HABITAT USE BY SYMPATRIC MALLARD ANASPLATYRHYNCHOS AND AMERICAN BLACK DUCK ANASRUBRIPES BROODS IN A FORESTED AREA OF QUEBEC,
CANADA
SUZANNE CARRIERE' and RODGER D TITMAN
Department of Natural Resource Sciences, Macdonald Campus of McGi11University
21,1 I I Lakeshore Road, Ste-Anne-de-Bellevue, Quebec H9X 3V9
I Present address: Department of Resources, Wildlife and Economic Development,
Government of the Northwest Territories, Yellowknife, NT, Canada
Habitat use by mallard Anas platlJfhynchos and American black duck Anas rubripesbroods was studied in AbitiVL Quebec, on four shared wetlands in June-July 1988 and1989. Average water levels were c. 0.5 m lower in 1988 than in 1989, changing theproportions of habitats available. Black duck broods used scrub-shrub, emergent andaquatic bed wetlands in 1988, whereas mallards used emergent and aquatic bed wet landsalmost exclusively. In 1989 before plant emergence occurred (prior to 25 June), both duckspecies used habitats according to their availability. After this, scrub-shrub wet landswere avoided by both species, and mallard preferred aquatic bed wetlands. The patternof use by mallard broods changed from 1988 to 1989 (from emergent wetlands to moreavailable scrub-shrub wetlands with emergents) but not for black duck. Forested scrub-shrub wet lands were used by black duck even during low availability in 1988 suggestingthat this species found dense cover desirable. The arrCllJof habitats most used by broodsof both species overlapped greatly in this study, but the extent of this overlap fluctuatedwith changes in water level and habitat availability.
Keywords: Sympatric Mallard, American Black Duck, Brood, Habitat
Introduction
Following a decline in numbers beginning in thelate 1950s, the status of the American BlackDuck Anas rubripes has been a matter ofincreasing concern for wildlife managers.Simultaneously, the distribution of Mallard Anasplatyrhynchos in North America has increasedeastward so that breeding and wintering areasof Black Duck and Mallard now overlap largely(Rusch et al. 1989). In the Clay Belt zone ofnorth-central Ontario, Ross & Fillman (1990)observed diminishing numbers of Black Duck
from 1973 to 1982-83, while Mallard numbersremained relatively constant. In the Clay Beltzone of Quebec, data from banding activitiesbetween 1965 and 1971 show an increasingproportion of Mallard captured relative toBlack Duck (Ministere du Loisir, de la Chasse etde la Pikhe du Quebec, Rouyn-Noranda,unpubl. data). Farther east and north in theboreal forest of central Quebec, Bordage(1988) noted similar changes in Black Duck andMallard numbers from aerial surveysperformed between 1985 and I987. Althoughthe highest concentrations of breeding Mallard
WILDFOWL (1998) 49: 150-160
HABITAT USE BY MALLARD AND AMERICAN BLACK DUCK BROODS 151
in North America occur in the prairie potholeregion (Bellrose 1979), Mallard also breedsuccessfully in forested regions (Gilmer et al.1975, Bellrose 1979, Kirby et al. 1985, Monda &Ratti 1988). However, there are few studies ofhabitat use by Mallard breeding in forestedareas (Gilmer et al. 1975, Merendino et al.1993).
Most Black Duck breed in the boreal forest(Bellrose 1979, Bordage 1988). Black Duckbroods typically use streams (Seymour 1984,Bordage 1988) and flooded thicket areas withemergent vegetation, which are pondsfrequently created by beaver Castor canadensis(Renouf 1972, Ringelman & Longcore 1982).
Studies of allopatric populations indicate thathabitat use by breeding Mallard and Black Duckmay overlap at landscape and wetland scales.Little is known, however, of the relative habitatpreferences of sympatric Mallard and BlackDuck (see Coulter & Miller 1968, Laperle 1974,Courcelles & Bedard 1979, Merendino et al.1993, Merendino & Ankney 1994).
Studies of relative habitat preference mayhelp us to understand the shift in Mallard andBlack Duck numbers in areas where they arecurrently sympatric. Brodsky et al. (1989)demonstrated that Mallard and Black Duckpreferred to mate with the phenotype withwhich they were raised. Sympatry may increasethe likelihood of interspecific pair formationdue to close contact or mixing of broods.Interspecific interactions of broods have ahigher probability of occurring if both species(colour morphs) use similar habitats at thewetland scale in the area of sympatry.Furthermore, management for habitatspreferred by Black Duck but less used byMallard may prove to be a means to maintainBlack Duck populations where they overlapwith Mallard.
The objective of this study was to examinehabitat use by brood-rearing Mallard and BlackDuck on shared wetlands in the Clay Belt zoneof north-western Quebec, and to enhance ourunderstanding of how increased interactionwith Mallard may have influenced Black Duckpopulations.
Study Site
This study was conducted on an upper sectionof the Harricana River and one tributary, theLaine River, situated 20 km northwest of Vald'Or, Quebec (48' 03'N, 77' 47'W). This regionlies on the extensive clay deposits of thesouthern Precambrian Shield (Remick 1969)near the eastern limit of Mallard breedingdistribution. Agriculture, forestry and mininghave reduced the extent of the original borealforest relatively recently in this area (Ministerede l'Energie et des Ressources, Quebec 1985).
The Harricana River is 45-75 m wide andforms floodplain wetlands which extend 300 mon average (max. 910 m) on both sides of theriver. The Laine River is 30 m wide withwetlands extending 160 m on both sides.Channels are 3-5 m deep with banks levellingquickly to form shallow basins that arepermanently flooded. Palustrine wetlandcomplexes modified by beaver are present.Aspen Papulus tremulaides and birch Betulapapyrifera dominate in areas surroundingwetlands. Mixed stands of hardwoods andconifers Picea spp., Abies balsam ea, old andcultivated hay fields Paa spp. and burned (in1984) conifer sites characterised the remainingupland areas.
Methods
Four palustrine wetlands were selected fordetailed observation of unmarked broods; theywere chosen as a representative sample ofhabitats available on the study site (Table I).Wetlands A and C were floodplains of theHarricana River (Figure I). Approximately 14ha(43%) of A was visible from a tree tower 18mhigh. Wetland B was on a section of the LaineRiver of which 21ha (95%) was visible from a15m tree tower. Wetland C was observed froma boat where about 13ha (24%) was visible. Inwetland D, about 16ha (70%) was visible from a7m metal frame tower. Two observers surveyeda wetland each for 3-4 hours per day from lateMay through July during 1988 and 1989. Daylighthours were divided into three observationperiods: morning (sunrise - 10:30), mid-day
152 HABITAT USE BY MALLARD AND AMERICAN BLACK DUCK BROODS
(10:35 - 16:00) and evening 16:05 - sunset). Wevisited wetlands in a different period each dayand systematically sampled the available daylightperiod in three days.
Water level fluctuations were recorded byregularly marking the water level on a bridgesupport and noting the changes from thehighest level observed.
Use-availability Analyses
We used the adjusted G statistic (Wil/iams'correction), which is regarded as moreappropriate for small samples than the Chi-square (Sokal & Rohlf 1981), to test whetherMallard and Black Duck broods used habitats inproportion to availability. When statisticallysignificant differences were detected, we usedthe Bonferroni confidence interval technique(Neu et al. 1974) to determine which habitatswere selected or avoided.
Availability was defined considering the totalsurface area of each habitat type (open waterexcluded) within the 4 observed wetlandscombined (Figure 2). Habitat boundaries
were delineated on I:2,700 maps made fromscale enlargements of I: 15,000 aerialphotographs, showing spatially distinct patchesof vegetation at a minimum size of 0.0 I ha.Each patch was classified based on dominantplant species as: forested, deciduous scrub-shrub, persistent emergent (P), non-persistentemergent (NP) and aquatic bed wetland(Cowardin et al. 1979, see Appendix A).
Use was defined in two ways, each requiringus to make different assumptions. First, wescanned the observed wetland every fiveminutes during the length of the observationperiod and noted the habitat where each broodwas present. This definition (hereafter termed'timed use') enabled us to weight habitat typesaccording to the relative amount of timebroods spent in each. Hens and broods couldswim through an area to reach another fordifferent activity. Thus habitat used duringswimming may not have represented habitatactively selected. Therefore use was recordedonly if the hen or at least one duckling waseither feeding, preening or resting in aparticular habitat. We assumed that hens and
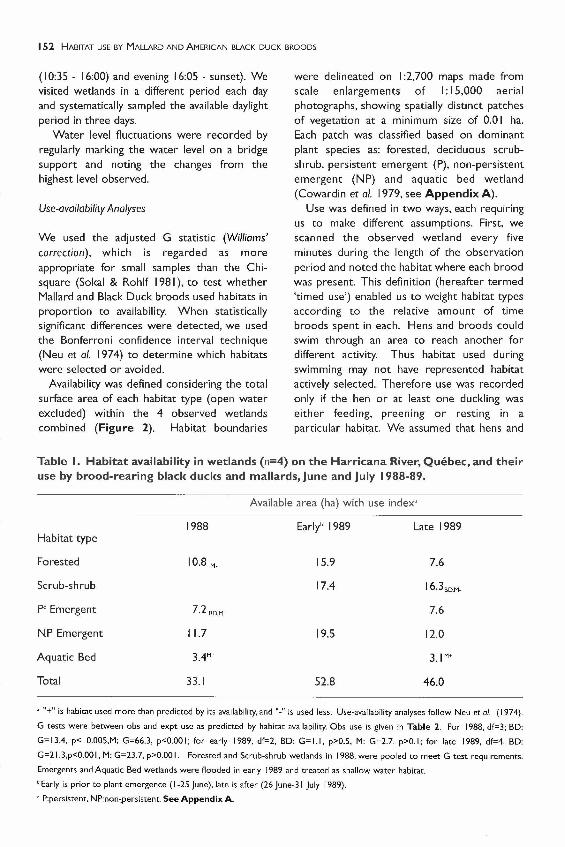
Table I. Habitat availability in wetlands (n=4) on the Harricana River, Quebec, and theiruse by brood-rearing black ducks and mallards, June and July 1988-89.
Available area (ha) with use index'
1988 Early' I989 Late 1989Habitat type
Forested 10.8 M- 15.9 7.6
Scrub-shrub 17.4 16.3BD,M_
P' Emergent 7.2 BD,M- 7.6
NP Emergent 11.7 19.5 12.0
Aquatic Bed 3.4M+ 3.1 M+
Total 33.1 52.8 46.0
, "+" is habitat used more than predicted by its availability, and "-" is used less, Use-availability analyses follow Neu et a/. (1974),
G tests were between obs and expt use as predicted by habitat availability, Obs use is given in Table 2, For 1988. df=3; BD:
G= 13.4, p< O,005.M: G=66,3, p<O,OOI; for early 1989, df=2. BD: G= 1.1. p>O,5, M: G=2.7, p>O, I; for late 1989. df=4, BD:
G=2\ ,3,p<O,OOI, M: G=23.7, p>O,OOI, Forested and Scrub-shrub wetlands in 1988, were pooled to meet G-test requirements,
Emergents and Aquatic Bed wetlands were flooded in early 1989 and treated as shallow water habitat,
"Early is prior to plant emergence (1-25 June), late is after (26 June-31 July 1989),
, P:persistent, NP:non-persistent, See Appendix A,
HABITAT USE BY MALLARD AND AMERICAN BLACK DUCK BROODS 153
Figure I. Four palustrine wetland complexes along the Harricana and laine Rivers,Quebec, were observed for Mallard and Black Duck broods, in June-July 1988 and 1989.Complexes were similarly composed of forested, scrub-shrub, emergent, and aquatic bed wetlands.Area visible from observation tower or boat:A: 14 ha B:21 ha, C: 13 ha, D: 16 ha. The locations ofobservation towers are marked by squares; the observation boat is marked by a diamond.
broods used habitat i at time t+5 minindependently of their use of the same habitatat time t. This is reasonable because during theminimum time interval (5 min), a brood couldswim across an entire observed wetland anduse all habitat types.
Second, use was recorded as the presence ofa brood in a habitat at least once during anobservation period. For example, a broodcould only have been recorded as using aparticular habitat twice if observed during twoperiods (ie on different days) or if a differenthabitat was used in between during the sameperiod. By collecting use data as presence!absence per brood-period (hereafter presence!absence use), we can abide by a secondassumption required for the application of Gtests; the probability of recording a presence issimilar for Black Duck and Mallard in allhabitats. Visibility of broods of an individualspecies in each habitat was not estimated butwe recognise that it may not have been equal in
each type, probably being poorer in forestedand scrub-shrub wetland than in the habitatswith shorter vegetation. This second usedefinition enabled us to decrease theheterogeneous visibility effect: a broodobserved during 5 min in a habitat producedthe same use value as a brood visible during 30min while using another habitat. We are,however, no longer able to provide an estimateof time spent in each habitat. According to thisdefinition, the first assumption (ofindependence of observations) is againreasonable as the minimum time intervalbetween sightings in the same habitat is 10 min(time necessary for a brood to use habitat i attime t, habitat j at time t + 5 min, and againhabitat i at time t + 10 min). At least 30% of useobservations were obtained from differentbroods (identified by a species-age index[Gollop & Marshall 1954]) or from the samebrood but in different observation periods.
154 HABITATUSEBYMALLARDANDAMERICANBLACKDUCKBROODS
1988!>ID20
18 mC16
DB1412 M+ M+
M- -A10 M- M-86
~
M - BD-::::::::::"::::~' ..4
~2 - ----,0
Aquatic bed NP emergent P emergent Scrub-shrub-Forested-m <25 June 1989.r:.-
~ 20 M - BO- M+18.c 16 M - BO-m 14'co 12
~
> 10
~
m+-' 8m 6~.c 4m 2:r: 0
Aquatic bed NP emergent P emergent Scrub-shrub Forested
>25 June 1989 BO-20 M - BD-18161412108
~~
6420
Aquatic bed NP emergent P emergent Scrub-shrub Forested
Habitat type
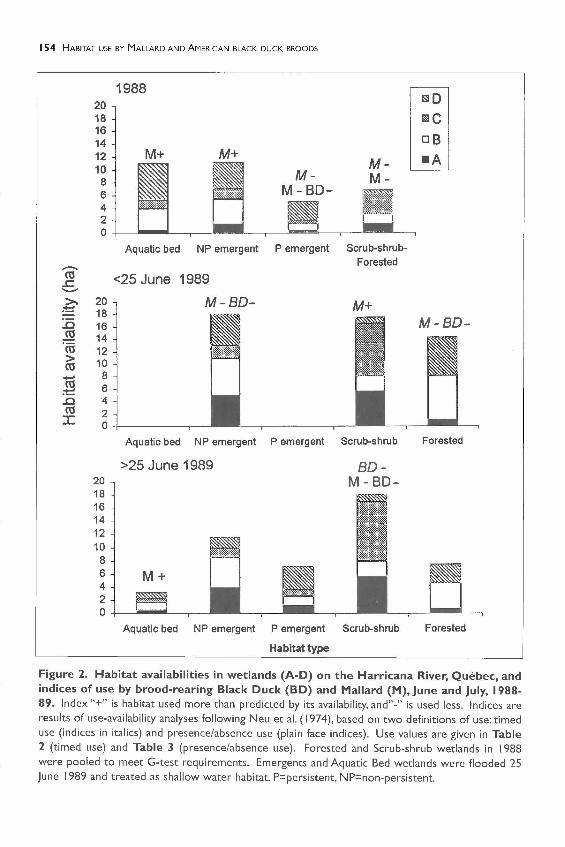
Figure 2. Habitat availabilities in wetlands (A-D) on the Harricana River, Quebec, andindices of use by brood-rearing Black Duck (BD) and Mallard (M), June and July, 1988-89. Index "+" is habitat used more than predicted by its availability, and"-" is used less. Indices areresults of use-availability analyses following Neu et al. (1974), based on two definitions of use: timeduse (indices in italics) and presence/absence use (plain face indices). Use values are given in Table2 (timed use) and Table 3 (presence/absence use). Forested and Scrub-shrub wetlands in 1988were pooled to meet G-test requirements. Emergents and Aquatic Bed wetlands were flooded 25June 1989 and treated as shallow water habitat. P=persistent, NP=non-persistent.
HABITAT USE BY MALLARD AND AMERICAN BLACK DUCK BROODS 155
Black Duck-Mallard use comparisons
The hypothesis that Mallard and Black Duckbroods use habitats differently was tested usingeach use definition with a G statistic. Uponobtaining significant results, we tested all pairsof habitats for significant differences in use byeach species (2x2 G tests, Sokal & Rohlf 1981).We are confident that visibility of Black Duckand Mallard broods in a particular habitat didnot differ, i.e. that the probability of beingsighted was independent of species.
Between year comparisons
Differences in habitat use between 1988 and1989, for both species, were analysed by using asimultaneous test procedure (STP; Sokal &Rohlf 1981:728).
Results
Annual differences in water level causeddifferences of habitat availability between 1988and 1989. Water levels were 0.3 - 0.8 m higherfrom early May to August 1989 compared to1988. Consequently, emergent plants did notappear above the water surface until late June.For this reason, we divided the 1989 data intotwo periods: (I) before and (2) after plantemergence, which occurred around 25 June,and performed separate habitat analyses.When the first broods appeared in early June1988, emergent plants had already formedrelatively dense patches. In 1989, NP-emergenthabitats were not available to early broods,however, some forested and scrub-shrubpatches (which were never flooded in 1988)were available to them until mid-June.
Use vs Availability
In 1988, timed use analyses showed that BlackDuck used all habitats according to theiravailability (G=8.807, p>0.05), whereaspresence/absence use analyses showed thatthey avoided P-emergent habitats, using otherhabitats according to their availability (G= 13.4,p<0.005) (Figure 2). Mallard broods usedsome habitats differently than expected from
their availability (timed use: G= 197.2, p<O.OO I;presence/absence use: G=66.6, p<O.OO I)(Figure 2). Mallard broods selected aquaticbeds and avoided other habitats according totimed use analyses. They either selected NP-emergent wetlands according to timed useanalyses or ignored it according to presence/absence analyses (Figure 2).
In 1989, before plant emergence (25 June),both species selected scrub-shrub habitats andavoided NP-emergents and forested habitatsaccording to timed use analyses (BD: G=48.7,p<0.005; M: G= I03.8, p<0.005). According topresence/absence use analyses, they used allhabitats according to their availability (BD:G= 1.1, p>O.I; M:G=2.7, p>O.I) (Figure 2).
After 25 June 1989, Black Duck broodsavoided scrub-shrub habitats (timed use:G= I7.26, p<0.005; presence/absence use:G=23.7, p<O.OO I) (Figure 2).
Black Duck-Mallard use comparisons
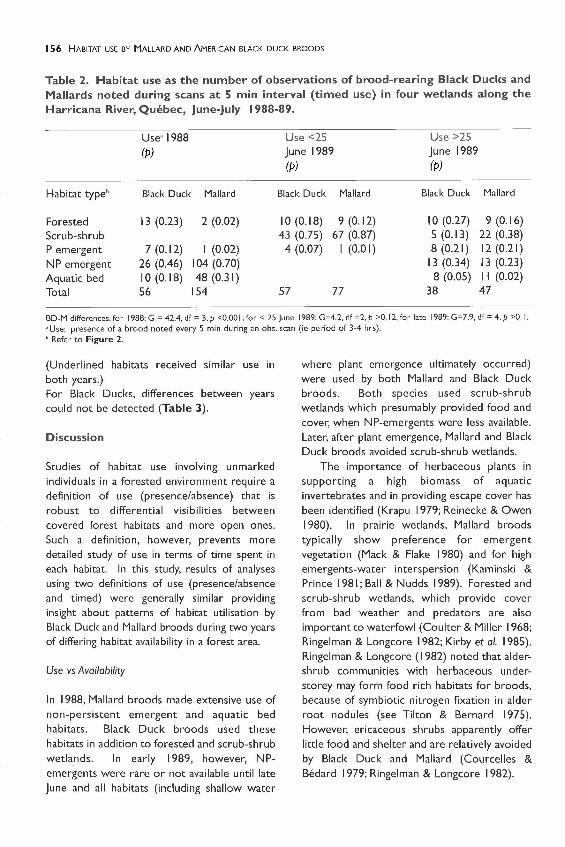
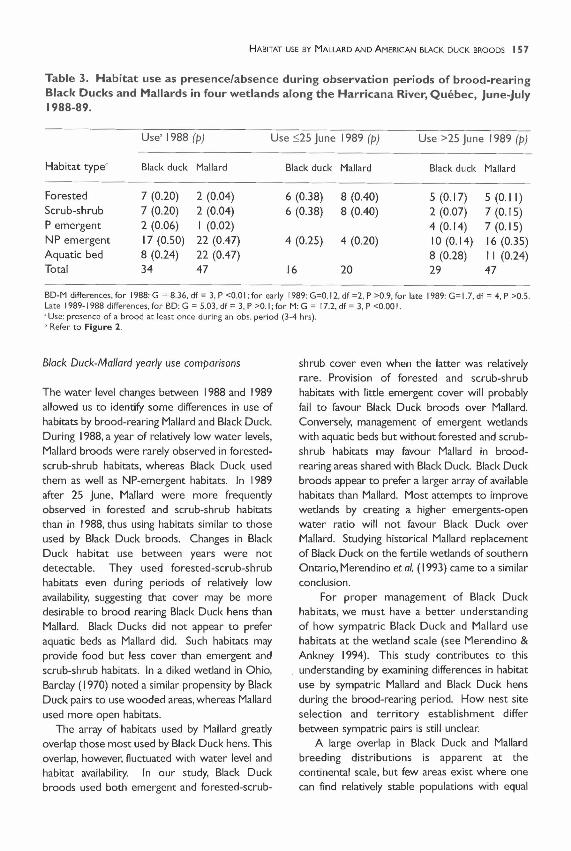
Habitat use by Black Ducks and Mallardsdiffered in 1988 according to both definitions ofuse (Table 2, timed use: G=42.44, p<O.OO I;Table 3, presence/absence use: G=8.36,p<O.OI). Black Duck broods were associatedwith forested scrub-shrub wetlands whileMallards were associated with aquatic bedhabitats (presence/absence use: 2x2 G=7.64,p<0.05). In 1989, both before and after 25 June,habitat use by Mallards and Black Ducks did notdiffer (Tables 2 and 3).
Between year comparisons
Analyses were performed using onlypresence/absence use, which is more robust toyearly differences in visibility and we onlycompared habitat use between 1988 and after25 June 1989, when all possible habitats wereavailable. Mallard broods showed significantdifferences in use patterns (Table 3). STP testsproduced the following array of habitatsordered (from left to right) as those associatedwith after 25 June 1989 to those associatedwith 1988:Forested scrub-shrub P-emer,gentNP-emer,gent Aquatic bed
156 HABITAT USE BY MALLARD AND AMERICAN BLACK DUCK BROODS
Table 2. Habitat use as the number of observations of brood-rearing Black Ducks andMallards noted during scans at 5 min interval (timed use) in four wetlands along theHarricana River, Quebec, June-July 1988-89.
BD-M differences, for 1988: G = 42.4, df = 3. P <0.00 I; for < 25 June 1989: G=4.2, df =2. P >0.12, for late 1989: G=7.9, df = 4. P >0.1 .•Use: presence of a brood noted every 5 min during an obs. scan (ie period of 3-4 hrs)." Refer to Figure 2.
(Underlined habitats received similar use inboth years.)For Black Ducks, differences between yearscould not be detected (Table 3).
Discussion
Studies of habitat use involving unmarkedindividuals in a forested environment require adefinition of use (presence/absence) that isrobust to differential visibilities betweencovered forest habitats and more open ones.Such a definition, however, prevents moredetailed study of use in terms of time spent ineach habitat. In this study, results of analysesusing two definitions of use (presence/absenceand timed) were generally similar providinginsight about patterns of habitat utilisation byBlack Duck and Mallard broods during two yearsof differing habitat availability in a forest area.
Use vs Availability
In 1988, Mallard broods made extensive use ofnon-persistent emergent and aquatic bedhabitats. Black Duck broods used thesehabitats in addition to forested and scrub-shrubwetlands. In early 1989, however, NP-emergents were rare or not available until lateJune and all habitats (including shallow water
where plant emergence ultimately occurred)were used by both Mallard and Black Duckbroods. Both species used scrub-shrubwetlands which presumably provided food andcover, when NP-emergents were less available.Later, after plant emergence, Mallard and BlackDuck broods avoided scrub-shrub wetlands.
The importance of herbaceous plants insupporting a high biomass of aquaticinvertebrates and in providing escape cover hasbeen identified (Krapu 1979; Reinecke & Owen1980). In prairie wetlands, Mallard broodstypically show preference for emergentvegetation (Mack & Flake 1980) and for highemergents-water interspersion (Kaminski &Prince 1981; Ball & Nudds 1989). Forested andscrub-shrub wetlands, which provide coverfrom bad weather and predators are alsoimportant to waterfowl (Coulter & Miller 1968;Ringelman & Longcore 1982; Kirby et al. 1985).Ringelman & Longcore (1982) noted that alder-shrub communities with herbaceous under-storey may form food rich habitats for broods,because of symbiotic nitrogen fixation in alderroot nodules (see Tilton & Bernard 1975).However, ericaceous shrubs apparently offerlittle food and shelter and are relatively avoidedby Black Duck and Mallard (Courcelles &Bedard 1979; Ringelman & Longcore 1982).
HABITAT USE BY MALLARD AND AMERICAN BLACK DUCK BROODS 157
Table 3. Habitat use as presence/absence during observation periods of brood-rearingBlack Ducks and Mallards in four wetlands along the Harricana River, Quebec, June-July1988-89.
Use' 1988 (p) Use ::;25 June 1989 (p) Use >25 June 1989 (p)
BD-M differences, for 1988: G = 8.36, df = 3, P <0.0 I; for early 1989: G=0.12, df =2, P >0.9, for late 1989: G= 1.7, df = 4. P >0.5.Late 1989-1988 differences. (or BD: G = 5.03. d( = 3, P >0.1; for M: G = 17.2. d( = 3, P <0.00 I.'Use: presence of a brood at least once during an obs. period (3·4 hrs).o Refer to Figure 2.
Black Duck-Mallard yearly use comparisons
The water level changes betWeen 1988 and 1989allowed us to identify some differences in use ofhabitats by brood-rearing Mallard and Black Duck.During 1988, a year of relatively low water levels,Mallard broods were rarely observed in forested-scrub-shrub habitats, whereas Black Duck usedthem as well as NP-emergent habitats. In 1989after 25 June, Mallard were more frequentlyobserved in forested and scrub-shrub habitatsthan in 1988, thus using habitats similar to thoseused by Black Duck broods. Changes in BlackDuck habitat use between years were notdetectable. They used forested-scrub-shrubhabitats even during periods of relatively lowavailability, suggesting that cover may be moredesirable to brood rearing Black Duck hens thanMallard. Black Ducks did not appear to preferaquatic beds as Mallard did. Such habitats mayprovide food but less cover than emergent andscrub-shrub habitats. In a diked wetland in Ohio,Barclay (1970) noted a similar propensity by BlackDuck pairs to use wooded areas, whereas Mallardused more open habitats.
The array of habitats used by Mallard greatlyoverlap those most used by Black Duck hens. Thisoverlap, however, fluctuated with water level andhabitat availability. In our stUdy, Black Duckbroods used both emergent and forested-scrub-
shrub cover even when the latter was relativelyrare. Provision of forested and scrub-shrubhabitats with liule emergent cover will probablyfail to favour Black Duck broods over Mallard.Conversely, management of emergent wetlandswith aquatic beds but without forested and scrub-shrub habitats may favour Mallard in brood-rearing areas shared with Black Duck. Black Duckbroods appear to prefer a larger array of availablehabitats than Mallard. Most attempts to improvewetlands by creating a higher emergents-openwater ratio will not favour Black Duck overMallard. Studying historical Mallard replacementof Black Duck on the fertile wetlands of southernOntario, Merendino et al. (1993) came to a similarconclusion.
For proper management of Black Duckhabitats, we must have a better understandingof how sympatric Black Duck and Mallard usehabitats at the wetland scale (see Merendino &Ankney 1994). This stUdy contributes to this
. understanding by examining differences in habitatuse by sympatric Mallard and Black Duck hensduring the brood-rearing period. How nest siteselection and territory establishment differbetWeen sympatric pairs is still unclear.
A large overlap in Black Duck and Mallardbreeding distributions is apparent at thecontinental scale, but few areas exist where onecan find relatively stable populations with equal
158 HABITAT USE BY MALLARD AND AMERICAN BLACK DUCK BROODS
numbers of breeding pairs (see Rusch et al. 1989;Ross & Fillman 1990). Small sample sizes for atleast one species and overall low densities maypreclude conclusive results for habitat preferenceanalyses (eg Barclay 1970). Areas of such overlapare often remote but studies of sympatricbreeding populations are needed, especially wherethe overlap is of recent occurrence such as incentral Quebec (Bordage 1988), whereincidentally, little is known about the possibleeffects of recent major habitat changes upon theBlack Duck population (Rusch et al. 1989).
We are grateful to J.R. Longcore,j.-F. Giroux,J.R. Ringelman and M. T. Merendino for advice andcomments on an earlier draft. We wish to thankQ. van Ginhoven, N. Raiche, S. Gagnon (DucksUnlimited, Rouyn-Noranda), G. Berube (MLCP, Vald'Or), J.-F. and C. Thiboutot and A.B. Carriere fortheir assistance in the field. Financial support wasprovided by FCAR (Quebec), the Natural Sciencesand Engineering Research Council of Canada(grant A9700 to ROT), the Canadian WildlifeFoundation (Orville Erickson Memorial ScholarshiPto SC), the Canadian Wildlife Service UniversityResearch Support Fund, the Alf Kelly Fund of theProvince of Quebec Society for the Preservation of .Birds and la Fondation Heritage-Faune (Quebec).
selection:An experiment and implications formanagement. In: Sharitz, R.R. & Gibbons, J.W.(Eds.) Proceedings of the SymposiumFreshwater wetlands and wildlife, Charleston,South Carolina, March 1986. U.S. Dept. OfEnergy, Office of Scientific and TechnicalInformation. Pp 659-671
Bellrose, F.e. 1979. Species distribution, habitatsand characteristics of breeding dabblingducks in North America. In: Bookhout, T.A.(Ed.). Waterfowl and wetlands: an integratedreview. La Crosse Printing Co., La Crosse,Wisconsin. Pp 1-15
Bordage, D. 1988. Suivi des couples nicheurs decanard noir en foret boreale - 1987. Serie derapports techniques. N. 35. Service canadiende la faune, region du Quebec, Quebec.
Brodsky, L.M., Ankney, CD. & Dennis, D.G.1989. Social experience influencespreferences in Black Ducks and Mallards.Can.J. Zool. 67: 1434-1438.
Coulter, M.W. & Miller, W.R. 1968. Nestingbiology of Black Ducks and Mallards innorthern New England. Vermont Fish andGame Dept. Bulletin 68-2.
Courcelles, R. & Bedard, J. 1979. Habitatselection by dabbling ducks in the Baie Noiremarsh, southwestern Quebec. Can. J. Zool.57:2230-2238.
Cowardin, L.M.Y.,Carter,v., Golet, F.e. & Laroe,E.T. 1979. Classification of wetlands anddeepwater habitats of the United States. USFish and Wildlife Serv., Washington, D.e.
Gilmer, D.S., Ball, I.J., Cowardin, L.M.,Reichmann, J.H. & Tester, J.R. 1975. Habitatuse and home range of Mallards breeding inMinnesota. J. Wildl. Manage. 39:781-789.
Gollop, J.B. & Marshall, W.H. 1954. A guide foraging duck broods in the field. Miss. FlywayCounc. Tech. Sect., Minneapolis, Minn.
Kaminski, R.M. & Prince, H.H. 1981. Dabblingduck and aquatic macroinvertebrateresponses to manipulated wetland habitat. J.Wildl. Manage. 45: 1-15.
Kirby,R.E.,Reichmann,J.H. & Cowardin, L.M.1985.Home range and habitat use of forest dwellingMallards in Minnesota. Wilson Bull. 97:215-219.
Krapu, G.L. 1979. Nutrition of female dabblingducks during reproduction. In: Bookhout,T.A.(Ed.). Waterfowl and wetlands: an integratedreview. La Crosse Printing Co., La Crosse,Wisconsin. Pp 59-70
Laperle, M. 1974. Effects of water levelfluctuations on duck breeding success. In:Boyd, H. (Ed.). Waterfowl studies. Can. Wildl.Serv. Rep. No. 29. Pp 18-30.
Mack, GoO. & Flake, LoO. 1980. Habitatrelationships of waterfowl broods on SouthDakota stock ponds. J. Wildl. Manage. 44:695-700.
HABITAT USE BY MALLARD AND AMERICAN BLACK DUCK BROODS I S9
Marie-Victorin, F. 1964. Flore laurentienne. 2'·Ed. Les Presses de l'Universite de Montreal,Montreal, Quebec.
Merendino, M.T.& Ankney, CD. 1994. Habitat useby Mallards and American Black Ducksbreeding in central Ontario. Condor 96:41 1-421.
Merendino, M.T.,Ankney, CD. & Dennis, D.G.1993. Increasing Mallards, decreasingAmerican Black Ducks: more evidence forcause and effect. J. Wildl. Manage. 57: 199-208.
Ministere de I'Energie et des RessourcesQuebec. 1985. Les regions ecologiques duQuebec meridional; deuxieme approxim.Cartel: 1,250,000. Quebec, Quebec.
Monda, M.J.& Ratti,J.T. 1988. Niche overlap and. habitat use by sympatric duck broods in
eastern Washington. J. Wildl. Manage. 52:95-103.
Neu, Cw., Byers, CR. & Peek, j.M. 1974. Atechnique for analysis of utilisation-availabilitydata. J. Wildl. Manage. 38:541-545.
Reinecke, K.J. & Owen, R.B. Jr. 1980. Food useand nutrition of Black Ducks nesting inMaine. J. Wild/. Manage. 44:549·558.
Remick, J.H. 1969. Geologie de la regiond'Harricana-Turgeon, comtes d'Abitibi-est etd'Abitibi-ouest, Quebec. Ministere desRichesses Naturelles, R. P. 564, Quebec,Quebec.
Renouf, R.N. 1972. Waterfowl utilization ofbeaver ponds in New Brunswick. J. Wildl.Manage. 36:740-744.
Ringelman, J.K. & Longcore, J.R. 1982.Movements and wetland selection by brood·rearing Black Ducks. J. Wildl. Manage. 46:615-621.
Ross, R.K. & Fillman, D. 1990. Distribution ofBlack Ducks and Mallards in northernOntario. Can. Wildl.Serv. Bioi. Notes, No. 189.
Rusch, D.H., Ankney, CD., Boyd, H., Longcore,J.R., Montalbano, Ell!, Ringelman,J.K. & Stotts,VD. 1989. Population ecology and harvest ofthe American Black Duck: a review. Wild!. Soc.Bull. 17:379-406.
Seymour, N.R. 1984. Activity of Black Ducksnesting along streams in northeastern NovaScotia. Wildfowl 35: 143-150.
Sokal, R.S. & Rohlf, EJ. 1981. Biometry: Theprinciples and practice of statistics in biologicalresearch. 2nd Ed.W.H. Freeman and Co., NewYork.
Tilton, D.L. & Bernard, J.M. 1975. Primaryproductivity and biomass distribution in analder shrub ecosystem. Amer. MidlandNaturalist 94:251-256.
160 HABITAT USE BY MALLARD AND AMERICAN BLACK DUCK BROODS
Appendix A. Plant species by habitat. Classification follows Cowardin et al. (1979) andtaxonomy follows Marie-Victorin (1964).
Habitat classesCommon name
ForestedBalsam firBirchPoplarSpruce blackTamarack
Scrub-shrubAlderDwarf-birchRed-osier dogwoodWillowBog rosemaryLeatherleafSpiraeaSheep LaurelSwamp LaurelSweet gale
Persistent emergentBlack-girded wool-rushBulrushCommon rushSedgesWater parsnipGrasses etc.
Species name
Abies balsam ea (L.) Mill.Betula papyrifera Marsh.Populus tremuloides Michx.Picea mariana (Mill.) BSP.Larix laricina (Du Roi) K. Koch.