Page 1
TAKING THE BULL BY THE HORNS: IDEOLOGY, MASCULINITY, AND CATTLE HORNS AT ÇATALHÖYÜK (TURKEY)
K.C. TWISS and N. RUSSELL
Abstract: Animal symbolism is a ubiquitous and powerful component of human ideology. Cattle were clearly the preeminent symbolic taxon in the Southwest Asian Neolithic, and archaeologists have argued that not just cattle, but specifi cally bulls, were key symbols. However, the biological attributes, including the sex, of the cattle used in Neolithic symbolic contexts remain largely unestablished. Furthermore, most of the cattle representations on which these arguments are based are not clearly male. In this article, we test the symbolic importance of taurine masculinity to early villagers by assessing the biological characteristics of cattle horns found in spe-cial deposits as well as in more prosaic contexts at the Anatolian Neolithic site of Çatalhöyük.
Résumé : Le symbolisme animal est une composante omniprésente et puissante de l’idéologie humaine. Les bovinés constituaient le taxon symbolique majeur dans le Néolithique de l’Asie du Sud-Ouest, et les archéologues ont affi rmé que ce n’étaient pas simplement les bovinés, mais plutôt les taureaux qui constituaient le symbole-clé. Pourtant, les attributs biologiques, dont celui du sexe, des bovinés utilisés dans des contextes symboliques néolithiques restent en grande partie indéterminés. De plus, la plupart des représentations de bovinés sur lesquelles ces arguments sont fondés n’attestent pas cette masculinité. Dans cet article, nous examinons l’importance symbolique de la masculinité taurine pour les premiers agriculteurs en évaluant les caractéristiques biologiques des cornes trouvées dans des dépôts spécifi ques de même que dans des contextes plus prosaïques du village néolithique anatolien de Çatalhöyük.
Keywords: Neolithic; Cattle; Bull; Symbolism; Çatalhöyük; Turkey.Mots-clés : Néolithique ; Bovinés ; Taureau ; Symbolisme ; Çatalhöyük ; Turquie.
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009 Manuscrit reçu le 4 août 2009, accepté le 19 janvier 2010
Animal symbolism is a ubiquitous and powerful compo-
nent of human ideology. Animals are commonly crucial ele-
ments of ritual activity, and their remains are frequently found
in ritual deposits. In the Southwest Asian Neolithic, cattle were
the preeminent symbolic taxon, and researchers have posited
that not just cattle, but specifi cally bulls, were key symbols.
Indeed, Neolithic populations have been called the “people of
the bull”.1 These accounts commonly interpret bulls as sym-
bols of masculinity and vitality2—traits that scholars have seen
as (a) propelling the diffusion of village life,3 or (b) sustaining
ties to hunter-gatherer lifeways4.
1. CAUVIN, 2000: 123.
2. Although see TESTART, 2006: 28.
3. CAUVIN, 2000.
4. VERHOEVEN, 2002.
However, the biological attributes, including the sex, of
the cattle used in Neolithic symbolic contexts remain largely
unestablished. Furthermore, most of the cattle representations
on which these arguments are based are not clearly male. A
recent paper therefore challenged the supposed masculine
associations of Çatalhöyük’s cattle horns, relying on modern
ethnographic comparisons with Southeast Asian societies that
also display bovine horns on their houses.5 We here marshal
archaeological evidence to test the trope, and by extension the
symbolic importance of masculinity to early villagers, assess-
ing the biological characteristics of cattle horns found in spe-
cial deposits such as feasts and architectural installations as
5. TESTART, 2006.
019-032-Twiss.indd 19019-032-Twiss.indd 19 22/07/10 12:30:3922/07/10 12:30:39
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 2
20 K.C. TWISS and N. RUSSELL
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
well as those recovered from more prosaic contexts at the Ana-
tolian Neolithic site of Çatalhöyük.
CATTLE CRANIA IN THE SOUTHWEST ASIAN NEOLITHIC
Cattle, and particularly cattle crania, are the dominant ani-
mal symbol of the Southwest Asian Neolithic, with the result
that scholars have posited a widespread Neolithic “cattle cult”.6
Cattle images dominate fi gurine assemblages and appear in
murals, reliefs, and stone carvings; special deposits of cattle
remains are found throughout the region.7
Cattle crania, and particularly horns, appear to have been
particularly symbolically weighted. At sites that have non-
domestic architecture (e.g., Çayönü, Aswad), these buildings
are often marked with cattle horns and skulls. Architectural
installations involving cattle skulls and/or horn cores are
reported from (proto-Neolithic) Hallan Çemi as well as Neo-
lithic Çayönü and Jerf el-Ahmar;8 cattle horns were concealed
in or under walls, fl oors, and benches at Mureybet, Halula,
Tell ‘Abr 3, Dja’de and Ginnig.9 A cache of both cattle and
goat skulls at Ghwair I lay nearly atop a plastered fl oor, under
which was a burial.10 Other sites with cattle horns and skulls
apparently placed as abandonment deposits include Tell ‘Ain
el-Kerkh (a pair of horns left on a building’s fl oor) and Tell
Aswad (an aurochs skull placed across a threshold).11 Mortuary
deposits involving cattle crania are likewise attested through-
out the Neolithic: aurochs skulls and horns in a sub-fl oor pit
with multiple human burials in Çayönü’s Skull Building, an
aurochs cranium plausibly associated with a burial at Hatoula,
and a fragmentary cattle horn in a burial at Menteşe.12
This symbolic richness underlies the long-powerful idea
that the bull was a core symbol of the Neolithic Near East.
J. Cauvin, the primary advocate for this view, interpreted the
bull as an image of masculine power, the associate and com-
plement to a feminine, civilizing goddess. He argued that the
power of the bull for Neolithic populations developed in asso-
ciation with human dominance over nature and the wild:
6. ROLLEFSON et al., 1992: 466.
7. GORING-MORRIS and HORWITZ, 2007: 913; CAUVIN, 2000.
8. ROSENBERG, 1994: 125; ÖZDOĞAN, 1999; HELMER et al., 2004: 151.
9. HELMER et al., 2004; CAUVIN, 2000: 28; CAMPBELL and BAIRD, 1990;
MOLIST, 1998; YARTAH, 2004 ; É. Coqueugniot, personal communication.
10. SIMMONS and NAJJAR, 1999.
11. MALLOWAN, 1946; TSUNEKI, 2002: 140.
12. LECHEVALLIER et RONEN, 1994; ÖZDOĞAN, 1999; ALPASLAN-
ROODENBERG, 2001: 4.
“only he who can control instinct [e.g., the ‘brute violence’
symbolized by bulls] can truly and precisely be a man (…). It
may be in that this dialectic of ‘virility’ (…) the real internal
dynamism of the PPNB culture (…) lived on.”13
The wild bull symbolized the masculine component of the
Neolithic ethos; it was this component that gave early agricul-
turalists their expansionist values and led to the diffusion of
village lifeways.14
M. Verhoeven,15 despite questioning the centrality of the
woman-and-the-bull cult in Neolithic ideology, has argued
that:
“(…) the presence of aurochs [sic] horns in PPNB contexts
may perhaps also be explained by invoking the concept of
vitality (…). Maybe, as Cauvin (2000) has argued, the bull
in general, and especially powerful and liminal elements like
horns and blood (Verhoeven 2000a, 57-9), were metaphors for
male dominance, power and vitality (…). [B]ulls do seem to be
related to vitality, i.e. domestication, life-force and fecundity.”
Verhoeven stresses that PPNB animal iconography over-
whelmingly involves wild animals rather than domesticates,
and posits:
“the set of links: human-wild-male seems to indicate that
within early agricultural societies in the Near East people
were symbolically attached to the wild, to nature, and that this
domain may have been generally regarded as male. In this res-
pect it is interesting to note that the bull, perhaps a fertility-
giving beast, when used in rituals was both male and wild”.16
According to these models, the wildness and the masculin-
ity of the animals are what makes them central to the early
agriculturalists’ ideology: links between humans and wild
male animals counterbalanced the ongoing domestication of
both resources and society, enabling the maintenance of ties to
hunter-gatherer lifeways.17
Although cattle masculinity and maturity are integral to
these widely-cited interpretations of early agricultural ideol-
ogy and the rise of domestication,18 the biological attributes of
the cattle found in Southwest Asian Neolithic special deposits
have been minimally queried. Their identity as bulls remains
an assumption, allowing arguments ranging from the above to
13. CAUVIN, 2000: 125.
14. Ibid.: 133.
15. VERHOEVEN, 2002: 251.
16. Ibid.: 252.
17. Ibid.: 252-253.
18. See also HELMER et al., 2004.
019-032-Twiss.indd 20019-032-Twiss.indd 20 22/07/10 12:30:4022/07/10 12:30:40
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 3
Taking the Bull by the Horns: Ideology, Masculinity, and Cattle Horns at Çatalhöyük 21
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
the idea that animal sex was irrelevant, as cattle—male and
female—were simply the Neolithic sacrifi cial species par excellence.19
Çatalhöyük’s relatively large assemblage of fairly com-
plete cattle horn cores, as well as abundant contextual infor-
mation, permit analyses that integrate data on animal age,
size, and sex; depositional context; and varying treatments of
the remains. It is thus uniquely well-suited to an investigation
of the centrality of the bull in Neolithic ideology. In order to
evaluate whether Neolithic populations chose primarily male,
large, or prime-age animals for symbolic use, we fi rst describe
the site and its horn core assemblage, including the various
types of cattle horn deposits and their probable social/ritual
signifi cance. We then compare and contrast the biological
characteristics of the horn cores found in apparently ritual/
special deposits with those from more prosaic contexts.
ÇATALHÖYÜK AND CATTLE SYMBOLISM
The Central Anatolian Neolithic site of Çatalhöyük was
originally excavated by J. Mellaart in the early 1960s; excava-
tions began again in 1995 and are ongoing.20 There are two
mounds at Çatalhöyük, but Neolithic occupation is limited to
the 34-acre East Mound. The occupation there spans approxi-
mately 7400-6000 cal. BC, and is associated with the regional
Early Ceramic Neolithic. It is estimated that the site’s peak
Neolithic population was between 3,500 and 8,000 people,
making it one of the largest Neolithic settlements in the ancient
Near East. In the initial years of the current project, effort was
focused on reaching the lowest levels of the site, now published
in a series of monographs.21 More recently, excavation has
targeted the later levels; analysis of this material is currently
incomplete.
The site was supported by a combination of crop agricul-
ture, caprine herding, intensive gathering, and hunting. So far,
only the animal remains from the earlier part of the sequence,
dating to between ca 7400 and 6500 cal. BC, have been fully
analyzed. In these levels, cattle constitute 20-25% of the faunal
assemblage, and there is no indication of domestication.22 Pre-
liminary analysis of the later levels shows that cattle drop to
ca 10%; domestication status has not yet been determined for
these levels. Villagers dwelt in mudbrick houses built so close
19. TESTART, 2006.
20. HODDER, 2006b.
21. e.g., HODDER, 2005a-b, 2006a and 2007.
22. RUSSELL et al., 2005.
together that occupants accessed them through their roofs. All
buildings are believed to have been occupied: there are no spe-
cialized shrines or temples at the site. Nonetheless, Çatalhöyük
is renowned for its rich and varied symbolic repertoire. Figu-
rines, wall paintings and moldings, and worked bone and stone
artifacts abound, as do burials, ritual caches, and other kinds
of structured deposits. There is evidence for feasting, complex
mortuary ceremonialism, and possibly deliberate incineration
of buildings.
The symbolic importance of cattle at Çatalhöyük has long
been recognized. Cattle are the most common animal repre-
sented in the zoomorphic fi gurine assemblage, and animal
(mostly cattle) horns make up 78% of the zoomorphic fi gu-
rine assemblage.23 Only two certain examples of bulls in wall
paintings have been found (one more animal may be a bull),
but both are immense cynosures surrounded by smaller fi g-
ures of humans and other animals.24 Limited numbers of cattle
appear to have been consumed, but they feature disproportion-
ately in feasts.25
Within the general body of cattle symbolism, a particu-
lar emphasis on heads and horns is apparent. As noted above,
cattle horns fi gure prominently in the site’s iconography, its rit-
ual activity, and even its architecture. Sometimes the “horns”
are mere artistic representations of cattle horns,26 but actual
Bos cranial remains are frequently used. They are embedded
in walls and benches, installed in the architecture, ceremoni-
ally deposited in caches and abandoned buildings, and even
included (rarely) in burials. Additional ideological weight was
provided to a minority of Çatalhöyük’s horns and bucrania via
plastering—a treatment associated regionally with selected
human skulls, and not reported on cattle horn cores from
other sites. Interestingly, at Çatalhöyük (as at Mureybet), some
cattle horns are incorporated into buildings in ways that leave
them invisible. These occurrences, as well as the abandonment
deposits, suggest that cattle horns were not just trophies for
display: they had non-display signifi cance as well.
It is possible that horns were distinctive visual shorthand
for complete crania, whole animals, or particular ceremonies.
It is also possible that horns had signifi cance apart from, or
above and beyond, that of complete heads. Because there is no
clear cut mark evidence for horn removal at Çatalhöyük, and
because many horn cores are found attached to skulls, it seems
unlikely that horns were typically disassociated from crania.
23. HAMILTON, 2005.
24. RUSSELL and MEECE, 2005.
25. RUSSELL and MARTIN, 2005; FRAME et al., 1999.
26. e.g., HAMILTON, 2005; LAST, 2006; RUSSELL and MEECE, 2005.
019-032-Twiss.indd 21019-032-Twiss.indd 21 22/07/10 12:30:4022/07/10 12:30:40
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 4
22 K.C. TWISS and N. RUSSELL
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
However, it remains possible that they had meaning beyond
that of the rest of the skull: e.g., Verhoeven’s27 suggestion that
horns often symbolize strength and dominance.
THE ÇATALHÖYÜK CATTLE HORN ASSEMBLAGE
All that remains today of Çatalhöyük’s many cattle horns
are their cores. Horn cores are the bony centers of horns, which
in life are covered by keratinous sheaths. The sheaths do not
preserve at Çatalhöyük. It is possible that they were present
during the horns’ use. Treatment of the horn cores is diffi cult
to assess, particularly since many horns are likely to have
passed through multiple stages of use. A core’s treatment—
its modifi cation, decoration, etc.—may have varied over the
course of its use-life. In general, there are few if any direct
indications of horn sheath removal, such as cut marks. This
suggests that sheaths were usually left on the horns. However,
27. VERHOEVEN, 2002.
in some specimens that include both horn and skull fragments,
the horn cores, but not the skulls, are plastered, and the plaster
is so tight on the core that it can only have been applied after
the sheath was gone. Either these sheaths were removed for
another use, or else these horns remained in use for so long that
the sheaths were lost or damaged. In the latter case, perhaps
the plastering represents an attempt to “replace” the missing
sheath.
Since 1995, excavations have yielded 12,466 Neolithic cat-
tle horn cores and horn core fragments. Because horn cores are
often quite fragile, most of these are heavily fragmented splin-
ters. Our comparatively low number of specimen records (716)
refl ects this fragmentation, as we group analytically identical
fragments (from the same context, and with all the same char-
acteristics) into single database records. The minimum number
of elements (MNE, based on horn core tips)28 represented is 71:
28. We recognize that the horn-tip measure for calculating MNE is not ideal.
However, horn cores are fragile, easily distorted in the ground, and
intrinsically unlikely to survive. Tips are the densest portion of the core,
Fig. 1 – Pillars with horn cores, Building 77. (Photo by J. QUINLAN; © Çatalhöyük Research Project.)
019-032-Twiss.indd 22019-032-Twiss.indd 22 22/07/10 12:30:4022/07/10 12:30:40
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 5
Taking the Bull by the Horns: Ideology, Masculinity, and Cattle Horns at Çatalhöyük 23
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
clearly a great underestimate. Our data tables include both the
number of identifi ed specimens (NISP, which heavy fragmen-
tation infl ates) and the MNE (which fragmentation decreases).
The total weight of the horn core assemblage cannot be pre-
cisely assessed: many specimens include fragments of cranial
remains; others have plaster or clay attached to their surfaces;
and a complete bucranium, a horned bench, and two horned
pillars have been left in situ for display (fi gs. 1-2). We estimate
the total weight of the Çatalhöyük horn core assemblage to be
on the order of 154 kg. Many fragments were broken during
excavation rather than predepositionally. The average length of
the undamaged specimens is 13.1 cm.
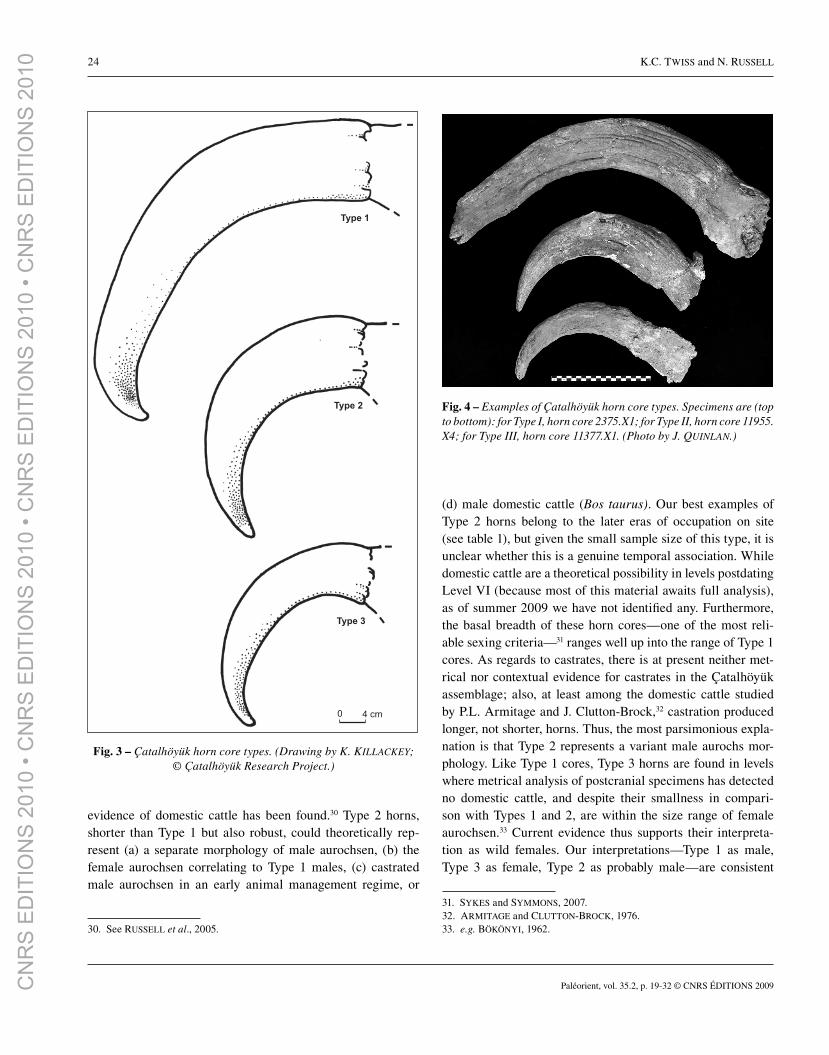
Three morphological types are apparent in the Çatalhöyük
horn core assemblage (fi gs. 3-4; table 1). Type 1 horns are long
(ca 0.5 m from base to tip in a straight line, not around the cur-
vature), robust, and curve outwards from the skull, dip down-
and one that doesn’t require an analyst to guess whether or not, for exam-
ple, > 50% of the circumference is present (as necessary with the bases),
or > 50% of a total core (of an often-unknown original length).
ward midway along the corpus, and then rise again at the tip.
Type 2 horns are shorter and more tightly curved (straight line
base-tip ca 35 cm), fatter, and have a simpler curve: they do not
dip down midway, and rise only slightly at their tips. They are
quite broad in relation to their length, as their greatest breadths
at the base are comparable to those of Type 1 horns, but Type 2
horns are far shorter. Type 3 horns are roughly comparable to
Type 2 horns in length (maximum base-tip line is ca 30 cm),
and have a similarly simple curvature (rising slightly at tip),
but they are distinctly slimmer than Type 2 horns. They also
appear to have round cross-sections at their bases rather than
the ovoid bases seen with Type 1 and Type 2 horns.
Type 1 horns display a classic male Bos primigenius (wild
cattle, or aurochs) morphology,29 and we interpret them as
such. Notably, they are also found throughout the entire Çatal-
höyük sequence of occupation, including early levels where no
29. GRIGSON, 1978; VAN VUURE, 2005: 129; UERPMANN, 1999; BÖKÖNYI,
1962.
Fig. 2 – Bucranium and collapsed bench with horns, Building 52. (Photo by J. QUINLAN; © Çatalhöyük Research Project.)
019-032-Twiss.indd 23019-032-Twiss.indd 23 22/07/10 12:30:4022/07/10 12:30:40
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 6
24 K.C. TWISS and N. RUSSELL
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
Type 1
Type 2
Type 3
0 4 cm
Fig. 3 – Çatalhöyük horn core types. (Drawing by K. KILLACKEY; © Çatalhöyük Research Project.)
evidence of domestic cattle has been found.30 Type 2 horns,
shorter than Type 1 but also robust, could theoretically rep-
resent (a) a separate morphology of male aurochsen, (b) the
female aurochsen correlating to Type 1 males, (c) castrated
male aurochsen in an early animal management regime, or
30. See RUSSELL et al., 2005.
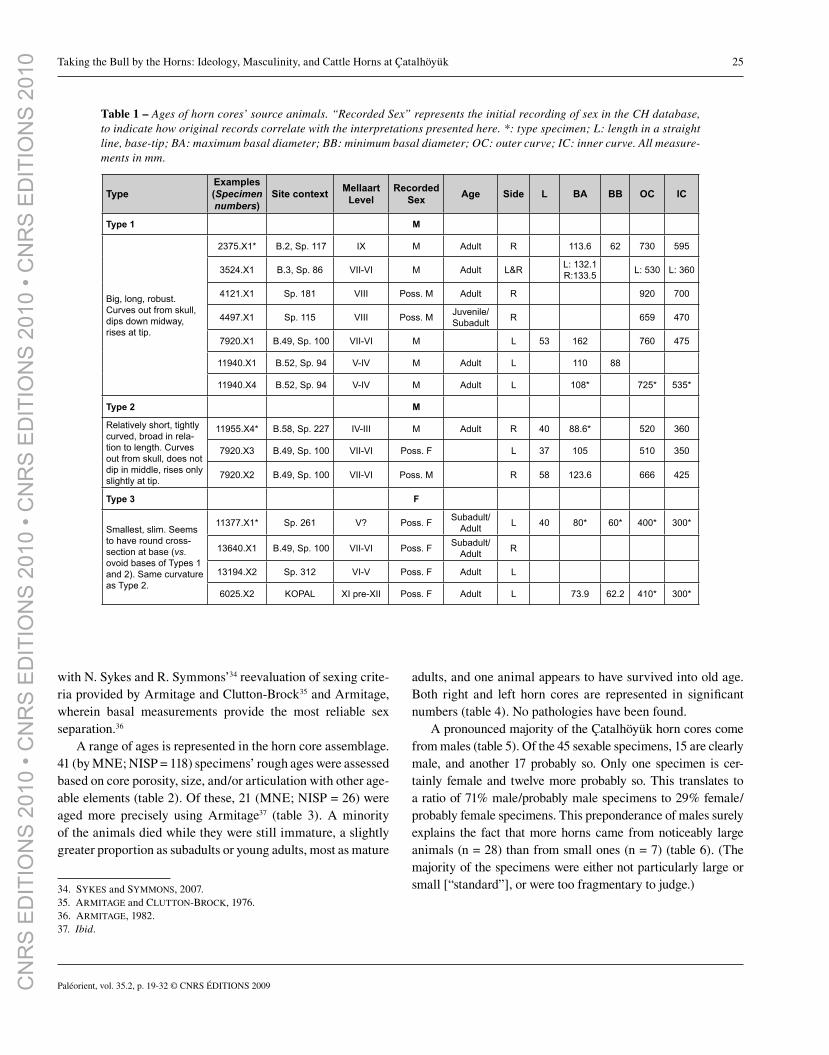
Fig. 4 – Examples of Çatalhöyük horn core types. Specimens are (top to bottom): for Type I, horn core 2375.X1; for Type II, horn core 11955.X4; for Type III, horn core 11377.X1. (Photo by J. QUINLAN.)
(d) male domestic cattle (Bos taurus). Our best examples of
Type 2 horns belong to the later eras of occupation on site
(see table 1), but given the small sample size of this type, it is
unclear whether this is a genuine temporal association. While
domestic cattle are a theoretical possibility in levels postdating
Level VI (because most of this material awaits full analysis),
as of summer 2009 we have not identifi ed any. Furthermore,
the basal breadth of these horn cores—one of the most reli-
able sexing criteria—31 ranges well up into the range of Type 1
cores. As regards to castrates, there is at present neither met-
rical nor contextual evidence for castrates in the Çatalhöyük
assemblage; also, at least among the domestic cattle studied
by P.L. Armitage and J. Clutton-Brock,32 castration produced
longer, not shorter, horns. Thus, the most parsimonious expla-
nation is that Type 2 represents a variant male aurochs mor-
phology. Like Type 1 cores, Type 3 horns are found in levels
where metrical analysis of postcranial specimens has detected
no domestic cattle, and despite their smallness in compari-
son with Types 1 and 2, are within the size range of female
aurochsen.33 Current evidence thus supports their interpreta-
tion as wild females. Our interpretations—Type 1 as male,
Type 3 as female, Type 2 as probably male—are consistent
31. SYKES and SYMMONS, 2007.
32. ARMITAGE and CLUTTON-BROCK, 1976.
33. e.g. BÖKÖNYI, 1962.
019-032-Twiss.indd 24019-032-Twiss.indd 24 22/07/10 12:30:4022/07/10 12:30:40
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 7
Taking the Bull by the Horns: Ideology, Masculinity, and Cattle Horns at Çatalhöyük 25
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
with N. Sykes and R. Symmons’34 reevaluation of sexing crite-
ria provided by Armitage and Clutton-Brock35 and Armitage,
wherein basal measurements provide the most reliable sex
separation.36
A range of ages is represented in the horn core assemblage.
41 (by MNE; NISP = 118) specimens’ rough ages were assessed
based on core porosity, size, and/or articulation with other age-
able elements (table 2). Of these, 21 (MNE; NISP = 26) were
aged more precisely using Armitage37 (table 3). A minority
of the animals died while they were still immature, a slightly
greater proportion as subadults or young adults, most as mature
34. SYKES and SYMMONS, 2007.
35. ARMITAGE and CLUTTON-BROCK, 1976.
36. ARMITAGE, 1982.
37. Ibid.
adults, and one animal appears to have survived into old age.
Both right and left horn cores are represented in signifi cant
numbers (table 4). No pathologies have been found.
A pronounced majority of the Çatalhöyük horn cores come
from males (table 5). Of the 45 sexable specimens, 15 are clearly
male, and another 17 probably so. Only one specimen is cer-
tainly female and twelve more probably so. This translates to
a ratio of 71% male/probably male specimens to 29% female/
probably female specimens. This preponderance of males surely
explains the fact that more horns came from noticeably large
animals (n = 28) than from small ones (n = 7) (table 6). (The
majority of the specimens were either not particularly large or
small [“standard”], or were too fragmentary to judge.)
Table 1 – Ages of horn cores’ source animals. “Recorded Sex” represents the initial recording of sex in the CH database, to indicate how original records correlate with the interpretations presented here. *: type specimen; L: length in a straight line, base-tip; BA: maximum basal diameter; BB: minimum basal diameter; OC: outer curve; IC: inner curve. All measure-ments in mm.
Type Examples(Specimen numbers)
Site context Mellaart Level
Recorded Sex Age Side L BA BB OC IC
Type 1 M
Big, long, robust. Curves out from skull, dips down midway, rises at tip.
2375.X1* B.2, Sp. 117 IX M Adult R 113.6 62 730 595
3524.X1 B.3, Sp. 86 VII-VI M Adult L&R L: 132.1R:133.5 L: 530 L: 360
4121.X1 Sp. 181 VIII Poss. M Adult R 920 700
4497.X1 Sp. 115 VIII Poss. M Juvenile/Subadult R 659 470
7920.X1 B.49, Sp. 100 VII-VI M L 53 162 760 475
11940.X1 B.52, Sp. 94 V-IV M Adult L 110 88
11940.X4 B.52, Sp. 94 V-IV M Adult L 108* 725* 535*
Type 2 M
Relatively short, tightly curved, broad in rela-tion to length. Curves out from skull, does not dip in middle, rises only slightly at tip.
11955.X4* B.58, Sp. 227 IV-III M Adult R 40 88.6* 520 360
7920.X3 B.49, Sp. 100 VII-VI Poss. F L 37 105 510 350
7920.X2 B.49, Sp. 100 VII-VI Poss. M R 58 123.6 666 425
Type 3 F
Smallest, slim. Seems to have round cross-section at base (vs. ovoid bases of Types 1 and 2). Same curvature as Type 2.
11377.X1* Sp. 261 V? Poss. F Subadult/Adult L 40 80* 60* 400* 300*
13640.X1 B.49, Sp. 100 VII-VI Poss. F Subadult/Adult R
13194.X2 Sp. 312 VI-V Poss. F Adult L
6025.X2 KOPAL XI pre-XII Poss. F Adult L 73.9 62.2 410* 300*
019-032-Twiss.indd 25019-032-Twiss.indd 25 22/07/10 12:30:4122/07/10 12:30:41
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 8
26 K.C. TWISS and N. RUSSELL
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
Table 2 – Ages of horn cores’ source animals. “Young” includes all specimens that were clearly immature but could not be aged more precisely than that. The more precise age classes are as follows: roughly up to 1 year for infantile; 1-3 years for juvenile; 3-4 years for subadult; over 4 years for adult. “Old” does not have a specifi c age range attached.
Age All horn cores Special horn cores Non-special horn cores
NISP % NISP MNE % MNE NISP % NISP MNE % MNE NISP % NISP MNE % MNE
Young 57 48.3 1 2.4 3 9.1 1 3.4 54 65.1 0 0.0
Infantile 5 4.2 0 0 0 0.0 0 0 5 6.0 0 0.0
Juvenile 1 0.8 1 2.4 0 0.0 0 0 1 1.2 1 8.3
Juvenile/Subadult 3 2.5 2 4.9 1 3.0 1 3.4 2 2.4 1 8.3
Subadult 2 1.7 0 0 2 6.1 0 0 0 0.0 0 0.0
Subadult/Adult 14 11.9 5 12.2 4 12.1 4 13.8 10 12.0 1 8.3
Adult 34 28.8 29 70.7 22 66.7 23 72.4 10 12.0 8 66.7
Old 2 1.7 3 7.3 1 3.0 2 6.9 1 1.2 1 8.3
TOTAL 118 41 33 31 83 12
HORN CORES IN CONTEXT
Sizeable horn core pieces have been recovered from a wide
variety of contexts. Some were incorporated either visibly or
invisibly into the architecture; others come from intramural
or extramural fi ll. While some horns probably moved through
more than one context during their use-lives, we consider their
fi nal locations to be signifi cant.
The most famous of Çatalhöyük horn cores are architectural
installations. Mellaart38 recorded three variants: frontlets (horn
cores and the intervening skull) set in plaster heads attached to
walls; frontlets set in small pillars along the edges of platforms;
and pairs of horns set into benches. Notable recent fi nds include
a bench with three left horns protruding from its northern side,
two frontlets set into pillars edging a platform, and the remains
of a plastered skull set on the fl oor against a wall.39 The current
project has also found single horns set into walls and concealed
in architecture. Horns set in walls and benches would have been
on display, but many installed horns were essentially invisible,
integrated into the architecture. One house contained a cattle
horn plastered over to form the arm of a bin; another house’s
wall had a horn segment and two cattle scapulae laid between
its brick rows.
38. MELLAART, 1967.
39. TWISS et al., 2008.
Feasting remains are concentrations of large pieces of
bone, processed for marrow but not bone grease: this treatment
contrasts with that of most animal bone at Çatalhöyük, which
is more heavily processed. Feasting deposits are found chiefl y
as pockets in middens or in between-wall spaces, but also as
abandonment deposits. Some include cattle horns.
Many cattle horns appear to have been placed in houses
at abandonment or at demolition and fi lling. Some of these
deposits include partial or complete skulls; others focus more
exclusively on horns. Recent examples include a skull with one
horn shoved into an oven alongside a partially articulated dog
carcass; multiple horns plus a frontlet in a deposit that also
included a fragmentary human skull and an elaborate, bone-
handled fl int dagger; and two pieces of horn and a partial cattle
skull placed on a fl oor together with a collection of cattle scap-
ulae. Mellaart40 also alludes to cattle horns in what seem to be
abandonment deposits.
Some horn cores found on fl oors and in fi ll were coated
with plaster, suggesting that they are dismantled installations.
A few of these are associated with probable feasting deposits,
e.g., a feasting spread in Building 2 that includes a large piece
of horn core as well as a complete horn leaning against a bin.
The horn core retained some plaster on its base, and had many
chop marks along its corpus. The placement of these marks
makes no sense in terms of horn sheath removal (which entails
40. MELLAART, 1962: 51, 1963 and 1964.
019-032-Twiss.indd 26019-032-Twiss.indd 26 22/07/10 12:30:4122/07/10 12:30:41
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 9
Taking the Bull by the Horns: Ideology, Masculinity, and Cattle Horns at Çatalhöyük 27
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
Table 3 – ARMITAGE, 1982: 38, age stages for the Çatalhöyük horn cores. Armitage suggests that classes 0-2: immature animals; class 3: young adult; class 4: adult; classes 5-6: old adults; but these correlations are tentative.
Age class All horn cores Special horn cores Non-special horn cores
NISP % NISP MNE % MNE NISP % NISP MNE % MNE NISP % NISP MNE % MNE
0-2 0 0 0 0 0 0 0 0 0 0 0 0
3-4 4 15.4 3 14.3 1 7.7 1 9.1 3 23.1 2 18.2
4 5 19.2 4 19.0 1 7.7 0 9.1 4 30.8 4 36.4
5 12 46.2 9 42.9 9 69.2 6 54.5 3 23.1 3 27.3
6 5 19.2 5 23.8 2 15.4 3 27.3 3 23.1 2 18.2
TOTAL 26 21 13 10 13 11
Table 4 – Horn core siding.
Side All horn cores Special horn cores Non-special horn cores
NISP % NISP MNE % MNE NISP % NISP MNE % MNE NISP % NISP MNE % MNE
Right 48 39.0 28 46.7 21 45.7 16 39.0 27 35.1 12 63.2
Left 62 50.4 15 25.0 12 26.1 8 19.5 50 64.9 7 36.8
Right and left 13 10.6 17 28.3 13 28.3 17 41.5 0 0.0 0 0.0
TOTAL 123 60 46 41 77 19
Table 5 – Sexes of horn cores’ source animals. Sexing based on ARMITAGE and CLUTTON-BROCK, 1976. The right-hand column provides comparative information about sex ratios metrically derived from postcranial remains (see RUSSELL and MARTIN, 2005: 51-53 for methodological details).
SexAll horn cores Special horn cores Non-special horn cores Postcrania
in special contexts
Postcrania in non-special
contextsNISP % NISP MNE % MNE NISP % NISP MNE % MNE NISP % NISP MNE % MNE
Male 18 35.3 15 33.3 13 37.1 12 36.4 5 31.3 3 25.0 — —
Probably male 21 41.2 17 37.8 15 42.9 13 39.4 6 37.5 4 33.3 — —
Female 1 2.0 1 2.2 0 0.0 0 0 1 6.3 1 8.3 — —
Probably female 11 21.6 12 26.7 7 20.0 8 24.2 4 25.0 4 33.3 — —
Males and proba-ble males 39 76.5 32 71.1 28 80.0 25 75.8 11 68.8 7 58.3 59
(67.8%)36
(52.2%)Females and pro-bable females 12 23.5 13 28.9 7 20.0 8 24.2 5 31.3 5 41.7 28
(32.2%)33
(47.8%)
TOTAL 51 45 35 33 16 12 87 69
Table 6 – Sizes of horn cores’ source animals.
Size All horn cores Special horn cores Non-special horn cores
NISP % NISP MNE % MNE NISP % NISP MNE % MNE NISP % NISP MNE % MNE
Large 112 0.9 28 40.0 60 0.6 19 39.6 52 2.5 9 40.9
Standard 11,709 99.0 35 50.0 9,729 99.4 25 52.1 1,979 97.0 10 45.5
Small 12 0.1 7 10.0 2 0.0 4 8.3 10 0.5 3 13.6
TOTAL 11,833 70 9,791 48 2,041 22
019-032-Twiss.indd 27019-032-Twiss.indd 27 22/07/10 12:30:4122/07/10 12:30:41
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 10
28 K.C. TWISS and N. RUSSELL
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
chopping around the base), so we interpret these as scars from
the demolition of the installation to which it once belonged.
A few cattle horns are associated with commemorative
deposits.41 Building 1 has two such deposits, plus an abandon-
ment deposit on the fl oor. A complete female aurochs horn
lay on a bed of small stones against the building’s outer wall:
this deposit, which is stratigraphically associated with Build-
ing 1’s construction, also included a dog skull, two wild goat
horn cores, a stone macehead, and a crane wing modifi ed for
suspension.42 The second deposit was buried inside a platform,
and included a large chunk of cattle skull and horn core, sev-
eral large potsherds, and some cattle postcrania.
Animal remains are rarely found in human burials at
Çatalhöyük,43 and we know of only two cases involving cat-
tle horns. Both come from the East Mound’s latest Neolithic
levels, so they may signal a diachronic change in burial prac-
tice. The fi rst example consists of a cattle frontlet (within the
size range of female aurochs) with a human skull nestled under
its center, as though the person were wearing the frontlet as a
hat. Unfortunately the exact context is unclear, as a Hellenistic
pit disturbed the area adjacent to the human skull. There are a
few human bones on the other side of the pit, so it was probably
a complete body rather than an isolated skull. The disturbance
makes confi dence impossible, but the excavators could fi nd no
trace of a pit, and believe that the body and cattle frontlet were
placed in the room as it was fi lled. Another burial in the same
room contains a cattle horn.
Finally, several cattle horns and crania have been found in
midden deposits. These may simply be butchery waste, but some
occur in bone concentrations, suggesting that they are dismantled
installations or part of feasting deposits. Examples include one
midden’s complete horn core found associated with a concentra-
tion of cattle bones, and a separate midden’s bone concentration
containing cattle, sheep (possibly wild) and goat horn cores.
The variety of special treatments commonly accorded cat-
tle horns indicate that they were symbolically laden on mul-
tiple fronts. Indeed, the abundance and contextual variety of
Çatalhöyük’s horn cores suggest that horns played a key role
in multiple forms of ritual at the site. They are thus uniquely
well-suited to an investigation of the ideological signifi cance
of wildness, masculinity, and/or other biological traits. We
turn now to the biological traits that characterize horn cores
found in “special” contexts, contrasting them with those from
domestic assemblages.
41. RUSSELL et al., 2009.
42. RUSSELL and MCGOWAN, 2003.
43. RUSSELL and DÜRING, 2006.
THE CHARACTERISTICS OF HORN CORES FOUND IN “SPECIAL” CONTEXTS
We defi ne special contexts as feasting spreads, installa-
tions, and caches, and discuss them using MNE counts. Fifty-
three horn cores were deemed “special” or probably special,
a pronounced majority of the horn core assemblage (table 7:
the distinction between certainly and probably special is based
on reliability of context, e.g., a horned pillar installation [cer-
tainly] vs horns found by a collection of minimally processed
limb bones [probably]). Of the 48 cores that are certainly spe-
cial, 33 are sexable: 76% of these are male or probably male.
A (weaker) bias in favor of males (58%) is apparent among the
12 “non-special” horn cores (table 5).
41 of the special cores could be sided. 39.0% were rights,
19.5% lefts, and 41.5% were part of bucrania. These proportions
Table 7 – Proportions of horn cores deemed special (i.e., belonging to feasting spreads, installations, and caches).
Special? NISP % NISP MNE % MNE
Yes 9,791 82.7 48 67.6
Probably yes 161 1.4 5 7.0
No 1,336 11.3 5 7.0
Probably no 40 0.3 0 0.0
Maybe 505 4.3 13 18.3
TOTAL 11,833 71
Yes and probably yes 9,951 87.9 53 91.4
No and probably no 1,376 12.1 5 8.6
TOTAL 11,327 58
are different from those found among the 19 sidable non-
special cores (63.2% rights, 36.8% lefts, 0% bucrania). How-
ever, if one counts the two horns of each bucranium separately
as one right and one left, then the right: left ratio among the
special horns is 56.9%:43.1%, fairly similar to that seen among
the non-special horn cores. There is therefore a bias toward
right-hand horn cores in both groups; this may be attributable
to statistical variation in a limited-size assemblage.
Forty-eight special horn cores were assessed as to animal
size (table 6). 52.1% were from animals within the standard
size range for Çatalhöyük Bos, while 39.6% were from particu-
larly large specimens, and 8.3% from small animals. Again,
the ratios are similar among the 22 non-special horn cores:
45.5% standard-size, 40.9% large, and 13.6% small.
019-032-Twiss.indd 28019-032-Twiss.indd 28 22/07/10 12:30:4122/07/10 12:30:41
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 11
Taking the Bull by the Horns: Ideology, Masculinity, and Cattle Horns at Çatalhöyük 29
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
Twenty-nine special horn cores could be aged, as could
twelve non-special cores (tables 2-3). Prime-age and older
adults overwhelmingly dominated both samples, while very
few young animals were represented among either group. In
contrast, subadult and adult age groups formed about half of
the cattle remains assessed by dental criteria and epiphyseal
fusion, indicating either taphonomic loss of younger horn cores
or failure to transport these horns to the site.
In sum, no pronounced differences exist between the spe-
cial and non-special horn core assemblages. What differences
there are can plausibly be attributed to either (a) sampling bias
in moderately-sized assemblages (e.g., the modest differences
between the two groups with respect to sex, size, and age ratios),
or (b) defi nitional/taphonomic bias (e.g., the lack of bucrania
among the non-special cores. Bucrania are unlikely to remain
intact unless sheltered from weathering, carnivores, trampling
and repeated redeposition; they are far more likely to receive
such protection as special deposits such as installations and
abandonment deposits than they are in middens). Both the spe-
cial and the non-special groups emphasized mature animals
and males; biases toward large animals and right-hand-side
horn cores were also apparent.
DISCUSSION
Zoological analysis of the entire Çatalhöyük horn core
assemblage indicates that it is not a random sample from a
natural population. Signifi cant selection biases exist in favor
of mature animals and males. Since masculinity and size are
correlated in Bos, it is possible that the emphasis on large
specimens was merely a byproduct of the focus on males—an
idea supported by the fact that the majority of the horns in
the special as well as the non-special assemblages are from
medium-sized aurochsen. Horn symmetry also appears to have
been relevant. This conclusion is reinforced by two fi nds in a
single burned building: all of the sidable specimens in a cache
of at least 13 tightly-bunched horn cores found in the fi ll were
rights, while the three large horn cores protruding from a plas-
tered bench were all lefts.44
There are, however, only limited differences between
the biological characteristics of the horns discovered in spe-
cial contexts (i.e., abandonment deposits, ceremonial trash,45
architectural installations, and caches) and those discovered
44. TWISS et al., 2008.
45. HILL, 2000.
in ostensibly prosaic contexts such as middens. This suggests
that the inhabitants of Çatalhöyük either targeted adult male
aurochsen for slaughter, or they preferentially brought the
crania of such animals on site over those of females and
juveniles.
Horn core sex and age patterns do not match those of the
general Çatalhöyük Bos assemblage (nearly 50% females in
daily contexts [table 5], and 18-28% juveniles).46 Therefore, the
bias towards adult male horn cores does not straightforwardly
refl ect hunting strategies—all ages and both sexes were taken.
However, the postcranial remains do reveal a pronounced bias
toward larger animals (presumably males) in feasting/special
deposits (table 5).47 Juvenile cattle are also slightly more fre-
quent among early levels’ daily consumption remains than in
special contexts: they constitute 18% of these levels’ total cat-
tle remains, but 26% of those found in non-special contexts.48
No such bias is apparent in later levels, in which juveniles con-
tribute 28% of daily remains, and 28% of all remains.49 The
Çatalhöyük Bos assemblage thus indicates that while aurochs
cows and calves were regularly taken, bulls were distinctly
preferred for special activities.
This suggests that many of the site’s horn cores—not just
those from clearly special deposits—derive from feasting and/
or other ritualized activities. A signifi cant percentage of cows’
and calves’ horns must have been either deposited off-site or
processed in a fashion that rendered them archaeologically
unidentifi able.50 Among those horns that were brought on site,
additional selection for males is apparent among the special-
context specimens. It is therefore clear that large adult males
were specially selected for use in ritual contexts. However,
the inclusion of small but signifi cant numbers of females and
young animals among the special-context remains indicates
that large males were not absolutely required in all cases.
Bulls are larger and fi ercer than cows, and if a herd is man-
aged fewer adult males are required for herd security. Selection
for adult males suggests a desire for (a) maximally prepossess-
ing physical specimens, (b) maximally dangerous game ani-
mals, (c) relatively expendable herd animals, (d) maximum
meat yield, or (e) some combination of these motives. Given
that the Çatalhöyük cattle were wild,51 biological expendabil-
ity was probably less important to the villagers than physical
presence and/or danger, but beyond that we cannot at present
46. RUSSELL and MARTIN, 2005: 53-54.
47. Ibid., 2005.
48. Ibid.: fi g. 2.10.
49. Ibid.50. cf. ARMITAGE and CLUTTON-BROCK, 1976: 329.
51. RUSSELL and MARTIN, 2005.
019-032-Twiss.indd 29019-032-Twiss.indd 29 22/07/10 12:30:4122/07/10 12:30:41
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 12
30 K.C. TWISS and N. RUSSELL
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
say which factor(s) shaped the selection of bulls for feasts and
other ritual activities at Çatalhöyük.
Our fi ndings confi rm that at Çatalhöyük, the “people of the
bull” did, in fact, prefer adult males over female and juvenile
aurochsen for feasts and symbolic use. Bulls were not an abso-
lute prerequisite for ritual activity, however, indicating that
masculinity was not the exclusive factor in the site’s taurine
symbolism. It would be interesting to learn whether bulls were
similarly preferred—but not indispensable—at other sites
throughout the Neolithic Near East. As stated above, cattle
horns have been recovered at numerous Southwest Asian sites,
although in most cases little age/sex/morphology informa-
tion has been published about them. Simple visual assessment
of horn core images (comparing them, for example, with the
typology provided here) could provide some idea of male:
female ratios among those cores deemed noteworthy enough
to photograph or draw. This would be a biased sample, though:
truly understanding the importance of the bull to Neolithic
populations will require systematic study of all horns, from all
kinds of contexts, and from multiple sites.
Whatever the emic symbolic meaning of bulls, the slaughter
and consumption of such large and dangerous animals would
have endowed hunters and feasters with considerable status,
well worthy of commemoration through display and depic-
tion.52 Younger animals and females would have constituted
less of a challenge as well as provided less meat. Finally, adult
52. RUSSELL and MARTIN, 2005; TWISS, 2008.
bulls have the largest, most impressive horns: at Çatalhöyük,
these were preferred for display and likely embodied the power
of the bull.
ACKNOWLEDGMENTS
We thank A. Bogaard, A. Demirergi, J. Henecke, H. Malko, and
four anonymous reviewers for comments; J. Quinlan for the photos;
K. Killackey for the illustration; and M.P. Charles for the title. This
paper is based upon work supported by the National Science Founda-
tion under Grant No. 0647131. Above all, thanks to L. Martin, who
contributed extensively to getting this research off the ground and
to preliminary work on description of the Çatalhöyük cattle horn
cores.
Katheryn C. TWISSDepartment of Anthropology
Stony Brook UniversityStony Brook, NY 11794-4364
[email protected]
Nerissa RUSSELLDepartment of Anthropology
Cornell UniversityIthaca, NY 14853
[email protected]
BIBLIOGRAPHY
ALPASLAN-ROODENBERG S.
2001 Newly Found Remains from Menteşe in the Yenişehir Plain:
The Season of 2000. Anatolica 27: 1-14.
ARMITAGE P.L.
1982 A System for Ageing and Sexing the Horn Cores of Cattle from
British Post-Medieval Sites (17th to Early 18th Century) with
Special Reference to Unimproved British Longhorn Cattle.
In: WILSON B., GRIGSON C. and PAYNE S. (eds.), Ageing and Sexing Animal Bones from Archaeological Sites: 37-54. Oxford
(BAR Int. Ser. 109).
ARMITAGE P.L. and CLUTTON-BROCK J.
1976 A System for Classification and Description of the Horn Cores
of Cattle from Archaeological Sites. Journal of Archaeological Science 3,4: 329-348.
BÖKÖNYI S.
1962 Zur Naturgeschichte des Ures in Ungarn und das Problem der
Domestikation des Hausrindes. Acta Archaeologica 14,3-4:
175-214.
CAMPBELL S. and BAIRD D.
1990 Excavations at Ginnig, the Aceramic to Early Ceramic Sequence
in North Iraq. Paléorient 16,2 : 65-78.
CAUVIN J.
2000 The Birth of the Gods and the Origins of Agriculture. Cam-
bridge: Cambridge University Press.
FRAME S., RUSSELL N. and MARTIN L.
1999 Animal Bone Report (Hayvan Kemiği Raporu). www.catalhoyuk.com/archive_reports/1999/ar99_12.html
GORING-MORRIS A.N. and HORWITZ L.K.
2007 Funerals and Feasts during the Pre-Pottery Neolithic B of the
Near East. Antiquity 81: 902-919.
019-032-Twiss.indd 30019-032-Twiss.indd 30 22/07/10 12:30:4122/07/10 12:30:41
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 13
Taking the Bull by the Horns: Ideology, Masculinity, and Cattle Horns at Çatalhöyük 31
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
GRIGSON C.
1978 The Craniology and Relationships of Four Species of Bos: 4.
The Relationship between Bos primigenius Boj. and B. taurus L. and its Implications for the Phylogeny of the Domestic Breeds.
Journal of Archaeological Science 5,2: 123-152.
HAMILTON N.
2005 The Figurines. In: HODDER I. (ed.), Changing Materialities at Çatalhöyük: Reports from the 1995-1999 Seasons: 187-213.
Cambridge: McDonald Institute Monographs/British Institute
of Archaeology at Ankara.
HELMER D., GOURICHON L. et STORDEUR D.
2004 À l’aube de la domestication animale. Imaginaire et symbolisme
animal dans les premières sociétés néolithiques du Nord du
Proche-Orient. Anthropozoologica 39,1 : 143-163.
HILL E.
2000 The Contextual Analysis of Animal Interments and Ritual Prac-
tice in Southwestern North America. Kiva 65,4: 361-398.
HODDER I. (ed.)
2005a Changing Materialities at Çatalhöyük: Reports from the 1995-1999 Seasons. Cambridge: McDonald Institute Monographs/
British Institute of Archaeology at Ankara.
2005b Inhabiting Çatalhöyük: Reports from the 1995-1999 Seasons. Cambridge: McDonald Institute for Archaeological Research/
British Institute of Archaeology at Ankara.
2006a Çatalhöyük Perspectives: Themes from the 1995-1999 Seasons. Cambridge: McDonald Institute Monographs/British Institute
of Archaeology at Ankara.
2007 Excavating Çatalhöyük: South, North and KOPAL Area Reports from the 1995-1999 Seasons. Cambridge: McDonald Institute
Monographs/British Institute of Archaeology at Ankara.
HODDER I.
2006b Çatalhöyük: The Leopard’s Tale. London: Thames and Hudson.
LAST J.A.
2006 Art. In: HODDER I. (ed.), Çatalhöyük Perspectives: Themes from the 1995-1999 Seasons: 197-208. Cambridge: McDon-
ald Institute Monographs/British Institute of Archaeology at
Ankara.
LECHEVALLIER M. et RONEN A. (éd.)
1994 Le gisement de Hatoula en Judée occidentale, Israël. Paris :
Association Paléorient (Mémoires et travaux du Centre de recherche français de Jérusalem 8).
MALLOWAN M.
1946 Excavations in the Balikh Valley, 1938. Iraq 8: 111-159.
MELLAART J.
1962 Excavations at Çatal Hüyük, First Preliminary Report, 1961.
Anatolian Studies 12: 41-65.
1963 Excavations at Çatal Hüyük, Second Preliminary Report, 1962.
Anatolian Studies 13: 43-103.
1964 Excavations at Çatal Hüyük, Third Preliminary Report, 1963.
Anatolian Studies 49: 39-119.
1967 Çatal Hüyük: A Neolithic Town in Anatolia. London: Thames
and Hudson.
MOLIST M.
1998 Espace collectif et espace domestique dans le Néolithique
des 9e et 8e millénaires B.P. au Nord de la Syrie : apports du
site de Tell Halula (vallée de l’Euphrate). In : FORTIN M. et
AURENCHE O. (éd.), Espace naturel, espace habité en Syrie du Nord (10e-2e millénaires av. J.-C.). Actes du colloque tenu à l’université de Laval (Québec) du 5 au 7 mai 1997 : 115-130. Toronto : Canadian Society for Mesopotamian Societies.
ÖZDOĞAN A.
1999 Çayönü. In: ÖZDOĞAN M. and BAŞGELEN N. (ed.), Neolithic in Turkey: 35-64. Istanbul: Arkeoloji ve Sanat Yayınları.
ROLLEFSON G.O., SIMMONS A. and KAFAFI Z.
1992 Neolithic Cultures at ‘Ain Ghazal, Jordan. Journal of Field Archaeology 19,4: 443-470.
ROSENBERG M.
1994 Hallan Çemi Tepesi: Some Further Observations Concerning
Stratigraphy and Material Culture. Anatolica 20: 121-140.
RUSSELL N. and DÜRING B.S.
2006 Worthy is the Lamb: A Double Burial at Neolithic Çatalhöyük
(Turkey). Paléorient 32,1 : 73-84.
RUSSELL N. and MARTIN L.
2005 The Çatalhöyük Mammal Remains. In: HODDER I. (ed.), Inha-biting Çatalhöyük: Reports from the 1995-1999 Seasons: 33-98.
Cambridge: McDonald Institute for Archaeological Research.
RUSSELL N. and MCGOWAN K.J.
2003 Dance of the Cranes: Crane Symbolism at Çatalhöyük and
Beyond. Antiquity 77,297: 445-455.
RUSSELL N. and MEECE S.
2005 Animal Representations and Animal Remains at Çatalhöyük.
In: HODDER I. (ed.), Çatalhöyük Perspectives: Themes from the 1995-1999 Seasons: 209-230. Cambridge: McDonald Institute
for Archaeological Research/British Institute of Archaeology at
Ankara.
RUSSELL N., MARTIN L. and BUITENHUIS H.
2005 Cattle Domestication at Çatalhöyük Revisited. Current Anthro-pology 46,S5: S101-S108.
RUSSELL N., MARTIN L. and TWISS K.C.
2009 Building Memories: Commemorative Deposits at Çatalhöyük.
Anthropozoologica 44,1: 103-128.
SIMMONS A.H. and NAJJAR M.
1999 Preliminary Field Report of the 1998-1999 Excavations at
Ghwair I, a Pre-Pottery Neolithic Community in the Wadi Fei-
nan Regions of Southern Jordan. Neo-Lithics 1/99: 4-6.
SYKES N. and SYMMONS R.
2007 Sexing Cattle Horn-Cores: Problems and Progress. Internatio-nal Journal of Osteoarchaeology 17,5: 514-523.
TESTART A.
2006 Interprétation symbolique et interprétation religieuse en archéo-
logie. L’exemple du taureau à Çatal Höyük. Paléorient 32,2 :
23-57.
TSUNEKI A.
2002 A Neolithic Foundation Deposit at Tell ‘Ain el-Kerkh. In:
GEBEL H.G.K., HERMANSEN B.D. and JENSEN C.H. (eds.),
Magic Practices and Ritual in the Near Eastern Neolithic: 133-
144. Berlin: ex oriente.
019-032-Twiss.indd 31019-032-Twiss.indd 31 22/07/10 12:30:4122/07/10 12:30:41
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
Page 14
32 K.C. TWISS and N. RUSSELL
Paléorient, vol. 35.2, p. 19-32 © CNRS ÉDITIONS 2009
TWISS K.C.
2008 Transformations in an Early Agricultural Society: Feasting
in the Southern Levantine Pre-Pottery Neolithic. Journal of Anthropological Archaeology 27,4: 418-442.
TWISS K.C., BOGAARD A., BOGDAN D., CARTER T., CHARLES M.P.,
FARID S., RUSSELL N., STEVANOVIĆ M., YALMAN E.N. and YEOMANS L.
2008 Arson or Accident? The Burning of a Neolithic House at Çatal-
höyük. Journal of Field Archaeology 33,1: 41-57.
UERPMANN H.-P.
1999 Der Rückzucht-Auerochse und sein ausgestorbenes Vorbild. In:
WENIGER G.-C. (ed.), Archaeology and Biology of the Aurochs:
93-102. Neandertal: Neandertal Museum.
VAN VUURE C.
2005 Retracing the Aurochs: History, Morphology and Ecology of an Extinct Wild Ox. Sofia (Bulgaria): Pensoft Publishers.
VERHOEVEN M.
2002 Ritual and Ideology in the Pre-Pottery Neolithic B of the Levant
and Southeast Anatolia. Cambridge Archaeological Jour-nal 12,2: 233-258.
YARTAH T.
2004 Tell ‘Abr 3, un village du Néolithique précéramique (PPNA) sur le
moyen Euphrate. Première approche. Paléorient 30,2 : 141-158.
019-032-Twiss.indd 32019-032-Twiss.indd 32 22/07/10 12:30:4122/07/10 12:30:41
CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010
• CN
RS
ED
ITIO
NS
201
0 • C
NR
S E
DIT
ION
S 2
010