Summer diet of European ShagsPhalacrocorax aristotelis desmarestiiin southern MallorcaSamer Al-Ismail1, Miguel McMinn2, Víctor Manuel Tuset3, Antoni Lombarte3 andJosep Antoni Alcover1*
07004 Palma, Balearic Islands, Spain;3 Institut de Ciències del Mar (CSIC), Passeig Marítim de la Barceloneta, 37–49,
08003 Barcelona, Catalonia, Spain.
AbstractAnalysis of pellets of European Shags Phalacrocorax aristotelis desmarestii collected ata non-breeding roost site in southern Mallorca identified 36 species of fish prey,belonging to 27 genera and to 16 families. This diversity is higher than in the diet ofP. a. aristotelis in the Atlantic, and higher than in the previous literature for P. a.desmarestii in the Mediterranean. European Shags in southern Mallorca foragedmainly on fishes with a mean estimated length of 11.6 cm (84.1% ranging from6.1–15.0 cm in estimated length), most being pelagic species (59.6 %). The most
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern Mallorca
important fish in numerical frequency (43.9%) and estimated biomass (37.2%) wasthe Bogue Boops boops (Sparidae). This species has not been reported in EuropeanShag diet in the Atlantic, and its importance was low in other Ph. a. desmarestiipopulations studied. The second most frequent prey was sand smelt Atherina (15%),but its contribution to biomass was low (1.4% of estimated biomass) because of itssmall size, as has been reported from other Mediterranean locations. The occurence ofScorpaenidae (10.7% by frequency, 17.4% of estimated biomass) was higher than inprevious studies of Ph. a. desmarestii. Scorpaenids have not been found in the diet ofPh. a. aristotelis. Sandeels (Ammodytidae), a key prey for Ph. a. aristotelis in theAtlantic, were very scarce in this study, as in other recent Mediterranean studies. Therelative abundance of species anatomically well protected against predation, such asscorpaenids and trachinids, and the diversity of prey probably reflects the scarcity orabsence of other preferred prey. This study reflects the opportunistic behaviour ofEuropean Shags in the Mediterranean Sea, foraging on fish with very differentecological requirements, in an environment that is poor but diverse.
Figure 1. The location of the Na Moltona study site in Mallorca. The locations of previous European ShagPhalacrocorax aristotelis desmarestii dietary studies in the Mediterranean Sea are shown upper-left: 1.Archipelago of Riou (Morat 2007; Morat et al. 2011); 2. Gulf of Trieste (Cosolo et al. 2011); 3. Losinj Archipelago(Cosolo et al. 2011); 4. Habibas Islands (Morat 2007); 5. Corsica (Guyot 1985).
SEABIRD 26 (2013): 8–2310
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern Mallorca
IntroductionThe European Shag Phalacrocorax aristotelis is distributed along the coasts of theWestern Palaearctic with three subspecies recognised: Ph. a. aristotelis, with66–73,000 pairs breeding along Atlantic coasts from the Kola Peninsula in Russiato southern Portugal (Wanless & Harris 2004); Ph. a. riggenbachi in south-westernMorocco; and Ph. a. desmarestii, which is endemic to the Mediterranean, from theIberian Peninsula to the Black Sea, with an overall breeding population estimatedat between 3,000 (Velando & Munilla 2008) and 10,000 (Muntaner & Mayol 2007)pairs. Ph. a. riggenbachi and Ph .a. desmarestii are morphologically smaller than thenominal subspecies, having a smaller crest, and a brighter yellow coloration at thebasis of the bill. However, differences between the subspecies are slight, and theirtaxonomic separation has not been evaluated genetically.
The European Shag is a coastal feeding seabird, showing a strong preference for rockycoasts and small islands with clear, shallow waters over sandy or rocky seabeds (delHoyo et al. 1992). Birds mainly feed on fish, with a few species dominating the diet,and within a foraging range of up to 20 km around their breeding and roosting sites(Wanless et al. 1991; Velando 1997). Although almost exclusively piscivorous, smallnumbers of polychaetes, cephalopods, other molluscs and small benthic crustaceanshave been reported in the diet (e.g. Barrett et al. 1990; Velando & Freire 1999;Hillersøy 2011). Prey-items taken can differ spatially, even between neighbouringcolonies (Velando & Freire 1999), or seasonally, between the breeding season and therest of the year (Lilliendahl & Solmundsson 2006), depending on availability,suggesting opportunistic foraging behaviour (Barrett 1991).
There is abundant literature on the diet of Atlantic/North European Shags. Withexception of the northernmost latitudes where gadoids are often their main prey(Barrett et al. 1990; Hillersøy 2011), different species of sandeels (Ammodytidae)are usually, but not always (Fortin et al. 2013), the main food source for Ph. a.aristotelis (Steven 1933; Lack 1945; Lumsden & Haddow 1946; Snow 1960;Pearson 1968; Harris & Wanless 1991, 1993; Velando & Freire 1999; Furness &Tasker 2000; Pennington et al. 2004; Lilliendahl & Solmundsson 2006). However,fewer dietary studies have been performed in the Mediterranean Sea, where thediet seems to be much more varied, especially during the breeding season, withsome fish families such as Atherinidae, Labridae, Sparidae or Gobiidae playing animportant role as prey items (Araujo et al. 1977; Guyot 1985; Morat 2007; Cosoloet al. 2011; Morat et al. 2011).
The Balearic Islands hold one of the most important populations of Ph. a.desmarestii (hereafter ‘Mediterranean Shag’). Censuses carried out on Menorca,Eivissa and Formentera in 2005, and on Mallorca and Cabrera in 2006, estimatedtheir populations at 1,800 breeding pairs, 95% of the Spanish population for thissubspecies, and 18% of its world population (after Muntaner & Mayol 2007); afurther census in 2006 and 2007 estimated 2,017 breeding pairs (Álvarez &Velando 2007). It has been suggested that the Balearic Islands could be a sourceregion for individuals dispersing to other areas (García et al. 2011). Despite the
11SEABIRD 26 (2013): 8–23
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern Mallorca
importance of this area, the only information on their diet in the Balearic Islandscomes from stomach contents analysis of eight specimens, with 16 individual fishidentified, and several prawns (Araujo et al. 1977). Shags regurgitate pelletscontaining fish bones, otoliths and scales, small invertebrates, marine vegetationand even small stones. Although several authors (Ainley et al. 1981; Duffy &Laurenson 1983; Johnstone et al. 1990) estimated that pellets were regurgitated atleast once a day, Russell et al. (1995) obtained a mean production of one pelletevery four days for birds in the wild. Here, we present diet data inferred from ananalysis of pellets from Mediterranean Shags collected on an islet off southernMallorca (Balearic Islands). This information contributes to our knowledge of thediet of this species in the Mediterranean Sea.
MethodsMediterranean Shags breed during the winter months and in this study pelletswere collected during the non-breeding period, in a single visit on 3 July 2009 toNa Moltona, a small islet in southern Mallorca. Pellets were produced by adult andjuvenile Shags, and the proportion belonging to each age-group was unknown. NaMoltona is used as a post-breeding roosting site by Shags from colonies on(mostly) Cabrera and Mallorca that feed in the channel between the two islands(Figure 1). They use the islet when sea conditions are not very favourable in theopen sea around Cabrera, but better close to the Mallorcan coast. We collected atotal of 60 pellets, of which at least eight were incomplete. Dislodged parts of the60 pellets were also collected and analysed to avoid information loss, and allpellets were stored frozen until analysis. Once taken out of the freezer, each pelletwas soaked in a solution of water and commercial washing powder for 24–48 h toseparate its content. Bones and otoliths were separated from the remaining mucuslayer and other soft tissues by precipitation, and rinsed through a sieve (1 mm).Remains of a few crustaceans, such as two small crabs and some legs of prawns,were also found, but they were not included in our study as they could havebelonged to fish stomach contents. After drying, the content of each pellet waslabelled and packed for subsequent identification.
Initially, two approaches were performed for the analysis of the diet: 1) identifi-cation of bones and teeth (pharyngeal, mandibular and premaxillar), and 2) identi-fication of otoliths (Figure 2). Both methods fitted well with each other for preyclassification. However, otoliths have highly diagnostic characters (Tuset et al.2008), show lower digestibility, and permit the identification of a larger number ofprey items (c. 800 specimens) than bones (c. 200 specimens). Results presentedhere are therefore derived from otolith identification only. In comparison withother sampling methods, such as analysis of stomach contents or directobservation of foraging activity, pellet analysis is a relatively easy method, allowingthe collection of a large number of samples, and involving little or no disturbanceto the birds. Nevertheless, the method is not free of biases. At least five sources oferror have been highlighted for the assessment of diet through pellet analyses(Carss et al. 1997). Thus, our work should be only considered as indicative of thediet of the Mediterranean Shag in Mallorca.
SEABIRD 26 (2013): 8–2312
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern MallorcaTable 1.P
aram
eter
s of
the
rel
atio
nshi
ps b
etw
een
Oto
lith
Leng
th (
OL)
and
Fis
h Le
ngth
(FL
) an
d be
twee
n Fi
sh L
engt
h an
d Fi
sh W
eigh
t (F
W),
esta
blis
hed
for
the
fish
spec
ies
foun
d w
ithi
n pe
llets
of
the
Euro
pean
Sha
g Ph
alac
roco
rax
aris
tote
lis d
esm
ares
tiiat
Na
Mol
tona
, Sou
ther
n M
allo
rca.
OL versus FL parameters F
L= a
+ b
*OL
FL versus FW parameters F
W =
a *
FLb
Species
ab
nR2
Source
ab
nR2
Source
Arnog
lossus im
perialis 1
NA
NA
NA
NA
Fish
base
.org
0.00
453.
1717
0.94
6Fi
shba
se.o
rg2
Atherina bo
yeri
38.8
4814
.295
50.
869
AFO
RO0.
0041
683.
1542
60.
95Le
onar
dos
& S
inis
200015
Blenniidae unidentified 3
-58.
145
77.6
578
0.88
3A
FORO
0.01
682.
9130
0.98
01M
erel
laet
al.
19972
Blennius ocella
ris
-58.
145
77.6
578
0.88
3A
FORO
0.01
682.
9130
0.98
01M
erel
laet
al.
19972
Boop
s bo
ops
35.6
4928
.111
100.
838
AFO
RO0.
0119
2.85
5422
80.
986
Mor
ey e
t al.
20034
Chrom
is chrom
is-2
7.47
228
.450
50.
978
AFO
RO0.
0189
2.93
369.
000.
935
Mor
ey e
t al.
20034
Citharus lin
guatula
-33.
120
41.5
9910
0.95
9A
FORO
0.00
33.
350
0.96
04M
erel
laet
al.
19972
Coris julis
-58.
289
76.7
857
0.97
0A
FORO
0.00
73.
0462
473
0.97
6M
orey
et a
l.20
034
Dicentrarchus labrax
-96.
743
38.8
569
0.96
0A
FORO
0.00
513.
1589
140.
992
Mor
ey e
t al.
20034
Diplodu
s annu
laris
-30.
716
35.5
2210
0.97
4A
FORO
0.01
153.
1668
848
0.97
2M
orey
et a
l.20
034
Diplodu
s pu
ntazzo
-192
.040
66.7
866
0.94
1A
FORO
0.02
62.
8188
430.
988
Mor
ey e
t al.
20034
Diplodu
s sargus
-81.
474
47.4
3211
0.82
1A
FORO
0.01
143.
1317
750.
989
Mor
ey e
t al.
20034
Diplodu
ssp.5
NA
NA
NA
NA
NA
Diplodu
s vulgaris
-82.
594
45.7
5813
0.94
0A
FORO
0.01
493.
0058
328
0.99
7M
orey
et a
l.20
034
Gob
iidae unidentified 6
NA
NA
NA
NA
NA
Gob
ius bu
cchichi 7
NA
NA
NA
NA
Fish
base
.org
0.01
13.
192
210.
996
Fish
base
.org
Gob
ius cruentatus
-16.
984
30.5
986
0.98
2A
FORO
0.00
443.
4108
490.
994
Mor
ey e
t al.
20034
Gob
ius pa
ganellu
s 7
NA
NA
NA
NA
Fish
base
.org
0.01
13.
192
210.
996
Fish
base
.org
Gym
nammod
ytes semisquam
atus
67.6
1426
.498
60.
079
AFO
RO0.
0006
3.47
623
NA
Fish
base
.org8
Labrus merula
-88.
272
88.4
737
0.88
5A
FORO
0.00
763.
1862
124
0.95
9M
orey
et a
l.20
034
Labrus viridis
-17.
920
62.7
076
0.98
4A
FORO
0.05
83.
2216
630.
97M
orey
et a
l.20
034
Lithog
nathus mormyrus
-69.
445
44.4
407
0.95
9A
FORO
0.01
023.
0327
441
0.98
6M
orey
et a
l.20
034
Merluccius merluccius
-16.
876
24.8
2616
0.98
6A
FORO
0.00
483.
055
960.
924
Mor
ey e
t al.
20034
Oblada melanura
-140
.770
48.2
196
0.92
8A
FORO
0.02
43.
567
250.
984
Fish
base
.org9
Pagellu
s erythrinus
-32.
382
33.0
489
0.92
4A
FORO
0.01
642.
8936
580.
983
Mor
ey e
t al.
20034
Parablenniusspp.10
-0.5
0347
.615
30.
590
NA
0.01
22.
769
270.
97Fi
shba
se.o
rg4
13SEABIRD 26 (2013): 8–23
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern Mallorca
Parablennius tentacularis
-0.5
0347
.615
30.
590
AFO
RO0.
012
2.76
927
0.97
Fish
base
.org4
Paralip
ophrys trigloides 11
-7.4
4059
.982
30.
897
AFO
RON
iede
r et
al.
19944
Sarpa salpa
-86.
983
50.9
156
0.95
7A
FORO
0.03
232.
7004
790.
966
Mor
ey e
t al.
20034
Scorpa
ena no
tata
2.32
818
.692
120.
964
AFO
RO0.
016
3.03
8483
0.98
8M
orey
et a
l.20
034
Scorpa
ena po
rcus
-123
.630
40.0
349
0.71
3A
FORO
0.01
833.
0202
980
0.96
9M
orey
et a
l.20
034
Scorpa
ena scrofa
-44.
830
26.6
939
0.98
2A
FORO
0.02
22.
9418
359
0.98
1M
orey
et a
l.20
034
Scorpa
enasp.12
NA
NA
NA
NA
NA
Serranus cabrilla
-25.
114
29.7
5710
0.92
7A
FORO
0.00
923.
0658
298
0.97
8M
orey
et a
l.20
034
Serranus hep
atus
-13.
121
24.5
599
0.98
3A
FORO
0.00
443.
5681
220.
928
Mor
ey e
t al.
20034
Sparus aurata
-169
.500
56.6
979
0.97
3A
FORO
0.00
533.
2393
140.
978
Mor
ey e
t al.
20034
Spicara sm
aris
36.0
9521
.966
60.
955
AFO
RO0.
0113
2.86
9652
0.98
2M
orey
et a
l.20
034
Spon
dylio
soma cantharus
-80.
069
38.8
419
0.92
5A
FORO
0.01
582.
9957
860.
994
Mor
ey e
t al.
20034
Symph
odus mediterraneus
7.97
141
.117
60.
960
AFO
RO0.
0123
3.06
5321
40.
988
Mor
ey e
t al.
20034
Symph
odusspp.13
NA
NA
NA
NA
NA
Synapturichthys kleinii
AFO
RO-5
2.51
086
.603
30.
961
0.05
82.
724
340.
889
Fish
base
.org14
Trachinu
s draco
AFO
RO-4
4.42
930
.966
80.
829
0.01
012.
8354
270.
989
Mor
ey e
t al.
20034
NA
not
avai
labl
e.
1O
nly
the
FL m
ean
valu
e w
as a
vaila
ble
(N=
17, R
2= 0
,946
).2
From
Bal
earic
Isla
nds
spec
imen
s.3
Blen
nius
oce
llaris
para
met
ers
from
AFO
RO a
nd M
erel
la e
t al.
1997
use
d.4
From
Spa
nish
Med
iter
rane
an c
oast
spe
cim
ens.
5FW
cal
cula
ted
as t
he m
ean
wei
ght
of a
ll th
e D
iplo
dus
sp. c
lass
ified
to
spec
ies
leve
l.6
FW c
alcu
late
d as
the
mea
n w
eigh
t of
all
the
Gob
ius
sp. c
lass
ified
to
spec
ies
leve
l.7
FL a
s m
ean
valu
e of
FL
forG
. pag
anel
lus
in M
ar M
enor
(Fi
shba
se.o
rg).
8Fr
om U
K sp
ecim
ens.
9Fr
om T
urki
sh (
Gök
cead
a Is
land
) sp
ecim
ens.
10Pa
rabl
enni
us te
ntac
ular
ispa
ram
eter
s fr
om F
ishb
ase.
org
used
for
FL.
11M
ean
wei
ght
from
Nie
der
et a
l. 19
94 u
sed.
12FW
cal
cula
ted
as t
he m
ean
wei
ght
of a
ll th
e Sc
orpa
ena
sp. c
lass
ified
at
spec
ies
leve
l.13
FW c
alcu
late
d as
the
mea
n w
eigh
t of
S. m
edite
rran
eus.
14Fr
om C
roat
ian
spec
imen
s.15
From
Gre
ek s
peci
men
s.
SEABIRD 26 (2013): 8–2314
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern Mallorca
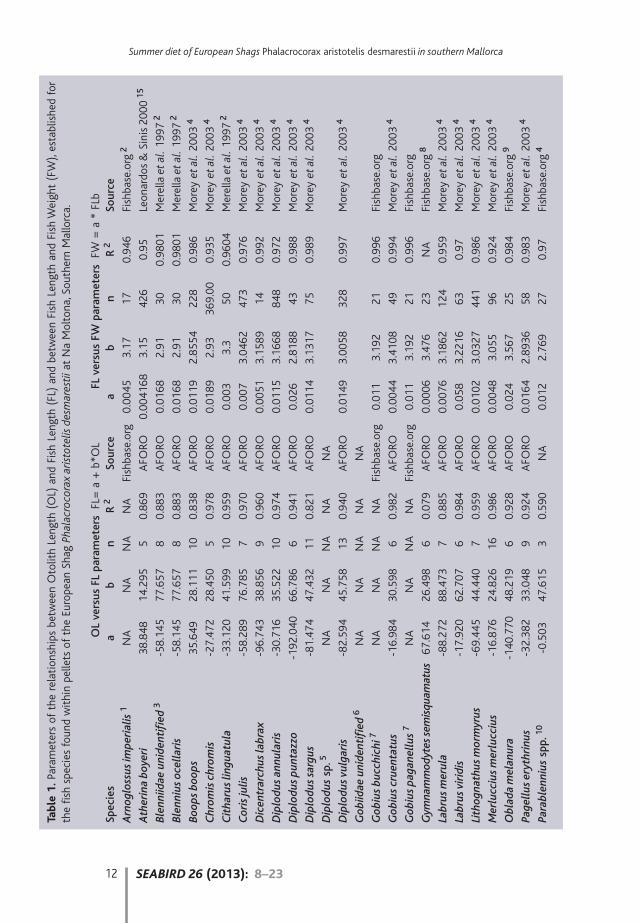
Otoliths were identified to species or genus level at the Institute of MarineSciences (ICM) in Barcelona, distinguishing their right or left sides. The mostnumerous side was used to calculate the minimum number of individuals of eachfish species in each pellet. Otoliths were measured with the software ImageJ on theimages obtained using a Marlin® b/w camera, adapted to a stereomicroscopeLeica® MZ160. Regression equations were used to estimate fish length fromotolith length with the AFORO database (http://www.cmima.csic.es/aforo/;Lombarte et al. 2006) for the species available on it (Table 1, left) whilstrelationships between fish estimated length and fish estimated weight wereobtained from the literature, using almost exclusively studies of Mediterranean fishpopulations (Table 1, right). When otoliths were only identified to genus or familylevel, we used the average weight of all the preyed specimens of the genus (orfamily) that were identified to species. An arbitrary size was applied to Gobiinae(length 5.6 cm, mass 2.7 g, based on average size for Gobius paganellus from MarMenor, Murcia, Spain; Fishbase.org), when adequate otolith/size relationships werenot available.
In order to address the ecological significance of Shag feeding behaviour, preyitems were classified according to their habitat preferences in three categories(benthic, demersal and pelagic) and seven arbitrary size classes.
Figure 2. Left sagitta otoliths obtained from European Shag Phalacrocorax aristotelis desmarestii pellets at NaMoltona, Mallorca. A. Atherina sp. B. Scorpaena porcus. C. Labrus merula. D. Boops boops. E. Diplodus vulgaris. F.Gobius bucchichi. Scalebars = 1 mm.
15SEABIRD 26 (2013): 8–23
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern Mallorca
ResultsA total of 808 otoliths were obtained, of which 68 were too eroded to beidentified. Of the remaining 740, 730 were identified to species and 10 to genuslevel (Table 2). A total of 36 different fish species were identified, belonging to 27different genera and to 16 families. The most numerous fish species was the BogueBoops boops (43.9%) followed by sand-smelts (Atherina) (15.0%), Scorpaenaporcus (6.1%), Diplodus sargus (4.5%), S. notata (3.5%), Serranus hepatus (3.0%),Lithognathus mormynus (3.0%), D. vulgaris (2.8%), Coris julis (2.0%), Spicara smaris(1.9%), Trachinus draco (1.8%), and D. annularis (1.5%). The remaining 24 individualspecies accounted each for < 1% of diet composition.
The most common genera were Boops (43.9%), followed by Atherina (15.0%),Scorpaena (10.7%), Diplodus (10.1%), Serranus (3.1%), Lithognathus (3.0%),Coris (2.0%), Spicara (1.9%), Trachinus (1.8%) and Gobius (1.2%). The mostrepresented families were Sparidae (58.9%), followed by Atherinidae (15.0%) andScorpaenidae (10.7%).
In terms of estimated biomass, 64.7% of all prey belonged to Sparidae.Scorpaenidae was also well represented, comprising 17.4% of the estimatedtotal mass of the sample studied. Other, less represented families were Labridae(9.8%), Trachinidae (2.3%), Atherinidae (1.4%), and Centracanthidae (1.3%). Allremaining families each accounted for < 1% of estimated total diet mass. At alower taxonomic level, five species belonging to the three most importantfamilies in terms of estimated biomass represented up to 75% of total diet mass:Bogue (37.2%), Scorpaena porcus (12.7%), Diplodus sargus (12.6%), Labrusmerula (6.3%), and Lithognathus mormyrus (6.0%).
Table 2. Number of otoliths (individuals), frequency, estimated biomass (g), and percentage in estimatedbiomass of the fish species found in pellets of European Shag Phalacrocorax aristotelis desmarestii at NaMoltona, Southern Mallorca.
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern Mallorca
Fish species recorded inthe diet were mostlypelagic (59.6% of thetotal), followed bydemersal (22.6%) andbenthic (17.8%)species, with 84.1% ofall prey being 6.1–15cm in estimated length(Figure 3). The mostfrequent estimated sizefor benthic anddemersal prey was9.1–12 cm. Theestimated size distri-bution of pelagic preywas bimodal, withhigher frequencies at6.1–9 cm and 12.1–15cm being due to therespective prevalence ofAtherina spp. and Boguein the diet, each with rather narrow size ranges. Shags at Na Moltona fed on fishof medium size, with only 3.7% of prey items estimated to be < 6.1 cm in length,and only 4.1% to be 18.1 cm or more (mean estimated length = 11.6 cm).
DiscussionThis study highlights the great variety of fish (36 different species belonging to 27genera and 16 families) present in the summer diet of Mediterranean Shags inMallorca, which reflects the fish diversity of the Balearic coast and confirms thegeneralist diet of these birds. Although more work is required to improve knowledgeof the spatio-temporal dynamics of diet choice in this species in Mallorca, the resultsobtained considerably enlarge available ecological information.
Our results are in agreement with other recent studies, showing a higherdiversity of prey caught by Mediterranean Shags (Table 3), compared with thatcaught by P. a. aristotelis on Atlantic coasts. At most Atlantic localities, sandeelis the dominant prey item (Barrett et al. 1986; Harris & Wanless 1991, 1993;Grémillet et al. 1998; Velando & Freire 1999; Furness & Tasker 2000; Lilliendahl& Solmundsson 2006), especially during chick-rearing (Harris & Wanless 1993),when it can represent 100% of the chick diet (Barrett et al. 1986). Further, thediversity of taxa found in pellets from Na Moltona was higher than in all otherstudies conducted in other localities in the Mediterranean Sea (Table 3), despitethe fact that our sampling was limited to a single day.
0.0
5.0
10.0
15.0
20.0
25.0
30.0
35.0
40.0
45.0
0–3 3.1–6 6.1–9 9.1–12 12.1–15 15.1–18 >18.1
% o
f ec
olo
gica
l cat
ego
ry w
ithi
n ea
ch s
ize
clas
s
Size classes (cm)
PelagicDemersalBenthic
Figure 3. Estimated size classes of fish preyed by European ShagsPhalacrocorax aristotelis desmarestii roosting at Na Moltona according thethree ecological categories considered.
Unlike European Atlantic Coasts, the Balearic Sea ischaracterised by a low productivity (UNEP 2008;Cognetti et al. 2001). The islet of Na Moltona isformed by sandstone substrate, and is separatedfrom the Mallorcan coast by a 5–6 m deep channelwith a sandy bottom colonised extensively by theseagrass Posidonia oceanica. Close to Na Moltonathere are also submerged sandstone blocks andremains of fossil dunes. In this habitat, there is not aclear dominance of one fish species as is the case inAtlantic waters where sandy substrates have highdensities of sandeels.
In our study, the most important fish, both infrequency of occurrence and estimated biomass, wasthe Bogue (43.9% and 37.2% respectively). Thissparid is a common pelagic inhabitant of coastalwaters of the Eastern Atlantic and MediterraneanSea, which can be found in shoals over rock, sand ormud substrates (Froese & Pauly 2011). In Balearicwaters, the species can be found over seagrassmeadows as well as over rocky and sandy bottoms atdepths of 2–30 m (Fischer et al. 2007). This specieshas not yet been found in the diet of P. a. aristotelisin the Atlantic. For Mediterranean Shag, it has onlybeen found in low frequency (0.2–2.8%) at RiouArchipelago (Morat 2007; Morat et al. 2011).
The second most frequent species found at NaMoltona (15% of otoliths) were sand smeltsAtherina spp. However, atherinids were irrelevant interms of biomass (1.4% of estimated biomass) dueto the small size of the captured specimens. Thesame was found in the Adriatic Sea during the non-breeding season, where atherinids represented10.7% of Shag prey but only 1.4% of estimatedbiomass; their frequency in the diet was higher onRiou (11.5–36.8%), and the Habibas Islands (28.5%).
Scorpaenids were less numerous (10.7%), but werethe second most important prey in terms ofestimated biomass (17.4%) after Bogue, and thecontribution to the diet of Shags of the three speciesidentified at Na Moltona was high compared withother locations. In Croatia, they were only found inMarch in the breeding season (one Scorpaena scrofa
SEABIRD 26 (2013): 8–2318
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern MallorcaTable 3.C
ompa
rison
of s
ampl
ing
perio
d, n
umbe
r of r
ecov
ered
oto
liths
, and
the
num
ber o
f fis
h fa
mili
es a
nd s
peci
es re
cord
ed in
die
tary
stu
dies
of t
he E
urop
ean
Shag
Pha
lacr
ocor
ax a
risto
telis
des
mar
estii
in t
he M
edit
erra
nean
Sea
.1Ra
nge
of m
ean
leng
ths
of t
he d
iffer
ent
spec
ies
cons
umed
.
Locality
Period
Number
Number of
Number
Mean
Range of
Author
of otoliths
families
of species
length (cm)lengths (cm)
Riou
Arc
hipe
lago
, Fra
nce
Apr
il–Se
ptem
ber
2004
–07
2,46
212
257.
4–14
.01
5.1–
21.9
Mor
at 2
007;
Mor
at e
t al.
2011
Hab
ibas
Isla
nds,
Alg
eria
.Ju
ly 2
005
284
612
Mor
at 2
007
Gul
f of
Trie
ste,
Ital
y.M
ay–O
ctob
er 2
005
20,7
1617
317.
61.
5–29
.0Co
solo
et a
l. 20
11
Losi
nj A
rchi
pela
go, C
roat
ia.
Janu
ary–
Apr
il 20
063,
272
1526
7.5
1.6–
25.1
Coso
lo e
t al.
2011
Na Moltona, Mallorca.
3 June 2009
808
1636
11.6
1.6–31.9
This study
19SEABIRD 26 (2013): 8–23
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern Mallorca
only, frequency 0.1%, estimated biomass 0.6%), while on Riou two species (S.notata and S. porcus) comprised 3.8% of prey items in the non-breeding season.Although most studies on the diet of the European Shag have been performed inthe Atlantic (e.g., Steven 1933; Lumsden & Haddow 1946; Furness & Barrett 1985;Barrett et al. 1990; Wanless et al. 1991; Harris & Wanless 1993; Álvarez 1998;Lilliendahl & Solmundsson 2006; Hillersøy 2011; Fortin et al. 2013), scorpaenidshave not been reported in the diet of P. a. aristotelis.
Other sparids, such as Diplodus sargus and Lithognathus mormyrus, and labrids,such as Labrus merula, were also important in terms of biomass (6–13% ofestimated biomass), and were extremely scarce or have not been reported in thediet of Atlantic Shags. Other closely related species such as Labrus bimaculatus andL. bergylta, Crenilabrus melops and Ctenolabrus rupestris were present in somestudies (Álvarez 1998; Hillersøy 2011), but, with the exception of C. melops in theCantabric sea, generally at low frequency. In the literature available from theMediterranean Sea, D. sargus and L. mormyrus have also been reported, but theircontribution to the diet was limited (Morat et al. 2011; Cosolo et al. 2011), and L.merula was not found.
Only two sandeel otoliths, representing 0.3% of prey items, were found in ourstudy. On Riou, sandeels represented only < 1.5% of prey items, and were notfound in the diet of Shags in the Adriatic Sea, either in the breeding or non-breeding seasons. In great contrast, sandeels are an essential food resource for thegrowth of many young seabird species in the Atlantic (Pearson 1968), especially forEuropean Shags (Harris & Wanless 1991; Velando & Freire 1999).
The ecological categories of fish prey at Na Moltona were in similar proportion tothose reported from the Archipelago of Riou (64.6% pelagic, 29.8% demersal, 5.6%benthic; Morat 2007), with differences in the last two categories being partly dueto different criteria used for classification as demersal or benthic for some species.Nevertheless, at other localities the diet of Mediterranean Shag can includedifferent proportions of the ecological categories used here, with demersal gobidscomprising 81.5% of prey in the northern Adriatic during the non-breeding season(Cosolo et al. 2011). This all further suggests opportunistic foraging behaviour ofMediterranean Shags.
The size distribution of the prey was similar to that found in other Mediterraneanstudies (Table 3), with Shags at Na Moltona feeding largely on medium-sized fish(84.1% of prey estimated at 6.1–15 cm in length, mean 11.6 cm).
The fish families preyed on by Mediterranean Shags roosting at Na Moltonainclude species with different ecological requirements. Gobies are strictlydemersal and poor swimmers (Louisy 2006), characteristic of sandy or rockybottoms (Fisher et al. 2007). Sparids are also demersal, but less linked tosubstrate, i.e., with species characteristic of different substrates (rock, sand orseagrasses), and with a few species, such as Bogue, more characteristically linked
to the water column (Fisher et al. 2007). Labrids are also typically demersal, anddo not occur in pelagic waters (Fisher et al. 2007). On the other hand, atherinidscan aggregate in schools lying at different depths in the water column (Riedl1991; Louisy 2006), and others such as the pomacentrid Chromis chromis aretypically pelagic in coastal areas (Fisher et al. 2007). This wide prey range reflectsthe opportunistic feeding behaviour of the European Shag, as suggested for Ph.a. aristotelis (Barrett et al. 1990; Velando & Freire 1999), and for MediterraneanShag (Morat et al. 2011). The relatively high abundance of species armed withhard scales and venomous spines (Scorpaenidae and Trachinidae), both wellcamouflaged on the sea bottom, suggests a scarcity of other more palatable preyfor Mediterranean Shags roosting at Na Moltona. The higher prey diversity foundin their pellets, compared with other locations, reinforces this hypothesis.
SEABIRD 26 (2013): 8–2320
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern Mallorca
Summer diet of European Shags Phalacrocorax aristotelis desmarestii in southern Mallorca
AcknowledgementsWe would like to express our gratitude to Dr Daniel Oro, Dr Alejandro Martínez-Abraín and Dr Damià Jaume for valuable comments, and to Silvia Pérez and ItzíarÁlvarez for their help and useful technical recommendations. This paper isincluded as a colateral research in the DGICYT Research ProjectsCGL2012–38089 and MINECO Research Project CTM2010–19701. Dr. Victor M.Tuset has a contract JAE-DOC (CSIC) co-funded by the European SocialFoundation. Finally, we thank Dr David Grémillet and an anonymous reviewer fortheir useful comments and improvements, and Martin Heubeck and Andy Webbfor their editorial improvements.
ReferencesAFORO Project (Shape Analysis of Fish Otoliths). Centre Mediterrani d’Investigacions
Marines i Ambientals CMIMA-CSIC (Barcelona), http://www.cmima.csic.es/aforo/index.jsp
Ainley, D. G., Anderson, D. W. & Kelly, P. R. 1981. Feeding ecology of marine Cormorants
in southwestern North America. Condor 83: 120–131.
Álvarez, D. 1998. The diet of Shags Phalacrocorax aristotelis in the Cantabrian Sea, northern
Spain, during the breeding season. Seabird 20: 22–30.
Álvarez, D. & Velando, A. 2007. El Cormorán moñudo en España y Gibraltar. Población en
2006–2007 y método de censo. SEO/BirdLife, Madrid.
Araujo J., Muñoz-Cobo, J. & Purroy, F. J. 1977. Las rapaces y aves marinas del archipiélago
de Cabrera. Naturalia Hispanica 12: 1–94.
Barrett, R. T. 1991. Shags (Phalacrocorax aristotelis L.) as potential samplers of juvenile