www.sciencemag.org/content/344/6187/992/suppl/DC1 Supplementary Materials for Crystal structure of a heterotetrameric NMDA receptor ion channel Erkan Karakas and Hiro Furukawa* *Corresponding author. E-mail: [email protected]Published 30 May 2014, Science 344, 992 (2014) DOI: 10.1126/science.1251915 This PDF file includes Materials and Methods Figs. S1 to S16 Table S1 References
Transcript
www.sciencemag.org/content/344/6187/992/suppl/DC1
Supplementary Materials for
Crystal structure of a heterotetrameric NMDA receptor ion channel
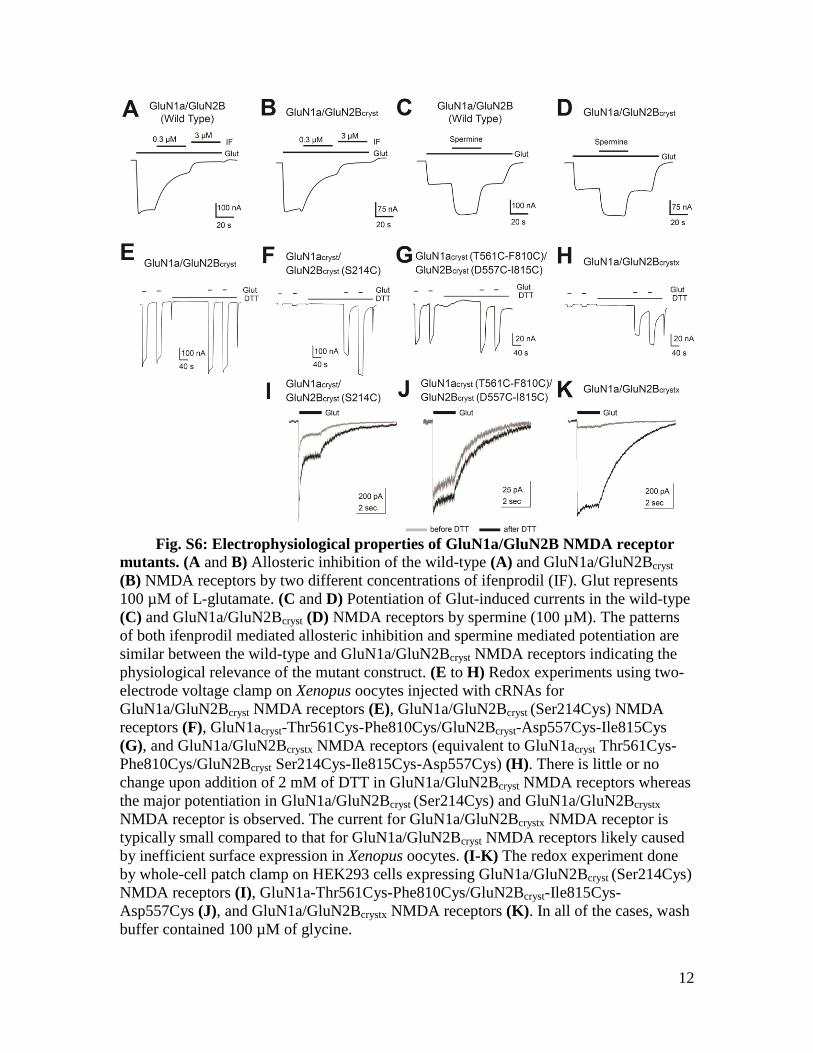
Asp557Cys (J), and GluN1a/GluN2Bcrystx NMDA receptors (K). In all of the cases, wash
buffer contained 100 µM of glycine.
13
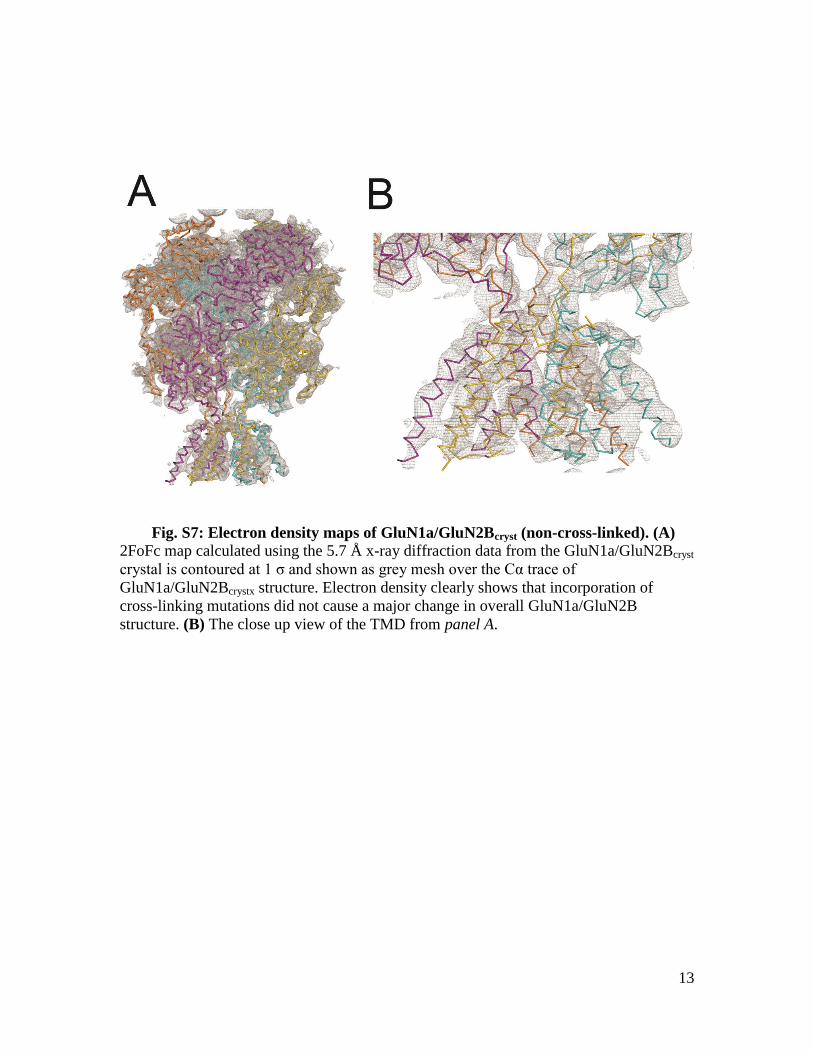
Fig. S7: Electron density maps of GluN1a/GluN2Bcryst (non-cross-linked). (A) 2FoFc map calculated using the 5.7 Å x-ray diffraction data from the GluN1a/GluN2Bcryst
crystal is contoured at 1 σ and shown as grey mesh over the Cα trace of
GluN1a/GluN2Bcrystx structure. Electron density clearly shows that incorporation of
cross-linking mutations did not cause a major change in overall GluN1a/GluN2B
structure. (B) The close up view of the TMD from panel A.
14
Fig. S8: Electron densities in the transmembrane domains in
GluN1a/GluN2Bcrystx. 2FoFc maps for the M1, M2, M3 and M4 helices of both GluN1a
(α) and GluN2B (β) TMDs are prepared with a B-factor sharpening factor of -90 Å2,
countered at 1.0 σ and shown as grey mesh. The models of GluN1a (α) and GluN2B (β)
are shown as yellow and cyan Cα traces, respectively. Cαs of methionine residues are
shown as spheres. Anomalous difference Fourier maps calculated from data collected on
two different selenomethionine-derivative crystals are averaged around the 2-fold non-
crystallographic axis using COOT (43) and shown as red mesh. The maps are contoured
at the following σ levels; 2.8 σ for GluN1a Met 555, 3.6 σ for GluN1a Met 576, 2.5 σ for
GluN1a Met 607, 2.5 σ for GluN1a Met 634, 4.2 σ for GluN1a Met 641, 3.3 σ for
GluN1a Met 813, 2.7 σ for GluN1a Met 818, 2.5 σ for GluN2B Met 561, 2.8 σ for
GluN2B Met 631, 2.5 σ for GluN2B Met 654, 3.0 σ for GluN2B Met 824 and 2.5 σ for
GluN2B Met 829. No significant signal was observed for residues GluN2B Met 562 and
Met 565.
15
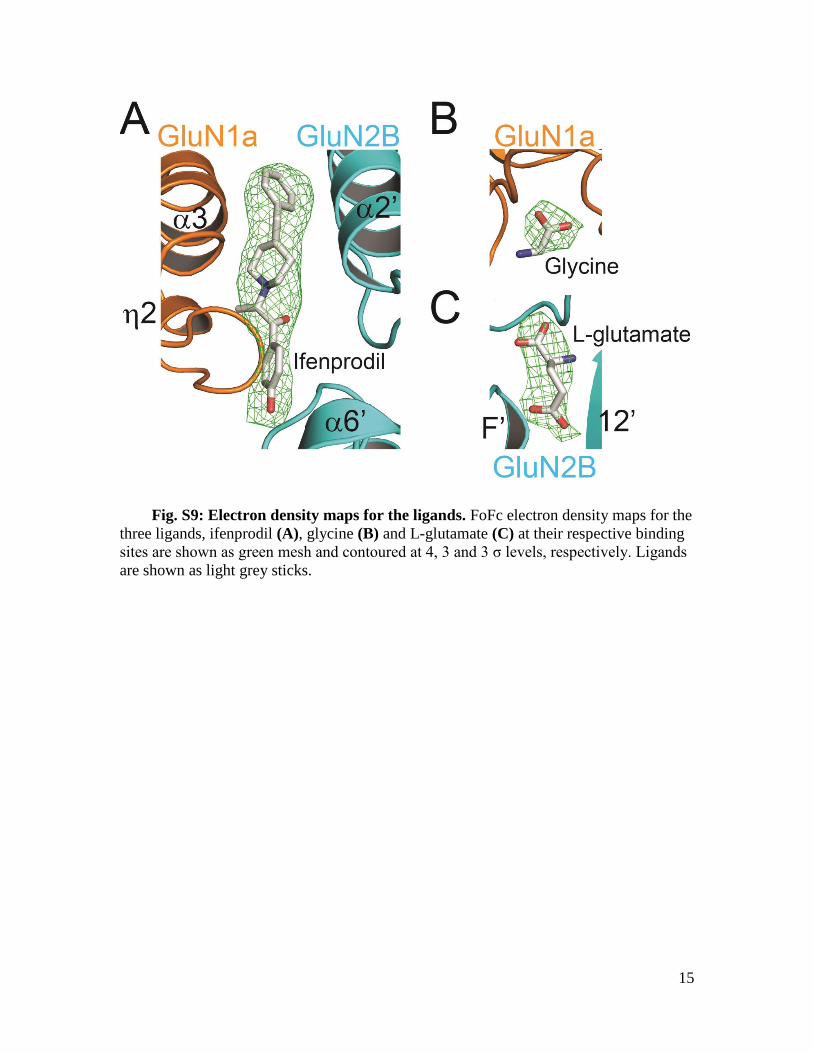
Fig. S9: Electron density maps for the ligands. FoFc electron density maps for the
three ligands, ifenprodil (A), glycine (B) and L-glutamate (C) at their respective binding
sites are shown as green mesh and contoured at 4, 3 and 3 σ levels, respectively. Ligands
are shown as light grey sticks.
16
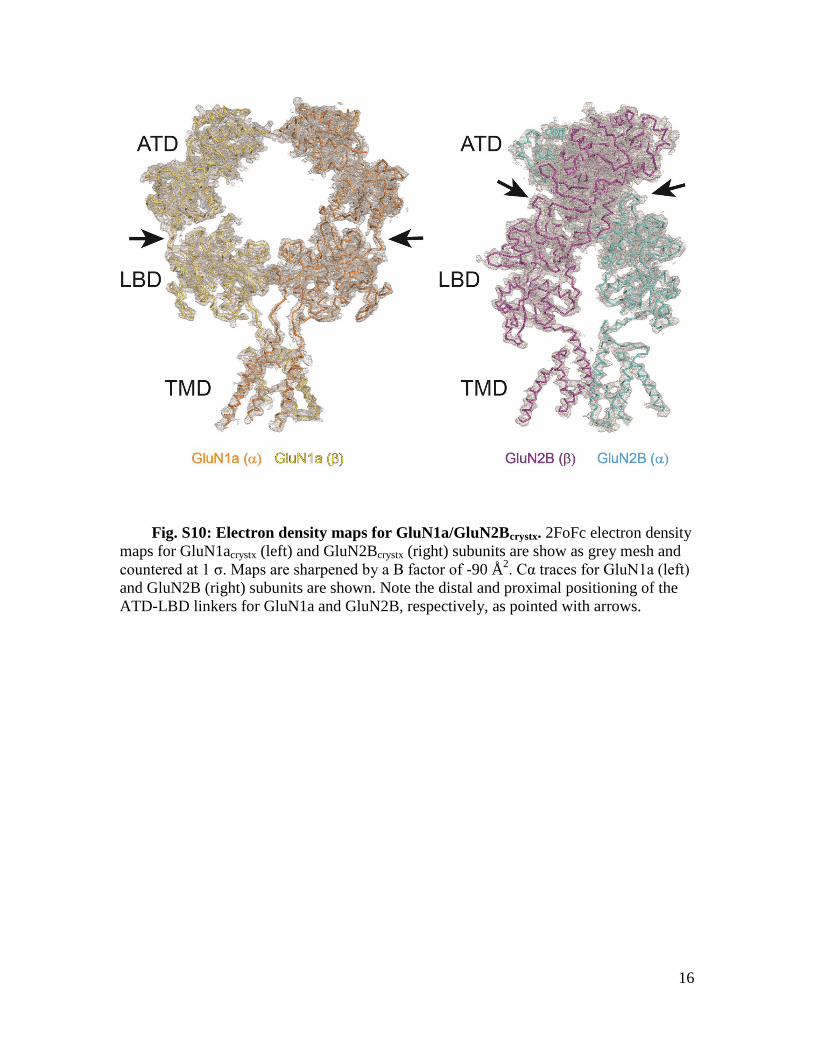
Fig. S10: Electron density maps for GluN1a/GluN2Bcrystx. 2FoFc electron density
maps for GluN1acrystx (left) and GluN2Bcrystx (right) subunits are show as grey mesh and
countered at 1 σ. Maps are sharpened by a B factor of -90 Å2. Cα traces for GluN1a (left)
and GluN2B (right) subunits are shown. Note the distal and proximal positioning of the
ATD-LBD linkers for GluN1a and GluN2B, respectively, as pointed with arrows.
17
Fig. S11: Dimer-of-dimers assembly of GluN1a/GluN2B NMDA receptors.
Tetrameric arrangement of ATD (A), LBD (B) and TMD (C) of GluN1a/GluN2Bcrystx
NMDA receptors viewed from top (left panels) and side (right panels). Regions at the
dimer-of-dimers interfaces are highlighted and residues in close proximity at ATD and
LBD are shown as sticks on the side view. Side view of LBD (panel B, right) is one
(between GluN1a (α) and GluN2B (α)) of the two equivalent sites of dimer-of-dimers
interaction at LBD.
18
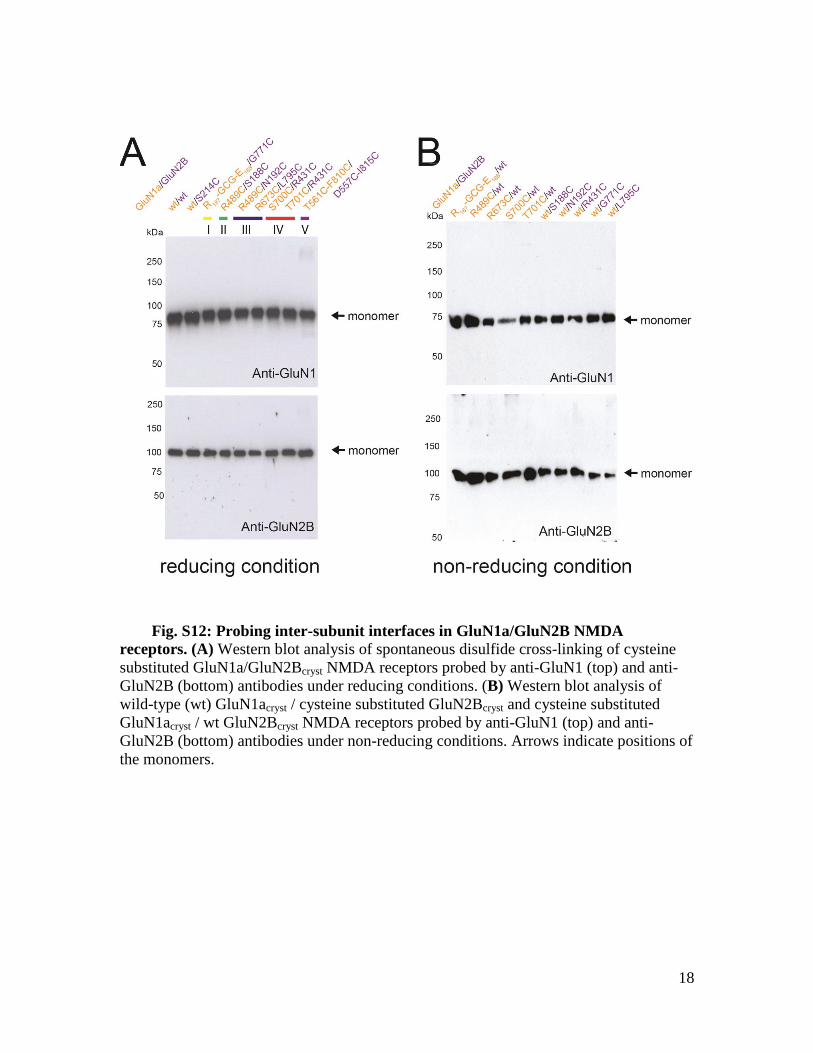
Fig. S12: Probing inter-subunit interfaces in GluN1a/GluN2B NMDA
receptors. (A) Western blot analysis of spontaneous disulfide cross-linking of cysteine
substituted GluN1a/GluN2Bcryst NMDA receptors probed by anti-GluN1 (top) and anti-
GluN2B (bottom) antibodies under reducing conditions. (B) Western blot analysis of
wild-type (wt) GluN1acryst / cysteine substituted GluN2Bcryst and cysteine substituted
GluN1acryst / wt GluN2Bcryst NMDA receptors probed by anti-GluN1 (top) and anti-
GluN2B (bottom) antibodies under non-reducing conditions. Arrows indicate positions of
the monomers.
19
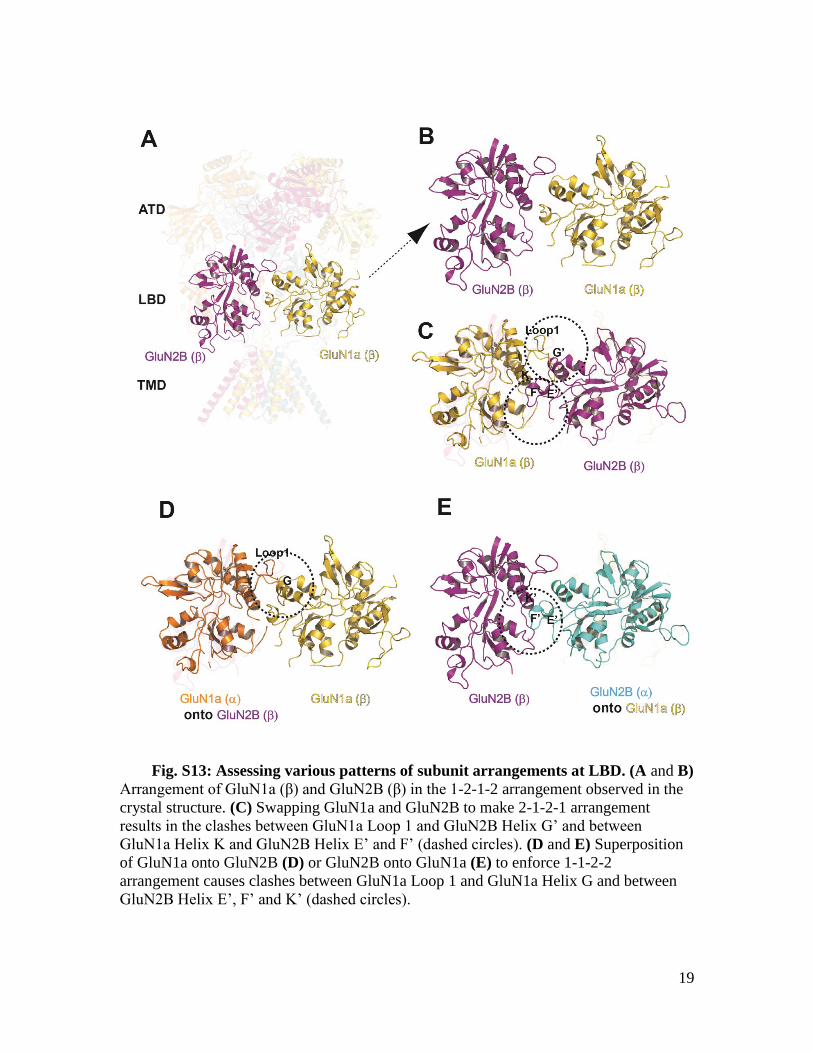
Fig. S13: Assessing various patterns of subunit arrangements at LBD. (A and B)
Arrangement of GluN1a (β) and GluN2B (β) in the 1-2-1-2 arrangement observed in the
crystal structure. (C) Swapping GluN1a and GluN2B to make 2-1-2-1 arrangement
results in the clashes between GluN1a Loop 1 and GluN2B Helix G’ and between
GluN1a Helix K and GluN2B Helix E’ and F’ (dashed circles). (D and E) Superposition
of GluN1a onto GluN2B (D) or GluN2B onto GluN1a (E) to enforce 1-1-2-2
arrangement causes clashes between GluN1a Loop 1 and GluN1a Helix G and between
GluN2B Helix E’, F’ and K’ (dashed circles).
20
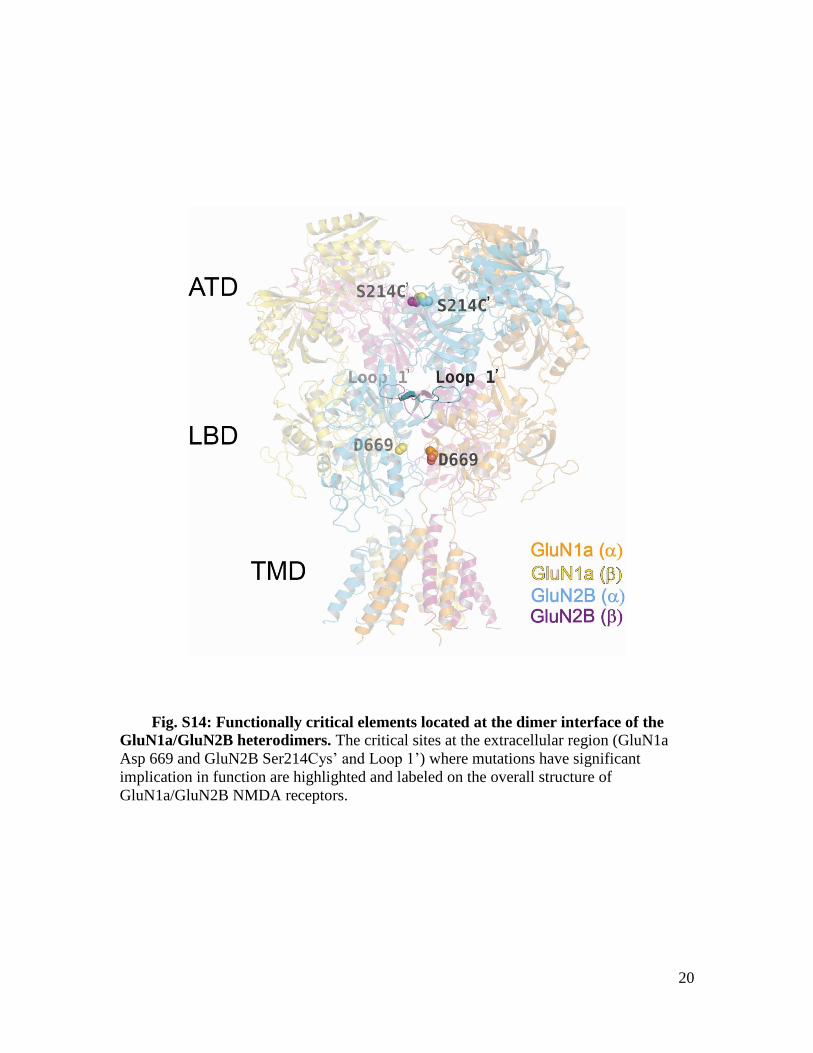
Fig. S14: Functionally critical elements located at the dimer interface of the
GluN1a/GluN2B heterodimers. The critical sites at the extracellular region (GluN1a
Asp 669 and GluN2B Ser214Cys’ and Loop 1’) where mutations have significant
implication in function are highlighted and labeled on the overall structure of
GluN1a/GluN2B NMDA receptors.
21
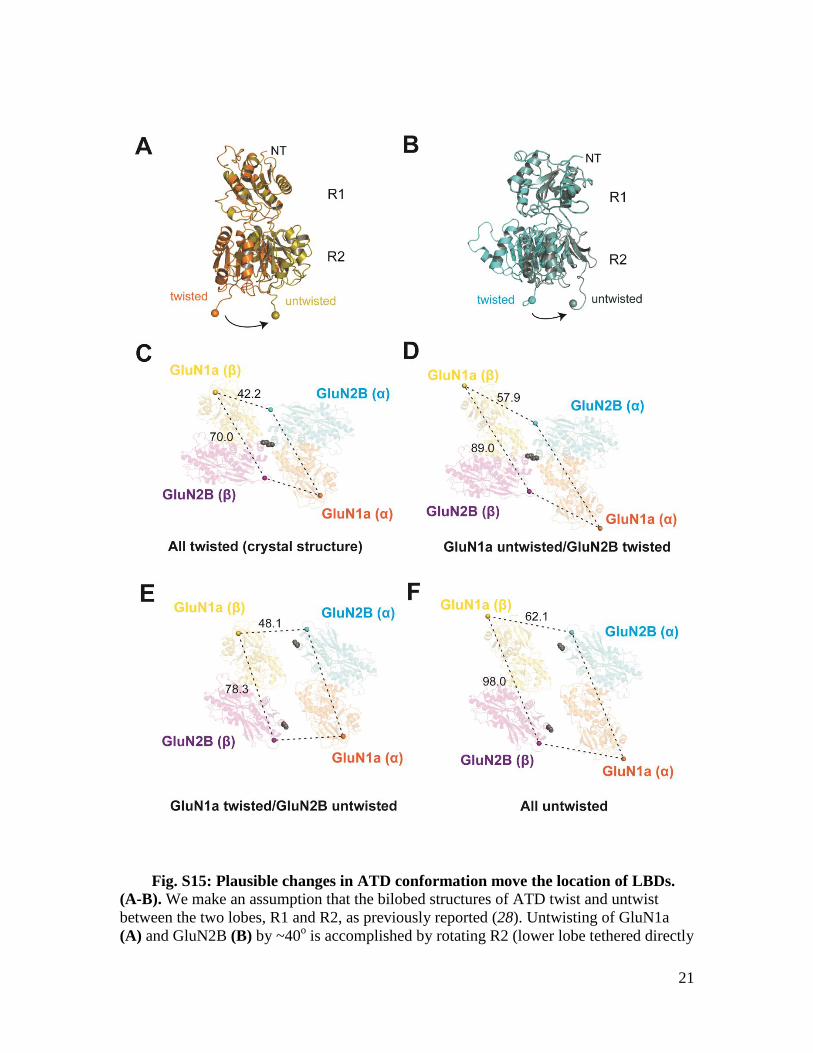
Fig. S15: Plausible changes in ATD conformation move the location of LBDs.
(A-B). We make an assumption that the bilobed structures of ATD twist and untwist
between the two lobes, R1 and R2, as previously reported (28). Untwisting of GluN1a
(A) and GluN2B (B) by ~40o is accomplished by rotating R2 (lower lobe tethered directly
22
to LBD) using GluK1 ATD structure (30) as a guide. (C to F) Shown here are structures
of ATD tetramers viewed “up” from the LBDs. The ATDs in the current crystal structure
and the previously published GluN1b/GluN2B ATD structures have “twisted”
conformations in both GluN1 and GluN2B (C). Untwisting of GluN1a (D), GluN2B (E),
or both (F) dramatically changes the location of LBDs. Numbers next to the dashed lines
represent distance (in Å) between the beginning points of LBD defined as the Cαs of
GluN1a Thr396 and GluN2B His405 (spheres). Note that untwisting of both GluN1a and
GluN2B ATDs result in separation of the LBDs in both length and width to significantly
more extent in GluN1a than GluN2B. This separation likely results in rearrangement of
subunits within and between the GluN1a/GluN2B LBD thereby affecting patterns of ion
channel gating. Shown in gray spheres are GluN2B Ser214 residues from both GluN2B
(α) and (β). Formation of a disulfide bond by the GluN2B Ser214Cys mutation at the
lower lobe (R2) not only traps the inter-subunit arrangement between the two GluN2B
subunits, but also movement in the GluN2B ATD lobes.
23
Fig. S16: Structural comparison of NMDA receptor ion channel with potassium
channels. TMDs of GluN1a subunits (yellow, left panel) and GluN2B subunits (cyan,
middle panel) are superposed onto the ion channel domains (red) of the closed
conformation of KcsA (PDB ID: 1K4C) (A), open conformation of MthK (PDB ID:
3LDC) (B). The superposed structures are viewed from the side (left and middle panels)
or from the extracellular side (right panel). Superposition is performed using Secondary-
structure matching (SSM) tool. Loops are excluded from the figure for clarity.
24
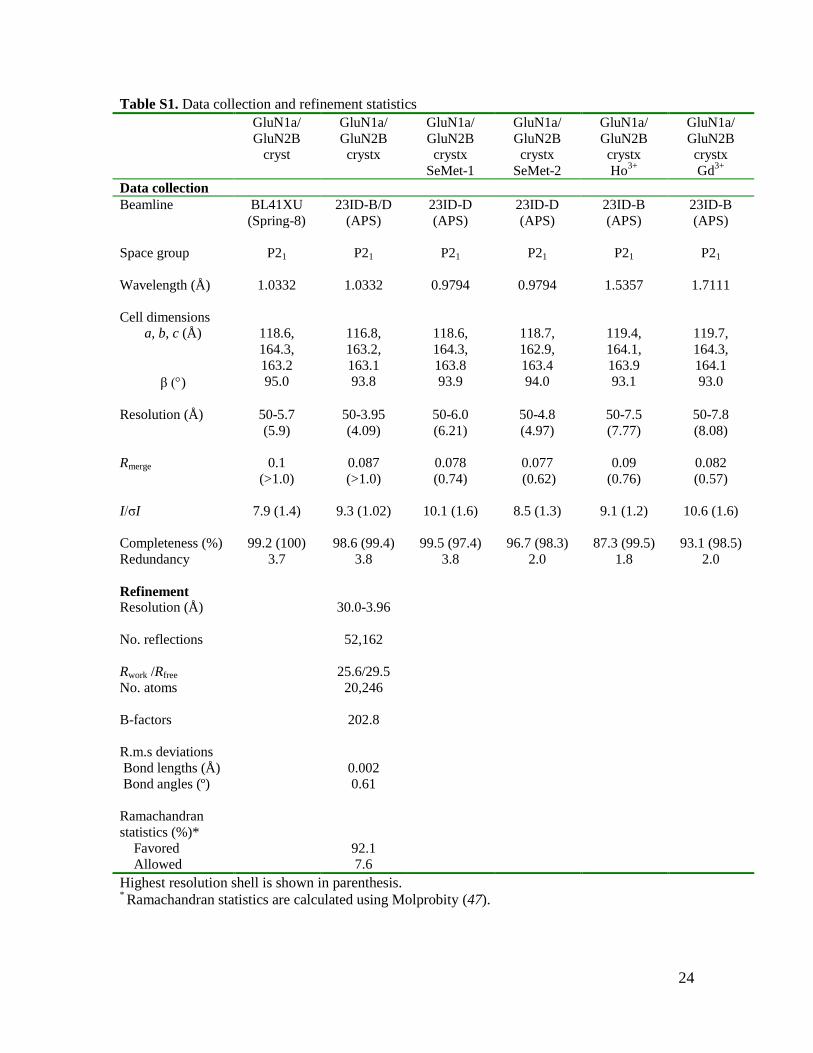
Table S1. Data collection and refinement statistics
Highest resolution shell is shown in parenthesis. * Ramachandran statistics are calculated using Molprobity (47).
25
References and Notes 1. T. Hayashi, Effects of sodium glutamate on the nervous system. Keio J. Med. 3, 183–192
(1954). doi:10.2302/kjm.3.183
2. M. L. Mayer, G. L. Westbrook, P. B. Guthrie, Voltage-dependent block by Mg2+ of NMDA responses in spinal cord neurones. Nature 309, 261–263 (1984). Medline doi:10.1038/309261a0
3. S. F. Traynelis, L. P. Wollmuth, C. J. McBain, F. S. Menniti, K. M. Vance, K. K. Ogden, K. B. Hansen, H. Yuan, S. J. Myers, R. Dingledine, Glutamate receptor ion channels: structure, regulation, and function. Pharmacol. Rev. 62, 405–496 (2010). Medline doi:10.1124/pr.109.002451
4. M. Gielen, B. Siegler Retchless, L. Mony, J. W. Johnson, P. Paoletti, Mechanism of differential control of NMDA receptor activity by NR2 subunits. Nature 459, 703–707 (2009). Medline doi:10.1038/nature07993
5. H. Yuan, K. B. Hansen, K. M. Vance, K. K. Ogden, S. F. Traynelis, Control of NMDA receptor function by the NR2 subunit amino-terminal domain. J. Neurosci. 29, 12045–12058 (2009). Medline doi:10.1523/JNEUROSCI.1365-09.2009
6. K. B. Hansen, H. Furukawa, S. F. Traynelis, Control of assembly and function of glutamate receptors by the amino-terminal domain. Mol. Pharmacol. 78, 535–549 (2010). Medline doi:10.1124/mol.110.067157
7. E. Karakas, N. Simorowski, H. Furukawa, Structure of the zinc-bound amino-terminal domain of the NMDA receptor NR2B subunit. EMBO J. 28, 3910–3920 (2009). Medline doi:10.1038/emboj.2009.338
8. E. Karakas, N. Simorowski, H. Furukawa, Subunit arrangement and phenylethanolamine binding in GluN1/GluN2B NMDA receptors. Nature 475, 249–253 (2011). Medline doi:10.1038/nature10180
9. L. Mony, S. Zhu, S. Carvalho, P. Paoletti, Molecular basis of positive allosteric modulation of GluN2B NMDA receptors by polyamines. EMBO J. 30, 3134–3146 (2011). Medline doi:10.1038/emboj.2011.203
10. A. I. Sobolevsky, M. P. Rosconi, E. Gouaux, X-ray structure, symmetry and mechanism of an AMPA-subtype glutamate receptor. Nature 462, 745–756 (2009). Medline doi:10.1038/nature08624
11. A. N. Farina, K. Y. Blain, T. Maruo, W. Kwiatkowski, S. Choe, T. Nakagawa, Separation of domain contacts is required for heterotetrameric assembly of functional NMDA receptors. J. Neurosci. 31, 3565–3579 (2011). Medline doi:10.1523/JNEUROSCI.6041-10.2011
12. H. Furukawa, S. K. Singh, R. Mancusso, E. Gouaux, Subunit arrangement and function in NMDA receptors. Nature 438, 185–192 (2005). Medline doi:10.1038/nature04089
13. K. M. Vance, N. Simorowski, S. F. Traynelis, H. Furukawa, Ligand-specific deactivation time course of GluN1/GluN2D NMDA receptors. Nat. Commun. 2, 294 (2011). Medline doi:10.1038/ncomms1295
26
14. Y. Yao, C. B. Harrison, P. L. Freddolino, K. Schulten, M. L. Mayer, Molecular mechanism of ligand recognition by NR3 subtype glutamate receptors. EMBO J. 27, 2158–2170 (2008). Medline doi:10.1038/emboj.2008.140
15. A. Jespersen, N. Tajima, G. Fernandez-Cuervo, E. C. Garnier-Amblard, H. Furukawa, Structural insights into competitive antagonism in NMDA receptors. Neuron 81, 366–378 (2014). Medline doi:10.1016/j.neuron.2013.11.033
16. Single-letter abbreviations for the amino acid residues are as follows: A, Ala; C, Cys; D, Asp; E, Glu; F, Phe; G, Gly; H, His; I, Ile; K, Lys; L, Leu; M, Met; N, Asn; P, Pro; Q, Gln; R, Arg; S, Ser; T, Thr; V, Val; W, Trp; and Y, Tyr[BPO1].
17. G. N. Murshudov, P. Skubák, A. A. Lebedev, N. S. Pannu, R. A. Steiner, R. A. Nicholls, M. D. Winn, F. Long, A. A. Vagin, REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. D Biol. Crystallogr. 67, 355–367 (2011). Medline doi:10.1107/S0907444911001314
18. G. F. Schröder, M. Levitt, A. T. Brunger, Super-resolution biomolecular crystallography with low-resolution data. Nature 464, 1218–1222 (2010). Medline doi:10.1038/nature08892
19. C. L. Salussolia, M. L. Prodromou, P. Borker, L. P. Wollmuth, Arrangement of subunits in functional NMDA receptors. J. Neurosci. 31, 11295–11304 (2011). Medline doi:10.1523/JNEUROSCI.5612-10.2011
20. M. Riou, D. Stroebel, J. M. Edwardson, P. Paoletti, An alternating GluN1-2-1-2 subunit arrangement in mature NMDA receptors. PLoS ONE 7, e35134 (2012). Medline doi:10.1371/journal.pone.0035134
21. U. Das, J. Kumar, M. L. Mayer, A. J. Plested, Domain organization and function in GluK2 subtype kainate receptors. Proc. Natl. Acad. Sci. U.S.A. 107, 8463–8468 (2010). Medline doi:10.1073/pnas.1000838107
22. J. Rachline, F. Perin-Dureau, A. Le Goff, J. Neyton, P. Paoletti, The micromolar zinc-binding domain on the NMDA receptor subunit NR2B. J. Neurosci. 25, 308–317 (2005). Medline doi:10.1523/JNEUROSCI.3967-04.2005
23. M. P. Regalado, A. Villarroel, J. Lerma, Intersubunit cooperativity in the NMDA receptor. Neuron 32, 1085–1096 (2001). Medline doi:10.1016/S0896-6273(01)00539-6
24. K. Kashiwagi, J. Fukuchi, J. Chao, K. Igarashi, K. Williams, An aspartate residue in the extracellular loop of the N-methyl-D-aspartate receptor controls sensitivity to spermine and protons. Mol. Pharmacol. 49, 1131–1141 (1996). Medline
25. A. Y. Lau, H. Salazar, L. Blachowicz, V. Ghisi, A. J. Plested, B. Roux, A conformational intermediate in glutamate receptor activation. Neuron 79, 492–503 (2013). Medline doi:10.1016/j.neuron.2013.06.003
26. F. Zheng, K. Erreger, C. M. Low, T. Banke, C. J. Lee, P. J. Conn, S. F. Traynelis, Allosteric interaction between the amino terminal domain and the ligand binding domain of NR2A. Nat. Neurosci. 4, 894–901 (2001). Medline doi:10.1038/nn0901-894
27. K. M. Vance, K. B. Hansen, S. F. Traynelis, GluN1 splice variant control of GluN1/GluN2D NMDA receptors. J. Physiol. 590, 3857–3875 (2012). Medline doi:10.1113/jphysiol.2012.234062
27
28. S. F. Traynelis, M. Hartley, S. F. Heinemann, Control of proton sensitivity of the NMDA receptor by RNA splicing and polyamines. Science 268, 873–876 (1995). Medline doi:10.1126/science.7754371
29. A. Inanobe, H. Furukawa, E. Gouaux, Mechanism of partial agonist action at the NR1 subunit of NMDA receptors. Neuron 47, 71–84 (2005). Medline doi:10.1016/j.neuron.2005.05.022
30. J. Zuo, P. L. De Jager, K. A. Takahashi, W. Jiang, D. J. Linden, N. Heintz, Neurodegeneration in Lurcher mice caused by mutation in delta2 glutamate receptor gene. Nature 388, 769–773 (1997). Medline doi:10.1038/42009
31. Y. Zhou, J. H. Morais-Cabral, A. Kaufman, R. MacKinnon, Chemistry of ion coordination and hydration revealed by a K+ channel-Fab complex at 2.0 A resolution. Nature 414, 43–48 (2001). Medline doi:10.1038/35102009
32. S. B. Long, X. Tao, E. B. Campbell, R. MacKinnon, Atomic structure of a voltage-dependent K+ channel in a lipid membrane-like environment. Nature 450, 376–382 (2007). Medline doi:10.1038/nature06265
33. S. Ye, Y. Li, Y. Jiang, Novel insights into K+ selectivity from high-resolution structures of an open K+ channel pore. Nat. Struct. Mol. Biol. 17, 1019–1023 (2010). Medline doi:10.1038/nsmb.1865
34. W. Li, R. W. Aldrich, Activation of the SK potassium channel-calmodulin complex by nanomolar concentrations of terbium. Proc. Natl. Acad. Sci. U.S.A. 106, 1075–1080 (2009). Medline doi:10.1073/pnas.0812008106
35. R. C. Liddington, Y. Yan, J. Moulai, R. Sahli, T. L. Benjamin, S. C. Harrison, Structure of simian virus 40 at 3.8-A resolution. Nature 354, 278–284 (1991). Medline doi:10.1038/354278a0
36. J. Watanabe, C. Beck, T. Kuner, L. S. Premkumar, L. P. Wollmuth, DRPEER: A motif in the extracellular vestibule conferring high Ca2+ flux rates in NMDA receptor channels. J. Neurosci. 22, 10209–10216 (2002). Medline
37. K. Imoto, C. Busch, B. Sakmann, M. Mishina, T. Konno, J. Nakai, H. Bujo, Y. Mori, K. Fukuda, S. Numa, Rings of negatively charged amino acids determine the acetylcholine receptor channel conductance. Nature 335, 645–648 (1988). Medline doi:10.1038/335645a0
38. T. Kawate, J. C. Michel, W. T. Birdsong, E. Gouaux, Crystal structure of the ATP-gated P2X(4) ion channel in the closed state. Nature 460, 592–598 (2009). Medline doi:10.1038/nature08198
39. B. Siegler Retchless, W. Gao, J. W. Johnson, A single GluN2 subunit residue controls NMDA receptor channel properties via intersubunit interaction. Nat. Neurosci. 15, 406–413, S1–S2 (2012). Medline doi:10.1038/nn.3025
40. D. J. Fitzgerald, P. Berger, C. Schaffitzel, K. Yamada, T. J. Richmond, I. Berger, Protein complex expression by using multigene baculoviral vectors. Nat. Methods 3, 1021–1032 (2006). Medline doi:10.1038/nmeth983
28
41. Z. Otwinowski, W. Minor, Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 276, 307–326 (1997). doi:10.1016/S0076-6879(97)76066-X
42. A. J. McCoy, R. W. Grosse-Kunstleve, P. D. Adams, M. D. Winn, L. C. Storoni, R. J. Read, Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007). Medline doi:10.1107/S0021889807021206
43. P. Emsley, K. Cowtan, Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126–2132 (2004). Medline doi:10.1107/S0907444904019158
44. R. A. Nicholls, F. Long, G. N. Murshudov, Low-resolution refinement tools in REFMAC5. Acta Crystallogr. D Biol. Crystallogr. 68, 404–417 (2012). Medline doi:10.1107/S090744491105606X
45. P. D. Adams, P. V. Afonine, G. Bunkóczi, V. B. Chen, I. W. Davis, N. Echols, J. J. Headd, L. W. Hung, G. J. Kapral, R. W. Grosse-Kunstleve, A. J. McCoy, N. W. Moriarty, R. Oeffner, R. J. Read, D. C. Richardson, J. S. Richardson, T. C. Terwilliger, P. H. Zwart, PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystallogr. 66, 213–221 (2010). Medline doi:10.1107/S0907444909052925
46. I. W. Davis, L. W. Murray, J. S. Richardson, D. C. Richardson, MOLPROBITY: Structure validation and all-atom contact analysis for nucleic acids and their complexes. Nucleic Acids Res. 32 (Web Server), W615–W619 (2004). Medline doi:10.1093/nar/gkh398