Page 1
SPECIFIC CLEAVAGE OF HEPATITIS C VIRUS RNA GENOME
BY HUMAN RNase P
Anna Nadal*, María Martell*, J.Robin Lytle†, Alita J.Lyons†, Hugh D. Robertson†, Beatriz
Cabot*, Juan I. Esteban*, Rafael Esteban*, Jaime Guardia* & Jordi Gómez*
*Servicio de Medicina Interna-Hepatología. Area de Investigación Básica, Hospital Valle de
Hebrón, 08035 Barcelona, Spain.
†Department of Biochemistry, Weill Medical College of Cornell University, New York NY
10021, U.S.A.
Author to whom correspondence should be sent:
Dr.Jordi Gómez
Laboratorio de Medicina Interna-Hepatología.
Area de Investigación Básica (B)
Hospital Vall d´Hebron
Paseo Vall d´Hebrón 119-129
08035 Barcelona
Spain
Phone: 34 93 4894034
Fax: 34 93 4894032
1
Copyright 2002 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on June 11, 2002 as Manuscript M203595200 by guest on A
pril 10, 2019http://w
ww
.jbc.org/D
ownloaded from
Page 2
E-mail: [email protected]
SUMMARY
We have found that RNase P from HeLa cells specifically and efficiently cleaves hepatitis
C virus (HCV) transcripts in vitro. The evidence includes identification of the 5’ phosphate
polarity of the newly generated termini at position A2860, as well as immunological and
biochemical assays. Active cleavage has been shown in five dominant sequences of HCV
quasispecies differing at or near the position of cleavage, demonstrating that this is a general
property of HCV RNA. During the analysis a second cleavage event was found in the 3’ domain
of the internal ribosome entry site (IRES). We have found that HCV RNA competitively inhibits
pre-tRNA cleavage by RNase P, suggesting that HCV RNA has structural similarities to tRNA.
This finding sets HCV apart from other pathogens causing serious human diseases and represents
the first description of human RNase P-viral RNA cleavage. Here we discuss the possible
meaning of these RNase P-accessible structures built into the viral genome and their possible role
in vivo. Moreover, such structures within the viral genome might be vulnerable to attack by
therapeutic strategies.
Keywords: Ribonuclease P, Hepatitis C Virus, Quasispecies, Ribozymes, tRNA, tRNA-like
structures
2
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 3
INTRODUCTION
Hepatitis C virus is a human pathogen causing chronic liver disease in 170 million people
worldwide. The virus is classified within the family Flaviviridae (1). The RNA genome is single-
stranded and functions as the sole mRNA species for translation (1) (Fig. 1A). It comprises a 5’-
untranslated region, which functions as an internal ribosome entry site (2), and a long open reading
frame, which encodes a polyprotein precursor of about 3010 amino acids, that is cleaved into
structural (core, envelope 1, envelope 2 and p7) and non-structural (NS-2, NS-3, NS-4 and
NS-5) proteins (3); followed by a 3’ non-coding region (4).
Analyzing significant numbers of cDNA clones of hepatitis C virus (HCV) from single
isolates provides unquestionable proof that the viral genome cannot be defined by a single
sequence, but rather by a population of variant sequences closely related to one another (5-7). In the
infected patient, a master (the most frequently represented sequence) and a spectrum of mutant
sequences (diverging by up to 5%) may be isolated at any given time during chronic infection (7).
This manner of organizing genetic information, which characterizes most RNA viruses, is
referred to as quasispecies (8). It has been proven that the use of this strategy provides RNA viruses
with a rapid increase of fitness while growing in cell culture conditions (9).
Many studies on genetic variability in recent years have focused on the analysis of HCV
quasispecies. Clinically relevant features, such as the ability to produce chronic infections and
severity of disease (including the frequency of hepatocellular carcinoma), have been related to
the interplay between host influences and the array of viral variants in each infected individual (10).
HCV resistance to interferon treatment (either alone or in combination with ribavirin) is one of
the most important clinical implications predicted by the quasispecies model (11-14) suggesting the
3
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 4
necessity to seek new therapies. HCV therapeutic strategies based on ribozyme cleavage are
leading candidates. It may be argued that a sequence-dependent ribozyme designed to cleave
viral RNA by interaction with a motif in the viral RNA, may, in fact select for (mismatching)
variants resistant to the ribozyme. However, strategies could be designed to take advantage of
ribozyme capabilities to minimize the effect of virus variability. Combination therapy with
multiple ribozymes directed against independent viral loci has been demonstrated to be efficient
in inhibiting influenza virus replication in cell culture (15). Making conserved motifs within the viral
genome accessible to therapy, as in the case of the HCV IRES, could be another promising
strategy.
The ribozyme activity of Ribonuclease P (RNase P) is among proposed antiviral agents (16).
RNase P is a ubiquitous cellular endonuclease and one of the most abundant and efficient
enzymes in the cell. This enzyme is a ribonucleoprotein complex that catalyzes a hydrolysis
reaction to remove the leader sequence of precursor tRNA to generate the mature tRNA (17). RNase
P from Escherichia coli contains a catalytic RNA subunit termed M1 RNA and a single
polypeptide known as C5 protein (18). In the presence of a high concentration of Mg2+, M1 RNA
itself can hydrolyze tRNA precursors in vitro (19). Human RNase P also contains an RNA subunit,
H1 RNA, but in the absence of protein factors, H1 RNA does not exhibit enzymatic activity by
itself in vitro (20,21). Substrate recognition by the RNase P ribozyme does not rely on sequence
requirements but on structural features of the RNA substrate. Custom-designed ribo-
oligonucleotides, which hybridize with the target, called external guide sequences (EGSs), may
provide the RNA structure which RNase P recognizes and cleaves in the hybridized complex (16).
Recognition of structures instead of sequences may represent a great advantage in the
4
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 5
fight against variable viruses because single or even double mutations in the target may be
tolerated for RNase P recognition (15). Also, it has already been shown that some forms of the
catalytic RNA moiety from E.coli RNase P, M1 RNA (either specifically modified or in vitro
selected) can be introduced into the cytoplasm of mammalian cells for the purpose of carrying
out targeted cleavage of mRNA molecules (22,23).
While performing targeting experiments on HCV RNA transcripts with RNase P we have
found that, surprisingly, purified RNase P (peak activity) from HeLa cells cleaved HCV genomic
RNA efficiently at two sites in the absence of EGSs. We report here the techniques used to prove
that the cleavage is specific to human RNase P, and to show where cleavage occurs. We further
report that cleavage is maintained in several variant sequences, which makes RNase P cleavage
an inherent property of HCV RNA. Since RNase P recognizes and cleaves tRNA-like structures,
these results suggest the presence of tRNA like structures within the viral genome.
EXPERIMENTAL PROCEDURES
Preparation of RNA transcripts
RNA transcripts used as substrates in the human RNase P assays derived from plasmids pN(1-
4728) Bluescript, which contains nt 1-4728 of hepatitis C virus under the T7 promoter, and
pUC19 TyrT which contains the sequence of the naturally occurring precursor to tRNATyr . To
obtain the radioactive substrates for peak RNase P activity from HeLa cells, one to two µg DNA
template were transcribed in vitro (1 h at 37ºC) with [α-32P]-GTP or [α-32P]-NTPs followed
by a 5 min treatment with RNase-free DNase I at 37ºC. We used cellulose CF11
chromatography to eliminate DNA fragments and non-incorporated nucleotides. Transcripts
5
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 6
were then purified by gel electrophoresis under denaturing conditions on 4% polyacrylamide gels
containing 7M urea. Bands were visualized by autoradiography, excised from the gel and eluted
in buffer (100mM Tris-HCl, pH 7.5 and 10mM EDTA, pH 7.5). The concentration of
radioactive transcripts was determined by calculating the amount of incorporated [α-32P]-GTP
based on scintillation counting.
Partial purification of human RNase P
RNase P was purified from 30 gr of HeLa cells according to the method of Bartkiewicz et
al. (20) with some modifications. Fractions eluted with a liner gradient of 100-350 mM from a
column of DEAE-Sepharose CL-6B (bed volume, 150 ml) were tested to determine i)
enzymatic activity using pre-tRNATyr as substrate ii) presence of the H1 RNA moiety from
RNase P but also presence of RNA from MRP RNase which could co-extract with RNase P
during the purification protocol. H1 RNA and MRP RNA were quantified by using Taqman
technology (Roche Molecular Systems) and real time RT-PCR (Abi Prism 7700, PE
Biosystems) following the protocol used for the quantification of HCV RNA from human serum
or liver samples (24) (data not shown). We have used one set of specific human RNase P primers
(PH1-213: 5’CCCGGCGGATGCCT3’ and PH1-274: 5’TTGAACTCACTTCGCTGGCC3’)
and a fluorogenic probe (PH1-228: 5’-(VIC)
CTTTGCCGGAGCTTGGAACAGACTCA(TAMRA)-3’), and a second set of specific human
MRP primers (MRP-90: 5’AGAGAGTGCCACGTGCATACG3’ and MRP-210:
5’TAACTAGAGGGAGCTGACGGATG3’) and a fluorogenic probe labeled with a different
reporter (MRP-145: 5’(FAM)CGCCAAGAAGCGTATCCCGCTGA(TAMRA)3’). Relative
quantitation of both RNase P RNA and MRP RNA was performed by comparing the
6
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 7
amplification results for the different fractions with those on standard curves generated from
serial dilutions of total RNA extracted from HeLa cells. Using the purification protocol described
above, RNase P and MRP co-extracted together but with an enrichment of RNase P versus MRP
of several orders of magnitude, in all the tested fractions (3.3 x 1011 molecules of RNase P RNA
versus 8.2 x 107 molecules of MRP RNA, on average, in the fractions from the ammonium
chloride gradient).
Fractions with coincident peaks of enzymatic activity and H1 RNA amplification were
pooled and concentrated using the Millipore Ultrafree-15 Centrifugal Filter Device, to a final
volume of approximately 6 ml. The concentrated fractions were subjected to linear glycerol
gradient centrifugation, as described (20). Relative quantitation of RNase P and MRP RNA
molecules at this point confirmed previous results, that is enrichment of RNase P versus MRP
during the purification process (6.7 x 1010 molecules of RNase P RNA versus 6.7 x 107
molecules of MRP RNA, on average, in the fractions from the glycerol gradient). Again,
fractions containing the peak of enzymatic activity were concentrated to a final volume of 0.1ml
and stored at –70ºC.
RNase P cleavage assay
Substrates for RNase P assays, SI , SII, SIII and SIV transcripts (1.8 nM final
concentration), were preheated at 90ºC for 1 min, before the addition of reaction buffer (10mM
HEPES-KOH, pH 7.5, 10mM MgOAc, 100mM NH4OAc) and left to cool to room temperature.
Cleavage reactions were performed with 4% PEG, 20U RNasin and 2 µl of the RNase P peak
activity, and carried out at 30ºC in a volume of 10 µl for 30 min. Samples were subjected to 2%
7
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 8
SDS and 5 min at 60ºC to disrupt aggregates before loading. Cleavage products were separated
on 4% denaturing polyacrylamide gels and visualized by autoradiography.
Determination of cleavage site and phosphate polarity
Oligonucleotides released from RNase T1 digestions of all four single or mixed labeled
SI , 5’PI and 3’PI RNAs, were fractionated to yield two-dimensional fingerprints (first separation
by gel electrophoresis and second by homochromatography) exactly as described (25). Standard
conditions for secondary analysis (with pancreatic RNase A, RNase U2, RNase T2 or 0.4 M
NaOH) (26) were followed permitting the oligonucleotides to be identified. One-dimensional
electrophoresis on DEAE or 3MM paper was done following Barrells’ protocol (26).
Immunoprecipitation of RNase P activity
Serum containing anti-Th antibodies from a patient with an autoimmune disease was
used to immunodeplete RNase P activity (27). Protein A Sepharose (PAS) beads (125µg of dry
weight, Pharmacia) were washed with TMKT buffer (10mM Tris-HCl, pH 7.5, 10mM MgCl2,
100mM KCl, 0.02% Tween 20) before incubation for 1 h at room temperature at different
concentrations of either normal serum or anti-Th serum (0.25, 0.5 or 1 µl) and in 30 µl of TMKT
buffer. After washing four times in TMKT, followed by three washes in RNase P Buffer (10mM
HEPES-KOH, pH 7.5, 10mM MgOAc, 100mM NH4OAc), the beads were incubated for 2 h at
4°C in 2.5 µl of RNase P extract. The suspension was centrifuged, and both the supernatant and
the immunoprecipitate were assayed for enzyme activity. To facilitate migration of products on
the acrylamide gel, an additional step of proteinase K treatment was carried out after phenol/SDS
treatment, on each reaction tube.
Construction of plasmids containing HCV variants
8
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 9
HCV variants were obtained from our library of cloned fragments encompassing HCV
nucleotides 2641-2872 from infected patients. Subsequently the fragments were extended by
PCR at their 3’ ends using primers HCV-2639:
5’ACAGGATCCAGTCCTTCCTTGTGTTCTTCT3’ and HCV-2871:
5’AACGAATTCCCACACATGCAAGTGCGCCTCAGCTCTGGTGATAAGATATTGTAACC
ACCA3’, corresponding to the "wild type" clone (this length of 3’ extension was required for
cleavage (data not shown)). Amplified DNA fragments were inserted in BamHI/EcoRI sites of
pGem-4Z. RNAs were synthesized from EcoRI linearized plasmids and contained an additional
45 nt stretch of plasmid polylinker. SI transcript, the DNA template for which has been cloned in
the same manner as the variant sequences, was referred as to wild type in these experiments.
Nucleotide sequence accession numbers
The nucleotide sequence for the HCV wild type genome is available in the GenBank database
under GenBank Accession Number S62220 (28).The nucleotide sequence for HCV variants
presented in this article can be accessed through EMBL database under EMBL database
accession number AJ248084, AJ391467, AJ391452 and AJ247989 (7).
RESULTS
Cleavage of HCV RNA transcripts
Initial RNase P cleavage experiments involved a 554-base HCV RNA transcript (SI nt
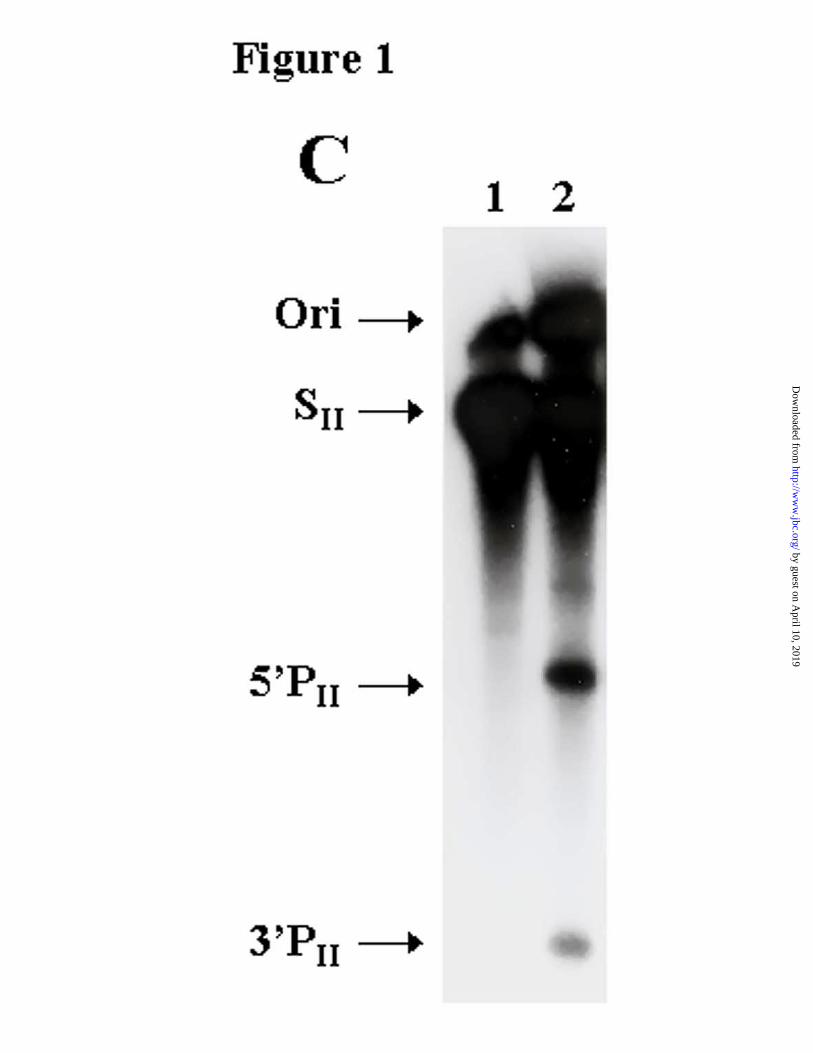
2486-3040). At the top of figure 1A is a schematic drawing of the HCV RNA genome, below
which appears the SI transcript located at the structural/non-structural junction region. The
9
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 10
autoradiogram in figure 1B shows that, using the SI transcript as a substrate, RNase P peak
activity alone, in the absence of any EGS, cleaved the HCV transcript very efficiently, producing
two intense cleavage products (5’PI and 3’PI). After this unexpected result we wanted to assess
if cleavage was maintained in a larger transcript, encompassing the first one third of the genome
(Fig.1A, SII nt 1-3032). In the course of that demonstration, we detected a second HCV
cleavage site in the IRES region (Fig. 1A and 1C). Subsequently, cleavage within the IRES was
confirmed using a third transcript corresponding to the first 1360 bases of HCV genome (Fig.
1A , SIII nt 1-1360) as well as in a shorter fragment, 641 nt long (SIV nt 1-641).
RNase P is responsible for the HCV RNA cleavages
The key experimental question of the HCV RNA cleavages obtained in the reactions
involving non-guided RNase P concerns demonstration that cleavages in different sized
transcripts are performed by RNase P itself, and not by a co-extracted contaminant. To prove
that, we have used direct and indirect methods.
Direct Method: End group determination and cleavage precision by RNA fingerprinting.
If RNase P were responsible for HCV RNA processing activity in our RNase P peak
activity (HeLa cell extract), the site of cleavage might be expected to occur between precise
nucleotide positions, and release products containing the 3’ hydroxyl and 5’ phosphate end
groups (17). Contaminating RNases almost invariably cleave to yield 5’ hydroxyl and 3’ phosphate
end groups (29), and very few other RNases and ribozymes cleave the phosphodiester backbone
through the same mechanism used by RNase P (30). SI RNA substrate: To allow direct and precise
10
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 11
determination of the cleavage site as well as to identify the phosphate polarity of the newly
cleaved termini, substrate SI was internally labeled either separately with [α-32 P] –GTP, -ATP,
-CTP and -UTP, or simultaneously with all four-labeled rNTPs and incubated with RNase P
peak activity under the conditions described in the experimental procedures. Both cleavage
products (5’PI and 3’PI) and control transcript (SI) were fractionated by gel electrophoresis,
eluted from the gel and subjected to RNase T1 digestion. Figure 2 shows two-dimensional
fingerprint analysis (25) of the oligonucleotides generated by RNase T1 digestion of single or mixed
labeled uncut SI substrate, 5’PI and 3’PI cleavage products, and identifies the exact
phosphodiester bond cleaved during the cleavage reaction to be between residues A2860 and
G2861 within the four base RNase T1-resistant oligonucleotide 2858CUAG2861 found to be
missing from both the 5’ PI and 3’ PI fingerprint patterns (data confirmed by secondary RNase
digestion, not shown). These experiments also demonstrate that the final position of the
phosphate group is 5’ to the cleavage. More precisely, the arrow in panel D indicates that a new
spot appears in the GTP-labeled 3’ PI fingerprint that does not appear in the UTP-labeled 3’ PI
(arrow in panel C). Neither does it appear in panels A and B nor in the ATP- nor CTP-labeled 3’
PI fingerprints (data not shown). This novel spot was eluted and confirmed to be the 5’ terminal
residue pGp, by its mobility with respect to markers in secondary analysis by one-dimensional
electrophoresis on DEAE and 3MM paper (26).
SIV RNA substrate: Figure 3 depicts RNA fingerprinting analysis of the substrate SIV
RNA, corresponding to the HCV IRES, as well as both cleavage products generated by RNase P.
11
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 12
In each case, T1-resistant oligonucleotides were eluted and subjected to further enzymatic
characterisation as described elsewhere (26). As summarized in the legend of figure 3, a spot (panel
A, spot 1) from the intact SIV RNA fingerprint was absent from the fingerprint pattern of both
cleavage products. This missing RNase T1-resistant oligonucleotide containing the RNase P
cleavage site(s) in the SIV RNA substrate is the 17mer (nt 351-367), thus indicating precise
cleavage of the RNA substrate in the HCV IRES domain. This 17mer is replaced by a new spot
in each one of the cleavage products’ fingerprints (panel B spot 2; panel C spot 3). Together
these two were found by secondary analysis to contain the sequence of the missing 17mer with
the expected composition indicating a cleavage site in the vicinity of bases 361-363. Further
experiments are in progress to pin down exact termini within this RNase P cleavage domain.
Indirect methods: Immunoprecipitation and competitive inhibition
Two indirect strategies were used to demonstrate that the activity which cleaves SI and
SII, and which exactly co-purified with RNase P peak activity, was in fact RNase P. The
experiments were carried out using SI , SII and SIII as substrates.
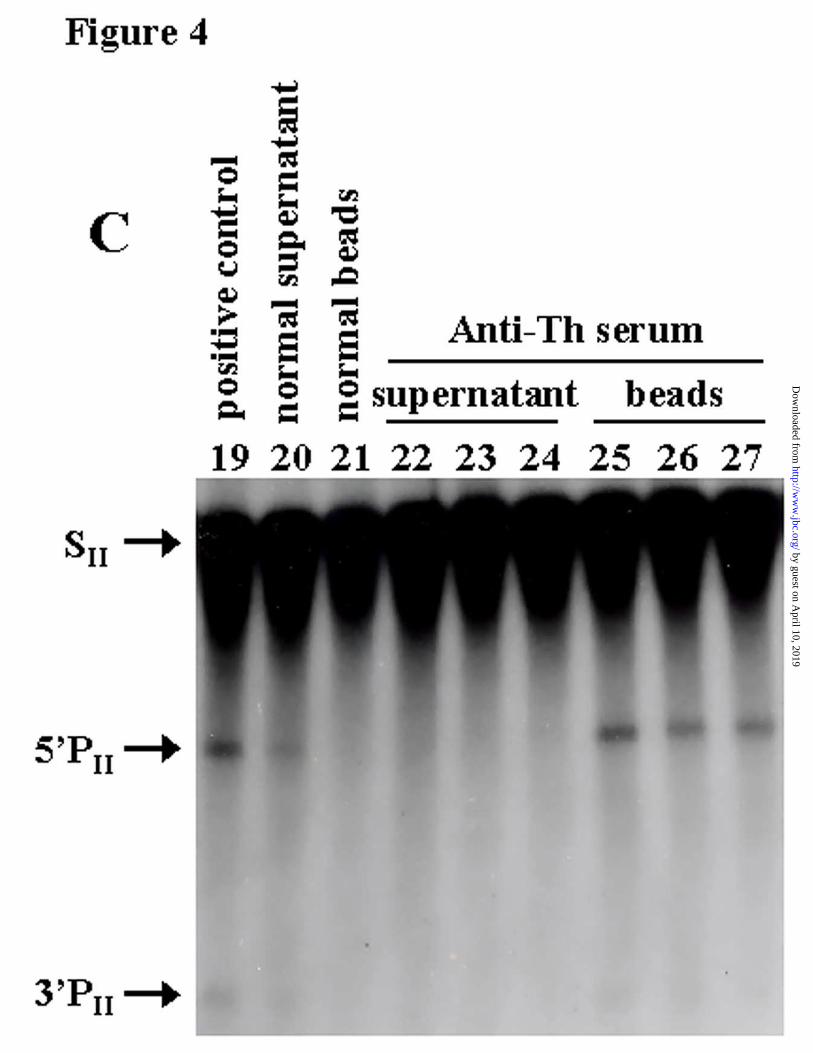
Immunoprecipitation: Some patients with autoimmune diseases produce antibodies
against a 38-40 kDa protein (designated Th antigen) which is an integral component of
eukaryotic RNase P (27). In the immunoprecipitation experiment we have incubated a serum
12
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 13
containing anti-Th antibodies with our peak activity (RNase P extract) and we have assayed both
the supernatant and the pellet. The anti-Th serum that immunodepleted pre-tRNATyr cleavage
activity from our glycerol gradient-purified enzyme (Fig. 4A, lane 2) also immunodepleted
cleavage of HCV transcripts (Fig. 4B, lanes 6-8; Fig. 4C, lanes 22-24). Moreover, the
autoimmune serum was able to precipitate the processing activity of HCV transcripts SI and SII
(Fig. 4B, lanes 9-11; Fig. 4C, lanes 25-27) as well as pre-tRNATyr (Fig. 4A, lane 3) and a
human suppressor pre-tRNA (data not shown). In contrast, the normal human serum failed to
precipitate the processing activity (Fig. 4B, lanes16-18; Fig. 4C, lane 21). Control reactions of
RNase P cleavage, using protein A-Sepharose beads with no added antiserum, showed that the
cleavage activity remains in the supernatant after the immunoprecipitation and is not found in the
pellet (Fig. 4B, lanes 5 and 12). An inverse correlation between the percent of cleavage with
increasing anti-Th sera (Fig. 4C, lanes 25, 26 and 27) may be due to inactivation of RNase P
activity due the presence of polyclonal antibodies reacting with important motifs for substrate
recognition.
Competitive Inhibition: like RNase P, MRP RNases cleave RNA to generate 5’ phosphate
and 3’ hydroxyl termini (31) and may be immunoprecipitated with anti-Th serum. As a
distinguishing feature, MRP RNase does not cleave pre-tRNATyr (32). To rule out that MRP
enzymes were responsible for HCV cleavage we carried out competitive inhibition experiments
between HCV RNA and pre-tRNA. The experiments consisted of incubating the same amount
of HCV RNA (SI, SII or SIII ) with increasing concentrations of pre-tRNA (from 1/8-fold to 4-
fold) in order to inhibit HCV RNA cleavages. When labeled pre-tRNATyr was included in the
13
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 14
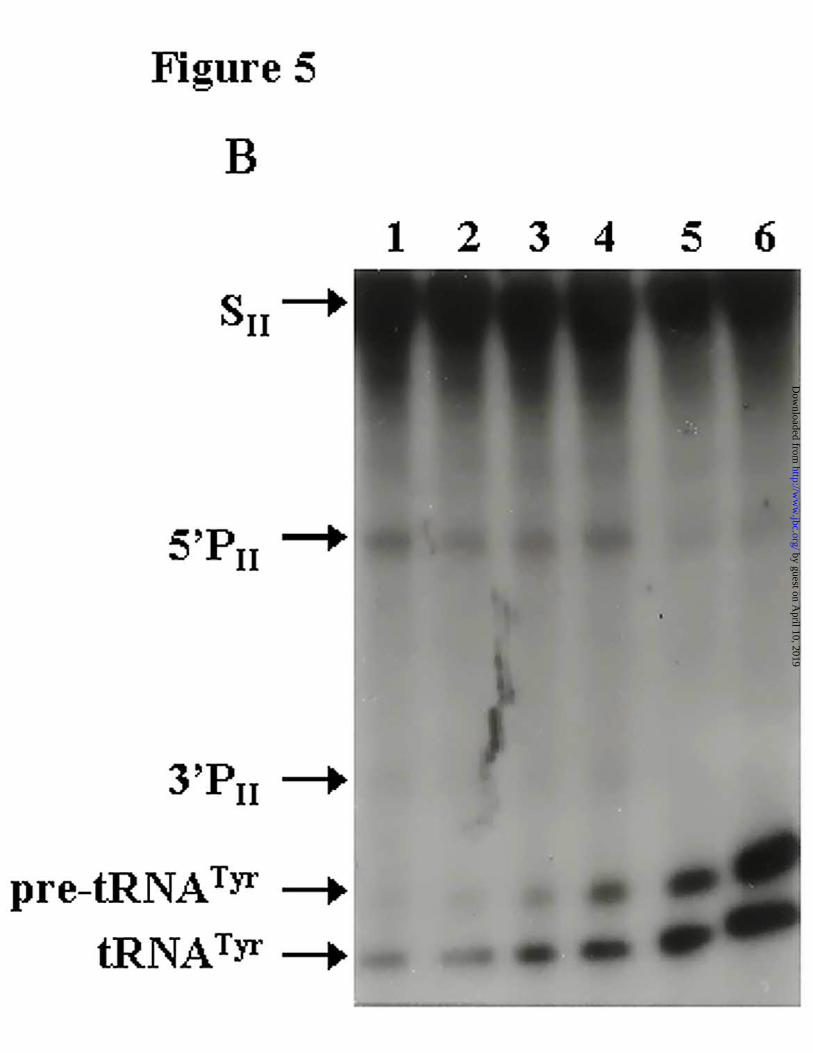
SI HCV cleavage reactions, the amount of HCV cleavage product decreased with an increasing
competitor concentration at a ratio near 1:1 (Fig. 5A, lane 5). With the long transcript (SII), a
decrease of products from the two cleavage reactions was more noticeable when the molar ratio
between pre-tRNATyr and SII reached 2:1 (Fig. 5B).
RNase P cleaves HCV variants
The direct consequence of the high mutation rate in HCV replication is that variant
genomes are continuously being generated. Thus, multiple variant sequences (quasispecies) co-
circulate within a patient and each patient carries a virus with a distinct "master" (the most
frequent) sequence (5,10). To define cleavage by RNase P as a general property of HCV, four viral
sequences obtained from different patients, together with SI sequence (referred to as wild type
here), were compared for RNase P cleavage accessibility (Fig. 6A). We used transcripts from
cloned HCV PCR fragments, representing the master sequence from infected patients’
quasispecies, with mutations at the vicinity or exactly at the nucleotides adjacent to the scissile
bond in the structural/non-structural junction. Cleavage was consistently observed in all
sequences tested although with different efficiencies (Fig. 6B).
HCV RNA competes with RNase P cleavage of pre-tRNA.
The concept of RNA mimicry has been defined for those cases where the structure of an
RNA molecule has evolved to fit a binding site on a protein or a macromolecular complex which
normally interacts with a different RNA (33). The specific cleavage of HCV RNA by RNase P
suggests that the viral RNA has structural similarities to tRNA. We wished to assess how much
these HCV RNA structures resemble tRNA. In competition experiments reciprocal to those
14
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 15
shown in figure 5, we tested the ability of HCV RNA transcripts SI and S III (0.9 nM to 180 nM)
to compete for RNase P activity with the natural substrate pre-tRNATyr (1.8 nM). Figure 7
shows that by using an RNase P concentration capable of cleaving around 25% of the pre-
tRNATyr in the reaction, the amount of pre-tRNA cleavage products decreased with increasing HCV
RNA concentration. The amount of HCV RNAs required for half inhibition were between 4 to 6
fold molar excess and were similar for both SI and SIII. In contrast, similar amounts of an
unrelated RNA of 400nt length corresponding to hepatitis B virus surface antigen mRNA (nt
1400 to nt 1800 of HBV adr subtype) (34) which is not cleaved by RNase P, had no observable
effect on RNase P activity on pre-tRNA (data not shown). The fact that the HCV RNA is a
competitive inhibitor of pre-tRNA cleavage within one order of magnitude range evidences
molecular mimicry between the HCV RNA motifs at the cleavage sites and those in pre-tRNA.
Furthermore, the inhibition of pre-tRNA cleavage provides strong evidence that the interaction
of HCV RNA is with RNase P and not with RNase MRP, in agreement with our previous
conclusion that RNase P is responsible for HCV RNA cleavage.
The results in Figures 2 and 3, which were first completed using RNAs produced and
cleaved in the Barcelona lab, have also been repeated in the New York lab. These experiments
were repeated using two different preparation of human RNase P (from Dr. Sidney Altman) and
the same results were reproduced.
DISCUSSION
15
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 16
We have defined a new specific interaction in vitro between HCV RNA and a host
component, RNase P; and we have confirmed that HCV RNA transcripts act as competitive
inhibitors of pre-tRNAtyr processing. This represents evidence for a similarity in structure and/or
function between both accessible motifs in HCV RNA and tRNA molecules.
RNase P specifically cleaves pre-tRNA in all organisms to produce mature 5’ ends. There
have been questions raised about the universal significance of RNase P cleavage in non-tRNA
molecules, especially concerning yeast RNase P. Chamberlain et al. (35) have suggested that yeast
RNase P can sometimes cleave 5.8S rRNA at sites which lack properties normally associated with
canonical tRNA. Given the uncertain evolutionary history of this rRNA species, however, it is
hard to prove or disprove the acquisition of internal tRNA-like domains, which are in fact present
in analogous prokaryotic spacer regions of rRNA precursors. It is also the case that RNase P
cleavage has reliably identified a number of authenticated tRNA-like domains in non-tRNA
molecules, including bacterial SRP and tmRNAs and various plant viral RNA genomes (36-40). Given
that our two HCV domains undergo RNase P cleavage with the same efficiency as pre-tRNA, we
believe that such recognition by RNase P is an indication for the presence of two possible tRNA-
like structures in the HCV genome.
The ability to mimic tRNA as we observe here in HCV RNA was first discovered 30
years ago at the 3’ end of the turnip yellow mosaic virus because its ability to undergo covalent
linkage with amino acids catalyzed by aminoacyl tRNA synthetase (41). Subsequently, this and other
plant viral RNAs were seen to be accessible to a battery of factors involved in other tRNA-
related activities (including accessibility of bacterial RNase P)(37,41,42). Nevertheless, in vivo functional
mimicry was not complete since viral RNAs were not amino acid donors for protein synthesis but
16
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 17
rather participated in virus replication.
This proposed idea is further supported by the presence of a pseudoknot near the HCV
IRES(43) cleavage site (a common element in tRNA-like structures including that which is known to
interact with E.coli RNase P in the case of the tRNA-like motifs of plant viral RNAs)(42) (Fig. 8).
Such a structure might contribute to the recruitment of the translational machinery in the absence
of a 5’ terminal cap. A recent study by J.R. Lytle et al. (44), confirms that this pseudoknot domain of
the HCV IRES is among those protected from extensive pancreatic RNase A digestion by
initiating 80S ribosomes.
During protein elongation, this RNA structure might also help a ribosomal frameshift
which has been described to happen within cleavage boundaries and that generates a new antigen
in HCV infected patients (45,46).
Concerning the internal cleavage site, the predicted secondary structure formed by the
RNA sequence flanking the cleavage site (the cleavage is 5’ to a G residue (G2861) and is
followed by two helices totaling 13 bases connected through a bulge (program: RNA structure
3.5) is in agreement with the characteristic features of the minimal model substrate for human
RNase P (47) (Fig. 8). In particular, cleavage determinants are confined to the tRNA domain that
contains the acceptor stem, the T stem and loop, and the junction between them (47), a recognition
feature also shared by the E.coli elongation factor EFTu (33). Also, the internal HCV RNase P
cleavage site resides in a highly structured domain of the viral RNA (data not shown), which
might be also compatible with a tRNA structure. Nevertheless, there is no readily obvious
functional explanation for such a structure at this site.
The presence of HCV RNA in the nucleus (where most RNase P is found) has not been
17
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 18
demonstrated (48), and no evidence of subgenomic HCV RNAs has been reported (49), arguing against an
active role of RNase P cleavage in HCV biology. Whatever the role of the RNase P-sensitive
structures, their importance for virus viability is apparent, despite the notorious heterogeneity and
dynamics of change in HCV quasispecies within the infected patient. A cleavage site at residue
A2860, present in the master sequences from individual patients (differing at or near the position
of cleavage), should be interpreted in a context where variants arise continuously, and are
repeatedly subjected to competition pressures. Thus the relative success of a mutant is the result
of its ability to replicate. This strongly supports that (i) the RNA structure that confers
accessibility to RNase P is not affected by mutations which become fixed within the cleavage
boundaries during error-prone replication; (ii) there is a continuous selective advantage for the
sequences within the quasispecies carrying the RNase P-accessible structure. Moreover,
conservation of RNase P-cleavable structures in the genomes of different patients implies that
this structure is even conserved during genetic bottlenecking of HCV quasispecies during host-
to-host transmission, in spite of the fact that this area is one of the most variable regions of the
HCV genome at the nucleotide sequence level (6). Altogether, this makes RNase P cleavability an
inherent property of HCV.
Higher-order structures of RNA play functional roles, and the mutations that alter such
higher-order structures must be subjected to negative selection. Such a strong tendency to
maintain RNase P-sensitive structures within the viral genome might be important in the
development of therapeutic strategies against the virus because they can represent highly
susceptible targets for E. coli RNase P M1 RNA (22,23).
The next phase of this work will involve investigation of the minimum requirements for
18
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 19
cleavage at both the IRES and internal site. Minimal length substrates will serve to define to what
extend tRNA processing enzymes like aminoacyl-tRNA synthetase, tRNA nucleotidyl
transferase, tRNA methyltransferases, and interacting factors (iEF2 and EF-1) react with these
HCV motifs. Comparison of such results at the two HCV RNase P cleavage sites should help us
to understand in greater detail HCV substrate structure, tRNA mimicry, rules underlying
recognition by human RNase P, and, in the particular case of the IRES motif, possible
participation in translation.
ACKNOWLEDGMENTS
We thanks Drs. C. Gelpí and J.L. Rodriguez from the Servei d’Immunologia, Hospital
Sant Pau (Barcelona), for providing the autoimmune serum. An HCV clone was kindly provided
by Drs. M. Honda and S. Lemon and tRNA precursors and RNase P by Drs. S. Altman and C.
Guerrier-Takada. We also thank Dra. E.Martinez-Salas for critical reading of the manuscript.
Work in New York was supported by the U.S. N.I.H. Work in Barcelona was funded by the
Ministerio de Ciencia y Tecnología (SAF1999-0108 and BIO00-0347), Ministerio de Sanidad y
Consumo (FISS-01/1351) , and by the Hospital Vall d´Hebron.
Correspondence and requests for material should be addressed to J. Gómez.
(e-mail: [email protected] )
19
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 20
REFERENCES
1. Houghton, M. (1996) Hepatitis C virus. In Fields, B. N., Knipe, D. N., and Howley, P. N., editors. Field´s Virology, Lippincott-Raven, Philadelphia,PA.
2. Tsukiyama-Kohara, K., Iizuka, N., Kohara, M., and Nomoto, A. (1992) J.Virol. 66, 1476-1483
3. Houghton, M., Selby, M., Weiner, A., and Choo, Q. L. (1994) Curr.Stud.Hematol.Blood Transfus. 1-11
4. Han, J. H., Shyamala, V., Richman, K. H., Brauer, M. J., Irvine, B., Urdea, M. S., Tekamp-Olson, P., Kuo, G., Choo, Q. L., and Houghton, M. (1991) Proc.Natl.Acad.Sci.U.S.A. 88, 1711-1715
5. Martell, M., Esteban, J. I., Quer, J., Genesca, J., Weiner, A., Esteban, R., Guardia, J., and Gomez, J. (1992) J.Virol. 66, 3225-3229
6. Bukh, J., Miller, R. H., and Purcell, R. H. (1995) Semin.Liver Dis. 15, 41-63
7. Cabot, B., Martell, M., Esteban, J. I., Sauleda, S., Otero, T., Esteban, R., Guardia, J., and Gomez, J. (2000) J.Virol. 74, 805-811
8. Domingo, E. and Holland, J. J. (1994) Mutation rates and rapid evolution of RNA viruses. In Morse, S. S., editor. Evolutionary biology of viruses, Raven Press, New York
9. Domingo, E. and Holland, J. J. (1997) Annu.Rev.Microbiol. 51, 151-178
10. Gomez, J., Martell, M., Quer, J., Cabot, B., and Esteban, J. I. (1999) J.Viral Hepat. 6, 3-16
11. Poynard, T., Marcellin, P., Lee, S. S., Niederau, C., Minuk, G. S., Ideo, G., Bain, V., Heathcote, J., Zeuzem, S., Trepo, C., and Albrecht, J. (1998) Lancet 352, 1426-1432
12. Enomoto, N., Kurosaki, M., Tanaka, Y., Marumo, F., and Sato, C. (1994) J.Gen.Virol. 75, 1361-1369
13. Le Guen, B., Squadrito, G., Nalpas, B., Berthelot, P., Pol, S., and Brechot, C. (1997) Hepatology 25, 1250-1254
14. Polyak, S. J., McArdle, S., Liu, S. L., Sullivan, D. G., Chung, M., Hofgartner, W. T., Carithers, R. L., Jr., McMahon, B. J., Mullins, J. I., Corey, L., and Gretch, D. R. (1998) J.Virol. 72, 4288-4296
15. Plehn-Dujowich, D. and Altman, S. (1998) Proc.Natl.Acad.Sci.U.S.A 95, 7327-7332
16. Altman, S. (1995) Biotechnology (N.Y.) 13, 327-329
20
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 21
17. Robertson, H. D., Altman, S., and Smith, J. D. (1972) J.Biol.Chem. 247, 5243-5251
18. Altman, S. (1989) Adv.Enzymol.Relat Areas Mol.Biol. 62, 1-36
19. Guerrier-Takada, C., Gardiner, K., Marsh, T., Pace, N., and Altman, S. (1983) Cell 35, 849-857
20. Bartkiewicz, M., Gold, H., and Altman, S. (1989) Genes Dev. 3, 488-499
21. Yuan, Y. and Altman, S. (1995) EMBO J. 14, 159-168
22. Kilani, A. F., Trang, P., Hsu, J. S., Kim, J., Nepomuceno, E., Liou, K., and Liu, F. (2000) J.Biol.Chem. 14, 10611-10622
23. Liu, F. and Altman, S. (1995) Genes Dev. 9, 471-480
24. Martell, M., Gómez, J., Esteban, J. I., Sauleda, S., Quer, J., Cabot, B., Esteban, R., and Guardia, J. (1999) J.Clin.Microbiol. 37, 327-332
25. Branch, A. D., Benenfeld, B. J., and Robertson, H. D. (1989) Methods Enzymol. 180, 130-154
26. Barrell, B. G. (1971) In Cantoni, G. L. and Davies, D. R., editors. Procedures in nucleic acid research, Harper and Row, New York
27. Eder, P. S., Kekuda, R., Stolc, V., and Altman, S. (1997) Proc.Natl.Acad.Sci.U.S.A 94, 1101-1106
28. Hayashi, N., Higashi, H., Kaminaka, K., Sugimoto, H., Esumi, M., Komatsu, K., Hayashi, K., Sugitani, M., Suzuki, K., and Tadao, O. (1993) J.Hepatol. 17 suppl 3, S94-S107
29. Adams, R. L. P., Knowler, J. T., and Leader, D. P. (1992) Degradation of nucleic acids. The biochemistry of the nucleic acids, Chapman and Hall Ltd., London
30. Pyle, A. M. (1993) Science. 261, 709-714
31. Chang, D. D. and Clayton, D. A. (1987) EMBO J. 6, 409-417
32. Karwan, R., Bennett, J. L., and Clayton, D. A. (1991) Genes Dev. 5, 1264-1276
33. Springer, M., Portier, C., and Grunberg-Manago, M. (1998) RNA mimicry in the translational apparatus. In Simons, R. W., editor. RNA structure and function, Cold Spring Harbor Laboratory Press, Cold Spring Harbor,NY
34. Ono, Y., Onda, H., Sasada, R., Igarashi, K., Sugino, Y., and Nishioka, K. (1983) Nucleic Acids Res. 11, 1747-1575
21
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 22
35. Chamberlain, J. R., Pagàn-Ramos, L., Kindelberger, D. W., and Engelke, D. R. (1996) Nucleic Acids Res. 24, 3158-3166
36. Peck-Miller, K. and Altman, S. (1991) J.Mol.Biol. 221, 1-5
37. Guerrier-Takada, C., van Belkum, A., Pleij, C. W., and Altman, S. (1988) Cell 53, 267-272
38. Baumstark, T. and Ahlquist, P. (2001) RNA 11, 1652-1670
39. Joshi, S., Chappeville, F., and Haenni, A. L. (1982) Nucleic Acids Research 10, 1947-1962
40. Komine, Y., Kitabatke, M., Yokogawa, T., Nishikawa, K., and Inokuchi, H. (1994) Proc.Natl.Acad.Sci.U.S.A. 91, 9923-9227
41. Giegé, R., Florentz, C., and Dreher, T. W. (1993) Biochimie 75, 569-582
42. Mans, R., Guerrier-Takada, C., Altman, S., and Pleij, C. (1990) Nucleic Acids Res. 18, 3479-3487
43. Wang, C., Le, S. Y., Ali, N., and Siddiqui, A. (1995) RNA. 1, 526-537
44. Lytle, J. R., Wu, L., and Robertson, H. D. (2001) J.Virol. 75, 7629-7636
45. Waleswsky, J. L., Keller, T. B., Stump D, D., and Branch, A. D. (2001) RNA 20, 3840-3848
46. Xu, Z., Choi, J., Yen, T. S., Lu, W., Stoecher , A., Govindarajan, S., Chien, D., Selby, M. J., and Ou, J.-H. (2001) EMBO J 20, 3840-3848
47. Yuan, Y. and Altman, S. (1995) EMBO J. 14, 159-168
48. Bartenschlager, R. and Lohmann, V. (2000) J Gen.Virol 81, 1631-1648
49. Moradpour, D., Kary, P., Rice, C. M., and Blum, H. E. (1998) Hepatology 28, 192-201
50. Lyons, A. J., Lytle, J. R., Gómez, J., and Robertson, H. D. (2001) Nucleic Acids Res. 29, 2535-2541
22
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 23
FIGURE LEGENDS
Figure 1. RNase P cleaves the HCV genome. A, Location of RNase P cleavage sites and HCV
transcripts in the viral genome. SI is a 554 nt transcript encompassing the junction fragment
between structural and non-structural regions; SII covers the first one-third of the HCV genome;
SIII is a 1360 nt long transcript containing the 5’ non-coding region and half of the structural
region. B, C and D, Autoradiograms of RNase P cleavage of SI , SII and SIII transcripts,
respectively. Lane 1 corresponds to the transcript alone. The arrows indicate the 5’ product (5’P)
and the 3’ product (3’P) (lane 2).
Figure 2. Determination of the cleavage site in the structural/non-structural junction region.
RNase T1 fingerprints of: A, uncut SI labeled by all four [α-32P]-rNTPs containing all of the
expected RNase T1-resistant oligonucleotide spots between residues 2486-3040. B, 5’ PI
labeled by all four [α-32P]-rNTPs containing spots that correspond to nucleotides 2486-2857.
C, and D, 3’ PI labeled by [α-32P]-rUTP or [α-32P]-rGTP, respectively, containing the
expected spots that correspond to residues 2862-3040 (data confirmed by secondary analysis of
eluted spots). Arrows in panels C and D indicate the position of RNase T1 generated pGp from
3’ PI.
23
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 24
Figure 3. Mapping the cleavage site in the 5’ region. RNase T1 fingerprints of: A, uncut substrate
containing HCV bases 1-641, transcribed from DNA template cleaved with SacII, labeled by all
four alpha 32P-rNTPs. This pattern contains all of the expected RNase T1-resistant
oligonucleotide spots as determined by secondary analysis (Branch et al.,1989). Spot 1,
containing the 17-base RNase T1-resistant product AAUCCUAAACCUCAAAG, is indicated,
along with positions 2 and 3, which are unoccupied in this panel. B, product labeled as in panel
A, containing spots corresponding to bases 1-361/362. Spot 1 is missing, and spot 2, which has
the sequence AAUCCUAAACC (bases 351-361) or AAUCCUAAACCU (bases 351-362), is
present instead. Spot 2 was identified by secondary analysis, but its 3’ end could not be identified
beyond the conclusion that it is either C361 or U362. C, 3’ product labeled as in panel A,
containing spots corresponding to bases 364-641. Spot 1 is missing and spot 3, which has the
sequence XAAAG (362/3-367, where x is C or UC), is present instead. Spot 3 was identified by
secondary analysis, and must have one or two additional bases upstream from the AAAG motif
(bases 364-367) which are not yet identified. Further experiments are in progress to pin down
exact termini within this RNase P cleavage domain.
Figure 4. Depletion of RNase P activity using an anti-Th serum. Cleavage reactions of three
different substrates: A, bacterial pre-tRNATyr ; B, SI transcript; and C, SII transcript, incubated
with immunoprecipitate from beads coated with 0.5µl (lane 3) and increasing concentrations
(0.25, 0.5 and 1 µl) of either anti-Th serum (lanes 9-11 and 25-27) or normal human serum
(lanes 16-18). Supernatant from beads coated with 0.5 µl (lane 2) and the same previous three
concentrations of anti-Th serum (lanes 6-8 and 22-24) or normal serum (lanes 13-15) were also
24
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 25
used to incubate the transcript. Only the intermediate concentration of normal serum (lane 20,
21) was tested with SII transcript. Lanes 5 (supernatant) and 12 (beads) are controls showing the
lack of affinity between PAS beads and RNase P without anti-Th serum addition. Lane 1 is pre-
tRNA alone, lanes 4 and 19 are standard reactions used as positive controls.
Figure 5. pre-tRNATyr competes with and inhibits the RNase P specific cleavage of HCV RNA.
Autoradiograms of the cleavage of A, SI transcript and B, SII transcript, by RNase P in the
presence of increasing concentrations of pre-tRNATyr. Lane c: pre-tRNA alone; Lanes 1-5:
cleavage reactions using a constant concentration of SI or SII transcript (1.8 nM) and increasing
concentrations of pre-tRNA (0.225 nM, 0.45 nM, 0.9 nM, 1.8 nM and 3.6 nM, respectively). An
additional assay using 7.2 nM pre-tRNATyr (lane 6) is shown in panel B.
Figure 6. RNase P cleavage of HCV 2658-2869 RNA variants. A, Schematic representation of
HCV variants cloned in a transcription vector. The open arrowhead indicates RNase P cleavage
site in the RNA sequence. Wild type (wt) corresponds to SI transcript. Wt and variant sequences
at +/- eight nts flanking the site of cleavage are shown beneath. B, Autoradiograms show the
RNase P specific cleavage of four different HCV variant and wt RNAs. (+) means presence of
RNase P in the reaction. The 5’ cleavage products are indicated by an arrow.
Figure 7. Competitive inhibition of pre-tRNA processing by the HCV RNA. A, Lanes 4-8:
different amounts of SI transcript (0.9nM, 1.8nM, 3.6nM, 7.2nM, 14.4nM and 28.8nM) were
25
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 26
premixed with 1.8 nM of pre-tRNATyr transcript and incubated with RNase P. In all the
reactions, a concentration of RNaseP able to cut the 20-30% pre-tRNATyr alone in 30 min was
used (Lane 2). Lane 1: pre-tRNATyr alone. B, the same protocol as in part A was carried out
with SIII transcript.
Figure 8. Positioning of the two RNAse P cleavage sites on the predicted secondary structure
model for the HCV RNA. HCV genome is represented schematically and the putative structures
of domains containing the two RNAse P cleavages sites are drawn. A, The 5’NCR containing the
IRES domain (nt 1-383) is shown as the currently accepted structure including the redrawing of
domain II (50) and the predicted pseudoknot structure in domain IV. The two-arrows indicates the
RNase P cleavage site in the vecinity of bases 361-363. B, The structure of a 70 nts portion (nt
2841-2890) of the folded structural/non-structural junction region, predicted by RNA structure
3.5 program, is represented. The single arrow indicates the exact position of human RNase P
cleavage.
26
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 27
by guest on April 10, 2019 http://www.jbc.org/ Downloaded from
Page 28
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 29
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 30
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 31
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 32
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 33
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 34
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 35
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 36
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 37
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 38
by guest on April 10, 2019 http://www.jbc.org/ Downloaded from
Page 39
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 40
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Page 41
by guest on April 10, 2019 http://www.jbc.org/ Downloaded from
Page 42
Cabot, Juan I. Esteban, Rafael Esteban, Jaime Guardia and Jordi GomezAnna Nadal, Maria Martell, J.Robin Lytle, Alita J. Lyons, Hugh D. Robertson, Beatriz
Specific cleavage of hepatitis C virus RNA genome by human RNase P
published online June 11, 2002J. Biol. Chem.
10.1074/jbc.M203595200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 10, 2019
http://ww
w.jbc.org/
Dow
nloaded from