Page 1
ORIGINAL PAPER
Systemic overexpression of antizyme 1 in mouse reducesornithine decarboxylase activity without major changesin tissue polyamine homeostasis
Marko Pietila • Hiramani Dhungana •
Anne Uimari • Reijo Sironen • Leena Alhonen
Received: 29 July 2013 / Accepted: 21 October 2013 / Published online: 31 October 2013
� Springer Science+Business Media Dordrecht 2013
Abstract Polyamines, spermidine, spermine and
their precursor putrescine, are ubiquitous cell compo-
nents essential for normal cell growth. Increased
polyamine levels and enhanced biosynthesis have been
associated with malignant transformation and tumor
formation, and thus, the polyamines have been con-
sidered to be a meaningful target to cancer therapies.
However, clinical cancer treatment trials using inhib-
itors of polyamine synthesis have been unsuccessful
probably due to compensatory uptake of polyamines
from extracellular sources. The antizyme proteins
regulate both polyamine biosynthesis and transport,
and thus, the antizymes could provide an efficient
approach to control cellular proliferation compared to
the mere inhibition of biosynthesis. To define the role
of antizymes in proliferative processes associated with
the whole animal, we have generated transgenic mice
overexpressing mouse antizyme 1 gene under its own
regulatory sequences. Antizyme 1 protein was abun-
dantly expressed in various organs and the expressed
antizyme protein was functional as ornithine decar-
boxylase activity was significantly reduced in all
tissues analyzed. However, antizyme 1 overexpression
caused only minor changes in tissue polyamine levels
demonstrating the challenges in using the ‘‘antizyme
approach’’ to deplete polyamines in a living animal.
Neither were there any changes in cellular proliferation
in the proliferative tissues of transgenic animals.
Interestingly though, there was occurrence of abnor-
mally high level of apoptosis in the non-proliferating
part of the colon epithelia. Otherwise, the transgenic
founder mice appeared healthy and out of seven
founders six were fertile. However, none of the
founders could transmit the transgene suggesting that
the antizyme 1 overexpression may be deleterious to
transgenic gametes.
Keywords Antizyme � Transgenic � Ornithine
decarboxylase � Polyamine � Proliferation
Introduction
Polyamines spermidine and spermine and their dia-
mine precursor, putrescine, are known to be essential
for cell growth and differentiation. Polyamines are
M. Pietila (&)
Institute of Biomedicine, University of Eastern Finland,
P.O.B 1627, 70211 Kuopio, Finland
e-mail: [email protected]
H. Dhungana � A. Uimari � L. Alhonen
Department of Biotechnology and Molecular Medicine,
A. I. Virtanen Institute for Molecular Sciences, Biocenter
Kuopio, University of Eastern Finland, Kuopio, Finland
R. Sironen
Institute of Clinical Medicine, Pathology and Forensic
Medicine, Cancer Center of Eastern Finland, University of
Eastern Finland, Kuopio, Finland
R. Sironen
Department of Pathology, Imaging Center, Kuopio
University Hospital, Kuopio, Finland
123
Transgenic Res (2014) 23:153–163
DOI 10.1007/s11248-013-9763-y
Page 2
positively charged at physiologically relevant ionic
and pH conditions and readily bind to negatively
charged macromolecules such as RNA, DNA and
proteins. In cells the polyamines are present in
millimolar concentrations and have been implicated
in many physiological functions including DNA
replication, transcription, translation, posttranslational
protein modification and membrane stability. Some of
these effects are specific for polyamines, while others
are less specific due to the general cationic nature of
these compounds. Irrespective of the specificity of their
effects, polyamines are indispensable cellular compo-
nents because their depletion, either by gene disruption
or inhibitors of their biosynthesis, results in severe
defects in cellular growth (Wallace and Fraser 2004).
The polyamine metabolism has long been an
attractive target for therapy of cancer (Kaiser et al.
2003; Casero et al. 2005) and parasitic diseases (Heby
et al. 2007). Ornithine decarboxylase (ODC) is a key
enzyme in polyamine synthesis and its overexpression
has been associated with cell transformation and
tumor formation. Although ODC is under tight control
in normal cells, its regulation is altered in neoplastic
cells, yielding constitutively high levels of ODC
expression and activity (Shantz and Levin 2007).
Furthermore, a polymorphism in ODC gene associates
with colon adenoma recurrence and the survival of
colon cancer patients (Martinez et al. 2003). Recently,
the inhibition of ODC has been successfully used in
the prevention of basal cell carcinoma of skin (Bailey
et al. 2010) and with the combination of anti-
inflammatory drug sulindac to prevent sporadic colo-
rectal adenomas (Meyskens et al. 2008). In experi-
mental animal studies, inhibition of ODC with its
specific inhibitor a-difluoromethylornithine (DFMO)
has proved to be effective in prevention of prostate
adenocarcinoma. In a study with transgenic adeno-
carcinoma model of mouse prostate (TRAMP),
DFMO treatment decreased malignant prostate growth
and totally prevented the formation of metastases,
which normally arise in these mice upon aging (Gupta
et al. 2000).
Despite these promising results, clinical trials to
treat advanced cancer with inhibitors of polyamine
synthesis have been unsuccessful due to compensatory
uptake of polyamines from extracellular sources and
complex regulation of polyamine metabolism. Even
during drastic changes in the cellular environment,
polyamine levels are maintained within a narrow
optimal range by a complex regulatory circuitry. For
example, the activity of ODC is controlled at the levels
of transcription, translation, and the stability of the
enzyme protein (Perez-Leal and Merali 2012). The
latter is regulated by the antizymes (AZ), and further
by antizyme inhibitor (AZIn) (Kahana et al. 2005).
The expression of AZ is controlled by intracellular
polyamine level whereby increased polyamine level
triggers ?1 ribosomal frameshift on the encoding AZ
mRNA, allowing functional AZ to be expressed
(Matsufuji et al. 1995). From three known mammalian
antizymes, the antizyme 1 (AZ1) is the best known and
most ubiquitously expressed. The AZ1 mRNA con-
tains two potential start codons near the 50 end and
produces a functional protein of either 29 kDa or
24 kDa depending on the start codon used (Mitchell
et al. 1998). AZ2 is also ubiquitously expressed but to
a lesser extent and is less efficient in targeting ODC to
proteosomal degradation. AZ3 is only expressed in
testis. There are also two antizyme inhibitor proteins
(AZIn) known to date, one ubiquitously expressed
(AZIn1) and the other which is restricted to brain and
testis (AZIn2). AZ decreases polyamine levels
through three mechanisms. First, AZ disrupts active
ODC dimers, resulting in decreased polyamine syn-
thesis. Second, AZ targets ODC for degradation by the
26S proteasome in a reaction that does not require
ubiquitination. Third, AZ inhibits polyamine uptake
from the microenvironment through a mechanism that
has yet to be completely characterized (Kahana 2007).
Taking together, the antizymes provide an approach
for cancer therapy which relies on to regulation of both
the synthesis and the uptake of the polyamines.
AZ1 has been shown to have tumor-suppressive
activity in mouse models. Targeted AZ1 overexpres-
sion to the skin using either the bovine keratin 5 or
keratin 6 promoter showed delayed tumor onset and
decreased tumor numbers relative to wild-type controls
after exposure to the chemical carcinogen DMBA
(7,12-dimethylbenz(a)anthracene) and the tumor pro-
moter TPA (12-O-tetradecanoylphorbol-13-acetate)
(Feith et al. 2001). Similarly, the K6/AZ and K5/AZ
mice were also more resistant to forestomach tumors
induced by N-nitrosomethylbenzylamine (NMBA)
(Fong et al. 2003). In the present study, we used mouse
transgenesis to study the physiological consequences
of systemic AZ1 overexpression. A genomic antizyme
gene constructs with a thymidine deletion, correspond-
ing to position 205 of antizyme cDNA was used. This
154 Transgenic Res (2014) 23:153–163
123
Page 3
converted the inframe UGA stop codon to UGG codon.
As a result, production of antizyme protein was
possible without the need of ribosomal frameshifting
(Matsufuji et al. 1995; Kankare et al. 1997).
Materials and methods
Generation of transgenic mice
The transgenic mice were generated using the standard
pronuclear microinjection technique (Hogan et al.
1986). Fertilized oocytes were obtained from supe-
rovulated BALB/c 9 DBA/2 (CD2F1) females mated
with CD2F1 males. The microinjected zygotes were
allowed to reach 2-cell stage and were then transferred
into oviducts of pseudopregnant foster females. The
gene construct used for transgenesis was a 5.5-kbp
genomic sequence isolated from 129 SVJ mouse
genomic library (Stratagene, La Jolla, CA) with a
thymidine deletion, corresponding to position 205 of
antizyme cDNA (Kankare et al. 1997) (Fig. 1). As the
transgene construct was of mouse origin, the method
for its detection was based on the integration of
multiple transgene copies in the form of concatamers
(head-to-tail) so that a minimum of two integrated
transgene copies were required for the detection. The
design of the oligonucleotides was based on the head-
to-tail recombination of the transgenes so that the
upper primer recognized the tail region and the lower
one the head region of the AZ1 transgene (Fig. 1). The
following oligonucleotide primers were used in the
PCR: 50-TGCCTATCTTCATGGAGAC-30 (from 30
tail) and 50-ATAGATGGTTGTGAGCCAC-30 (from
50 head). A positive control form the founder animal
was always used when the transgenity of progeny was
tested. The founder animals with no transgenic
offspring were sacrificed at the age of 8–10 months
for the phenotype analysis.
Analytical methods
Western blotting was performed according to a
laboratory manual, Current Protocols in Molecular
Biology (Ausubel et al. 2007). Shortly, 25 lg of
protein was used for SDS-PAGE gel electrophoresis
and transferred onto the PVDF membranes. Mem-
branes were blocked with 2 % ECL advance blocking
reagent (GE Healthcare, Fairfield, CT, USA) in
phosphate-buffered saline containing 0.1 % Tween
and probed with Antizyme 1 [described in Kankare
et al. (1997)], Antizyme inhibitor 1 (Cosmo bio,
Tokyo, Japan), HRP conjugated anti-PCNA (PC-10,
Santa Cruz Biotechnology, Santa Cruz, CA) and rabbit
polyclonal against cleaved Caspase 3 (Cell Signaling
Technology, Danvers, MA) and b-actin (Santa Cruz
Biotechnology) antibodies. DyLight 680 and 800-con-
jugated secondary antibodies (Thermo Fisher Scien-
tific) were used in 1:10,000 dilutions. Antibody-bound
protein was detected by the Odyssey Infrared Imaging
System (Li-Cor Biosciences, Lincoln, NE, USA).
The tissue specimens for histology were fixed in
10 % formalin in phosphate buffer, embedded in
paraffin and cut into 5-lm sections and stained with
hematoxylin/eosin (HE). For immunohistochemistry,
rabbit polyclonal anti-antizyme (1:1,000), HRP con-
jugated anti-PCNA (1:750) and rabbit polyclonal
against cleaved Caspase 3 (1:1,000) were used. PCNA
and cleaved Caspase 3 antigen retrieval was carried
out by boiling samples in 0.01 M citric acid buffer pH
6.0 for 10 min. To visualize the rabbit polyclonal
primary antibodies, we used Power Vision Poly-HRP
Rabbit IgG IHC kit (ImmunoLogic, Duiven, The
Netherlands).
Polyamines were quantified by using high-perfor-
mance liquid chromatography as described earlier
(Hyvonen et al. 1992). The activity of ornithine
decarboxylase was assayed by the method described in
(Janne and Williams-Ashman 1971).
Results
We obtained seven transgenic founder mice out of 57
pups. As the success rate was relatively good, it can be
considered that the transgene, if expressed in the early
embryos of the founders, was compatible with mouse
embryogenesis. All but one of the founders were
fertile, but none of the progeny (185 pups) were
transgenic. The unfertile male founder mouse (406F0)
was first to be sacrificed at the age of 8 months and
analyzed. A macroscopic postmortem examination
and histopathological evaluation of 13 different
tissues was performed but no morphological differ-
ences were detected when compared with age matched
wild type mouse. However, 406F0 overexpressed AZ1
protein in various organs (Fig. 2). Strong overexpres-
sion of full-length AZ1 proteins (24 and 29 kDa) were
Transgenic Res (2014) 23:153–163 155
123
Page 4
found in testes, epididymes, pancreas and brain. Also
liver, kidneys and salivary glands overexpressed the
transgene. In these organs, however, the majority of
the AZ1 protein was degraded into smaller fragments.
Eventually, the rest of the founder animals were
sacrificed at the age of 10 months and analyzed.
Similarly to 406F0, none of the animals showed any
morphological changes in their tissues. The protein
expression in four tissues was analyzed from each
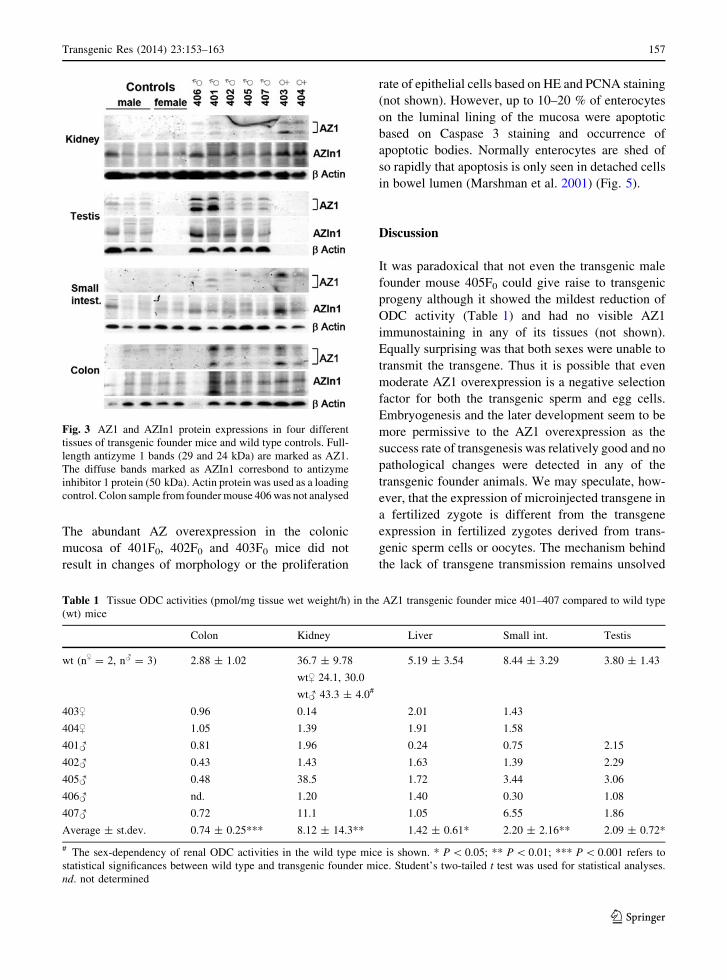
founder mouse (Fig. 3). The testis, small intestine and
colon were chosen for the analysis because we were
interested in the effect of AZ1 on these highly
proliferative tissues. Also kidneys were included
because it is known to have high sex-depended renal
ODC activity in rodents. This sex-dependency was also
seen in our ODC activity measurement where male
mice showed twice as high renal ODC activity than
female mice (Table 1). Similarly to 406F0, the founder
401F0 showed high AZ1 protein expression in the
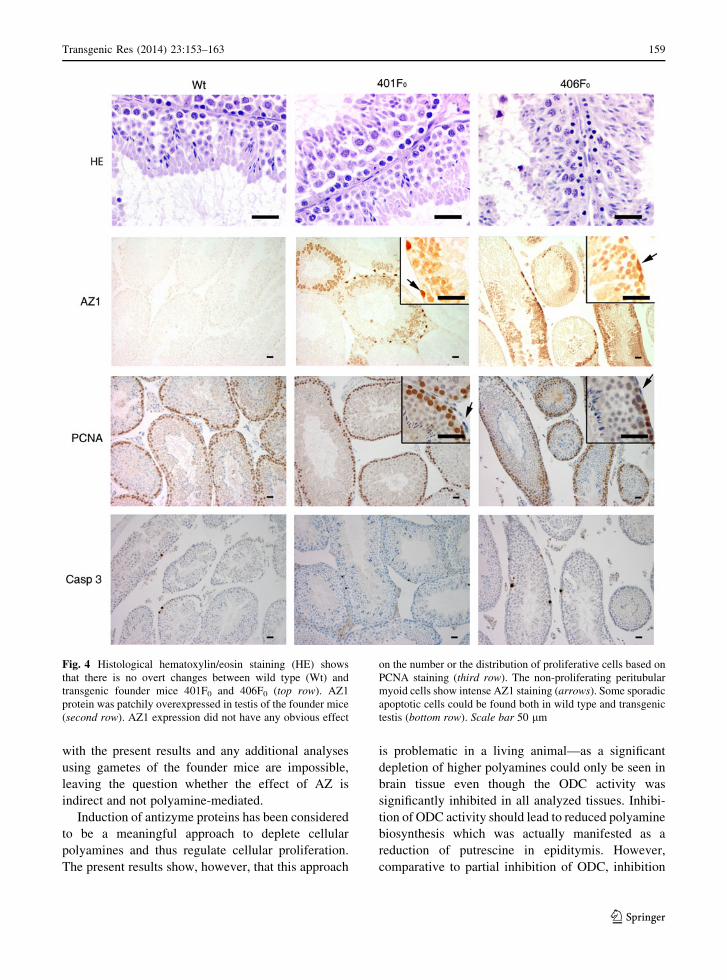
testis, which was also evident in immunohistochemis-
try as a patchy cytosolic staining in spermatocytes and
intense nuclear staining in the peritubular myoid cells
(Fig. 4). Also the male founders 402F0, 405F0, and
407F0 showed slight AZ1 protein overexpression in
their testis by immunoblotting analysis (Fig. 3). How-
ever, the level of expression was not high enough to
distinguish those animals from the wild type controls
by immunohistochemistry (not shown). Nor did the
ovaries of the two female founders (403F0 and 404F0)
show any AZ1 immunostaining above the wild type
controls (not shown).
Even the relatively weak overexpression of AZ1
protein in colon, kidney and small intestine (Fig. 3)
resulted in significant reduction of ODC activity in
transgenic founder mice. The most dramatic reduction
was seen in the kidneys of both sexes of the transgenic
mice (405F0 being an exception) (Table 1). The
reduction of ODC activity in these organs was not,
however, accompanied with significant depletion of
polyamine pools (Table 2). The only organ showing
reduction of higher polyamines was brain where
spermidine levels in transgenic mice were lower than
in wild type mice. Relevant putrescine concentrations
were only detected in epiditymis where it was also
reduced in transgenic mice. Surprisingly, hepatic
spermine levels were increased in transgenic animals.
A clear compensatory increase in antizyme inhibitor 1
(AZIn1) protein expression (Fig. 3) was wound in
kidney colon and testis. AZ immunohistochemistry
confirmed the results from western analysis and
showed most intense staining in the colon epithelia
of the founder mice 401F0, 402F0 and 403F0 (Fig. 5).
In these animals the staining was evenly spread
through the colonic mucosa whereas in the other
founders and wild type controls the staining was more
restricted to the luminal part of mucosa—as shown
earlier in wild type mice by Gritli-Linde et al. (2001).
Fig. 1 a The transgene was a 5.5-kbp Eco R1- Eco R1 fragment
isolated from 129 SVJ mouse genomic library (Stratagene, La
Jolla, CA) with a thymidine deletion (DT), corresponding to
position 205 of antizyme cDNA. Exon and intron sequences are
specified by boxes and solid lines respectively, and the protein
coding parts in the exons are in black (adopted from Kankare
et al. 1997). b. The PCR detection of transgenic pups based on
integration of multiple transgene copies to the genome in the
form of concatamers. Thus, only pups with two or more
integration were identified as transgenic. The position of PCR
primers are shown as small arrows
Fig. 2 A western blot analysis of a wild type (wt) and a transgenic mouse 406F0. The bands corresponding to the full-length antizyme 1
(29 and 24 kDa) are marked as AZ1. Actin protein was used as a loading control
156 Transgenic Res (2014) 23:153–163
123
Page 5
The abundant AZ overexpression in the colonic
mucosa of 401F0, 402F0 and 403F0 mice did not
result in changes of morphology or the proliferation
rate of epithelial cells based on HE and PCNA staining
(not shown). However, up to 10–20 % of enterocytes
on the luminal lining of the mucosa were apoptotic
based on Caspase 3 staining and occurrence of
apoptotic bodies. Normally enterocytes are shed of
so rapidly that apoptosis is only seen in detached cells
in bowel lumen (Marshman et al. 2001) (Fig. 5).
Discussion
It was paradoxical that not even the transgenic male
founder mouse 405F0 could give raise to transgenic
progeny although it showed the mildest reduction of
ODC activity (Table 1) and had no visible AZ1
immunostaining in any of its tissues (not shown).
Equally surprising was that both sexes were unable to
transmit the transgene. Thus it is possible that even
moderate AZ1 overexpression is a negative selection
factor for both the transgenic sperm and egg cells.
Embryogenesis and the later development seem to be
more permissive to the AZ1 overexpression as the
success rate of transgenesis was relatively good and no
pathological changes were detected in any of the
transgenic founder animals. We may speculate, how-
ever, that the expression of microinjected transgene in
a fertilized zygote is different from the transgene
expression in fertilized zygotes derived from trans-
genic sperm cells or oocytes. The mechanism behind
the lack of transgene transmission remains unsolved
Fig. 3 AZ1 and AZIn1 protein expressions in four different
tissues of transgenic founder mice and wild type controls. Full-
length antizyme 1 bands (29 and 24 kDa) are marked as AZ1.
The diffuse bands marked as AZIn1 corresbond to antizyme
inhibitor 1 protein (50 kDa). Actin protein was used as a loading
control. Colon sample from founder mouse 406 was not analysed
Table 1 Tissue ODC activities (pmol/mg tissue wet weight/h) in the AZ1 transgenic founder mice 401–407 compared to wild type
(wt) mice
Colon Kidney Liver Small int. Testis
wt (n$ = 2, n# = 3) 2.88 ± 1.02 36.7 ± 9.78
wt$ 24.1, 30.0
wt# 43.3 ± 4.0#
5.19 ± 3.54 8.44 ± 3.29 3.80 ± 1.43
403$ 0.96 0.14 2.01 1.43
404$ 1.05 1.39 1.91 1.58
401# 0.81 1.96 0.24 0.75 2.15
402# 0.43 1.43 1.63 1.39 2.29
405# 0.48 38.5 1.72 3.44 3.06
406# nd. 1.20 1.40 0.30 1.08
407# 0.72 11.1 1.05 6.55 1.86
Average ± st.dev. 0.74 ± 0.25*** 8.12 ± 14.3** 1.42 ± 0.61* 2.20 ± 2.16** 2.09 ± 0.72*
# The sex-dependency of renal ODC activities in the wild type mice is shown. * P \ 0.05; ** P \ 0.01; *** P \ 0.001 refers to
statistical significances between wild type and transgenic founder mice. Student’s two-tailed t test was used for statistical analyses.
nd. not determined
Transgenic Res (2014) 23:153–163 157
123
Page 6
Ta
ble
2T
issu
ep
oly
amin
ep
oo
ls(p
mo
l/m
gti
ssu
ew
etw
eig
ht)
inth
eA
Z1
tran
sgen
icfo
un
der
mic
e4
01
–4
07
com
par
edto
wil
dty
pe
(wt)
mic
e
Pu
tres
cin
e#S
per
mid
ine
Ep
idit
ym
isB
rain
Co
lon
Ep
idit
ym
isK
idn
eyL
iver
Pan
crea
sS
mal
lin
t.T
esti
s
wt
n$
=2
,n#
=3
18
5±
36
35
8±
15
44
7±
55
72
2±
68
24
4±
37
63
1±
47
4,9
90
±4
86
61
±3
92
89
±9
2
40
3$
32
04
69
30
35
57
5,0
30
66
4
40
4$
33
74
25
30
17
99
5,0
20
65
4
40
1#
10
83
50
46
16
61
23
55
91
4,0
30
67
52
79
40
2#
11
93
20
40
55
46
24
96
15
4,6
00
65
83
03
40
5#
14
33
36
47
58
26
19
77
50
5,1
40
63
53
12
40
6#
10
33
07
nd
.7
31
15
84
21
3,3
60
89
23
28
40
7#
17
93
28
34
57
03
21
57
26
4,7
40
67
42
52
Av
erag
e±
st.d
ev.
13
0±
31
*3
28
±1
4*
*4
30
±5
06
93
±1
02
23
6±
53
63
7±
13
14
,56
0±
65
69
3±
89
29
5±
30
Sp
erm
ine
Bra
inC
olo
nE
pid
ity
mis
Kid
ney
Liv
erP
ancr
eas
Sm
all
int.
Tes
tis
wt
n$
=2
,n#
=3
25
0±
18
39
2±
48
16
4±
46
62
2±
59
74
4±
35
1,1
30
±2
45
49
9±
52
45
2±
15
40
3$
24
93
79
68
69
50
1,2
50
48
2
40
4$
24
84
12
59
48
55
1,4
50
46
9
40
1#
22
74
14
23
57
06
77
91
,38
04
99
55
7
40
2#
23
54
13
28
27
04
1,0
30
1,6
60
50
55
29
40
5#
24
44
75
22
56
85
1,1
20
1,4
60
48
65
54
40
6#
27
1n
d.
13
66
93
87
01
,03
05
88
56
9
40
7#
22
84
22
37
06
86
97
21
,43
06
55
51
3
Av
erag
e±
st.d
ev.
24
3±
16
41
9±
31
25
0±
86
67
9±
39
93
9.9
±1
16
**
1,3
80
±1
98
52
6±
69
54
4±
23
#R
elev
ant
pu
tres
cin
eco
nce
ntr
atio
ns
wer
eo
nly
det
ecte
dfr
om
epid
ity
mis
.*
P\
0.0
5,
**
P\
0.0
1,
refe
rsto
stat
isti
cal
sig
nifi
can
ces
bet
wee
nw
ild
typ
ean
dtr
ansg
enic
fou
nd
er
mic
e.S
tud
ent’
stw
o-t
aile
dt
test
was
use
dfo
rst
atis
tica
lan
aly
ses.
nd
.n
ot
det
erm
ined
158 Transgenic Res (2014) 23:153–163
123
Page 7
with the present results and any additional analyses
using gametes of the founder mice are impossible,
leaving the question whether the effect of AZ is
indirect and not polyamine-mediated.
Induction of antizyme proteins has been considered
to be a meaningful approach to deplete cellular
polyamines and thus regulate cellular proliferation.
The present results show, however, that this approach
is problematic in a living animal—as a significant
depletion of higher polyamines could only be seen in
brain tissue even though the ODC activity was
significantly inhibited in all analyzed tissues. Inhibi-
tion of ODC activity should lead to reduced polyamine
biosynthesis which was actually manifested as a
reduction of putrescine in epiditymis. However,
comparative to partial inhibition of ODC, inhibition
Fig. 4 Histological hematoxylin/eosin staining (HE) shows
that there is no overt changes between wild type (Wt) and
transgenic founder mice 401F0 and 406F0 (top row). AZ1
protein was patchily overexpressed in testis of the founder mice
(second row). AZ1 expression did not have any obvious effect
on the number or the distribution of proliferative cells based on
PCNA staining (third row). The non-proliferating peritubular
myoid cells show intense AZ1 staining (arrows). Some sporadic
apoptotic cells could be found both in wild type and transgenic
testis (bottom row). Scale bar 50 lm
Transgenic Res (2014) 23:153–163 159
123
Page 8
Fig. 5 AZ1 protein was
overexpressed evenly in the
colon epithelia in the
transgenic founders 401F0,
402F0 and 403F0. Some
sporadic apoptotic cells
could be found both in wild
type and transgenic colon
epithelia. However, based
on caspase 3 staining, the
transgenic founders 401F0,
402F0 and 403F0 showed
increased amount of
apoptotic cells on the
epithelial mucosa facing the
lumen of colon
160 Transgenic Res (2014) 23:153–163
123
Page 9
of polyamine uptake by AZ was probably also only
partial although we have no evidence for it. Therefore,
polyamine uptake from external sources, like from gut
bacteria and diet, is expected to contribute to the
maintenance of polyamine pools in a whole body
setting. The slight but statically significant reduction
of spermidine in brain tissue backs up this assumption
as the transport of polyamines to brain tissue is limited
due to the blood brain barrier (Shin et al. 1985) and
thus the reduction in polyamine biosynthesis is more
readily manifested in brain. The increased hepatic
spermine in transgenic mice may also reflect the
compensatory intake of polyamine from these external
sources. It is also possible that in our animals, during
long-term, ubiquitous and constitutive expression of
the transgene, the tissues have adapted to the meta-
bolic condition and the remaining level of ODC
activity was sufficient to maintain the normal pools of
spermidine and spermine. One explanation for the
unaffected polyamine levels is the compensatory
increase in the AZIn1 protein level in transgenic
founders, which attenuates the effect of AZ1
overexpression.
We chose to use a mouse-derived transgene con-
struct with its own regulatory sequences, as this
approach can give valuable information about the
tissue dependency of AZ1 action. For example, it is
remarkable how strongly AZ1 expression was directed
to the testes of the transgenic mice. It has been shown
earlier that polyamine synthesis follows a fine tuned
temporal and spatial pattern during spermatogenesis
where ODC transcripts are especially abundant in late
pachytene of the first meiotic division and in the early-
stage round spermatids (Kaipia et al. 1990). The
polyamine biosynthesis needs to be strictly regulated
as disturbance of this regulation e.g. by ODC overex-
pression leads to reduction of meiotic cells and to
disruption of spermatogenesis (Halmekyto et al.
1991). Remarkably, a testis specific antizyme (AZ3)
has specially evolved to shut down the polyamine
synthesis at the time of late meiosis and spermiogen-
esis. It has also been shown that there is a transition of
AZ expression from AZ1 in spermatogonias and
spermatocytes to AZ3 in haploid spermatids (Tosaka
et al. 2000; Ivanov et al. 2000). Our results support
these findings as the AZ1 overexpression was mainly
directed to the basal compartment of semiferous
tubules (Fig. 4). However, a mysterious patchy
expression pattern was found where some of the
tubular cross sections were devoid of staining and
some had patches of more and less intense staining.
The latter can be partly explained by the syncytium
where developing sperm cells, originated from single
spermatogonia, are attached by cytoplasmic bridges
forming clusters of cells shearing their cytoplasmic
proteins. It is also possible, but highly unlikely, that
each founder animal was a gonadal mosaic due to late
transgene integration occurring after the one cell stage
of the embryo, which could easily lead to uneven
distribution of the transgene. However, in other
organs, such as in intestine, kidney and liver, the
overexpressed transgene resulted to even staining
without any visible pattern of somatic mosaicism.
We and others have shown that it is possible to
hinder (Murakami et al. 1994) or halt (Pietila et al.
2012) the cellular proliferation by overexpressing AZ1
in vitro. Moreover, AZ1 overexpression inhibits
malignant growth in chemical skin carcinoma model
(Feith et al. 2001). There is also evidence that AZ1 can
induce apoptosis through mitochondrial membrane
depolarization in haematopoietic cells (Liu et al.
2006). Based on this background, we were interested
to see whether AZ overexpression could have an
impact on highly proliferative intestinal epithelia
where apoptosis has also an important role in the
mucosal regeneration. Based on histological exami-
nation, no obvious changes were seen in proliferative
base of intestinal epithelia. However, increased apop-
tosis was evident on the luminal lining of the mucosa
of transgenic animals. This can be regarded as
abnormal, as in normal undamaged epithelia, apopto-
sis is triggered by the detachment of epithelial cells
from the underlying basement membrane and is rarely
seen in cells still attached to the epithelia (Marshman
et al. 2001). Further research is needed to conclude
whether this observation is due premature apoptosis or
defect in detachment of cells, as the polyamines have
been linked both to regulation of apoptosis of intes-
tinal epithelia by HuR dependent p53 mRNA stabil-
ization (Wang et al. 2007) and to integrity of epithelia
by c-Myc regulated E-catherin expression (Liu et al.
2009).
Acknowledgments We thank Prof. Olli Janne for providing
the mutated mouse AZ1 gene construct and the rabbit polyclonal
anti-AZ1 antibody. We also thank Ms. Marita Heikkinen, Sisko
Juutinen, Arja Korhonen, Anne Karppinen and Tuula Reponen
for their skillful technical assistance. This work was financially
supported by the Academy of Finland.
Transgenic Res (2014) 23:153–163 161
123
Page 10
References
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG,
Smith JA, Struhl K (2007) Current protocols in molecular
biology. Wiley, Hoboken
Bailey HH, Kim K, Verma AK et al (2010) A randomized,
double-blind, placebo-controlled phase 3 skin cancer pre-
vention study of a-difluoromethylornithine in subjects with
previous history of skin cancer. Cancer Prev Res 3:35–47
Casero R Jr, Frydman B, Stewart TM, Woster PM (2005) Sig-
nificance of targeting polyamine metabolism as an anti-
neoplastic strategy: unique targets for polyamine
analogues. Proc West Pharmacol Soc 48:24–30
Feith DJ, Shantz LM, Pegg AE (2001) Targeted antizyme expres-
sion in the skin of transgenic mice reduces tumor promoter
induction of ornithine decarboxylase and decreases sensitivity
to chemical carcinogenesis. Cancer Res 61:6073–6081
Fong LYY, Feith DJ, Pegg AE (2003) Antizyme overexpression in
transgenic mice reduces cell proliferation, increases apop-
tosis, and reduces N-nitrosomethylbenzylamine-induced
forestomach carcinogenesis. Cancer Res 63:3945–3954
Gritli-Linde A, Nilsson J, Bohlooly-Y M, Heby O, Linde A
(2001) Nuclear translocation of antizyme and expression of
ornithine decarboxylase and antizyme are developmentally
regulated. Dev Dyn 220:259–275
Gupta S, Ahmad N, Marengo SR, MacLennan GT, Greenberg
NM, Mukhtar H (2000) Chemoprevention of prostate car-
cinogenesis by a-difluoromethylornithine in TRAMP
mice. Cancer Res 60:5125–5133
Halmekyto M, Hyttinen JM, Sinervirta R et al (1991) Trans-
genic mice aberrantly expressing human ornithine decar-
boxylase gene. J Biol Chem 266:19746–19751
Heby O, Persson L, Rentala M (2007) Targeting the polyamine
biosynthetic enzymes: a promising approach to therapy of
African sleeping sickness, Chagas’ disease, and leish-
maniasis. Amino Acids 33:359–366
Hogan B, Constantini F, Lacy E (1986) Manipulating the mouse
embryo. Cold Spring Harbor Laboratory, Cold Spring
Harbor, NY, pp 1–322
Hyvonen T, Keinanen TA, Khomutov AR, Khomutov RM, El-
oranta TO (1992) Monitoring of the uptake and metabolism
of aminooxy analogues of polyamines in cultured cells by
high-performance liquid chromatography. J Chromatogr
574:17–21
Ivanov IP, Rohrwasser A, Terreros DA, Gesteland RF, Atkins JF
(2000) Discovery of a spermatogenesis stage-specific
ornithine decarboxylase antizyme: antizyme 3. Proc Natl
Acad Sci USA 97:4808–4813
Janne J, Williams-Ashman HG (1971) On the purification of L-
ornithine decarboxylase from rat prostate and effects of thiol
compounds on the enzyme. J Biol Chem 246:1725–1732
Kahana C (2007) Ubiquitin dependent and independent protein
degradation in the regulation of cellular polyamines.
Amino Acids 33:225–230
Kahana C, Asher G, Shaul Y (2005) Mechanisms of protein
degradation: an odyssey with ODC. Cell Cycle 4:1461–1464
Kaipia A, Toppari J, Mali P, Kangasniemi M, Alcivar AA, Hecht
NB, Parvinen M (1990) Stage- and cell-specific expression
of the ornithine decarboxylase gene during rat and mouse
spermatogenesis. Mol Cell Endocrinol 73:45–52
Kaiser A, Gottwald A, Maier W, Seitz HM (2003) Targeting
enzymes involved in spermidine metabolism of parasitic
protozoa—a possible new strategy for anti-parasitic treat-
ment. Parasitol Res 91:508–516
Kankare K, Uusi-Oukari M, Janne OA (1997) Structure, orga-
nization and expression of the mouse ornithine decarbox-
ylase antizyme gene. Biochem J 324:807–813
Liu GY, Liao YF, Hsu PC, Chang WH, Hsieh MC, Lin CY, Hour
TC, Kao MC, Tsay GJ, Hung HC (2006) Antizyme, a
natural ornithine decarboxylase inhibitor, induces apopto-
sis of haematopoietic cells through mitochondrial mem-
brane depolarization and caspases’ cascade. Apoptosis
11:1773–1788
Liu B, Sun H, Wang W, Li W, Yan Y, Chen S, Yang Y, Xu C,
Xin J, Liu X (2009) Adenovirus vector-mediated upregu-
lation of spermidine/spermineN1-acetyltransferase impairs
human gastric cancer growth in vitro and in vivo. Cancer
Sci 100:2126–2132
Marshman E, Ottewell PD, Potten CS, Watson AJM (2001) Cas-
pase activation during spontaneous and radiation-induced
apoptosis in the murine intestine. J Pathol 195:285–292
Martinez ME, O’Brien TG, Fultz KE, Babbar N, Yerushalmi H,
Qu N, Guo Y, Boorman D, Einspahr J, Alberts DS, Gerner
EW (2003) Pronounced reduction in adenoma recurrence
associated with aspirin use and a polymorphism in the
ornithine decarboxylase gene. Proc Natl Acad Sci USA
100:7859–7864
Matsufuji S, Matsufuji T, Miyazaki Y, Murakami Y, Atkins JF,
Gesteland RF, Hayashi S (1995) Autoregulatory frame-
shifting in decoding mammalian ornithine decarboxylase
antizyme. Cell 80:51–60
Meyskens J, McLaren CE, Pelot D et al (2008) Difluorometh-
ylornithine plus sulindac for the prevention of sporadic
colorectal adenomas: a randomized placebo-controlled,
double-blind trial. Cancer Prev Res 1:32–38
Mitchell JL, Judd GG, Leyser A, Choe C (1998) Osmotic stress
induces variation in cellular levels of ornithine decarbox-
ylase-antizyme. Biochem J 329:453–459
Murakami Y, Matsufuji S, Miyazaki Y, Hayashi S (1994)
Forced expression of antizyme abolishes ornithine decar-
boxylase activity, suppresses cellular levels of polyamines
and inhibits cell growth. Biochem J 304:183–187
Perez-Leal O, Merali S (2012) Regulation of polyamine
metabolism by translational control. Amino Acids 42:
611–617
Pietila M, Lampinen A, Pellinen R, Alhonen L (2012) Inducible
expression of antizyme 1 in prostate cancer cell lines after
lentivirus mediated gene transfer. Amino Acids 42:559–564
Shantz LM, Levin VA (2007) Regulation of ornithine decar-
boxylase during oncogenic transformation: mechanisms
and therapeutic potential. Amino Acids 33:213–223
Shin WW, Fong WF, Pang SF, Wong PC (1985) Limited blood-
brain barrier transport of polyamines. J Neurochem
44:1056–1059
Tosaka Y, Tanaka H, Yano Y, Masai K, Nozaki M, Yomogida
K, Otani S, Nojima H, Nishimune Y (2000) Identification
and characterization of testis specific ornithine decarbox-
ylase antizyme (OAZ-t) gene: expression in haploid germ
cells and polyamine-induced frameshifting. Genes Cells
5:265–276
162 Transgenic Res (2014) 23:153–163
123
Page 11
Wallace HM, Fraser AV (2004) Inhibitors of polyamine
metabolism: review article. Amino Acids 26:353–365
Wang X, Feith DJ, Welsh P, Coleman CS, Lopez C, Woster PM,
O’Brien TG, Pegg AE (2007) Studies of the mechanism by
which increased spermidine/spermine N1-acetyltransfer-
ase activity increases susceptibility to skin carcinogenesis.
Carcinogenesis 28:2404–2411
Transgenic Res (2014) 23:153–163 163
123