www.sciencemag.org/content/342/6160/863/suppl/DC1 Supplementary Materials for Staphylococcus aureus Degrades Neutrophil Extracellular Traps to Promote Immune Cell Death Vilasack Thammavongsa, Dominique M. Missiakas, Olaf Schneewind* *Corresponding author. E-mail: [email protected]Published 15 November 2013, Science 342, 863 (2013) DOI: 10.1126/science.1242255 This PDF file includes: Materials and Methods Supplementary Text Figs. S1 to S8 Table S1 Full Reference List
Transcript
www.sciencemag.org/content/342/6160/863/suppl/DC1
Supplementary Materials for
Staphylococcus aureus Degrades Neutrophil Extracellular Traps to
Promote Immune Cell Death
Vilasack Thammavongsa, Dominique M. Missiakas, Olaf Schneewind*
Published 15 November 2013, Science 342, 863 (2013)
DOI: 10.1126/science.1242255
This PDF file includes:
Materials and Methods
Supplementary Text
Figs. S1 to S8
Table S1
Full Reference List
2
Materials and Methods Bacterial strains
S. aureus strains were grown in TSB at 37°C. All mutants used in this study were obtained from the Phoenix (ΦΝΞ) library (10). Each Phoenix isolate is a derivative of the clinical isolate Newman (21). All bursa aurealis insertions were transduced into wild-type S. aureus Newman using bacteriophage φ85 and verified by PCR analysis. Erythromycin was used at 10 mg l−1 for allele selection during transduction into S. aureus Newman. Chloramphenicol was used at 10 mg l−1 for plasmid and allele selection with padsA and pnuc.
Plasmids
The plasmid padsA was described earlier (13). PCR amplification reactions with the oligonucleotide primers VT30 (5’-GTTTCAGGATCCTTATTAGACGTGATCATG-TTAAAGAAGCAC-3’) and VT31 (5’-GTTTCAGGATCCTTATTGACCTGAATCAG-CGT-3’) were used to amplify the nuclease gene. Ligation of VT30/VT31 (the structural nuc gene and 500 bp upstream from start site) PCR product into the BamH1 site of pOS1 (22) generated the plasmid pnuc. Animal experiments
Experimental protocols involving animals were reviewed, approved and performed under regulatory supervision of The University of Chicago’s Institutional Biosafety Committee (IBC) and Institutional Animal Care and Use Committee (IACUC). BALB/c mice were purchased from Charles River Laboratories. Overnight cultures of S. aureus strains were diluted 1:100 into fresh TSB and grown for 3 h at 37°C. Staphylococci were centrifuged, washed twice and diluted in PBS to yield an OD600 of 0.5 (1×108 CFU ml−1). Viable staphylococci were enumerated by colony formation on tryptic soy agar plates to quantify the infectious dose. Mice were anaesthetized by intraperitoneal injection of 50–65 mg of ketamine and 3–6 mg of xylazine per kilogram of body weight. One hundred µl of bacterial suspension (1×107 CFU) were administered intravenously via retro-orbital injection into BALB/c mice (6-week old female). On day 5, mice were killed by compressed CO2 inhalation and kidneys dissected. Histology
Kidney tissues were incubated at room temperature in 10% formalin for 24 hours. Tissues were embedded in paraffin, thin-sectioned, stained with hematoxylin/eosin and examined by microscopy. For immuno-histochemical staining, kidneys of infected animals were dissected, placed in 1×PBS on ice, and flash frozen in Tissue Tek OCT within cryomolds. Samples were cryo-sectioned (10 μm cuts), mounted on slides, and stored at -80ºC. Prior to staining, slides were warmed to room temperature for 30 minutes, fixed in ice cold acetone for 10 minutes, and washed twice with ice cold PBS. The slides were blocked in 1% BSA, 1:20 human IgG (Sigma), 1×PBS for 1 hour at room temperature in a humidified chamber. F4/80 and Ly-6G rat antibodies (1:100) and anti-caspase-3 rabbit antibody (1:10) were added to the slides and incubated for one hour with αF4/80 and αLy-6G or for six hours with anti-caspase-3. Immuno-histochemistry staining solutions were decanted and glass slides washed three times with PBS. Slides
3
were placed in 1×PBS 1:200 goat anti-rat HRP secondary antibody and allowed to incubate at room temperature. The solution was decanted, slides were washed three times with PBS, and developed with diaminobenzidine (Molecular Probes, Inc.). The slides were washed once more with PBS, mounted in N-propylgallate, and viewed with a Zeiss Axioskop light microscope. Chemicals
PMA (Sigma) was suspended at a concentration of 20 mM in DMSO and stored at –20 °C. dCF (pentostatin, NIPENT) was suspended at a concentration of 2 mg ml-1 in sterile PBS and stored at –20 °C. EHNA hydrochloride (Sigma) was suspended at a concentration of 3 mM in PBS and stored at –20 °C. Staphylococcal nuclease (Sigma) was suspended at a concentration of 1 unit µl-1 in sterile water. Z-DEVD-FMK (BD Biosciences) was suspended at a concentration of 10 mM in DMSO and stored at –20 °C. Type XV thymus DNA (Sigma) was suspended at a concentration of 1 mg ml-1 and stored at –20 °C. Purified adenosine, deoxyadenosine, 2’-deoxyadenosine 5’-monophosphate, 2’-deoxyadenosine 3’-monophosphate and recombinant adenosine deaminase were purchased from Sigma. Propidium iodide and trypan blue were purchased from Invitrogen. [14C]adenosine and [14C]inosine were purchased from Moravek Biochemicals. Adenosine deaminase activity For enzyme activity measurements in tissue culture media, 105 cells in 1 ml RPMI media (10% FBS, penicillin/streptomycin) were incubated at 37 °C 5% CO2 for 16 hours. Cells were then sedimented by centrifugation and 20 µl media incubated with 3 µCi [14C] adenosine for 60 minutes in the presence or absence of 300 µM EHNA or 100 µM dCF. Samples were then spotted on a silica plate followed by separation by TLC using a (75:25 isopropanol: ddH2O) 0.2 M ammonium bicarbonate solvent. For analyses with recombinant ADA1, 1 unit ADA1 was incubated with 3 µCi [14C] adenosine in 50 mM Tris-HCl, 150 mM NaCl (pH 7.5) for 60 minutes in the presence or absence of 300 µM EHNA. rpHPLC and mass spectrometry
Deoxyadenosine synthesis was analyzed by rpHPLC. Samples were chromatographed on a 250 mm×3 mm column (BDS Hypersil C18, 5 µm particle size, Thermoscientific). The mobile phase consisted of solution A (65 mM potassium phosphate, pH 6.0) and solution B (65 mM potassium phosphate, 25% methanol, pH 6.0). Deoxyadenosine was eluted with a solvent B gradient of 1 to 100% from 5 to 50 min. The solvent flow rate was 0.5 ml/min. Peaks were detected by their UV absorbance at 260 nm. The peak of deoxyadenosine in the rpHPLC chromatogram was identified by comparison of its retention time to the retention time of purified deoxyadenosine used as a standard sample. Fractions containing deoxyadenosine were co-spotted with matrix (α-cyano-4-hydroxycinnamic acid) and subjected to MALDI-MS under reflector positive conditions. Stimulation of neutrophil NETs
4
Blood from human volunteers was drawn using protocols that were reviewed, approved and performed under regulatory supervision of The University of Chicago’s Institutional Review Board (IRB). Human neutrophils were isolated from peripheral blood of healthy donors using a dextran-Ficoll method. Extracellular NET release was conducted as described earlier (23). Neutrophils (2×105 cells) were seeded in 500 μl RPMI (containing 2 % human serum albumin) per well and incubated for 1 hour with 5% CO2 at 37°C. PMA (100 μl 600 nM PMA in RPMI) was added per well and incubated for 3 hours at 37°C 5% CO2. Isolation of primary human monocytes
Fifteen mL of sodium citrate uncoagulated blood from human volunteers was diluted into PBS to a final volume of 35 ml and overlayed onto 15 ml of Ficoll-Paque. The sample was then centrifuged at 400 ×g for 30 minutes. The upper layer was aspirated and the peripheral blood mononuclear cells (PBMC) enriched in the interphase was carefully pipetted and collected. The cells were suspended in PBS and washed by centrifuging at 300 ×g for 15 minutes. Platelets were removed by an additional wash step and centrifugation at 200 ×g for 15 minutes. Peripheral blood mononuclear cells (PBMCs) were suspended in PBS and monoctyes isolated by depletion of lymphocytes via magnetic beads (Miltenyl Biotec). Purified monocytes (>90% confirmed by CD14 positive staining by FACS) were suspended in RPMI and subjected to further analysis. Isolation of bone marrow derived macrophages
Six-eight week-old mice were killed and bone marrow was collected from the femur. Bone marrow cells were cultured in Dulbecco modified Eagle medium (DMEM) containing 10% 3T3-MCSF cell supernatant for 6 days, resulting in the differentiation into naïve bone marrow-derived macrophages. Wild-type C57BL/6 mice were purchased from Jackson Laboratories (Bar Harbor, ME). Caspase1−/− macrophages were a gift from JD Sauer (University of Wisconsin, Madison, WI). Digestion of NETs and DNA
Overnight cultures of S. aureus strains were diluted 1:100 into fresh TSB and grown for 3 h at 37°C to an OD600 of 0.5 (1×108 CFU ml−1). One hundred µl staphylococci culture (107 CFU) was then added to PMA stimulated neutrophils and incubated in 5% CO2 at 37°C. Digested NET supernatants were then isolated by centrifugation. For digestion of DNA: 50 µg of thymus DNA (Sigma) was incubated with 5 units staphylococcal nuclease in 200 µl buffer (final concentration 240 mM Tris, 2.4 mM MgCl2, 2.4 mM CaCl2, pH 7.4) for 1 hour followed by the addition of 10 µM recombinant AdsA and 2.5 mM MnCl2 (final concentrations). The reaction was allowed to proceed for an additional 2 hours and was then analyzed by rpHPLC. Cellular cytoxicity assays
To measure cytotoxicity, 1×105 U937 cells, 1×105 HL60 cells, 1×105 THP-1 cells, 5 × 104 primary human neutrophils or 5 × 104 primary human monocytes in 2 ml RPMI media (10% FBS, penicillin/streptomycin) were incubated overnight with 300 µl digested NET samples from PMA stimulated neutrophils or purified nuclease / AdsA digested
5
thymus DNA (200 µl) in the presence or absence of 100 µM dCF as indicated. The cells were then stained with trypan blue and visualized by light microscopy. Apoptosis assays
Cells were treated as described for cellular cytoxicity with digested DNA and dAdo and incubated for 5 hours. Cells were then stained with Annexin V-FITC and propidium iodide using the ApoDETECT reagent kit (Invitrogen) and subjected to FACS analysis with a FACSCanto instrument (BD Biosciences).
Supplementary Text Abbreviations
NETs, neutrophil extracellular traps; DNA, deoxyribonucleic acid; Ado, adenosine; dAdo, deoxyadenosine; ADA, adenosine deaminase; dCF, deoxycoformycin; dATP, deoxyadenosine triphosphate; CFU, colony forming unit; PMA, phorbol 12-myristate 13-acetate; rpHPLC, reverse phase high pressure liquid chromatography; MALDI-MS, matrix assisted laser desorption / ionization mass spectrometry; AdsA, adenosine synthase A; ATP, adenosine triphosphate; EHNA, erythro-9-(2-hydroxy-3-nonyl)adenine. References 21. T. Baba, T. Bae, O. Schneewind, F. Takeuchi, K. Hiramatsu, Genome sequence of Staphylococcus aureus strain Newman and comparative analysis of staphylococcal genomes. J. Bacteriol. 190, 300 (2007). 22. O. Schneewind, P. Model, V. A. Fischetti, Sorting of protein A to the staphylococcal cell wall. Cell 70, 267 (1992). 23. V. Brinkmann, B. Laube, U. Abu Abed, C. Goosmann, A. Zychlinsky, Neutrophil extracellular traps: how to generate and visualize them. J. Vis. Exp. 36, 1724 (2010).
6
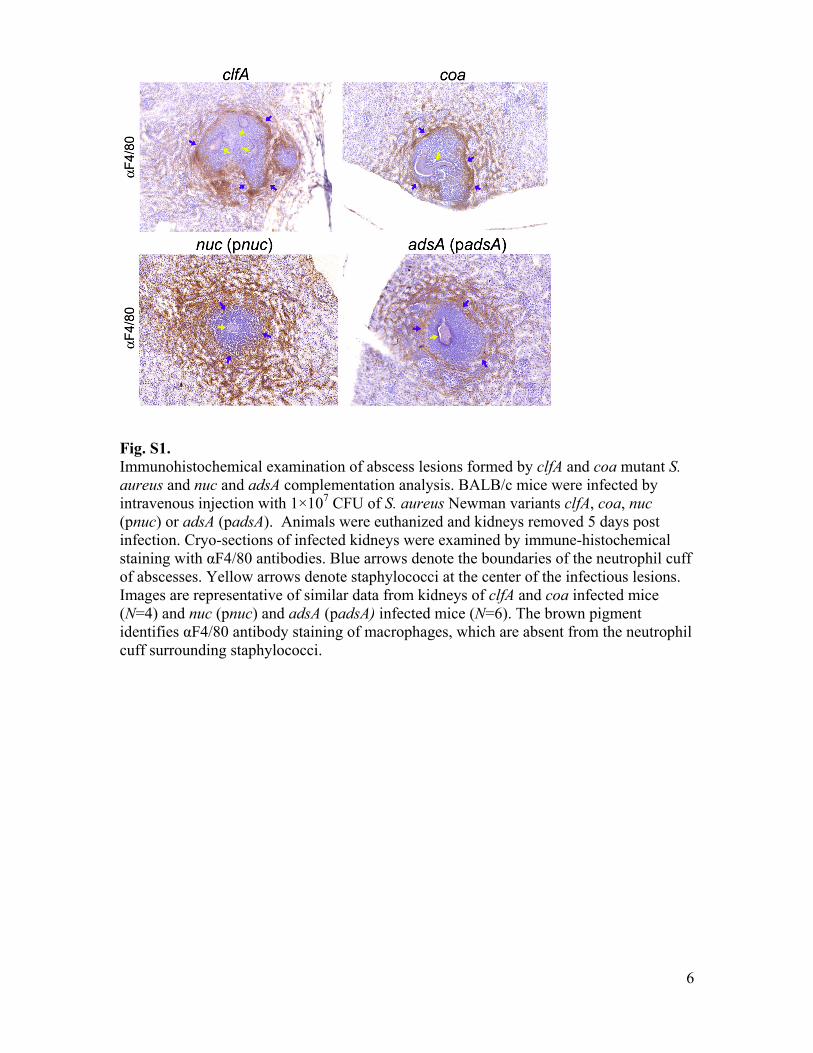
Fig. S1. Immunohistochemical examination of abscess lesions formed by clfA and coa mutant S. aureus and nuc and adsA complementation analysis. BALB/c mice were infected by intravenous injection with 1×107 CFU of S. aureus Newman variants clfA, coa, nuc (pnuc) or adsA (padsA). Animals were euthanized and kidneys removed 5 days post infection. Cryo-sections of infected kidneys were examined by immune-histochemical staining with αF4/80 antibodies. Blue arrows denote the boundaries of the neutrophil cuff of abscesses. Yellow arrows denote staphylococci at the center of the infectious lesions. Images are representative of similar data from kidneys of clfA and coa infected mice (N=4) and nuc (pnuc) and adsA (padsA) infected mice (N=6). The brown pigment identifies αF4/80 antibody staining of macrophages, which are absent from the neutrophil cuff surrounding staphylococci.
7
Fig. S2. Complementation analysis of nuclease activity in cultures of Staphylococcus aureus strains. Sterile tryptic soy broth culture medium (TSB) or conditioned culture medium derived from wild-type S. aureus Newman or its nuc and nuc (pnuc) variants were incubated with thymus DNA for one hour and subjected to agarose gel electrophoresis and ethidium bromide staining of nucleic acids. Nuclease activity was observed in conditioned culture medium of wild type and nuc (pnuc) variant S. aureus strains (N=2).
8
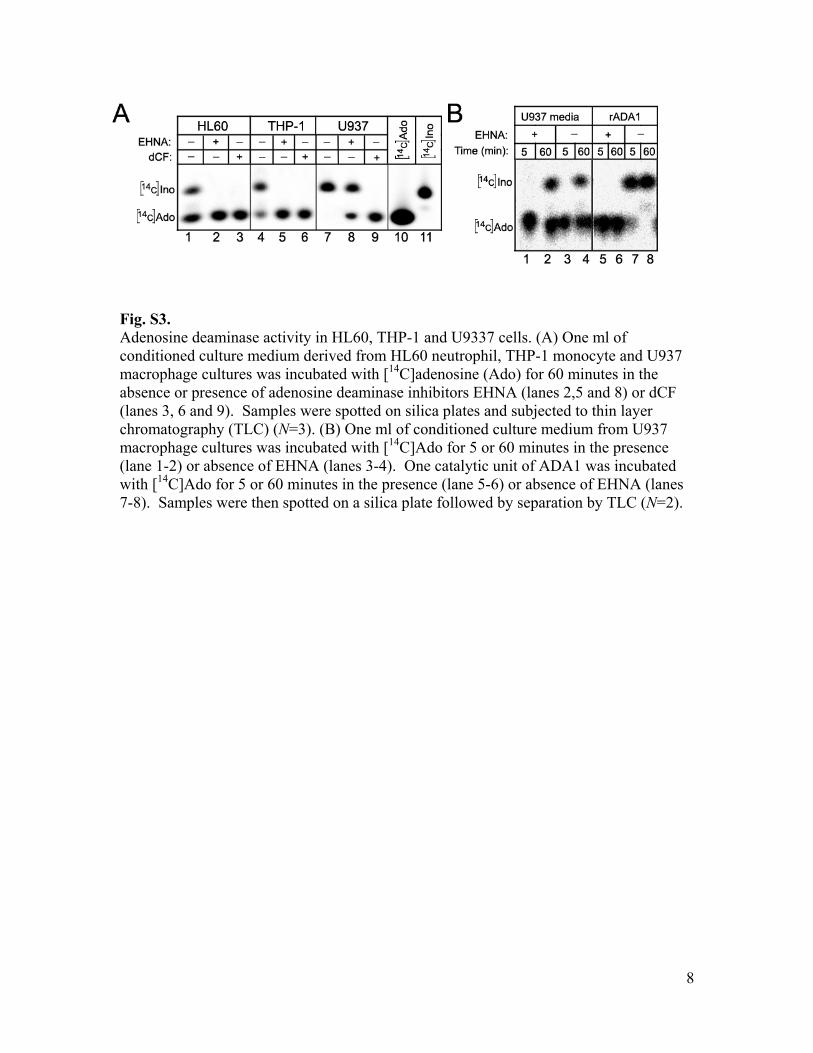
Fig. S3. Adenosine deaminase activity in HL60, THP-1 and U9337 cells. (A) One ml of conditioned culture medium derived from HL60 neutrophil, THP-1 monocyte and U937 macrophage cultures was incubated with [14C]adenosine (Ado) for 60 minutes in the absence or presence of adenosine deaminase inhibitors EHNA (lanes 2,5 and 8) or dCF (lanes 3, 6 and 9). Samples were spotted on silica plates and subjected to thin layer chromatography (TLC) (N=3). (B) One ml of conditioned culture medium from U937 macrophage cultures was incubated with [14C]Ado for 5 or 60 minutes in the presence (lane 1-2) or absence of EHNA (lanes 3-4). One catalytic unit of ADA1 was incubated with [14C]Ado for 5 or 60 minutes in the presence (lane 5-6) or absence of EHNA (lanes 7-8). Samples were then spotted on a silica plate followed by separation by TLC (N=2).
9
Fig. S4. AdsA displays 5’ and 3’ nucleotidase activity. The compounds 2’-deoxyadenosine 5’-monophosphate (A) and 2’-deoxyadenosine 3’-monophosphate (C) were each incubated with purified AdsA for 30 minutes (B and D) and reaction products subjected to rpHPLC. Deoxyadenosine adsorption peaks were identified by mass spectrometry (N=2).
10
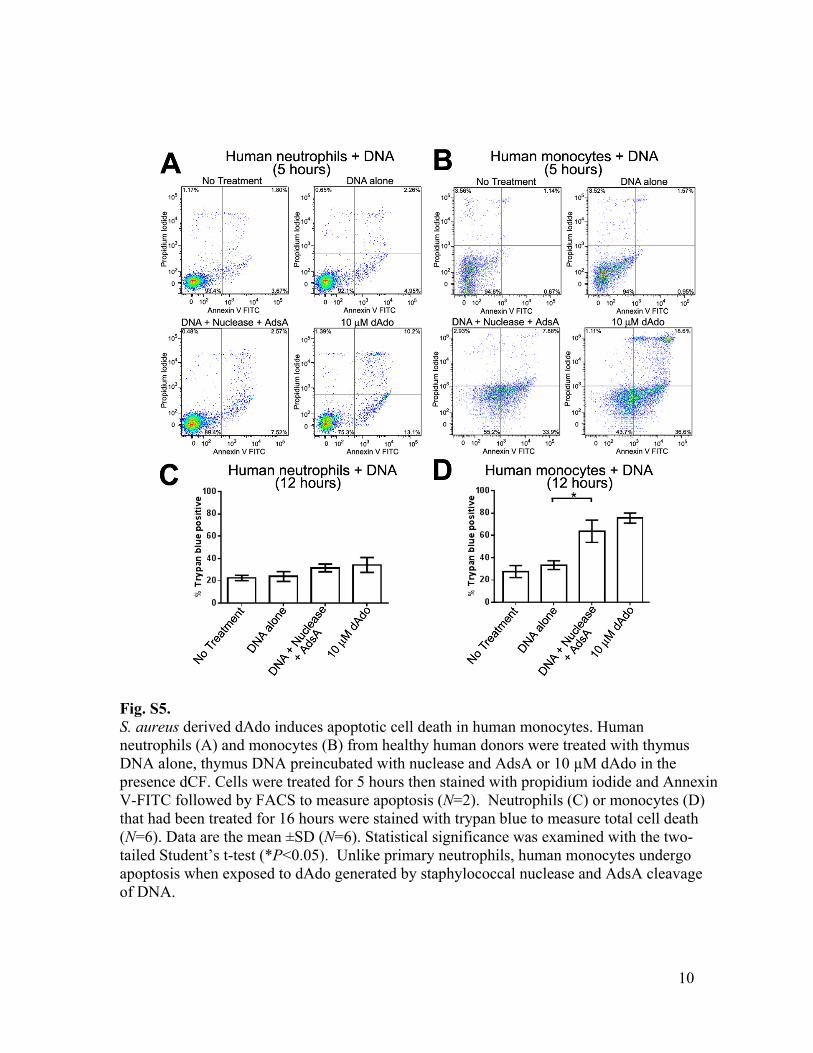
Fig. S5. S. aureus derived dAdo induces apoptotic cell death in human monocytes. Human neutrophils (A) and monocytes (B) from healthy human donors were treated with thymus DNA alone, thymus DNA preincubated with nuclease and AdsA or 10 µM dAdo in the presence dCF. Cells were treated for 5 hours then stained with propidium iodide and Annexin V-FITC followed by FACS to measure apoptosis (N=2). Neutrophils (C) or monocytes (D) that had been treated for 16 hours were stained with trypan blue to measure total cell death (N=6). Data are the mean ±SD (N=6). Statistical significance was examined with the two-tailed Student’s t-test (*P<0.05). Unlike primary neutrophils, human monocytes undergo apoptosis when exposed to dAdo generated by staphylococcal nuclease and AdsA cleavage of DNA.
11
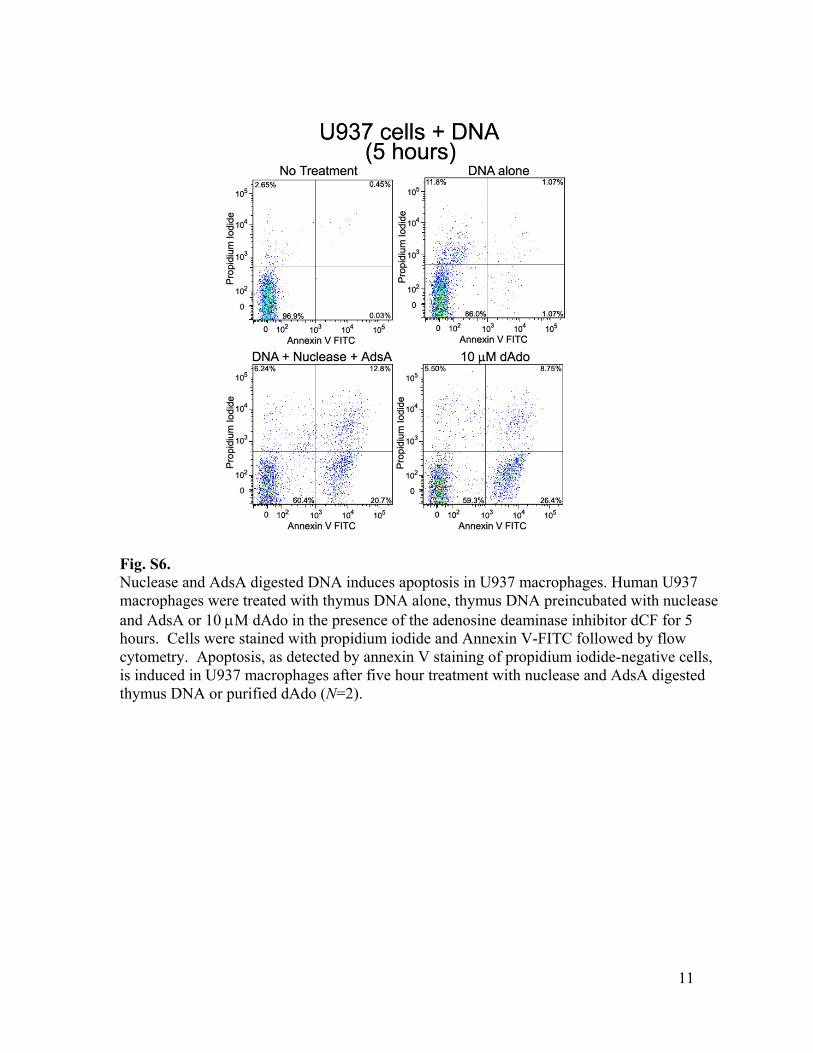
Fig. S6. Nuclease and AdsA digested DNA induces apoptosis in U937 macrophages. Human U937 macrophages were treated with thymus DNA alone, thymus DNA preincubated with nuclease and AdsA or 10 µM dAdo in the presence of the adenosine deaminase inhibitor dCF for 5 hours. Cells were stained with propidium iodide and Annexin V-FITC followed by flow cytometry. Apoptosis, as detected by annexin V staining of propidium iodide-negative cells, is induced in U937 macrophages after five hour treatment with nuclease and AdsA digested thymus DNA or purified dAdo (N=2).
12
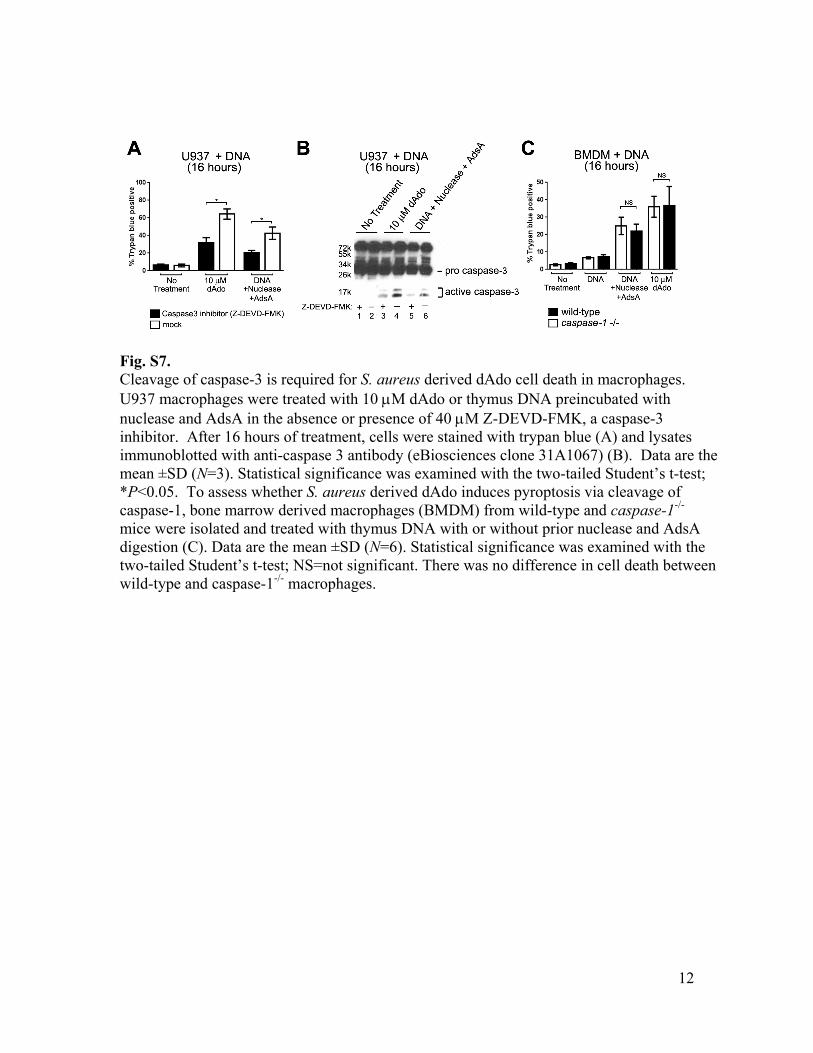
Fig. S7. Cleavage of caspase-3 is required for S. aureus derived dAdo cell death in macrophages. U937 macrophages were treated with 10 µM dAdo or thymus DNA preincubated with nuclease and AdsA in the absence or presence of 40 µM Z-DEVD-FMK, a caspase-3 inhibitor. After 16 hours of treatment, cells were stained with trypan blue (A) and lysates immunoblotted with anti-caspase 3 antibody (eBiosciences clone 31A1067) (B). Data are the mean ±SD (N=3). Statistical significance was examined with the two-tailed Student’s t-test; *P<0.05. To assess whether S. aureus derived dAdo induces pyroptosis via cleavage of caspase-1, bone marrow derived macrophages (BMDM) from wild-type and caspase-1-/- mice were isolated and treated with thymus DNA with or without prior nuclease and AdsA digestion (C). Data are the mean ±SD (N=6). Statistical significance was examined with the two-tailed Student’s t-test; NS=not significant. There was no difference in cell death between wild-type and caspase-1-/- macrophages.
13
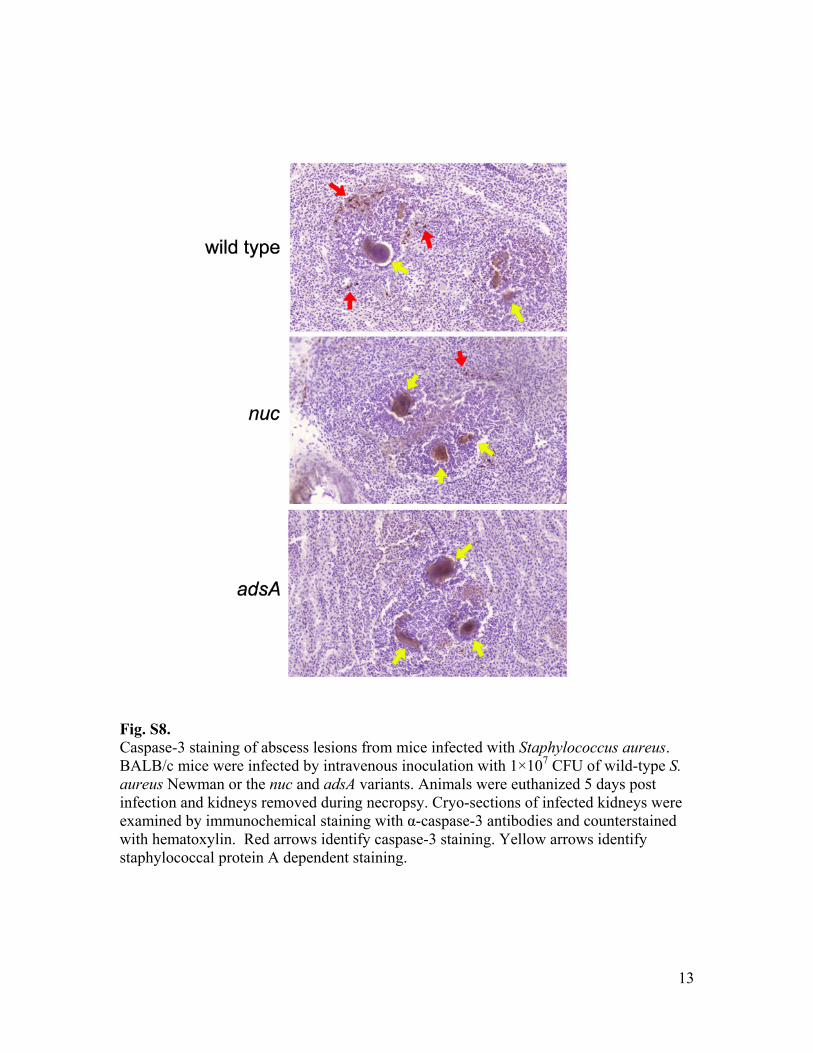
Fig. S8. Caspase-3 staining of abscess lesions from mice infected with Staphylococcus aureus. BALB/c mice were infected by intravenous inoculation with 1×107 CFU of wild-type S. aureus Newman or the nuc and adsA variants. Animals were euthanized 5 days post infection and kidneys removed during necropsy. Cryo-sections of infected kidneys were examined by immunochemical staining with α-caspase-3 antibodies and counterstained with hematoxylin. Red arrows identify caspase-3 staining. Yellow arrows identify staphylococcal protein A dependent staining.
14
Table S1. Histopathological examination of kidneys from mice infected with Staphylococcus aureus strains. S. aureus Newman straina Number of abscessesb P valuec wild-type 1.78 (±0.28) - nuc 0.65 (±0.18) 0.0035 adsA 0.45 (±0.12) 0.0001 nuc (pnuc) 1.61 (±0.38) 0.1198 adsA (padsA) 1.14 (±0.28) 0.6134 aBALB/c mice (N=15) were infected by intravenous injection with 1×107 CFU S. aureus Newman (wild-type) or its variants. On day 5 following infection, animals were euthanized, necropsied and kidneys removed for histopathology analyses. bHistopathology of hematoxylin-eosin stained thin-sectioned kidney tissues from 15 infected BALB/c mice in each cohort. The data are mean ±SD of the number of abscesses per kidney. cStatistical significance was analyzed with the unpaired two-tailed Student’s t-test and P values recorded.
References and Notes
1. A. J. Kallen, Y. Mu, S. Bulens, A. Reingold, S. Petit, K. Gershman, S. M. Ray, L. H. Harrison,
R. Lynfield, G. Dumyati, J. M. Townes, W. Schaffner, P. R. Patel, S. K. Fridkin; Active
Bacterial Core surveillance (ABCs) MRSA Investigators of the Emerging Infections
Program, Health care-associated invasive MRSA infections, 2005-2008. JAMA 304, 641–
648 (2010). Medline doi:10.1001/jama.2010.1115
2. A. N. Spaan, B. G. J. Surewaard, R. Nijland, J. A. G. van Strijp, Neutrophils versus
Staphylococcus aureus: A biological tug of war. Annu. Rev. Microbiol. 67, 629–650




















![4.1 WED ! (3T3) 100 *EYRAV4 2.0 Adventure (5A*GJ.4WD.CVT ... · 4.1 WED ! (3T3) 100 *EYRAV4 2.0 Adventure (5A*GJ.4WD.CVT) (6BA-MXAA54-ANXVBl [20V] Hybrid RALL COROUA T-Connect74Yñ-e](https://static.documents.pub/doc/80x56/60253b0c218dd170dc469fb2/41-wed-3t3-100-eyrav4-20-adventure-5agj4wdcvt-41-wed-3t3-100.jpg)




![Increased Growth of NIH/3T3 Cells by Transfection with Human pi … · [CANCER RESEARCH 52. 428-436. January 15. 1992] Increased Growth of NIH/3T3 Cells by Transfection with Human](https://static.documents.pub/doc/80x56/60b9042eb86d34336e7964ac/increased-growth-of-nih3t3-cells-by-transfection-with-human-pi-cancer-research.jpg)

