90
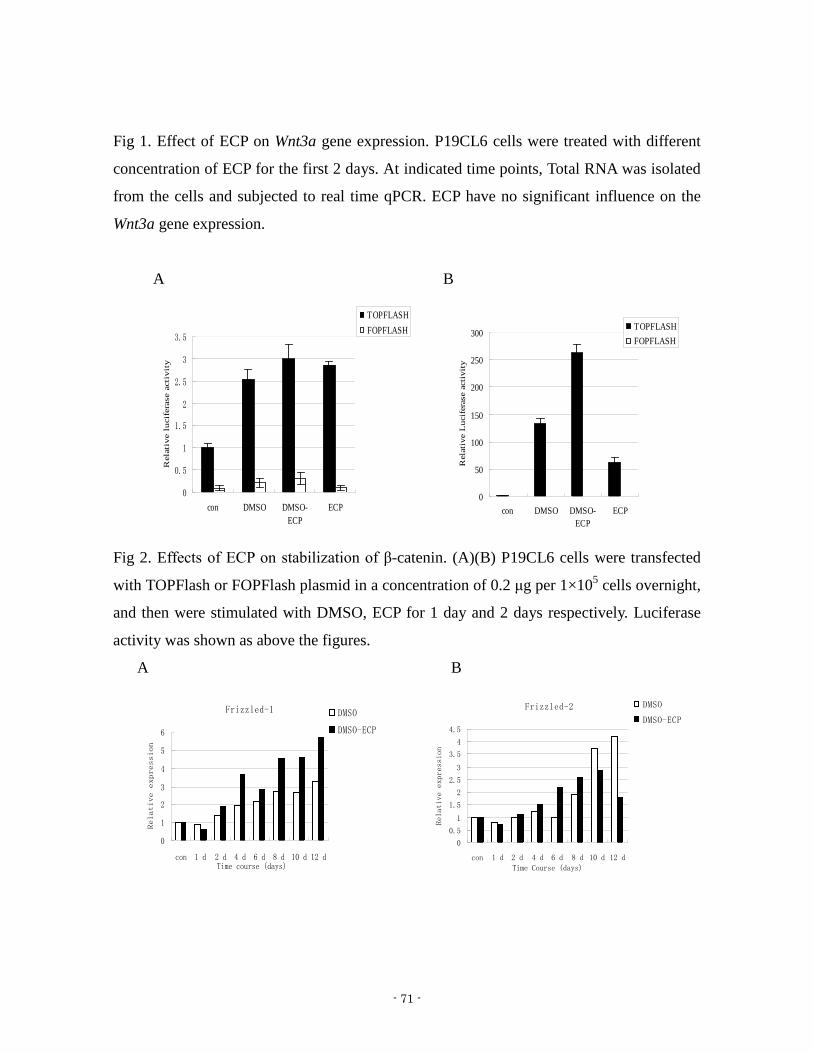
- 1 - Study on Cardiomyocyte Differentiation Induced by ECP 2012, September Guoliang JIN Graduate School of Natural Science and Technology (Doctor Course) OKAYAMA UNIVERSITY

- 1 -
Study on Cardiomyocyte Differentiation Induced by ECP
2012 September
Guoliang JIN
Graduate School of Natural Science and Technology
(Doctor Course)
OKAYAMA UNIVERSITY
- 2 -
- 3 -
CONTENTS
General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip4
Chapter 1 Effect of Eosinophil cationic protein (ECP) on
the cardiomyocyte differentiation in P19CL6 cells helliphelliphelliphelliphelliphelliphelliphellip21
Chapter 2 Effect of Eosinophil cationic protein (ECP) on
activation of FGF receptor 1 signaling during
cardiomyocyte differentiationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip37
Chapter 3 Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells helliphelliphelliphelliphelliphellip63
Concluding Remarkshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip88
Acknowledgmentshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip89
- 4 -
Introduction
The human eosinophil cationic protein (ECP) also known as RNase 3 is an eosinophil
secretion protein that is involved in innate immunity and displays antipathogen and
proinflammatory activities ECP is released during degranulation of eosinophils In human
ECP encoded by the RNase3 gene belongs to the RNase A superfamily ECP is a
single-chain zinc-containing protein with a molecular weight ranging from 16 to 22 kDa
The heterogeneity of the molecule is partially due to differences in glycosylation of three
potential sites in its amino acid chain
1 Molecular characteristics of ECP
Eosinophil cationic protein (ECP) generally speaking is a heterogeneous molecule
originating from activated eosinophil granulocytes At present as shown in figure 1 103
kinds of ECP or ECP similar proteins have been sequenced And most of them are from
mammalian especially from hominid
Fig1 Taxonomic group of organisms that ECP been sequenced (From
httpwwwncbinlmnihgovprotein)
- 5 -
Eosinophil cationic protein (ECP) that from human eosinophil granules was first isolated
and characterized by Gleich at al [1] They isolated ECP and another basic protein
eosinophil-derived neurotoxin (EDN) by gel filtration and ion exchange chromatography
on heparin-Sepharose NH2-terminal amino acid sequences of EDN and ECP from residue
1 through residue 55 showed a high similarity (67 homology) They also estimated that
the molecular mass of human ECP was 18-21 kD depending on the degree of glycosylation
Then Rosenberg et al did molecular cloning for ECP and reported the full-length cDNA
sequence The complete cDNA clone contains a Kozak-like translation initiation sequence
an open reading frame and a polyadenylation signal with a 15-base spacer preceding the
poly A tail The amino acid sequence encoded by the ECP cDNA confirms the identity of
residues identified by Gleich et al The mature ECP polypeptide (without leader sequence)
contains 133 amino acids which is one fewer than that found in EDN with a sequence
similarity between the two polypeptides of 66 The calculated molecular mass of mature
ECP is 15 6 kD and pI of ECP is 108 [2]
ECP and EDN sequences showed marked similarities to RNase from various species And
the amino acid sequence of both proteins showed a striking similarity with pancreatic-type
ribonucleases As shown in figure 2 they kept the specific residues of the active site (Gln11
His12 Lys41 and His119 RNase A numbering) and the eight Cys residues that form four
disulphide bridges Accordingly ECP and EDN were grouped in the human ribonuclease
family and have also been referred to as ribonuclease (RNase) 2 and 3 respectively [3 4]
In 2000 ECP has been cloned heterologously overexpressed purified and crystallized by
Mallorqui-Fernandez et al Its crystal structure has been determined and refined using data
up to 175 Aring resolutions The molecule displays the α+ β folding topology typical for
members of the ribonuclease A superfamily [5]
- 6 -
Fig2 Alignment of the amino acid sequences of ECP and the bovine pancreatic RNase A
The α-helices and β-strand of ECP are displayed as labelled dark rods and light arrows
respectively The topologically equivalent secondary structure elements in the enzymes with
known three-dimensional structures are shaded α-helices with dark grey background and
β-strands with light grey The upper umbering corresponds to ECP and the lower one to
RNase A (From Goretti et al 2000)
Table 1 Crystalized recombinant ECP for July 2012
Complex
crystalized together
Resolution PDB-code Reference
no 200 Aring 1QMT Biox et al 1999
no 175 Aring 1DYT Mallorqui-Fernandez et al 2000
2rsquo 5rsquo-ADP 240 Aring 1H1H Mohan et al 2002
sulfate anions 170 Aring - Biox et al 2012
- 7 -
Fig 3 Three-dimensional structure of human eosinophil cationic protein From Research
Collaboratory for Structural Bioinformatics (RCSB) protein data bank (PDB) accession
code 1DYT
Until now four crystalized ECP has been reported As shown in table 1 All the ECP were
recombinant type expressed in E coli BL21 (DE3) cell Crystals were grown by the
vapour diffusion hanging drop method
2 Functional characteristics and application of ECP
ECP as one of eosinophil secretion proteins involved in the host immune defense response
[6 7] has only been found in eosinophils granules and has not been detected in other
human tissues [8] Eosinophils were identified more than hundred years ago however their
roles in homeostasis and in disease still remain unclear The most prominent feature of the
eosinophils is their large secondary granules In the case of inflammation and asthma there
are increased levels of ECP in the body Therefore ECP has been developed as a biomarker
for eosinophilic disease and quantified in biological fluids including serum [9-12] saliva
[13 14] bronchoalveolar lavage [15] and nasal secretions [16] ECP as a biomarker was
studied firstly in Bronchial-Asthma by Bjornsson et al Then lots of researchers focus on it
- 8 -
elevated ECP levels have been found in activated asthma [9-11 14 15 17-22] and other
respiratory tract diseases such as allergic rhinitis and bacterial sinusitis [16] In recent years
researchers also found that ECP could be biomarkers in other diseases such as inflammatory
bowel diseases [23] collagenous colitis [24] atopic keratoconjunctivitis [25 26] and
toxocara infection [27]
On the other hand ECP as ribonuclease has very weak ribonuclease activity Compared
with EDN ECP shows 100-fold less activity than EDN against a yeast RNA substrate [2
28] With single-stranded RNA or synthetic polyribonucleotide substrates ECP showed
significant but low activity 70- to 200-fold less than that of bovine RNase A [29]
The low ribonuclease activity was related to its substrate subsites Boix et al investigated
ribonuclease activity of ECP through kinetic study Poly(C) poly(U) uridylyl(3rsquo5rsquo)
adenosine (UpA) uridylyl(3rsquo5rsquo) guanosine (UpG) cytidylyl(3rsquo5rsquo) adenosine (CpA)
uridine 2rsquo3rsquo-cyclic monophosphate (Ugtp) cytidine 2rsquo3rsquo-cyclic monophosphate (Cgtp) and
oligouridylic acids (Up)nUgtp n= 1ndash4 were used as substrates and the kinetic parameters
were determined In comparison with bovine pancreatic RNase A reduced catalytic
efficiency of ECP and the shift from an endonuclease to an exonuclease-type mechanism
may be caused by the differences in the multisubsites structure[30] Mallorqui-Fernandez et
al got consistent conclusion they crystallized ECP and found that the catalytic active site
residues are conserved with respect to other ribonucleases of the superfamily but some
differences appear at substrate recognition subsites which may account in part for the low
catalytic activity [5] In 2002 Mohan et al reported crystal structure of ECP in complex
with 2rsquo 5rsquo-ADP at 20 Aring resolution The study is the first detailed structural analysis of the
nucleotide recognition site in ECP Residues Gln-14 His-15 and Lys-38 make hydrogen
bond interactions with the phosphate at the P1 site while His-128 interacts with the purine
ring at the B2 site A new phosphate binding site P-1 has been identified which involves
Arg-34 [31]
Moreover ECP shows a wide range of biological activities in addition to its weak
ribonuclease activity ECPs biological activities related with eosinophils in most cases are
not associated with ribonuclease activity It has been shown to possess cytotoxic activity
- 9 -
neurotoxic activity and antimicrobial activity against bacteria pathogens and fungi
antiviral function and fibrosis promoting functions [6 32-35] Young et al reported that
purified human ECP could damage schistosomula of Schistosoma mansoni at low
concentrations Membrane damage was mediated by ECP through forming transmembrane
pores [33] Sorrentino et al treated EDN liver RNase and ECP with iodoacetic acid at
pH 55 which resulted in inactivation of their RNase activity and also destroyed their
neurotoxicity So they concluded that RNase activity is necessary but not sufficient to
induce neurotoxic action[36] However Rosenberg got the different conclusion Mutant
recombinant ECP that have no ribonuclease activity still shows antibacterial activity He
believed that ribonuclease activity and cytotoxicity are in that case independent functions
of ECP[37]About antiviral activity Domachowske et al prepared recombinant human ECP
(rhECP) prepared in baculovirus The rhECP was N-glycosylated and had similar to
100-fold more ribonuclease activity than nonglycosylated rhECP prepared in bacteria The
enzymatic activity of rhECP was sensitive to inhibition by placental ribonuclease inhibitor
(RI) And they found that rhECP promotes ribonuclease-dependent toxicity toward
extracellular virions of the single-stranded RNA virus respiratory syncytial virus group B
(RSV-B) [34]
On the basis of structure analysis ECP variants modified at basic and hydrophobic residues
have been constructed The bactericidal activity of both native ECP and point-mutated
variants were tested against Escherichia coli and Staphylococcus aureus Changes in the
leakage of liposome vesicles suggests that basic amino acids play in addition to the effect
on the disruption of the cellular membrane other roles such as specific binding on the
surface of the bacteria cell[38] Navarro et al analyzed ECP cytotoxic activity on
eukaryotic cell lines They found that ECP effects begin with its binding and aggregation to
the cell surface and then induce cell-specific morphological and biochemical changes such
as chromatin condensation reversion of membrane asymmetry reactive oxygen species
production and activation of caspase-3-like activity and eventually cell death They also
reported that the ribonuclease activity component of ECP is not involved in cytotoxic
activity as no RNA degradation is observed [39] Sanchez et al applied enzymatic and
- 10 -
chemical limited cleavage to search for active sequence determinants for antimicrobial
activity and confirm the main role of the protein N-terminal The results corroborated that
the 11-35 internal sequence and in particular the 28-35 stretch is essential The identified
region is involved in both ECP aggregation and bacteria envelope binding properties[40]
Torrent et al studied ECP aggregation they demonstrated that ECP is able to form amyloid
aggregates The amyloid protofibrils formed by ECP bind amyloid-diagnostic dyes as Th-T
and Congo Red present a fibrilar structure under electron microscopy They identified an
N-terminus hydrophobic patch (residues 8-16) that is required for the amyloid aggregation
process [41] Pulido et al found ECP has a remarkable affinity for lipopolysaccharide (LPS)
and a distinctive agglutinating activity By using a battery of LPS-truncated E coli mutant
strains they demonstrated that the polysaccharide moiety of LPS is essential for
ECP-mediated bacterial agglutination thereby modulating its antimicrobial action The
mechanism of action of ECP at the bacterial surface is drastically affected by the LPS
structure and in particular by its polysaccharide moiety They also analyzed an N-terminal
fragment that retains the whole protein activity and displays similar cell agglutination
behavior A fragment with further minimization of the antimicrobial domain though
retaining the antimicrobial capacity significantly loses its agglutinating activity exhibiting
a different mechanism of action which is not dependent on the LPS composition [42]
In 2012 Boix et al crystallized ECP in complex with sulfate anions in a new momoclinic
crystal form The study provides direct evidence of the main protein sulfate binding sites
Three main sites (S1-S3) are located in the protein active site involved in sulfated
heterosaccharide binding S1 and S2 overlap with the phosphate binding sites involved in
RNase nucleotide recognition A new site (S3) is one of the key anchoring points for
sulfated ligands In particular site S3 is unique to ECP in the RNase A superfamily This
may explaine ECP reduced ribonuclease activity and high affinity for glycosaminoglycans
[43]
Some researchers studied the effect of ECP on mammalian tumor cells Maeda et al
evaluated the effect of ECP on 13 mammalian cell lines They found that ECP inhibited the
growth of several cell lines including those derived from carcinoma and leukemia in a
- 11 -
dose-dependent manner ECP significantly suppressed the size of colonies of A431 cells
and decreased K562 cells in G(1)G(0) phase However there was little evidence that ECP
killed cells in either cell line Those results suggest that growth inhibition by ECP is
dependent on cell type and is cytostatic [44] Similar studies were also done on gastric
cancer cells[45] oral malignant tumor cells[46] and Hodgkin lymphoma cell[47]
Very recently we have demonstrated that eosinophil cationic protein (ECP) affected the
development of cytoskeleton in normal fibroblast cells and the differentiation of rat
neonatal cardiomyocytes[48] This finding implies that ECP might be useful as a
cardiomyocyte differentiation factor in the repair of cardiac tissue by stimulating the
differentiation of cardiomyocyte progenitor cells[48]
3 Cardiomyocyte differentiation
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [49 50] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [51 52] In this
context alternative approaches may include restoring heart function via induction of
endogenous regenerative processes and transferring progenitor cells to produce new
myocardium Cardiomyocytes are currently thought to be postmitotic cells that withdraw
from the cell cycle soon after birth Nevertheless mitoses in the myocyte population have
been detected in the human heart after myocardial infarction [53] Recent studies suggest
that cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [54] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
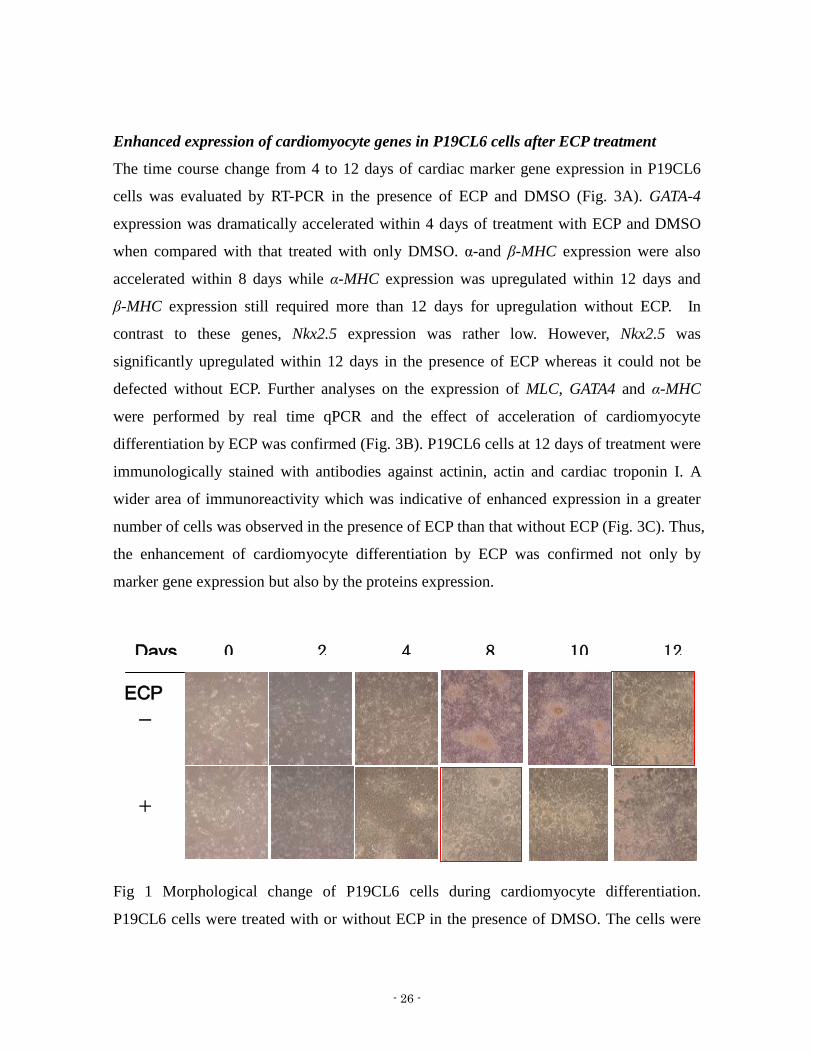
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
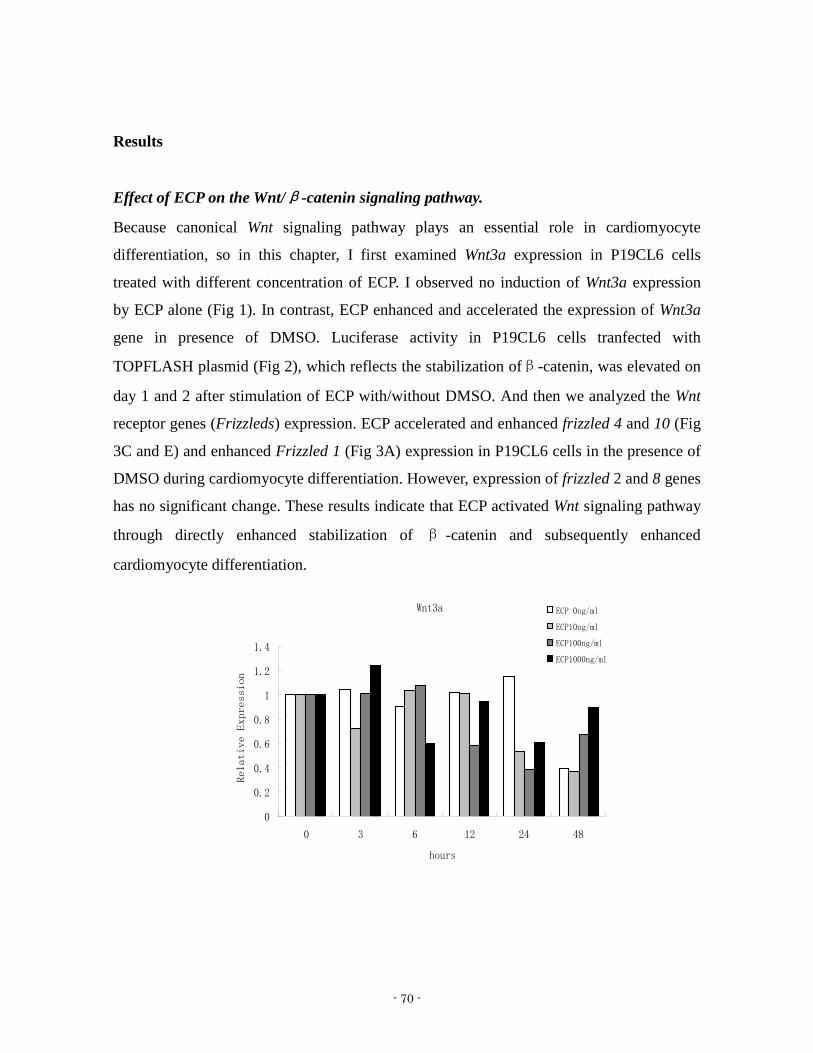
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
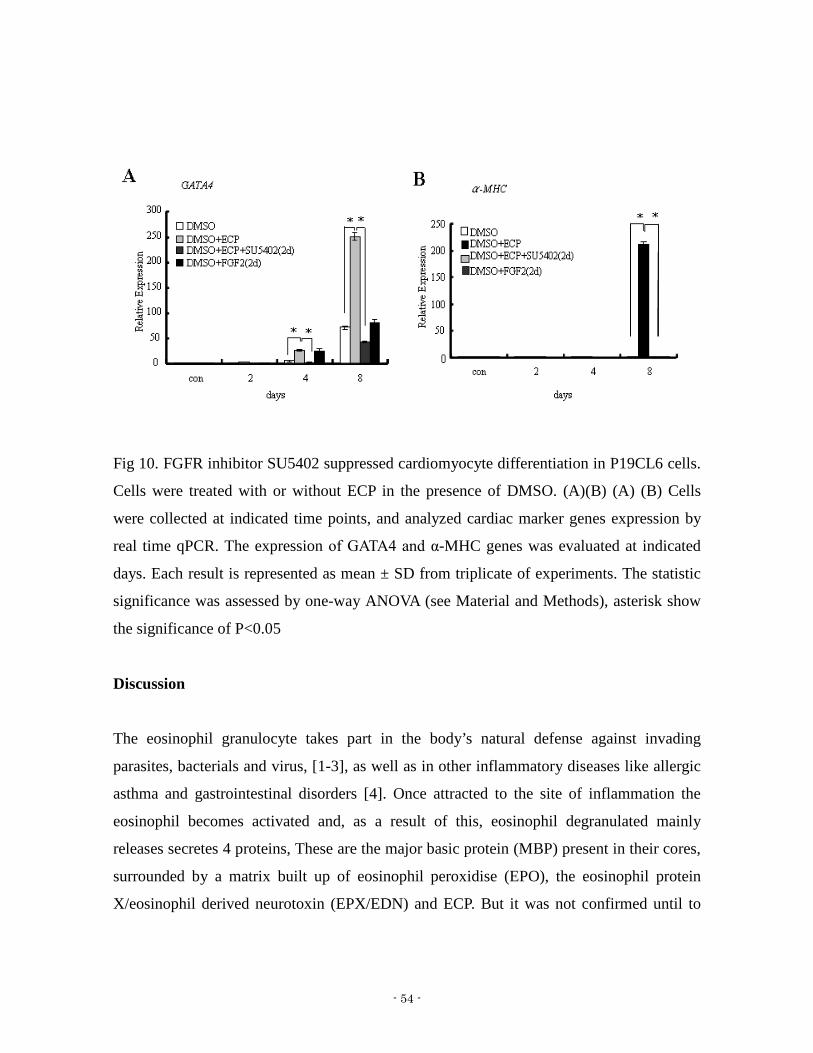
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 2 -
- 3 -
CONTENTS
General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip4
Chapter 1 Effect of Eosinophil cationic protein (ECP) on
the cardiomyocyte differentiation in P19CL6 cells helliphelliphelliphelliphelliphelliphelliphellip21
Chapter 2 Effect of Eosinophil cationic protein (ECP) on
activation of FGF receptor 1 signaling during
cardiomyocyte differentiationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip37
Chapter 3 Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells helliphelliphelliphelliphelliphellip63
Concluding Remarkshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip88
Acknowledgmentshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip89
- 4 -
Introduction
The human eosinophil cationic protein (ECP) also known as RNase 3 is an eosinophil
secretion protein that is involved in innate immunity and displays antipathogen and
proinflammatory activities ECP is released during degranulation of eosinophils In human
ECP encoded by the RNase3 gene belongs to the RNase A superfamily ECP is a
single-chain zinc-containing protein with a molecular weight ranging from 16 to 22 kDa
The heterogeneity of the molecule is partially due to differences in glycosylation of three
potential sites in its amino acid chain
1 Molecular characteristics of ECP
Eosinophil cationic protein (ECP) generally speaking is a heterogeneous molecule
originating from activated eosinophil granulocytes At present as shown in figure 1 103
kinds of ECP or ECP similar proteins have been sequenced And most of them are from
mammalian especially from hominid
Fig1 Taxonomic group of organisms that ECP been sequenced (From
httpwwwncbinlmnihgovprotein)
- 5 -
Eosinophil cationic protein (ECP) that from human eosinophil granules was first isolated
and characterized by Gleich at al [1] They isolated ECP and another basic protein
eosinophil-derived neurotoxin (EDN) by gel filtration and ion exchange chromatography
on heparin-Sepharose NH2-terminal amino acid sequences of EDN and ECP from residue
1 through residue 55 showed a high similarity (67 homology) They also estimated that
the molecular mass of human ECP was 18-21 kD depending on the degree of glycosylation
Then Rosenberg et al did molecular cloning for ECP and reported the full-length cDNA
sequence The complete cDNA clone contains a Kozak-like translation initiation sequence
an open reading frame and a polyadenylation signal with a 15-base spacer preceding the
poly A tail The amino acid sequence encoded by the ECP cDNA confirms the identity of
residues identified by Gleich et al The mature ECP polypeptide (without leader sequence)
contains 133 amino acids which is one fewer than that found in EDN with a sequence
similarity between the two polypeptides of 66 The calculated molecular mass of mature
ECP is 15 6 kD and pI of ECP is 108 [2]
ECP and EDN sequences showed marked similarities to RNase from various species And
the amino acid sequence of both proteins showed a striking similarity with pancreatic-type
ribonucleases As shown in figure 2 they kept the specific residues of the active site (Gln11
His12 Lys41 and His119 RNase A numbering) and the eight Cys residues that form four
disulphide bridges Accordingly ECP and EDN were grouped in the human ribonuclease
family and have also been referred to as ribonuclease (RNase) 2 and 3 respectively [3 4]
In 2000 ECP has been cloned heterologously overexpressed purified and crystallized by
Mallorqui-Fernandez et al Its crystal structure has been determined and refined using data
up to 175 Aring resolutions The molecule displays the α+ β folding topology typical for
members of the ribonuclease A superfamily [5]
- 6 -
Fig2 Alignment of the amino acid sequences of ECP and the bovine pancreatic RNase A
The α-helices and β-strand of ECP are displayed as labelled dark rods and light arrows
respectively The topologically equivalent secondary structure elements in the enzymes with
known three-dimensional structures are shaded α-helices with dark grey background and
β-strands with light grey The upper umbering corresponds to ECP and the lower one to
RNase A (From Goretti et al 2000)
Table 1 Crystalized recombinant ECP for July 2012
Complex
crystalized together
Resolution PDB-code Reference
no 200 Aring 1QMT Biox et al 1999
no 175 Aring 1DYT Mallorqui-Fernandez et al 2000
2rsquo 5rsquo-ADP 240 Aring 1H1H Mohan et al 2002
sulfate anions 170 Aring - Biox et al 2012
- 7 -
Fig 3 Three-dimensional structure of human eosinophil cationic protein From Research
Collaboratory for Structural Bioinformatics (RCSB) protein data bank (PDB) accession
code 1DYT
Until now four crystalized ECP has been reported As shown in table 1 All the ECP were
recombinant type expressed in E coli BL21 (DE3) cell Crystals were grown by the
vapour diffusion hanging drop method
2 Functional characteristics and application of ECP
ECP as one of eosinophil secretion proteins involved in the host immune defense response
[6 7] has only been found in eosinophils granules and has not been detected in other
human tissues [8] Eosinophils were identified more than hundred years ago however their
roles in homeostasis and in disease still remain unclear The most prominent feature of the
eosinophils is their large secondary granules In the case of inflammation and asthma there
are increased levels of ECP in the body Therefore ECP has been developed as a biomarker
for eosinophilic disease and quantified in biological fluids including serum [9-12] saliva
[13 14] bronchoalveolar lavage [15] and nasal secretions [16] ECP as a biomarker was
studied firstly in Bronchial-Asthma by Bjornsson et al Then lots of researchers focus on it
- 8 -
elevated ECP levels have been found in activated asthma [9-11 14 15 17-22] and other
respiratory tract diseases such as allergic rhinitis and bacterial sinusitis [16] In recent years
researchers also found that ECP could be biomarkers in other diseases such as inflammatory
bowel diseases [23] collagenous colitis [24] atopic keratoconjunctivitis [25 26] and
toxocara infection [27]
On the other hand ECP as ribonuclease has very weak ribonuclease activity Compared
with EDN ECP shows 100-fold less activity than EDN against a yeast RNA substrate [2
28] With single-stranded RNA or synthetic polyribonucleotide substrates ECP showed
significant but low activity 70- to 200-fold less than that of bovine RNase A [29]
The low ribonuclease activity was related to its substrate subsites Boix et al investigated
ribonuclease activity of ECP through kinetic study Poly(C) poly(U) uridylyl(3rsquo5rsquo)
adenosine (UpA) uridylyl(3rsquo5rsquo) guanosine (UpG) cytidylyl(3rsquo5rsquo) adenosine (CpA)
uridine 2rsquo3rsquo-cyclic monophosphate (Ugtp) cytidine 2rsquo3rsquo-cyclic monophosphate (Cgtp) and
oligouridylic acids (Up)nUgtp n= 1ndash4 were used as substrates and the kinetic parameters
were determined In comparison with bovine pancreatic RNase A reduced catalytic
efficiency of ECP and the shift from an endonuclease to an exonuclease-type mechanism
may be caused by the differences in the multisubsites structure[30] Mallorqui-Fernandez et
al got consistent conclusion they crystallized ECP and found that the catalytic active site
residues are conserved with respect to other ribonucleases of the superfamily but some
differences appear at substrate recognition subsites which may account in part for the low
catalytic activity [5] In 2002 Mohan et al reported crystal structure of ECP in complex
with 2rsquo 5rsquo-ADP at 20 Aring resolution The study is the first detailed structural analysis of the
nucleotide recognition site in ECP Residues Gln-14 His-15 and Lys-38 make hydrogen
bond interactions with the phosphate at the P1 site while His-128 interacts with the purine
ring at the B2 site A new phosphate binding site P-1 has been identified which involves
Arg-34 [31]
Moreover ECP shows a wide range of biological activities in addition to its weak
ribonuclease activity ECPs biological activities related with eosinophils in most cases are
not associated with ribonuclease activity It has been shown to possess cytotoxic activity
- 9 -
neurotoxic activity and antimicrobial activity against bacteria pathogens and fungi
antiviral function and fibrosis promoting functions [6 32-35] Young et al reported that
purified human ECP could damage schistosomula of Schistosoma mansoni at low
concentrations Membrane damage was mediated by ECP through forming transmembrane
pores [33] Sorrentino et al treated EDN liver RNase and ECP with iodoacetic acid at
pH 55 which resulted in inactivation of their RNase activity and also destroyed their
neurotoxicity So they concluded that RNase activity is necessary but not sufficient to
induce neurotoxic action[36] However Rosenberg got the different conclusion Mutant
recombinant ECP that have no ribonuclease activity still shows antibacterial activity He
believed that ribonuclease activity and cytotoxicity are in that case independent functions
of ECP[37]About antiviral activity Domachowske et al prepared recombinant human ECP
(rhECP) prepared in baculovirus The rhECP was N-glycosylated and had similar to
100-fold more ribonuclease activity than nonglycosylated rhECP prepared in bacteria The
enzymatic activity of rhECP was sensitive to inhibition by placental ribonuclease inhibitor
(RI) And they found that rhECP promotes ribonuclease-dependent toxicity toward
extracellular virions of the single-stranded RNA virus respiratory syncytial virus group B
(RSV-B) [34]
On the basis of structure analysis ECP variants modified at basic and hydrophobic residues
have been constructed The bactericidal activity of both native ECP and point-mutated
variants were tested against Escherichia coli and Staphylococcus aureus Changes in the
leakage of liposome vesicles suggests that basic amino acids play in addition to the effect
on the disruption of the cellular membrane other roles such as specific binding on the
surface of the bacteria cell[38] Navarro et al analyzed ECP cytotoxic activity on
eukaryotic cell lines They found that ECP effects begin with its binding and aggregation to
the cell surface and then induce cell-specific morphological and biochemical changes such
as chromatin condensation reversion of membrane asymmetry reactive oxygen species
production and activation of caspase-3-like activity and eventually cell death They also
reported that the ribonuclease activity component of ECP is not involved in cytotoxic
activity as no RNA degradation is observed [39] Sanchez et al applied enzymatic and
- 10 -
chemical limited cleavage to search for active sequence determinants for antimicrobial
activity and confirm the main role of the protein N-terminal The results corroborated that
the 11-35 internal sequence and in particular the 28-35 stretch is essential The identified
region is involved in both ECP aggregation and bacteria envelope binding properties[40]
Torrent et al studied ECP aggregation they demonstrated that ECP is able to form amyloid
aggregates The amyloid protofibrils formed by ECP bind amyloid-diagnostic dyes as Th-T
and Congo Red present a fibrilar structure under electron microscopy They identified an
N-terminus hydrophobic patch (residues 8-16) that is required for the amyloid aggregation
process [41] Pulido et al found ECP has a remarkable affinity for lipopolysaccharide (LPS)
and a distinctive agglutinating activity By using a battery of LPS-truncated E coli mutant
strains they demonstrated that the polysaccharide moiety of LPS is essential for
ECP-mediated bacterial agglutination thereby modulating its antimicrobial action The
mechanism of action of ECP at the bacterial surface is drastically affected by the LPS
structure and in particular by its polysaccharide moiety They also analyzed an N-terminal
fragment that retains the whole protein activity and displays similar cell agglutination
behavior A fragment with further minimization of the antimicrobial domain though
retaining the antimicrobial capacity significantly loses its agglutinating activity exhibiting
a different mechanism of action which is not dependent on the LPS composition [42]
In 2012 Boix et al crystallized ECP in complex with sulfate anions in a new momoclinic
crystal form The study provides direct evidence of the main protein sulfate binding sites
Three main sites (S1-S3) are located in the protein active site involved in sulfated
heterosaccharide binding S1 and S2 overlap with the phosphate binding sites involved in
RNase nucleotide recognition A new site (S3) is one of the key anchoring points for
sulfated ligands In particular site S3 is unique to ECP in the RNase A superfamily This
may explaine ECP reduced ribonuclease activity and high affinity for glycosaminoglycans
[43]
Some researchers studied the effect of ECP on mammalian tumor cells Maeda et al
evaluated the effect of ECP on 13 mammalian cell lines They found that ECP inhibited the
growth of several cell lines including those derived from carcinoma and leukemia in a
- 11 -
dose-dependent manner ECP significantly suppressed the size of colonies of A431 cells
and decreased K562 cells in G(1)G(0) phase However there was little evidence that ECP
killed cells in either cell line Those results suggest that growth inhibition by ECP is
dependent on cell type and is cytostatic [44] Similar studies were also done on gastric
cancer cells[45] oral malignant tumor cells[46] and Hodgkin lymphoma cell[47]
Very recently we have demonstrated that eosinophil cationic protein (ECP) affected the
development of cytoskeleton in normal fibroblast cells and the differentiation of rat
neonatal cardiomyocytes[48] This finding implies that ECP might be useful as a
cardiomyocyte differentiation factor in the repair of cardiac tissue by stimulating the
differentiation of cardiomyocyte progenitor cells[48]
3 Cardiomyocyte differentiation
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [49 50] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [51 52] In this
context alternative approaches may include restoring heart function via induction of
endogenous regenerative processes and transferring progenitor cells to produce new
myocardium Cardiomyocytes are currently thought to be postmitotic cells that withdraw
from the cell cycle soon after birth Nevertheless mitoses in the myocyte population have
been detected in the human heart after myocardial infarction [53] Recent studies suggest
that cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [54] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 3 -
CONTENTS
General Introductionhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip4
Chapter 1 Effect of Eosinophil cationic protein (ECP) on
the cardiomyocyte differentiation in P19CL6 cells helliphelliphelliphelliphelliphelliphelliphellip21
Chapter 2 Effect of Eosinophil cationic protein (ECP) on
activation of FGF receptor 1 signaling during
cardiomyocyte differentiationhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip37
Chapter 3 Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells helliphelliphelliphelliphelliphellip63
Concluding Remarkshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip88
Acknowledgmentshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip89
- 4 -
Introduction
The human eosinophil cationic protein (ECP) also known as RNase 3 is an eosinophil
secretion protein that is involved in innate immunity and displays antipathogen and
proinflammatory activities ECP is released during degranulation of eosinophils In human
ECP encoded by the RNase3 gene belongs to the RNase A superfamily ECP is a
single-chain zinc-containing protein with a molecular weight ranging from 16 to 22 kDa
The heterogeneity of the molecule is partially due to differences in glycosylation of three
potential sites in its amino acid chain
1 Molecular characteristics of ECP
Eosinophil cationic protein (ECP) generally speaking is a heterogeneous molecule
originating from activated eosinophil granulocytes At present as shown in figure 1 103
kinds of ECP or ECP similar proteins have been sequenced And most of them are from
mammalian especially from hominid
Fig1 Taxonomic group of organisms that ECP been sequenced (From
httpwwwncbinlmnihgovprotein)
- 5 -
Eosinophil cationic protein (ECP) that from human eosinophil granules was first isolated
and characterized by Gleich at al [1] They isolated ECP and another basic protein
eosinophil-derived neurotoxin (EDN) by gel filtration and ion exchange chromatography
on heparin-Sepharose NH2-terminal amino acid sequences of EDN and ECP from residue
1 through residue 55 showed a high similarity (67 homology) They also estimated that
the molecular mass of human ECP was 18-21 kD depending on the degree of glycosylation
Then Rosenberg et al did molecular cloning for ECP and reported the full-length cDNA
sequence The complete cDNA clone contains a Kozak-like translation initiation sequence
an open reading frame and a polyadenylation signal with a 15-base spacer preceding the
poly A tail The amino acid sequence encoded by the ECP cDNA confirms the identity of
residues identified by Gleich et al The mature ECP polypeptide (without leader sequence)
contains 133 amino acids which is one fewer than that found in EDN with a sequence
similarity between the two polypeptides of 66 The calculated molecular mass of mature
ECP is 15 6 kD and pI of ECP is 108 [2]
ECP and EDN sequences showed marked similarities to RNase from various species And
the amino acid sequence of both proteins showed a striking similarity with pancreatic-type
ribonucleases As shown in figure 2 they kept the specific residues of the active site (Gln11
His12 Lys41 and His119 RNase A numbering) and the eight Cys residues that form four
disulphide bridges Accordingly ECP and EDN were grouped in the human ribonuclease
family and have also been referred to as ribonuclease (RNase) 2 and 3 respectively [3 4]
In 2000 ECP has been cloned heterologously overexpressed purified and crystallized by
Mallorqui-Fernandez et al Its crystal structure has been determined and refined using data
up to 175 Aring resolutions The molecule displays the α+ β folding topology typical for
members of the ribonuclease A superfamily [5]
- 6 -
Fig2 Alignment of the amino acid sequences of ECP and the bovine pancreatic RNase A
The α-helices and β-strand of ECP are displayed as labelled dark rods and light arrows
respectively The topologically equivalent secondary structure elements in the enzymes with
known three-dimensional structures are shaded α-helices with dark grey background and
β-strands with light grey The upper umbering corresponds to ECP and the lower one to
RNase A (From Goretti et al 2000)
Table 1 Crystalized recombinant ECP for July 2012
Complex
crystalized together
Resolution PDB-code Reference
no 200 Aring 1QMT Biox et al 1999
no 175 Aring 1DYT Mallorqui-Fernandez et al 2000
2rsquo 5rsquo-ADP 240 Aring 1H1H Mohan et al 2002
sulfate anions 170 Aring - Biox et al 2012
- 7 -
Fig 3 Three-dimensional structure of human eosinophil cationic protein From Research
Collaboratory for Structural Bioinformatics (RCSB) protein data bank (PDB) accession
code 1DYT
Until now four crystalized ECP has been reported As shown in table 1 All the ECP were
recombinant type expressed in E coli BL21 (DE3) cell Crystals were grown by the
vapour diffusion hanging drop method
2 Functional characteristics and application of ECP
ECP as one of eosinophil secretion proteins involved in the host immune defense response
[6 7] has only been found in eosinophils granules and has not been detected in other
human tissues [8] Eosinophils were identified more than hundred years ago however their
roles in homeostasis and in disease still remain unclear The most prominent feature of the
eosinophils is their large secondary granules In the case of inflammation and asthma there
are increased levels of ECP in the body Therefore ECP has been developed as a biomarker
for eosinophilic disease and quantified in biological fluids including serum [9-12] saliva
[13 14] bronchoalveolar lavage [15] and nasal secretions [16] ECP as a biomarker was
studied firstly in Bronchial-Asthma by Bjornsson et al Then lots of researchers focus on it
- 8 -
elevated ECP levels have been found in activated asthma [9-11 14 15 17-22] and other
respiratory tract diseases such as allergic rhinitis and bacterial sinusitis [16] In recent years
researchers also found that ECP could be biomarkers in other diseases such as inflammatory
bowel diseases [23] collagenous colitis [24] atopic keratoconjunctivitis [25 26] and
toxocara infection [27]
On the other hand ECP as ribonuclease has very weak ribonuclease activity Compared
with EDN ECP shows 100-fold less activity than EDN against a yeast RNA substrate [2
28] With single-stranded RNA or synthetic polyribonucleotide substrates ECP showed
significant but low activity 70- to 200-fold less than that of bovine RNase A [29]
The low ribonuclease activity was related to its substrate subsites Boix et al investigated
ribonuclease activity of ECP through kinetic study Poly(C) poly(U) uridylyl(3rsquo5rsquo)
adenosine (UpA) uridylyl(3rsquo5rsquo) guanosine (UpG) cytidylyl(3rsquo5rsquo) adenosine (CpA)
uridine 2rsquo3rsquo-cyclic monophosphate (Ugtp) cytidine 2rsquo3rsquo-cyclic monophosphate (Cgtp) and
oligouridylic acids (Up)nUgtp n= 1ndash4 were used as substrates and the kinetic parameters
were determined In comparison with bovine pancreatic RNase A reduced catalytic
efficiency of ECP and the shift from an endonuclease to an exonuclease-type mechanism
may be caused by the differences in the multisubsites structure[30] Mallorqui-Fernandez et
al got consistent conclusion they crystallized ECP and found that the catalytic active site
residues are conserved with respect to other ribonucleases of the superfamily but some
differences appear at substrate recognition subsites which may account in part for the low
catalytic activity [5] In 2002 Mohan et al reported crystal structure of ECP in complex
with 2rsquo 5rsquo-ADP at 20 Aring resolution The study is the first detailed structural analysis of the
nucleotide recognition site in ECP Residues Gln-14 His-15 and Lys-38 make hydrogen
bond interactions with the phosphate at the P1 site while His-128 interacts with the purine
ring at the B2 site A new phosphate binding site P-1 has been identified which involves
Arg-34 [31]
Moreover ECP shows a wide range of biological activities in addition to its weak
ribonuclease activity ECPs biological activities related with eosinophils in most cases are
not associated with ribonuclease activity It has been shown to possess cytotoxic activity
- 9 -
neurotoxic activity and antimicrobial activity against bacteria pathogens and fungi
antiviral function and fibrosis promoting functions [6 32-35] Young et al reported that
purified human ECP could damage schistosomula of Schistosoma mansoni at low
concentrations Membrane damage was mediated by ECP through forming transmembrane
pores [33] Sorrentino et al treated EDN liver RNase and ECP with iodoacetic acid at
pH 55 which resulted in inactivation of their RNase activity and also destroyed their
neurotoxicity So they concluded that RNase activity is necessary but not sufficient to
induce neurotoxic action[36] However Rosenberg got the different conclusion Mutant
recombinant ECP that have no ribonuclease activity still shows antibacterial activity He
believed that ribonuclease activity and cytotoxicity are in that case independent functions
of ECP[37]About antiviral activity Domachowske et al prepared recombinant human ECP
(rhECP) prepared in baculovirus The rhECP was N-glycosylated and had similar to
100-fold more ribonuclease activity than nonglycosylated rhECP prepared in bacteria The
enzymatic activity of rhECP was sensitive to inhibition by placental ribonuclease inhibitor
(RI) And they found that rhECP promotes ribonuclease-dependent toxicity toward
extracellular virions of the single-stranded RNA virus respiratory syncytial virus group B
(RSV-B) [34]
On the basis of structure analysis ECP variants modified at basic and hydrophobic residues
have been constructed The bactericidal activity of both native ECP and point-mutated
variants were tested against Escherichia coli and Staphylococcus aureus Changes in the
leakage of liposome vesicles suggests that basic amino acids play in addition to the effect
on the disruption of the cellular membrane other roles such as specific binding on the
surface of the bacteria cell[38] Navarro et al analyzed ECP cytotoxic activity on
eukaryotic cell lines They found that ECP effects begin with its binding and aggregation to
the cell surface and then induce cell-specific morphological and biochemical changes such
as chromatin condensation reversion of membrane asymmetry reactive oxygen species
production and activation of caspase-3-like activity and eventually cell death They also
reported that the ribonuclease activity component of ECP is not involved in cytotoxic
activity as no RNA degradation is observed [39] Sanchez et al applied enzymatic and
- 10 -
chemical limited cleavage to search for active sequence determinants for antimicrobial
activity and confirm the main role of the protein N-terminal The results corroborated that
the 11-35 internal sequence and in particular the 28-35 stretch is essential The identified
region is involved in both ECP aggregation and bacteria envelope binding properties[40]
Torrent et al studied ECP aggregation they demonstrated that ECP is able to form amyloid
aggregates The amyloid protofibrils formed by ECP bind amyloid-diagnostic dyes as Th-T
and Congo Red present a fibrilar structure under electron microscopy They identified an
N-terminus hydrophobic patch (residues 8-16) that is required for the amyloid aggregation
process [41] Pulido et al found ECP has a remarkable affinity for lipopolysaccharide (LPS)
and a distinctive agglutinating activity By using a battery of LPS-truncated E coli mutant
strains they demonstrated that the polysaccharide moiety of LPS is essential for
ECP-mediated bacterial agglutination thereby modulating its antimicrobial action The
mechanism of action of ECP at the bacterial surface is drastically affected by the LPS
structure and in particular by its polysaccharide moiety They also analyzed an N-terminal
fragment that retains the whole protein activity and displays similar cell agglutination
behavior A fragment with further minimization of the antimicrobial domain though
retaining the antimicrobial capacity significantly loses its agglutinating activity exhibiting
a different mechanism of action which is not dependent on the LPS composition [42]
In 2012 Boix et al crystallized ECP in complex with sulfate anions in a new momoclinic
crystal form The study provides direct evidence of the main protein sulfate binding sites
Three main sites (S1-S3) are located in the protein active site involved in sulfated
heterosaccharide binding S1 and S2 overlap with the phosphate binding sites involved in
RNase nucleotide recognition A new site (S3) is one of the key anchoring points for
sulfated ligands In particular site S3 is unique to ECP in the RNase A superfamily This
may explaine ECP reduced ribonuclease activity and high affinity for glycosaminoglycans
[43]
Some researchers studied the effect of ECP on mammalian tumor cells Maeda et al
evaluated the effect of ECP on 13 mammalian cell lines They found that ECP inhibited the
growth of several cell lines including those derived from carcinoma and leukemia in a
- 11 -
dose-dependent manner ECP significantly suppressed the size of colonies of A431 cells
and decreased K562 cells in G(1)G(0) phase However there was little evidence that ECP
killed cells in either cell line Those results suggest that growth inhibition by ECP is
dependent on cell type and is cytostatic [44] Similar studies were also done on gastric
cancer cells[45] oral malignant tumor cells[46] and Hodgkin lymphoma cell[47]
Very recently we have demonstrated that eosinophil cationic protein (ECP) affected the
development of cytoskeleton in normal fibroblast cells and the differentiation of rat
neonatal cardiomyocytes[48] This finding implies that ECP might be useful as a
cardiomyocyte differentiation factor in the repair of cardiac tissue by stimulating the
differentiation of cardiomyocyte progenitor cells[48]
3 Cardiomyocyte differentiation
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [49 50] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [51 52] In this
context alternative approaches may include restoring heart function via induction of
endogenous regenerative processes and transferring progenitor cells to produce new
myocardium Cardiomyocytes are currently thought to be postmitotic cells that withdraw
from the cell cycle soon after birth Nevertheless mitoses in the myocyte population have
been detected in the human heart after myocardial infarction [53] Recent studies suggest
that cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [54] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 4 -
Introduction
The human eosinophil cationic protein (ECP) also known as RNase 3 is an eosinophil
secretion protein that is involved in innate immunity and displays antipathogen and
proinflammatory activities ECP is released during degranulation of eosinophils In human
ECP encoded by the RNase3 gene belongs to the RNase A superfamily ECP is a
single-chain zinc-containing protein with a molecular weight ranging from 16 to 22 kDa
The heterogeneity of the molecule is partially due to differences in glycosylation of three
potential sites in its amino acid chain
1 Molecular characteristics of ECP
Eosinophil cationic protein (ECP) generally speaking is a heterogeneous molecule
originating from activated eosinophil granulocytes At present as shown in figure 1 103
kinds of ECP or ECP similar proteins have been sequenced And most of them are from
mammalian especially from hominid
Fig1 Taxonomic group of organisms that ECP been sequenced (From
httpwwwncbinlmnihgovprotein)
- 5 -
Eosinophil cationic protein (ECP) that from human eosinophil granules was first isolated
and characterized by Gleich at al [1] They isolated ECP and another basic protein
eosinophil-derived neurotoxin (EDN) by gel filtration and ion exchange chromatography
on heparin-Sepharose NH2-terminal amino acid sequences of EDN and ECP from residue
1 through residue 55 showed a high similarity (67 homology) They also estimated that
the molecular mass of human ECP was 18-21 kD depending on the degree of glycosylation
Then Rosenberg et al did molecular cloning for ECP and reported the full-length cDNA
sequence The complete cDNA clone contains a Kozak-like translation initiation sequence
an open reading frame and a polyadenylation signal with a 15-base spacer preceding the
poly A tail The amino acid sequence encoded by the ECP cDNA confirms the identity of
residues identified by Gleich et al The mature ECP polypeptide (without leader sequence)
contains 133 amino acids which is one fewer than that found in EDN with a sequence
similarity between the two polypeptides of 66 The calculated molecular mass of mature
ECP is 15 6 kD and pI of ECP is 108 [2]
ECP and EDN sequences showed marked similarities to RNase from various species And
the amino acid sequence of both proteins showed a striking similarity with pancreatic-type
ribonucleases As shown in figure 2 they kept the specific residues of the active site (Gln11
His12 Lys41 and His119 RNase A numbering) and the eight Cys residues that form four
disulphide bridges Accordingly ECP and EDN were grouped in the human ribonuclease
family and have also been referred to as ribonuclease (RNase) 2 and 3 respectively [3 4]
In 2000 ECP has been cloned heterologously overexpressed purified and crystallized by
Mallorqui-Fernandez et al Its crystal structure has been determined and refined using data
up to 175 Aring resolutions The molecule displays the α+ β folding topology typical for
members of the ribonuclease A superfamily [5]
- 6 -
Fig2 Alignment of the amino acid sequences of ECP and the bovine pancreatic RNase A
The α-helices and β-strand of ECP are displayed as labelled dark rods and light arrows
respectively The topologically equivalent secondary structure elements in the enzymes with
known three-dimensional structures are shaded α-helices with dark grey background and
β-strands with light grey The upper umbering corresponds to ECP and the lower one to
RNase A (From Goretti et al 2000)
Table 1 Crystalized recombinant ECP for July 2012
Complex
crystalized together
Resolution PDB-code Reference
no 200 Aring 1QMT Biox et al 1999
no 175 Aring 1DYT Mallorqui-Fernandez et al 2000
2rsquo 5rsquo-ADP 240 Aring 1H1H Mohan et al 2002
sulfate anions 170 Aring - Biox et al 2012
- 7 -
Fig 3 Three-dimensional structure of human eosinophil cationic protein From Research
Collaboratory for Structural Bioinformatics (RCSB) protein data bank (PDB) accession
code 1DYT
Until now four crystalized ECP has been reported As shown in table 1 All the ECP were
recombinant type expressed in E coli BL21 (DE3) cell Crystals were grown by the
vapour diffusion hanging drop method
2 Functional characteristics and application of ECP
ECP as one of eosinophil secretion proteins involved in the host immune defense response
[6 7] has only been found in eosinophils granules and has not been detected in other
human tissues [8] Eosinophils were identified more than hundred years ago however their
roles in homeostasis and in disease still remain unclear The most prominent feature of the
eosinophils is their large secondary granules In the case of inflammation and asthma there
are increased levels of ECP in the body Therefore ECP has been developed as a biomarker
for eosinophilic disease and quantified in biological fluids including serum [9-12] saliva
[13 14] bronchoalveolar lavage [15] and nasal secretions [16] ECP as a biomarker was
studied firstly in Bronchial-Asthma by Bjornsson et al Then lots of researchers focus on it
- 8 -
elevated ECP levels have been found in activated asthma [9-11 14 15 17-22] and other
respiratory tract diseases such as allergic rhinitis and bacterial sinusitis [16] In recent years
researchers also found that ECP could be biomarkers in other diseases such as inflammatory
bowel diseases [23] collagenous colitis [24] atopic keratoconjunctivitis [25 26] and
toxocara infection [27]
On the other hand ECP as ribonuclease has very weak ribonuclease activity Compared
with EDN ECP shows 100-fold less activity than EDN against a yeast RNA substrate [2
28] With single-stranded RNA or synthetic polyribonucleotide substrates ECP showed
significant but low activity 70- to 200-fold less than that of bovine RNase A [29]
The low ribonuclease activity was related to its substrate subsites Boix et al investigated
ribonuclease activity of ECP through kinetic study Poly(C) poly(U) uridylyl(3rsquo5rsquo)
adenosine (UpA) uridylyl(3rsquo5rsquo) guanosine (UpG) cytidylyl(3rsquo5rsquo) adenosine (CpA)
uridine 2rsquo3rsquo-cyclic monophosphate (Ugtp) cytidine 2rsquo3rsquo-cyclic monophosphate (Cgtp) and
oligouridylic acids (Up)nUgtp n= 1ndash4 were used as substrates and the kinetic parameters
were determined In comparison with bovine pancreatic RNase A reduced catalytic
efficiency of ECP and the shift from an endonuclease to an exonuclease-type mechanism
may be caused by the differences in the multisubsites structure[30] Mallorqui-Fernandez et
al got consistent conclusion they crystallized ECP and found that the catalytic active site
residues are conserved with respect to other ribonucleases of the superfamily but some
differences appear at substrate recognition subsites which may account in part for the low
catalytic activity [5] In 2002 Mohan et al reported crystal structure of ECP in complex
with 2rsquo 5rsquo-ADP at 20 Aring resolution The study is the first detailed structural analysis of the
nucleotide recognition site in ECP Residues Gln-14 His-15 and Lys-38 make hydrogen
bond interactions with the phosphate at the P1 site while His-128 interacts with the purine
ring at the B2 site A new phosphate binding site P-1 has been identified which involves
Arg-34 [31]
Moreover ECP shows a wide range of biological activities in addition to its weak
ribonuclease activity ECPs biological activities related with eosinophils in most cases are
not associated with ribonuclease activity It has been shown to possess cytotoxic activity
- 9 -
neurotoxic activity and antimicrobial activity against bacteria pathogens and fungi
antiviral function and fibrosis promoting functions [6 32-35] Young et al reported that
purified human ECP could damage schistosomula of Schistosoma mansoni at low
concentrations Membrane damage was mediated by ECP through forming transmembrane
pores [33] Sorrentino et al treated EDN liver RNase and ECP with iodoacetic acid at
pH 55 which resulted in inactivation of their RNase activity and also destroyed their
neurotoxicity So they concluded that RNase activity is necessary but not sufficient to
induce neurotoxic action[36] However Rosenberg got the different conclusion Mutant
recombinant ECP that have no ribonuclease activity still shows antibacterial activity He
believed that ribonuclease activity and cytotoxicity are in that case independent functions
of ECP[37]About antiviral activity Domachowske et al prepared recombinant human ECP
(rhECP) prepared in baculovirus The rhECP was N-glycosylated and had similar to
100-fold more ribonuclease activity than nonglycosylated rhECP prepared in bacteria The
enzymatic activity of rhECP was sensitive to inhibition by placental ribonuclease inhibitor
(RI) And they found that rhECP promotes ribonuclease-dependent toxicity toward
extracellular virions of the single-stranded RNA virus respiratory syncytial virus group B
(RSV-B) [34]
On the basis of structure analysis ECP variants modified at basic and hydrophobic residues
have been constructed The bactericidal activity of both native ECP and point-mutated
variants were tested against Escherichia coli and Staphylococcus aureus Changes in the
leakage of liposome vesicles suggests that basic amino acids play in addition to the effect
on the disruption of the cellular membrane other roles such as specific binding on the
surface of the bacteria cell[38] Navarro et al analyzed ECP cytotoxic activity on
eukaryotic cell lines They found that ECP effects begin with its binding and aggregation to
the cell surface and then induce cell-specific morphological and biochemical changes such
as chromatin condensation reversion of membrane asymmetry reactive oxygen species
production and activation of caspase-3-like activity and eventually cell death They also
reported that the ribonuclease activity component of ECP is not involved in cytotoxic
activity as no RNA degradation is observed [39] Sanchez et al applied enzymatic and
- 10 -
chemical limited cleavage to search for active sequence determinants for antimicrobial
activity and confirm the main role of the protein N-terminal The results corroborated that
the 11-35 internal sequence and in particular the 28-35 stretch is essential The identified
region is involved in both ECP aggregation and bacteria envelope binding properties[40]
Torrent et al studied ECP aggregation they demonstrated that ECP is able to form amyloid
aggregates The amyloid protofibrils formed by ECP bind amyloid-diagnostic dyes as Th-T
and Congo Red present a fibrilar structure under electron microscopy They identified an
N-terminus hydrophobic patch (residues 8-16) that is required for the amyloid aggregation
process [41] Pulido et al found ECP has a remarkable affinity for lipopolysaccharide (LPS)
and a distinctive agglutinating activity By using a battery of LPS-truncated E coli mutant
strains they demonstrated that the polysaccharide moiety of LPS is essential for
ECP-mediated bacterial agglutination thereby modulating its antimicrobial action The
mechanism of action of ECP at the bacterial surface is drastically affected by the LPS
structure and in particular by its polysaccharide moiety They also analyzed an N-terminal
fragment that retains the whole protein activity and displays similar cell agglutination
behavior A fragment with further minimization of the antimicrobial domain though
retaining the antimicrobial capacity significantly loses its agglutinating activity exhibiting
a different mechanism of action which is not dependent on the LPS composition [42]
In 2012 Boix et al crystallized ECP in complex with sulfate anions in a new momoclinic
crystal form The study provides direct evidence of the main protein sulfate binding sites
Three main sites (S1-S3) are located in the protein active site involved in sulfated
heterosaccharide binding S1 and S2 overlap with the phosphate binding sites involved in
RNase nucleotide recognition A new site (S3) is one of the key anchoring points for
sulfated ligands In particular site S3 is unique to ECP in the RNase A superfamily This
may explaine ECP reduced ribonuclease activity and high affinity for glycosaminoglycans
[43]
Some researchers studied the effect of ECP on mammalian tumor cells Maeda et al
evaluated the effect of ECP on 13 mammalian cell lines They found that ECP inhibited the
growth of several cell lines including those derived from carcinoma and leukemia in a
- 11 -
dose-dependent manner ECP significantly suppressed the size of colonies of A431 cells
and decreased K562 cells in G(1)G(0) phase However there was little evidence that ECP
killed cells in either cell line Those results suggest that growth inhibition by ECP is
dependent on cell type and is cytostatic [44] Similar studies were also done on gastric
cancer cells[45] oral malignant tumor cells[46] and Hodgkin lymphoma cell[47]
Very recently we have demonstrated that eosinophil cationic protein (ECP) affected the
development of cytoskeleton in normal fibroblast cells and the differentiation of rat
neonatal cardiomyocytes[48] This finding implies that ECP might be useful as a
cardiomyocyte differentiation factor in the repair of cardiac tissue by stimulating the
differentiation of cardiomyocyte progenitor cells[48]
3 Cardiomyocyte differentiation
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [49 50] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [51 52] In this
context alternative approaches may include restoring heart function via induction of
endogenous regenerative processes and transferring progenitor cells to produce new
myocardium Cardiomyocytes are currently thought to be postmitotic cells that withdraw
from the cell cycle soon after birth Nevertheless mitoses in the myocyte population have
been detected in the human heart after myocardial infarction [53] Recent studies suggest
that cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [54] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 5 -
Eosinophil cationic protein (ECP) that from human eosinophil granules was first isolated
and characterized by Gleich at al [1] They isolated ECP and another basic protein
eosinophil-derived neurotoxin (EDN) by gel filtration and ion exchange chromatography
on heparin-Sepharose NH2-terminal amino acid sequences of EDN and ECP from residue
1 through residue 55 showed a high similarity (67 homology) They also estimated that
the molecular mass of human ECP was 18-21 kD depending on the degree of glycosylation
Then Rosenberg et al did molecular cloning for ECP and reported the full-length cDNA
sequence The complete cDNA clone contains a Kozak-like translation initiation sequence
an open reading frame and a polyadenylation signal with a 15-base spacer preceding the
poly A tail The amino acid sequence encoded by the ECP cDNA confirms the identity of
residues identified by Gleich et al The mature ECP polypeptide (without leader sequence)
contains 133 amino acids which is one fewer than that found in EDN with a sequence
similarity between the two polypeptides of 66 The calculated molecular mass of mature
ECP is 15 6 kD and pI of ECP is 108 [2]
ECP and EDN sequences showed marked similarities to RNase from various species And
the amino acid sequence of both proteins showed a striking similarity with pancreatic-type
ribonucleases As shown in figure 2 they kept the specific residues of the active site (Gln11
His12 Lys41 and His119 RNase A numbering) and the eight Cys residues that form four
disulphide bridges Accordingly ECP and EDN were grouped in the human ribonuclease
family and have also been referred to as ribonuclease (RNase) 2 and 3 respectively [3 4]
In 2000 ECP has been cloned heterologously overexpressed purified and crystallized by
Mallorqui-Fernandez et al Its crystal structure has been determined and refined using data
up to 175 Aring resolutions The molecule displays the α+ β folding topology typical for
members of the ribonuclease A superfamily [5]
- 6 -
Fig2 Alignment of the amino acid sequences of ECP and the bovine pancreatic RNase A
The α-helices and β-strand of ECP are displayed as labelled dark rods and light arrows
respectively The topologically equivalent secondary structure elements in the enzymes with
known three-dimensional structures are shaded α-helices with dark grey background and
β-strands with light grey The upper umbering corresponds to ECP and the lower one to
RNase A (From Goretti et al 2000)
Table 1 Crystalized recombinant ECP for July 2012
Complex
crystalized together
Resolution PDB-code Reference
no 200 Aring 1QMT Biox et al 1999
no 175 Aring 1DYT Mallorqui-Fernandez et al 2000
2rsquo 5rsquo-ADP 240 Aring 1H1H Mohan et al 2002
sulfate anions 170 Aring - Biox et al 2012
- 7 -
Fig 3 Three-dimensional structure of human eosinophil cationic protein From Research
Collaboratory for Structural Bioinformatics (RCSB) protein data bank (PDB) accession
code 1DYT
Until now four crystalized ECP has been reported As shown in table 1 All the ECP were
recombinant type expressed in E coli BL21 (DE3) cell Crystals were grown by the
vapour diffusion hanging drop method
2 Functional characteristics and application of ECP
ECP as one of eosinophil secretion proteins involved in the host immune defense response
[6 7] has only been found in eosinophils granules and has not been detected in other
human tissues [8] Eosinophils were identified more than hundred years ago however their
roles in homeostasis and in disease still remain unclear The most prominent feature of the
eosinophils is their large secondary granules In the case of inflammation and asthma there
are increased levels of ECP in the body Therefore ECP has been developed as a biomarker
for eosinophilic disease and quantified in biological fluids including serum [9-12] saliva
[13 14] bronchoalveolar lavage [15] and nasal secretions [16] ECP as a biomarker was
studied firstly in Bronchial-Asthma by Bjornsson et al Then lots of researchers focus on it
- 8 -
elevated ECP levels have been found in activated asthma [9-11 14 15 17-22] and other
respiratory tract diseases such as allergic rhinitis and bacterial sinusitis [16] In recent years
researchers also found that ECP could be biomarkers in other diseases such as inflammatory
bowel diseases [23] collagenous colitis [24] atopic keratoconjunctivitis [25 26] and
toxocara infection [27]
On the other hand ECP as ribonuclease has very weak ribonuclease activity Compared
with EDN ECP shows 100-fold less activity than EDN against a yeast RNA substrate [2
28] With single-stranded RNA or synthetic polyribonucleotide substrates ECP showed
significant but low activity 70- to 200-fold less than that of bovine RNase A [29]
The low ribonuclease activity was related to its substrate subsites Boix et al investigated
ribonuclease activity of ECP through kinetic study Poly(C) poly(U) uridylyl(3rsquo5rsquo)
adenosine (UpA) uridylyl(3rsquo5rsquo) guanosine (UpG) cytidylyl(3rsquo5rsquo) adenosine (CpA)
uridine 2rsquo3rsquo-cyclic monophosphate (Ugtp) cytidine 2rsquo3rsquo-cyclic monophosphate (Cgtp) and
oligouridylic acids (Up)nUgtp n= 1ndash4 were used as substrates and the kinetic parameters
were determined In comparison with bovine pancreatic RNase A reduced catalytic
efficiency of ECP and the shift from an endonuclease to an exonuclease-type mechanism
may be caused by the differences in the multisubsites structure[30] Mallorqui-Fernandez et
al got consistent conclusion they crystallized ECP and found that the catalytic active site
residues are conserved with respect to other ribonucleases of the superfamily but some
differences appear at substrate recognition subsites which may account in part for the low
catalytic activity [5] In 2002 Mohan et al reported crystal structure of ECP in complex
with 2rsquo 5rsquo-ADP at 20 Aring resolution The study is the first detailed structural analysis of the
nucleotide recognition site in ECP Residues Gln-14 His-15 and Lys-38 make hydrogen
bond interactions with the phosphate at the P1 site while His-128 interacts with the purine
ring at the B2 site A new phosphate binding site P-1 has been identified which involves
Arg-34 [31]
Moreover ECP shows a wide range of biological activities in addition to its weak
ribonuclease activity ECPs biological activities related with eosinophils in most cases are
not associated with ribonuclease activity It has been shown to possess cytotoxic activity
- 9 -
neurotoxic activity and antimicrobial activity against bacteria pathogens and fungi
antiviral function and fibrosis promoting functions [6 32-35] Young et al reported that
purified human ECP could damage schistosomula of Schistosoma mansoni at low
concentrations Membrane damage was mediated by ECP through forming transmembrane
pores [33] Sorrentino et al treated EDN liver RNase and ECP with iodoacetic acid at
pH 55 which resulted in inactivation of their RNase activity and also destroyed their
neurotoxicity So they concluded that RNase activity is necessary but not sufficient to
induce neurotoxic action[36] However Rosenberg got the different conclusion Mutant
recombinant ECP that have no ribonuclease activity still shows antibacterial activity He
believed that ribonuclease activity and cytotoxicity are in that case independent functions
of ECP[37]About antiviral activity Domachowske et al prepared recombinant human ECP
(rhECP) prepared in baculovirus The rhECP was N-glycosylated and had similar to
100-fold more ribonuclease activity than nonglycosylated rhECP prepared in bacteria The
enzymatic activity of rhECP was sensitive to inhibition by placental ribonuclease inhibitor
(RI) And they found that rhECP promotes ribonuclease-dependent toxicity toward
extracellular virions of the single-stranded RNA virus respiratory syncytial virus group B
(RSV-B) [34]
On the basis of structure analysis ECP variants modified at basic and hydrophobic residues
have been constructed The bactericidal activity of both native ECP and point-mutated
variants were tested against Escherichia coli and Staphylococcus aureus Changes in the
leakage of liposome vesicles suggests that basic amino acids play in addition to the effect
on the disruption of the cellular membrane other roles such as specific binding on the
surface of the bacteria cell[38] Navarro et al analyzed ECP cytotoxic activity on
eukaryotic cell lines They found that ECP effects begin with its binding and aggregation to
the cell surface and then induce cell-specific morphological and biochemical changes such
as chromatin condensation reversion of membrane asymmetry reactive oxygen species
production and activation of caspase-3-like activity and eventually cell death They also
reported that the ribonuclease activity component of ECP is not involved in cytotoxic
activity as no RNA degradation is observed [39] Sanchez et al applied enzymatic and
- 10 -
chemical limited cleavage to search for active sequence determinants for antimicrobial
activity and confirm the main role of the protein N-terminal The results corroborated that
the 11-35 internal sequence and in particular the 28-35 stretch is essential The identified
region is involved in both ECP aggregation and bacteria envelope binding properties[40]
Torrent et al studied ECP aggregation they demonstrated that ECP is able to form amyloid
aggregates The amyloid protofibrils formed by ECP bind amyloid-diagnostic dyes as Th-T
and Congo Red present a fibrilar structure under electron microscopy They identified an
N-terminus hydrophobic patch (residues 8-16) that is required for the amyloid aggregation
process [41] Pulido et al found ECP has a remarkable affinity for lipopolysaccharide (LPS)
and a distinctive agglutinating activity By using a battery of LPS-truncated E coli mutant
strains they demonstrated that the polysaccharide moiety of LPS is essential for
ECP-mediated bacterial agglutination thereby modulating its antimicrobial action The
mechanism of action of ECP at the bacterial surface is drastically affected by the LPS
structure and in particular by its polysaccharide moiety They also analyzed an N-terminal
fragment that retains the whole protein activity and displays similar cell agglutination
behavior A fragment with further minimization of the antimicrobial domain though
retaining the antimicrobial capacity significantly loses its agglutinating activity exhibiting
a different mechanism of action which is not dependent on the LPS composition [42]
In 2012 Boix et al crystallized ECP in complex with sulfate anions in a new momoclinic
crystal form The study provides direct evidence of the main protein sulfate binding sites
Three main sites (S1-S3) are located in the protein active site involved in sulfated
heterosaccharide binding S1 and S2 overlap with the phosphate binding sites involved in
RNase nucleotide recognition A new site (S3) is one of the key anchoring points for
sulfated ligands In particular site S3 is unique to ECP in the RNase A superfamily This
may explaine ECP reduced ribonuclease activity and high affinity for glycosaminoglycans
[43]
Some researchers studied the effect of ECP on mammalian tumor cells Maeda et al
evaluated the effect of ECP on 13 mammalian cell lines They found that ECP inhibited the
growth of several cell lines including those derived from carcinoma and leukemia in a
- 11 -
dose-dependent manner ECP significantly suppressed the size of colonies of A431 cells
and decreased K562 cells in G(1)G(0) phase However there was little evidence that ECP
killed cells in either cell line Those results suggest that growth inhibition by ECP is
dependent on cell type and is cytostatic [44] Similar studies were also done on gastric
cancer cells[45] oral malignant tumor cells[46] and Hodgkin lymphoma cell[47]
Very recently we have demonstrated that eosinophil cationic protein (ECP) affected the
development of cytoskeleton in normal fibroblast cells and the differentiation of rat
neonatal cardiomyocytes[48] This finding implies that ECP might be useful as a
cardiomyocyte differentiation factor in the repair of cardiac tissue by stimulating the
differentiation of cardiomyocyte progenitor cells[48]
3 Cardiomyocyte differentiation
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [49 50] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [51 52] In this
context alternative approaches may include restoring heart function via induction of
endogenous regenerative processes and transferring progenitor cells to produce new
myocardium Cardiomyocytes are currently thought to be postmitotic cells that withdraw
from the cell cycle soon after birth Nevertheless mitoses in the myocyte population have
been detected in the human heart after myocardial infarction [53] Recent studies suggest
that cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [54] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 6 -
Fig2 Alignment of the amino acid sequences of ECP and the bovine pancreatic RNase A
The α-helices and β-strand of ECP are displayed as labelled dark rods and light arrows
respectively The topologically equivalent secondary structure elements in the enzymes with
known three-dimensional structures are shaded α-helices with dark grey background and
β-strands with light grey The upper umbering corresponds to ECP and the lower one to
RNase A (From Goretti et al 2000)
Table 1 Crystalized recombinant ECP for July 2012
Complex
crystalized together
Resolution PDB-code Reference
no 200 Aring 1QMT Biox et al 1999
no 175 Aring 1DYT Mallorqui-Fernandez et al 2000
2rsquo 5rsquo-ADP 240 Aring 1H1H Mohan et al 2002
sulfate anions 170 Aring - Biox et al 2012
- 7 -
Fig 3 Three-dimensional structure of human eosinophil cationic protein From Research
Collaboratory for Structural Bioinformatics (RCSB) protein data bank (PDB) accession
code 1DYT
Until now four crystalized ECP has been reported As shown in table 1 All the ECP were
recombinant type expressed in E coli BL21 (DE3) cell Crystals were grown by the
vapour diffusion hanging drop method
2 Functional characteristics and application of ECP
ECP as one of eosinophil secretion proteins involved in the host immune defense response
[6 7] has only been found in eosinophils granules and has not been detected in other
human tissues [8] Eosinophils were identified more than hundred years ago however their
roles in homeostasis and in disease still remain unclear The most prominent feature of the
eosinophils is their large secondary granules In the case of inflammation and asthma there
are increased levels of ECP in the body Therefore ECP has been developed as a biomarker
for eosinophilic disease and quantified in biological fluids including serum [9-12] saliva
[13 14] bronchoalveolar lavage [15] and nasal secretions [16] ECP as a biomarker was
studied firstly in Bronchial-Asthma by Bjornsson et al Then lots of researchers focus on it
- 8 -
elevated ECP levels have been found in activated asthma [9-11 14 15 17-22] and other
respiratory tract diseases such as allergic rhinitis and bacterial sinusitis [16] In recent years
researchers also found that ECP could be biomarkers in other diseases such as inflammatory
bowel diseases [23] collagenous colitis [24] atopic keratoconjunctivitis [25 26] and
toxocara infection [27]
On the other hand ECP as ribonuclease has very weak ribonuclease activity Compared
with EDN ECP shows 100-fold less activity than EDN against a yeast RNA substrate [2
28] With single-stranded RNA or synthetic polyribonucleotide substrates ECP showed
significant but low activity 70- to 200-fold less than that of bovine RNase A [29]
The low ribonuclease activity was related to its substrate subsites Boix et al investigated
ribonuclease activity of ECP through kinetic study Poly(C) poly(U) uridylyl(3rsquo5rsquo)
adenosine (UpA) uridylyl(3rsquo5rsquo) guanosine (UpG) cytidylyl(3rsquo5rsquo) adenosine (CpA)
uridine 2rsquo3rsquo-cyclic monophosphate (Ugtp) cytidine 2rsquo3rsquo-cyclic monophosphate (Cgtp) and
oligouridylic acids (Up)nUgtp n= 1ndash4 were used as substrates and the kinetic parameters
were determined In comparison with bovine pancreatic RNase A reduced catalytic
efficiency of ECP and the shift from an endonuclease to an exonuclease-type mechanism
may be caused by the differences in the multisubsites structure[30] Mallorqui-Fernandez et
al got consistent conclusion they crystallized ECP and found that the catalytic active site
residues are conserved with respect to other ribonucleases of the superfamily but some
differences appear at substrate recognition subsites which may account in part for the low
catalytic activity [5] In 2002 Mohan et al reported crystal structure of ECP in complex
with 2rsquo 5rsquo-ADP at 20 Aring resolution The study is the first detailed structural analysis of the
nucleotide recognition site in ECP Residues Gln-14 His-15 and Lys-38 make hydrogen
bond interactions with the phosphate at the P1 site while His-128 interacts with the purine
ring at the B2 site A new phosphate binding site P-1 has been identified which involves
Arg-34 [31]
Moreover ECP shows a wide range of biological activities in addition to its weak
ribonuclease activity ECPs biological activities related with eosinophils in most cases are
not associated with ribonuclease activity It has been shown to possess cytotoxic activity
- 9 -
neurotoxic activity and antimicrobial activity against bacteria pathogens and fungi
antiviral function and fibrosis promoting functions [6 32-35] Young et al reported that
purified human ECP could damage schistosomula of Schistosoma mansoni at low
concentrations Membrane damage was mediated by ECP through forming transmembrane
pores [33] Sorrentino et al treated EDN liver RNase and ECP with iodoacetic acid at
pH 55 which resulted in inactivation of their RNase activity and also destroyed their
neurotoxicity So they concluded that RNase activity is necessary but not sufficient to
induce neurotoxic action[36] However Rosenberg got the different conclusion Mutant
recombinant ECP that have no ribonuclease activity still shows antibacterial activity He
believed that ribonuclease activity and cytotoxicity are in that case independent functions
of ECP[37]About antiviral activity Domachowske et al prepared recombinant human ECP
(rhECP) prepared in baculovirus The rhECP was N-glycosylated and had similar to
100-fold more ribonuclease activity than nonglycosylated rhECP prepared in bacteria The
enzymatic activity of rhECP was sensitive to inhibition by placental ribonuclease inhibitor
(RI) And they found that rhECP promotes ribonuclease-dependent toxicity toward
extracellular virions of the single-stranded RNA virus respiratory syncytial virus group B
(RSV-B) [34]
On the basis of structure analysis ECP variants modified at basic and hydrophobic residues
have been constructed The bactericidal activity of both native ECP and point-mutated
variants were tested against Escherichia coli and Staphylococcus aureus Changes in the
leakage of liposome vesicles suggests that basic amino acids play in addition to the effect
on the disruption of the cellular membrane other roles such as specific binding on the
surface of the bacteria cell[38] Navarro et al analyzed ECP cytotoxic activity on
eukaryotic cell lines They found that ECP effects begin with its binding and aggregation to
the cell surface and then induce cell-specific morphological and biochemical changes such
as chromatin condensation reversion of membrane asymmetry reactive oxygen species
production and activation of caspase-3-like activity and eventually cell death They also
reported that the ribonuclease activity component of ECP is not involved in cytotoxic
activity as no RNA degradation is observed [39] Sanchez et al applied enzymatic and
- 10 -
chemical limited cleavage to search for active sequence determinants for antimicrobial
activity and confirm the main role of the protein N-terminal The results corroborated that
the 11-35 internal sequence and in particular the 28-35 stretch is essential The identified
region is involved in both ECP aggregation and bacteria envelope binding properties[40]
Torrent et al studied ECP aggregation they demonstrated that ECP is able to form amyloid
aggregates The amyloid protofibrils formed by ECP bind amyloid-diagnostic dyes as Th-T
and Congo Red present a fibrilar structure under electron microscopy They identified an
N-terminus hydrophobic patch (residues 8-16) that is required for the amyloid aggregation
process [41] Pulido et al found ECP has a remarkable affinity for lipopolysaccharide (LPS)
and a distinctive agglutinating activity By using a battery of LPS-truncated E coli mutant
strains they demonstrated that the polysaccharide moiety of LPS is essential for
ECP-mediated bacterial agglutination thereby modulating its antimicrobial action The
mechanism of action of ECP at the bacterial surface is drastically affected by the LPS
structure and in particular by its polysaccharide moiety They also analyzed an N-terminal
fragment that retains the whole protein activity and displays similar cell agglutination
behavior A fragment with further minimization of the antimicrobial domain though
retaining the antimicrobial capacity significantly loses its agglutinating activity exhibiting
a different mechanism of action which is not dependent on the LPS composition [42]
In 2012 Boix et al crystallized ECP in complex with sulfate anions in a new momoclinic
crystal form The study provides direct evidence of the main protein sulfate binding sites
Three main sites (S1-S3) are located in the protein active site involved in sulfated
heterosaccharide binding S1 and S2 overlap with the phosphate binding sites involved in
RNase nucleotide recognition A new site (S3) is one of the key anchoring points for
sulfated ligands In particular site S3 is unique to ECP in the RNase A superfamily This
may explaine ECP reduced ribonuclease activity and high affinity for glycosaminoglycans
[43]
Some researchers studied the effect of ECP on mammalian tumor cells Maeda et al
evaluated the effect of ECP on 13 mammalian cell lines They found that ECP inhibited the
growth of several cell lines including those derived from carcinoma and leukemia in a
- 11 -
dose-dependent manner ECP significantly suppressed the size of colonies of A431 cells
and decreased K562 cells in G(1)G(0) phase However there was little evidence that ECP
killed cells in either cell line Those results suggest that growth inhibition by ECP is
dependent on cell type and is cytostatic [44] Similar studies were also done on gastric
cancer cells[45] oral malignant tumor cells[46] and Hodgkin lymphoma cell[47]
Very recently we have demonstrated that eosinophil cationic protein (ECP) affected the
development of cytoskeleton in normal fibroblast cells and the differentiation of rat
neonatal cardiomyocytes[48] This finding implies that ECP might be useful as a
cardiomyocyte differentiation factor in the repair of cardiac tissue by stimulating the
differentiation of cardiomyocyte progenitor cells[48]
3 Cardiomyocyte differentiation
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [49 50] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [51 52] In this
context alternative approaches may include restoring heart function via induction of
endogenous regenerative processes and transferring progenitor cells to produce new
myocardium Cardiomyocytes are currently thought to be postmitotic cells that withdraw
from the cell cycle soon after birth Nevertheless mitoses in the myocyte population have
been detected in the human heart after myocardial infarction [53] Recent studies suggest
that cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [54] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 7 -
Fig 3 Three-dimensional structure of human eosinophil cationic protein From Research
Collaboratory for Structural Bioinformatics (RCSB) protein data bank (PDB) accession
code 1DYT
Until now four crystalized ECP has been reported As shown in table 1 All the ECP were
recombinant type expressed in E coli BL21 (DE3) cell Crystals were grown by the
vapour diffusion hanging drop method
2 Functional characteristics and application of ECP
ECP as one of eosinophil secretion proteins involved in the host immune defense response
[6 7] has only been found in eosinophils granules and has not been detected in other
human tissues [8] Eosinophils were identified more than hundred years ago however their
roles in homeostasis and in disease still remain unclear The most prominent feature of the
eosinophils is their large secondary granules In the case of inflammation and asthma there
are increased levels of ECP in the body Therefore ECP has been developed as a biomarker
for eosinophilic disease and quantified in biological fluids including serum [9-12] saliva
[13 14] bronchoalveolar lavage [15] and nasal secretions [16] ECP as a biomarker was
studied firstly in Bronchial-Asthma by Bjornsson et al Then lots of researchers focus on it
- 8 -
elevated ECP levels have been found in activated asthma [9-11 14 15 17-22] and other
respiratory tract diseases such as allergic rhinitis and bacterial sinusitis [16] In recent years
researchers also found that ECP could be biomarkers in other diseases such as inflammatory
bowel diseases [23] collagenous colitis [24] atopic keratoconjunctivitis [25 26] and
toxocara infection [27]
On the other hand ECP as ribonuclease has very weak ribonuclease activity Compared
with EDN ECP shows 100-fold less activity than EDN against a yeast RNA substrate [2
28] With single-stranded RNA or synthetic polyribonucleotide substrates ECP showed
significant but low activity 70- to 200-fold less than that of bovine RNase A [29]
The low ribonuclease activity was related to its substrate subsites Boix et al investigated
ribonuclease activity of ECP through kinetic study Poly(C) poly(U) uridylyl(3rsquo5rsquo)
adenosine (UpA) uridylyl(3rsquo5rsquo) guanosine (UpG) cytidylyl(3rsquo5rsquo) adenosine (CpA)
uridine 2rsquo3rsquo-cyclic monophosphate (Ugtp) cytidine 2rsquo3rsquo-cyclic monophosphate (Cgtp) and
oligouridylic acids (Up)nUgtp n= 1ndash4 were used as substrates and the kinetic parameters
were determined In comparison with bovine pancreatic RNase A reduced catalytic
efficiency of ECP and the shift from an endonuclease to an exonuclease-type mechanism
may be caused by the differences in the multisubsites structure[30] Mallorqui-Fernandez et
al got consistent conclusion they crystallized ECP and found that the catalytic active site
residues are conserved with respect to other ribonucleases of the superfamily but some
differences appear at substrate recognition subsites which may account in part for the low
catalytic activity [5] In 2002 Mohan et al reported crystal structure of ECP in complex
with 2rsquo 5rsquo-ADP at 20 Aring resolution The study is the first detailed structural analysis of the
nucleotide recognition site in ECP Residues Gln-14 His-15 and Lys-38 make hydrogen
bond interactions with the phosphate at the P1 site while His-128 interacts with the purine
ring at the B2 site A new phosphate binding site P-1 has been identified which involves
Arg-34 [31]
Moreover ECP shows a wide range of biological activities in addition to its weak
ribonuclease activity ECPs biological activities related with eosinophils in most cases are
not associated with ribonuclease activity It has been shown to possess cytotoxic activity
- 9 -
neurotoxic activity and antimicrobial activity against bacteria pathogens and fungi
antiviral function and fibrosis promoting functions [6 32-35] Young et al reported that
purified human ECP could damage schistosomula of Schistosoma mansoni at low
concentrations Membrane damage was mediated by ECP through forming transmembrane
pores [33] Sorrentino et al treated EDN liver RNase and ECP with iodoacetic acid at
pH 55 which resulted in inactivation of their RNase activity and also destroyed their
neurotoxicity So they concluded that RNase activity is necessary but not sufficient to
induce neurotoxic action[36] However Rosenberg got the different conclusion Mutant
recombinant ECP that have no ribonuclease activity still shows antibacterial activity He
believed that ribonuclease activity and cytotoxicity are in that case independent functions
of ECP[37]About antiviral activity Domachowske et al prepared recombinant human ECP
(rhECP) prepared in baculovirus The rhECP was N-glycosylated and had similar to
100-fold more ribonuclease activity than nonglycosylated rhECP prepared in bacteria The
enzymatic activity of rhECP was sensitive to inhibition by placental ribonuclease inhibitor
(RI) And they found that rhECP promotes ribonuclease-dependent toxicity toward
extracellular virions of the single-stranded RNA virus respiratory syncytial virus group B
(RSV-B) [34]
On the basis of structure analysis ECP variants modified at basic and hydrophobic residues
have been constructed The bactericidal activity of both native ECP and point-mutated
variants were tested against Escherichia coli and Staphylococcus aureus Changes in the
leakage of liposome vesicles suggests that basic amino acids play in addition to the effect
on the disruption of the cellular membrane other roles such as specific binding on the
surface of the bacteria cell[38] Navarro et al analyzed ECP cytotoxic activity on
eukaryotic cell lines They found that ECP effects begin with its binding and aggregation to
the cell surface and then induce cell-specific morphological and biochemical changes such
as chromatin condensation reversion of membrane asymmetry reactive oxygen species
production and activation of caspase-3-like activity and eventually cell death They also
reported that the ribonuclease activity component of ECP is not involved in cytotoxic
activity as no RNA degradation is observed [39] Sanchez et al applied enzymatic and
- 10 -
chemical limited cleavage to search for active sequence determinants for antimicrobial
activity and confirm the main role of the protein N-terminal The results corroborated that
the 11-35 internal sequence and in particular the 28-35 stretch is essential The identified
region is involved in both ECP aggregation and bacteria envelope binding properties[40]
Torrent et al studied ECP aggregation they demonstrated that ECP is able to form amyloid
aggregates The amyloid protofibrils formed by ECP bind amyloid-diagnostic dyes as Th-T
and Congo Red present a fibrilar structure under electron microscopy They identified an
N-terminus hydrophobic patch (residues 8-16) that is required for the amyloid aggregation
process [41] Pulido et al found ECP has a remarkable affinity for lipopolysaccharide (LPS)
and a distinctive agglutinating activity By using a battery of LPS-truncated E coli mutant
strains they demonstrated that the polysaccharide moiety of LPS is essential for
ECP-mediated bacterial agglutination thereby modulating its antimicrobial action The
mechanism of action of ECP at the bacterial surface is drastically affected by the LPS
structure and in particular by its polysaccharide moiety They also analyzed an N-terminal
fragment that retains the whole protein activity and displays similar cell agglutination
behavior A fragment with further minimization of the antimicrobial domain though
retaining the antimicrobial capacity significantly loses its agglutinating activity exhibiting
a different mechanism of action which is not dependent on the LPS composition [42]
In 2012 Boix et al crystallized ECP in complex with sulfate anions in a new momoclinic
crystal form The study provides direct evidence of the main protein sulfate binding sites
Three main sites (S1-S3) are located in the protein active site involved in sulfated
heterosaccharide binding S1 and S2 overlap with the phosphate binding sites involved in
RNase nucleotide recognition A new site (S3) is one of the key anchoring points for
sulfated ligands In particular site S3 is unique to ECP in the RNase A superfamily This
may explaine ECP reduced ribonuclease activity and high affinity for glycosaminoglycans
[43]
Some researchers studied the effect of ECP on mammalian tumor cells Maeda et al
evaluated the effect of ECP on 13 mammalian cell lines They found that ECP inhibited the
growth of several cell lines including those derived from carcinoma and leukemia in a
- 11 -
dose-dependent manner ECP significantly suppressed the size of colonies of A431 cells
and decreased K562 cells in G(1)G(0) phase However there was little evidence that ECP
killed cells in either cell line Those results suggest that growth inhibition by ECP is
dependent on cell type and is cytostatic [44] Similar studies were also done on gastric
cancer cells[45] oral malignant tumor cells[46] and Hodgkin lymphoma cell[47]
Very recently we have demonstrated that eosinophil cationic protein (ECP) affected the
development of cytoskeleton in normal fibroblast cells and the differentiation of rat
neonatal cardiomyocytes[48] This finding implies that ECP might be useful as a
cardiomyocyte differentiation factor in the repair of cardiac tissue by stimulating the
differentiation of cardiomyocyte progenitor cells[48]
3 Cardiomyocyte differentiation
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [49 50] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [51 52] In this
context alternative approaches may include restoring heart function via induction of
endogenous regenerative processes and transferring progenitor cells to produce new
myocardium Cardiomyocytes are currently thought to be postmitotic cells that withdraw
from the cell cycle soon after birth Nevertheless mitoses in the myocyte population have
been detected in the human heart after myocardial infarction [53] Recent studies suggest
that cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [54] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 8 -
elevated ECP levels have been found in activated asthma [9-11 14 15 17-22] and other
respiratory tract diseases such as allergic rhinitis and bacterial sinusitis [16] In recent years
researchers also found that ECP could be biomarkers in other diseases such as inflammatory
bowel diseases [23] collagenous colitis [24] atopic keratoconjunctivitis [25 26] and
toxocara infection [27]
On the other hand ECP as ribonuclease has very weak ribonuclease activity Compared
with EDN ECP shows 100-fold less activity than EDN against a yeast RNA substrate [2
28] With single-stranded RNA or synthetic polyribonucleotide substrates ECP showed
significant but low activity 70- to 200-fold less than that of bovine RNase A [29]
The low ribonuclease activity was related to its substrate subsites Boix et al investigated
ribonuclease activity of ECP through kinetic study Poly(C) poly(U) uridylyl(3rsquo5rsquo)
adenosine (UpA) uridylyl(3rsquo5rsquo) guanosine (UpG) cytidylyl(3rsquo5rsquo) adenosine (CpA)
uridine 2rsquo3rsquo-cyclic monophosphate (Ugtp) cytidine 2rsquo3rsquo-cyclic monophosphate (Cgtp) and
oligouridylic acids (Up)nUgtp n= 1ndash4 were used as substrates and the kinetic parameters
were determined In comparison with bovine pancreatic RNase A reduced catalytic
efficiency of ECP and the shift from an endonuclease to an exonuclease-type mechanism
may be caused by the differences in the multisubsites structure[30] Mallorqui-Fernandez et
al got consistent conclusion they crystallized ECP and found that the catalytic active site
residues are conserved with respect to other ribonucleases of the superfamily but some
differences appear at substrate recognition subsites which may account in part for the low
catalytic activity [5] In 2002 Mohan et al reported crystal structure of ECP in complex
with 2rsquo 5rsquo-ADP at 20 Aring resolution The study is the first detailed structural analysis of the
nucleotide recognition site in ECP Residues Gln-14 His-15 and Lys-38 make hydrogen
bond interactions with the phosphate at the P1 site while His-128 interacts with the purine
ring at the B2 site A new phosphate binding site P-1 has been identified which involves
Arg-34 [31]
Moreover ECP shows a wide range of biological activities in addition to its weak
ribonuclease activity ECPs biological activities related with eosinophils in most cases are
not associated with ribonuclease activity It has been shown to possess cytotoxic activity
- 9 -
neurotoxic activity and antimicrobial activity against bacteria pathogens and fungi
antiviral function and fibrosis promoting functions [6 32-35] Young et al reported that
purified human ECP could damage schistosomula of Schistosoma mansoni at low
concentrations Membrane damage was mediated by ECP through forming transmembrane
pores [33] Sorrentino et al treated EDN liver RNase and ECP with iodoacetic acid at
pH 55 which resulted in inactivation of their RNase activity and also destroyed their
neurotoxicity So they concluded that RNase activity is necessary but not sufficient to
induce neurotoxic action[36] However Rosenberg got the different conclusion Mutant
recombinant ECP that have no ribonuclease activity still shows antibacterial activity He
believed that ribonuclease activity and cytotoxicity are in that case independent functions
of ECP[37]About antiviral activity Domachowske et al prepared recombinant human ECP
(rhECP) prepared in baculovirus The rhECP was N-glycosylated and had similar to
100-fold more ribonuclease activity than nonglycosylated rhECP prepared in bacteria The
enzymatic activity of rhECP was sensitive to inhibition by placental ribonuclease inhibitor
(RI) And they found that rhECP promotes ribonuclease-dependent toxicity toward
extracellular virions of the single-stranded RNA virus respiratory syncytial virus group B
(RSV-B) [34]
On the basis of structure analysis ECP variants modified at basic and hydrophobic residues
have been constructed The bactericidal activity of both native ECP and point-mutated
variants were tested against Escherichia coli and Staphylococcus aureus Changes in the
leakage of liposome vesicles suggests that basic amino acids play in addition to the effect
on the disruption of the cellular membrane other roles such as specific binding on the
surface of the bacteria cell[38] Navarro et al analyzed ECP cytotoxic activity on
eukaryotic cell lines They found that ECP effects begin with its binding and aggregation to
the cell surface and then induce cell-specific morphological and biochemical changes such
as chromatin condensation reversion of membrane asymmetry reactive oxygen species
production and activation of caspase-3-like activity and eventually cell death They also
reported that the ribonuclease activity component of ECP is not involved in cytotoxic
activity as no RNA degradation is observed [39] Sanchez et al applied enzymatic and
- 10 -
chemical limited cleavage to search for active sequence determinants for antimicrobial
activity and confirm the main role of the protein N-terminal The results corroborated that
the 11-35 internal sequence and in particular the 28-35 stretch is essential The identified
region is involved in both ECP aggregation and bacteria envelope binding properties[40]
Torrent et al studied ECP aggregation they demonstrated that ECP is able to form amyloid
aggregates The amyloid protofibrils formed by ECP bind amyloid-diagnostic dyes as Th-T
and Congo Red present a fibrilar structure under electron microscopy They identified an
N-terminus hydrophobic patch (residues 8-16) that is required for the amyloid aggregation
process [41] Pulido et al found ECP has a remarkable affinity for lipopolysaccharide (LPS)
and a distinctive agglutinating activity By using a battery of LPS-truncated E coli mutant
strains they demonstrated that the polysaccharide moiety of LPS is essential for
ECP-mediated bacterial agglutination thereby modulating its antimicrobial action The
mechanism of action of ECP at the bacterial surface is drastically affected by the LPS
structure and in particular by its polysaccharide moiety They also analyzed an N-terminal
fragment that retains the whole protein activity and displays similar cell agglutination
behavior A fragment with further minimization of the antimicrobial domain though
retaining the antimicrobial capacity significantly loses its agglutinating activity exhibiting
a different mechanism of action which is not dependent on the LPS composition [42]
In 2012 Boix et al crystallized ECP in complex with sulfate anions in a new momoclinic
crystal form The study provides direct evidence of the main protein sulfate binding sites
Three main sites (S1-S3) are located in the protein active site involved in sulfated
heterosaccharide binding S1 and S2 overlap with the phosphate binding sites involved in
RNase nucleotide recognition A new site (S3) is one of the key anchoring points for
sulfated ligands In particular site S3 is unique to ECP in the RNase A superfamily This
may explaine ECP reduced ribonuclease activity and high affinity for glycosaminoglycans
[43]
Some researchers studied the effect of ECP on mammalian tumor cells Maeda et al
evaluated the effect of ECP on 13 mammalian cell lines They found that ECP inhibited the
growth of several cell lines including those derived from carcinoma and leukemia in a
- 11 -
dose-dependent manner ECP significantly suppressed the size of colonies of A431 cells
and decreased K562 cells in G(1)G(0) phase However there was little evidence that ECP
killed cells in either cell line Those results suggest that growth inhibition by ECP is
dependent on cell type and is cytostatic [44] Similar studies were also done on gastric
cancer cells[45] oral malignant tumor cells[46] and Hodgkin lymphoma cell[47]
Very recently we have demonstrated that eosinophil cationic protein (ECP) affected the
development of cytoskeleton in normal fibroblast cells and the differentiation of rat
neonatal cardiomyocytes[48] This finding implies that ECP might be useful as a
cardiomyocyte differentiation factor in the repair of cardiac tissue by stimulating the
differentiation of cardiomyocyte progenitor cells[48]
3 Cardiomyocyte differentiation
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [49 50] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [51 52] In this
context alternative approaches may include restoring heart function via induction of
endogenous regenerative processes and transferring progenitor cells to produce new
myocardium Cardiomyocytes are currently thought to be postmitotic cells that withdraw
from the cell cycle soon after birth Nevertheless mitoses in the myocyte population have
been detected in the human heart after myocardial infarction [53] Recent studies suggest
that cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [54] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 9 -
neurotoxic activity and antimicrobial activity against bacteria pathogens and fungi
antiviral function and fibrosis promoting functions [6 32-35] Young et al reported that
purified human ECP could damage schistosomula of Schistosoma mansoni at low
concentrations Membrane damage was mediated by ECP through forming transmembrane
pores [33] Sorrentino et al treated EDN liver RNase and ECP with iodoacetic acid at
pH 55 which resulted in inactivation of their RNase activity and also destroyed their
neurotoxicity So they concluded that RNase activity is necessary but not sufficient to
induce neurotoxic action[36] However Rosenberg got the different conclusion Mutant
recombinant ECP that have no ribonuclease activity still shows antibacterial activity He
believed that ribonuclease activity and cytotoxicity are in that case independent functions
of ECP[37]About antiviral activity Domachowske et al prepared recombinant human ECP
(rhECP) prepared in baculovirus The rhECP was N-glycosylated and had similar to
100-fold more ribonuclease activity than nonglycosylated rhECP prepared in bacteria The
enzymatic activity of rhECP was sensitive to inhibition by placental ribonuclease inhibitor
(RI) And they found that rhECP promotes ribonuclease-dependent toxicity toward
extracellular virions of the single-stranded RNA virus respiratory syncytial virus group B
(RSV-B) [34]
On the basis of structure analysis ECP variants modified at basic and hydrophobic residues
have been constructed The bactericidal activity of both native ECP and point-mutated
variants were tested against Escherichia coli and Staphylococcus aureus Changes in the
leakage of liposome vesicles suggests that basic amino acids play in addition to the effect
on the disruption of the cellular membrane other roles such as specific binding on the
surface of the bacteria cell[38] Navarro et al analyzed ECP cytotoxic activity on
eukaryotic cell lines They found that ECP effects begin with its binding and aggregation to
the cell surface and then induce cell-specific morphological and biochemical changes such
as chromatin condensation reversion of membrane asymmetry reactive oxygen species
production and activation of caspase-3-like activity and eventually cell death They also
reported that the ribonuclease activity component of ECP is not involved in cytotoxic
activity as no RNA degradation is observed [39] Sanchez et al applied enzymatic and
- 10 -
chemical limited cleavage to search for active sequence determinants for antimicrobial
activity and confirm the main role of the protein N-terminal The results corroborated that
the 11-35 internal sequence and in particular the 28-35 stretch is essential The identified
region is involved in both ECP aggregation and bacteria envelope binding properties[40]
Torrent et al studied ECP aggregation they demonstrated that ECP is able to form amyloid
aggregates The amyloid protofibrils formed by ECP bind amyloid-diagnostic dyes as Th-T
and Congo Red present a fibrilar structure under electron microscopy They identified an
N-terminus hydrophobic patch (residues 8-16) that is required for the amyloid aggregation
process [41] Pulido et al found ECP has a remarkable affinity for lipopolysaccharide (LPS)
and a distinctive agglutinating activity By using a battery of LPS-truncated E coli mutant
strains they demonstrated that the polysaccharide moiety of LPS is essential for
ECP-mediated bacterial agglutination thereby modulating its antimicrobial action The
mechanism of action of ECP at the bacterial surface is drastically affected by the LPS
structure and in particular by its polysaccharide moiety They also analyzed an N-terminal
fragment that retains the whole protein activity and displays similar cell agglutination
behavior A fragment with further minimization of the antimicrobial domain though
retaining the antimicrobial capacity significantly loses its agglutinating activity exhibiting
a different mechanism of action which is not dependent on the LPS composition [42]
In 2012 Boix et al crystallized ECP in complex with sulfate anions in a new momoclinic
crystal form The study provides direct evidence of the main protein sulfate binding sites
Three main sites (S1-S3) are located in the protein active site involved in sulfated
heterosaccharide binding S1 and S2 overlap with the phosphate binding sites involved in
RNase nucleotide recognition A new site (S3) is one of the key anchoring points for
sulfated ligands In particular site S3 is unique to ECP in the RNase A superfamily This
may explaine ECP reduced ribonuclease activity and high affinity for glycosaminoglycans
[43]
Some researchers studied the effect of ECP on mammalian tumor cells Maeda et al
evaluated the effect of ECP on 13 mammalian cell lines They found that ECP inhibited the
growth of several cell lines including those derived from carcinoma and leukemia in a
- 11 -
dose-dependent manner ECP significantly suppressed the size of colonies of A431 cells
and decreased K562 cells in G(1)G(0) phase However there was little evidence that ECP
killed cells in either cell line Those results suggest that growth inhibition by ECP is
dependent on cell type and is cytostatic [44] Similar studies were also done on gastric
cancer cells[45] oral malignant tumor cells[46] and Hodgkin lymphoma cell[47]
Very recently we have demonstrated that eosinophil cationic protein (ECP) affected the
development of cytoskeleton in normal fibroblast cells and the differentiation of rat
neonatal cardiomyocytes[48] This finding implies that ECP might be useful as a
cardiomyocyte differentiation factor in the repair of cardiac tissue by stimulating the
differentiation of cardiomyocyte progenitor cells[48]
3 Cardiomyocyte differentiation
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [49 50] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [51 52] In this
context alternative approaches may include restoring heart function via induction of
endogenous regenerative processes and transferring progenitor cells to produce new
myocardium Cardiomyocytes are currently thought to be postmitotic cells that withdraw
from the cell cycle soon after birth Nevertheless mitoses in the myocyte population have
been detected in the human heart after myocardial infarction [53] Recent studies suggest
that cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [54] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 10 -
chemical limited cleavage to search for active sequence determinants for antimicrobial
activity and confirm the main role of the protein N-terminal The results corroborated that
the 11-35 internal sequence and in particular the 28-35 stretch is essential The identified
region is involved in both ECP aggregation and bacteria envelope binding properties[40]
Torrent et al studied ECP aggregation they demonstrated that ECP is able to form amyloid
aggregates The amyloid protofibrils formed by ECP bind amyloid-diagnostic dyes as Th-T
and Congo Red present a fibrilar structure under electron microscopy They identified an
N-terminus hydrophobic patch (residues 8-16) that is required for the amyloid aggregation
process [41] Pulido et al found ECP has a remarkable affinity for lipopolysaccharide (LPS)
and a distinctive agglutinating activity By using a battery of LPS-truncated E coli mutant
strains they demonstrated that the polysaccharide moiety of LPS is essential for
ECP-mediated bacterial agglutination thereby modulating its antimicrobial action The
mechanism of action of ECP at the bacterial surface is drastically affected by the LPS
structure and in particular by its polysaccharide moiety They also analyzed an N-terminal
fragment that retains the whole protein activity and displays similar cell agglutination
behavior A fragment with further minimization of the antimicrobial domain though
retaining the antimicrobial capacity significantly loses its agglutinating activity exhibiting
a different mechanism of action which is not dependent on the LPS composition [42]
In 2012 Boix et al crystallized ECP in complex with sulfate anions in a new momoclinic
crystal form The study provides direct evidence of the main protein sulfate binding sites
Three main sites (S1-S3) are located in the protein active site involved in sulfated
heterosaccharide binding S1 and S2 overlap with the phosphate binding sites involved in
RNase nucleotide recognition A new site (S3) is one of the key anchoring points for
sulfated ligands In particular site S3 is unique to ECP in the RNase A superfamily This
may explaine ECP reduced ribonuclease activity and high affinity for glycosaminoglycans
[43]
Some researchers studied the effect of ECP on mammalian tumor cells Maeda et al
evaluated the effect of ECP on 13 mammalian cell lines They found that ECP inhibited the
growth of several cell lines including those derived from carcinoma and leukemia in a
- 11 -
dose-dependent manner ECP significantly suppressed the size of colonies of A431 cells
and decreased K562 cells in G(1)G(0) phase However there was little evidence that ECP
killed cells in either cell line Those results suggest that growth inhibition by ECP is
dependent on cell type and is cytostatic [44] Similar studies were also done on gastric
cancer cells[45] oral malignant tumor cells[46] and Hodgkin lymphoma cell[47]
Very recently we have demonstrated that eosinophil cationic protein (ECP) affected the
development of cytoskeleton in normal fibroblast cells and the differentiation of rat
neonatal cardiomyocytes[48] This finding implies that ECP might be useful as a
cardiomyocyte differentiation factor in the repair of cardiac tissue by stimulating the
differentiation of cardiomyocyte progenitor cells[48]
3 Cardiomyocyte differentiation
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [49 50] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [51 52] In this
context alternative approaches may include restoring heart function via induction of
endogenous regenerative processes and transferring progenitor cells to produce new
myocardium Cardiomyocytes are currently thought to be postmitotic cells that withdraw
from the cell cycle soon after birth Nevertheless mitoses in the myocyte population have
been detected in the human heart after myocardial infarction [53] Recent studies suggest
that cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [54] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 11 -
dose-dependent manner ECP significantly suppressed the size of colonies of A431 cells
and decreased K562 cells in G(1)G(0) phase However there was little evidence that ECP
killed cells in either cell line Those results suggest that growth inhibition by ECP is
dependent on cell type and is cytostatic [44] Similar studies were also done on gastric
cancer cells[45] oral malignant tumor cells[46] and Hodgkin lymphoma cell[47]
Very recently we have demonstrated that eosinophil cationic protein (ECP) affected the
development of cytoskeleton in normal fibroblast cells and the differentiation of rat
neonatal cardiomyocytes[48] This finding implies that ECP might be useful as a
cardiomyocyte differentiation factor in the repair of cardiac tissue by stimulating the
differentiation of cardiomyocyte progenitor cells[48]
3 Cardiomyocyte differentiation
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [49 50] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [51 52] In this
context alternative approaches may include restoring heart function via induction of
endogenous regenerative processes and transferring progenitor cells to produce new
myocardium Cardiomyocytes are currently thought to be postmitotic cells that withdraw
from the cell cycle soon after birth Nevertheless mitoses in the myocyte population have
been detected in the human heart after myocardial infarction [53] Recent studies suggest
that cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [54] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 12 -
cardiac injury [55] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in a large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [56] One of
the major obstacles hindering the clinical use of ES cells derived cardiomyocytes for
cardiac regenerative therapy is inefficient cardiac differentiation leading to insufficient
amount of cardiomyocytes [57] and the potential development of teratomas The most
common approach to solve these problems are to apply knowledge of developmental
biology in employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [58-60]
According to previous study ECP enhanced neonatal rat cardiomyocyte differentiation
However the role of ECP on the cardiomyocyte differentiation and detail mechanism were
still unclear These unclear problems provided me with motivation to initiate this study
This thesis consists of 3 chapters in addition to Introduction and Conclusions
In chapter 1 the functional role of ECP in the cardiogenesis was investigated by mouse
P19CL6 embryonic carcinoma cells ECP was confirmed to accelerate the cardiomyocyte
differentiation of P19CL6 cells by the rate and area size of beating and the expression of
cardiomyocyte specific genes
In chapter 2 I detected the molecular mechanism of ECP on the cardiomyocyte
differentiation of P19CL6 cells focus on FGFR signaling I concluded that ECP induced
mesoderm differentiation by stimulating FGF signaling pathway and then enhanced
subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6 cells
In chapter 3 I checked effects of ECP on the other signaling pathways correlated with
cardiomyocyte differentiation such as Wntβ-catenin PI3KAkt EphA1 ET-1 and so on
- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 13 -
4 References
1 Gleich GJ et al Biochemical and Functional Similarities between Human
Eosinophil-Derived Neurotoxin and Eosinophil Cationic Protein - Homology with
Ribonuclease Proceedings of the National Academy of Sciences of the United
States of America 1986 83(10) p 3146-3150
2 Rosenberg HF SJ Ackerman and DG Tenen Human Eosinophil Cationic
Protein - Molecular-Cloning of a Cyto-Toxin and Helminthotoxin with
Ribonuclease-Activity Journal of Experimental Medicine 1989 170(1) p
163-176
3 Beintema JJ The ribonuclease A superfamily - Introduction Cellular and
Molecular Life Sciences 1998 54(8) p 763-765
4 Beintema JJ and RG Kleineidam The ribonuclease A superfamily general
discussion Cellular and Molecular Life Sciences 1998 54(8) p 825-832
5 Mallorqui-Fernandez G et al Three-dimensional crystal structure of human
eosinophil cationic protein (RNase 3) at 175 angstrom resolution Journal of
Molecular Biology 2000 300(5) p 1297-1307
6 Boix E et al The antipathogen activities of eosinophil cationic protein Current
Pharmaceutical Biotechnology 2008 9(3) p 141-152
7 Venge P et al Eosinophil cationic protein (ECP) molecular and biological
properties and the use of ECP as a marker of eosinophil activation in disease
Clinical and Experimental Allergy 1999 29(9) p 1172-1186
8 Futami J et al Tissue-specific expression of pancreatic-type RNases and RNase
inhibitor in humans DNA and Cell Biology 1997 16(4) p 413-419
9 Sorva R et al Eosinophil cationic protein in induced sputum as a marker of
inflammation in asthmatic children Pediatric Allergy and Immunology 1997 8(1)
p 45-50
- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 14 -
10 Niimi A et al Serum eosinophil cationic protein as a marker of eosinophilic
inflammation in asthma Clinical and Experimental Allergy 1998 28(2) p
233-240
11 Kang H et al Serum Eosinophil Cationic Protein a Useful Follow-up Marker in
Bakers Asthma Journal of Allergy and Clinical Immunology 2009 123(2) p
S235-S235
12 Guilpain P et al Serum eosinophil cationic protein - A marker of disease activity
in Churg-Strauss syndrome Autoimmunity Pt C 2007 1107 p 392-399
13 Kurklu E and K Guven Eosinophil cationic protein in saliva a marker for disease
activity in oral lesions Oral Diseases 2010 16(6) p 540-540
14 Schmekel B et al Eosinophil cationic protein (ECP) in saliva a new marker of
disease activity in bronchial asthma Respiratory Medicine 2001 95(8) p 670-675
15 Robinson DS et al Eosinophil Cationic Protein (Ecp) and Eosinophil-Derived
Neurotoxin (Edn) Levels Are Elevated in Blood and Bronchoalveolar Lavage from
Asthmatics Journal of Allergy and Clinical Immunology 1994 93(1) p 228-228
16 Rasp G et al IL-5 IgE ECP and ICAM-1 in nasal secretions of patients suffering
from chronic non-allergic sinusitis allergic rhinitis and non-allergic nasal
polyposis Journal of Allergy and Clinical Immunology 1999 103(1) p
S247-S247
17 Terada A et al Eosinophil Cationic Protein (Ecp) Is a Useful Marker for
Differential-Diagnosis of Wheezing in Children Journal of Allergy and Clinical
Immunology 1995 95(1) p 277-277
18 McGill KA et al Use of eosinophil cationic protein as a predictive marker for
exacerbations of asthma Journal of Allergy and Clinical Immunology 1996 97(1)
p 511-511
19 Vatrella A et al Serum eosinophil cationic protein (ECP) as a marker of disease
activity and treatment efficacy in seasonal asthma Allergy 1996 51(8) p 547-555
20 Terada A et al Serum eosinophil cationic protein as a guiding marker for
initiation of inhaled corticosteroid in children with asthma Journal of Allergy and
- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 15 -
Clinical Immunology 1997 99(1) p 1636-1636
21 Iikura Y et al Evaluation of serum eosinophil cationic protein (ECP) monitoring
childhood asthma ECP as a marker for indication and discontinuation of inhaled
steroid therapy Journal of Allergy and Clinical Immunology 1998 101(1) p
S8-S8
22 Bystrom J et al Dissecting the role of eosinophil cationic protein in upper airway
disease Current Opinion in Allergy and Clinical Immunology 2012 12(1) p
18-23
23 Dainese R et al Role of serological markers of activated eosinophils in
inflammatory bowel diseases European Journal of Gastroenterology amp Hepatology
2012 24(4) p 393-397
24 Wagner M et al Fecal eosinophil cationic protein as a marker of active disease
and treatment outcome in collagenous colitis A pilot study Scandinavian Journal of
Gastroenterology 2011 46(7-8) p 849-854
25 Wakamatsu TH et al IgE and eosinophil cationic protein (ECP) as markers of
severity in the diagnosis of atopic keratoconjunctivitis British Journal of
Ophthalmology 2012 96(4) p 581-586
26 Wakamatsu TH et al Eosinophil cationic protein as a marker for assessing the
efficacy of tacrolimus ophthalmic solution in the treatment of atopic
keratoconjunctivitis Molecular Vision 2011 17(103) p 932-938
27 Magnaval JF et al Eosinophil cationic protein as a possible marker of active
human Toxocara infection Allergy 2001 56(11) p 1096-1099
28 Gullberg U et al The Cytotoxic Eosinophil Cationic Protein (Ecp) Has
Ribonuclease-Activity Biochemical and Biophysical Research Communications
1986 139(3) p 1239-1242
29 Sorrentino S and DG Glitz Ribonuclease-Activity and Substrate Preference of
Human Eosinophil Cationic Protein (Ecp) Febs Letters 1991 288(1-2) p 23-26
30 Boix E et al Kinetic and product distribution analysis of human eosinophil
cationic protein indicates a subsite arrangement that favors exonuclease-type
- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012

- 16 -
activity Journal of Biological Chemistry 1999 274(22) p 15605-15614
31 Mohan CG et al The crystal structure of eosinophil cationic protein in complex
with 25-ADP at 20 A resolution reveals the details of the ribonucleolytic active
site Biochemistry 2002 41(40) p 12100-6
32 Meyer JS and JM Foley The Encephalopathy Produced by Extracts of
Eosinophils and Bone Marrow Journal of Neuropathology and Experimental
Neurology 1953 12(4) p 349-362
33 Young JD et al Mechanism of Membrane Damage Mediated by Human
Eosinophil Cationic Protein Nature 1986 321(6070) p 613-616
34 Domachowske JB et al Eosinophil cationic protein RNase 3 is another
RNaseA-family ribonuclease with direct antiviral activity Nucleic Acids Research
1998 26(14) p 3358-3363
35 Torrent M et al Topography studies on the membrane interaction mechanism of
the eosinophil cationic protein Biochemistry 2007 46(3) p 720-733
36 Sorrentino S et al Eosinophil-Derived Neurotoxin and Human Liver
Ribonuclease - Identity of Structure and Linkage of Neurotoxicity to Nuclease
Activity Journal of Biological Chemistry 1992 267(21) p 14859-14865
37 Rosenberg HF Recombinant Human Eosinophil Cationic Protein -
Ribonuclease-Activity Is Not Essential for Cytotoxicity Journal of Biological
Chemistry 1995 270(14) p 7876-7881
38 Carreras E et al Both aromatic and cationic residues contribute to the
membrane-lytic and bactericidal activity of eosinophil cationic protein
Biochemistry 2003 42(22) p 6636-6644
39 Navarro S et al The cytotoxicity of eosinophil cationic proteinribonuclease 3 on
eukaryotic cell lines takes place through its aggregation on the cell membrane
Cellular and Molecular Life Sciences 2008 65(2) p 324-337
40 Sanchez D et al Mapping the eosinophil cationic protein antimicrobial activity by
chemical and enzymatic cleavage Biochimie 2011 93(2) p 331-338
41 Torrent M et al Eosinophil Cationic Protein Aggregation Identification of an
- 17 -
N-Terminus Amyloid Prone Region Biomacromolecules 2010 11(8) p
1983-1990
42 Pulido D et al Antimicrobial Action and Cell Agglutination by the Eosinophil
Cationic Protein Are Modulated by the Cell Wall Lipopolysaccharide Structure
Antimicrobial Agents and Chemotherapy 2012 56(5) p 2378-2385
43 Boix E et al The sulfate-binding site structure of the human eosinophil cationic
protein as revealed by a new crystal form Journal of Structural Biology 2012
179(1) p 1-9
44 Maeda T et al Growth inhibition of mammalian cells by eosinophil cationic
protein European Journal of Biochemistry 2002 269(1) p 307-316
45 Koo JW et al Association of of RNase3 Polymorphisms with the Susceptibility
of Gastric Cancer Journal of the Korean Surgical Society 2010 78(5) p 283-289
46 Pereira MC DT Oliveira and LP Kowalski The role of eosinophils and
eosinophil cationic protein in oral cancer A review Archives of Oral Biology 2011
56(4) p 353-358
47 Glimelius I et al Effect of eosinophil cationic protein (ECP) on Hodgkin
lymphoma cell lines Experimental Hematology 2011 39(8) p 850-858
48 Fukuda T et al Human eosinophil cationic protein enhances stress fiber formation
in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes Growth
Factors 2009 27(4) p 228-236
49 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009)
24ndash32
50 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
51 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
52 JG Augoustides H Riha Recent progress in heart failure treatment and heart
- 18 -
transplantation J Cardiothorac Vasc Anesth (2009)
53 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that
human cardiac myocytes divide after myocardial infarction N Engl J Med 344
(2001) 1750ndash1757
54 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
55 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007)
970ndash974
56 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
57 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
58 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function
of infarcted rat hearts Nat Biotechnol 251015-1024
59 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
60 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition
of BMP signaling by Noggin induces cardiomyocyte differentiation of mouse
embryonic stem cells Nature Biotechnology 23897-897
- 19 -
Chapter 1
Effect of Eosinophil cationic protein (ECP) on the
cardiomyocyte differentiation in P19CL6 (mouse
teratocarcinoma) cells
- 20 -
Abstract
We previously demonstrated that eosinophil cationic protein (ECP) promoted the
differentiation of the rat primary cardiac cells Here we investigated the functional role of
ECP in the cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells which are
well known as a model of cardiomyocyte differentiation in the presence of DMSO ECP
was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by the rate
and area size of beating and the expression of cardiomyocyte specific genes such as
GATA-4 and α-MHC P19CL6 cells at 12 days of treatment were immunologically stained
with antibodies against actinin actin ANF and cardiac troponin I The wider size of
immunoreactive area was observed in the presence of ECP than that without ECP This is
the first to report the function of ECP on cardiomyocyte differentiation from stem cells in
vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be devoted in the development of novel therapies for cardiac regeneration
- 21 -
Introduction
Unlike other organs such as the skin and liver the heart is not able to regenerate sufficient
cardiomyocytes to undergo extensive repair Shortly after birth most cardiomyocytes stop
dividing and are terminally differentiated Ischemic heart disease and congestive heart
failure are major reasons of morbidity and mortality in the world and place a substantial
economic burden on most health systems [1] Heart failure occurs as the end result of
pathological remodeling of the myocardium in response to either ischemic or nonischemic
injury and a core component of this process is cardiomyocyte death and loss of myocardial
cell mass [2] Cardiac transplantation is a major treatment of choice for end-stage heart
failure at present However the number of available donor organs limits this treatment
option to a minority of patients [3] The development of new therapeutic paradigms for
heart failure and alternative therapies such as cardiac cell replacement has therefore
become imperative Stem cell therapy is a relatively new frontier in the battle against
cardiovascular disease that has sparked intense research and criticism In theory ES cells
could be produced in a large numbers differentiated into cardiomyocytes which could be
used as a renewable source for cardiac cell therapy [23] Over the past few years several
promising results have been reported but many hurdles remain before stem cells can
actually be used to treat patients with a damaged heartsuch as low efficiency of
differentiation find a inducer which can enhance cardiomyocyte differentiation is hot point
on this research
P19 embryonal carcinoma cells
P19 mouse embryonal carcinoma cell line has been reported to differentiate into an
embryonic cardiac muscle phenotype in vitro [23 24] upon the addition of dimethyl
sulfoxide (DMSO) Differentiated P19 cells have been reported to retain the ability to
spontaneously contract and shown to express transcripts in a temporal manner during
culture suggestive of a cardiac muscle phenotype [28-30] and as such these cells have
therefore been extensively used to study cardiac cell physiology [2425282931] although
- 22 -
with the caveat that these cells are embryonic instead However in addition to these cardiac
muscle specific properties P19 cells also display pluripotent properties and can be
differentiated into cells displaying either a skeletal muscle or neural phenotype [24 26-28]
There has thus been some concern about the homogeneity of DMSO differentiated P19
cultures with a heterogeneous cell population following differentiation significantly
reducing the utility of these cells as a cardiac-muscle-specific model There has thus been
much interest in identifying subclones of P19 cells that more robustly differentiate into
cardiomyocytes The P19CL6 cells line a sub-clone of P19 embryonal cells has been
reported to efficiently differentiate into beating cardiomyocytes upon exposure to DMSO
under adherent culture conditions [32] and has been widely used as an in vitro model of
cardiovascular cells [242533-39] It is clear that the P19CL6 cell line regards as a model
system for the study of cardiomyocyte development and differentiation In this chapter we
used this cell line to examine the function of ECP on the cardiomyocyte differentiation
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[13 14] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
- 23 -
Recombinant human ECP was expressed in bacteria and prepared as described previously
[15 16] SU5402 which is an FGF receptor (FGFR) inhibitor was purchased from
Calbiochem (San Diego CA) Anti-actin rabbit polyclonal antibody anti-actinin mouse
monoclonal antibody and anti-cardiac troponin I mouse monoclonal antibody were from
Sigma-Aldrich
Reverse transcription polymerase chain reaction (RT-PCR) and real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For semiquantitative analysis reverse
transcribed products were pooled and fourfold serial dilutions were used for RT-qPCR
(Lyte-Cycler 480 Roche Diagnostics Germany) PCR was performed in a reaction volume
of 20 μL with 200 nM deoxynucleoside triphosphates 500 nM each of sense and antisense
primers and 5 units100 μL Taq polymerase (Takara Japan) The amplification reaction
was carried out in an authorized thermal cycler (Eppendorf Germany) The sequences of
primers used for the RT-PCR are following (forward and reverse) GATA4
5-ACTCTGGAGGCGAGATGGG-3 and 5-CTCGGCATTACGACGCCACAG-3
α-MHC 5-GGAAGAGTGAGCGGCGCATCAAGGA-3 and
5-TCTGCTGGAGAGGTTATTCCTCG-3 β-MHC
5-CGGAGGAGCAGGCCAACACCAACT-3 and
5-GCAAAGGCTCCAGGTCTGAGGGCTT-3 Nkx25
5-TGGCAGAGCTGCGCGCGGAGATG-3 and
5-CGTGGCTTCCGTCGCCGCCGTGC-3 glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) 5-CCCTTCATTGACCTCAACTAC-3 and
5-CCACCTTCTTGATGTCATCAT-3 PCR was performed for 1 cycle at 94degC for 5 min
followed by 25ndash35 cycles of denaturation at 94degC for 30 sec annealing at 56ndash62degC
- 24 -
depending on the melting temperature of each pair of primers for 30 sec followed by
extension at 72degC for 1 min
For quantitative analysis of gene expression levels real time qPCR were performed and the
data were normalized to GAPDH Primers used for the real time qPCR were as following
(forward and reverse) 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Immunostaining of P19CL6 cells
P19CL6 cells were seeded into 12-well tissue culture plate (TPP Switzerland) with a
sterilized cover slip at 105 cells per well in growth medium Twenty-four hours later the
medium was changed to differentiation medium with or without 1microgmL (66 nM) of ECP
On the 12th day the cells were washed with PBS twice fixed with 4 (wv)
paraformaldehyde at room temperature for 30 min permeabilized in phosphate-buffered
saline buffer containing 005 Tween-20 (PBST) and then blocked with PBST containing
5 BSA (blocking buffer) for one hour The cells were incubated with anti-actin and
anti-actinin antibodies diluted in blocking buffer at a dilution of 1200 at 4 for 16 h The
cells were washed twice with PBST and then incubated with both Alexa 488-conjugated
anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG secondary
antibody (Amersham-Pharmacia) in blocking buffer at 25 ˚C for 1 h For cardiac troponin I
staining mouse anti-troponin I antibody was used at a dilution of 1200 Fluorescent
- 25 -
images of immunostaining were observed under a confocal laser microscope LSM510-V30
(Carl Zeiss UK) at 488 nm and 543 nm
Statistical analysis
All experiments were replicated at least three times Statistical data analysis was performed
using the Studentrsquos t-test by Excel software Values are reported as means plusmn SD The data
were considered statistically significant when p lt 005
Results
Enhanced cardiomyocyte differentiation of P19CL6 cells after ECP treatment
P19CL6 cells differentiate into cardiomyocyte in the presence of 1 DMSO
DMSO-treated P19CL6 cells are considered as a model of cardiomyocyte differentiation of
which the molecular mechanism has been extensively analyzed [17-19] Since we found
that ECP promoted the differentiation of cardiomyocytes in the rat neonate we tried to
assess the activity of ECP using P19CL6 cells First P19CL6 cells were cultured in the
presence or absence of ECP and DMSO The medium was changed every two days and
then morphology of cells was observed under microscope On the 8th day the cells
spontaneously started beating in the presence of both ECP and DMSO while it took 12 days
for the cells to start beating with only DMSO (Fig 1) ECP was found to accelerate
cardiomyocyte differentiation by almost 4 days On day 12 the beating rate of the cells was
50plusmn8 timesmin in the presence of ECP and 30plusmn10 timesmin without ECP (Fig 2A)
Simultaneously the area of beating in the presence of ECP was nearly 8-fold wider than
that without ECP The average area of beating was 48 plusmn 1047 per field (n = 24 different
fields field size = 08 mm2) with ECP versus 617 plusmn 123 (n = 24 different fields) without
ECP (Fig 2B and C) Thus ECP was judged to enhance cardiomyocyte differentiation in
both beating rate and area However ECP did not induce cardiomyocyte differentiation
without DMSO treatment
- 26 -
Enhanced expression of cardiomyocyte genes in P19CL6 cells after ECP treatment
The time course change from 4 to 12 days of cardiac marker gene expression in P19CL6
cells was evaluated by RT-PCR in the presence of ECP and DMSO (Fig 3A) GATA-4
expression was dramatically accelerated within 4 days of treatment with ECP and DMSO
when compared with that treated with only DMSO α-and β-MHC expression were also
accelerated within 8 days while α-MHC expression was upregulated within 12 days and
β-MHC expression still required more than 12 days for upregulation without ECP In
contrast to these genes Nkx25 expression was rather low However Nkx25 was
significantly upregulated within 12 days in the presence of ECP whereas it could not be
defected without ECP Further analyses on the expression of MLC GATA4 and α-MHC
were performed by real time qPCR and the effect of acceleration of cardiomyocyte
differentiation by ECP was confirmed (Fig 3B) P19CL6 cells at 12 days of treatment were
immunologically stained with antibodies against actinin actin and cardiac troponin I A
wider area of immunoreactivity which was indicative of enhanced expression in a greater
number of cells was observed in the presence of ECP than that without ECP (Fig 3C) Thus
the enhancement of cardiomyocyte differentiation by ECP was confirmed not only by
marker gene expression but also by the proteins expression
-
+ 0
Fig 1 Morphological change of P19CL6 cells during cardiomyocyte differentiation
P19CL6 cells were treated with or without ECP in the presence of DMSO The cells were
ECP
Days 0 2 4 8 10 12
- 27 -
observed under microscope at indicated days The red frame shown the time that cells
started beating Magnification is times 20
Fig 2 Beating rate (A) and area (B) of cardiomyocytes differentiated from p19CL6 cells
treated for 12 days with or without ECP in the presence of DMSO (A) Beating rate was
calculated by beating frequency per minute of P19CL6 cells (A) (B) data represent mean plusmn
SD from triplicate of experiments Each asterisk shows the significance of Plt005 (C)
Typical area of beating cells (in white circle) counted in (B) The magnification is times 20
- 28 -
Fig 3 Induction of cardiac-specific genes (GATA4 Nkx25 MHC) assessed by RT-PCR
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to RT-PCR
A B C
Fig 4 Expression of cardiac marker genes (GATA4 α-MHC MLC) assessed by real time
qPCR (A)(B)(C) P19CL6 cells were treated with or without ECP in the presence of DMSO
Total RNA was isolated at indicated time points and subjected to real time qPCR Each
result is represented as mean plusmn SD from triplicate of experiments
Fig 5 Immunostaining of cardiac specific proteins (Actinin and actin) P19CL6 cells were
treated with or without ECP in the presence of DMSO On 12 days the cells were
incubated with anti-actin and anti-actinin antibodies and then incubated with both Alexa
- 29 -
488-conjugated anti-mouse IgG antibody and Alexa Fluorreg 555-conjugated anti-rabbit IgG
secondary antibody Magnification is times 63 Scale Bars = 50 μm (C) Actinin (green) and
actin (red)
A B
Fig 6 Immunostaining of cardiac specific proteins (Cardiac troponin I and ANF) (A)
P19CL6 cells were treated with or without ECP in the presence of DMSO On 12 days the
cells were incubated with anti- cardiac troponin I actin antibodies and then incubated with
both Alexa 488-conjugated anti-mouse IgG antibody (B) The cells incubated as in (A) on
12 days the cells were incubated with anti- ANF and then incubated with Alexa
555-conjugated anti-mouse IgG antibody Magnification is times 63 Scale Bars = 50 μm
Discussion
Heart failure is currently the most common cardiac disease largely due to the increasing
average age of the population [1 2] Heart failure is a progressive disorder initiated by an
acute or gradual loss of functional cardiomyocytes resulting in diminished cardiac output
and cardiac performance for instance after myocardial infarction or in patients suffering
from hypertension Current therapies include lifestyle modification drug treatments
surgery and ultimately heart transplantation
ANF
ANF
- 30 -
Cardiac transplantation remains the only therapeutic option for end-stage heart failure but
the low number of organ donors and the side effects of immunosuppressive drugs limit the
access to a transplantation program to a few thousand patients a year [3 4] In this context
alternative approaches may include restoring heart function via induction of endogenous
regenerative processes and transferring progenitor cells to produce new myocardium
Cardiomyocytes are currently thought to be postmitotic cells that withdraw from the cell
cycle soon after birth Nevertheless mitoses in the myocyte population have been detected
in the human heart after myocardial infarction [5] Recent studies suggest that
cardiomyocytes may be continually replaced in the heart through processes involving
differentiation maturation senescence and death [6] Cardiomyocyte turnover in the
mammalian heart has been demonstrated experimentally and shown to be triggered by
cardiac injury [7] Regenerative stem cell therapy is a relatively new frontier in the battle
against cardiovascular disease that has sparked intense research and criticism In theory
embryonic stem (ES) cells could be produced in large numbers differentiated to
cardiomyocytes and then used as a renewable source for cardiac cell therapy [8] One of the
major obstacles hindering the clinical use of ES cells-derived cardiomyocytes for cardiac
regenerative therapy is inefficient cardiac differentiation leading to insufficient amount of
cardiomyocytes [9] and the potential development of teratomas The most common
approach to solve this problem is to apply knowledge of developmental biology in
employing cardiogenesis-related factors for inducing stem cells cardiomyocyte
differentiation [10-12]
In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 5C) This is
the first study to report a function for ECP in cardiomyocyte differentiation from stem cells
in vitro We are now trying to develop ECP as cardiomyocyte differentiation factor which
could be used in the development of novel therapies for cardiac regeneration
- 31 -
Reference
1 P Hodges Heart failure epidemiologic update Crit Care Nurs Q 32 (2009) 24ndash32
2 Y Sun Myocardial repairremodelling following infarction roles of local factors
Cardiovasc Res 81 (2009) 482ndash490
3 DO Taylor LB Edwards P Aurora JD Christie F Dobbels R Kirk AO
Rahmel AY Kucheryavaya MI Hertz Registry of the International Society for
Heart and Lung Transplantation twenty-fifth official adult heart transplant
reportmdash2008 J Heart Lung Transplant 27 (2008) 943ndash956
4 JG Augoustides H Riha Recent progress in heart failure treatment and heart
transplantation J Cardiothorac Vasc Anesth (2009)
5 AP Beltrami K Urbanek J Kajstura SM Yan N Finato R Bussani B
Nadal-Ginard F Silvestri A Leri CA Beltrami P Anversa Evidence that human
cardiac myocytes divide after myocardial infarction N Engl J Med 344 (2001)
1750ndash1757
6 P Anversa B Nadal-Ginard Myocyte renewal and ventricular remodelling Nature
415 (2002) 240ndash243
7 PC Hsieh VF Segers ME Davis C MacGillivray J Gannon JD Molkentin J
Robbins RT Lee Evidence from a genetic fate-mapping study that stem cells
refresh adult mammalian cardiomyocytes after injury Nat Med 13 (2007) 970ndash974
8 Habara-Ohkubo A 1996 Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Struct Funct 21101-110
9 Segers VF Lee RT 2008 Stem-cell therapy for cardiac disease Nature 451937-942
10 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
derived from human embryonic stem cells in pro-survival factors enhance function of
- 32 -
infarcted rat hearts Nat Biotechnol 251015-1024
11 Singh AM Li FQ Hamazaki T Kasahara H Takemaru KI Terada N 2007 Chibby
an antagonist of the Wntbeta-catenin pathway facilitates cardiomyocyte
differentiation of murine embryonic stem cells Circulation 115617-626
12 Yuasa S Itabashi Y Koshimizu U Tanaka T Sugimura K Kinoshita M Hattori F
Fukami S Shimazaki T Okano H Ogawa S Fukuda K 2005 Transient inhibition of
BMP signaling by Noggin induces cardiomyocyte differentiation of mouse embryonic
stem cells Nature Biotechnology 23897-897
13 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
14 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
15 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
16 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
17 Clement CA Kristensen SG Mollgard K Pazour GJ Yoder BK Larsen LA
Christensen ST 2009 The primary cilium coordinates early cardiogenesis and
hedgehog signaling in cardiomyocyte differentiation J Cell Sci 1223070-3082
18 Laflamme MA Chen KY Naumova AV Muskheli V Fugate JA Dupras SK
Reinecke H Xu C Hassanipour M Police S OSullivan C Collins L Chen Y
Minami E Gill EA Ueno S Yuan C Gold J Murry CE 2007 Cardiomyocytes
- 33 -
derived from human embryonic stem cells in pro-survival factors enhance function of
infarcted rat hearts Nat Biotechnol 251015-1024
19 Harada K Ogai A Takahashi T Kitakaze M Matsubara H Oh H 2008
Crossveinless-2 controls bone morphogenetic protein signaling during early
cardiomyocyte differentiation in P19 cells J Biol Chem 28326705-26713
20 Cohn JN Bristow MR Chien KR et al Report of the National Heart Lung And
Blood Institute Special Emphasis Panel On Heart Failure Research Circulation 1997
95766ndash770
21 Fedak PW Verma S Weisel RD et al Cardiac remodeling and failure From
molecules to man (Part I) Cardiovasc Pathol 2005 141ndash11
22 HABARA-OHKUBO A (1996) Differentiation of beating cardiac muscle cells from
a derivative of P19 embryonal carcinoma cells Cell StructFunct 21 101-110
23 Jackson KA Majka SM Wang H Pocius J Hartley CJ Majesky MW
Entman ML Michael LH Hirschi KK and Goodell MA (2001) Regeneration
of ischemic cardiac muscle and vascular endothelium by adult stem cells J Clin
Invest 107 1ndash8
24 Van der Heyden MA and Defize LH (2003) Twenty one years of P19 cells what
an embryonal carcinoma cell line taught us about cardiomyocyte differentiation
Car-diovascular Research 58 292-302
25 Anisimov SV Tarasov KV Riordon D Wobus AM and Boheler KR (2002)
SAGE identification of differ-entiation responsive genes in P19 embryonic cells
induced to form cardiomyocytes in vitro Mechanisms of Devel-opment 117 25-74
26 McBurney MW Jones-Villeneuve EM Edwards MK and Anderson PJ (1982)
Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell
line Nature 299 165-167
27 McBurney MW (1993) P19 embryonal carcinoma cells International Journal of
Developmental Biology 37 135-140
28 Skerjanc IS (1999) Cardiac and skeletal muscle development in P19 embryonal
carcinoma cells Trends Car-diovascular Medicine 9 139-143
- 34 -
29 Skerjanc IS Petropoulos H Ridgeway AG and Wil-ton S (1998) Myocyte
enhancer factor 2C and Nkx2-5 up-regulate each others expression and initiate
cardio-myogenesis in P19 cells Journal of Biological Chemistry 273 34904-34910
30 Wobus AM Kleppisch T Maltsev V and Hescheler J (1994) Cardiomyocyte like
cells differentiated in vitro from embryonic carcinoma cells P19 are characterized by
functional expression of adrenoceptors and Ca2+ channels In Vitro Cellular
Development and Biology 30A 425-434
31 Rudnicki MA Jackowski G Saggin L and McBurney MW (1990) Actin and
myosin expression during devel-opment of cardiac muscle from cultured embryonal
car-cinoma cells Developmental Biology 138 348-358
32 Habara-Ohkubo A (1996) Differentiation of beating cardiac muscle cells from a
derivative of P19 embryonal carcinoma cells Cell Structure and Function 21
101-110
33 Van der Heyden MA van Kempen MJ Tsuji Y Rook MB Jongsma HJ and
Opthof T (2003) P19 embryonal carcinoma cells a suitable model system for cardiac
electrophysiological differentiation at the molecular and functional level
Cardiovascular Research 58 410-422
34 Eaton S Chatziandreou I Krywawych S Pen S Clayton PT and Hussain K
(2003) Short-chain 3-hydroxyacyl-CoA dehydrogenase deficiency associated with
hyperinsulinism a novel glucose-fatty acid cycle Biochemical Society Transactions
31 1137-1139
35 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D
Hosoda T Kawabata M Miyazono K Ishii S Yazaki Y Nagai R and Komuro
I (2001) Smads TAK1 and their common target ATF-2 play a critical role in
cardiomyocyte differentiation Journal of Cell Biology 153 687-698
36 Paquin J Danalache BA Jankowski M McCann SM and Gutkowska J (2002)
Oxytocin induces differ-entiation of P19 embryonic stem cells to cardiomyocytes
Proceedings of the National Academy of Sciences USA 99 9550-9555
37 Peng CF Wei Y Levsky JM McDonald TV Childs G and Kitsis RN (2002)
- 35 -
Microarray analysis of global changes in gene expression during cardiac myocyte
dif-ferentiation Physioogical Genomics 9 145-155
38 Ridgeway AG Wilton S and Skerjanc IS (2000) Myocyte enhancer factor 2C
and myogenin up-regulate each others expression and induce the development of
skeletal muscle in P19 cells Journal of Biological Chemistry 275 41-46
39 Young DA Gavrilov S Pennington CJ Nuttall RK Edwards DR Kitsis RN
and Clark IM (2004) Expression of metalloproteinases and inhibitors in the
differentiation of P19CL6 cells into cardiac myocytes Biochemical and Biophysical
Research Communications 322 759-765
- 36 -
- 37 -
Chapter 2
Effect of eosinophil cationic protein (ECP) on activation
of FGF receptor 1 signaling during cardiomyocyte
differentiation
- 38 -
Abstract
I investigated the functional role of eosinophil cationic protein (ECP) in regulating
cardiomyogenesis using mouse P19CL6 embryonic carcinoma cells From previous chapter
ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells by
enhancing the rate and area size of beating of cardiomyocyte and by facilitating the
expression of cardiomyocyte specific genes such as GATA-4 and α-MHC In this chapter I
detected the molecular mechanism of ECP on the cardiomyocyte differentiation of P19CL6
cells Since cardiomyocyte differentiation in vivo is considered to follow mesoderm
induction the induction of Brachyury a marker of mesoderm was assessed Brachyury
expression was found to be enhanced after the addition of ECP This enhancement was due
to the stimulation of ERK12 phosphorylation by ECP In this context treatment with
SU5402 an inhibitor of FGFR1 suppressed Brachyury expression phosphorylation of
ERK12 and cardiomyocyte differentiation induced by ECP We concluded that ECP might
induce mesoderm differentiation through FGF signaling pathway and enhance subsequent
cardiomyocyte differentiation in concert with DMSO in P19CL6 cells ECP may be a novel
factor for cardiomyocyte differentiation which should be very useful to prepare adequate
numbers of cardiomyocytes for therapeutic cell transplantation
- 39 -
Introduction
The heart is formed through multiple developmental steps which include the determination
of the cardiac field in the mesoderm differentiation of cardiovascular progenitor cells
differentiation of cardiac precursor cells and maturation of the heart Entry of cells into the
cardiac lineage is dependent upon appropriate external signals coupled to the expression of
a set of transcription factors that initiates the program for cardiac genes expression and
drives the morphogenic events involved in formation of the multichambered heart It is well
known that this process is complicated and so many signaling pathways included Research
in Mice Birds Amphibians Flies and Mammals as well as in various cell culture systems
has led to the identification of multiple transcription factors and extracellular growth factors
whose concerted actions specify the cardiac lineage in mesodermal progenitor cells The
earliest expressed transcription factors that initiate cardiac fate are the homeobox
transcription factor NKX25 and members of the GATA family of zinc finger transcription
factors GATA4 GATA5 and GATA6 Equally important roles in heart development have
been shown for members of the Tbx5 Tbx20 basic helix-loop-helix eHANDHAND1
(Heart and Neural crest Derivatives expressed-1)) and MADS (MCMI Agamous
Deficiens Serum response factor) domain (MEF2) families Extracellular signals that act
upstream of these factors have been primarily identified by their ability to induce cardiac
differentiation in non-cardiac mesoderm These signals belong to the BMP (Bone
Morphogenetic Protein) FGF (Fibroblast Growth Factor) and Wnt (Wingless-related
MMTV integration site) families of Growth Factors and also include secreted Wnt
antagonists such as Dkk1 (Dickkopf1) and Crescent [29-33]
The fibroblast growth factor (FGF) family is essential to normal heart development FGF2
and BMP signaling pathway play a crucial role in early Cardiomyogenesis Kawai T et al
reported FGF2 is required for the expression of Cardiac transcription factors and the
differentiation of mesoderm explants induced by BMP2 (Bone Morphogenetic Protein-2)
[34] FGF2 induces mesenchymal cell formation from precardiac mesoderm explants Other
- 40 -
members of the FGF family compensate for the lack of FGF2 expression in the embryo
such as FGF 4 and FGF 8 FGF-10 induced cardiomyocyte differentiation from ES cells
and iPS cells [28] In this chapter I detected the effect of ECP on activation of FGF
signaling pathway
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[20 21] Briefly the cells were grown in
100-mm tissue culture dishes under adherent conditions with α-minimum essential medium
(α-MEM) (Invitrogen Tokyo Japan) containing 10 fetal bovine serum (FBS)
(Invitrogen) as growth medium in a 5 CO2 atmosphere at 37degC To induce
cardiomyocyte differentiation under adherent conditions P19CL6 cells were plated at a
density of 5times105 in 60-mm tissue culture dishes in growth medium Twenty four hours later
the medium was replaced with growth medium containing 1 DMSO (Nacalai tesque
Kyoto Japan) as differentiation medium The medium was changed to every 2 days and the
cells were maintained under fresh conditions until they started beating
Reagents and antibodies
Recombinant human ECP and recombinant human FGF-2 were expressed in bacteria and
prepared as described previously [22-24] SU5402 which is an FGF receptor (FGFR)
inhibitor was purchased from Calbiochem (San Diego CA) Anti-ERK and
anti-phospho-ERK12 mouse monoclonal antibodies were from New England Biolabs
(Beverly MA)All other reagents were of analytical grade and were purchased from Wako
or Sigma-Aldrich unless otherwise noted
Real time qPCR
P19CL6 cells cultured under various conditions were harvested and total RNA was
- 41 -
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA were then used to synthesize first-strand cDNA with
oligo-dT18 and 200 uunits SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse) Wnt3a
5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
Brachyury 5-AGCTCTCCAACCTATGCGGACAAT-3 and
5-TATCATGGGACTGCAGCATGGACA-3
GATA4 5-CAGCCCAGTCCTGCACAGCC-3 and
5-GGGCCGGTTGATGCCGTTCA-3 α-MHC
5-GCCATCACAGATGCCGCCATGA-3 and 5-TGCGCTTTTGCTCAGCCTCCA-3
Every PCR condition was confirmed to be within the linear range and within the
semiquantitative range for these specific genes and primer pairs To confirm that the
obtained bands were not derived from contaminated genomic DNA a negative assay was
performed on each sample without reverse transcriptase before PCR Amplified samples
were electrophoresed on 2 agarose gels and stained with ethidium bromide GAPDH
mRNA levels were need as an internal control
Western blotting
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
- 42 -
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 01 Tween 20 (TBST)
membranes were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-ERK or anti-phospho-ERK12 antibodies diluted at 11000 at 4 ˚C
for 16 h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Statistical analysis
All experiments were replicated at least three times Results are described as means plusmn SD
To compare the differentiation in P19CL6 cells stimulated with ECP at different
concentration and durations One-way ANOVA followed by post hoc turkey analysis was
employed to assess the significance between the groups more than two Marker gene
expression beating rate beating area were analyzed by paired T-tests The statistical
software used for analysis was SPSS (version 170) P lt 005 was considered statistically
significant
Results
Effect of ECP on early cardiomyocyte differentiation in P19CL6 cells
During the early stages of cardiomyocyte differentiation P19CL6 cells express the
mesenndomerm genes Wnt-3a and Brachyury [25 26] The expression of these two genes
was therefore assessed in the presence of ECP (Fig 1) Expression of Wnt-3a and
Brachyury were transiently increased within 2 days after treatment with both ECP and
DMSO However expression of both genes was delayed by 2 days without ECP with
maximum expression at 4 days with only DMSO In addition the level of Wnt-3a and
- 43 -
Brachyury expression after 2 days of ECP treatment was significantly higher than
expression levels ascertained after 4 days of treatment with only DMSO This delay may
suggest that ECP accelerated the process of cardiomyocyte differentiation from
mesendoderm up to beating cardiomyocytes with respect to generating a greater number of
beating cardiomyocytes at earlier time
We therefore evaluated the effect of ECP on the early stages of cardiomyocyte
differentiation in P19CL6 cells First the expression profiles of GATA-4 and α-MHC genes
were assessed when ECP treatment was limited for the first 2 days or 4 days (Fig2) Both
genes were upregulated showing a similar pattern with those when treated with ECP and
DMSO whole through the entire period This suggests that ECP should engage a rapid
change in gene expression with the first 48 hours following treatment with DMSO
To determine concentration of ECP ECP at 10100 and 1000 ngmL in the presence of
DMSO was evaluated for its ability to stimulate cardiomyocyte differentiation of P19CL6
cells in the presence of 1 DMSO (Fig 3) Cardiomyocyte differentiation was monitored
by assessing the expression of GATA-4 and α-MHC genes by real time qPCR ECP at 1000
ngmL was found to maximally induce the expression of GATA4 and α-MHC at the earliest
time of differentiation on day 8(Fig 3) The effect of different concentration of ECP was
further assessed on the expression of Brachyury at the earlier stages of within a 48 h
window in the absence of 1 DMSO treatment Brachyury gene expression was
upregulated not only in a dose-dependent manner of ECP (Fig 4A) but also in a
time-dependent manner(Fig 4B) Maximum upregulation occurred at 24 h following
treatment with ECP at 1000 ngmL However ECP alone did not induce end stage
cardiomyocyte differentiation as reflected by beating cardiomyocyte Stimulation with ECP
at 1000 ngmL for only 24 h was able to significantly induce GATA-4 and α-MHC
expression in P19CL6 cells in the presence of DMSO (Fig 5) as well as a beating
phenotype after 8 days Therefore cardiomyocyte differentiation of P19CL6 cells can be
induced by 1 DMSO and enhanced by the stimulation with ECP at 1000ngmL following
a relatively short 24 h treatment
- 44 -
Stimulation of ERK12 phosphorylation by ECP
Considering the result that ECP could induce Brachyury expression in P19CL6 cells within
24h we hypothesized that ECP might induce mesendoderm formation leading to
subsequent cardiomyocyte differentiation Since Brachyury is a marker for mesendoderm
ERK12 phosphorylation has been described important for mesoderm induction [26] (Yao
et al 2003) Hence the time course change in the phosphorylation of ERK12 was assessed
in P19CL6 cells when stimulated with ECP (Fig 6) ECP upregulated the phosphorylation
of ERK12 after 5 minutes stimulation while DMSO alone did not show any effect on
ERK12 phosphorylation FGF2 also stimulated phosphorylation of ERK12 in a pattern
which was similar to ECP stimulation of ERK12 phosphorylation The effect of FGF2 on
enhancing ERK12 phosphorylation was delayed by the inclusion of DMSO
FGFR signaling pathway is involved in the stimulation of p-ERK by ECP
Brachyury expression is regulated by FGF through an ERK12-dependent signaling
pathway that is involved in mesoderm induction [27] To investigate the possible
involvement of an FGF receptor (FGFR) signaling pathway in ECP stimulation inhibition
of FGFR1 with SU5402 was evaluated SU5402 suppressed the phosphorylation of ERK12
that was induced by ECP at 5 minute (Fig 7B and C) In addition the expression of
Brachyury that was induced by ECP was suppressed in the presence of SU5402 (Fig 8)
These results suggest that ECP utilize an FGFR signaling pathway to initiate mesoderm
induction in P19CL6 cells Furthermore the protractive differentiation of P19CL6 cells into
cardiomyocytes was evaluated in the continuous presence of ECP and SU5402 for 2 days
by the expression of GATA-4 and α-MHC These two genes were downregulated by
SU5402 even in the presence of DMSO or DMSO and ECP (Fig 9) In the presence of
SU5402 for 2 or 4 days no beating cardiomyocyte phenotype appeared at day 8 and 12 in
the presence of DMSO even after ECP stimulation Collectively the induction of mesoderm
lineages through an FGFR1 signaling pathway is essentially dependent on DMSO and
enhanced by ECP Interestingly FGF2 unlike ECP did not induce cardiomyocyte
differentiation in P19CL6 cells either in the presence or absence of DMSO In contrast
- 45 -
Brachyury expression was induced additively on the effect of DMSO by ECP while FGF2
did not induce this additive effect (Fig 8)
Fig 1 Effect of ECP on Expression of genes associated with mesoderm in the presence of
DMSO (A)(B) Expression of Wnt3a and Brachyury genes P19CL6 cells were treated with
or without ECP in the presence of DMSO Total RNA was isolated at indicated time points
- 46 -
and subjected to real time qPCR Each result is represented as mean plusmn SD from triplicate of
experiments
Fig 2 The effects of different duration of stimulation by ECP on cardiomyocyte
differentiation of P19CL6 cells (A) P19CL6 cells were treated with ECP for the first 2 and
4 days in the presence of DMSO as shown in the schematic diagram(B)(C) P19CL6 cells
were treated with or without ECP in the presence of DMSO Total RNA was isolated at
indicated time points and subjected to real time qPCR The expression of GATA4 and
α-MHC genes was evaluated at indicated days Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 47 -
Fig 3 Cardiomyocyte differentiation at different concentration of ECP (A)(B) P19CL6
cells were treated with indicated concentration of ECP for first 2 days in the presence of
DMSO The expression of GATA4 and α-MHC genes was evaluated at indicated days
Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) Each asterisk
shows the significance of Plt005
- 48 -
Fig 4 ECP induced mesoderm marker gene Brachyury expression without DMSO (A)
Time course of Brachyury induction by ECP P19CL6 cells were treated with different
concentration of ECP At indicated time points Total RNA was isolated from the cells and
subjected to real time qPCR (B) Brachyury induction by ECP P19CL6 cells were treated
for 24 hours with different concentration of ECP The expression of Brachyury was
upregulated in a dose-dependent manner (A) (B) Each result is represented as mean plusmn SD
from triplicate of experiments The statistic significance was assessed by one-way ANOVA
(see Material and Methods) Each asterisk shows the significance of Plt005
- 49 -
Fig 5 Cardiomyocyte differentiation of P19CL6 cells treated with ECP for one day (A)
Schematic diagram of the culturing conditions for P19CL6 cells with or without ECP for
one day in the presence of DMSO Total RNA was isolated at day 4 and day 8 (indicated by
asterisks) (B) Results from real time qPCR for the expression of cardiomyocyte marker
genes (GATA4 and α-MHC)
- 50 -
- 51 -
Fig 6 Effects of ECP FGF2 and DMSO on the phosphorylation of ERK12 in P19CL6
cells P19CL6 cells treated with ECP FGF2 and DMSO P19CL6 cells were analyzed by
western blot with anti-ERK antibody and anti-p-ERK12 antibody Each blot was
densitometrically quantified by Image J software In each panel the level of p-ERK12
after stimulated are presented as relative values against it at time 0 Total ERK12 was used
for normalization Each asterisk shows the significance of Plt005
- 52 -
Fig 7 Induction and phosphorylation of ERK12 by ECP is dependent on FGFR1 signaling
pathway (A) (B) Phosphorylation of ERK12 in P19CL6 cells by FGF2 ECP and its
inhibition by SU5402 (C) Phosphorylation of ERK12 in the P19CL6 cells by ECP in the
presence of DMSO All western blots were densitometrically quantified by Image J
software In each panel the level of p-ERK12 after stimulated are presented as relative
values against it in unstimulated cells Total ERK12 was used for normalization Each
asterisk shows the significance of Plt005
Fig 8 ECP induced Brachyury expression was suppressed by SU5402 in P19CL6 cells
P19CL6 cells were treated with DMSO ECP FGF and SU5402 for different combinations
Cells total RNA isolated at 24 hours and expression of Brachyury (a mesoderm marker)
was analyzed by real time qPCR The results are represented by mean plusmn SD from triplicate
of experiments Single-asterisk and double-asterisk show the significance of Plt005
- 53 -
Fig 9 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells in
the presence of DMSO (A)(B) The P19CL6 cells were treated with inhibitor SU5402 for
the first 2 and 4 days in the presence of DMSOGATA4 and α-MHC by real time qPCR
Cells were collected at indicated time points and analyzed cardiac marker genes expression
by real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
- 54 -
Fig 10 FGFR inhibitor SU5402 suppressed cardiomyocyte differentiation in P19CL6 cells
Cells were treated with or without ECP in the presence of DMSO (A)(B) (A) (B) Cells
were collected at indicated time points and analyzed cardiac marker genes expression by
real time qPCR The expression of GATA4 and α-MHC genes was evaluated at indicated
days Each result is represented as mean plusmn SD from triplicate of experiments The statistic
significance was assessed by one-way ANOVA (see Material and Methods) asterisk show
the significance of Plt005
Discussion
The eosinophil granulocyte takes part in the bodyrsquos natural defense against invading
parasites bacterials and virus [1-3] as well as in other inflammatory diseases like allergic
asthma and gastrointestinal disorders [4] Once attracted to the site of inflammation the
eosinophil becomes activated and as a result of this eosinophil degranulated mainly
releases secretes 4 proteins These are the major basic protein (MBP) present in their cores
surrounded by a matrix built up of eosinophil peroxidise (EPO) the eosinophil protein
Xeosinophil derived neurotoxin (EPXEDN) and ECP But it was not confirmed until to
- 55 -
1975 it become clear ECP was of eosinophil origin Since the discovery of ECP in 1971
more than 3000 articles have been published about ECP focus on dealing with molecular
and genetic characteristics and functions of ECP and the majority describing the
distribution of ECP in tissues and body fluids in patients with diseases such as asthma and
other inflammatory diseases
The major challengeproblem of using ES cell-based cardiomyocytes replacement therapy
is the inefficiency of the conventional protocol to generate cardiomyocytes [5 6] It is
crucial for researchers to find novel inducers of cardiomyocyte differentiation and to clearly
understand the mechanism of differentiation ECP can upregulate transforming growth
factor alpha expression in human lung fibroblast cells [7] and can enhance stress fiber
formation in mouse fibroblasts [8] Simultaneously we demonstrated that ECP induced the
differentiation of cardiomyocytes in rat neonates as judged by a increase in both the beating
rate and the expression of cardiac muscle specific markers such as atrial natriuretic factor
[8] In the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor ECP upregulated the expression of cardiomyocyte
marker genes such as GATA4 α-MHC β-MHC Nkx25 and MLC in the presence of DMSO
together with the immunostaining for actinin actin and cardiac troponin I (Fig 2C) In
the present study using P19CL6 cells we evaluated the potential of ECP as a
cardiomyocyte differentiation factor Actually ECP shortened the period of differentiation
by approximately at least 4 days until the appearance of a beating phenotype in P19CL6
cells while DMSO essentially primes differentiation in this system Early stages of
cardiomyocyte differentiation were reported to involve the transient upregulation of Wnt-3a
and Brachyury genes which were the typical markers of mesendoderm induction [9-11] As
shown in Fig 4 Brachyury expression was dependent on the dose of ECP in P19CL6 cells
The expression of Brachyury was maximally induced at 1000ngmL of ECP within 24 h
without DMSO
Wnt-3a induces mesoderm formation and cardiomyogenesis in embryonic stem cells [12]
Consistent with the result by Nakamula et al we observed temporal up-regulation of wnt3a
- 56 -
expression during DMSO induced the differentiation (Fig 3a) Wnt3a should activate
Wntβ-catenin signaling pathway presumably in autocrineparacrine manner which is
required for the cardiomyocyte differentiation in P19CL6 cells [13] We observed no
induction of Wnt3a expression by ECP alone (data not shown) In contrast ECP enhanced
and accelerated the expression of Wnt3a gene in presence of DMSO implicating that this
should be an indirect effect of ECP resulting from enhanced mesoderm differentiation
Extracellular signal-regulated kinase (ERK) was also shown to be necessary for mesoderm
differentiation in ES cells [14 15] In this study ECP was found to up-regulate the
phosphorylation of ERK12 in P19CL6 cells SU5402 a specific inhibitor of FGFR1 not
only downregulated phosphorylation of ERK12 but also inhibited Brachyury expression
abolishing the cardiomyocyte differentiation of P19CL6 cells following treatment with
ECP in the presence of DMSO ECP was found to stimulate the FGFR signaling pathway
However FGF2 did not enhance the cardiomyocyte differentiation in P19CL6 cells in the
presence of DMSO as ECP did FGF4 had the same negative result as FGF2 (data not
shown) FGF2 failed to enhance the Brachyury expression (Fig 8) and the phosphorylation
of ERK12 induced by FGF2 was delayed in the presence of DMSO (Fig 6) which differs
from the responses observed after ECP treatment These results might be due to the
binding of ECP to the FGFR which is different from FGF a region as conformation of the
FGF In this respect ECP has been reported to bind heparin and other glycosaminoglycans
like lectins through its RNase active site [16] FGF2 also binds to extracellular matrix
through syndecan 4 to exert some of its distinct biological activity [17] Binding of ECP to
the extracellular matrix components or cell surface GAG-containing proteins such as
syndecans and glypicans of P19CL6 cells should be explored to potentially explain the
modulation of FGFR signaling by ECP to enhance Brachyury expression The optimal
concentration of FGF2 to induce proliferation in fibroblasts is around 06 nM In this study
the optimal concentration of ECP was 1000ngmL which is equivalent to approximately 60
nM This 100-fold difference in biological activity may be due to different in binding
affinity to the FGFR by each protein ECP might be interacting with additional some
extracellular matrix proteins like syndecans which could alter the affinity of binding to the
- 57 -
FGFR Further studies are necessary to identify the effect of ECP on the activation of
FGFR signaling
SU5402 inhibited cardiomyocyte differentiation of P19CL6 cells in the presence of DMSO
Cardiomyocyte differentiation of P19CL6 cells appeared to depend on signaling through
the FGFR1 (Fig 9A) ECP induced ERK phosphorylation through FGFR1 while DMSO
alone did not affect the phosphorylation of ERK12 in P19CL6 cells (Fig 6)
Phosphatidylinositol 3-kinase (PI3K) plays an important role on the cardiomyocyte
differentiation of P19CL6 cells [18] Downstream intracellular signaling which is activated
by the FGFR could be due to both of PI3K-AKT and RAS-MAPK-ERK12 pathways [19]
ECP alone is not able to induce the cardiomyocyte differentiation in P19CL6 cells while
DMSO is essential for cardiomyocyte differentiation of P19CL6 cells The enhanced
phosphorylation of ERK12 that is induced by ECP through the FGFR1 should be closely
related with the accelerated cardiomyocyte differentiation because this phosphorylation was
not affected by DMSO while DMSO delayed the phosphorylation stimulated by FGF2
The expression of FGFR-1 in P19CL6 cells was confirmed by real time qPCR
(supplementary data) This might implicate our result that FGFR1 signaling pathway
triggered by ECP is involved in the accelerated differentiation of P19CL6 cell into
cardiomyocytes
This is the first study to report a function for ECP in cardiomyocyte differentiation from
stem cells in vitro We are now trying to develop ECP as cardiomyocyte differentiation
factor which could be used in the development of novel therapies for cardiac regeneration
- 58 -
Reference
1 McLaren DJ Peterson CG Venge P Schistosoma mansoni further studies of the
interaction between schistosomula and granulocyte-derived cationic proteins in vitro
Parasitol 1984 88491 503
2 Domachowske JB Dyer KD Adams AG Leto TL Rosenberg HF Eosinophil cationic
proteinRNase 3 is another RNase A-family ribonuclease with direct antiviral activity
(In process citation) Nucl Acids Res 1998 26 3358 63
3 Lehrer RI Szklarez D Barton A Ganz T Hamann KJ Gleich GJ Anti-bacterial
properties of eosinophil major basic protein (MBP) and eosinophil cationic protein
(ECP) J Immunol 1989 1424428 34
4 Venge P Eosinophils In Barnes P Rodger IA Thomson NC eds Asthma basic
Mechanisms and clinical management 3rd edn London Academic Press 1998141
57
5 Chan SS Chen JH Hwang SM Wang IJ Li HJ Lee RT Hsieh PC 2009 Salvianolic
acid B-vitamin C synergy in cardiac differentiation from embryonic stem cells
Biochem Biophys Res Commun 387723-728
6 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
embryonic and induced pluripotent stem cells PLoS One 5e14414
7 Chang HT Kao YL Wu CM Fan TC Lai YK Huang KL Chang YS Tsai JJ Chang
MD 2007 Signal peptide of eosinophil cationic protein upregulates transforming
growth factor-alpha expression in human cells J Cell Biochem 1001266-1275
8 Fukuda T Iwata M Kitazoe M Maeda T Salomon D Hirohata S Tanizawa K
Kuroda S Seno M 2009 Human eosinophil cationic protein enhances stress fiber
formation in Balbc 3T3 fibroblasts and differentiation of rat neonatal cardiomyocytes
Growth Factors 27228-236
- 59 -
9 Martin BL Kimelman D 2010 Brachyury establishes the embryonic mesodermal
progenitor niche Genes amp Development 242778-2783
10 Mauritz C Schwanke K Reppel M Neef S Katsirntaki K Maier LS Nguemo F
Menke S Haustein M Hescheler J Hasenfuss G Martin U 2008 Generation of
functional murine cardiac myocytes from induced pluripotent stem cells Circulation
118507-517
11 Mueller RL Huang C Ho RK 2010 Spatio-temporal regulation of Wnt and retinoic
acid signaling by tbx16spadetail during zebrafish mesoderm differentiation Bmc
Genomics 11492
12 Tran TH Wang X Browne C Zhang Y Schinke M Izumo S Burcin M 2009
Wnt3a-induced mesoderm formation and cardiomyogenesis in human embryonic
stem cells Stem Cells 271869-1878
13 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
14 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97144-151
15 Yao Y Li W Wu J Germann UA Su MS Kuida K Boucher DM 2003 Extracellular
signal-regulated kinase 2 is necessary for mesoderm differentiation Proc Natl Acad
Sci U S A 10012759-12764
16 Torrent M Odorizzi F Nogues MV Boix E 2010 Eosinophil cationic protein
aggregation identification of an N-terminus amyloid prone region
Biomacromolecules 111983-1990
17 Freund C Ward-van Oostwaard D Monshouwer-Kloots J van den Brink S van
Rooijen M Xu X
18 Giembycz MA Lindsay MA 1999 Pharmacology of the eosinophil Pharmacol Rev
51213-340
19 Gleich GJ 2000 Mechanisms of eosinophil-associated inflammation J Allergy Clin
- 60 -
Immunol 105651-663
20 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
21 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
22 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
23 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
24 Newton DL Nicholls PJ Rybak SM Youle RJ 1994 Expression and
Characterization of Recombinant Human Eosinophil-Derived Neurotoxin and
Eosinophil-Derived Neurotoxin-Anti-Transferrin Receptor Sfv J Biol Chem
26926739-26745
25 Jamali M Rogerson PJ Wilton S Skerjanc IS 2001 Nkx2-5 activity is essential for
cardiomyogenesis J Biol Chem 27642252-42258
26 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and
beta-catenin-dependent pathway for mammalian cardiac myogenesis Proc Nat Acad
Sci USA 1005834-5839
27 Schultemerker S Smith JC 1995 Mesoderm Formation in Response to Brachyury
Requires FGF Signaling Curr Biol 562-67
28 Chan SS Li HJ Hsueh YC Lee DS Chen JH Hwang SM Chen CY Shih E Hsieh
PC 2010 Fibroblast growth factor-10 promotes cardiomyocyte differentiation from
- 61 -
embryonic and induced pluripotent stem cells PLoS One 5e14414
29 Schultheiss TM Burch JB Lassar AB A role for bone morphogenetic proteins in the
induction of cardiac myogenesis Genes Dev 1997 11 451ndash462
30 Alsan BH Schultheiss TM Regulation of avian cardiogenesis by Fgf8 signaling
Development 2002 129 1935ndash1943
31 Marvin MJ Di Rocco G Gardiner A Bush SM Lassar AB Inhibition of Wnt activity
induces heart formation from posterior mesoderm Genes Dev 2001 15 316ndash327
32 Schneider VA Mercola M Wnt antagonism initiates cardiogenesis in Xenopus laevis
Genes Dev 2001 15 304ndash315
33 Tzahor E Lassar AB Wnt signals from the neural tube block ectopic cardiogenesis
Genes Dev 2001 15 255ndash260
34 Kawai T Takahashi T Esaki M Ushikoshi H Nagano S Fujiwara H Kosai K 2004
Efficient cardiomyogenic differentiation of embryonic stem cell by fibroblast growth
factor 2 and bone morphogenetic protein 2 Circ J 68691ndash702
- 62 -
- 63 -
Chapter 3
Effect of ECP on the expression of genes correlated
with cardiomyocyte differentiation in P19CL6 cells
- 64 -
Abstract
We all know that cardiomyocyte differentiation is a complex process and so many signaling
pathways are included (such as FGF signaling Wnt signaling Eendothelin 1 (ET-1)
endothelin receptor A (ET-A) criptonodal signaling and so on) ECP effects are dependent
on DMSO on inducing cardiomyocyte differentiation in P19CL6 cells From chapter 3 we
have shown that ECP enhanced cardiomyocyte differentiation by stimulating FGFR1
signaling pathway In this chapter we checked effects of ECP on other signaling pathways
DMSO up-regulated Wnt3a expression and Wnt3a activated Wntβ-catenin signaling
pathway in autocrine manner This is required for the cardiomyocyte differentiation in
P19CL6 cells ECP alone can not induce Wnt3a expression However in the presence of
DMSO ECP accelerated Wnt3a suggested ECP stimulation have some positive effects on
the Wntβ-catenin signaling pathway in the presence of DMSO ECP activated PI3KAkt
signaling pathway It was well known that PI3KAkt signaling pathway is pivotal for
β-catenin (a down-stream molecule of Wnt) stabilization DMSO and ECP alone activated
this signaling respectively and ECP increased effects of DMSO on this signaling pathway
Through TOPFOP FLASH luciferase assay even if ECP did not induced Wnt3a expression
but it increased luciferase activity From above we get summary that ECP directly activated
PI3KAkt signaling and Wntβ-catenin pathway which was activated by DMSO
subsequently promote differentiation In addition we also confirmed that ECP also
enhanced EphA1 and ET-1 expression by DMSO both of signaling pathway correlated
with cardiomyocyte differentiation From above data it suggested that ECP through direct
and indirect manner activated signaling pathway subsequently influenced cardiomyocyte
differentiation
- 65 -
Introduction
It is well known that cardiomyocyte differentiation is complicated and so many signaling
pathways are included Proteins of the Wnt family are known regulators of
Cardiomyogenesis Wnts could bind to their transmembrane receptors Frizzled and
co-receptors LRP 56 on target cells to activate different signaling pathways Activation of
the canonical pathway leads to stabilization of β-catenin through inactivation of GSK3β
(Glycogen Synthase Kinase-3-β) β-catenin -dependent activation of TCF (T Cell Factor)
LEF (Lymphoid Enhancer Factor) transcription factors and induction of Wnt-responsive
genes This is canonical Wntβ-catenin signaling pathway In contrast Wnt11 signals
through a β-catenin -independent non-canonical pathway involving PKC (Protein
Kinase-C) and JNK (Jun-N-terminal kinase) Wnts induced Myogenic specification and
mammalian Cardiac myogenesis Wnt3a (Wingless-related MMTV integration site-3a)
upregulated early Cardiac marker genes before gastrulation in Mouse through stabilizing
β-catenin but inhibits heart formation during gastrulation [9] It suggests that
Wntβ-catenin signaling pathway plays a biphasic role in cardiomyocyte differentiation
Activation is required to commit mesenchymal cells to the cardiac lineage downregulation
of β-catenin is needed for cardiomyocyte differentiation at later stages Activation of
Wntβ-catenin signaling pathway during early EB formation enhances mouse ES cell
differentiation into cardiomyocytes and suppresses the differentiation into hematopoietic
and vascular cell lineages [10]
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including humans [14 17 19] mice [16 20 21] rats [13 15] zebrafish [23] and
chickens [12 16 18] Addictively EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In addition to both of signaling above
- 66 -
NodalCripto signaling Endothelin 1 (ET-1) Endothelin 1 receptor A (ET-A) signaling
and Notch signaling have been investigated that these signaling are correlated with
cardiomyocyte differentiation during development stage In this report we are deciphering
the roles of ECP on these signaling pathways during cardiomyocyte differentiation in
P19CL6 cells
Materials and Methods
Cell culture
P19CL6 cells which were obtained from RIKEN Bioresource Center Cell Bank Japan
were cultured essentially as described previously[25 26] (Monzen et al 2001 Monzen et
al 1999) Briefly the cells were grown in 100-mm tissue culture dishes under adherent
conditions with α-minimum essential medium (α-MEM) (Invitrogen Tokyo Japan)
containing 10 fetal bovine serum (FBS) (Invitrogen) as growth medium in a 5 CO2
atmosphere at 37degC To induce cardiomyocyte differentiation under adherent conditions
P19CL6 cells were plated at a density of 5times105 in 60-mm tissue culture dishes in growth
medium Twenty four hours later the medium was replaced with growth medium
containing 1 DMSO (Nacalai tesque Kyoto Japan) as differentiation medium The
medium was changed to every 2 days and the cells were maintained under fresh conditions
until they started beating
Reagents and antibodies
Recombinant human ECP was expressed in bacteria and prepared as described previously
[27 28] Epha1 siRNA and transfection reagent were purchased from Qiagen Anti-Akt was
prepared in our lab and anti-phospho-Akt mouse monoclonal antibodies were from New
England Biolabs (Beverly MA) All other reagents were of analytical grade and were
purchased from Wako or Sigma-Aldrich unless otherwise noted
Real time qPCR
- 67 -
P19CL6 cells cultured under various conditions were harvested and total RNA was
extracted with RNeasy Mini Kit (QIAGEN MD) In order to exclude the contamination of
genomic DNA the extracted RNA was treated with RNase-Free DNase1 (Takara Japan)
Five micrograms of total RNA was then used to synthesize first-strand cDNA with
oligo-dT18 and 5 unitsμL of SuperScript Ⅱ (Invitrogen CA) in a reaction volume of 20 μL
following the manufacturerrsquos instructions For quantitative analysis of gene expression
levels real time qPCR were performed and the data were normalized to GAPDH Primers
used for the real time qPCR were as following (forward and reverse)
Wnt3a 5-AAGCAGGCTCTGGGCAGCTA-3 and 5-GACGGTGGTGCAGTTCCA-3
OCT4 5-TCTTTCCACCAGGCCCCCGGCTC-3 and
5-TGCGGGCGGACATGGGGAGATCC-3 FIk1
5-TAGCTGTCGCTCTGTGGTTCTG-3 and 5-GTCTTTCTGTTGTGCTGAGCTTGG-3
Nodal 5-TGCTGAAACGATACCAACCC-3 and
5-CCTGCCATTGTCCACATAAAGC-3 Frizzled 1
5-GCGACGTACTGAGCGGAGTG-3 and 5-TGATGGTGCGGATGCGGAAG-3
Frizzled 2 5-CTCAAGGTGCCGTCCTATCTCAG-3 and
5-GCAGCACAACACCGACCATG-3 Frizzled 4
5-GACAACTTTCACGCCGCTCATC-3 and
5-CCAGGCAAACCCAAATTCTCTCAG-3 Frizzled 8
5-GTTCAGTCATCAAGCAGCAAGGAG-3 and 5-AAGGCAGGCGACAACGACG-3
Frizzled 10 5-ATCGGCACTTCCTTCATCCTGTC-3 and
5-TCTTCCAGTAGTCCATGTTGAG-3 ET-1 5-ACTTCTGCCACCTGGACATC-3 and
5-ACTTTGGGCCCTGAGTTCTT-3 ET-A 5-ACCGTCTTGAACCTCTGTGC-3 and
5-AGCCACCAGTCCTTCACATC-3 Notch
5-AGATTGAGGCCGTGAAGAGTGAGC-3 and
5-CCACAAAGAACAGGAGCACGAAGG-3 EphA1
5-AACCTTATGCCAACTACACC-3 and 5-TTCCCCAAACTCTCCTTCTC-3
Luciferase assays
- 68 -
Plasmids TOPFlash contains TCF-LEF binding site upstream of the thymidine kinase
minimal promoter and luciferase cDNA Negative control counterpart FOPFlash has a
mutated TcfLef binding site (Upstate Biotechnology) P19CL6 cells were transfected by
using FuGENE 6 (Roche) with 02 μg of plasmid per 1times105 cells Luciferase activity was
determined by using a commercially available assay system (Promega) and was normalized
for transfection efficiency with pcDNA3-β-galactosidase
Western blot
Cells grown on culture dishes were washed twice with ice-cold PBS and then suspended in
50 mM TrisndashHCl pH 74 containing 150 mM NaCl 1 mM EDTA 1 NP-40
supplemented with phosphatase inhibitors 10 mM sodium fluoride and 1 mM
phenylmethylsulfonyl fluoride 1 mM sodium orthovanadate and 10 mM sodium
pyrophosphate The cells were then disrupted by a sonicator (Ultras Homogenizer VP-53
TAITEC JP) with a microprobe setting at level 2 for 30 sec on ice and centrifuged at
13000g for 20 min at 4degC The supernatants were then carefully collected and stored at
-80degC until use Protein concentration was determined by BCA Protein Assay kit (Pierce
IL) and 20 microg of protein was subjected to 125 SDS polyacrylamide gel electrophoresis
and subsequently transferred onto PVDF membranes (Millipore MA) by semidry blotting
After being washed with Tris-buffered saline pH 76 containing 005 Tween 20 (TBST)
membrane were incubated for 1 h at 25 ˚C in TBST containing 5 BSA and the antigens
were probed with anti-AKT or anti-phospho-Akt antibodies diluted at 11000 at 4 ˚C for 16
h After three washes with TBST membranes were probed with
horseradish-peroxidase-conjugated anti-rabbit antibody diluted at 12000 for 1 h at 25 ˚C
Following by three washes with TBST horseradish-peroxidase on the membrane was
reacted with the substrate kit (Western lightening plus-ECL Western blotting detection
reagents PerkinElmer US) and then detected by LSA-4000 (FUJIFILM Japan)
Microrarray analysis
Expression of genes in P19CL6 cells was analyzed by microarray procedure Total RNA
- 69 -
preparation and analysis was performed as described previously [29 30] DNA microarray
was carrying 1428-oligonuclotide probes for human cell surface proteins cDNAs were
synthesized with Superescript Ⅱ reverse transcriptase (invitrogen) with oligo dT primers
Amino-allyl-dUTP was incorprated into cDNAs followed by coupling with Cy-3 dye
(Ambion TX) and were processed for hybridization at 55 for 15 hours The fluorescent
images for the hybridization were captured using FLA8000 scanner (Fuji film Japan) and
analyzed with GenePix Pro51 software (Axon Instruments CA) We found ECP or DMSO
only upregulated EphA1 expression in P19CL6 cells Moreover EphA1 expression was
enhanced in P19CL6 cells treated with ECP in the presence of DMSO Real time qPCR
further confirmed the results (Fig) Real time qPCR was performed with SYBR Green Real
time Master Mix (toyobo) in triplicates containing 5 ng of cDNA along with 400 nM
primers using LightCycler (Roche) The thermal cycling condition for real time qPCR was
as follows 95degC for 5 min followed by 40 cycles of denaturation at 95degC for 10 sec
annealing at 58degC depending on the melting temperature of each pair of primers for 10 sec
followed by extension at 72degC for 10s The following sets of primers were used for the
PCR reaction EphA1 5-TCACGCTGCCTCTGCTGTCT-3 and
5-GCCCACCAGTTTCCAGAAGCCT-3
EphA1 siRNA transfection
P19CL6 cells were plated at a density of 04times105 well in 24 well plates in growth medium
and incubated one hour in a 5 CO2 atmosphere at 37degC EphA1 siRNA and control
siRNA 75 ng were diluted with 100 μl OPTI-MEM culture medium with no FBS final
concentration is 10 nM and mix with 3μl HiperFect Transfection Reagent by vortexing
then incubated 10 minutes at room temperature The mixtures were then added dropwise
into the well with cells and siRNA concentrations were obtained to 10 nM in the medium
After 24 hours incubation the cells were induced differentiation in the presence of DMSO
with or without ECP Real time qPCR was performed to analysis the knockdown efficiency
of EphA1 and cardiomyocyte differentiation
- 70 -
Results
Effect of ECP on the Wntβ-catenin signaling pathway
Because canonical Wnt signaling pathway plays an essential role in cardiomyocyte
differentiation so in this chapter I first examined Wnt3a expression in P19CL6 cells
treated with different concentration of ECP I observed no induction of Wnt3a expression
by ECP alone (Fig 1) In contrast ECP enhanced and accelerated the expression of Wnt3a
gene in presence of DMSO Luciferase activity in P19CL6 cells tranfected with
TOPFLASH plasmid (Fig 2) which reflects the stabilization ofβ-catenin was elevated on
day 1 and 2 after stimulation of ECP withwithout DMSO And then we analyzed the Wnt
receptor genes (Frizzleds) expression ECP accelerated and enhanced frizzled 4 and 10 (Fig
3C and E) and enhanced Frizzled 1 (Fig 3A) expression in P19CL6 cells in the presence of
DMSO during cardiomyocyte differentiation However expression of frizzled 2 and 8 genes
has no significant change These results indicate that ECP activated Wnt signaling pathway
through directly enhanced stabilization of β -catenin and subsequently enhanced
cardiomyocyte differentiation
Wnt3a
0
02
04
06
08
1
12
14
0 3 6 12 24 48
hours
Relative Expression
ECP 0ngml
ECP10ngml
ECP100ngml
ECP1000ngml
- 71 -
Fig 1 Effect of ECP on Wnt3a gene expression P19CL6 cells were treated with different
concentration of ECP for the first 2 days At indicated time points Total RNA was isolated
from the cells and subjected to real time qPCR ECP have no significant influence on the
Wnt3a gene expression
A B
0
05
1
15
2
25
3
35
con DMSO DMSO-ECP
ECP
Rel
ativ
e lu
cife
rase
act
ivit
y
TOPFLASHFOPFLASH
0
50
100
150
200
250
300
con DMSO DMSO-ECP
ECP
Rel
ativ
e L
ucif
eras
e ac
tivi
ty
TOPFLASHFOPFLASH
Fig 2 Effects of ECP on stabilization of β-catenin (A)(B) P19CL6 cells were transfected
with TOPFlash or FOPFlash plasmid in a concentration of 02 μg per 1times105 cells overnight
and then were stimulated with DMSO ECP for 1 day and 2 days respectively Luciferase
activity was shown as above the figures
A B
Frizzled-1
0
1
2
3
4
5
6
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-2
0
05
1
15
2
25
3
35
4
45
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime Course (days)
Relative expression
DMSO
DMSO-ECP
- 72 -
C D
Frizzled-4
0
5
10
15
20
25
30
35
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Frizzled-8
0
2
4
6
8
10
12
14
con 1 d 2 d 4 d 6 d 8 d 10 d 12 dTime course (days)
Relative expression
DMSO
DMSO-ECP
E
Frizzled-10
0
10
20
30
40
50
60
con 1 d 2 d 4 d 6 d 8 d 10 d 12 d
Time course (days)
Relative expression
DMSO
DMSO-ECP
Fig 3 Effect of ECP on expression of Wnt receptor genes (Frizzleds ) in the presence of
DMSO (A) Frizzled 1 (B) Frizzled 2 (C) Frizzled 4 (D) Frizzled 8 (E) Frizzled 10
(A)-(E) P19CL6 cells were treated with or without ECP in the presence of DMSO Total
RNA was isolated at indicated time points and subjected to real time qPCR
Effects of ECP and DMSO on the phosphorylation of AKT in P19CL6 cells
- 73 -
Recently it has been reported that activation of PI3K is required for DMSO-induced
cardiomyocyte differentiation of P19CL6 cells Akt is one of the important downstream
molecules of PI3K Hence the time course change in the phosphorylation of Akt was
assessed in P19CL6 cells when stimulated with ECP (Fig 4) Phosphorylation of Akt was
upregulated in P19CL6 cells by ECP stimulation after 5 minutes in the presence of DMSO
or not
Fig 4 Effects of ECP and DMSO on the phosphorylation of Akt in P19CL6 cells P19CL6
cells treated with ECP and DMSO P19CL6 cells were analyzed by western blot with
anti-AKT antibody and anti-p-AKT antibody
Effect of ECP on Expression of Endothelin 1 (ET-1) and Endothelin receptor A genes
(ET-A) in the presence of DMSO
Papers have already published that ET-1 and ET-A are essential for cardiomyocyte
differentiation in P19 cells [35] Here we examined the expression of ET-1 and ET-A genes
by real time qPCR ECP enhanced both of genes expression in P19CL6 cells in the
presence of DMSO from 4 days (Fig 5) And then we used antagonist of ET-A (BQ123)
- 74 -
and ET-A and ET-B (another receptor of endothelins) (Bosentan) to treat P19CL6 cells in
the presence of DMSO then check the cardiomyocyte differentiation by real time qPCR
BQ123 and Bosentan suppressed the expression of cardiomyocyte marker gene in the
presence of DMSO (GATA4) (Fig 6) It suggested that ECP have a positive effect on
endothelins signaling
A B
ET-1
0
20
40
60
80
100
120
0 1 2 4 6 8 10 12
Time Course (day)
Rel
ativ
e ex
pres
sion
DMSO
DMSO-ECP
ET-A
0
50
100
150
200
250
300
0 1 2 4 6 8 10 12
Time Course (day)
Relative expression
DMSO
DMSO-ECP
Fig 5 Effect of ECP on Expression of ET-1 and its receptor genes (ET-A ) in the presence
of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence of
DMSO Total RNA was isolated at indicated time points and subjected to real time qPCR
A B
GATA4
0
10
20
30
40
50
60
70
0 4 8
Time Course (day)
Rela
tive
exp
ress
ion
DMSO-BQ123
DMSO-BS
DMSO
MLC2
0
5
10
15
20
25
0 4 8
Time course(day)
Rela
tive
exp
ress
ion
DMSO
DMSO-BQ123
DMSO-BS
- 75 -
Fig 6 Effect of endothelins receptor A and receptor B antagonists (BQ123 and Bosentan)
on cardiomyocyte differentiation in P19CL6 cells in the presence of DMSO (A)(B) The
P19CL6 cells were treated with antagonist of endothelins receptor Bosentan (TRC) and
BQ-123(MERCK) in the presence of DMSO Total RNA was isolated at indicated time
points and analyzed cardiac marker genes expression by real time qPCR The expression of
GATA4 and α-MHC genes was evaluated at indicated days
Effect of ECP on NodalCripto Notch signaling pathway
Cripto an essential co-receptor of Nodal acts together with activin receptors to transmit the
Nodal signal via Smad2 and Smad3 [31-34] Previous studies have shown a requirement for
Nodal signaling in mesoderm differentiation in P19cells According to this point we
examined the expression of Nodal and its co-receptor Cripto genes Before 4 days both of
genes expression level was higher in P19CL6 cells treated with ECP in the presence of
DMSO over 4 days expression of two genes in the cells with ECP was significantly
downregulated however without ECP expression of two genes still maintained high level
from 4 days to eight days(Fig 7) Li et al reported that activation of Notch signaling in
undifferentiated P19CL6 cells promoted cardiac differentiation in this chapter ECP
accelerated Notch expression in the presence of DMSO (Fig 8)
A B
Fig 7 Effect of ECP on the expression of Nodal and its receptor (Cripto) gene in the
- 76 -
presence of DMSO (A) (B) P19CL6 cells were treated with or without ECP in the presence
of DMSO Total RNA was isolated at indicated time points and subjected to real time
qPCR
Fig 8 Effect of ECP on Expression of notch genes in the presence of DMSO (A) (B)
P19CL6 cells were treated with or without ECP in the presence of DMSO Total RNA was
isolated at indicated time points and subjected to real time qPCR
Effect of ECP on EphA1 induction in P19CL6 cells with or without DMSO
Jason D et al investigated the potential role of Eph-related molecules during very early
embryonic development [36] In this chapter first of all we examined the Epha1
expression by using microarray at indicated days ECP upregulated the gene expression in
P19CL6 cells in the presence of DMSO or not To confirm this we checked the gene
expression by real time qPCR at indicated days (Fig 9) and got similar result with
microarray assays ECP treatment upregulated the expression of Epha1 gene Moreover
ECP enhanced Epha1 expression in the presence of DMSO compare to DMSO alone The
highest expression level of EphA1 was observed at one day These results indicated that
ECP activated EphA1 signaling pathway during very early embryonic development
- 77 -
Fig 9 Effect of ECP on the expression of EphA1 gene in the presence of DMSO P19CL6
cells were treated with or without ECP in the presence of DMSO Total RNA was isolated
at indicated time points and subjected to real time qPCR
Fig 10 EphA1 siRNA transfection P19CL6 cells were transfected with EphA1 siRNA
control means mock siRNA Total RNA was isolated at indicated time Knockdown
efficiency was examined by real time qPCR Results showed that EphA1 siRNA
suppressed EphA1 expression significantly
The roles of EphA1 signaling in the cardiomyocyte differentiation of P19CL6 cells
In order to investigate the roles of EphA1 signaling in the cardiomyocyte differentiation of
P19CL6 cells I knocked down the expression of EphA1 by siRNA Knockdown efficiency
- 78 -
was examined by real time qPCR EphA1 siRNA suppressed EphA1 expression
significantly (Fig 10) It is well known that cardiomyocyte differentiation of embryonic
stem cells is a very complicated procedure Cardiomyocyte differentiation of P19CL6 cells
underwent several stages denoted by the temporal expression of specific markers in the
presence of DMSO (Figure 11A and 11B) Undifferentiated P19CL6 cells (day 0) highly
expressed Oct4 which gradually diminished after differentiation Mesodermal cells
expressed Brachyury from day 1 to day 4 and cardiovascular progenitors the offspring of
mesodermal cells expressed Flk-1 from day 4 to day 8 Cardiac precursors started to appear
on day 4 indicated by the expression of Gata4 Cardiomyocytes as denoted by the
expression of the cardiac structural markers α-MHC expressed α-MHC appeared on day 12
However when the cells were treated with ECP the marker genes expression was
accelerated almost 4 days In order to examine effect of Epha1 signaling on cardiomyocyte
differentiation from P19CL6 cells we transfected P19CL6 with EphA1 siRNA after 1 day
induced P19CL6 cells with or without ECP in the presence of DMSO As shown in Fig 1
expression of these marker genes was suppressed (Fig 12) when the EphA1 gene was
knocked down even if mesoderm marker gene Brachyury expression These results
indicated that EphA1 signaling have a positive effect on mesoderm formation and then
influenced subsequent cardiomyocyte differentiation This is consistent with Jason D et al
reported that Eph-related molecules have a potential role during very early embryonic
development
A
- 79 -
B
Fig 11 Effects of ECP on different development stage of cardiomyocytes (A)The schemic
showed that marker genes expression at different stage of cardiomyocyte differentiation
(B) Marker genes expression was analyzed by real time qPCR P19CL6 cells were treated
with or without ECP in the presence of DMSO Total RNA was isolated at indicated time
points and subjected to real time qPCR
- 80 -
- 81 -
Fig 12 Effect of EphA1 siRNA on cardiomyocyte differentiation of P19CL6 cells
RT-qPCR results showed temporal expression patterns of these marker genes on days 1 2
4 and 8 during cardiomyocyte differentiation from P19CL6 cells Mock siRNA serve as
control
Discussion
Wntβ-catenin signaling plays pivotal roles in development of multiple tissues through
regulation of cell proliferation differentiation migration and gene expression [4]
Wntβ-catenin signaling pathway is focused on β-catenin stabilization In the inactivation of
canonical Wnt signaling pathway β-catenin complexed with APC and AXIN is
phosphorylated by glycogen synthase kinase 3β (GSK3β) in a degradation box which is
polyubiquitinated for proteasomal degradation [5] In the presence of Wnt ligands binding
to its receptor complex GSK3β is inhibited leading to stabilization of cytoplasmic
β-catenin Accumulated cytoplasmic β-catenin subsequently translocates to the nucleus and
initiates target gene transcription through T-cell factor (TCF)-lymphoid enhancer factor
(LEF) transcription factors [6 7] Wntβ-catenin signaling is critical for vertebrate cardiac
development This is same with our results Ablation of β-catenin in Isl1- expressing
cardiovascular progenitor cells disrupts multiple aspects of cardiogenesis resulting in
embryonic lethality at E13 [8] Other studies have shown a biphasic role of β-catenin in
cardiac specification The early activation of Wntβ-catenin signaling promotes cardiac
differentiation whereas the activation of Wntβ-catenin signaling at the later stage inhibits
heart formation [9] It has also been shown that the canonical Wnt signaling pathway
promotes commitment of P19CL6 cells into cardiac lineage at the early stage inhibiting
further differentiation into mature cardiomyocytes at the later stage [10] In this chapter we
examined the luciferase activity (Fig 2) which reflects the activity of canonical Wnt
signaling In P19CL6 cells luciferase activity was elevated on day 1 and 2 It implicated
that ECP had ability to activate Wnt signaling To confirm this we further examined the
- 82 -
expression of Frizzled genes Frizzled 1 4 10 three genes were upregulated in P19CL6
cells treated with ECP in the presence of DMSO (Fig 3) However after checked the
expression of Wnt3a by real time qPCR I found the gene expression was not upregulated in
P19CL6 cells treated with ECP These results suggested that activation of Wnt signaling is
required for the cardiomyocyte differentiation early event ECP activated Wnt signaling
through enhancing stabilization of β-catenin
Many studies have indicated that PI3KAkt signaling is involved in the differentiation of
various kinds of cells such as myoblasts[1] osteoblasts[2] and adipocytes[3] PI3K is
critically involved in cardiomyocyte differentiation in vitro It was reported that activation
of this signaling was required for cardiomyocyte differentiation of P19CL6 cells [10]
Activation of PI3KAkt pathway increased the content of cytoplasmic and nuclear β-catenin
and the activity of canonical Wnt pathway through increased GSK-3β phosphorylation So
in this study we examined phosphorylation of Akt which is important molecule
downstream of PI3K DMSO and ECP upregulated phosphorylation of Akt in P19CL6 cells
at 30 minutes This is consistent with Atsuhiko T et al reported that activation of
phosphatidylinositol 3-kinase (PI3K) plays a critical role in the early stage of
cardiomyocyte differentiation of P19CL6 cells [10]
Juan Carlos Monge et al reported that ET-1 and its receptor ET-A play critical role in
cardiomyocyte differentiation in P19 cells [35] In this chapter we detected both of genes
expression in P19CL6 cells with or without ECP in the presence of DMSO ECP enhanced
expression of two genes after 4 days compared to DMSO only It suggested that ECP had
positive effects on ET-1 and ET-A signaling and then enhanced cardiomyocyte
differentiation This is consistent with Juan Carlos Monge et al reported In addition we
used antagonist of ET-A to treat P19CL6 cells in the presence of DMSO GATA4
expression was downregulated And then we checked other genes expression correlated
with cardiomyocyte differentiation such as Nodal and its co-receptor Cripto genes
expression Notch expression ECP accelerated these genes expression Previous studies
have shown a requirement for Nodal Cripto signaling in mesoderm differentiation in P19
- 83 -
cells it implicated that ECP accelerated the mesoderm formation This is similar with our
result in chapter 2
Eph was the first isolated member of what is currently the largest subfamily of RTKs
[11-23] This group is distinguished by a cysteine-rich region and two fibronectin type III
repeatsin the extracellular domain [12] Eph-subfamily kinases have been found in diverse
species including zebrafish [23] chickens [12 16 18] mice [16 20 21] rats [13 15] and
humans [14 17 19] Additionally EphA1 have a high expression level in embryonic stem
cells Features of their expression pattern suggest key functions during embryonic
development Jason D et al reported that Eph-related molecules have a potential role
during very early embryonic development [36] In this chapter I found that ECP enhanced
the expression of EphA1 in P19CL6 cells treated with ECP ECP with DMSO and DMSO
alone by microarray assay To further confirm this I used real time qPCR to examine again
I got the same results (Fig 9) as microarray assayed Furthermore I used siRNA of EphA1
to treated P19CL6 cells with or without ECP in the presence of DMSO checked the temple
profile marker genes expression during cardiomyocyte differentiation Knockdown of
EphA1 expression of stem cells marker gene OCT4 keep a high level In contract
Brachyury gene an early mesoderm marker gene expression was suppressed subsequently
influenced cardiomyocyte differentiation (Fig 11) These results were similar with Jason D
et al reported that Eph-related molecules have a potential role during very early embryonic
development [36]
From above data we summarize that ECP enhanced cardiomyocyte differentiation induced
by DMSO at early stage through initiating mesoderm differentiation by influencing several
signaling pathwaysuch as FGF Wnt PI3K EphA1
- 84 -
Reference
1 Jiang BH Aoki M Zheng JZ Li J Vogt PK Myogenic signaling of
phosphatidylinositol 3-kinase requires the serine-threonine kinase Aktprotein kinase
B Proc Natl Acad Sci U S A 1999 96 2077ndash2081
2 Ghosh-Choudhury N Abboud SL Nishimura R Celeste A Mahimainathan L
Choudhury GG Requirement of BMP-2-induced phosphatidylinositol 3-kinase and
Akt serinethreonine kinase in osteoblast differentiation and Smad-dependent BMP-2
gene transcription J Biol Chem 2002 277 33361ndash33368Epub 2002 Jun 25
3 Magun R Burgering BM Coffer PJ Pardasani D Lin Y Chabot J Sorisky A
Expression of a constitutively activated form of protein kinase B (c-Akt) in 3T3ndashL1
preadipose cells causes spontaneous differentiation Endocrinology 1996 137
3590ndash3593
4 Logan CY Nusse R 2004 The Wnt signaling pathway in development and disease
Annu Rev Cell Dev Biol 20 781ndash810
5 Katoh M Katoh M 2007 WNT signaling pathway and stem cell signaling network
Clin Cancer Res 13 4042ndash4045
6 Nakamura T Sano M Songyang Z Schneider MD 2003 A Wnt- and beta
-catenin-dependent pathway for mammalian cardiac myogenesis Proc Natl Acad Sci
USA 100 5834ndash5839
7 Deb A Davis BH Guo J Ni A Huang J Zhang Z Mu H Dzau VJ 2008 SFRP2
regulates cardiomyogenic differentiation by inhibiting a positive transcriptional
autofeedback loop of Wnt3a Stem Cells 26 35ndash44
8 Lin L Cui L Zhou W Dufort D Zhang X Cai CL Bu L Yang L Martin J Kemler R
Rosenfeld MG Chen J Evans SM 2007 Beta-catenin directly regulates Islet1
expression in cardiovascular progenitors and is required for multiple aspects of
cardiogenesis Proc Natl Acad Sci USA 104 9313ndash9318
9 Ueno S Weidinger G Osugi T Kohn AD Golob JL Pabon L Reinecke H Moon RT
- 85 -
Murry CE 2007 Biphasic role for Wntbeta-catenin signaling in cardiac specification
in zebrafish and embryonic stem cells Proc Natl Acad Sci USA 104 9685ndash9690
10 Naito AT Akazawa H Takano H Minamino T Nagai T Aburatani H Komuro I 2005
Phosphatidylinositol 3-kinase-Akt pathway plays a critical role in early
cardiomyogenesis by regulating canonical Wnt signaling Circ Res 97 144ndash151
11 Hirai H Maru Y Hagiwara K Nishida J amp Takaku F 1987 A novel
putativetyrosine kinase receptor encoded by the eph gene Science 238 1717-1720
12 Pasquale E B 1991 Identification of chicken embryo kinase 5 a developmentally
regulated receptor-type tyrosine kinase of the Eph family Cell Regul 2 523-534
13 Letwin K Yee S P amp Pawson T 1988 Novel protein-tyrosine kinase cDNAs
related to fpsfes and eph cloned using anti-phosphotyrosine antibody Oncogene 3
621-627
14 Lindberg R A amp Hunter T 1990 cDNA cloning and characterization of eck an
epithelial cell receptor protein-tyrosine kinase in the ephelk family of protein kinases
Mol Cell Biol 10 6316-6324
15 Chan J amp Watt V M 1991 eek and erk new members of the eph subclass of
receptor protein-tyrosine kinases Oncogene 6 1057-1061
16 Sajjadi F G Pasquale E B amp Subramani S 1991 Identification of a new
eph-related receptor tyrosine kinase gene from mouse and chicken that is
developmentally regulated and encodes at least two forms of the receptor New Biol
3769-3778
17 Boyd A W Ward L D Wicks I P Simpson R J Salvaris E Wilks A Welch
K Loudovaris M Rockman S ampBusmanis I 1992 Isolation and characterization
of a novel receptor-type protein tyrosine kinase (hek) from a human pre-B cell line J
Biol Chem 267 3262-3267
18 Wicks I P Wilkinson D Salvaris E amp Boyd A W 1992 Molecular cloning of
HEK the gene encoding a receptor tyrosine kinase expressed by human lymphoid
tumor cell lines Proc Natl Acad Sci USA 89 1611-1615
19 Sajjadi F G amp Pasquale E B 1993 Five novel avian Eph-related tyrosine kinases
- 86 -
are differentially expressed Oncogene 8 1807-1813
20 Bohme B Holtrich U Wolf G Luzius H Grzeschik K H Strebhardt K amp
Rubsamen W H 1993 PCR mediated detection of a new human
receptor-tyrosine-kinase HEK 2 Oncogene 8 2857-2862
21 Maisonpierre P C Barrezueta N X amp Yancopoulos G D 1993 Ehk-1 and Ehk-2
two novel members of the Eph receptor-like tyrosine kinase family with distinctive
structures and neuronal expression Oncogene 8 3277-3288
22 Henkemeyer M Marengere L E McGlade J Olivier J P Conlon R A
Holmyard D P Letwin K amp Pawson T 1994 Immunolocalization of the Nuk
receptor tyrosine kinase suggests roles in segmental patterning of the brain and
axonogenesis Oncogene9 1001-1014
23 Xu Q Holder N Patient R amp Wilson S W 1994 Spatially regulated expression
of three receptor tyrosine kinase genes during gastrulation in the zebrafish
Development Cambridge UK 120 287-299
24 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd 1996 Embryonic
stem cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad
Sci U S A January 9 93(1) 145ndash150
25 Monzen K Hiroi Y Kudoh S Akazawa H Oka T Takimoto E Hayashi D Hosoda T
Kawabata M Miyazono K Ishii S Yazaki Y Nagai R Komuro I 2001 Smads
TAK1 and their common target ATF-2 play a critical role in cardiomyocyte
differentiation J Cell Biol 153687-698
26 Monzen K Shiojima I Hiroi Y Kudoh S Oka T Takimoto E Hayashi D Hosoda T
Habara-Ohkubo A Nakaoka T Fujita T Yazaki Y Komuro I 1999 Bone
morphogenetic proteins induce cardiomyocyte differentiation through the
mitogen-activated protein kinase kinase kinase TAK1 and cardiac transcription factors
CsxNkx-25 and GATA-4 Mol Cell Biol 197096-7105
27 Iwane M Kurokawa T Sasada R Seno M Nakagawa S Igarashi K 1987 Expression
of cDNA-encoding human basic fibroblast growth factor in Escherichia coli Biochem
Biophys Res Commun 146470-477
- 87 -
28 Maeda T Kitazoe M Tada H de Llorens R Salomon DS Ueda M Yamada H Seno
M 2002 Growth inhibition of mammalian cells by eosinophil cationic protein Eur J
Biochem 269307-316
29 Tuoya Hirayama K Nagaoka T Yu D Fukuda T Tada H Yamada H Seno M 2005
Identification of cell surface marker candidates on SV-T2 cells using DNA microarray
on DLC-coated glass Biochem Biophys Res Commun Aug 19 334(1)263-8
30 Samah Abou-Sharieha Yuh Sugii Tuoya Dongwei Yu Ling Chen Heizou Tokutaka
and Masaharu Seno 2009 Identification of TM9SF2 as a candidate of the cell surface
marker common to breast carcinoma cells Clin Oncol Cancer Res (2009) 6 1-9
31 Reissmann E H Jornvall A Blokzijl O Andersson C Chang G Minchiotti MG
Persico CF Ibanez and AH Brivanlou 2001 The orphan receptor ALK7 and the
Activin receptor ALK4 mediate signaling by Nodal proteins during vertebrate
development Genes Dev 152010ndash2022
32 Gritsman K J Zhang S Cheng E Heckscher WS Talbot and AF Schier 1999
The EGF-CFC protein one-eyed pinhead is essential for nodal signaling Cell
97121ndash132
33 Yeo C and M Whitman 2001 Nodal signals to Smads through Cripto-dependent
and Cripto-independent mechanisms Mol Cell 7949ndash957
34 Yan YT JJ Liu Y Luo C E RS Haltiwanger C Abate-Shen and MM Shen
2002 Dual roles of Cripto as a ligand and coreceptor in the nodal signaling pathway
Mol Cell Biol 224439ndash4449
35 Juan Carlos Monge Duncan J Stewart Peter Cernacek Differentiation of Embryonal
Carcinoma Cells to a Neural or Cardiomyocyte Lineage Is Associated with Selective
Expression of Endothelin Receptors J Biol Chem 1995 270 15385-15390
36 J D Lickliter F M Smith J E Olsson K L Mackwell A W Boyd Embryonic stem
cells express multiple Eph-subfamily receptor tyrosine kinases Proc Natl Acad Sci U
S A 1996 93(1) 145ndash150
- 88 -
Conclusions
In this thesis the role of ECP on the cardiomyocyte differentiation was investigated in
mouse P19CL6 embryonic carcinoma cells I got conclusions as follows
1 ECP was confirmed to accelerate the cardiomyocyte differentiation of P19CL6 cells
This is the first to report the function of ECP on cardiomyocyte differentiation from
stem cells in vitro
2 Since cardiomyocyte differentiation in vivo is considered to follow mesoderm induction
the induction of Brachyury a marker gene of mesoderm was assessed ECP enhanced
Brachyury expression
3 ECP induced mesoderm differentiation by stimulating FGF signaling pathway and
enhanced subsequent cardiomyocyte differentiation in concert with DMSO in P19CL6
cells
4 ECP alone can not induce Wnt3a expression However in the presence of DMSO ECP
accelerated Wnt3a expression increased TOPFLASH luciferase activity upregulated
some co-receptor frizzleds genes expression It suggested ECP stimulation have some
positive effects on the Wntβ-catenin signaling pathway in the presence of DMSO
5 ECP activated PI3KAkt signaling pathway through upregulating phosphorylation of
Akt
6 ECP enhanced EphA1 expression Aactivation of EphA1 signaling is required for the
cardiomyocyte differentiation in the P19CL6 cells
- 89 -
Acknowledgements
Completion of this doctoral dissertation was possible with the support of several people I
would like to express my sincere gratitude to all of them First of all I am extremely
grateful to my research guide Professor Masaharu Seno in Laboratory of Bioengineering
Graduate School of Nature Science and Technology Okayama University for his valuable
guidance discussion with inordinate patience reviewing and correction of the manuscript
and consistent encouragement I received throughout the research work I consider it as a
great opportunity to do my doctoral program under his guidance and to learn from his
research expertise The successful accomplishment of this thesis is mainly attributed to his
selfless contribution and excellent guidance
I am grateful to Dr Hiroshi Murakami for his guidance in statistical analysis and discussion
with inordinate patience I am also grateful to DrTakayuki Kudoh for his support and
motivation
I am deeply grateful to Dr Akifumi Mizutani for valuable advice kind technical assistance
reviewing and correction of the manuscript continuing encouragement and motivation I
am also grateful to Dr Tomonari Kasai for his kind helpful suggestion kind assistant and
humorous words
I would like to extend my sincere gratitude to the former post doc Dr Ling Chen and
Takayuki Fukuda for their support and motivation and contribution to the experiment
I would also like to extend my gratefulness to Dr Masashi Okada Dr Shuichi Matsuda Dr
Sreeja C Sekhar Dr Ting Yan and all the members of the laboratory for their kindly
- 90 -
discussion friendliness and encouragement I am also grateful to friends in Japan for their
encouragement warmth and friendly entertainments
I owe a lot to my parents who endured with extraordinary missing encouraged and helped
me at every stage of my personal and academic life and longed to see this achievement
come true
I am very much indebted to my wife who supported me in every possible way to see the
completion of this work
The financial support of the Japanese government is gratefully acknowledged
Guoliang Jin
Okayama University
September 2012