Page 1
www.sciencemag.org/content/348/6230/120/suppl/DC1
Supplementary Materials for
Suppression of endogenous gene silencing by bidirectional cytoplasmic
RNA decay in Arabidopsis
Xinyan Zhang, Ying Zhu, Xiaodan Liu, Xinyu Hong, Yang Xu, Ping Zhu, Yang Shen,
Huihui Wu, Yusi Ji, Xing Wen, Chen Zhang, Qiong Zhao, Yichuan Wang, Jian Lu,
Hongwei Guo*
*Corresponding author. E-mail: [email protected]
Published 3 April 2015, Science 348, 120 (2015)
DOI: 10.1126/science.aaa2618
This PDF file includes:
Materials and Methods
Figs. S1 to S19
Captions for Tables S1 to S4
Full Reference List
Other Supplementary Material for this manuscript includes the following:
(available at www.sciencemag.org/content/348/6230/120/suppl/DC1)
Tables S1 to S4 (Excel file)
Page 2
2
Materials and Methods
Plant Materials and Growth Conditions
Commercially-available Murashige and Skoog (MS) medium and nitrogen-depleted
MS salt (PhytoTechnology Laboratories; Catalog: M524, M531) were used in preparing
the full-nutrition MS medium and nitrogen-depleted medium (pH 5.7-5.8, 1% sucrose, 10
g/L agar), respectively. Seeds were surface-sterilized and plated on the medium. Seeds
pretreated with stratification for 2-4 days at 4°C were kept in the greenhouse for another
5-7 days (22°C, 16 h/8 h photoperiod) before transferring the seedlings to the soil or
phenotyping. Ethylene response assays and phenotypic analysis were performed as
described previously (26, 27).
Genetic Screen and Map-Based Cloning
Map-based cloning for each mutant was done with publicly available markers (28).
EMS-mutagenized M2 seeds in EIN3ox background were pooled in 50 families and 3000
M2 seeds in each family were screened on MS medium supplemented with 10 μM ACC.
Large-cotyledon seedlings were selected as suppressor mutants and phenotypically
confirmed in the next generation. The suppressor mutants with stable large-cotyledon
phenotypes were designated according to the original family number; for example, s28
was a suppressor mutant from the pool 28.
To identify the causal genes associated with the EIN3ox suppressor phenotypes in
s28, s37 and s40 mutants, map-based cloning strategy was used. Genomic DNA was
extracted using the simplified CTAB method described previously (29). Rough mapping
was performed using about 50 large-cotyledon seedlings selected from the F2 mapping
populations that phenocopied each mutant on Murashige and Skoog medium
supplemented with ACC. The s28 mutation was mapped to an interval around T6H20 on
chromosome 3 (17123 kb, 2 recombinants/758; 17294 kb, 0 recombinants/758; 17312 kb,
2 recombinants/758). We sequenced the coding sequences of 22 genes (At3g46960-
At3g47160) and found one point mutation in At3g46960 as described in the main text.
Page 3
3
The s37 mutant was mapped ~0.5 cM on the upper side of a marker (28837 kb) on
chromosome 1. However, no recombinants were found in 468 individual plants within the
large interval further on the upper side (27944-28724 kb), because the T-DNA insertion
of EIN3ox resided at 27885 kb on chromosome 1, and we found the large-cotyledon
phenotype manifested by s37 and s40 was dependent on the existence of two copies of
the EIN3 transgene. We sequenced the candidate AtSKI3 gene (AT1G76630) in both s37
and s40, and found two distinct nonsense mutations described in the main text (Fig. 1D).
Genetic Analysis and Genotyping
Mutants and transgenic materials used in this study were either maintained in our
laboratory or purchased from ABRC. The embryo defects were analyzed by examining
both the ratio of aborted seeds within heterozygous siliques and the segregation ratios in
the filial generation of the heterozygous individuals (30). The T-DNA insertional Salkline
mutants ski2-2 (Salk_129982) and ski2-3 (Salk_063541) were obtained from ABRC and
verified by PCR amplification (31). The EIN3ox transgene were genotyped by PCR, and
the point mutations such as rdr6-11 and ago1-45 were genotyped as described previously
(19, 32).
Homozygous double and triple mutants were generated by genetic crosses and were
identified from the F2 or F3 population. Each mutation was confirmed by PCR-based
genotyping and phenotypic analysis or by using antibiotic-resistant markers. To generate
the ein5-1 ski2-2 dcl4-2 dcl2-1 and ein5-1 ski2-3 dcl4-2 dcl2-1 quadruple mutants, we
genotyped the F2 and F3 plants propagated from the ein5-1 ski2-2 (ski2-3) hemizygote
and dcl4-2 dcl2-1 cross. In the filial generation of ein5-1 (+/-) ski2-2 (+/-) dcl4-2 (+/-)
dcl2-1 (+/-), no ein5-1 ski2-2 double mutant, ein5-1 ski2-2 dcl2-1 or ein5-1 ski2-2 dcl4-2
triple mutants were verified, while the combinations of other genotypes were viable.
Similarly, no ein5-1 ski2-3 dcl4-2 plant was verified from the segregating population
derived from the ein5-1 ski2-3 hemizygote and dcl4-2 dcl2-1 cross. In these experiments,
the ski2-2, ski2-3, dcl4-2, dcl2-1 loci were genotyped by PCR, and the ein5-1 mutation (1
bp deletion, frame shift) was identified by ethylene-related phenotyping (27) and
confirmed by Sanger sequencing.
Page 4
4
Gene Expression Analysis by RT-PCR
Total RNA extracted with the TRIzol reagent (Invitrogen) was used for the reverse
transcription reaction (M-MLV reverse transcription system, Promega) after Dnase I
(Promega) digestion. Real-time PCR was performed using SYBR GreenMix (Takara)
with the UBQ10 gene as a reference on the Roche realtime PCR platform
(LightCycler®480). The gene-specific primers used in real-time PCR were designed
using the QuantPrime tool (33) and listed in Supplement Table S4 online.
Transformation Vectors and Construction of Transgenic Plants
To generate the 35S:GFP-AtSKI2 transgenic lines, the full-length AtSKI2 coding
region was amplified and constructed into the pEGAD binary vector in-frame with an
upstream GFP. The primers for making these constructs are listed in Supplement Table
S4 online. The floral-dipping transformation method was performed using Agrobacterium
tumefaciens strain GV3101. Transgenic plants were selected on MS medium
supplemented with 20 μg/L glufosinate and then transferred to soil. Homozygous lines
were used in this study.
Microscopy
GFP fluorescence was acquired using a confocal laser microscope (LSM 710; Carl
Zeiss) at an excitation wavelength of 488 nm. Live seedlings were stained for 15 min in
DAPI solution (1 μg/mL in water) before mounting and inspection.
In Vitro Pull-Down Assay
GST pull-down assays were performed following the procedure described
previously (34). The coding sequences corresponding to the 344 amino-acids of the SKI2
amino terminus (SKI2N) and 240 amino-acids of the SKI3 carboxyl terminus (SKI3C)
were constructed in frame into the pGEX-6p-1 (GE Healthcare) and pCold-TF (Takara
Bio) vectors, respectively. The GST-tagged SKI2N and His-tagged SKI3C proteins were
expressed in the E.coli BL21 (DE3) strain with isopropylbeta-D-thiogalactopyranoside-
induction at 0.1 mM and 0.5 mM, respectively, and purified using glutathione Sepharose
Page 5
5
4B (GE Healthcare) and Ni-NTA Agarose (Qiagen) following the manufacturer’s
instructions.
Immunoprecipitation Assay and Mass Spectrometry
Tissues (6-day-old light-grown seedlings of homozygous 35S:GFP-SKI2 transgenic
plants) were ground into a fine powder in liquid nitrogen. Two grams of powder was
homogenized in 4 mL lysis buffer [10 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.5 mM
EDTA, 0.5% NP-40, 1× complete protease inhibitor cocktail (Roche)] and centrifuged at
12,000 g for 10 min after 30 min incubation at 4°C. GFP-Trap_A (50 μL; Chromotek)
beads were added into the supernatant. The following steps of immunoprecipitation were
carried out according to the manufacturer’s instructions. Precipitates were eluted by
adding 50 μL 4×LDS sampling buffer (Invitrogen) with 50 mM DTT (Invitrogen) and
heated at 65°C for 10 min followed by SDS-PAGE separation. Silver staining and mass
spectrometry analysis were performed as described previously (35, 36). Silver-stained gel
pieces were excised and then digested following the procedure described previously (37).
Peptides were further extracted from the trypsin-digested gel pieces and separated with
the EASYnLCIITM
integrated nano-HPLC system (Proxeon, Denmark) and the eluate was
used in the Thermo LTQ-Orbitrap velos mass spectrometer as described previously (36).
MS/MS analysis was performed with a spray voltage of 2.8 kV. The range of the MS
scan was 350-20000 m/z.
RNA-seq and Small RNA-seq
Both total RNA (for mRNA-seq) and small RNA were extracted from the shoot
tissues of 16-day-old plants. For mRNA-seq, total RNA was extracted using the RNeasy
Plant Mini Kit (QIAGEN). Enriched mRNA using the Epicentre Ribo-Zero™ rRNA
Removal Kits (Plant Leaf) from 10 μg total RNA was sheared before random-primed
cDNA synthesis and amplification using the TruSeq mRNA preparation kit (Illumina)
following the standard protocol. The size-selected 100-bp single-ended libraries were
sequenced with Illumina HiSeq 2000. Deep sequencing data were generated on the
Illumina Hiseq 2000 platform and filtered reads were aligned to the Col-0 transcriptome
(main gene models, TAIR 9.0 release) using the BWA program (38) with a seed length of
Page 6
6
32 and up to 4% mismatches and 1 gap allowed. The clustering was run with the log2
ratios of 15663 genes using Cluster 3.0 software (39) and visualized with the Java
Treeview program (40). The small RNAs (< 200 nt) were isolated using the miRNeasy
Mini Kit (Qiagen) and then used in constructing the sequence library. The small RNA seq
was performed with Illumina HiSeq 2500. The 21-22 nt clean reads after adaptor-removal
and filtering were mapped to the genome (Tair 10.0) using the Bowtie program (41).
Reads mapped to each gene model were counted, then followed by normalization and
differential expression analysis as described previously (IHGSC, 2004). Manual
correction was done for the reads mapped to miRNA target genes over which some
miRNA reads piled up at the miRNA target sites. Visualization of the mapped reads was
done using the Integrated Genome Browser, version 6.7.2 (42). Geneset enrichment
analysis was performed using an online webserver PlantGSEA (43) and the KOBAS 2.0
program (44). The primers for validation of the RNA seq results are listed in Supplement
Table S4 online.
Small RNA Northern Blotting
Small RNA was extracted through TRIzol-isolation method followed by a PEG-NaCl
enrichment [0.5 M NaCl, 10% polyethylene glycol 8000 (w/v)] and precipitated by
isopropanol. For each sample, 10 μg small RNA was separated in a denaturing 17%
polyacrylamide gel and then transferred to a neutral nylon membrane (Hybond-NX, GE
Healthcare) and immobilized by EDC fixation method as previously described (45). We
used the PerfectHyb™ plus Hybridization Buffer (Sigma) for small RNA hybridization.
End-labled DNA oligo probes were used in the detection of U6, and in vitro transcribed
RNA probes were used in the detection of siRNAs except for the EIN3-siRNAs with
DNA probes from random primed labeling.
Page 7
7
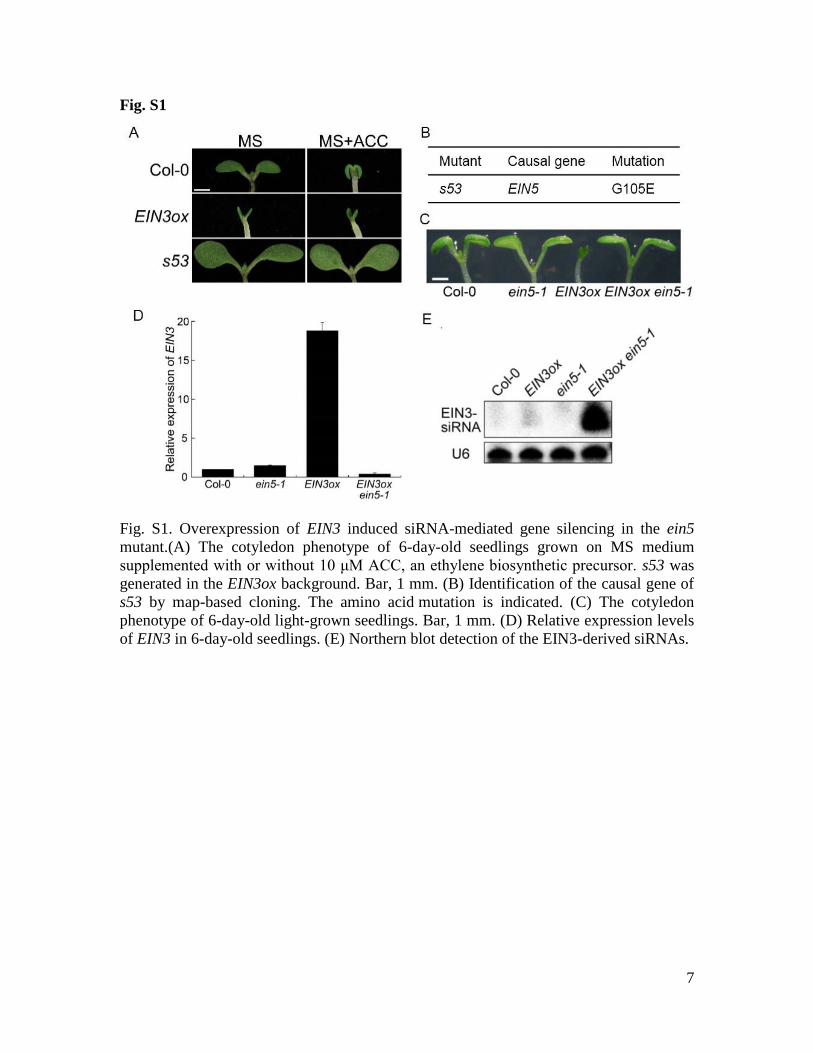
Fig. S1
Fig. S1. Overexpression of EIN3 induced siRNA-mediated gene silencing in the ein5
mutant.(A) The cotyledon phenotype of 6-day-old seedlings grown on MS medium
supplemented with or without 10 μM ACC, an ethylene biosynthetic precursor. s53 was
generated in the EIN3ox background. Bar, 1 mm. (B) Identification of the causal gene of
s53 by map-based cloning. The amino acid mutation is indicated. (C) The cotyledon
phenotype of 6-day-old light-grown seedlings. Bar, 1 mm. (D) Relative expression levels
of EIN3 in 6-day-old seedlings. (E) Northern blot detection of the EIN3-derived siRNAs.
Page 8
8
Fig. S2
Fig. S2. Identification of the SKI complex in Arabidopsis by map-based cloning. (A)
Positional cloning of AtSKI2 and AtSKI3. Mutations of s28 (ski2-1), two T-DNA
insertional mutants (ski2-2 and ski2-3), and s37, s40 (ski3) are indicated. The ski2-1
mutation resulted in a T389I amino-acid substitution in the well-defined motif-I in the
AtSKI2 protein, which is essential for its helicase activity. The two ski3 mutants resulted
in a non-sense mutation at Q20 and W158, respectively. (B) Schematic diagram of the
SKI complex (SKI2/3/8), SKI7 protein and the exosome engaged in 3’-5’ RNA decay in
the cytoplasm.
Page 9
9
Fig. S3
Fig. S3.Transgenic expression of GFP-AtSKI2 complements the mutant phenotype of
s28. (A) The cotyledon phenotype of 6-day-old seedlings. Bar, 1 mm. Three independent
transgenic lines were shown. (B) Relative expression levels of EIN3 in 6-day-old
seedlings.
Page 10
10
Fig. S4
Fig. S4. Verification of two T-DNA insertional ski2 mutants. (A) PCR-verification of the
T-DNA insertions in ski2-2 and ski2-3. The ACT2 gene was used as an internal control.
LB was a T-DNA specific primer while LP and RP were designed according to the
genomic sequences. LB+RP was used to detect the T-DNA insertions while LP+RP was
used to amplify the genomic DNA fragments that harbor the T-DNA insertions. (B)
Relative expression levels of AtSKI2 in 16-day-old plants indicating that ski2-2 failed to
express AtSKI2 RNA while ski2-3 had reduced expression, so ski2-3 seemed a weak
allele while ski2-2 could be a null allele.
Page 11
11
Fig. S5
Fig. S5. The insertional ski2 mutants show normal ethylene response. (A) The phenotype
of 3-day-old etiolated seedlings treated with or without 10 μM ACC. Bar, 2 mm. (B-C)
Quantification of the hypocotyl (B) and root (C) lengths of 3-day-old etiolated seedlings.
Mean ±SD, n > 20.
Page 12
12
Fig. S6
Fig. S6. Immunoprecipitation-mass spectrometry detection of proteins associated with
GFP-AtSKI2. (A)Silver staining of the proteins that immunoprecipitated with GFP
antibody. Key proteins in the SKI complex or associated with GFP-AtSKI2 protein
detected by mass spectrometry are indicated. (B) Peptide counts of the SKI complex
proteins detected by mass spectrometry.
Page 13
13
Fig. S7
Fig. S7. Overexpression of APT1 in the ski2 mutant phenocopies the apt1 loss-of-
function mutant. The phenotype of five-day-old light-grown seedlings treated with or
without 200 μM kinetin (KT, an active form of plant hormone cytokinin), which caused
marked growth and greening suppression. It is noted that overexpression of APT1 in
wild-type background (Col-0) results in a hypersensitive response to toxic dose of KT
manifested by retarded germination. In contrast, overexpression of APT1 in ski2-2
background results in KT hyposensitivity manifested by enhanced growth and green
cotyledons, reminiscent of apt1 mutant. Bar, 2 mm.
Page 14
14
Fig. S8
Fig. S8. Rescue of the embryo-lethality of ein5-1 ski2-2 by the rdr6 mutation. (A) The
phenotypes of immature siliques. Colorless seeds segregated in the siliques of ein5-1-/-
ski2-2+/- hemizygous parents are indicated by arrows. About one quarter (80/354, 2 =
0.666, p > 0.41) of seeds aborted in the ein5-1-/- ski2-2+/- siliques, and the filial
generation conformed to a 1:2 ratio between wild type and ski2-2 heterozygous plants
(25:45; 2 = 0.179, p > 0.67), indicating a recessive lethal segregation of the embryos
homozygous ein5-1 ski2-2 (see supplemental Methods). The ein5-1 ski2-2 rdr6-11 line
was fertile and was maintained homozygously. Bar, 0.5 mm. (B) Rosette morphology of
18-day-old plants of indicated genotypes. Bar, 1 cm.
Page 15
15
Fig. S9
Fig. S9. Restoration of the disturbed transcriptome of ein5-1 ski2-3 by the rdr6 mutation.
(A) Venn diagram of differentially-expressed genes (2-fold cutoff, RPKM>3) among
indicated comparisons identifies 596 genes that are defined as EIN5/AtSKI2-coregulated
genes. C: Col-0; e: ein5-1, s: ski2-3; es: ein5-1 ski2-3; res: rdr6-11 ein5-1 ski2-3. (B)
Comparisons of the expression levels of EIN5/AtSKI2-coregulated genes (111 up-
regulated and 485 down-regulated in ein5-1 ski2-3 ) between ein5-1 ski2-3 (es) and rdr6-
11 ein5-1 ski2-3 (res) (2-fold cutoff).
Page 16
16
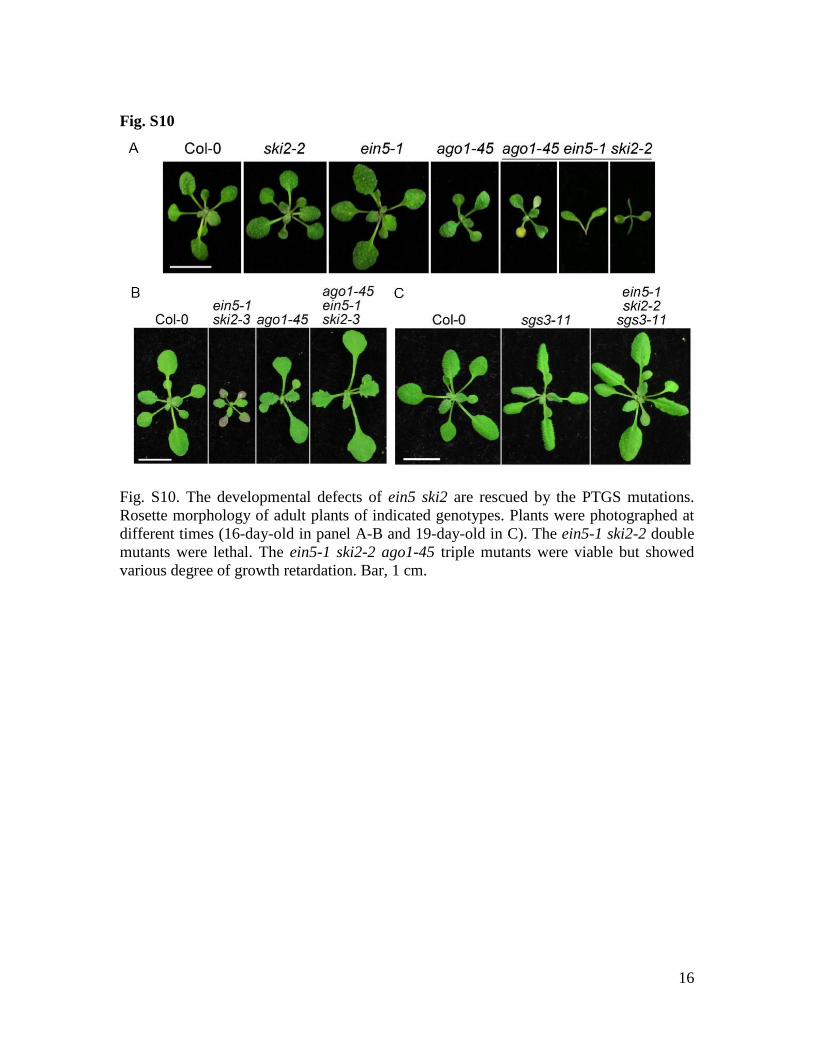
Fig. S10
Fig. S10. The developmental defects of ein5 ski2 are rescued by the PTGS mutations.
Rosette morphology of adult plants of indicated genotypes. Plants were photographed at
different times (16-day-old in panel A-B and 19-day-old in C). The ein5-1 ski2-2 double
mutants were lethal. The ein5-1 ski2-2 ago1-45 triple mutants were viable but showed
various degree of growth retardation. Bar, 1 cm.
Page 17
17
Fig. S11
Fig. S11. Restoration of select gene expression in ein5-1 ski2-3 by ago1-45.(A) Relative
expression levels of anthocyanin biosynthesis genes in 16-day-old plants.(B) Relative
expression levels of ARF6/8 and HD-ZIPIII family genes in 16-day-old plants.(C)
Relative expression levels of NIA1/2 and nitrogen-responsive genes in 16-day-old plants.
Page 18
18
Fig. S12
Fig. S12. Statistics of small RNA profiling. (A) Statistics of the small RNA sequencing
data and detected ct-siRNA reads. (B-C) tasiRNA reads detected in small RNA
sequencing (B) and normalized tasiRNA abundance in each genotype (C).
Page 19
19
Fig. S13
Fig. S13. Relative expression levels of TAS4 in 16-day-old plants.
Page 20
20
Fig. S14
Fig. S14. Northern blot detection of small RNAs derived from ARF6 and PHB in 16-day-
old plants.
Page 21
21
Fig. S15
Fig. S15. Distribution of ct-siRNA biogenesis along the transcripts that are cleaved by
miRNA. (A) and (B) 21-22 nt siRNA reads mapped over the 5’ and 3’ miRNA cleavage
fragments of 39 miRNA target genes. RPM: reads per million (21-22 nt) reads mapped to
all genes.
Page 22
22
Fig. S16
Fig. S16. The ct-siRNA loci are significantly enriched in the genes that are either up- or
down-regulated in the ein5-1 ski2-3 mutant. The expression levels of the genes were
normalized and the up- and down-regulated genes were defined with a two-fold cutoff.
RDR6-dependent differentially expressed gene sets (up- and down-regulated genes) were
obtained by comparing the ein5-1 ski2-3 transcriptome with that of rdr6-11 ein5-1 ski2-3.
χ2 test was used to calculate the significance of enrichment.
Page 23
23
Fig. S17
Fig. S17. The top-scoring 20 of 441 ct-siRNA-generating genes ranked by the fold-
change of ct-siRNA abundance between Col-0 (C) and ein5-1 ski2-3 (es).
Page 24
24
Fig. S18
Fig. S18. The 441 ct-siRNA loci have significantly higher expression levels than the rest
of the genome in ein5-1 ski2-3. 23700 transcripts that have expression level of RPKM >
0.5 were considered. The difference is highly significant (p < 10-16
, Kolmogorov-Smirnov
test).
Page 25
25
Fig. S19
Fig. S19. A proposed model depicting that bidirectional cytoplasmic RNA decay
suppresses endogenous PTGS. The cytoplasmic aberrant mRNAs are efficiently
processed by 5’-3’ (EIN5) and 3’-5’ (SKI-Exosome) RNA decay machineries. Upon
RNA decay deficiency, aberrant mRNAs are channeled into the SGS3/RDR6-dependent
double-strand synthesis and the production of 21-22 nt ct-siRNAs. The DCL4-mediated
21 nt ct-siRNA pathway could serve as an decoy to compete with the more destructive
DCL2-mediated 22 nt ct-siRNA pathway that is capable of triggering amplified PTGS.
Page 26
26
Table S1 to S4
These tables are provided as a separate Excel document.
Table S1. Geneset enrichment analysis with the gene set whose expression was co-
regulated by EIN5 and SKI2.
Table S2. Quantitation of 21-22 nt smRNA reads arising from the 441 genes from which
EIN5- and SKI2-affected smRNAs are produced.Three biological repeats for
each genotypes: c1-c3, Col-0; es1-es3, ein5-1 ski2-3; r1-r3, rdr6-11; res1-res3,
rdr6-11 ein5-1 ski2-3.
Table S3. A list of 39 ct-siRNA-generating miRNA target genes.
Table S4. Primers used in this research.
Page 27
References and Notes
1. C. Cogoni, G. Macino, Post-transcriptional gene silencing across kingdoms. Curr. Opin.
Genet. Dev. 10, 638–643 (2000). Medline doi:10.1016/S0959-437X(00)00134-9
2. R. C. Wilson, J. A. Doudna, Molecular mechanisms of RNA interference. Annu. Rev. Biophys.
42, 217–239 (2013). Medline doi:10.1146/annurev-biophys-083012-130404
3. M. Ghildiyal, P. D. Zamore, Small silencing RNAs: An expanding universe. Nat. Rev. Genet.
10, 94–108 (2009). Medline doi:10.1038/nrg2504
4. M. Incarbone, P. Dunoyer, RNA silencing and its suppression: Novel insights from in planta
analyses. Trends Plant Sci. 18, 382–392 (2013). Medline
doi:10.1016/j.tplants.2013.04.001
5. N. Pumplin, O. Voinnet, RNA silencing suppression by plant pathogens: Defence, counter-
defence and counter-counter-defence. Nat. Rev. Microbiol. 11, 745–760 (2013). Medline
doi:10.1038/nrmicro3120
6. A. B. Moreno, A. E. Martínez de Alba, F. Bardou, M. D. Crespi, H. Vaucheret, A. Maizel, A.
C. Mallory, Cytoplasmic and nuclear quality control and turnover of single-stranded
RNA modulate post-transcriptional gene silencing in plants. Nucleic Acids Res. 41, 4699–
4708 (2013). Medline doi:10.1093/nar/gkt152
7. S. Gazzani, T. Lawrenson, C. Woodward, D. Headon, R. Sablowski, A link between mRNA
turnover and RNA interference in Arabidopsis. Science 306, 1046–1048 (2004). Medline
doi:10.1126/science.1101092
8. Q. Chao, M. Rothenberg, R. Solano, G. Roman, W. Terzaghi, J. R. Ecker, Activation of the
ethylene gas response pathway in Arabidopsis by the nuclear protein ETHYLENE-
INSENSITIVE3 and related proteins. Cell 89, 1133–1144 (1997). Medline
doi:10.1016/S0092-8674(00)80300-1
9. H. Guo, J. R. Ecker, Plant responses to ethylene gas are mediated by SCF(EBF1/EBF2)-
dependent proteolysis of EIN3 transcription factor. Cell 115, 667–677 (2003). Medline
doi:10.1016/S0092-8674(03)00969-3
10. G. Olmedo, H. Guo, B. D. Gregory, S. D. Nourizadeh, L. Aguilar-Henonin, H. Li, F. An, P.
Guzman, J. R. Ecker, ETHYLENE-INSENSITIVE5 encodes a 5′→3′ exoribonuclease
required for regulation of the EIN3-targeting F-box proteins EBF1/2. Proc. Natl. Acad.
Sci. U.S.A. 103, 13286–13293 (2006). Medline doi:10.1073/pnas.0605528103
11. R. Solano, A. Stepanova, Q. Chao, J. R. Ecker, Nuclear events in ethylene signaling: A
transcriptional cascade mediated by ETHYLENE-INSENSITIVE3 and ETHYLENE-
RESPONSE-FACTOR1. Genes Dev. 12, 3703–3714 (1998). Medline
doi:10.1101/gad.12.23.3703
12. E. Dorcey, A. Rodriguez-Villalon, P. Salinas, L. Santuari, S. Pradervand, K. Harshman, C. S.
Hardtke, Context-dependent dual role of SKI8 homologs in mRNA synthesis and
turnover. PLOS Genet. 8, e1002652 (2012). Medline doi:10.1371/journal.pgen.1002652
Page 28
13. J. T. Brown, X. Bai, A. W. Johnson, The yeast antiviral proteins Ski2p, Ski3p, and Ski8p
exist as a complex in vivo. RNA 6, 449–457 (2000). Medline
doi:10.1017/S1355838200991787
14. X. Zhang, Y. Chen, X. Lin, X. Hong, Y. Zhu, W. Li, W. He, F. An, H. Guo, Adenine
phosphoribosyl transferase 1 is a key enzyme catalyzing cytokinin conversion from
nucleobases to nucleotides in Arabidopsis. Mol. Plant 6, 1661–1672 (2013). Medline
doi:10.1093/mp/sst071
15. T. Dalmay, A. Hamilton, S. Rudd, S. Angell, D. C. Baulcombe, An RNA-dependent RNA
polymerase gene in Arabidopsis is required for posttranscriptional gene silencing
mediated by a transgene but not by a virus. Cell 101, 543–553 (2000). Medline
doi:10.1016/S0092-8674(00)80864-8
16. E. Grotewold, The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 57,
761–780 (2006). Medline doi:10.1146/annurev.arplant.57.032905.105248
17. N. G. Bologna, O. Voinnet, The diversity, biogenesis, and activities of endogenous silencing
small RNAs in Arabidopsis. Annu. Rev. Plant Biol. 65, 473–503 (2014). Medline
doi:10.1146/annurev-arplant-050213-035728
18. F. Vazquez, T. Hohn, Biogenesis and biological activity of secondary siRNAs in plants.
Scientifica 2013, 783253 (2013). Medline doi:10.1155/2013/783253
19. A. Peragine, M. Yoshikawa, G. Wu, H. L. Albrecht, R. S. Poethig, SGS3 and
SGS2/SDE1/RDR6 are required for juvenile development and the production of trans-
acting siRNAs in Arabidopsis. Genes Dev. 18, 2368–2379 (2004). Medline
doi:10.1101/gad.1231804
20. E. Allen, Z. Xie, A. M. Gustafson, J. C. Carrington, microRNA-directed phasing during
trans-acting siRNA biogenesis in plants. Cell 121, 207–221 (2005). Medline
doi:10.1016/j.cell.2005.04.004
21. C. F. Hwang, Y. Lin, T. D’Souza, C. L. Cheng, Sequences necessary for nitrate-dependent
transcription of Arabidopsis nitrate reductase genes. Plant Physiol. 113, 853–862 (1997).
Medline doi:10.1104/pp.113.3.853
22. G. Rubin, T. Tohge, F. Matsuda, K. Saito, W. R. Scheible, Members of the LBD family of
transcription factors repress anthocyanin synthesis and affect additional nitrogen
responses in Arabidopsis. Plant Cell 21, 3567–3584 (2009). Medline
doi:10.1105/tpc.109.067041
23. C. Diaz, V. Saliba-Colombani, O. Loudet, P. Belluomo, L. Moreau, F. Daniel-Vedele, J. F.
Morot-Gaudry, C. Masclaux-Daubresse, Leaf yellowing and anthocyanin accumulation
are two genetically independent strategies in response to nitrogen limitation in
Arabidopsis thaliana. Plant Cell Physiol. 47, 74–83 (2006). Medline
doi:10.1093/pcp/pci225
24. D. Schubert, B. Lechtenberg, A. Forsbach, M. Gils, S. Bahadur, R. Schmidt, Silencing in
Arabidopsis T-DNA transformants: The predominant role of a gene-specific RNA
sensing mechanism versus position effects. Plant Cell 16, 2561–2572 (2004). Medline
doi:10.1105/tpc.104.024547
Page 29
25. C. A. Beelman, R. Parker, Degradation of mRNA in eukaryotes. Cell 81, 179–183 (1995).
Medline doi:10.1016/0092-8674(95)90326-7
26. F. An, Q. Zhao, Y. Ji, W. Li, Z. Jiang, X. Yu, C. Zhang, Y. Han, W. He, Y. Liu, S. Zhang, J.
R. Ecker, H. Guo, Ethylene-induced stabilization of ETHYLENE INSENSITIVE3 and
EIN3-LIKE1 is mediated by proteasomal degradation of EIN3 binding F-box 1 and 2 that
requires EIN2 in Arabidopsis. Plant Cell 22, 2384–2401 (2010). Medline
doi:10.1105/tpc.110.076588
27. P. Guzmán, J. R. Ecker, Exploiting the triple response of Arabidopsis to identify ethylene-
related mutants. Plant Cell 2, 513–523 (1990). Medline doi:10.1105/tpc.2.6.513
28. X. Hou, L. Li, Z. Peng, B. Wei, S. Tang, M. Ding, J. Liu, F. Zhang, Y. Zhao, H. Gu, L. J. Qu,
A platform of high-density INDEL/CAPS markers for map-based cloning in Arabidopsis.
Plant J. 63, 880–888 (2010). Medline doi:10.1111/j.1365-313X.2010.04277.x
29. J. Doebley, A. Stec, Inheritance of the morphological differences between maize and
teosinte: Comparison of results for two F2 populations. Genetics 134, 559–570 (1993).
Medline
30. D. W. Meinke, “Seed development in Arabidopsis thaliana.” in Arabidopsis, E. M.
Meyerowitz, C. R. Somerville, Eds. (Cold Spring Harbor Laboratory Press, Cold Spring
Harbor, 1994); pp. 253–295.
31. J. M. Alonso, A. N. Stepanova, T. J. Leisse, C. J. Kim, H. Chen, P. Shinn, D. K. Stevenson,
J. Zimmerman, P. Barajas, R. Cheuk, C. Gadrinab, C. Heller, A. Jeske, E. Koesema, C.
C. Meyers, H. Parker, L. Prednis, Y. Ansari, N. Choy, H. Deen, M. Geralt, N. Hazari, E.
Hom, M. Karnes, C. Mulholland, R. Ndubaku, I. Schmidt, P. Guzman, L. Aguilar-
Henonin, M. Schmid, D. Weigel, D. E. Carter, T. Marchand, E. Risseeuw, D. Brogden,
A. Zeko, W. L. Crosby, C. C. Berry, J. R. Ecker, Genome-wide insertional mutagenesis
of Arabidopsis thaliana. Science 301, 653–657 (2003). Medline
doi:10.1126/science.1086391
32. M. R. Smith, M. R. Willmann, G. Wu, T. Z. Berardini, B. Möller, D. Weijers, R. S. Poethig,
Cyclophilin 40 is required for microRNA activity in Arabidopsis. Proc. Natl. Acad. Sci.
U.S.A. 106, 5424–5429 (2009). Medline doi:10.1073/pnas.0812729106
33. S. Arvidsson, M. Kwasniewski, D. M. Riaño-Pachón, B. Mueller-Roeber, QuantPrime – a
flexible tool for reliable high-throughput primer design for quantitative PCR. BMC
Bioinformatics 9, 465 (2008). Medline doi:10.1186/1471-2105-9-465
34. C. Fankhauser, K. C. Yeh, J. C. Lagarias, H. Zhang, T. D. Elich, J. Chory, PKS1, a substrate
phosphorylated by phytochrome that modulates light signaling in Arabidopsis. Science
284, 1539–1541 (1999). Medline doi:10.1126/science.284.5419.1539
35. E. Mortz, T. N. Krogh, H. Vorum, A. Görg, Improved silver staining protocols for high
sensitivity protein identification using matrix-assisted laser desorption/ionization-time of
flight analysis. Proteomics 1, 1359–1363 (2001). Medline doi:10.1002/1615-
9861(200111)1:11<1359::AID-PROT1359>3.0.CO;2-Q
36. H. Tabara, E. Yigit, H. Siomi, C. C. Mello, The dsRNA binding protein RDE-4 interacts with
RDE-1, DCR-1, and a DExH-box helicase to direct RNAi in C. elegans. Cell 109, 861–
871 (2002). Medline doi:10.1016/S0092-8674(02)00793-6
Page 30
37. A. Shevchenko, M. Wilm, O. Vorm, M. Mann, Mass spectrometric sequencing of proteins
silver-stained polyacrylamide gels. Anal. Chem. 68, 850–858 (1996). Medline
doi:10.1021/ac950914h
38. H. Li, R. Durbin, Fast and accurate short read alignment with Burrows-Wheeler transform.
Bioinformatics 25, 1754–1760 (2009). Medline doi:10.1093/bioinformatics/btp324
39. M. J. L. de Hoon, S. Imoto, J. Nolan, S. Miyano, Open source clustering software.
Bioinformatics 20, 1453–1454 (2004). Medline doi:10.1093/bioinformatics/bth078
40. A. J. Saldanha, Java Treeview—Extensible visualization of microarray data. Bioinformatics
20, 3246–3248 (2004). Medline doi:10.1093/bioinformatics/bth349
41. B. Langmead, C. Trapnell, M. Pop, S. L. Salzberg, Ultrafast and memory-efficient alignment
of short DNA sequences to the human genome. Genome Biol. 10, R25 (2009). Medline
doi:10.1186/gb-2009-10-3-r25
42. J. W. Nicol, G. A. Helt, S. G. Blanchard Jr., A. Raja, A. E. Loraine, The Integrated Genome
Browser: Free software for distribution and exploration of genome-scale datasets.
Bioinformatics 25, 2730–2731 (2009). Medline doi:10.1093/bioinformatics/btp472
43. X. Yi, Z. Du, Z. Su, PlantGSEA: A gene set enrichment analysis toolkit for plant community.
Nucleic Acids Res. 41, W98–W103 (2013). Medline doi:10.1093/nar/gkt281
44. C. Xie, X. Mao, J. Huang, Y. Ding, J. Wu, S. Dong, L. Kong, G. Gao, C. Y. Li, L. Wei,
KOBAS 2.0: A web server for annotation and identification of enriched pathways and
diseases. Nucleic Acids Res. 39 (suppl. 2), W316–W322 (2011). Medline
doi:10.1093/nar/gkr483
45. G. S. Pall, C. Codony-Servat, J. Byrne, L. Ritchie, A. Hamilton, Carbodiimide-mediated
cross-linking of RNA to nylon membranes improves the detection of siRNA, miRNA and
piRNA by northern blot. Nucleic Acids Res. 35, e60 (2007). Medline
doi:10.1093/nar/gkm112