Page 1
Streptococcus pneumoniae secretes a glyceraldehyde-3-phosphate dehydrogenase, which binds haemoglobinand haem
Zelene Edith Vazquez-Zamorano • Marco Antonio Gonzalez-Lopez •
Marıa Elena Romero-Espejel • Elisa Irene Azuara-Liceaga •
Mavil Lopez-Casamichana • Jose de Jesus Olivares-Trejo
Received: 2 December 2013 / Accepted: 2 June 2014
� Springer Science+Business Media New York 2014
Abstract Streptococcus pneumoniae is a gram posi-
tive encapsulated bacterium responsible of septicaemia
and upper respiratory infections in children. This
pathogen requires iron to survive in the host, which it
can obtain of haemoglobin (Hb) or haem. Only two Hb-
binding membrane proteins have been identified up to
now. However it is unknown whether this pathogen
secretes proteins in order to scavenge iron from the Hb or
haem. Therefore, in order to explore these possibilities,
cellular growth of S. pneumoniae was tested with several
alternative iron supplies. The bacterial growth was
supported with iron, Hb and haem. Additionally, S.
pneumoniae expressed and secreted a protein of 38 kDa
which was purified and characterized as Hb and haem-
binding protein. This protein was also identified by mass
spectrometry as glyceraldehyde-3-phosphate dehydro-
genase. Our overall results suggest that S. pneumoniae
secretes a protein capable of binding two usefull iron
sources for this bacterium (Hb and haem). This protein
could be playing a dynamic role in the success of the
invasive and infective processes of this pathogen.
Keywords Streptococcus pneumoniae � Haem �Haemoglobin � Glyceraldehyde-3-phospate
dehydrogenase � Secreted
Introduction
The capacity of microbial pathogens to obtain iron from
humans is essential for the infection establishment. Iron
can be acquired from lactoferrin (Lf), transferrin (Tf)
and ferritin (Ft) (Andrews et al. 2003). In addition, there
are other iron sources called haemoproteins such as
haemoglobin (Hb), which binds iron via haem (Wan-
dersman and Delepelaire 2004). Hb is a human iron
source found in the microenvironment of many tissues
and accessible to almost all pathogens (Wandersman
and Stojiljkovic 2000). Several mechanisms describing
these processes have been described in different bacte-
ria. In one of these processes, proteins called haemo-
phores are released to bind Hb or haem from the
extracellular media and deliver it to membrane recep-
tors for its subsequent internalisation (Brown et al.
2004). Although the amino acid sequence of haem-
binding proteins varies significantly between patho-
gens, it has been proposed that some of these receptor
Zelene Edith Vazquez-Zamorano and Marco Antonio
Gonzalez-Lopez have contributed equally to this work.
Z. E. Vazquez-Zamorano � M. A. Gonzalez-Lopez �M. E. Romero-Espejel � E. I. Azuara-Liceaga �M. Lopez-Casamichana � J. J. Olivares-Trejo (&)
Posgrado en Ciencias Genomicas, Universidad Autonoma
de la Ciudad de Mexico, San Lorenzo 290, Del Valle,
C.P. 03100 Ciudad de Mexico, D.F., Mexico
e-mail: [email protected]
M. E. Romero-Espejel
Departamento de Infectomica y Patogenesis Molecular,
Centro de investigacion y de estudios avanzados del IPN,
Ciudad de Mexico, Mexico
123
Biometals
DOI 10.1007/s10534-014-9757-0
Page 2
proteins bind the iron source via the two motifs FRAP
and NPNL (Simpson et al. 2000) or the motif KVAFDH,
which is found in Haemophilus influenzae (Reidl and
Mekalanos 1996). These mechanisms have been
described in more detail in gram negative bacteria than
in gram positive bacteria. In Staphylococcus aureus, at
least four proteins have been suggested to be involved in
haem binding (IsdA, B, C and H) (Torres et al. 2006).
Interestingly, in Streptococcus pneumoniae, which is a
gram positive encapsulated bacterium responsible for
septicaemia and upper respiratory tract infections in
children (Tuomanen et al. 2004), not all of the available
iron from the host can be acquired by this pathogen
because S. pneumoniae can use Hb and haem but not Lf
and Tf (Tai et al. 1993). In this pathogen, only two
membrane proteins have been identified as Hb and
haem-binding proteins thus far (Romero-Espejel et al.
2013). Unfortunately, the complete mechanism utilised
by this bacterium to acquire iron is poorly understood
because no iron scavenging proteins secreted by this
bacterium has been described to date. In this study, we
have isolated a protein that is secreted by S. pneumoniae
and binds both Hb and haem. Although this protein was
revealed to be glyceraldehyde-3-phosphate dehydroge-
nase (GAPDH), which is an enzyme with well-estab-
lished metabolic roles, our findings suggest an addition
role for this protein in iron-binding.
Materials and methods
Conditions for bacterial growth
The R6 strain of S. pneumoniae was grown under
microaerophilic conditions in 5 % CO2 for 24 h at
37 �C using agar supplemented with 5 % sheep blood.
The cultures were then inoculated in Todd Hewitt-
Broth (THB) supplemented with 0.5 % yeast extract
(THB-Y) under the same conditions until the optical
density (600 nm) reached 0.3. To test the effect of
alternative iron sources such as haem (Sigma
51280-5G), Hb (Sigma H7379-10G), FeCl3 or holo-
Tf (Sigma T-4132), a chelating agent (500 lM 2,
2-dipyridyl) (Sigma D216305-25G) was used, and the
bacteria were incubated for 16 h at 37 �C in 5 % CO2.
To synchronise cellular growth, the optical density
(measured at 600 nm) was adjusted to 0.1 and mon-
itored every hour. After 3 h under iron starvation the
culture medium was supplemented with haem, Hb,
FeCl3, Tf or fresh medium (THB), with the concentra-
tion adjusted to 10 lM of iron. E. coli O157:H7 strain
EDL 933 was grown in LB broth for 12 h at 37 �C.
Separation of secreted protein from total proteins
To prevent cellular lysis, the bacteria were cultivated for
7 h in THB under iron-starved conditions. Cells were
then collected by centrifugation at 6,0009g for 10 min,
and the supernatant was filtered through a 0.45 lm pore
size membrane filter to remove residual bacteria and
then concentrated by centrifugation using Amicon
Ultra-15 filtration units at 5,0009g for 40 min to obtain
the S. pneumoniae secretome. The cellular pellet was
washed three times with THB by centrifugation at
6,0009g for 3 min and resuspended in medium supple-
mented with 1 mM PMSF. The bacterial cells were
lysed by sonication using 30 s pulses for 6 min. Samples
were centrifuged at 12,0009g for 20 min to remove
unbroken cells, and the supernatant was taken as the
total protein fraction (Gonzalez-Lopez et al. 2013).
Determination of b-galactosidase enzymatic
activity
b-Galactosidase activity was determined following pro-
tocol of Miller (Miller 1972), the bacteria were incubated
overnight, cells were then diluted in fresh medium, grew
to mid-log, the cultures were incubated for 20 min on ice
to stop growth and washed, 2 ml cells were centrifuged at
6,0009g for 10 min to separate the supernatant, the cell
pellet was resuspended in the same volume of Z buffer
chilled (60 mM Na2HPO4.7H2O, 40 mM NaH2PO4.H2-
O, 10 mM KCl, 1 mM MgSO4 and 50 mM b-mercap-
toethanol) and measured the optical density at 600 nm.
1 ml of Z buffer containing bacteria was mixed with
200 ll (4 mg/ml) of ortho-nitrophenyl-b-galactoside
(ONPG used as substrate). The reaction was developed
adding 500 ll 1 M Na2CO3. The colorimetric signal was
monitored by spectrophotometry at 420 and 550 nm.
Activity units were calculated using the following
equation: Miller units = 1,000 9 [(OD420 -1.75 9
OD550)]/(T 9 V 9 OD600), OD420 and OD550. OD600;
reflects cell density, T; time of the reaction (min) and V;
volume used in the assay (ml). The experiments were
performed by triplicate in three occasions.
Biometals
123
Page 3
Protein quantification
The concentration of proteins was determined by
spectrophotometry at 595 nm using the Bradford
method; a standard curve was generated using Brad-
ford reagent and bovine serum albumin at concentra-
tions of 1, 5, 10, 25 and 50 lg/ml, the concentration of
samples was determined by interpolation.
Densitometry
The proteins separated by SDS-PAGE were submitted
to Chemi Doc (BioRad) equip and the protein bands
were quantified using the Quantity One program
version 4.6.3 with the tool volume analysis report to
determine the concentration of a protein by their
intensity in the gel.
Coomassie brilliant blue and hydrogen peroxide
staining
Proteins were resolved in gels and used for subsequent
mass spectrometry analysis were visualised by a
Coomassie brilliant blue R-250 method; the gel was
immersed in a staining solution (0.1 % Coomassie
brilliant blue R-250, 50 % methanol and 10 % glacial
acetic acid) for 15 min with gentle agitation and then a
destaining solution was added (40 % methanol and
10 % glacial acetic acid) until the gel background was
fully clear. To identify haem interaction, native gels
were revealed with hydrogen peroxide (3.5 %).
SDS-PAGE and native gels
50 lg proteins were prepared in loading buffer (1 %
SDS, 10 % glycerol, 10 mM Tris–Cl, pH 6.8, 0.5 M
dithiothreitol, 1 % Bromophenol blue) and loading
onto SDS-PAGE gel (30 % acrylamide/bisacrylamide
(29:1), 1.5 M Tris–HCl buffer (pH 8.8), 10 % ammo-
nium persulfate solution, 0.08 % TEMED, 0.1 % SDS,
in the case of native gel the sample was mixed with
binding buffer (250 mM Tris/HCl, pH 8.0, 5 mM
EDTA and 10 % glycerol) for 30 min at 25 �C, after
that the sample was loaded in the native gel (7 %
acrylamide/bisacrylamide, 1.5 M Tris–HCl buffer (pH
8.8), 10 % ammonium persulfate solution and 0.08 %
TEMED, the sample was resolving at 35 mA.
Western blotting
A Western blot was performed using anti-b-galacto-
sidase antibodies as an internal control to verify that no
intracellular proteins were released under our cellular
culture conditions. Proteins were transferred to nitro-
cellulose membranes in a semi-dry trans-blot cell for
1 h at 100 mA in a solution of 30 mM Tris, 1.4 %
glycine and 20 % methanol. The membranes were
soaked in PBST buffer (137 mM NaCl, 2.7 mM KCl,
10 mM Na2HPO4, 2 mM KH2PO4 and 0.1 % Tween
20) containing 5 % non-fat milk overnight to saturate
all remaining active binding sites. The membrane was
washed three times with PBST and then was incubated
with anti-b-galactosidase antibody (Millipore)
(1:40,000) for 1 h. Five washes were performed with
PBST, and the secondary antibody was added (anti
rabbit coupled to horseradish peroxidase; 1:10,000).
The antibody was specific for E. coli protein but cross-
react with homologous protein from S. pneumoniae.
Purification of secreted proteins by haem-affinity
chromatography
The secretome of bacteria grown in THB media under
iron-starved supplemented with Hb was loaded onto
haem-affinity chromatography resin (Sigma H6390)
and was incubated overnight at 4 �C. The chromato-
graphic fraction was centrifuged at 1,0009g for
Table 1 List of the bacterial species and UniProt access
numbers of GAPDH protein used to perform the alignment
illustrated in Fig. 6
Bacteria GAPDH Access
number UniProt
Streptococcus pneumoniae Q8CWN6
Bacillus anthracis Q81X74
Bacillus subtilis P09124
Clostridium botulinium C1FQW2
Escherichia coli P58072
Haemophilus influenzae E1X8N0
Haemophilus influenzae e-P4 C9MJZ6
Helicobacter pylori Q9ZJP0
Neisseria maningitidis E7BH93
Staphylococcus aureus Q6GIL8
Streptococcus pneumoniae 24 Q97P40
Streptococcus pneumoniae 37 Q97RH40
Biometals
123
Page 4
1 min, the flow-through was collected, and the resin
was washed three times with wash buffer (50 mM
Tris–HCl pH 7.0, 300 mM NaCl) to eliminate non-
specific interactions. Secreted proteins bound to haem
were eluted using a 6 M guanidine hydrochloride
solution. The samples were then cleaned of salts, lipids
and nucleic acids by precipitation using a kit (Cleanup
kit, Bio-Rad), and the proteins were recovered in the
pellet. Protein concentration was determined by the
Bradford method. A total of 50 ll of protein was
resolved by SDS-PAGE and visualised by Coomassie
blue staining to evaluate the integrity and resolution of
the proteins in the mixture (Asuthkar 2007).
Hb-binding experiments
The secreted fraction purified by haem-affinity was
used to evaluate the Hb-binding capability of the
38 kDa protein. First, 30 lg of purified proteins was
suspended in binding buffer (250 mM Tris/HCl,
5 mM EDTA and 10 % glycerol, pH 6.8) and was
incubated with 20 lM Hb at 37 �C for 30 min. The
complex was then separated by native PAGE (7 %
gel). To identify the interaction of the 38 kDa protein
with the iron source (Hb or Tf) the native gels were
treated with hydrogen peroxide (3.5 %) and protein
was visualised by Coomassie blue staining (Cruz-
Castaneda et al. 2011).
Competition assays
Competition experiments were performed in the same
way as haem-affinity chromatography, but the bound
proteins were eluted using several possible competi-
tors such as haem (180 lmol), Hb (180 lmol) or Tf
(180 lmol) instead of guanidine hydrochloride. Com-
petitors were used at an iron concentration ten-fold
higher than that used in haem-affinity chromatogra-
phy. Finally, 30 ll of each sample was resolved by
SDS-PAGE and visualised by Coomassie blue stain-
ing (Romero-Espejel et al. 2013).
Mass spectrometry analysis
The 38 kDa protein band was excised from the gel and
was digested with trypsin. LC–MS/MS was performed
on a Micromass QTof I equipped with a LC Packings
nanoflow LC. A total of 5 ll of digested protein
solution was injected onto a LC Packings C18 PepMap
column (0.75 lm 9 15 cm) and was eluted with a
linear acetonitrile gradient at a flow rate of 200 nL/
min. The peptides eluted from the column were
introduced into the mass spectrometer through a New
Objective PicoTip held by a New Objective adapter.
The experimental conditions were as follows: capil-
lary voltage of 1.8 kV, cone voltage of 32 V and
collision energy according to the mass and charge of
the ion, ranging from 14 eV to 50 eV. Raw data files
were processed using the MassLynx ProteinLynx
software and pkl files were analysed by www.
matrixscience.com using the Mascot algorithm (Pro-
tein Core Facility, Columbia University Medical
Center).
Multiple amino acid sequence alignment
The amino acid sequence of the S. pneumoniae
GAPDH (Q8CWN6) protein was compared with the
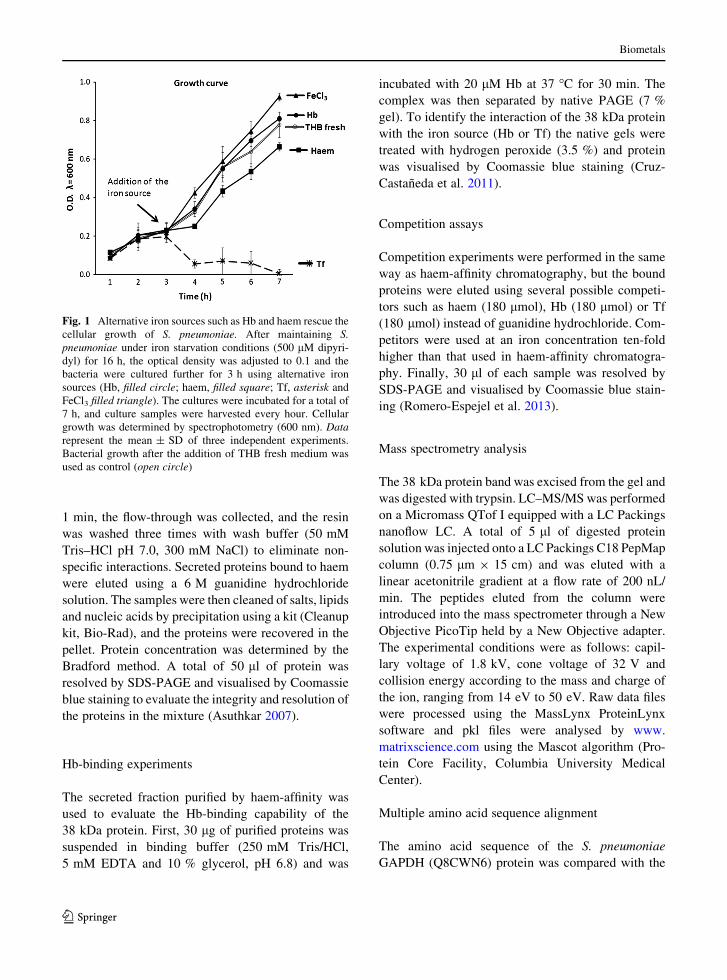
Fig. 1 Alternative iron sources such as Hb and haem rescue the
cellular growth of S. pneumoniae. After maintaining S.
pneumoniae under iron starvation conditions (500 lM dipyri-
dyl) for 16 h, the optical density was adjusted to 0.1 and the
bacteria were cultured further for 3 h using alternative iron
sources (Hb, filled circle; haem, filled square; Tf, asterisk and
FeCl3 filled triangle). The cultures were incubated for a total of
7 h, and culture samples were harvested every hour. Cellular
growth was determined by spectrophotometry (600 nm). Data
represent the mean ± SD of three independent experiments.
Bacterial growth after the addition of THB fresh medium was
used as control (open circle)
Biometals
123
Page 5
protein sequences of GAPDH from Bacillus anthracis,
Bacillus subtilis, Clostridium botulinum, Escherichia
coli, Helicobacter pylori, Neisseria meningitidis, S.
aureus and two membrane proteins of S. pneumoniae
of 22 and 37 kDa (Romero-Espejel et al. 2013) and the
protein e-P4 of H. influenza, which had the haem-
binding motif (Reidl and Mekalanos 1996) (Table 1).
The amino acid sequences were submitted to the http://
www.ebi.ac.uk/Tools/msa/clustalw2/ server to obtain
an alignment. The residues were highlighted in bold-
face using the JalView program 2.8. The amino acid
sequence of S. pneumoniae GAPDH (Q8CWN6)
protein was submitted to the CPH models 3.2 server
(http://www.cbs.dtu.dk/services/CPHmodels/) to
obtain the PDB file, which was used to construct a 3-D
model using PyMol program 0.9.
Results
Hb and haem support the cellular growth of
S. pneumoniae but transferrin does not
It has been proposed that S. pneumoniae uses Hb or
haem as iron sources, and under these conditions two
membrane proteins are expressed by a direct mecha-
nism (Romero-Espejel et al. 2013). However, an
indirect mechanism explaining the role of haemophore
secretion in iron acquisition by the bacterium has not
been documented (Brown et al. 2001). To investigate
whether S. pneumoniae secretes proteins capable of
scavenging iron from Hb or haem, this bacterium was
grown using different iron sources Hb (filled circle),
haem (filled square), FeCl3 (filled triangle), Tf
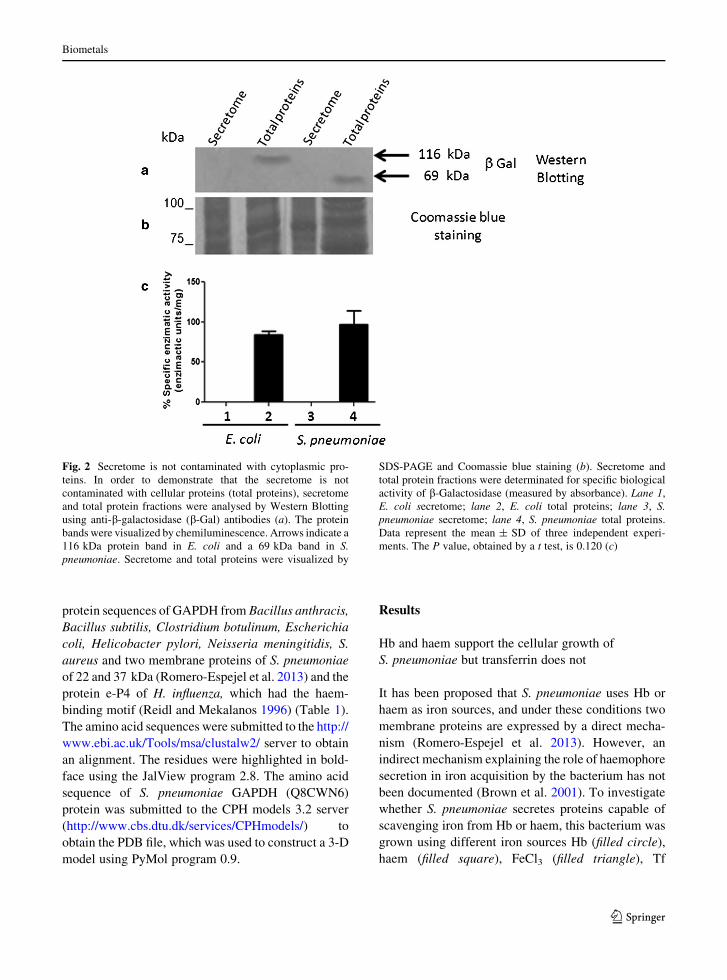
Fig. 2 Secretome is not contaminated with cytoplasmic pro-
teins. In order to demonstrate that the secretome is not
contaminated with cellular proteins (total proteins), secretome
and total protein fractions were analysed by Western Blotting
using anti-b-galactosidase (b-Gal) antibodies (a). The protein
bands were visualized by chemiluminescence. Arrows indicate a
116 kDa protein band in E. coli and a 69 kDa band in S.
pneumoniae. Secretome and total proteins were visualized by
SDS-PAGE and Coomassie blue staining (b). Secretome and
total protein fractions were determinated for specific biological
activity of b-Galactosidase (measured by absorbance). Lane 1,
E. coli secretome; lane 2, E. coli total proteins; lane 3, S.
pneumoniae secretome; lane 4, S. pneumoniae total proteins.
Data represent the mean ± SD of three independent experi-
ments. The P value, obtained by a t test, is 0.120 (c)
Biometals
123
Page 6
(asterisk) and free iron (open circle) after previous
iron depletion using dipyridyl addition (iron chelator).
Our results showed that this pathogen uses selective
iron sources for its cellular growth, and while iron, Hb
and haem support the cellular growth of S. pneumo-
niae, Tf does not (Fig. 1).
S. pneumoniae expresses a 38 kDa haem-binding
protein
To determine whether cytoplasmic proteins released
upon lysis of S. pneumoniae contaminate the secreted
proteins, the total and secreted protein fractions from S.
pneumoniae were resolved by SDS–PAGE (Fig. 2b),
and the presence of b-galactosidase (b-Gal), an exclu-
sive cytoplasmic protein, was detected by Western blot
analysis. b-Gal was detected in total protein samples
(Fig. 2a, lane 4), but not in the secreted protein fraction
(Fig. 2a, lane 3). These results indicated that under our
conditions of bacterial culture, the cytoplasmic proteins
do not contaminate the secreted proteins. As an
experimental control, this protocol was also performed
using E. coli (Fig. 2a and b, lanes 1 and 2). To
corroborate that the secretome is not contaminated with
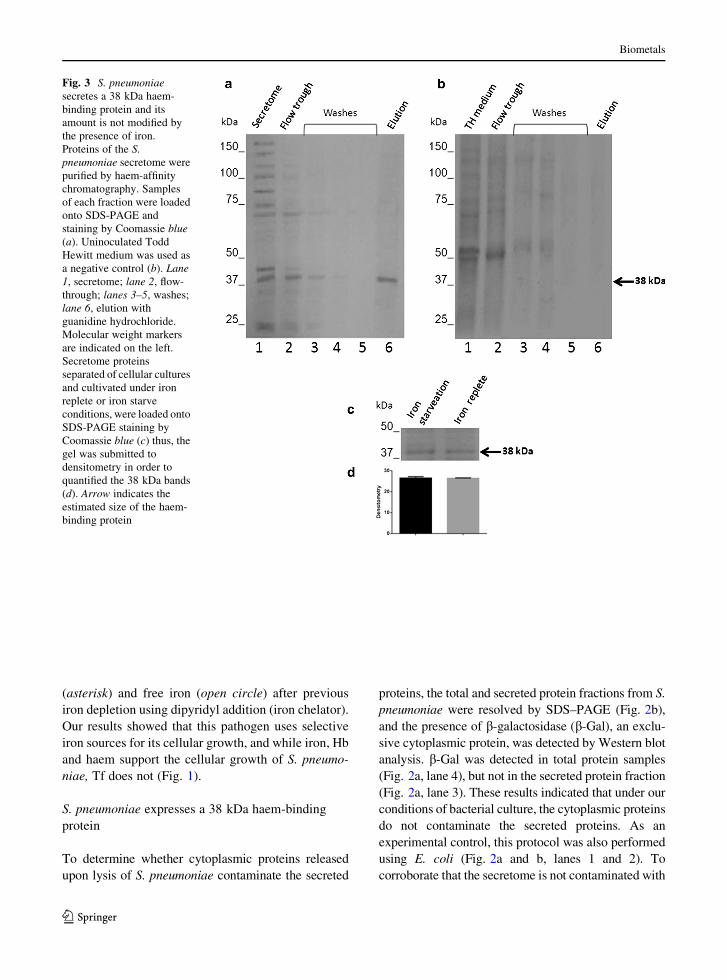
Fig. 3 S. pneumoniae
secretes a 38 kDa haem-
binding protein and its
amount is not modified by
the presence of iron.
Proteins of the S.
pneumoniae secretome were
purified by haem-affinity
chromatography. Samples
of each fraction were loaded
onto SDS-PAGE and
staining by Coomassie blue
(a). Uninoculated Todd
Hewitt medium was used as
a negative control (b). Lane
1, secretome; lane 2, flow-
through; lanes 3–5, washes;
lane 6, elution with
guanidine hydrochloride.
Molecular weight markers
are indicated on the left.
Secretome proteins
separated of cellular cultures
and cultivated under iron
replete or iron starve
conditions, were loaded onto
SDS-PAGE staining by
Coomassie blue (c) thus, the
gel was submitted to
densitometry in order to
quantified the 38 kDa bands
(d). Arrow indicates the
estimated size of the haem-
binding protein
Biometals
123
Page 7
cellular proteins (total proteins), secretome and total
protein fractions were determined for b-galactosidase
activity following protocol of Miller (Fig. 2c), the b-
galactosidase activity was detected only in total protein
samples, the values obtained were 83.93 Miller units to
E. coli and 97.03 Miller units to S. pneumoniae (Fig. 2c,
lanes 2 y 4) and 0 Miller units in the secreted proteins
(Fig. 2c, lanes 1 y 3). Our results clearly showed that
under these culture conditions the secreted and cyto-
plasmic proteins do not mix.
To investigate whether S. pneumoniae secretes
proteins with an affinity for haem, S. pneumoniae was
cultivated in THB. The supernatant (secretome) was
separated and loaded onto a haem-affinity chromatog-
raphy column (Fig. 3a, lane 1). The flow-through
containing unbound proteins of the bacterial secre-
tome (Fig. 3a, lane 2) and three subsequent wash
fractions (Fig. 3a, lanes 3–5) were collected, and
finally, proteins bound to the resin were eluted by
increasing the stringency of the elution buffer (Fig. 3a,
lane 6). These results showed the presence of a major
protein of 38 kDa (obtained by RF calculation). To
exclude the possibility that this protein could be a
contaminating component from the Todd Hewitt
Broth, a negative control was performed with this
medium. As expected, no protein was eluted from the
control sample (Fig. 3b, lane 6). Our results clearly
showed that an S. pneumoniae 38 kDa protein was
secreted, but we did not know if the growing
conditions modified the amount of 38 kDa protein
secreted, in order to investigate this assumption S.
pneumoniae was cultivated under iron starvation or
iron replete condition, after that, the secreted proteins
were separated and loaded onto SDS-PAGE. Gels
were analysed by densitometry (Fig. 3c). However,
we did not find difference between both conditions, it
seems to be, that the amount of 38 kDa protein
secreted remains without changes (Fig. 3d). There-
fore, we think that the iron source and growth
condition do not modify the amount of 38 kDa protein.
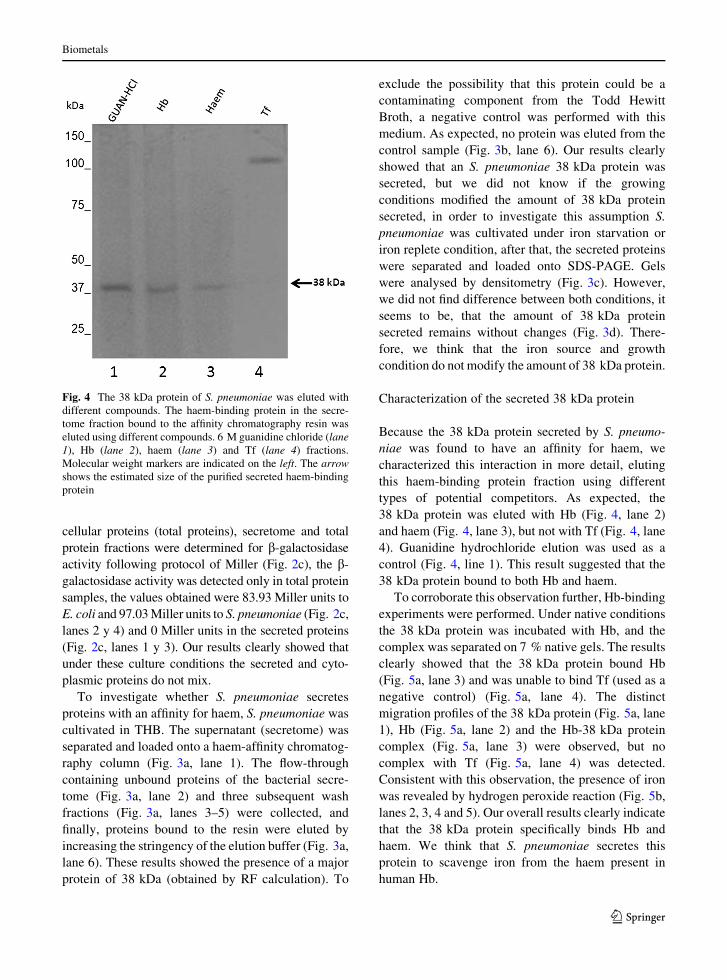
Characterization of the secreted 38 kDa protein
Because the 38 kDa protein secreted by S. pneumo-
niae was found to have an affinity for haem, we
characterized this interaction in more detail, eluting
this haem-binding protein fraction using different
types of potential competitors. As expected, the
38 kDa protein was eluted with Hb (Fig. 4, lane 2)
and haem (Fig. 4, lane 3), but not with Tf (Fig. 4, lane
4). Guanidine hydrochloride elution was used as a
control (Fig. 4, line 1). This result suggested that the
38 kDa protein bound to both Hb and haem.
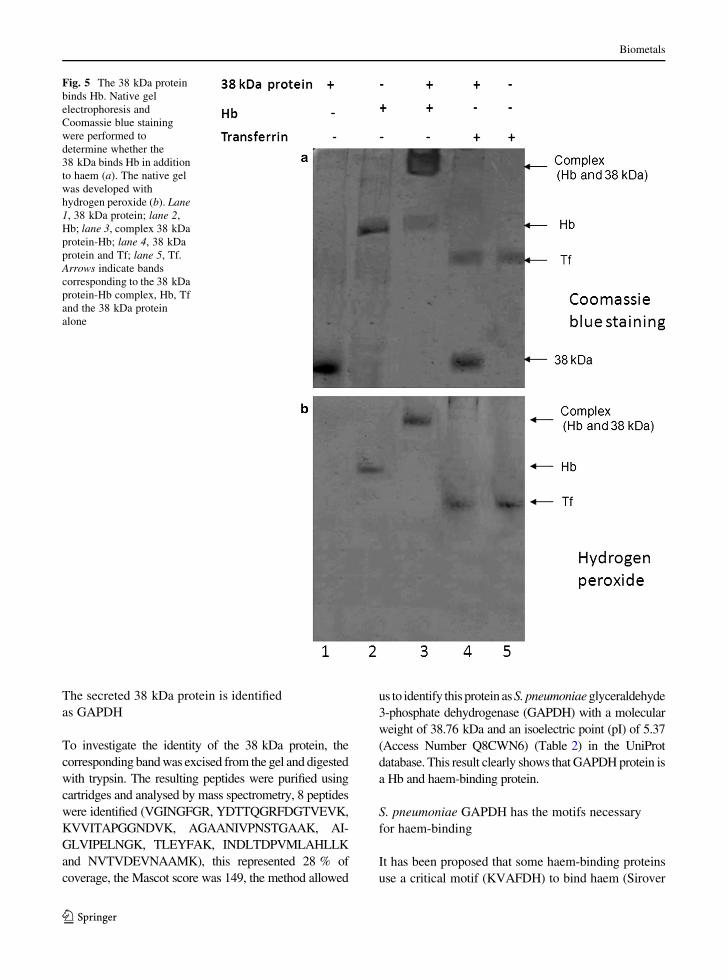
To corroborate this observation further, Hb-binding
experiments were performed. Under native conditions
the 38 kDa protein was incubated with Hb, and the
complex was separated on 7 % native gels. The results
clearly showed that the 38 kDa protein bound Hb
(Fig. 5a, lane 3) and was unable to bind Tf (used as a
negative control) (Fig. 5a, lane 4). The distinct
migration profiles of the 38 kDa protein (Fig. 5a, lane
1), Hb (Fig. 5a, lane 2) and the Hb-38 kDa protein
complex (Fig. 5a, lane 3) were observed, but no
complex with Tf (Fig. 5a, lane 4) was detected.
Consistent with this observation, the presence of iron
was revealed by hydrogen peroxide reaction (Fig. 5b,
lanes 2, 3, 4 and 5). Our overall results clearly indicate
that the 38 kDa protein specifically binds Hb and
haem. We think that S. pneumoniae secretes this
protein to scavenge iron from the haem present in
human Hb.
Fig. 4 The 38 kDa protein of S. pneumoniae was eluted with
different compounds. The haem-binding protein in the secre-
tome fraction bound to the affinity chromatography resin was
eluted using different compounds. 6 M guanidine chloride (lane
1), Hb (lane 2), haem (lane 3) and Tf (lane 4) fractions.
Molecular weight markers are indicated on the left. The arrow
shows the estimated size of the purified secreted haem-binding
protein
Biometals
123
Page 8
The secreted 38 kDa protein is identified
as GAPDH
To investigate the identity of the 38 kDa protein, the
corresponding band was excised from the gel and digested
with trypsin. The resulting peptides were purified using
cartridges and analysed by mass spectrometry, 8 peptides
were identified (VGINGFGR, YDTTQGRFDGTVEVK,
KVVITAPGGNDVK, AGAANIVPNSTGAAK, AI-
GLVIPELNGK, TLEYFAK, INDLTDPVMLAHLLK
and NVTVDEVNAAMK), this represented 28 % of
coverage, the Mascot score was 149, the method allowed
us to identify this protein as S. pneumoniae glyceraldehyde
3-phosphate dehydrogenase (GAPDH) with a molecular
weight of 38.76 kDa and an isoelectric point (pI) of 5.37
(Access Number Q8CWN6) (Table 2) in the UniProt
database. This result clearly shows that GAPDH protein is
a Hb and haem-binding protein.
S. pneumoniae GAPDH has the motifs necessary
for haem-binding
It has been proposed that some haem-binding proteins
use a critical motif (KVAFDH) to bind haem (Sirover
Fig. 5 The 38 kDa protein
binds Hb. Native gel
electrophoresis and
Coomassie blue staining
were performed to
determine whether the
38 kDa binds Hb in addition
to haem (a). The native gel
was developed with
hydrogen peroxide (b). Lane
1, 38 kDa protein; lane 2,
Hb; lane 3, complex 38 kDa
protein-Hb; lane 4, 38 kDa
protein and Tf; lane 5, Tf.
Arrows indicate bands
corresponding to the 38 kDa
protein-Hb complex, Hb, Tf
and the 38 kDa protein
alone
Biometals
123
Page 9
2011). To explore if S. pneumoniae GAPDH has this
motif, amino acid sequence multiple alignment was
performed. The S. pneumoniae GAPDH sequence was
compared with sequences of other proteins including
the S. pneumoniae 22 kDa protein, S. pneumoniae
37 kDa protein and GAPDH proteins from B. anthracis,
B. subtilis, C. botulinum, E. coli, H. influenzae, H.
pylori, N. meningitidis and S. aureus (Table 1). This
analysis revealed the existence of the essential haem-
binding motif (KVAFDH) in the S. pneumoniae GAP-
DH sequence (Fig. 6). To determine the localization of
this motif in the 3D structure of the protein, 3D
modelling was performed using PyMol 0.9. Our
analysis revealed that this motif (KVAFDH) is exposed
in the GAPDH 3D structure (Fig. 7).
Discussion
S. pneumoniae is a pathogenic bacterium capable of
utilising Hb or haem as its only iron source (Tai et al.
1993, Tuomanen et al. 2004). Unfortunately, very
little is known about the mechanisms involved in its
iron acquisition from Hb or haem. Here, we present the
first evidence suggesting that this pathogen secretes an
Table 2 The 38 kDa protein was identified as GAPDH
Access
number
Protein name Coverage Mass
(kDa/
pl)
Mascot
score
Peptides Function
Q8CWN6 Glyceraldehyde-3-
phosphate
dehydrogenase
GAPDH
28 % 38.76/
5.38
149 VGIGFGR, YDTTQGRFDGTVENK,
KVVITAPGGNDVK,
AGAANUVPNSTGAAK,
AIGLVIPELINGK, TLEYFAK,
INDLTDPVMLAHLLK,
NVTVDEVNAAMK
Glucose metabolic
process NAD
binding NADP
binding
The purified protein was resolved by SDS-PAGE. The band corresponding to the protein was then excised from the gel, digested with
trypsin and the purified peptides were analysed and identified by mass spectrometry
Fig. 6 The 38 kDa protein of S. pneumoniae has the motif
necessary for haem-binding. Multiple amino acid sequence
alignment was performed to compare the sequence of S.
pneumoniae GAPDH with that of GAPDH from B. anthracis,
B. subtilis, C. botulinum, E. coli, Haemophilus influenzae, H.
pylori, N. meningitidis, S. aureus and two membrane proteins
containing the haem-binding motif (S. pneumoniae 22 kDa and
S. pneumoniae 37 kDa). The KVAFDH motif is essential for
haem-binding (shown as dark boxes). Val (V) is the most
conserved residue followed by Lys (K) and His (H). Multi-
alignment was performed using Clustal W server and marked
residues were obtained using Jalview program 2.8
Fig. 7 The motif necessary for haem-binding is exposed in
GAPDH as observed from the 3D analysis of the spatial
structure of the protein. The 3D GAPDH structure was
determined using PyMol 0.9 program. The haem-binding motif
is indicated in black
Biometals
123
Page 10
38 kDa protein which binds Hb and haem and its
amount was not modified under iron starvation
condition. This protein was identified as GAPDH.
GAPDH protein is a cytoplasmic enzyme involved
in the glycolytic pathway, which is an essential
metabolic pathway utilised by a wide variety of
organisms ranging from prokaryotic to eukaryotic.
Nevertheless, in addition to metabolic role, the
eukaryotic GAPDH participates in several cellular
functions including transcription, oxidative stress,
apoptosis, autophagy (Sirover 1996, 2011) and haem
binding (Hannibal et al. 2012). Our results suggest that
S. pneumoniae also secretes GAPDH to bind Hb and
haem. We believe that S. pneumoniae GAPDH has
unique characteristics and they differ from rabbit
muscle GAPDH. Rabbit-GAPDH binds haem, but its
tetramer form has a higher haem affinity than the
monomer (Hannibal et al. 2012). In contrast, mono-
meric S. pneumoniae GAPDH binds Hb and haem
efficiently, as was demonstrated by overlay experi-
ments (data no showed). Because these experiments
were performed with high stringency and nevertheless
GAPDH still bound Hb (These conditions maintain the
monomeric form of GAPDH. We suggest that forma-
tion of its quaternary structure of GAPDH is not
necessary for Hb- or haem-binding.
Despite the lack of secretory signals in its amino
acid sequence, GAPDH has been localized on the
surface of several bacteria, fungi and even protozoans
(Pancholi and Chhatwal 2003). Although the mecha-
nism of secretion is not known, there is report in which
GAPDH is releases of the bacterium using a method
detergent-assisted lysis (Oliveira et al. 2012). In the
present study, the bacteria culturing method designed
allowed the growth of bacteria without lysis, our
results suggest that this protein could be secreted using
a different unknown pathway. This notion was further
corroborated by the b-galactosidase control experi-
ment in which the exclusively cytoplasmic protein b-
galactosidase with enzymatic activity was not detected
in the growth media. This observation was consistent
with experiments performed in H. Pylori and S.
pneumoniae (Vanet and Labigne 1998; Choi et al.
2012; Gonzalez-Lopez et al. 2013). Although the
experimentally obtained molecular size of 38 kDa for
GAPDH was higher than that of the other GAPDH
proteins described in the multiple alignment (Fig. 6),
we cannot exclude the possibility that all of these
proteins have this function (Hb and haem-binding).
Multiple alignment revealed the essential motif for
haem-binding (KVAFDH) previously described for
the e-P4 lipoprotein of H. influenzae (Reidl and
Mekalanos 1996). This result was corroborated by
3D modeling in which the spatial location of the
functional motif was elucidated. Our hypothesis is
supported by current evidence suggesting that GAP-
DH is a multifunctional protein displaying diverse
activities distinct from its conventional metabolic role.
In humans, new roles for GAPDH may be dependent
on its subcellular localization, oligomeric state (Han-
nibal et al. 2012) or the proliferative state of the cell
(Mazzola and Sirover 2001). Furthermore, the GAP-
DH of Chlamydomonas reinhardtii has been shown to
be a bifunctional protein (Erales et al. 2009).
In summary, S. pneumoniae GAPDH could play
another role distinct from its role in carbohydrate
metabolism. We propose that S. pneumoniae GAPDH
is an Hb- and haem-binding protein that participates in
iron acquisition and is essential for the successful
invasion of S. pneumoniae in several tissues during the
infection process.
Acknowledgments This work was supported by CONACyT
(Consejo Nacional de Ciencia y Tecnologıa) (Grant numbers
SALUD-2010-01-139945, SALUD-2012-01-181641). We thank
Ms. D. Alma Lidia Olivares from Hospital Infantil de Mexico
‘‘Federico Gomez’’ for providing the R6 strain used in this study.
References
Andrews SC, Robinson AK, Rodrıguez-Quinones F (2003) Bac-
terial iron homeostasis. FEMS Microbiol Rev 27:215–237
Asuthkar S (2007) Expression and characterization of an iron-
regulated hemin-binding protein, HbpA, from Lepto-
spira interrogans serovar. Lai. Infect Immun 75:4582–
4591
Brown JS, Gilliland SM, Holden DW (2001) A Streptococcus
pneumoniae pathogenicity island encoding an ABC trans-
porter involved in iron uptake and virulence. Mol Micro-
biol 40:572–585
Brown JS, Gilliland SM, Ruiz-Alber J, Holden DW (2004)
Characterization of pit, a Streptococcus pneumoniae iron
uptake ABC transporter. Infect Immun 70:4389–4398
Choi CW, Lee YG, Kwon SO, Kim HY, Lee JC, Chung YH,
Yun CY, Kim SI (2012) Analysis of Streptococcus pneu-
moniae secreted antigens by immuno-proteomic approach.
Diagn Microbiol Infect Dis 4:318–327
Cruz-Castaneda A, Lopez-Casamichana M, Olivares-Trejo JJ
(2011) Entamoeba histolytica secretes two haem-binding
proteins to scavenge haem. Biochem J 434:105–111
Erales J, Lignon S, Gontero B (2009) CP12 from Chlamydo-
monas reinhardtii, a permanent specific ‘‘chaperone-like’’
Biometals
123
Page 11
protein of glyceraldehyde-3- phosphate dehydrogenase.
J Biol Chem 284:12735–12744
Gonzalez-Lopez MA, Velazquez-Guadarrama N, Romero-Es-
pejel ME, Olivares-Trejo JJ (2013) Helicobacter pylori
secretes the chaperonin GroEL (HSP60), which binds iron.
FEBS Lett 587:1823–1828
Hannibal L, Collins D, Brassard J, Chakravarti R, Vempati R,
Dorlet P, Santolini J, Dawson JH, Stuehr DJ (2012) Heme
binding properties of glyceraldehyde-3-phosphate dehy-
drogenase. Biochemistry 51:8514–8529
Mazzola JL, Sirover MA (2001) Reduction of glyceraldehyde-
3-phosphate dehydrogenase activity in Alzheimer’s dis-
ease and in Huntington’s disease fibroblasts. J Neurochem
76:442–449
Miller JH (1972) Experiments in molecular genetics. Cold
Spring Harbor Laboratory, Cold Spring Harbor
Oliveira L, Madureira P, Andrade EB, Bouaboud A, Morello E,
Ferreira P, Poyart C, Trieu-Cuot P, Dramsi S (2012) Group
B streptococcus GAPDH is released upon cell lysis, asso-
ciates with bacterial surface, and induces apoptosis in
murine macrophages. PLoS One 7(1):e29963
Pancholi V, Chhatwal GS (2003) Housekeeping enzymes as
virulence factors for pathogens. Int J Med Microbiol
293:391–401
Reidl J, Mekalanos JJ (1996) Lipoprotein e(P4) is essential for
hemin uptake by Haemophilus influenzae. J Exp Med
183:621–629
Romero-Espejel ME, Gonzalez-Lopez MA, Olivares-Trejo JJ
(2013) Streptococcus pneumoniae requires iron for its
viability and expresses two membrane proteins that bind
haemoglobin and haem. Metallomics 5:384–389
Simpson W, Olczak T, Genco CA (2000) Characterization and
expression of HmuR, a TonB-dependent hemoglobin
receptor of Porphyromonas gingivalis. J Bacteriol
182:5737–5748
Sirover MA (1996) Minireview. Emerging new functions of the
glycolytic protein, glyceraldehyde-3-phosphate dehydro-
genase, in mammalian cells. Life Sci 58:2271–2277
Sirover MA (2011) On the functional diversity of glycer-
aldehyde-3-phosphate dehydrogenase: biochemical mech-
anisms and regulatory control. Biochim Biophys Acta
10:741–751
Tai SS, Lee CJ, Winter RE (1993) Hemin utilization is related to
virulence of Streptococcus pneumoniae. Infect Immun
61:5401–5405
Torres VJ, Pishchany G, Humayun M, Schneewind O, Skaar EP
(2006) Staphylococcus aureus IsdB is a hemoglobin
receptor required for heme iron utilization. J Bacteriol
188:8421–8429
Tuomanen EI, Mitchell TJ, Morrison D, Spratt BG (2004) The
Pneumococcus. ASM Press, Washington, DC, USA
Vanet A, Labigne A (1998) Evidence for specific secretion
rather than autolysis in the release of some Helicobacter
pylori proteins. Infect Immun 66:1023–1027
Wandersman C, Delepelaire P (2004) Bacterial iron sources:
from siderophores to hemophores. Annu Rev Microbiol
58:611–647
Wandersman C, Stojiljkovic I (2000) Bacterial heme sources:
the role of heme, hemoprotein receptors and hemophores.
Curr Opin Microbiol 3:215–220
Biometals
123