Spontaneous electrical and Ca2+ signals in themouse renal pelvis that drive pyeloureteric peristalsis
Richard J. Lang,* Hikaru Hashitani, † Mary A. Tonta, *
Justin L. Bourke,* Helena C. Parkington* & H ikaru Suzuki†
*Department of Physiology, School of Biomedical Sciences, Monash University, Clayton, VIC 3800, Australia and†Department of Cell Physiology, Nagoya City University, Graduate School of Medical Sciences, Nagoya 467-8601, Japan.
Summary
1. Peristalsis in the smooth muscle cell (SMC) wallof the pyeloureteric system is unique in physiology in thatthe primary pacemaker resides in a population of atypicalSMCs situated near the border of the renal papilla.
2. Atypical SMCs display high frequency Ca2+
transients upon the spontaneous release of Ca2+ fromIP3-dependent stores that trigger cation-selectivespontaneous transient depolarizations (STDs).Innifedipine, these Ca2+ transients and STDS seldompropagate >100 µm. Synchronization of STDs inneighbouring atypical SMCs into an electrical signal thatcan trigger action potential discharge and contraction in thetypical SMC layer involves a coupled oscillator mechanismdependent on Ca2+ entry through L-type voltage-operatedCa2+ channels.
3. A population of spindle- or stellate-shaped cells,immuno-positive for the tyrosine receptor kinase kit, aresparsely distributed throughout the pyeloureteric system.Ca2+ transients and action potentials of long durationoccurring at low frequencies are also recorded in apopulation of fusiform cells, which we have termedinterstitial cells of Cajal-like cells (ICC-like cells).
4. The electrical and Ca2+ signals in ICC-like cellsare abolished upon blockade of Ca2+ release from eitherIP3- or ryanodine-dependent Ca2+ stores. However, thespontaneous Ca2+ signals in atypical SMCs or ICC-likecells are little affected inW/W−v transgenic mice whichhave extensive lesions of their intestinal ICC networks.
5. In summary, we hav e developed a model ofpyeloureteric pacemaking in which atypical SMCs areindeed the primary pacemakers, the function of ICC-likecells has yet to be determined.
Pyeloureteric peristalsis
Since the earliest descriptions by Englemann (1869)1
of pyeloureteric peristalsis, it has been recognised that thespontaneous propagating contractions which transport urineexpressed in the kidney along the ureter to the bladderoriginate within the most proximal regions of the renalpelvis. Inmost mammals, including the mouse, the kidneycontains a single papilla which is surrounded by a funnel-shaped calyx or renal pelvis (Figure 1A). This renal pelvisconsists of a urothelium-lined lumen and a plexus of long,spindle-shaped ‘typical’ smooth muscle cells (SMCs)which form an inner layer, originating near the base of the
papilla and extending through into the ureter.2,3 Thedensity of typical SMCs increases with distance from thepapilla base resulting in a gradual thickening of the pelvicwall.3-5 In human and pig, the kidney is multi-papillate sothat a number of major and minor calyces fuse to form aseparate renal pelvis which extends to the ureter.2-4,6 Eventhough there is evidence of extensive networks ofparasympathetic and sympathetic nerves within the wall ofthe pelviureteric system,7 it has been difficult todemonstrate that these networks play any efferent role inmaintaining or modulating pyeloureteric peristalsis.Electrical nerve stimulation evokes in the guinea pig renalpelvis a transient increase in the amplitude and frequency ofthe spontaneous contractions which is followed by aprolonged negative inotropic effect that are little affected byNG-nitro-L-arginine, guanethidine or atropine,1,8,9 but hav ebeen attributed to the release of tachykinins and calcitoningene related peptide (CGRP), respectively, from sensorynerves.9,10
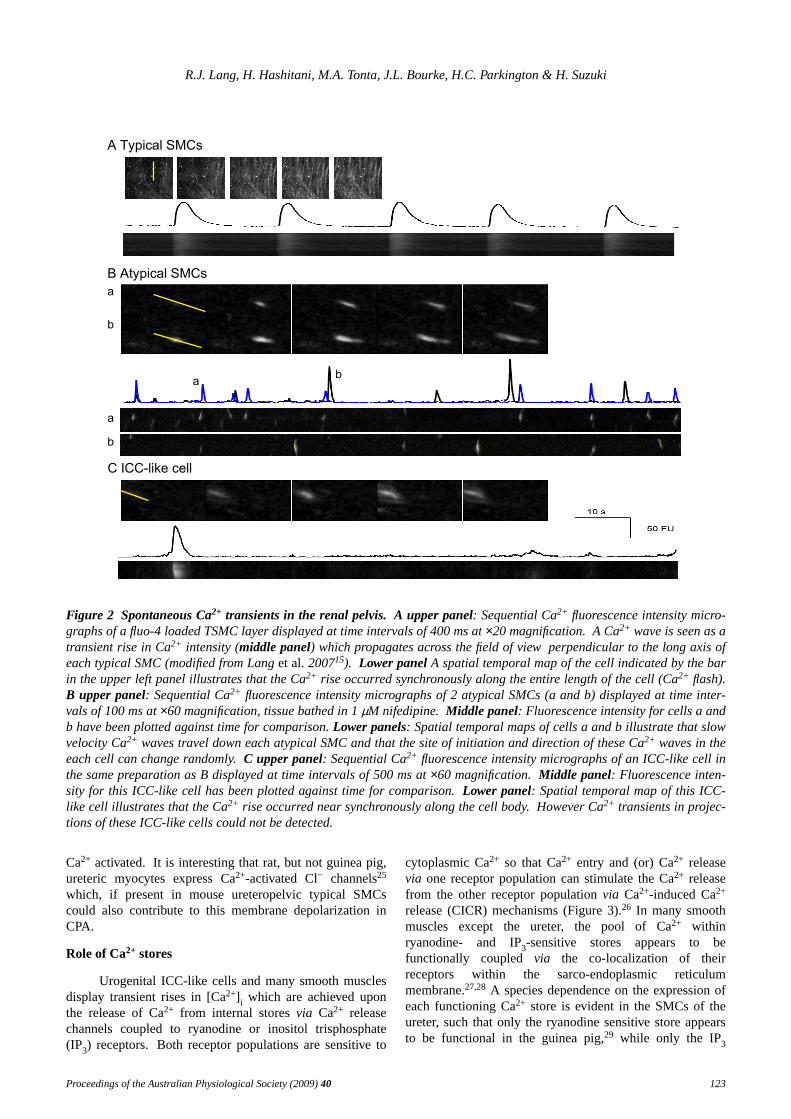
The recent use of intracellular microelectroderecording techniques (Figure 1Bi) and fluorescenceindicators of Ca2+ concentration (Figure 2A) haveunequivocally established that nifedipine-sensitive actionpotentials and Ca2+ waves within the smooth muscle wallare responsible for the peristaltic contractions thatpropagate the length of the upper urinary system.5,11-15Thespatial temporal map in the lower panel of Figure 2A alsoillustrates that rises of Ca2+ occur instantaneously(‘flashes’) along the entire length of each typical SMCinsitu and that propagation of the Ca2+ wave occurs in thedirection perpendicular to the long axis of individual typicalSMCs (Figure 2A upper panel).
Pacemaking in the pyleoureteric system
In both uni- and multi-calyceal kidneys, a singlepacemaker region at any one time on the papilla-calycealborder initiates the wav e of contraction, which conductsradially across the pelvis to form a crescent shaped wav ethat then conducts distally. In uni-papillate kidneys, thepacemaker region shifts spontaneously along the pelvi-calyceal border; in multi-papillate kidneys this ‘primary’pacemaker shifts spontaneously between calyces.16,17
Atypical SMCs, which are shorter in their long axisthan typical SMCs, form a relatively sparse outer layer inuni-calyceal kidneys and an inner layer in multi-calycealkidneys that wraps around the most proximal regions of thepapilla and terminates at the pelviureteric junction.2,3,5,6,18
Proceedings of the Australian Physiological Society (2009)40 121
Autorhythmicity in the mouse pyeloureteric system
���
��� �����
�� ���µ�
�� �
��
�
��
��
��
����
����������������
����������
��
����
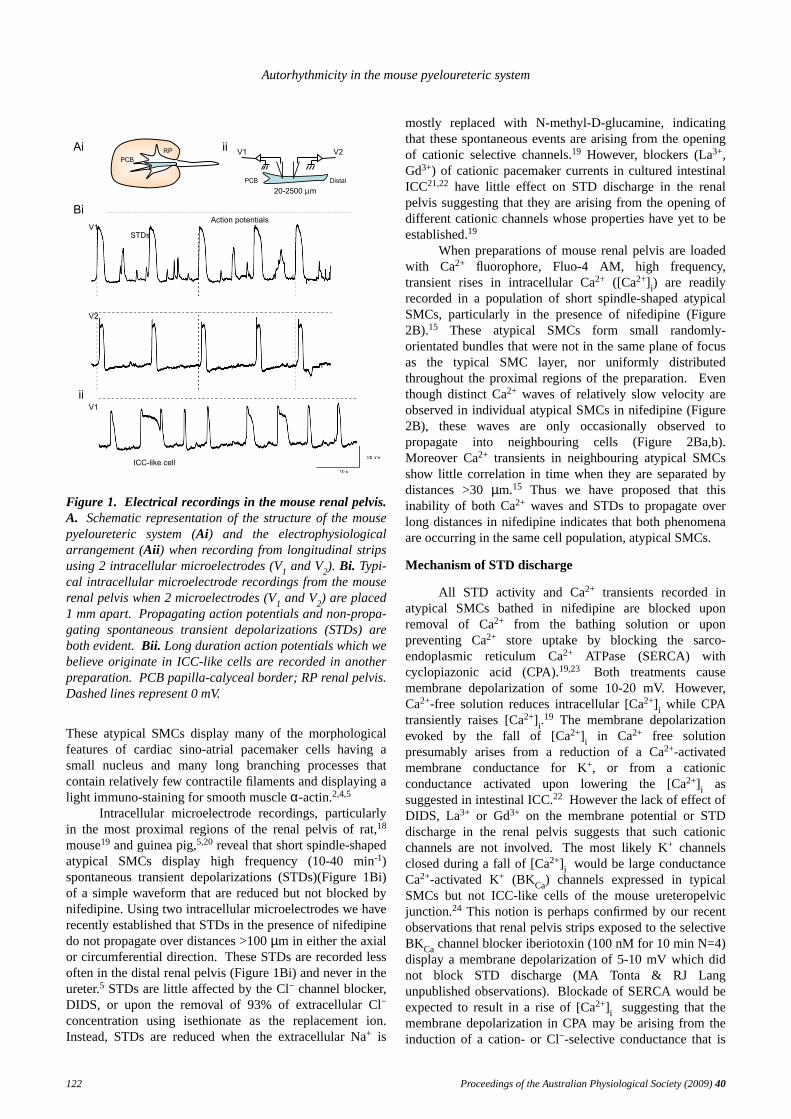
Figure 1. Electrical recordings in the mouse renal pelvis.A. Schematic representation of the structure of the mousepyeloureteric system (Ai) and the electrophysiologicalarrangement (Aii) when recording from longitudinal stripsusing 2 intracellular microelectrodes (V1 and V2). Bi. Typi-cal intracellular microelectrode recordings from the mouserenal pelvis when 2 microelectrodes (V1 and V2) are placed1 mm apart. Propagating action potentials and non-propa-gating spontaneous transient depolarizations (STDs) areboth evident. Bii. Long duration action potentials which webelieve originate in ICC-like cells are recorded in anotherpreparation. PCBpapilla-calyceal border; RP renal pelvis.Dashed lines represent 0 mV.
These atypical SMCs display many of the morphologicalfeatures of cardiac sino-atrial pacemaker cells having asmall nucleus and many long branching processes thatcontain relatively few contractile filaments and displaying alight immuno-staining for smooth muscleα-actin.2,4,5
Intracellular microelectrode recordings, particularlyin the most proximal regions of the renal pelvis of rat,18
mouse19 and guinea pig,5,20 reveal that short spindle-shapedatypical SMCs display high frequency (10-40 min-1)spontaneous transient depolarizations (STDs)(Figure 1Bi)of a simple wav eform that are reduced but not blocked bynifedipine. Using two intracellular microelectrodes we haverecently established that STDs in the presence of nifedipinedo not propagate over distances >100µm in either the axialor circumferential direction.These STDs are recorded lessoften in the distal renal pelvis (Figure 1Bi) and never in theureter.5 STDs are little affected by the Cl− channel blocker,DIDS, or upon the removal of 93% of extracellular Cl−
concentration using isethionate as the replacement ion.Instead, STDs are reduced when the extracellular Na+ is
mostly replaced with N-methyl-D-glucamine, indicatingthat these spontaneous events are arising from the openingof cationic selective channels.19 However, blockers (La3+,Gd3+) of cationic pacemaker currents in cultured intestinalICC21,22 have little effect on STD discharge in the renalpelvis suggesting that they are arising from the opening ofdifferent cationic channels whose properties have yet to beestablished.19
When preparations of mouse renal pelvis are loadedwith Ca2+ fluorophore, Fluo-4 AM, high frequency,transient rises in intracellular Ca2+ ([Ca2+] i) are readilyrecorded in a population of short spindle-shaped atypicalSMCs, particularly in the presence of nifedipine (Figure2B).15 These atypical SMCs form small randomly-orientated bundles that were not in the same plane of focusas the typical SMC layer, nor uniformly distributedthroughout the proximal regions of the preparation.Eventhough distinct Ca2+ waves of relatively slow velocity areobserved in individual atypical SMCs in nifedipine (Figure2B), these wav es are only occasionally observed topropagate into neighbouring cells (Figure 2Ba,b).Moreover Ca2+ transients in neighbouring atypical SMCsshow little correlation in time when they are separated bydistances >30µm.15 Thus we have proposed that thisinability of both Ca2+ waves and STDs to propagate overlong distances in nifedipine indicates that both phenomenaare occurring in the same cell population, atypical SMCs.
Mechanism of STD discharge
All STD activity and Ca2+ transients recorded inatypical SMCs bathed in nifedipine are blocked uponremoval of Ca2+ from the bathing solution or uponpreventing Ca2+ store uptake by blocking the sarco-endoplasmic reticulum Ca2+ ATPase (SERCA) withcyclopiazonic acid (CPA).19,23 Both treatments causemembrane depolarization of some 10-20 mV. Howev er,Ca2+-free solution reduces intracellular [Ca2+] i while CPAtransiently raises [Ca2+] i.
19 The membrane depolarizationev oked by the fall of [Ca2+] i in Ca2+ free solutionpresumably arises from a reduction of a Ca2+-activatedmembrane conductance for K+, or from a cationicconductance activated upon lowering the [Ca2+] i assuggested in intestinal ICC.22 However the lack of effect ofDIDS, La3+ or Gd3+ on the membrane potential or STDdischarge in the renal pelvis suggests that such cationicchannels are not involved. Themost likely K+ channelsclosed during a fall of [Ca2+] i would be large conductanceCa2+-activated K+ (BKCa) channels expressed in typicalSMCs but not ICC-like cells of the mouse ureteropelvicjunction.24 This notion is perhaps confirmed by our recentobservations that renal pelvis strips exposed to the selectiveBKCa channel blocker iberiotoxin (100 nM for 10 min N=4)display a membrane depolarization of 5-10 mV which didnot block STD discharge (MA Tonta & RJ Langunpublished observations). Blockadeof SERCA would beexpected to result in a rise of [Ca2+] i suggesting that themembrane depolarization in CPA may be arising from theinduction of a cation- or Cl−-selective conductance that is
122 Proceedings of the Australian Physiological Society (2009)40
R.J. Lang, H. Hashitani, M.A. Tonta, J.L. Bourke, H.C. Parkington & H. Suzuki
����������� �
�� �������
�
�
�
���������� �
Figure 2 Spontaneous Ca2+ transients in the renal pelvis. A upper panel: Sequential Ca2+ fluorescence intensity micro-graphs of a fluo-4 loaded TSMC layer displayed at time intervals of 400 ms at×20 magnification. ACa2+ wave is seen as atransient rise in Ca2+ intensity (middle panel) which propagates across the field of view perpendicular to the long axis ofeach typical SMC (modified from Langet al.200715). Lower panel A spatial temporal map of the cell indicated by the barin the upper left panel illustrates that the Ca2+ rise occurred synchronously along the entire length of the cell (Ca2+ flash).B upper panel: Sequential Ca2+ fluorescence intensity micrographs of 2 atypical SMCs (a and b) displayed at time inter-vals of 100 ms at×60 magnification, tissue bathed in 1µM nifedipine. Middle panel: Fluorescence intensity for cells a andb have been plotted against time for comparison.Lower panels: Spatial temporal maps of cells a and b illustrate that slowvelocity Ca2+ waves travel down each atypical SMC and that the site of initiation and direction of these Ca2+ waves in theeach cell can change randomly. C upper panel: Sequential Ca2+ fluorescence intensity micrographs of an ICC-like cell inthe same preparation as B displayed at time intervals of 500 ms at×60 magnification. Middle panel: Fluorescence inten-sity for this ICC-like cell has been plotted against time for comparison.Lower panel: Spatial temporal map of this ICC-like cell illustrates that the Ca2+ rise occurred near synchronously along the cell body. However Ca2+ transients in projec-tions of these ICC-like cells could not be detected.
Ca2+ activated. It is interesting that rat, but not guinea pig,ureteric myocytes express Ca2+-activated Cl− channels25
which, if present in mouse ureteropelvic typical SMCscould also contribute to this membrane depolarization inCPA.
Role of Ca2+ stores
Urogenital ICC-like cells and many smooth musclesdisplay transient rises in [Ca2+] i which are achieved uponthe release of Ca2+ from internal storesvia Ca2+ releasechannels coupled to ryanodine or inositol trisphosphate(IP3) receptors. Bothreceptor populations are sensitive to
cytoplasmic Ca2+ so that Ca2+ entry and (or) Ca2+ releasevia one receptor population can stimulate the Ca2+ releasefrom the other receptor populationvia Ca2+-induced Ca2+
release (CICR) mechanisms (Figure 3).26 In many smoothmuscles except the ureter, the pool of Ca2+ withinryanodine- and IP3-sensitive stores appears to befunctionally coupled via the co-localization of theirreceptors within the sarco-endoplasmic reticulummembrane.27,28 A species dependence on the expression ofeach functioning Ca2+ store is evident in the SMCs of theureter, such that only the ryanodine sensitive store appearsto be functional in the guinea pig,29 while only the IP3
Proceedings of the Australian Physiological Society (2009)40 123
Autorhythmicity in the mouse pyeloureteric system
receptor-sensitive Ca2+ store can be detected in the rat.29,30
Blockers of, phospholipase C (PLC) function and IP3formation (neomycin, U73122, xestospongin C) orIP3receptors (2-APB and heparin) have all been shown toblock or reduce electrical oscillations in urogenital23,31 andgastrointestinal32-35 preparations. However, the mostcompelling evidence establishing the essential role ofIP3-sensitive Ca2+ release channels has come from theabsence of slow wav es in mice which lack IP3 type 1receptors in their smooth muscle.36 In our experiments,STDs and Ca2+ transient discharge in atypical SMCs areblocked by 2-APB and reduced by blockers of PLCfunction, both in a manner associated with membranedepolarization of some 5-10 mV.19
2-APB has been suggested to both deplete Ca2+ storesby blocking store-operated channels without affecting IP3dependent Ca2+ release37 and block SERCA in a mannersimilar to CPA.38 However, the disconnect betweenmembrane potential and Ca2+ levels seen with 2-APB, whencompared with CPA, and the lack of effect of TRP channelsblockers in our hands suggest that 2-APB is indeedblocking IP3 receptors. These observations are consistentwith our previous notion that the intrinsic release ofprostaglandins and tachykinins provides a constantexcitatory PLC drive of IP3 production in our pacemakercells, which is essential in maintaining pelviuretericperistalsis.39,40
Blockade of ryanodine receptors, usingconcentrations (<1 mM) thought to block ryanodine-sensitive Ca2+ release channels without causing storedepletion,41 abolishes spontaneous electrical and Ca2+
signalling in ICC-like cells in the rabbit urethra42 andguinea-pig corporal tissue,43 but not the spontaneouselectrical activity in portions of the mouse small intestine44
or guinea renal pelvis.23 In mouse renal pelvis, theamplitude and ½ width of STD recordedin situ aresignificantly reduced to 582% and 746%, respectively, after20 min exposure to 100µM ryanodine. However the Ca2+
transients recorded in individual atypical SMCsin situ arenot significantly affected, even after 30-60 min exposure.19
Model of pyeloureteric pacemaking
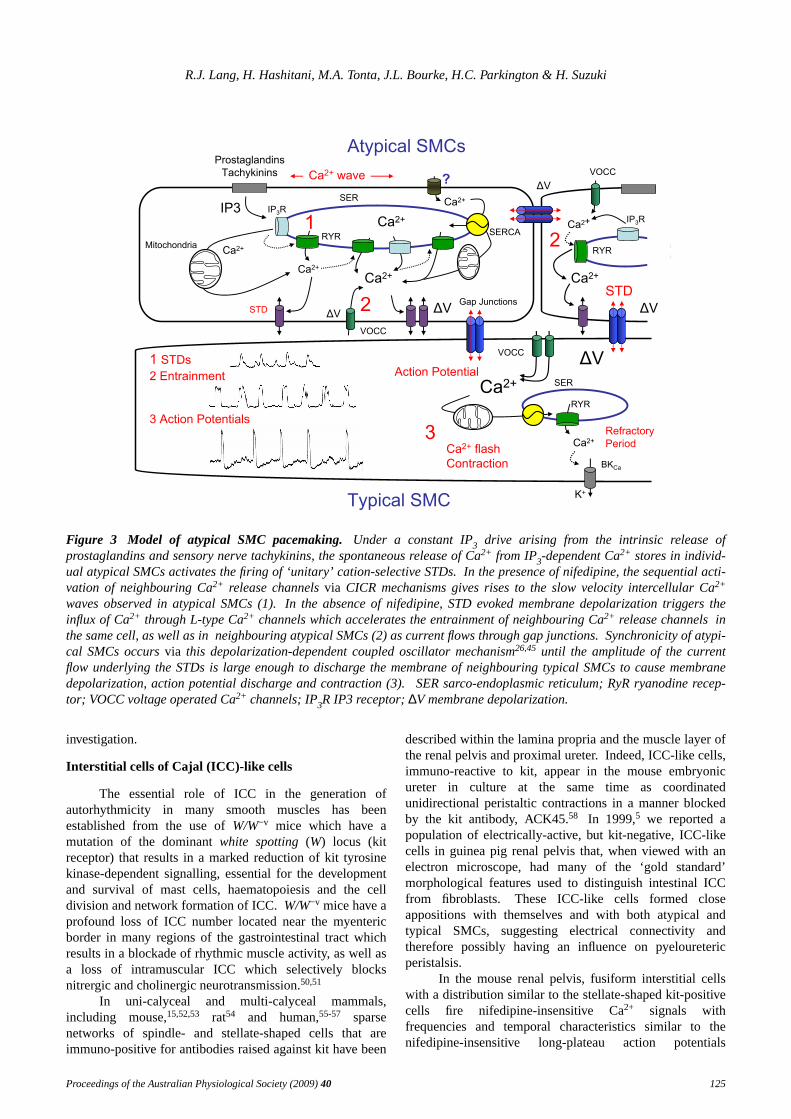
We hav einterpreted our data to suggest that STDs arelikely to represent the summation of the electrical activitygenerated by more than one atypical SMC.We envisagethat, in the presence of nifedipine, the spontaneous releaseof Ca2+ from IP3-dependent Ca2+ stores in individualatypical SMCs underlies the generation of the Ca2+
transients and ‘unitary’ STDs, while the subsequentactivation of neighbouring Ca2+ release channels and CIRCmechanisms represent the fundamental mechanismunderlying the slow velocity Ca2+ waves (Figure 3).26
In the absence of nifedipine, STD discharge in anindividual atypical SMC induces the entry of Ca2+ through‘L-type’ voltage-operated Ca2+ channels (VOCCs) whichleads to the accelerated activation (entrainment) ofneighbouring IP3-primed Ca2+ stores both within the samecell and in neighbouring atypical SMCs (Figure 3).This
depolarization-dependent entrainment couples neighbouringatypical SMCs within a pacemaker bundle to fire (releaseCa2+ and generate STDs) synchronouslyvia a coupledoscillator mechanism (as described in the stomach by vanHelden & Imtiaz45) to create a depolarizing signalsufficiently rapid and large enough to propagate into thetypical SMC layer and trigger action potential dischargeand contraction (Figure 3).The atypical SMC bundle firingSTDs at the highest frequency would presumably driveneighbouring atypical and typical smooth muscle bundlesso that only one pacemaker region would be apparent at anyone time. However, as each atypical SMC is capable ofeither initiating or contributing to an entrainment signalgenerated by its neighbours, the site of the dominantpacemaker region can spontaneously move within the mostproximal region of the uni-calyceal renal pelvis, evenbetween the minor calyces in multi-calyceal kidneys.26,45
Action potential discharge in the SMC wall of thepyeloureteric system is characterized by a prolongedrefractory period of many seconds so that contractionfrequency of the muscle wall is generally ¼ to ½ of thefrequency of atypical SMC STD discharge (10-40min-1).5,11,46 Moreover, the frequency of contraction ofcircumferentially-cut muscle strips decreases with distancefrom the base of papilla12,47,48 which we have previouslycorrelated with an increasing ratio of typical SMCs,compared to atypical SMCs.5 This difference infrequencies between atypical and typical SMCs has recentlybeen explained in terms of the refractory mechanismswithin typical SMCs that are dependent on the state of theinternal Ca stores.11,26 It has been suggested that Ca2+ entrythrough VOCCs during typical SMC action potentialdischarge and contraction is sequestered into theendoplasmic reticulum, which, once loaded, sensitizes itsryanodine receptors to generate Ca2+ sparks. Thisreleaseof stored Ca2+, in turn, activates plasmalemmal BKCachannels which hyperpolarize the membrane of the typicalSMCs creating the refractory periods observed. Ca2+
release declines as the Ca2+ levels in the store returns tonormal which results in a decreased BKCa channel activityand a repolarization of the membrane into a voltage rangethat is responsive to the STD drive coming fromneighbouring atypical SMCs (Figure 3).26
The role of the mitochondria in the generation of theatypical SMC STDs or in the control of smooth musclecontraction remains unclear. Whether the mitochondriaplay a passive role, temporarily and rapidly storing Ca2+
before slowly releasing it so that it can be taken up into theendoplasmic reticulum by SERCA,26 or whether it has amore active inv olvement, lowering Ca2+ concentrations inmicro-domains near the plasmalemma membrane so thatthe cationic conductance underlying STD generation isactually activated by a fall33,49 in Ca2+ has yet to beestablished. However, preliminary experiments havedemonstrated that disruption of the mitochondrial functionwith CCCP (1µM) abolishes Ca2+ transients in the typicalSMC layer in the absence of nifedipine (H.Hashitaniunpublished observations). Therole of the mitochondria inatypical and typical SMC function is under intensive
124 Proceedings of the Australian Physiological Society (2009)40
R.J. Lang, H. Hashitani, M.A. Tonta, J.L. Bourke, H.C. Parkington & H. Suzuki
IP3
Ca2+
2 Entrainment
Ca2+
RYR
RYR
IP3R
Ca2+
VOCC
Prostaglandins
Tachykinins
Atypical SMCs
Typical SMC
SERCA
Gap Junctions
1 STDs
Ca2+ flash
Contraction
1Mitochondria
SER
3 Action Potentials
Ca2+
RYR
V V
Ca2+
V
V
STD
STD
Action PotentialSER
2
3
2
V
IP3R
VOCC
K+
Refractory
Period
BKCa
Ca2+
Ca2+
VOCC?
Ca2+
Ca2+
Ca2+ wave
Figure 3 Model of atypical SMC pacemaking. Under a constant IP3 drive arising from the intrinsic release ofprostaglandins and sensory nerve tachykinins, the spontaneous release of Ca2+ from IP3-dependent Ca2+ stores in individ-ual atypical SMCs activates the firing of ‘unitary’ cation-selective STDs. In the presence of nifedipine, the sequential acti-vation of neighbouring Ca2+ release channelsvia CICR mechanisms gives rises to the slow velocity intercellular Ca2+
waves observed in atypical SMCs (1). In the absence of nifedipine, STD evoked membrane depolarization triggers theinflux of Ca2+ through L-type Ca2+ channels which accelerates the entrainment of neighbouring Ca2+ release channels inthe same cell, as well as in neighbouring atypical SMCs (2) as current flows through gap junctions.Synchronicity of atypi-cal SMCs occurs via this depolarization-dependent coupled oscillator mechanism26,45 until the amplitude of the currentflow underlying the STDs is large enough to discharge the membrane of neighbouring typical SMCs to cause membranedepolarization, action potential discharge and contraction (3). SER sarco-endoplasmic reticulum; RyR ryanodine recep-tor; VOCC voltage operated Ca2+ channels; IP3R IP3 receptor;∆V membrane depolarization.
investigation.
Interstitial cells of Cajal (ICC)-lik e cells
The essential role of ICC in the generation ofautorhythmicity in many smooth muscles has beenestablished from the use ofW/W−v mice which have amutation of the dominantwhite spotting(W) locus (kitreceptor) that results in a marked reduction of kit tyrosinekinase-dependent signalling, essential for the developmentand survival of mast cells, haematopoiesis and the celldivision and network formation of ICC.W/W−v mice have aprofound loss of ICC number located near the myentericborder in many regions of the gastrointestinal tract whichresults in a blockade of rhythmic muscle activity, as well asa loss of intramuscular ICC which selectively blocksnitrergic and cholinergic neurotransmission.50,51
In uni-calyceal and multi-calyceal mammals,including mouse,15,52,53 rat54 and human,55-57 sparsenetworks of spindle- and stellate-shaped cells that areimmuno-positive for antibodies raised against kit have been
described within the lamina propria and the muscle layer ofthe renal pelvis and proximal ureter. Indeed, ICC-like cells,immuno-reactive to kit, appear in the mouse embryonicureter in culture at the same time as coordinatedunidirectional peristaltic contractions in a manner blockedby the kit antibody, ACK45.58 In 1999,5 we reported apopulation of electrically-active, but kit-negative, ICC-likecells in guinea pig renal pelvis that, when viewed with anelectron microscope, had many of the ‘gold standard’morphological features used to distinguish intestinal ICCfrom fibroblasts. These ICC-like cells formed closeappositions with themselves and with both atypical andtypical SMCs, suggesting electrical connectivity andtherefore possibly having an influence on pyelouretericperistalsis.
In the mouse renal pelvis, fusiform interstitial cellswith a distribution similar to the stellate-shaped kit-positivecells fire nifedipine-insensitive Ca2+ signals withfrequencies and temporal characteristics similar to thenifedipine-insensitive long-plateau action potentials
Proceedings of the Australian Physiological Society (2009)40 125
Autorhythmicity in the mouse pyeloureteric system
recorded with intracellular microelectrodes (Figure 1Bii).15
However, we hav e not been able to demonstrate that theinterstitial cells pre-labelled with a kit antibody that binds atan extracellular site(s) are in fact the interstitial cellsdisplaying low frequency Ca2+ signals. Moreover, wecannot find any noticeable difference between the renalpelvis obtained from wild type andW/W−v mice,particularly in terms of their Ca2+ signalling, contractility orresponses to electrical nerve stimulation. Finally, thekidneys of W/W−v mice did not display any changes in theirshape or size, consistent with the lack of anyhydronephrosis in this phenotype (RJ Lang & H Hasihtaniunpublished observations).
Autorhythmicity in ICC-lik e cells
The major limitation of recording the membranepotential of the renal pelvis using intracellularmicroelectrodes is that ICC-like cells and atypical andtypical SMCs are likely to be in electrical continuity and,therefore, it is difficult to ascertain the cellular origin of ourelectrical signals.The infrequency of impaling an ICC-likecell with an intracellular microelectrode with confidence,rather than a cell electrically close to an ICC-like cell, hasprevented the examination of the effects of our modifiers ofCa2+ mobilization on their electrical behaviour. Howev er ithas been possible to examine the effects of these agents onthe low frequency Ca2+ transients in fusiform interstitialcells at the same time as our examination of the Ca2+
transients in atypical SMCs.15 In Figure 2C, it is evidentthat, in contrast to atypical SMCs, the Ca2+ signal in ICC-like cells appears as a long-lasting signal along the entirelength of the cell body. Interestingly, even though this Ca2+
transient in nifedipine is relatively larger and longer thanthe Ca2+ wave in atypical SMCs, it fails to propagate intoany neighbouring cells, be they other ICC-like cells oratypical and typical SMCs.In addition, even though someelectrical coupling between the cells firing the long lastingaction potentials and the typical SMC layer is often evident,we have not yet seen an individual interstitial cell trigger atypical SMC Ca2+ transient or muscle contraction.15
The agents described above that interrupted storeuptake and IP3-dependent release of Ca2+ in atypical SMCswere equally effective on Ca2+ signalling in ICC-like cellsin the mouse renal pelvis.However, there was a cleardifference in the action of ryanodine in atypical SMCs andICC-like cells within the same preparation. In comparisonto the relatively small effect on Ca2+ transients in atypicalSMCs in nifedipine, ryanodine completely blocked Ca2+
transient discharge in ICC-like cells within several minutesexposure.19
We hav epreviously suggested that ICC-like cells inguinea pig renal pelvis could well be acting as integrators ofthe atypical SMC pacemaker drive.5,8 In our presentexperiments, we have been unable to demonstrate anysynchronicity between neighbouring ICC-like cells, orbetween any atypical SMCs and ICC-like cells inpreparations bathed in 1µM nifedipine.15 This situationmay well be quite different in the absence of nifedipine
when Ca2+ influx through VOCCs could again be the agentof entrainment in ICC-like cells such that Ca2+ transientsgenerated in the cell body or neighbouring cells rapidlytravel down the interstitial cell projectionsvia CICR fromCa2+ release channels as described above.26
Our measurements of [Ca2+] i in cells of the renalpelvis are at a relatively low resolution so that we have notbeen able to detect Ca2+ transients in the stellate projectionsof our ICC-like cells. However, we envisage that the longprocesses of ICC-like cells could serve an integrative role torapidly distribute a pacemaker signal over a relatively widearea of the renal pelvis. This may well be critical in theproximal regions of the renal pelvis where the distributionof both atypical and typical SMCs is relatively sparse andnot as well organized as in the more distal regions. Eventhough the intrinsically low frequency of Ca2+ transientdischarge in ICC-like cells bathed in nifedipine makes it isunlikely that these cells are acting as a primary pacemaker,it is tempting to suggest that ICC-like cells could provide asecondary drive which can take over pacemaking duringconditions that dislocate the ureter from the proximalpacemaker drive, e.g. during ureteric obstruction56 or afterkidney transplantation.
Conclusions
Except for regions distal of the ureteropelvic junctionit is likely that atypical SMCs and ICC-like cells bothinfluence the spontaneous electrical and contractile activityof the upper urinary tract.We believe that atypical SMCsare indeed the primary pacemakers for pyelouretericperistalsis, the function of ICC-like cells has yet to bedetermined. We hav edemonstrated that the mechanisms ofCa2+ transient generation in these 2 cell populations can bepharmacologically distinguished on the basis of their(in)sensitivity to ryanodine.Elucidation of the fundamentalmechanisms by which ICC-like cells and atypical SMCscontrol muscle wall contractility will lead to thedevelopment of functional/ electrical models that combinethe autorhythmicity of atypical SMCs and ICC-like cellswith the ultrastructure and electrical coupling of all of thecells involved under normal physiological and pathologicalconditions. To date, non-surgical pharmaceutical treatmentsto alleviate the consequences of obstruction-inducedremodelling in the upper urinary system have not beendeveloped due to a lack of a basic understanding of thefundamental physiology underlying the initiation andpropagation of pyeloureteric contractions. Thuspyeloureteric ICC-like cells and atypical SMCs with theirunique distribution and autorhythmicity may well provideselective pharmacological targets when considering non-surgical interventions to alleviate hydronephrosis arisingfrom pelviureteric remodelling during and after uretericblockade or pyeloplasty.
Acknowledgements
This work was supported in part by the NationalHealth & Medical Research Council (Australia) and Grant-in-Aids from Japan Society for the Promotion of Science to
126 Proceedings of the Australian Physiological Society (2009)40
R.J. Lang, H. Hashitani, M.A. Tonta, J.L. Bourke, H.C. Parkington & H. Suzuki
H.H. (No.17390443).
References
1. Golenhofen K, Hannappel J. Normal spontaneousactivity of the pyeloureteral system in the guinea-pig. Pflügers Arc h. 1973;341: 257-70.
2. GoslingJA, Dixon JS. Species variation in the locationof upper urinary tract pacemaker cells.Invest. Urol.1974;11: 418-23.
3. Dixon JS, Gosling JA. The musculature of the humanrenal calices, pelvis and upper ureter. J. Anat. 1982;135: 129-39.
4. Dixon JS, Gosling JA. Ultrastructure of smooth musclecells in the urinary system. In: Ultrastructure ofsmooth muscles. Edited by Motta PM. Boston:Kluwer Academic Publishers, p. 153-70, 1990.
5. Klemm MF, Exintaris B, Lang RJ. Identification of thecells underlying pacemaker activity in the guinea-pigupper urinary tract.J. Physiol.1999;519: 867-84.
6. Dixon JS, Gosling, JA. The fine structure of pacemakercells in the pig renal calices.Anat. Res.1973; 175:139-53.
7. Rolle U, Brylla E, Tillig B et al. Demonstration ofintrinsic innervation of the guinea pig upper urinarytract using whole-mount preparation.Neurourol.Urodyn.2008;27: 341-7.
8. LangRJ, Exintaris B, Teele MEet al.Electrical basis ofperistalsis in the mammalian upper urinary tract.Clin. Exp. Pharmacol. Physiol.1998;25: 310-21.
9. Santicioli P, Maggi CA. Myogenic and neurogenicfactors in the control of pyeloureteral motility andureteral peristalsis.Pharmacol. Rev. 1998; 50:683-722.
10. Teele ME, Lang RJ. Stretch-evoked inhibition ofspontaneous migrating contractions in a wholemount preparation of the guinea-pig upper urinarytract.Br. J. Pharmacol.1998;123: 1143-53.
11. Burdyga T, Wray S. Action potential refractory periodin ureter smooth muscle is set by Ca sparks and BKchannels.Nature2005;436: 559-62.
12. ZhangY, Lang RJ. Effects of intrinsic prostaglandinson the spontaneous contractile and electrical activityof the proximal renal pelvis of the guinea-pig.Br. J.Pharmacol.1994;113: 431-8.
13. LangRJ, Zhang Y, Exintaris B et al. Effects of nervestimulation on the spontaneous action potentialsrecorded in the proximal renal pelvis of the guinea-pig. Urol. Res.1995;23: 343-50.
14. LangRJ, Zhang Y. The effects of K+ channel blockerson the spontaneous electrical and contractile activityin the proximal renal pelvis of the guinea pig.J.Urol. 1996;155: 332-6.
15. LangRJ, Hashitani H, Tonta MAet al. Spontaneouselectrical and Ca2+ signals in typical and atypicalsmooth muscle cells and interstitial cell of Cajal-likecells of mouse renal pelvis.J. Physiol. 2007; 583:1049-68.
16. LammersWJ, Ahmad HR, Arafat K. Spatial and
temporal variations in pacemaking and conduction inthe isolated renal pelvis.Am. J. Physiol. 1996;270:F567-74.
17. Yamaguchi O, Constantinou CE. Renal calyceal andpelvic contraction rhythms. Am. J. Physiol. 1989;257: 788-95.
18. Lang RJ, Takano H, Davidson ME et al.Characterization of the spontaneous electrical andcontractile activity of smooth muscle cells in the ratupper urinary tract.J. Urol. 2001;166: 329-34.
19. LangRJ, Hashitani H, Tonta MA et al. Role of Ca2+
entry and Ca2+ stores in atypical smooth muscle cellautorhythmicity in the mouse renal pelvis.Br. J.Pharmacol.2007;152: 1248-59.
20. Tsuchida S, Suzuki T. Pacemaker activity of thepelvicalyceal border recorded by an intracellularglass microelectrode.Urol. Intern.1992;48: 121-4.
21. ChangSJ, Cho ET, Heo GSet al. Characterization ofpacemaking currents in cultured interstitial cells ofCajal from mice small intestine.Kor. J. Gastrol.2003;42: 121-7.
22. Walker RL, Koh SD, Sergeant GPet al. TRPC4currents have properties similar to the pacemakercurrent in interstitial cells of Cajal.Am. J. Physiol.2002; 283: C1637-45.
23. LangRJ, Hashitani H, Keller Set al. Modulators ofinternal Ca2+ stores and the spontaneous electricaland contractile activity of the guinea-pig renalpelvis.Br. J. Pharmacol.2002;135: 1363-674.
24. LangRJ, Zoltkowski BZ, Hammer JMet al. Electricalcharacterization of interstitial cells of Cajal-like cellsand smooth muscle cells isolated from the mouseureteropelvic junction.J. Urol. 2007; 177: 1573-80.
25. SmithRD, Borisova L, Wray Set al. Characterisationof the ionic currents in freshly isolated rat uretersmooth muscle cells: evidence for species-dependentcurrents.Pflügers Arc h. 2002;445: 444-53.
28. Boittin FX, Macrez N, Halet Get al. Norepinephrine-induced Ca2+ waves depend on InsP3 and ryanodinereceptor activation in vascular myocytes. Am. J.Physiol.1999;277: C139-51.
29. Burdyga TV, Taggart MJ, Crichton C.et al. Themechanism of Ca2+ release from the SR ofpermeabilised guinea-pig and rat ureteric smoothmuscle.Biochim. Biophys. Acta1998;1402: 109-14.
30. Boittin FX, Coussin F, Morel JL et al. Ca2+ signalsmediated by Ins1,4,5P3-gated channels in ratureteric myocytes.Biochem. J.2000;349: 323-32.
31. Sergeant GP, Hollywood MA, McCloskey KD et al.Role of IP3 in modulation of spontaneous activity inpacemaker cells of rabbit urethra.Am. J. Physiol.2001;280: C1349-56
Proceedings of the Australian Physiological Society (2009)40 127
Autorhythmicity in the mouse pyeloureteric system
32. Van Helden DF, Imtiaz MS, Nurgaliyeva K et al. Roleof calcium stores and membrane voltage in thegeneration of slow wav eaction potentials in guinea-pig gastric pylorus.J. Physiol.2000;524: 245-65.
33. Ward SM, Ordog T, Koh SD et al. Pacemaking ininterstitial cells of Cajal depends upon calciumhandling by endoplasmic reticulum andmitochondria.J. Physiol.2000;525: 355-61.
34. Aoyama M, Yamada A, Wang Jet al. Requirement ofryanodine receptors for pacemaker Ca2+ activity inICC and HEK293 cells.J. Cell Sci. 2004; 117:2813-25.
35. Liu HN, Ohya S, Furuzono Set al. Co-contribution ofIP3R and Ca2+ influx pathways to pacemaker Ca2+
activity in stomach ICC.J. Bio. Rhyth.2005;20: 1536. SuzukiH, Takano H, Yamamoto Yet al. Properties of
gastric smooth muscles obtained from mice whichlack inositol trisphosphate receptor. J. Physiol.2000;525: 105-11.
37. Gregory RB, Rychkov G, Barritt GJ. Evidence that2-aminoethyl diphenylborate is a novel inhibitor ofstore-operated Ca2+ channels in liver cells, and actsthrough a mechanism which does not involveinositol trisphosphate receptors.Biochem. J. 2001;354: 285-90.
38. Gordienko DV, Bolton, TB. Crosstalk betweenryanodine receptors and IP3 receptors as a factorshaping spontaneous Ca2+-release events in rabbitportal vein myocytes.J. Physiol.2002;542: 743-62.
39. Lang RJ, Davidson ME, Exintaris B. Pyeloureteralmotility and ureteral peristalsis: essential role ofsensory nerves and endogenous prostaglandins.Exp.Physiol.2002;87: 129-46.
40. Weiss RM, Tamarkin FJ, Wheeler MA. Pacemakeractivity in the upper urinary tract.J. Smooth Musc.Res.2006;42: 103-15.
41. Sutko JL, Airey JA, Welch Wet al.The pharmacologyof ryanodine and related compounds.Pharmacol.Rev. 1997;49: 53-98.
42. McHaleN, Hollywood M, Sergeant Get al. Origin ofspontaneous rhythmicity in smooth muscle.J.Physiol.2006;570: 23-8.
43. HashitaniH, Suzuki H. Identification of interstitialcells of Cajal in corporal tissues of the guinea-pigpenis.Br. J. Pharmacol.2004;141: 199-204.
44. Malysz J, Donnelly G, Huizinga JD. Regulation ofslow wav e frequency by IP3-sensitive calciumrelease in the murine small intestine.Am. J. Physiol.2001;280: G439-48.
45. van Helden DF, Imtiaz MS. Ca2+ phase wav es: a basisfor cellular pacemaking and long-rangesynchronicity in the guinea-pig gastric pylorus. J.Physiol.2003;548: 271-96.
46. ExintarisB, Lang RJ. K+ channel blocker modulationof the refractory period in spontaneously activeguinea-pig ureters.Urol. Res.1999;27: 319-27.
47. HannappelJ, Lutzeyer W. Pacemaker localization inthe renal pelvis of the unicalyceal kidney. In vitrostudy in the rabbit.Eur. Urol. 19784: 192-4.
48. ConstantinouCE, Neubarth JL, Mensah-Dwumah M.Frequency gradient in the autorhythmicity of thepyeloureteral pacemaker system.Experientia1978;34: 614-5.
49. SandersKM, Ordog T, Koh SD et al. A novelpacemaker mechanism drives gastrointestinalrhythmicity.News Physiol.Sci.2000;15: 291-8.
50. Ward SM, Burns AJ, Torihashi Set al.Mutation of theproto-oncogene c-kit blocks development ofinterstitial cells and electrical rhythmicity in murineintestine.J. Physiol.2004;480: 91-7.
51. Huizinga JD, Thuneberg L, Kluppel M et al. W/kitgene required for interstitial cells of Cajal and forintestinal pacemaker activity. Nature 1995; 373:347-9.
52. Pezzone MA, Watkins SC, Alber SM et al.Identification of c-kit-positive cells in the mouseureter: the interstitial cells of Cajal of the urinarytract.Am. J. Physiol.2003;284: F925-9.
53. LangRJ, Klemm MF. Interstitial cell of Cajal-like cellsin the upper urinary tract.J. Cell. Molec. Med.2005;9: 543-56.
54. MetzgerR, Schuster T, Till H et al. Cajal-like cells inthe upper urinary tract: comparative study in variousspecies.Pediat. Surg. Inter.2005;21: 169-74.
55. Van der Aa F, Roskams T, Blyweert W et al.Identification of kit positive cells in the humanurinary tract.J. Urol. 2004;171: 2492-2496.
56. SolariV, Piotrowska AP, Puri P. Altered expression ofinterstitial cells of Cajal in congenital ureteropelvicjunction obstruction.J. Urol. 2003;170: 2420-2.
57. MetzgerR, Schuster T, Till H et al. Cajal-like cells inthe human upper urinary tract.J. Urol. 2004; 172:769-72.








![W E L C O M E []€¦ · 4. Registering Property 112 112 108 121 122 117 102 0 -10 5. Getting Credit 118 109 104 86 129 126 128-9 10 6. Protecting Investors 137 155 154 128 128 133](https://static.documents.pub/doc/80x56/5f0582267e708231d4134f73/w-e-l-c-o-m-e-4-registering-property-112-112-108-121-122-117-102-0-10-5-getting.jpg)








![Young Mathematicians Conference · Polynomial Sequence, Congressus Numerantium, 184(2007), 121-128. [3]Molina, R., Zeleke, A. Generalizing Results On the Convergence of the Maximum](https://static.documents.pub/doc/80x56/6023c3baa039c169985e9c28/young-mathematicians-conference-polynomial-sequence-congressus-numerantium-1842007.jpg)




