family endemic to the Palearctic. – Ecol. Evol. 3: 1518–1528.
Drummond, A. J., Suchard, M., Xie, D. and Rambaut, A. 2012. Bayesian phylogenetics with BEAUti and the BEAST 1.7. – Mol. Biol. Evol. 29: 1969–1973.
Edwards, S. V. and Bensch, S. 2009. Looking forwards or looking backwards in avian phylogeography? A comment on Zink and Barrowclough 2008. – Mol. Ecol. 18: 2930–2933.
Favre, A., Päckert, M., Pauls, S. U., Jähnig, S. C., Uhl, D., Michalak, I. and Muellner-Riehl, A. N. 2015. The role of the uplift of the Qinghai-Tibetan Plateau for the evolution of Tibetan biotas. – Biol. Rev. 90: 236–253.
Fjeldså, J., Bowie, R. C. and Rahbek, C. 2012. The role of mountain ranges in the diversification of birds. – Annu. Rev. Ecol. Evol. Syst. 43: 249–265.
Fregin, S., Haase, M., Olsson, U. and Alström, P. 2012. Pitfalls in comparisons of genetic distances: a case study of the avian family Acrocephalidae. – Mol. Phylogenet. Evol. 62: 319–328.
Fujita, M. K., Leaché, A. D., Burbrink, F. T., Mcguire, J. A. and Moritz, C. 2012. Coalescent-based species delimitation in an integrative taxonomy. – Trends Ecol. Evol. 27: 480–488.
Galewski, T., Tilak, M.-K., Sanchez, S., Chevret, P., Paradis, E. and Douzery, E. 2006. The evolutionary radiation of Arvicolinae rodents (voles and lemmings): relative contribution of nuclear and mitochondrial DNA phylogenies. – BMC Evol. Biol. 6: 80.
Gelang, M., Cibois, A., Pasquet, E., Olsson, U., Alström P. and Ericson, P. G. P. 2009. Phylogeny of babblers (Aves, Passeri-formes): major lineages, family limits and classification. – Zool. Scripta 35: 225–236.
Gill, F. and Donsker, D. 2015. IOC World bird list. – Version 5.4.
Groth, J. G. 1998. Molecular phylogenetics of finches and sparrows: consequences of character state removal in cytochrome b sequences. – Mol. Phylogenet. Evol. 10: 377–390.
Gu, L. Y., Liu, Y., Que, P. J. and Zhang, Z. W. 2012. Quaternary climate and environmental changes have shaped genetic differentiation in a Chinese pheasant endemic to the eastern margin of the Qinghai-Tibetan Plateau. – Mol. Phylogenet. Evol. 67: 129–139.
(Alström et al. 2007, 2008, 2015) and treecreepers (Tietze et al. 2006). Several other candidates for species status exist, and taxonomic studies of these are necessary. Taken together, this study highlights the important applications of integra-tive taxonomy in understanding avian species limits and diversity in the Sino-Himalayan mountains.
Acknowledgements – We are grateful to Jun Gou and Pengcheng Wang for DNA samples; Arend Wassink, Desmond Allen, Ding Li Yong, Frank Lambert, Jan Hein van Steenis, Manuel Schweizer, Mike Nelson, Raffael Ayé, Ruud van Beusekom, Thijs Fijen and Tomas Carlberg for sound recordings, Peng He for his help to access specimen in IOZ, Chinese Academy of Sciences (CAS). This research is supported by the National Natural Science Foundation of China (no. 31301875 and 31572251 to YL, and 31272300 to CXJ), the CAS Visiting Professorship for Senior International Sci-entists (no. 2011T2S04) to PA, and Jornvall Foundation and The Sound Approach to PA and UO, and the Ministry of Science and Technology of the People’s Republic of China (no. 2015DFR30790) to CXJ. We also thank Menxiu Tong, Andrew Lassey and Junhua Hu for the preparation of Fig. 1.
References
Akaike, H. 1974. A new look at the statistical model identification. – IEEE Trans. Auto. Control 19: 716–723.
Aliabadian, M., Kaboli, M., Nijman, V. and Vences, M. 2009. Molecular identification of birds: performance of distance-based DNA barcoding in three genes to delimit parapatric species. – PLoS One 4: e4119.
Allen, S. E., Omland, K. E. and Prum, R. 2003. Novel intron phylogeny supports plumage convergence in orioles (Icterus). – Auk 120: 961–969.
Alström, P., Olsson, U., Rasmussen, P. C., Yao, C. T., Ericson, P. G. and Sundberg, P. 2007. Morphological, vocal and genetic divergence in the Cettia acanthizoides complex (Aves: Cettiidae). – Zool. J. Linn. Soc. 149: 437–452.
Alström, P., Rasmussen, P. C., Olsson, U. and Sundberg, P. 2008. Species delimitation based on multiple criteria: the spotted bush warbler Bradypterus thoracicus complex (Aves: Megaluridae). – Zool. J. Linn. Soc. 154: 291–307.
Alström, P., Fregin, S., Norman, J. A., Ericson, P. G., Christidis, L. and Olsson, U. 2011a. Multilocus analysis of a taxonomi-cally densely sampled dataset reveal extensive non-monophyly in the avian family Locustellidae. – Mol. Phylogenet. Evol. 58: 513–526.
Alström, P., Höhna, S., Gelang, M., Ericson, P. G. and Olsson, U. 2011b. Non-monophyly and intricate morphological evolu-tion within the avian family Cettiidae revealed by multilocus analysis of a taxonomically densely sampled dataset. – BMC Evol. Biol. 11: 352.
Alström, P., Olsson, U. and Lei, F. 2013a. A review of the recent advances in the systematics of the avian superfamily Sylvioidea. – Chinese Birds 4: 99–131.
Alström, P., Song, G., Zhang, R., Gao, X., Holt, P. I., Olsson, U. and Lei, F. 2013b. Taxonomic status of blackthroat Calliope obscura and firethroat C. pectardens. – Forktail 29: 94–99.
Alström, P., Hooper, D. M., Liu, Y., Olsson, U., Mohan, D., Gelang, M., Le Manh, H., Zhao, J., Lei, F. and Price, T. D. 2014. Discovery of a relict lineage and monotypic family of passerine birds. – Biol. Lett. 10: 20131067.
Alström, P., Xia, C., Rasmussen, P. C., Olsson, U., Dai, B., Zhao, J., Leader, P. J., Carey, G. J., Dong, L., Cai, T., Holt, P., Le Manh, H., Song, G., Liu, Y., Zhang, Y. Y. and Lei, F. M. 2015. Integrative taxonomy of the russet bush warbler Locustella
909
Päckert, M., Blume, C., Sun, Y. H., Liang, W. and Martens, J. 2009. Acoustic differentiation reflects mitochondrial lineages in Blyth’s leaf warbler and white-tailed leaf warbler complexes (Aves: Phylloscopus reguloides, Phylloscopus davisoni). – Biol. J. Linn. Soc. 96: 584–600.
Päckert, M., Martens, J. and Sun, Y. H. 2010. Phylogeny of long-tailed tits and allies inferred from mitochondrial and nuclear markers (Aves: Passeriformes, Aegithalidae). – Mol. Phylogenet. Evol. 55: 952–967.
Päckert, M., Martens, J., Sun, Y. H., Severinghaus, L. L., Nazarenko, A. A., Ting, J., Töpfer, T. and Tietze, D. T. 2012. Horizontal and elevational phylogeographic patterns of Himalayan and southeast Asian forest passerines (Aves: Passeriformes). – J. Biogeogr. 39: 556–573.
Padial, J. M., Miralles, A., De La Riva, I. and Vences, M. 2010. Review. The integrative future of taxonomy. – Front. Zool. 7: 16.
Papakostas, S., Michaloudi, E., Proios, K., Brehm, M., Verhage, L., Rota, J. and Declerck, S. A. 2016. Integrative taxonomy recognizes evolutionary units despite widespread mitonuclear discordance: evidence from a rotifer cryptic species complex. – Syst. Biol. doi: 10.1093/sysbio/syw016
Posada, D. 2008. jModelTest: phylogenetic model averaging. – Mol. Biol. Evol. 25: 1253–1258.
Price, T. D., Hooper, D. M., Buchanan, C. D., Johansson, U. S., Tietze, D. T., Alström, P., Olsson, U., Ghosh-Harihar, M., Ishtiaq, F. and Gupta, S. K. 2014. Niche filling slows the diversification of Himalayan songbirds. – Nature 509: 222–225.
Qu, Y. and Lei, F. 2009. Comparative phylogeography of two endemic birds of the Tibetan plateau, the white-rumped snow finch (Onychostruthus taczanowskii) and the Hume’s ground tit (Pseudopodoces humilis). – Mol. Phylogenet. Evol. 51: 312–326.
Qu, Y., Luo, X., Zhang, R., Song, G., Zou, F. and Lei, F. 2011. Lineage diversification and historical demography of a montane bird Garrulax elliotii – implications for the Pleis-tocene evolutionary history of the eastern Himalayas. – BMC Evol. Biol. 11: 174.
Rambaut, A. 2006. FigTree. – Version 1.42, < http://tree.bio.ed.ac.uk/software/figtree/ >.
Rambaut, A., Suchard, M., Xie, D. and Drummond, A. 2014. Tracer v1.6. – Computer program and documentation distrib-uted by the author, < http://beast.bio.ed.ac.uk/Tracer >.
Rasmussen, P. C. and Anderton, J. C. 2005. Birds of south Asia: the Ripley guide. – Lynx Edision.
Ripley, S. 1964. Family Muscicapidae, subfamily Turdinae. – In: Mayr, E. and Paynter, R. A. E. (eds), Check-list of birds of the World. Museum of Comparative Zoology, Harvard Univ. Press, pp. 13–227.
Ronquist, F. and Huelsenbeck, J. P. 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models. – Bioinformatics 19: 1572–1574.
Sangster, G., Alström, P., Forsmark, E. and Olsson, U. 2010. Multi-locus phylogenetic analysis of Old World chats and flycatchers reveals extensive paraphyly at family, subfamily and genus level (Aves: Muscicapidae). – Mol. Phylogenet. Evol. 57: 380–392.
Sibley, C. G. and Monroe, B. L. 1990. Distribution and taxonomy of birds of the World. – Yale Univ. Press.
Song, G., Alström, P., Zhang, Y., Gao, X., Gong, H., Holt, P. I., Qing, Q., Yin, Z. and Lei, F. 2014. Rediscovery of an enigmatic Chinese passerine, the blackthroat Calliope obscura: plumage, vocalizations, distribution, habitat choice, nesting and conservation. – J. Ornithol. 155: 347–356.
Spiridonova, L., Valchuk, O., Red’kin, Y. A. and Saitoh, T. 2014. Phylogeography of mtDNA haplotypes and range settling in the Siberian rubythroat. – Russian J. Genet. 49: 735–742.
Srinivasan, U., Tamma, K. and Ramakrishnan, U. 2014. Past climate and species ecology drive nested species richness
Hasegawa, M., Kishino, H. and Yahno, T. 1985. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. – J. Mol. Evol. 21: 160–174.
Hebert, P. D. N., Penton, E. H., Burns, J. M., Janzen, D. H. and Hallwachs, W. 2004. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. – Proc. Natl Acad. Sci. USA 101: 14812–14817.
Helbig, A. J., Knox, A. G., Parkin, D. T., Sangster, G. and Collinson, M. 2002. Guidelines for assigning species rank. – Ibis 144: 518–525.
Hung, C.-M., Hung, H.-Y., Yeh, C.-F., Fu, Y.-Q., Chen, D., Lei, F., Yao, C.-T., Yao, C.-J., Yang, X.-J., Lai, Y.-T. and Li, S.-H. 2014. Species delimitation in the Chinese bamboo partridge Bambusicola thoracica (Phasianidae; Aves). – Zool. Scripta 43: 562–575.
Kimball, R. T., Braun, E. L., Barker, F. K., Bowie, R. C., Braun, M. J., Chojnowski, J. L., Hackett, S. J., Han, K.-L., Harshman, J. and Heimer-Torres, V. 2009. A well-tested set of primers to amplify regions spread across the avian genome. – Mol. Phylogenet. Evol. 50: 654–660.
Lei, F., Qu, Y. and Song, G. 2014. Species diversification and phylogeographical patterns of birds in response to the uplift of the Qinghai-Tibet Plateau and Quaternary glaciations. – Curr. Zool. 60: 149–161.
Li, J. J., Fang, X. M., Ma, H. Z., Zhu, J. J., Pan, B. T. and Chen, H. L. 1996. Geomorphological and environmental evolution in the upper reaches of the Yellow River during the late Cenozoic. – Sci. China (Ser. D) 39: 380–390.
Liu, Y., Hu, J. H., Li, S. H., Dunchen, P., Wegmann, D. and Schweizer, M. 2016. Sino-Himalayan mountains act as cradles of diversity and immigration centres in the diversification of parrotbills (Paradoxornithidae). – J. Biogeogr. doi: 10.1111/jbi.12738
Luo, D., Yue, J. P., Sun, W. G., Xu, B., Li, Z. M., Comes, H. P. and Sun, H. 2016. Evolutionary history of the subnival flora of the Himalaya–Hengduan Mountains: first insights from comparative phylogeography of four perennial herbs. – J. Biogeogr. 43: 31–43.
Mahood, S. P., John, A. J. I., Eames, J. C., Oliveros, C. H., Moyle, R. G., Hong, C., Poole, C. M., Nielsen, H. and Sheldon, F. H. 2013. A new species of lowland tailorbird (Passeriformes: Cisticolidae: Orthotomus) from the Mekong floodplain of Cambodia. – Forktail 29: 1–14.
Martens, J., Sun, Y. H. and Päckert, M. 2008. Intraspecific differentiation of Sino-Himalayan bush-dwelling Phylloscopus leaf warblers, with description of two new taxa. – Vertebr. Zool. 58: 233–265.
Martens, J., Tietze, D. T. and Päckert, M. 2011. Phylogeny, biodiver-sity, and species limits of passerine birds in the Sino-Himalayan region – a critical review. – Ornithol. Monogr. 70: 64–94.
Mckay, B. D., Mays, H. L., Wan, D., Higuchi, H. and Nishiumi, I. 2014. Incorporating color into integrative taxonomy: analy-sis of the varied tit (Sittiparus varius) complex in east Asia. – Syst. Biol. 63: 505–517.
Moyle, R. G., Andersen, M. J., Oliveros, C. H., Steinheimer, F. and Reddy, S. 2012. Phylogeny and biogeography of the core babblers (Aves: Timaliidae). – Syst. Biol. 61: 631–651.
Nylander, J. A., Ronquist, F., Huelsenbeck, J. P. and Nieves-Aldrey, J. 2004. Bayesian phylogenetic analysis of combined data. – Syst. Biol. 53: 47–67.
Nylander, J. A., Olsson, U., Alström, P. and Sanmartín, I. 2008. Accounting for phylogenetic uncertainty in biogeography: a Bayesian approach to dispersal-vicariance analysis of the thrushes (Aves: Turdus). – Syst. Biol. 57: 257–268.
Olsson, U., Alström, P. and Sundberg, P. 2004. Non-monophyly of the avian genus Seicercus (Aves: Sylviidae) revealed by mitochondrial DNA. – Zool. Scripta 33: 501–510.
910
Waddell, P. J. and Steel, M. A. 1997. General time-reversible dis-tances with unequal rates across sites: mixing and inverse Gaussian distributions with invariant sites. – Mol. Phylogenet. Evol. 8: 398–414.
Wang, W., Dai, C., Alström, P., Zhang, C., Qu, Y., Li, S.-H., Yang, X., Zhao, N., Song, G. and Lei, F. 2014. Past hybridization between two east Asian long-tailed tits (Aegithalos bonvaloti and A. fuliginosus). – Front. Zool. 11: 40.
Weir, J. T. and Schluter, D. 2008. Calibrating the avian molecular clock. – Mol. Ecol. 17: 2321–2328.
Yang, Z. and Rannala, B. 2010. Bayesian species delimitation using multilocus sequence data. – Proc. Natl Acad. Sci. USA 107: 9264–9269.
Yeung, C. K., Lin, R.-C. Lei, F., Robson, C., Liang, W., Zhou, F., Han, L., Li, S.-H. and Yang, X. 2011. Beyond a morphologi-cal paradox: complicated phylogenetic relationships of the parrotbills (Paradoxornithidae, Aves). – Mol. Phylogenet. Evol. 61: 192–202.
Zheng, B. X., Xu, Q. Q. and Shen, Y. P. 2002. The relationship between climate change and Quaternary glacial cycles on the Qinghai-Tibetan Plateau: review and speculation. – Quat. Int. 97–98: 93–101.
Zink, R. M. and Barrowclough, G. F. 2008. Mitochondrial DNA under siege in avian phylogeography. – Mol. Ecol. 17: 2107–2121.
Zuccon, D., Prŷs-Jones, R., Rasmussen, P. C. and Ericson, P. G. 2012. The phylogenetic relationships and generic limits of finches (Fringillidae). – Mol. Phylogenet. Evol. 62: 581–596.
patterns along an east-west axis in the Himalaya. – Global Ecol. Biogeogr. 23: 52–60.
Stamatakis, A. 2006. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. – Bioinformatics 22: 2688–2990.
Stattersfield, A. J., Crosby, M. J., Long, A. J. and Wege, D. C. 1998. Endemic bird areas of the World: priorities for biodiversity conservation. – Birdlife Conservation Series, BirdLife International, Cambridge, UK.
Tamura, K., Stecher, G., Peterson, D., Filipski, A. and Kumar, S. 2013. MEGA6: molecular evolutionary genetics analysis version 6.0. – Mol. Biol. Evol. 30: 2725–2729.
Thompson, J., Higgins, D. and Gibson, T. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. – Nucleic Acids Res. 22: 4673–4680.
Tietze, D. T., Martens, J. and Sun, Y. H. 2006. Molecular phylog-eny of treecreepers (Certhia) detects hidden diversity. – Ibis 148: 477–488.
Tietze, D. T., Päckert, M., Martens, J., Lehmann, H. and Sun, Y. H. 2013. Complete phylogeny and historical biogeography of true rosefinches (Aves: Carpodacus). – Zool. J. Linn. Soc. 169: 215–234.
Tobias, J. A., Seddon, N., Spottiswoode, C. N., Pilgrim, J. D., Fishpool, L. D. C. and Collar, N. J. 2010. Quantitative criteria for species delimitation. – Ibis 152: 724–746.
Toews, D. P. and Brelsford, A. 2012. The biogeography of mitochondrial and nuclear discordance in animals. – Mol. Ecol. 21: 3907–3930.
Supplementary material (Appendix JAV-01015 at < www.avianbiology.org/appendix/jav-01015 >). Appendix 1.