Spermine modulates neuronal excitability and NMDA receptors in juvenile gerbil auditory thalamus Israeli Ran a , Robert M. Miura b , Ernest Puil a;c ; a Department of Pharmacology and Therapeutics, The University of British Columbia, Vancouver, BC, Canada V6T 1Z3 b Departments of Mathematical Sciences and of Biomedical Engineering, New Jersey Institute of Technology, Newark, NJ 07102, USA c Department of Anesthesia, The University of British Columbia, Vancouver, BC, Canada V6T 1Z3 Received 27 June 2002; accepted 28 October 2002 Abstract Medial geniculate body (MGB) neurons process synaptic inputs from auditory cortex. Corticothalamic stimulation evokes glutamatergic excitatory postsynaptic potentials (EPSPs) that vary markedly in amplitude and duration during development. The EPSP decay phase is prolonged during second postnatal week but then shortens, significantly, until adulthood. The EPSP prolongation depends on spermine interactions with a polyamine-sensitive site on receptors for N-methyl-D-aspartate (NMDA). We examined effects of spermine application on EPSPs, firing modes, and membrane properties in gerbil MGB neurons during the P14 period of highest polyamine sensitivity. Spermine slowed EPSP decay and promoted firing on EPSPs, without changing passive membrane properties. Spermine increased membrane rectification on depolarization, which is mediated by tetrodotoxin (TTX)-sensitive, persistent Na þ conductance. As a result, spermine lowered threshold and increased tonic firing evoked with current injection by up to V150%. These effects were concentration-dependent (ED 50 = 100 WM), reversible, and eliminated by NMDA receptor antagonist, 2-amino-5-phosphonovalerate (APV). In contrast, spermine increased dV/dt of the low threshold Ca 2þ spike (LTS) and burst firing, evoked from hyperpolarized potentials. LTS enhancement was greater at 355 mV than at hyperpolarized potentials and did not result from persistent Na þ conductance or glutamate receptor mechanisms. In summary, spermine increased excitability by modulating NMDA receptors in juvenile gerbil neurons. ß 2002 Published by Elsevier Science B.V. Key words: Spermine; Auditory thalamic neuron; Medial geniculate body; N-methyl-D-aspartate ; NR2B 1. Introduction Neurons of the medial geniculate body (MGB) inte- grate synaptic information from glutamatergic projec- tions of the auditory cortex and inferior colliculus (Gonzalez-Lima and Scheich, 1984; Morel and Imig, 1987; Hu et al., 1994; Metherate and Cruikshank, 1999). Stimulation of the corticothalamic pathway acti- vates postsynaptic N-methyl-D-aspartate (NMDA) and non-NMDA receptors on MGB neurons (Bartlett and Smith, 1999). The receptors for NMDA, robustly ex- pressed in the MGB (Sucher et al., 1995; Khan et al., 2000), become functional at an early age. By the end of the second postnatal week in the rat, NMDA receptors on thalamocortical neurons mediate the transformation of sound-related synaptic inputs to output ¢ring pat- terns (Heierli et al., 1987). The glutamatergic responses of neurons in visual cortex depend on the subunit com- position of the NMDA receptor, including the NR2B subunit (Roberts and Ramoa, 1999). The NR2B sub- unit, which MGB neurons express in high levels at post- natal day 14 (P14; Hsieh et al., 2002), confers an NMDA receptor sensitivity to spermine and other poly- amines which enhance the response to glutamatergic stimulation (Williams et al., 1994). Abnormal polyamine modulation of glutamatergic systems may occur in pathological states of auditory function (reviewed by McCann and Pegg, 1992). Phar- macological inhibition of polyamine synthesis decreases 0378-5955 / 02 / $ ^ see front matter ß 2002 Published by Elsevier Science B.V. PII:S0378-5955(02)00746-3 * Corresponding author. Tel.: +1 (604) 822-5080; Fax: +1 (604) 822-6012. E-mail address: [email protected](E. Puil). Hearing Research 176 (2003) 65^79 R Available online at www.sciencedirect.com www.elsevier.com/locate/heares
Israeli Ran a, Robert M. Miura b, Ernest Puil a;c;!
a Department of Pharmacology and Therapeutics, The University of British Columbia, Vancouver, BC, Canada V6T 1Z3b Departments of Mathematical Sciences and of Biomedical Engineering, New Jersey Institute of Technology, Newark, NJ 07102, USA
c Department of Anesthesia, The University of British Columbia, Vancouver, BC, Canada V6T 1Z3
Received 27 June 2002; accepted 28 October 2002
Abstract
Medial geniculate body (MGB) neurons process synaptic inputs from auditory cortex. Corticothalamic stimulation evokesglutamatergic excitatory postsynaptic potentials (EPSPs) that vary markedly in amplitude and duration during development. TheEPSP decay phase is prolonged during second postnatal week but then shortens, significantly, until adulthood. The EPSPprolongation depends on spermine interactions with a polyamine-sensitive site on receptors for N-methyl-D-aspartate (NMDA).We examined effects of spermine application on EPSPs, firing modes, and membrane properties in gerbil MGB neurons during theP14 period of highest polyamine sensitivity. Spermine slowed EPSP decay and promoted firing on EPSPs, without changingpassive membrane properties. Spermine increased membrane rectification on depolarization, which is mediated by tetrodotoxin(TTX)-sensitive, persistent Na" conductance. As a result, spermine lowered threshold and increased tonic firing evoked withcurrent injection by up to V150%. These effects were concentration-dependent (ED50 = 100 WM), reversible, and eliminated byNMDA receptor antagonist, 2-amino-5-phosphonovalerate (APV). In contrast, spermine increased dV/dt of the low thresholdCa2" spike (LTS) and burst firing, evoked from hyperpolarized potentials. LTS enhancement was greater at 355 mV than athyperpolarized potentials and did not result from persistent Na" conductance or glutamate receptor mechanisms. In summary,spermine increased excitability by modulating NMDA receptors in juvenile gerbil neurons.! 2002 Published by Elsevier Science B.V.
Neurons of the medial geniculate body (MGB) inte-grate synaptic information from glutamatergic projec-tions of the auditory cortex and inferior colliculus(Gonzalez-Lima and Scheich, 1984; Morel and Imig,1987; Hu et al., 1994; Metherate and Cruikshank,1999). Stimulation of the corticothalamic pathway acti-vates postsynaptic N-methyl-D-aspartate (NMDA) andnon-NMDA receptors on MGB neurons (Bartlett andSmith, 1999). The receptors for NMDA, robustly ex-pressed in the MGB (Sucher et al., 1995; Khan et al.,
2000), become functional at an early age. By the end ofthe second postnatal week in the rat, NMDA receptorson thalamocortical neurons mediate the transformationof sound-related synaptic inputs to output ¢ring pat-terns (Heierli et al., 1987). The glutamatergic responsesof neurons in visual cortex depend on the subunit com-position of the NMDA receptor, including the NR2Bsubunit (Roberts and Ramoa, 1999). The NR2B sub-unit, which MGB neurons express in high levels at post-natal day 14 (P14; Hsieh et al., 2002), confers anNMDA receptor sensitivity to spermine and other poly-amines which enhance the response to glutamatergicstimulation (Williams et al., 1994).
Abnormal polyamine modulation of glutamatergicsystems may occur in pathological states of auditoryfunction (reviewed by McCann and Pegg, 1992). Phar-macological inhibition of polyamine synthesis decreases
0378-5955 / 02 / $ ^ see front matter ! 2002 Published by Elsevier Science B.V.PII: S 0 3 7 8 - 5 9 5 5 ( 0 2 ) 0 0 7 4 6 - 3
polyamine concentrations in the cochlea (Schweitzer etal., 1986) and induces a temporary hearing loss in hu-mans and rats (McCann and Pegg, 1992). On the otherhand, excessive NMDA-mediated excitation contributesto absence epilepsy in an animal model (Koerner et al.,1996). Audiogenic seizure activity, initiated in brain-stem nuclei (Faingold et al., 1989), enhances MGB re-sponsiveness to acoustic stimuli (N’Gouemo and Fain-gold, 1997). Hence, high concentrations of polyaminesmay induce excessive NMDA receptor activation inMGB neurons which can impair auditory informationprocessing.
Despite the potential signi¢cance, there is little infor-mation about the e¡ects of polyamines on the ¢ringmodes as well as other functional aspects of thalamo-cortical neurons, especially during postnatal stages ofdevelopment. The present study on MGB neurons ex-amines, for the ¢rst time, the e¡ects of spermine andreceptor antagonists on the tonic and burst ¢ringmodes, excitatory and inhibitory postsynaptic poten-tials (EPSPs and IPSPs), and the electrical membraneproperties. For these experiments, we used gerbilswhich have a hearing capacity similar to humans(Kraus et al., 1987). At P14, the gerbil’s thalamus hasabundant NR2B subunits, implying a neuronal sensitiv-ity to polyamines (Hsieh et al., 2002).
2. Materials and methods
The procedures and conditions for thalamic slicepreparation were similar to those described previouslyfor rats (Tennigkeit et al., 1996). In brief, young gerbils(10^14-day-old) were decapitated while under deep iso-£urane anesthesia. For the preparation of slices, thecerebral hemispheres were removed rapidly (V1 min)from the cranial vault and immersed for 1^2 min in ice-cold (0^2‡C) arti¢cial cerebrospinal £uid (ACSF). TheACSF contained (in mM): NaCl, 124; NaHCO3, 26;glucose, 10; KCl, 4; CaCl2, 2; MgCl2, 2; and KH2PO4,1.25. The ACSF, on saturation with 95% O2 and 5%CO2, had a pH of 7.3. The brain tissue was trimmedinto a cube (V0.125 cm3) that contained the cortex andthalamus. We used a Vibroslicer (Campden Instru-ments, London, UK) to cut slices (thickness, 300^500Wm) in a horizontal orientation. The slices includedportions of the medial geniculate, inferior collicular,and reticularis thalamic nuclei. The slices were incuba-ted for 2^3 h in ACSF at room temperature (21^25‡C),until required for recording, which was carried out at21^25‡C.
Whole-cell patch-clamp techniques were used to rec-ord from thalamocortical neurons of a slice situated ona nylon mesh, and perfused with oxygenated ACSF(Tennigkeit et al., 1996). The electrode pipettes were
pulled from borosilicate glass tubing (WP-Instruments,Sarasota, FL, USA), using a Narishige puller (Narish-ige Instruments, Tokyo, Japan, Model PP83). The pi-pette solution contained (in mM): K-gluconate, 140;N-2-hydroxyethylpiperazine-N-2-ethanesulfonate(HEPES), 10; KCl, 5; NaCl, 4; adenosine 5-triphos-phate (disodium salt), 3; guanosine 5-triphosphate (tri-sodium salt), 0.3; ethylene glycol-bis-(L-aminoethyl-ether)-N,N,NP,NP-tetraacetic acid (EGTA), 10; andCaCl2, 1. This combination of EGTA and Ca2" yieldeda ¢nal [Ca2"] of 10 nM (calculated with Max Chela-tor software). In experiments with the Ca2" chelator1,2-bis(2-aminophenoxy)ethane-N,N,NP,NP-tetraacetate(BAPTA), we substituted EGTA with an equimolarconcentration of BAPTA, which yielded a ¢nal [Ca2"]of 1 nM. The calculated ECl was 355 mV. The pipettesolution had a pH of 7.3. The tip resistances of thepipettes were 5^10 M6 and access resistances were be-low 25 M6.
Guided by di¡erential interference contrast microsco-py (100U objective) and using a micromanipulator, wepositioned the electrode tip in the ventral partition ofthe medial geniculate nucleus. The electrical recordingswere performed in the current-clamp mode with an Ax-oclamp 2A ampli¢er (Axon Instruments, Foster City,CA, USA). We used Digidata 1320A hardware withpClamp 8 software (Axon Instruments), running on aPentium computer for data acquisition, storage, andanalysis. The input resistance (Ri) of the neuron wascomputed from the voltage displacements (range, 5^8mV) to intracellular injection of hyperpolarizing currentpulses or from the linear slope of current^voltage (I^V)relationships. The membrane time constant was esti-mated from a single exponential ¢t to a voltage re-sponse of 5^10 mV to a hyperpolarizing current pulse.The voltage values have been corrected for a measuredjunction potential of 310 mV.
We evoked tonic ¢ring of action potentials from neu-rons at DC-held potentials near V360 mV by injectingdepolarizing current pulses with an amplitude of 1.5times the amplitude of ‘just-threshold’ pulses, whichwas estimated from the voltage de£ection that resultedin action potentials in 50% of the trials. We evokedEPSPs by electrical stimulation, using a bipolar tung-sten electrode (tip diameter V100 Wm), placed in theslice at 0.2^0.3 mm mediodorsal to the MGB and nearcorticothalamic axons. Stimulation at this position re-sulted only in EPSPs. The stimuli consisted of singlepulses of approximately 30 V in amplitude (range, 10^100 V) and 100^200 Ws in duration. The stimulationrate was 0.5 Hz. Using these stimulus parameters, weevoked inhibitory postsynaptic potentials (IPSPs) whenthe electrode was placed in the brachium, midway be-tween the inferior colliculus and MGB. The postsynap-tic potentials were averaged and ¢tted with an K func-
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^7966
tion (pClamp 8 software), yielding the rise and decaytime constants for the EPSPs.
The drugs were prepared in distilled water as stocksolutions. The stock solutions were frozen for storageand thawed just before the experiment. Spermine, tetro-dotoxin (TTX), 6-cyano-7-nitroquinoxaline-2,3-dione(CNQX), and D-2-amino-5-phosphono-valerate (APV)were diluted V1000 times with ACSF to the concen-trations required for the particular experiment. Thesesubstances, ATP, EGTA, GTP, HEPES, and the inor-ganic chloride salts were obtained from Sigma (St.Louis, MO, USA). As with the ACSF, the drugs wereapplied with a roller-type pump at a rate of 2 ml/minthrough a submersion-type chamber that had a volumeof V1.0 cc.
In most cases, we analyzed data from neurons of 14-day-old gerbils ; however, some exceptions were madedue to the availability of the age group. We incorpo-rated four neurons from P10, P12 and P13 in the dataanalysis, including: (1) one out of 19 neurons from a13-day-old gerbil in the spermine group in studies oftonic ¢ring; (2) two out of eight neurons from a 12-day-old gerbil in the APV group; and (3) one out offour neurons from a 13-day-old gerbil in the BAPTAgroup. All ¢gures are from neurons of 14-day-old ger-bils. The data are presented as means" S.E.M. A Stu-dent’s paired t-test was used for comparison of re-sponses to drug application between two groups. Forcomparison of more than two groups, we used analysisof variance (ANOVA). P6 0.05 was considered signi¢-cant.
3. Results
3.1. Spermine application increases tonic ¢ring
Spermine application reversibly increased the numberof action potentials in all neurons depolarized from restby current pulse injection. Spermine (100 WM) appliedfor 3^6 min induced tonic ¢ring of action potentials ontop of subthreshold responses. When action potentialswere present in the control, spermine application in-creased the rate of ¢ring (Fig. 1A). Long recovery timesof 35^45 min characterized spermine’s e¡ects on thala-mic ¢ring modes after 6 min applications. In the neuronof Fig. 1A, substantial recovery occurred at V32 minafter discontinuing the spermine application.
The spermine-induced increase in the ¢ring frequencywas concentration-dependent over the range between 50and 500 WM (n=19, Fig. 1B). In addition, applicationof a spermine concentration of 1 WM did not a¡ect¢ring (n=2); however, at 1 mM, there was a markedincrease in the ¢ring rate (n=2), without any apparentrecovery (data not shown). Spermine, applied at an
ED50 of 100 WM (cf. Fig. 1B), reversibly increased thenumber of action potentials per pulse by an average ofV80% in nine neurons (control, 1.8" 0.3 action poten-tials/pulse and spermine application, 3.3" 0.4 actionpotentials/pulse, paired t-test, P6 0.01).
The increased ¢ring due to spermine did not likelyresult from changes in the passive membrane propertieswhich did not greatly change during 3^6 min spermineapplications (cf. subthreshold responses in Figs. 1A and3A). The average resting potentials were 367" 4 mVduring the control period, and 366" 5 mV during ap-plications of spermine at 50^500 WM (n=19). Spermineapplication did not signi¢cantly change the mean mem-brane time constant (dm = 64" 6 ms in control, and76" 6 ms during spermine application, paired t-test)and mean input resistance (Ri = 772" 38 M6 in control,and 756" 61 M6 during spermine application, paired t-test), computed from the responses to hyperpolarizingcurrent pulse injections in 19 neurons held at 365 mV.Hence, the spermine-induced e¡ects on the passiveproperties could not account for the changes in ¢ringthreshold.
Spermine (100 WM) decreased the latency to tonic¢ring by decreasing the threshold (Fig. 2). Sperminedecreased the ¢ring threshold from 350.9" 0.6 mV to
Fig. 1. Spermine enhanced tonic ¢ring in a concentration-dependentmanner in MGB neurons. (A) Spermine application (100 WM, 3 min)enhanced action potential ¢ring evoked by current pulses (25 and50 pA, 500 ms, 1.5Uthreshold). Holding potential, 365 mV. Verti-cal upper bar, 30 mV and lower bar, 60 pA; horizontal bar, 150ms. (B) Increase in number of action potentials per pulse was con-centration dependent. The control ¢ring was 1.8" 0.3 action poten-tials/pulse (n=9). The ED50 was V100 WM for spermine-enhanced¢ring which approached saturation at 200 WM.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^79 67
357.1" 2.2 mV (Fig. 2B). Signi¢cant changes in actionpotential amplitude did not accompany the decreasedthreshold. Fig. 2B summarizes the e¡ects of spermineon ¢ring threshold for six neurons.
3.2. NMDA receptors mediate the spermine-inducedincrease in ¢ring
We examined the possibility that NMDA receptorsmediated the e¡ects on the ¢ring threshold and tonic¢ring rate by determining the interactions of spermineand the competitive antagonist, APV (50 WM). Asshown in Fig. 2B, spermine reduced the threshold vol-tage for an action potential evoked with a 500 ms cur-rent pulse, by an average of 6.2" 1.1 mV. On recoveryfrom spermine (washout, Fig. 2B), application of APValone, or in combination with spermine, did not sig-ni¢cantly change the ¢ring threshold (during APV,352.1" 1.9 mV and APV+spermine, 353.0" 1.7 mV;n=6) or changes in membrane properties that couldaccount for the blockade of the spermine-induced re-duction in ¢ring threshold. This signi¢ed that APVacted on NMDA receptors to completely block sper-mine action. Since these neurons had received a sper-mine application prior to APV, we also applied APV to¢ve neurons that had not previously received a drugapplication in order to assess the possibility of consti-tutive release of glutamate in the slice. Here, APV pro-duced an increase in threshold, which remained largelyunaltered by a subsequent, combined application withspermine (Fig. 2C). All neurons showed substantial re-covery at 15 min after discontinuing the application.These data implicated an NMDA receptor mechanismin the spermine-induced decrease in the threshold.
NMDA receptors also mediated the spermine-in-duced increase in ¢ring rate. In the neuron of Fig.3A, spermine application (100 WM, 3 min) reversiblyincreased the number of action potentials during a500 ms current pulse injection from one action potentialin the initial control, to three action potentials. In theneurons that had not previously received a spermineapplication, an increase in the current pulse amplitudeduring action potential blockade due to APV applica-tion, produced a return of the action potential (Fig.3B). The APV blockade of spermine-induced increasein ¢ring also was overcome by an increase in the cur-rent amplitude. The APV-induced blockades of actionpotentials and spermine enhancement of ¢ring were notattributable to an increased input conductance andwere completely reversible. The graph of Fig. 3C sum-marizes the data that implicate NMDA receptor medi-ation.
We assessed the possibility that non-NMDA recep-tors for glutamate may have contributed to the in-creased ¢ring during spermine application. In these
Fig. 2. Spermine (100 WM) decreased the action potential threshold.(A) Spermine application produced a leftward shift in action poten-tial latency (current pulse duration, 500 ms). Arrows point tothreshold in control and spermine. (B) NMDA receptors mediatedspermine e¡ects on action potential threshold. The control thresholdwas 350.9" 0.6 mV, which spermine reduced to 357.1" 2.2 mV(n=6). Partial recovery was observed after 15 min (352.4" 0.6).Blockade of NMDA receptors by APV (50 WM) reduced the thresh-old by 6 1 mV. A reduction in threshold was not observed duringco-application of APV and spermine (30.9" 0.6 mV, n=6). (C)APV (50 WM) increased ¢ring threshold in ¢ve neurons from352.3" 0.7 to 48.7" 0.5 mV. Subsequent spermine application didnot alter the increased threshold (349.1" 0.6 mV). Two-way AN-OVA; * in (B) indicates P6 0.01 and in (C) P6 0.05. Vertical barin (A), 15 mV; horizontal bar, 50 ms.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^7968
studies, we determined the interactions of spermine withan AMPA receptor antagonist, CNQX. Application ofCNQX (30 WM) for 6 min did not result in signi¢cantchanges in evoked action potential ¢ring, con¢guration,or membrane electrical properties. A subsequent, com-bined CNQX and spermine application did not greatlyalter the reduction in threshold and V200% increasein ¢ring rate evoked by current pulses (amplitudeV1.5Uthreshold), as observed with prior spermine ap-plication in all ¢ve neurons (CNQX, 1.4" 0.3 actionpotentials/pulse, and CNQX+spermine, 4.2" 0.4 actionpotentials/pulse; data not shown). Hence, the increasein tonic ¢ring rate due to spermine application did notlikely involve AMPA receptors.
3.3. E¡ects on membrane recti¢cation
We examined the possibility that the spermine-in-duced increase in tonic ¢ring involved voltage-depen-dent membrane properties. For example, thalamocor-tical neurons exhibit larger responses to depolarizing,compared to hyperpolarizing current pulses (Tennigkeitet al., 1996; Parri and Crunelli, 1998; cf. Fig. 4B).Spermine (100 WM) application for 3 min increasedthis recti¢cation in a range between the rest and ¢ringthreshold but did not appreciably change the responsesat hyperpolarized potentials, down to 3100 mV (n=19;Fig. 4A). Quanti¢cation of the increase in recti¢cationon depolarization was di⁄cult because spermine appli-cation shortened the latency to ¢ring (cf. arrows inFigs. 2A and 4A). Application of APV (50 WM, 6 min)completely blocked the recti¢cation in the upper rightquadrant of the current^voltage (I^V) relationship. Asubsequent co-application with spermine (100 WM) didnot greatly change this curve. The graph of Fig. 4A(right) summarizes these ¢ndings for six neurons.
There was little or no involvement of AMPA recep-tors in the spermine-induced (100 WM, 3 min) enhance-ment of recti¢cation produced by depolarizing currentpulses. The application of CNQX (30 WM, 6 min) didnot a¡ect spermine’s e¡ects on the recti¢cation in ¢veneurons. The average voltage response during co-appli-cation of CNQX and spermine (15.9" 0.6 mV) was sig-ni¢cantly di¡erent from control (11.5" 0.5 mV) orCNQX application (11.3" 0.4 mV; ANOVA, P6 0.05).Hence, the spermine-induced increase in the depolar-izing responses involved NMDA receptors, but notlikely AMPA receptors.
We investigated the possibility that spermine in-creased the recti¢cation in the upper right quadrant ofthe I^V relationship by interacting with a persistentNa" conductance. The recti¢cation observed on depo-larization from V370 mV to threshold involves a per-sistent Na" conductance, sensitive to TTX blockade(Tennigkeit et al., 1996; Parri and Crunelli, 1998).
Fig. 3. Spermine increased tonic ¢ring by interacting with NMDAreceptors. (A) Spermine application (100 WM, 3 min) reversibly in-duced ¢ring. After a 15 min washout from spermine, APV (50 WM,6 min), an NMDA receptor antagonist, blocked the evoked actionpotential. Firing was not observed during co-application of APVand spermine (3 min). Washout shows recovery at 10 min after dis-continuing the co-application. Lower traces show hyperpolarizingtests for input resistance. (B) Application of APV (50 WM, 6 min)abolished ¢ring induced by just-threshold current pulse (40 pA). Asubsequent 3 min co-application of spermine and APV did not alterthis suppression (lower superimposed traces in middle panel). Atwo-fold increase in current amplitude overcame the blockade dur-ing APV application, alone, or during co-application with spermine(upper superimposed traces in middle panel). Recovery was ob-served after 10 min washout. (C) Summary of spermine e¡ects on¢ring in six neurons. ANOVA; *P6 0.01, **P6 0.05. Vertical bar,30 mV; horizontal bar, 200 ms.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^79 69
Blockade of voltage-dependent Na" channels with TTX(0.6 WM, 6 min) decreased the slope of the I^V relation-ship, more in the depolarizing quadrant than in thehyperpolarizing quadrant. The blockade with TTX nul-li¢ed the ability of spermine (100 WM, 3 min) to in-crease recti¢cation on depolarization (n=6; Fig. 4B).The results imply that spermine increased recti¢cationin the upper right quadrant of the I^V relationshipby increasing a TTX-sensitive, voltage-dependent Na"
conductance.The spermine-induced enhancement of recti¢cation
on depolarization of the neuron also may depend on
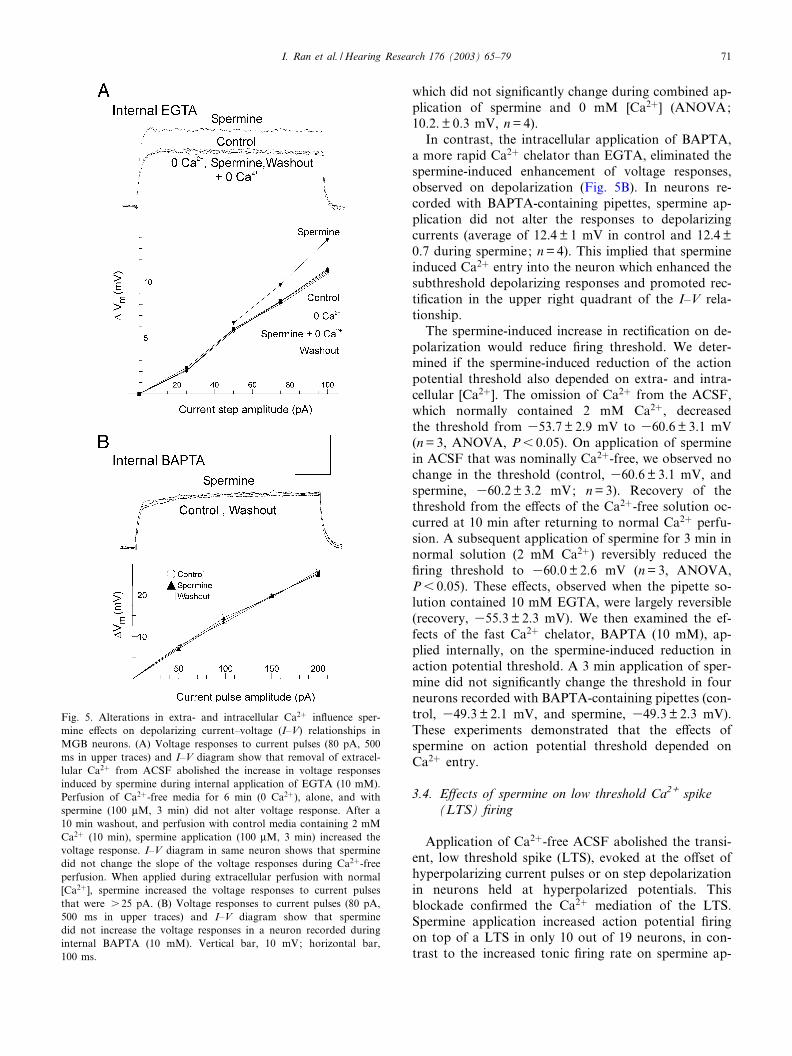
extra- or intracellular Ca2", as demonstrated for neo-cortical neurons (Crill, 1996). Hence, we measured thespermine-induced enhancement of voltage responses todepolarizing current injections during intracellular ap-plication of BAPTA (10 mM) and extracellular perfu-sion with Ca2"-free ACSF. In the neuron of Fig. 5A,perfusion of Ca2"-free ACSF did not greatly alter thesedepolarizing responses. In four neurons, a 50 pA cur-rent pulse evoked average responses of 9.8" 0.6 mV incontrol ACSF and 10.0" 0.5 mV in 0 mM [Ca2"] ; theresponse increased to 15.2" 1 mV during spermine ap-plication (in 2 mM Ca2" ACSF; ANOVA, P6 0.01)
Fig. 4. E¡ects of spermine (100 WM, 3 min) on membrane recti¢cation. (A) Current^voltage (I^V) relationship of a neuron shows that spermineincreased depolarizing response which was abolished during combined application (3 min) with APV. APV (50 WM, 6 min), alone, reduced rec-ti¢cation in upper right quadrant. I^V curve after 15 min washout shows substantial recovery. Graph at right summarizes e¡ects of spermine,APV, and their co-application on recti¢cation. The response on depolarization increased from 12.5" 0.5 mV (control) to 17.5" 0.8 mV duringspermine application, and after 15 min washout, recovered to 12.9" 0.7 mV. Subsequent APV application reduced the response on depolariza-tion from 12.9" 0.7 mV (¢rst washout) to 9.2" 0.4 mV (APV). Co-applied APV and spermine did not greatly alter recti¢cation (8.7" 0.3 mV).Recovery from APV and spermine occurred after 15 min (12.7" 0.5 mV). Holding potential, 370 mV. (B) I^V relationship for a neuron showsthat TTX application (0.6 WM, 6 min) decreased recti¢cation on depolarization over a s 10 mV range. Co-applied with TTX, spermine did notalter recti¢cation in upper right quadrant. A 20 pA pulse was su⁄cient to observe recti¢cation on depolarization, whereas a 350 pA pulse pro-duced little or no recti¢cation on hyperpolarization. Graph at right summarizes e¡ects of TTX and spermine on recti¢cation. Recti¢cation ondepolarization decreased from 13.1" 0.8 mV (control) to 10.2" 0.8 mV during TTX application. A subsequent co-application with spermine didnot greatly alter depolarizing responses (10.8" 0.7 mV). Inserts in upper left quadrants of (A) and (B) show superimposed responses (7 mV) todepolarizing and hyperpolarizing current pulses (duration 500 ms) of 60 and 360 pA, during control (C), spermine (S), and at 3 min of co-ap-plication of TTX and spermine (TTX+S). Bar graph values are mean" S.E.M. ANOVA; * and **P6 0.05.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^7970
which did not signi¢cantly change during combined ap-plication of spermine and 0 mM [Ca2"] (ANOVA;10.2." 0.3 mV, n=4).
In contrast, the intracellular application of BAPTA,a more rapid Ca2" chelator than EGTA, eliminated thespermine-induced enhancement of voltage responses,observed on depolarization (Fig. 5B). In neurons re-corded with BAPTA-containing pipettes, spermine ap-plication did not alter the responses to depolarizingcurrents (average of 12.4" 1 mV in control and 12.4"0.7 during spermine; n=4). This implied that spermineinduced Ca2" entry into the neuron which enhanced thesubthreshold depolarizing responses and promoted rec-ti¢cation in the upper right quadrant of the I^V rela-tionship.
The spermine-induced increase in recti¢cation on de-polarization would reduce ¢ring threshold. We deter-mined if the spermine-induced reduction of the actionpotential threshold also depended on extra- and intra-cellular [Ca2"]. The omission of Ca2" from the ACSF,which normally contained 2 mM Ca2", decreasedthe threshold from 353.7" 2.9 mV to 360.6" 3.1 mV(n=3, ANOVA, P6 0.05). On application of sperminein ACSF that was nominally Ca2"-free, we observed nochange in the threshold (control, 360.6" 3.1 mV, andspermine, 360.2" 3.2 mV; n=3). Recovery of thethreshold from the e¡ects of the Ca2"-free solution oc-curred at 10 min after returning to normal Ca2" perfu-sion. A subsequent application of spermine for 3 min innormal solution (2 mM Ca2") reversibly reduced the¢ring threshold to 360.0" 2.6 mV (n=3, ANOVA,P6 0.05). These e¡ects, observed when the pipette so-lution contained 10 mM EGTA, were largely reversible(recovery, 355.3" 2.3 mV). We then examined the ef-fects of the fast Ca2" chelator, BAPTA (10 mM), ap-plied internally, on the spermine-induced reduction inaction potential threshold. A 3 min application of sper-mine did not signi¢cantly change the threshold in fourneurons recorded with BAPTA-containing pipettes (con-trol, 349.3" 2.1 mV, and spermine, 349.3" 2.3 mV).These experiments demonstrated that the e¡ects ofspermine on action potential threshold depended onCa2" entry.
3.4. E¡ects of spermine on low threshold Ca2+ spike(LTS) ¢ring
Application of Ca2"-free ACSF abolished the transi-ent, low threshold spike (LTS), evoked at the o¡set ofhyperpolarizing current pulses or on step depolarizationin neurons held at hyperpolarized potentials. Thisblockade con¢rmed the Ca2" mediation of the LTS.Spermine application increased action potential ¢ringon top of a LTS in only 10 out of 19 neurons, in con-trast to the increased tonic ¢ring rate on spermine ap-
Fig. 5. Alterations in extra- and intracellular Ca2" in£uence sper-mine e¡ects on depolarizing current^voltage (I^V) relationships inMGB neurons. (A) Voltage responses to current pulses (80 pA, 500ms in upper traces) and I^V diagram show that removal of extracel-lular Ca2" from ACSF abolished the increase in voltage responsesinduced by spermine during internal application of EGTA (10 mM).Perfusion of Ca2"-free media for 6 min (0 Ca2"), alone, and withspermine (100 WM, 3 min) did not alter voltage response. After a10 min washout, and perfusion with control media containing 2 mMCa2" (10 min), spermine application (100 WM, 3 min) increased thevoltage response. I^V diagram in same neuron shows that sperminedid not change the slope of the voltage responses during Ca2"-freeperfusion. When applied during extracellular perfusion with normal[Ca2"], spermine increased the voltage responses to current pulsesthat were s 25 pA. (B) Voltage responses to current pulses (80 pA,500 ms in upper traces) and I^V diagram show that sperminedid not increase the voltage responses in a neuron recorded duringinternal BAPTA (10 mM). Vertical bar, 10 mV; horizontal bar,100 ms.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^79 71
plication, observed in all neurons. As shown in Fig. 6A,spermine induced an action potential on the rebounddepolarizing response at the termination of hyperpolar-izing current pulses. In ¢ve out of the 10 neurons hyper-polarized with DC to 380 mV, spermine applicationinduced one or two action potentials on a subthresholdrebound response to hyperpolarizing current pulses.Spermine application increased the amplitude of the re-bound LTS that did not reach action potential thresh-old in the remaining ¢ve neurons. The e¡ects of sper-mine were reversible, requiring 20^40 min for recovery.
Blockade of voltage-dependent Na" channels withTTX did not signi¢cantly alter the ability of spermineto enhance the LTS in six neurons (cf. Fig. 6A and B).During TTX blockade, the spermine enhancement ofthe LTS depended on the holding potential. In theseexperiments, we evoked the LTS by injecting hyperpo-
larizing currents of di¡erent amplitude into neuronsheld at di¡erent holding potentials (Fig. 6C). Co-appli-cation of spermine (100 WM) and TTX induced an LTSin neurons at potentials that caused marked inactiva-tion of the LTS. At potentials where an LTS waspresent, a spermine application increased its amplitudeand rate of rise (dV/dt). There was a greater increase inthe dV/dt of the LTS when the neuron was held at 355mV than at 385 mV (Fig. 6C). After spermine applica-tion, the LTS evoked in a neuron held at 355 mV hadan average dV/dt of 3.1" 0.2 mV/ms, compared to1.5" 0.3 mV/ms in the control during TTX application.The average rate of decay was 31.7" 0.3 mV/ms (n=6)with fast (26" 5 ms, n=5) and slow (146" 12 ms, n=5)components (trace 2, Fig. 6B). Fig. 6C summarizes thee¡ects of spermine on the dV/dt of the LTS, showing amaximal e¡ect at a holding potential (Vh) =355 mV
Fig. 6. E¡ects of spermine (100 WM, 3 min) on the low threshold Ca2" spike (LTS) ¢ring. (A) Superimposed voltage responses (control, sper-mine, and recovery) show that spermine induced an LTS on termination of a hyperpolarizing current pulse (340 pA, 500 ms). (B) Spermine in-creased the rate of rise and amplitude of the LTS at the end of a hyperpolarizing current pulse (380 pA) just before (1), during (2), and after(3) spermine application during TTX blockade (0.6 WM) of voltage-dependent Na" conductances. Holding potential, 355 mV. (C) Bar graphsummarizes spermine e¡ects on dV/dt of the LTS at the end of hyperpolarizing responses and during TTX blockade. Spermine increased dV/dtin neurons held at 385, 375, 365, and, maximally, at 355 mV (n=6, paired t-test, *P6 0.01, **P6 0.005). (D) APV (50 WM) did not blockthe e¡ects of spermine on the LTS, as shown by sub- and suprathreshold responses to current pulses (60, 120 pA) during application of APV,alone, and co-application with spermine. Application of APV reduced the subthreshold response, LTS rate of rise, and number of action poten-tials. Co-application (3 min) of APV and spermine transformed a subthreshold response to an LTS, increased LTS rate of rise, and shortenedthe latency to the action potential. Recovery (in APV) was observed after a 10 min washout. Vertical bar, 15 mV in (A) and (B); 30 mV in(D). Horizontal bar, 150 ms. Insert in (B) (right): Vertical bar, 3 mV; horizontal bar, 30 ms.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^7972
and a minimal increase at Vh =385 mV in six neurons(paired t-test, P6 0.01).
Since the hyperpolarization-activated current in£uen-ces the rate of rise of the LTS, we examined whetherspermine a¡ected the voltage sag, mediated by this cur-rent (Tennigkeit et al., 1996). The voltage sag was notprominent in the majority of neurons. Spermine appli-cation (100 WM) produced no change in the voltage saginduced by a hyperpolarizing current pulse in three neu-rons. Hence, the increase in the rate of rise of the LTSdid not likely involve interactions with this hyperpola-rization-activated conductance.
We determined if the spermine potentiation of theLTS involved NMDA receptors in eight neurons. Insix of these neurons, APV application decreased theamplitude of the subthreshold responses to currentpulses (cf. Fig. 6D, lower traces). As shown for theneuron of Fig. 6D, APV application also decreased anLTS burst to a single action potential and LTS rate ofrise, increasing the latency to the ¢rst action potentialon top of the LTS. Despite APV antagonism of NMDAreceptors, spermine application transformed the sub-threshold response into an LTS, as well as decreasedthe latency to an action potential on top of the LTS (cf.APV and spermine traces in Fig. 6D). In eight out ofeight neurons, APV application (50 WM, 6 min) reducedthe average rate of rise of the LTS from 1.6" 0.3 mV/ms in na|«ve controls to 1.2" 0.2 mV/ms. A subsequentco-application of APV and spermine caused a signi¢-
cant increase in the rate of rise of the LTS to 2.2" 0.1mV/ms (ANOVA, P6 0.05).
We determined if the spermine potentiation of theLTS involved interactions with AMPA receptors. Dur-ing blockade of AMPA receptors with CNQX (30 WM,6 min), spermine signi¢cantly increased the dV/dt of theLTS (control, 1.8" 0.1 mV/ms and spermine, 3.6" 0.2mV/ms; n=5, ANOVA, P6 0.05). Hence, the e¡ects ofspermine on the LTS did not likely involve NMDA orAMPA receptors.
3.5. E¡ects of spermine on excitatory and inhibitorypostsynaptic potentials
Spermine application (100 WM) to 18 neurons re-sulted in bursts of action potentials on EPSPs evokedby electrical stimulation of corticothalamic projections(Fig. 7A). Spermine had little or no e¡ects on the rateof rise of the EPSP, but always enhanced the late decayphase amplitude. The EPSP amplitude increasedslightly (3^5 mV) during spermine application to ¢veneurons, but this was not a consistent ¢nding in the18 neurons. The spermine-induced action potentialson the EPSPs were reversible in all neurons. Completerecovery was observed in 13 of 18 neurons at 35 minafter terminating the application.
Spermine prolonged the EPSP decay time constant(ddecay), as estimated with an K function ¢t of the EPSPs(Fig. 7B). This promoted the occurrence of action po-
Fig. 7. Spermine (100 WM, 3 min) prolonged late component of corticothalamic EPSPs mediated by NMDA receptors. (A) Spermine increasedEPSP amplitude and duration, resulting in three action potentials. (B) Spermine delayed the late component (2) of the EPSPs. The bar graphsummarizes the spermine-induced prolongation of EPSP decay time constant (ddecay), expressed as % of the control. Control ddecay was 142" 8.5ms (n=15, paired t-test, *P6 0.01). (C) Spermine did not a¡ect EPSPs during NMDA receptor blockade by 50 WM APV or signi¢cantlychange remaining EPSP components. Bar graph summarizes the reduction in EPSP ddecay by APV and lack of spermine e¡ect during APVblockade, expressed as % of control. Control ddecay was 143.6" 14 ms (n=8, ANOVA, *P6 0.01). Vertical bar, 5 mV; horizontal bar, 200 ms.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^79 73
tentials on top of the EPSPs (Fig. 7A). The ED50 forthe spermine-induced increase in ddecay of EPSPs wasV100 WM which was approximately the same for thespermine-induced increase in ¢ring (cf. Fig. 1B). Recov-ery to the control value occurred after 30 min (148" 15ms). Fig. 7B summarizes these results for 15 neurons.Table 1 summarizes the e¡ects of single or cumulativeapplications of spermine on the 90^10% decay time in18 neurons.
Spermine application (100 WM) did not signi¢cantlya¡ect the amplitude or time course of depolarizing po-tentials evoked by stimulation of the brachium colliculiinferioris (n=6; data not shown). These potentials,100^200 ms in duration, were likely IPSPs mediatedby GABAA receptors because they reversed at poten-tials near ECl (355 mV) and were sensitive to blockadeby bicuculline (50 WM, n=6).
Spermine prolonged the decay of the late EPSP com-ponent mediated by an NMDA-type receptor. The ap-plication of APV (50 WM), blocked the late component,resulting in shorter rise (10^90%) and decay (90^10%)times of the EPSPs (Table 1). During NMDA receptorblockade, EPSP mean ddecay was 100" 14 ms, comparedto 143" 15 ms in control (Fig. 7C; n=8, ANOVA,P6 0.01). This mean value did not change signi¢cantlyduring combined spermine and APV application (Fig.7B).
By applying spermine in combination with CNQX(30 WM, 6 min), we determined if the prolongation ofEPSPs resulted from interactions with AMPA recep-tors. We found that spermine prolonged the EPSP90^10% decay time during CNQX blockade to thesame extent as in the absence of AMPA receptor block-ade (Table 1). In three additional neurons, sperminewas applied before the co-application with CNQX.The co-application resulted in a signi¢cant prolonga-tion of the EPSP to the same extent as in the absenceof CNQX blockade of AMPA receptors (Table 1). Thiscon¢rmed that spermine a¡ected only the NMDA-
mediated component. Co-application of CNQX (30WM) and APV (50 WM) then abolished the early andlate components of the EPSP which remained absentdespite a subsequent spermine application (n=4).From these results, we suggest that spermine increasedthe duration of the EPSP decay phase by interactingwith NMDA receptors.
We considered the possibility that spermine pro-longed the EPSPs by acting on an extracellular poly-amine-sensitive site of the NMDA receptor (cf. Benve-niste and Mayer, 1993). We studied the interactions ofspermine and arcaine, an antagonist that acts at thepolyamine-sensitive site on the NMDA receptor (Rey-nolds, 1990). In these studies, we sequentially appliedarcaine (40 WM), spermine (100 WM), arcaine and sper-mine, each for 3 min (n=3). Arcaine, alone, did notgreatly alter the con¢guration of the EPSP (Fig. 8A)or produce changes in the EPSP amplitude, 90^10%decay time, and half-width (Table 1). After a 15 minwashout from arcaine application, spermine signi¢-cantly prolonged the EPSP ddecay to 180" 32 ms from101" 16 ms in the control (Fig. 8A). A subsequent co-application of spermine and arcaine abolished the ac-tions of spermine, resulting in ddecay of 118" 16 ms (Fig.8A). The graph of Fig. 8A summarizes the spermine-induced increases in EPSP ddecay and arcaine blockadeof spermine e¡ects.
We also determined if spermine increased theNMDA-mediated component of the EPSP by potenti-ating the actions of glycine on the NMDA receptor. Inthe presence of a saturating concentration of glycine (40WM), spermine still prolonged the EPSP by V49% (Fig.8B). In three neurons, spermine increased EPSP ddecayfrom 255" 44 ms to 379" 53 ms (ANOVA, P6 0.05).In summary, spermine actions on the EPSP likely in-volved an extracellular polyamine-sensitive site, and nota glycine-sensitive site of the NMDA receptor.
We attempted to assess a contribution of extracellu-lar Ca2" or Mg2" to the spermine-induced enhancement
Table 1E¡ects of spermine on EPSP variables
Amplitude (mV) 10^90% Rise time (ms) 90^10% Decay time (ms) Half-width (ms) n
Values are means" S.E.M. The spermine group was analyzed with a paired t-test and all other groups with an ANOVA test. *P6 0.05,**P6 0.01.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^7974
of EPSPs. Spermine application did not alter the am-plitude or duration of the EPSPs during a 6 min perfu-sion of Ca2"-free ACSF in three neurons (data notshown). Hence, spermine e¡ects on NMDA-mediatedEPSPs likely depended on Ca2" entry. In two neurons,the omission of Mg2" from ACSF perfusion resulted insubthreshold oscillations of the membrane potentialand spontaneous ¢ring of action potentials. These ob-servations were consistent with previous studies on tha-lamocortical neurons (Jacobsen et al., 2001) which pre-vented critical assessment of an agonist role of Mg2" atthe polyamine-sensitive site on the NR2B receptor sub-unit (Kew and Kemp, 1998).
4. Discussion
These studies demonstrate that extracellular sperminehas distinctive actions on thalamocortical neurons, con-sistent with a neuromodulator role in the medial genic-ulate body. Spermine actions on NMDA receptors pro-duced a heightened state of excitability which weviewed as prolonged corticothalamic EPSPs, and in-creased EPSP bursting and tonic ¢ring of action poten-
tials. To a large extent, these e¡ects resulted from in-creased membrane recti¢cation on depolarization andreduction in threshold for action potential genesis.Spermine also modulated the burst ¢ring mode by in-creasing the rate of rise and amplitude of low thresholdCa2" spikes (LTSs). This unusual e¡ect did not involveinteraction with glutamate receptors. The modulationof corticothalamic excitation and LTSs of MGB neu-rons may be critical in the transformation of auditorysignals in gerbil thalamus at the P14 stage of develop-ment.
4.1. Enhancement of late component of NMDA-mediatedEPSPs
Spermine application increased the decay time con-stant of corticothalamic EPSPs mediated by NMDAreceptors. This ¢nding is consistent with the increasedamplitude of NMDA-evoked currents during spermineapplication to cultured hippocampal and spinal neurons(Lerma, 1992; Benveniste and Mayer, 1993). In thepresent studies, the actions of spermine were selectiveand required extracellular Ca2" because they were abol-ished in three neurons by brief Ca2"-free perfusion.
Fig. 8. Spermine (100 WM) prolonged the EPSPs by interacting with the polyamine-sensitive site on NMDA receptor. (A) Arcaine (40 WM), ablocker at the polyamine-sensitive site on NMDA receptors, almost eliminated the spermine-induced prolongation of the EPSP, expressed as %control ddecay which was 100.4" 16 ms (n=3, ANOVA, *P6 0.05). Note that there was a 10 min washout after arcaine, prior to spermine ap-plication. (B) Spermine (100 WM) prolonged the EPSP ddecay during co-application with glycine (10 WM). Control ddecay (glycine present) was255" 44 ms (n=3, paired t-test, *P6 0.05). Bar graphs summarize the e¡ects of spermine on EPSP ddecay during arcaine and glycine applica-tions. Holding potential, 360 mV. Vertical bar, 5 mV; horizontal bar, 100 ms.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^79 75
Spermine also did not signi¢cantly alter the early EPSPcomponent mediated by AMPA receptors, or appreci-ably a¡ect IPSPs mediated by GABA receptors thatwere sensitive to bicuculline antagonism. Applicationof APV completely blocked the spermine-induced in-crease in the EPSP decay time constant. This implicatedNMDA receptors in spermine actions.
The e¡ects of spermine on MGB neurons involveda polyamine-sensitive site on the NR2B subtype ofNMDA receptors, as we demonstrated with arcaineand glycine applications. Arcaine, itself, did not havesigni¢cant e¡ects on the passive and active membraneproperties but reversed the spermine-induced decreaseof the EPSP decay. Previous studies have shown thatarcaine blocks spermine actions by inverse agonism,antagonism, and open-channel blockade of the poly-amine-sensitive site on NMDA receptors (Reynolds,1990; Pritchard et al., 1994). The actions of spermineat this site decreased the EPSP decay, despite saturatingconcentrations of glycine. These observations are con-sistent with the glycine-independent potentiation ofNMDA currents by spermine at the NR2B receptorsubunit in cultured hippocampal neurons (Benvenisteand Mayer, 1993). In thalamocortical neurons, the per-sistence during high glycine concentrations and arcainereversal imply that spermine acted independently of theglycine site at a speci¢c polyamine-sensitive site on theNR2B receptor subunit of the NMDA receptor.
The NR2B subunit may modulate the decay timeconstant of the NMDA receptor-mediated EPSP duringthe development in MGB neurons. At the end of thesecond postnatal week, thalamocortical neurons expressan abundance of the NR2B polyamine-sensitive recep-tor subtype in the MGB and lateral geniculate body(LGB) of the rat (Chen and Regehr, 2000). The dura-tion of EPSPs mediated by NMDA receptors in LGBneurons of the rat is similar at P14 to that in gerbilMGB neurons. The decay time constant in LGB neu-rons is longer at P14 in rats than at earlier (P7^P13) orlater (P16^P28) stages of development (Chen and Re-gehr, 2000; cf. also rat MGB at P21^P42, Bartlett andSmith, 1999). Hence, spermine modulation of theNR2B subunit may cause the longer EPSP durationin MGB neurons at P14.
4.2. Enhancement of membrane recti¢cation ondepolarization
Spermine enhanced excitability by increasing inwardrecti¢cation on depolarization, without greatly a¡ectingthe passive properties of MGB neurons. It is not knownif the passive and active membrane properties of MGBneurons mature by P14 in gerbil, as in the rat (Tennig-keit et al., 1998). Thalamocortical neurons of the adultguinea pig and P7^P28 rat inwardly rectify because the
activation of persistent Na" conductance on depolariza-tion results in an ampli¢cation of the voltage response(Jahnsen and Llinas, 1984; Tennigkeit et al., 1996; Par-ri and Crunelli, 1998). In the present studies, blockadeof the TTX-sensitive recti¢cation or NMDA receptorseliminated the spermine-induced enhancement of recti-¢cation on depolarization. These ¢ndings imply thatspermine interactions with NMDA receptors led to ac-tivation of a persistent Na" conductance in MGB neu-rons.
An elevation in intracellular Ca2" concentration([Ca2"]i) due to NMDA receptor activation (Jahr,1992) may have enhanced recti¢cation on depolariza-tion. We did not observe a spermine-induced increasein voltage responses on depolarization during Ca2"-freeperfusion or rapid chelation of Ca2" with intracellularBAPTA. It seems likely that an elevation of [Ca2"]iinitiated by spermine actions at NMDA receptors acti-vated intracellular messengers and increased this recti-fying behavior. In neocortical neurons, transmitter ac-tivation of dendritic NMDA receptors increases Ca2"
entry (Schwindt and Crill, 1995) that may increasechannel phosphorylation (Siekevitz, 1991) and a persis-tent Na" conductance (Schwindt et al., 1992). Hence,the spermine-induced enhancement of TTX-sensitiverecti¢cation on depolarization may result from NMDA-mediated Ca2" entry in MGB neurons.
The e¡ects of spermine on membrane recti¢cationand ¢ring threshold may involve the recruitment of aCa2"-dependent second messenger, subsequent toNMDA receptor activation. Activation of NMDA re-ceptors enhances Ca2" entry, resulting in a Ca2" gra-dient in the dendrites (Connor et al., 1988) and activa-tion of a protein kinase C (PKC) pathway. A rise inintracellular [Ca2"] also may activate calmodulin kinaseII which enhances Na" currents (Carlier et al., 2000).PKC activation increases membrane excitability byshifting the activation curve for the persistent Na" cur-rent along the voltage axis to more hyperpolarized po-tentials (Astman et al., 1998; Franceschetti et al., 2000).This voltage range is consistent with the range of sper-mine enhancement of voltage recti¢cation in our experi-ments.
The increased recti¢cation on depolarization mayhave reduced the threshold for an action potential inMGB neurons (cf. neocortical neurons, Stafstrom et al.,1982). Antagonism of NMDA receptors, perfusion withCa2"-free ACSF or rapid chelation of Ca2" with BAP-TA, eliminated the reduction in threshold and increasedtonic ¢ring due to spermine application. Hence, themodulation of NMDA receptor-mediated Ca2" entrylikely increased membrane recti¢cation on depolariza-tion and reduced ¢ring threshold. This mechanism ex-plains the ability of spermine to increase postsynapticexcitability and tonic ¢ring in MGB neurons.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^7976
4.3. Facilitation of low threshold Ca2+ spike (LTS)¢ring
Spermine facilitated LTS ¢ring by a mechanism thatdid not involve interactions with NMDA receptors.Spermine increased the rate of rise and amplitude ofthe LTS, despite APV blockade of NMDA receptors.This was evident on depolarization to action potentialthreshold where there is a smaller gradient for Ca2" aswell as greater inactivation of T-type Ca2" channels(Hernandez-Cruz and Pape, 1989). Spermine enhancedthe LTS during blockade of voltage-dependent Na"
channels by TTX. Hence, a change in some parameterof the T-type Ca2" current, e.g. voltage dependence ofthe inactivation^activation relationship, may have in-creased the LTS.
4.4. Signi¢cance
Spermine is widely distributed in rat and humanbrain (Harman and Shaw, 1981; Morrison et al.,1995). A membrane transporter appears to maintainlow extracellular concentrations of 6 1 WM (Dot etal., 2000). These concentrations may increase onNMDA stimulation to s 50 WM in striatal neurons ofadult rat brain (Fage et al., 1992). The e¡ects of sper-mine on NMDA receptors and low threshold Ca2"
spikes in juvenile MGB neurons (ED50 =V100 WM)are consistent with neuromodulatory actions at highmicromolar concentrations (Williams, 1997). Giventhe role of NMDA receptors during development,such modulation by spermine is likely important forlearning processes (Chida et al., 1992).
The present results are relevant to the normal func-tion of the central auditory system. The NMDA recep-tor-mediated e¡ects of spermine would enhance theability of MGB neurons to detect simultaneous inputs,as in coincidence detection. For example, an overex-pression of the spermine-sensitive NR2B subunit (Wil-liams et al., 1994) prolongs EPSPs and shortens thetime window between two coincident signals in hippo-campal neurons (Tang et al., 1999). In thalamic neu-rons, the generation of synchronous activity may in-volve coincidence detection (Roy and Alloway, 2001)as well as amplitude selectivity in the MGB neurons(Kuwabara and Suga, 1993).
The e¡ects of spermine on the low threshold Ca2"
spikes of MGB neurons may have relevance for con-scious or sleep states and disorders of consciousness.The LTS is essential in the generation of bursting andoscillatory activity in the auditory nuclei (Hu, 1995;Tennigkeit et al., 1996). By increasing the rate of riseand amplitude of the LTS and slowing its decay, sper-mine modulation may increase an MGB neuron’s re-sponsiveness of neurons at hyperpolarized potentials
(Hu et al., 1994) to inputs during these states (He andHu, 2002). Modulation by spermine may have impor-tance for bursting behavior during sleep states whereasexcessive modulation may occur in absence epilepsy asin audiogenic seizures (Porta et al., 1981), sensitive toblockade by polyamine antagonists (Kotlinska and Lil-jequist, 1996).
The present study has shown that spermine enhancedthe excitability of thalamocortical neurons in speci¢cways that were consistent with a neuromodulator rolein the medial geniculate body at P13^P15 stage of de-velopment. Spermine acted on a polyamine site ofNMDA receptors, to increase membrane recti¢cationon depolarization, reduce ¢ring threshold and to slowthe decay of corticothalamic EPSPs in MGB neurons.The heightened excitability increased tonic ¢ring evokedby depolarizing current pulses and EPSP bursts of ac-tion potentials. Spermine also increased the rates of riseand amplitudes of low threshold Ca2" spikes by anunknown mechanism, not mediated by NMDA recep-tors. By increasing the e⁄cacy of corticothalamic exci-tation, spermine actions have importance in the trans-formation of auditory signals to tonic and burst ¢ringduring the juvenile stage of development.
Acknowledgements
The authors thank Dr. D.W.F. Schwarz for adviceon the initial experiments and Dr. D.A. Mathers forkindly reviewing earlier versions of the manuscript.We gratefully acknowledge ¢nancial grant support ofthe Mathematics in Information Technology and Com-plex Systems to I.R., the Natural Sciences and Engi-neering Research Council of Canada to R.M.M., andthe Canadian Institutes for Health Research to E.P. Wealso appreciate the excellent technical support of Chris-tian Caritey.
References
Astman, N., Gutnick, M.J., Fleidervish, I.A., 1998. Activation ofprotein kinase C increases neuronal excitability by regulating per-sistent Na" current in mouse neocortical slices. J. Neurophysiol.80, 1547^1551.
Bartlett, E.L., Smith, P.H., 1999. Anatomic, intrinsic, and synapticproperties of dorsal and ventral division neurons in rat medialgeniculate body. J. Neurophysiol. 81, 1999^2016.
Benveniste, M., Mayer, M.L., 1993. Multiple e¡ects of spermine onN-methyl-D-aspartic acid receptor responses of rat cultured hippo-campal neurones. J. Physiol. 464, 131^163.
Carlier, E., Dargent, B., De Waard, M., Couraud, F., 2000. Na"
channel regulation by calmodulin kinase II in rat cerebellar gran-ule cells. Biochem. Biophys. Res. 274, 392^399.
Chen, C., Regehr, W.G., 2000. Developmental remodeling of the ret-inogeniculate synapse. Neuron 28, 955^966.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^79 77
Chida, N., Saito, H., Abe, K., 1992. Spermine facilitates the gener-ation of long-term potentiation of evoked potential in the dentategyrus of anesthetized rats. Brain Res. 593, 57^62.
Connor, J., Wadman, W.J., Hockberger, P.E., Wong, R.K., 1988.Sustained dendritic gradients of Ca2" induced by excitatory aminoacids in CA1 hippocampal neurons. Science 240, 649^653.
Crill, W.E., 1996. Persistent sodium current in mammalian centralneurons. Annu. Rev. Physiol. 58, 349^362.
Dot, J., Lluch, M., Blanco, I., Rodriguez-Alvarez, J., 2000. Polyamineuptake in cultures astrocytes: characterization and modulation byprotein kinases. J. Neurochem. 75, 1917^1926.
Fage, D., Voltz, C., Scatton, B., Carter, C., 1992. Selective release ofspermine and spermidine from the rat striatum by N-methyl-D-aspartate receptor activation in vivo. J. Neurochem. 58, 2170^2175.
Faingold, C.L., Millan, M.H., Boersma Anderson, C.A., Meldrum,B.S., 1989. Induction of audiogenic seizures in normal and genet-ically epilepsy-prone rats following focal microinjection of an ex-citant amino acid into reticular formation and auditory nuclei.Epilepsy Res. 3, 199^205.
Franceschetti, S., Taverna, G., Sancini, G., Panzica, F., Lombardi,R., Avanzini, G., 2000. Protein kinase C-dependent modulation ofNa+ currents increases the excitability of rat neocortical pyramidalneurons. J. Physiol. 528, 291^304.
Gonzalez-Lima, F., Scheich, H., 1984. Functional activation in theauditory system of the rat produced by arousing reticular stimu-lation: a 2-deoxyglucose study. Brain Res. 299, 201^214.
Harman, R.J., Shaw, G.G., 1981. High-a⁄nity uptake of spermine byslices of rat cerebral cortex. J. Neurochem. 36, 1609^1615.
He, J., Hu, B., 2002. Di¡erential distribution of burst and single-spike responses in auditory thalamus. J. Neurophysiol. 88, 2152^2156.
Heierli, P., de Ribaupierre, F., de Ribaupierre, Y., 1987. Functionalproperties and interactions of neuron pairs simultaneously re-corded in the medial geniculate body of the cat. Hear. Res. 25,209^225.
Hernandez-Cruz, A., Pape, H.C., 1989. Identi¢cation of two calciumcurrents in acutely dissociated neurons from the rat lateral genic-ulate nucleus. J. Neurophysiol. 61, 1270^1283.
Hsieh, C.Y., Leslie, F.M., Metherate, R., 2002. Nicotine exposureduring a postnatal critical period alters NR2A and NR2BmRNA expression in rat auditory forebrain. Brain Res. Dev.Brain Res. 133, 19^25.
Hu, B., 1995. Cellular basis of temporal synaptic signalling: an invitro electrophysiological study in rat auditory thalamus. J. Phys-iol. 483, 167^182.
Hu, B., Senatorov, V., Mooney, D., 1994. Lemniscal and non-lemnis-cal synaptic transmission in rat auditory thalamus. J. Physiol. 479,217^231.
Jacobsen, R.B., Ulrich, D., Huguenard, J.R., 2001. GABA(B) andNMDA receptors contribute to spindle-like oscillations in rat thal-amus in vitro. J. Neurophysiol. 86, 1365^1375.
Jahnsen, H., Llinas, R., 1984. Ionic basis for the electro-responsive-ness and oscillatory properties of guinea-pig thalamic neurons invitro. J. Physiol. 349, 227^247.
Jahr, C.E., 1992. High probability opening of NMDA receptor chan-nels by L-glutamate. Science 255, 470^472.
Kew, J.N., Kemp, J.A., 1998. An allosteric interaction between theNMDA receptor polyamine and ifenprodil sites in rat culturedcortical neurons. J. Physiol. 512, 17^28.
Khan, A.M., Stanley, B.G., Bozzetti, L., Chin, C., Stivers, C., Curras-Collazo, M.C., 2000. N-methyl-D-aspartate receptor subunitNR2B is widely expressed throughout the rat diencephalon: animmunohistochemical study. J. Comp. Neurol. 428, 428^449.
Koerner, C., Danober, L., Boehrer, A., Marescaux, C., Vergnes, M.,
1996. Thalamic NMDA transmission in a genetic model of ab-sence epilepsy in rats. Epilepsy Res. 25, 11^19.
Kraus, N., Smith, D.I., McGee, T., Stein, L., Cartee, C., 1987. Devel-opment of the middle latency response in an animal model and itsrelation to the human response. Hear. Res. 27, 165^176.
Kuwabara, N., Suga, N., 1993. Delay lines and amplitude selectivityare created in subthalamic auditory nuclei: the brachium of theinferior colliculus of the mustached bat. J. Neurophysiol. 69,1713^1724.
McCann, P.P., Pegg, A.E., 1992. Ornithine decarboxylase as an en-zyme target for therapy. Pharmacol. Ther. 54, 195^215.
Metherate, R., Cruikshank, S.J., 1999. Thalamocortical inputs triggera propagating envelope of gamma-band activity in auditory cortexin vitro. Exp. Brain Res. 126, 160^174.
Morel, A., Imig, T.J., 1987. Thalamic projections to ¢elds A, AI, P,and VP in the cat auditory cortex. J. Comp. Neurol. 265, 119^144.
Morrison, L.D., Becker, L., Ang, L.C., Kish, S.J., 1995. Polyaminesin human brain: regional distribution and in£uence of aging.J. Neurochem. 65, 636^642.
N’Gouemo, P., Faingold, C.L., 1997. Audiogenic kindling increasesneuronal responses to acoustic stimuli in neurons of the medialgeniculate body of the genetically epilepsy-prone rat. Brain Res.761, 217^224.
Parri, H.R., Crunelli, V., 1998. Sodium current in rat and cat thala-mocortical neurons: role of a non-inactivating component in tonicand burst ¢ring. J. Neurosci. 18, 854^867.
Porta, R., Doyle, R.L., Tatter, S.B., Wilens, T.E., Schatz, R.A., Sell-inger, O.Z., 1981. The biosynthesis of polyamines in the brain ofaudiogenic seizure-susceptible and -resistant deermice. J. Neuro-chem. 37, 723^729.
Pritchard, G.A., Fahey, J.M., Minocha, S.C., Conaty, C., Miller,L.G., 1994. Polyamine potentiation and inhibition of NMDA-mediated increases of intracellular free Ca2" in cultured chick cor-tical neurons. Eur. J. Pharmacol. 266, 107^115.
Reynolds, I.J., 1990. Arcaine uncovers dual interactions of poly-amines with the N-methyl-D-aspartate receptor. J. Pharmacol.Exp. Ther. 255, 1001^1007.
Roberts, E.B., Ramoa, A.S., 1999. Enhanced NR2A subunit expres-sion and decreased NMDA receptor decay time at the onset ofocular dominance plasticity in the ferret. J. Neurophysiol. 81,2587^2591.
Roy, S.A., Alloway, K.D., 2001. Coincidence detection or temporalintegration? What the neurons in somatosensory cortex are doing.J. Neurosci. 21, 2462^2473.
Schweitzer, L., Casseday, J.H., Sjoerdsma, A., McCann, P.P., Barto-lome, J.V., 1986. Identi¢cation of polyamines in the cochlea of therat and their potential role in hearing. Brain Res. Bull. 16, 215^218.
Schwindt, P.C., Crill, W.E., 1995. Ampli¢cation of synaptic currentby persistent sodium conductance in apical dendrite of neocorticalneurons. J. Neurophysiol. 74, 2220^2224.
Schwindt, P.C., Spain, W.J., Crill, W.E., 1992. E¡ects of intracellularcalcium chelation on voltage-dependent and calcium-dependentcurrents in cat neocortical neurons. Neuroscience 47, 571^578.
Siekevitz, P., 1991. Possible role for calmodulin and the Ca2+/cal-modulin-dependent protein kinase II in postsynaptic neurotrans-mission. Proc. Natl. Acad. Sci. USA 88, 5374^5378.
Stafstrom, C.E., Schwindt, P.C., Crill, W.E., 1982. Negative slopeconductance due to a persistent subthreshold sodium current incat neocortical neurons in vitro. Brain Res. 236, 221^226.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^7978
Sucher, N.J., Akbarian, S., Chi, C.L., Leclerc, C.L., Awobuluyi, M.,Deitcher, D.L., Wu, M.K., Yuan, J.P., Jones, E.G., Lipton, S.A.,1995. Developmental and regional expression pattern of a novelNMDA receptor-like subunit (NMDAR-L) in the rodent brain.J. Neurosci. 15, 6509^6520.
Tang, Y.P., Shimizu, E., Dube, G.R., Rampon, C., Kerchner, G.A.,Zhuo, M., Liu, G., Tsien, J.Z., 1999. Genetic enhancement oflearning and memory in mice. Nature 401, 63^69.
Tennigkeit, F., Schwarz, D.W., Puil, E., 1996. Mechanisms for signaltransformation in lemniscal auditory thalamus. J. Neurophysiol.76, 3597^3608.
Tennigkeit, F., Schwarz, D.W., Puil, E., 1998. Postnatal developmentof signal generation in auditory thalamic neurons. Brain Res. Dev.Brain Res. 109, 255^263.
Williams, K., 1997. Interactions of polyamines with ion channels.Biochem. J. 325, 289^297.
Williams, K., Zappia, A.M., Pritchett, D.B., Shen, Y.M., Molino¡,P.B., 1994. Sensitivity of the N-methyl-D-aspartate receptor topolyamines is controlled by NR2 subunits. Mol. Pharmacol. 45,803^809.
HEARES 4047 22-1-03
I. Ran et al. /Hearing Research 176 (2003) 65^79 79