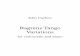Tapping to Ragtime: Cues to Pulse Finding Author(s): Joel Snyder and Carol L. Krumhansl Source: Music Perception: An Interdisciplinary Journal, Vol. 18, No. 4 (Summer, 2001), pp. 455-489 Published by: University of California Press Stable URL: http://www.jstor.org/stable/40285847 Accessed: 03/12/2010 05:47 Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at http://www.jstor.org/action/showPublisher?publisherCode=ucal. Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. University of California Press is collaborating with JSTOR to digitize, preserve and extend access to Music Perception: An Interdisciplinary Journal. http://www.jstor.org
Transcript
Tapping to Ragtime: Cues to Pulse FindingAuthor(s): Joel Snyder and Carol L. KrumhanslSource: Music Perception: An Interdisciplinary Journal, Vol. 18, No. 4 (Summer, 2001), pp.455-489Published by: University of California PressStable URL: http://www.jstor.org/stable/40285847Accessed: 03/12/2010 05:47
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available athttp://www.jstor.org/page/info/about/policies/terms.jsp. JSTOR's Terms and Conditions of Use provides, in part, that unlessyou have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and youmay use content in the JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained athttp://www.jstor.org/action/showPublisher?publisherCode=ucal.
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
University of California Press is collaborating with JSTOR to digitize, preserve and extend access to MusicPerception: An Interdisciplinary Journal.
JOEL SNYDER & CAROL L. KRUMHANSL Cornell University
Two experiments investigated cues to pulse finding using a relatively unconstrained, naturalistic paradigm. Participants tapped what they felt was a comfortable pulse on a keyboard playing a percussive sound. The stimulus materials were based on ragtime excerpts, played metronomically (i.e., without expressive timing or tempo variation). The first experiment, with 8 musically experienced and 8 musically inexperienced subjects, played each excerpt in two versions: a pitch-varied version (the original excerpt) and a monotonic version (with all tones changed to middle C) that was designed to remove all melodic and harmonic cues to pulse. Neither the absence of pitch information nor musical experience signifi- cantly affected performance. The second experiment tested 12 musically experienced subjects on shorter excerpts from the same ragtime pieces. Full (right-hand and left-hand parts together) and right-hand-only ver- sions of the excerpts were each played in pitch-varied and monotonic versions. Removing the left-hand part significantly affected tapping per- formance on a number of measures, causing a lower percentage of tap- ping on the downbeat, more off-beat taps, more aperiodic taps, more switches between tapping modes, a higher variability of the intertap in- terval, and larger deviations from the beat. As a whole, these indicate a negative effect of removing the left-hand part. Again, differences between pitch- varied and monotonic versions were generally small. Analysis of the music revealed the following cues to pulse finding: a predictable al- ternating bass pattern in the left-hand part and a majority of notes on metrically strong positions in both the right-hand and left-hand parts. These results suggest that, for piano ragtime music, temporal cues are prominently available for finding and following the pulse and that pulse finding is largely independent of pitch information. Implications of the experimental measures and music-analytic techniques for models of pulse perception are considered.
Received March 30, 2000; accepted December 21, 2000.
Pulse finding is a basic ability used during musical listening, performance,
and dancing. Pulse, which can be established after 2-4 cycles (Desain & Honing, 1999; Fraisse, 1982), serves as a basic rhythmic unit allowing
Address correspondence to Joel Snyder, Department of Psychology, Uris Hall, Cornell University, Ithaca, NY 14853. (e-mail: [email protected])
ISSN: 0730-7829. Send requests for permission to reprint to Rights and Permissions, University of California Press, 2000 Center St., Ste. 303, Berkeley, CA 94704-1223.
455
456 Joel Snyder & Carol L. Krumhansl
listeners, performers, and dancers to synchronize with music and each other. Evidence that such periodic behavior is ubiquitous comes from studies of tapping to simple acoustic patterns (Essens &c Povel, 1985; Helmuth &c Ivry, 1996; Ivry & Hazeltine, 1995; Jones & Pfordresher, 1997; Mates, Radii, & Poppel, 1992; Parncutt, 1994; Povel, 1981, 1984, 1985; Povel & Collard, 1982; Povel & Essens, 1985; Robertson et al., 1999; Vos, Mates, &C van Kruysbergen, 1995), tapping to music (Drake, Penel, & Bigand, 2000; Scheirer, 1998; van Noorden & Moelants, 1999; Vos, van Dijk, & Schomaker, 1994), tapping to polyrhythms (Deutsch, 1983; Klapp, Hill, Tyler, Martin, Jagacinski, &c Jones, 1985; Peper, Beek, & van Wieringen, 1995), speech rhythms (Cummins & Port, 1998), limb coordination (Diedrich & Warren, 1995; Schmidt, Beek, Treffner, &: Turvey, 1991; Schmidt, Bienvenu, Fitzpatrick, & Amazeen, 1998), time perception (Large & Jones, 1999; McAuley, 1995; McAuley & Kidd, 1998), and infants' postural adjustments (Bertenthal, Rose, & Bai, 1997; Schmuckler, 1997). In addition, theorists have hypothesized about the possible connections among pulse perception, time perception, motor behavior, and locomotion (Large & Jones, 1999; Parncutt, 1987; Todd, 1994, 1999; Todd, O'Boyle, &C Lee, 1999). Computational approaches to pulse include applying a set of rules for updating the current pulse hypothesis (Desain & Honing, 1999; Longuet-Higgins & Lee, 1982; Steedman, 1977), using equations that cal- culate the pulse strength on the basis of musical features (Parncutt, 1994), implementing mechanisms that oscillate in synchrony with the pulse (Eck, Gasser, & Port, 2000; Gasser, Eck, &c Port, 1999; Large, 1994; Large &c Kolen, 1994; Large & Jones, 1999; McAuley, 1995; Toiviainen, 1998), and simulating filtering mechanisms that extract periodicities in the music (Scheirer, 1998; Todd, 1994, 1999; Todd et al., 1999).
Past experimental and computational studies of pulse finding in music generally assume that only temporal information is important for pulse perception (but see Steedman, 1977). It is clear from behavioral and com- putational studies that temporal information is often sufficient for pulse finding, but any additional contribution of pitch information is largely unknown. Few behavioral experiments use musical stimuli to address this question, and even fewer make experimental manipulations to determine what cues listeners use. Another limitation of some past studies is that their experimental measures do not assess multiple aspects of performance. In other words, few studies combine measures of tapping period, tapping phase, and timing variability, thus giving a limited description of pulse-tracking performance. The following review focuses on investigations of meter per- ception and synchronization with auditory patterns, particularly focusing on the possible role of pitch in perceiving temporal structure, the perfor- mance measures used, and benchmarks suggested by the studies to date. Studies using musical and nonmusical patterns will be considered, in addi- tion to studies using perceptual and perceptual-motor tasks.
Tapping to Ragtime 457
The research of Dawe, Platt, and Racine (1993, 1994, 1995) has pointed to the importance of harmonic changes in rhythm perception. The first two studies pitted harmonic, temporal (Dawe et al., 1993, 1994), and melodic (Dawe et al., 1994) cues against one another by varying the position of a chord change in a repeating sequence consisting of four short durations and a long duration. Subjects identified the perceived rhythmic pattern (Dawe et al., 1993) and identified the meter (Dawe et al., 1994). Harmonic factors primarily determined the judgments in each study, and musicians weighted harmonic information more heavily than did nonmusicians. The final study in this series (Dawe et al., 1995) quantitatively modeled the contributions to meter perception of temporal accents, the onset of the long-duration note, the termination of the long-duration note, and the po- sition of the harmonic change. The latter factor showed a consistent differ- ence between nonmusicians and musicians, with the harmonic change be- ing the primary influence on musicians' judgments of rhythm. These results suggest that removing pitch information would diminish the ability to find and follow the pulse in music, especially for musicians. Such a finding would be a major challenge to models of pulse finding that rely solely on temporal information.
To address the problem of synchronizing with performed music, Drake et al. (2000) asked musicians and nonmusicians to tap to six classical piano works, each in three different versions. The versions were mechanical (with no timing or amplitude variations), accented (with 10-dB-amplitude ac- cents for notes on the first beat of the measure), and expressive (performed by a professional pianist in a "natural, musical fashion"). Musicians showed a higher percentage of successful synchronization across pieces (i.e., at least nine consecutive intertap intervals [ITIs] within 10% of the beat duration), a higher percentage of ITIs within 10% of the beat duration in measures 2- 11, a higher percentage of ITIs within 10% of the beat duration in the 10 most successful measures for each piece, a tendency to tap at a slower rate, more metrically consistent tapping, and greater flexibility in synchroniza- tion rate. In measures 2-11 and in the most successful 10 measures, sub- jects performed better for the mechanical and accented versions than for the expressive version. These results suggest that expressive music can cause difficulty in some aspects of pulse finding, that musicians have a synchro- nization advantage over nonmusicians, and that musicians focus on higher metrical levels than do nonmusicians.
In another study of musical synchronization, Vos et al. (1994) had sub- jects tap at the beginning of the perceived measure for excerpts from solo instrumental pieces by J. S. Bach. Subjects differed in judged measure length (although subjects usually tapped at the first metrical position) and showed increasing judged measure length as a function of tempo. Parncutt (1994) similarly found the phase and period of tapping to differ across subjects for rhythmic monotonic stimuli, suggesting that ambiguity is a general phe-
458 Joel Snyder & Carol L. Krumhansl
nomenon of pulse and meter. Vos et al. (1994) also tested whether subjects' judgments of meter could be explained by autocorrelating the melodic in- terval series (see also Brown, 1993). The metrical cues for period length found with autocorrelation predicted subjects' responses poorly, casting doubt on whether this particular melodic cue is used by subjects.
An important early study by Essens and Povel (1985) examined the re- production of monotonic patterns consisting of short and long interonset intervals (see also Povel, 1981). Subjects accurately reproduced patterns in which the long and short temporal intervals had a 2:1 ratio, but their re- productions of patterns with other ratios gravitated toward interval ratios of 2:1. This was even the case for stimuli with interval ratios of 3:1, which is a relatively simple ratio. This study suggests that subjects can better cognitively represent patterns with 2:1 interval ratios and that patterns may be misrepresented when they contain other ratios. These results could be interpreted as an inability to establish a pulse when time-interval ratios are larger or smaller than 2:1.
A recent study by van Noorden and Moelants (1999) asked subjects to tap to a wide range of music (e.g., 16th century Flemish polyphony, classic jazz, and contemporary popular music from the radio). Analyses of the tapping tempo showed partial dependence on style and a bias for tapping at rates around 500 ms ITI. This result is consonant with Fraisse's (1982) review of findings showing that preferred tempo, spontaneous tempo, and the indifference interval (i.e., the point of most veridical time perception) each resides in the 500-700 ms range.
In another study of musical synchronization (Scheirer, 1998), subjects tapped to seven performed musical excerpts, each from a different style. The author evaluated two measures, root-mean-square (RMS) deviation from the beat (as judged by the author) and the variance of ITI. These measures respectively index absolute deviation from the beat and consis- tency of the tapping period. Although there were large differences between subjects and between stimuli on RMS deviation from the beat, a represen- tative value was around 60 ms. Subjects differed from each other on vari- ance of ITI but were fairly consistent across the different stimuli. The range of values for this measure was around 10-100 ms.
Vos et al. (1995) examined the phenomenon known as negative asynchrony , the tendency to tap slightly ahead of a series of isochronous auditory stimuli. The negative asynchrony was reduced when the stimuli were longer in duration and when their rise times were longer. The authors concluded that subjects synchronize to a perceptual center rather than to the physical onset of a tone. Recently, Wohlschlager and Koch (2000) re- ported that filling the interval between the isochronous pacing tones with additional tones or with bodily movements greatly reduced the negative asynchrony. Thus, one might not expect to observe negative asynchrony while tapping to music containing notes in between perceived pulses.
Tapping to Ragtime 459
To summarize, the above studies represent a considerable body of re- search on synchronization to auditory stimuli. Some studies suggest a pos- sible role of pitch information in temporal perception. Performance mea- sures developed by studies of sensory-motor synchronization include indices of the phase and period of tapping, variability of the tapping period, the magnitude of anticipation, and the magnitude of deviation from the target. They suggest the following generalizations concerning tapping: a prefer- ence toward simple ratios, a preferred tapping rate in the range of 500-700 ms, a tapping period that may not correspond to the notated musical meter, a tendency to tap slightly ahead of pacing tones defining isochronous inter- vals, and a pulse-finding advantage with greater musical experience. With these results in mind, we now turn to the present experiments.
The two experiments reported next asked listeners to tap at a comfort- able rate to relatively complex extended musical excerpts based on piano ragtime pieces. Although during its most active period (ca. 1890-1930) ragtime was dominated by vocal music, the piano ragtime of Scott Joplin and his contemporaries is the only form that is widely known today (Ber- lin, 1980, 1985; Jasen & Tichenor, 1978). Piano ragtime music has a num- ber of features that were desirable for our purposes (see top of Figure 1). It is written in duple meter and is played at a moderate tempo suitable for dancing (originally cakewalks, marches, and two-steps). Its form is rela- tively obvious, consisting of pairs of 16-measure sections that subdivide into 4-measure phrases. The final of these four phrases typically contains a clear full cadence. The harmony is conventional, based largely on tonic, dominant, and subdominant chords in major. The left-hand part, often consisting of alternating bass notes and chords, defines simple, strong har- monic progressions and a clear duple meter. However, the right-hand melody is often syncopated against the left-hand accompaniment, providing me- lodic and rhythmic interest.
Fig. 1. The introduction from Glad Cat, showing pitch-varied and monotonic versions. For the monotonic version, all notes from the pitch-varied version are simply changed to middle C (duplicated notes are not shown).
460 Joel Snyder & Carol L. Krumhansl
One of our central objectives was to study the contributions of pitch and temporal cues to pulse finding, taking advantage of the characteristics of ragtime just described. The first experiment presented the excerpts in the original form and in a monotonic version (with all notes of the original changed to the same note, middle C). Removing the pitch variation in this way might have a large effect on tapping performance given the very clear harmonic structure of ragtime music. This experiment also compared two groups of listeners, musicians and nonmusicians, to assess possible effects of musical training on performance. The results might differ between these groups because musicians presumably have greater knowledge of harmony, meter, and musical form, as well as possibly more precise motor control than nonmusicians. The second experiment, using musicians only, again presented the music with and without pitch variation. In this experiment, the excerpts either contained both the regular left-hand part and the more syncopated right-hand part, or only the right-hand part.
A second objective of the study was to develop a battery of performance measures. These were all based on the tap time recorded by the computer. The first performance measure was tapping mode, illustrated in Figure 2, for the duple meter of the materials used in this study. There are four pos- sibilities: tapping on the downbeat, tapping on the upbeat, tapping off- beat (periodically but on neither the downbeat nor the upbeat), and tap-
Fig. 2. Subjects could tap in four different modes. Three of these were periodic: tapping on the downbeat (indicated by D), the upbeat (indicated by U), or off-beat. The other mode was aperiodic. Asterisks indicate tap positions.
Tapping to Ragtime 461
ping aperiodically. To determine if the tapping was to the downbeat or the upbeat, we matched each tap where possible to the closest downbeat posi- tion (the first and third eighth-note metrical positions) or upbeat position (the second and fourth eighth-note metrical positions) that was within 100 ms of the tap. Tapping off-beat meant tapping with a steady ITI but not near the downbeat or the upbeat (i.e., not within 100 ms of either). Aperi- odic taps included single deviant taps and prolonged tapping with an in- consistent or drifting period. We analyzed each participant's tapping per- formance for tapping mode throughout the excerpt, making it possible to determine the percentage of time spent in each tapping mode and the num- ber of switches between tapping modes. We defined switches as changes to a new tapping mode for at least two consecutive taps, and did not include changes to or from aperiodic tapping.
Figure 3 illustrates the other calculated performance measures. For these, we only used periodic taps. The first of these was based on the ITIs, shown in the top of Figure 3. This performance measure, called the coefficient of variability (CV), was the standard deviation of the ITIs, divided by the mean ITI. We excluded ITIs that were 100 ms greater or lesser than the
Fig. 3. Intertap interval (ITI) is the average time in milliseconds between taps (top). Coeffi- cient of variability (CV) is the SD of ITI, divided by mean ITI. Beats to start tapping (BST) is the time in milliseconds from the first note of a trial to the first periodic tap, as a propor- tion of the quarter-note interbeat interval (middle). In the example shown, BST is equal to 2, since downbeats correspond to the quarter-note level. Coefficient of delay (CDel) is the average time deviation in milliseconds of a tap from the nearest matched beat, as a propor- tion of mean ITI. Coefficient of deviation (CDev) is the average absolute time deviation in milliseconds of a tap from the nearest matched beat, as a proportion of mean ITI (bottom).
462 Joel Snyder & Carol L. Krumhansl
tapped interbeat interval (IBI) to reduce the effect of outliers. CV was a measure of the consistency of tapping independent of the relationship be- tween the taps and the events in the music. Shown in the middle of Figure 3 is beats to start tapping (BST), defined as the time in milliseconds from the first note of the stimulus to when subjects began tapping periodically, divided by the quarter-note IBI. This assessed how much musical informa- tion the subject needed to find the pulse, controlling for musical tempo. The final measures were based on the deviations of the tap from the corre- sponding events in the music, shown at the bottom of Figure 3. The first of these, called the coefficient of delay (CDel), was the average time between a tap and the matched beat (downbeat, upbeat, or off-beat), divided by the mean ITI. A positive value meant that the tap occurred after the corre- sponding beat, whereas a negative value indicated that the tap occurred before the beat. The second, called the coefficient of deviation (CDev), was the average absolute time between the tap and the closest beat divided by the mean ITI, a measure of deviation from the beat independent of sign (i.e., delay or anticipation). For CDel and CDev, we only used taps within 100 ms of a downbeat, an upbeat, or some other metrical position (i.e., off- beat). We normalized the three coefficients (CV, CDel, CDev) using the subjects' mean ITI to control for tapping rate.
Experiment 1
This experiment used the eight ragtime excerpts listed in Appendix A. They were played at slightly different tempos, but each had a quarter-note IBI in a comfortable tapping range. Appendix A also shows the form of the selections, which always ended at a section boundary so that the excerpts were of approximately equal duration and avoided awkward endings. The music was played in a metronomic fashion, with no expressive timing or tempo variations. These are of considerable interest in that they reflect structural characteristics of the music (see, for example, the review by Palmer, 1997). However, timing variability was not included in the current study to avoid complicating our analyses of tapping consistency with respect to pre- vious taps and to events in the music. Two versions of each excerpt were presented, one with pitch variations and the other monotonic. Musically experienced and inexperienced subjects participated in order to assess the effects of musical training.
METHOD
Subjects
Eight musically experienced (male = 2, female = 6; mean years playing music = 13) and eight musically inexperienced (male = 4, female = 4; mean years playing music = 3) Cornell
Tapping to Ragtime 463
University students participated for extra credit in psychology courses. No subject reported hearing problems, and two reported having absolute pitch. On a 7-point scale, the average familiarity with ragtime music was 2.6 for musically experienced subjects and 2.2 for inex- perienced subjects. One other subject participated but was unable to perform the task, and two other subjects' data were incomplete because of computer malfunction.
Materials and Stimuli
We selected eight Musical Instrument Digital Interface (MIDI) piano ragtime pieces in 2/4 meter (see Appendix A), which are available from the corresponding author upon re- quest. The tempo range was 587-697 ms quarter-note IBIs, or 86-102 beats per minute. For each MIDI file, there was a constant time between the offset and onset of adjacent notes (range across excerpts = 3.5-8.5% of the quarter-note IBI). All stimuli were played to sub- jects using an object-oriented MAX interface on a Macintosh Quadra 680AV computer controlled by the experimenter, a Yamaha TX816 MIDI rack, a Yamaha 1204 mixing con- sole, and AKG K-141 headphones.
We prepared each excerpt using Vision, a MIDI editing program, to have the following characteristics: (1) metronomic timing, (2) equalized MIDI velocity (related to acoustic amplitude and spectrum), (3) MIDI voice set to a synthesized piano timbre, (4) beginning at the start of the original piece, (5) lasting approximately 1.5 min in duration, and (6) termi- nating at a section end. We created two versions of each excerpt, as illustrated in Figure 1 for the beginning of one of the excerpts: (1) the original version with the previously men- tioned modifications, and (2) one identical to the original but with all notes changed to middle C. For this experiment, we term the first of the versions as pitch varied, and the second as monotonic. All melodic and harmonic cues to pulse in the lower and the upper voices were absent in the monotonic versions. Although not shown in Figure 1, we dupli- cated Cs when multiple tones appeared in the original, so the two versions had the same number of tones. Duplication of simultaneous identical tones results in increased loudness compared with single tones in MAX.
Procedure
The experimenter asked each subject to "tap the beat of the music with the index finger of your dominant hand on the keyboard the way you normally tap your foot while listening to music" and to "find the most comfortable beat but do not begin tapping until you have found the beat mentally." Subjects tapped on a Yamaha KX88 keyboard, with tap times collected by the MAX interface.1 A percussive hi-hat sound resulted from subjects' tapping on the keyboard. Before the experiment began, subjects practiced tapping to a pitch- varied version and a monotonic version of an excerpt from a ragtime piece not used in the experi- ment. All subjects tapped to both versions of each of the eight ragtime excerpts. Prior to each experimental session, the experimenter chose a random order for the eight excerpts. The MAX interface ran through this order twice, interleaving the two versions of the eight excerpts. For example, if the first two numbers selected were 5 and 3, the first stimuli presented would be the pitch-varied version of excerpt 5 and the monotonic version of excerpt 3. After eight trials, the order would repeat with excerpt 5 presented in its mono- tonic version, followed by excerpt 3 in its pitch-varied version. The experimenter counter- balanced whether a pitch-varied version or a monotonic version came first, across subjects.
1. To test the timing accuracy of MAX, we constructed, played, and recorded the output for an isochronous series of tones with 600-ms interonset intervals. Testing showed that the interonset intervals had an average absolute deviation of ~1 ms from the specified value and a maximum deviation of 5 ms. Thus, most of the effect sizes and means we report exceed the temporal resolution of MAX. In addition, MAX played the 600 ms interonset intervals ~1 ms too fast.
464 Joel Snyder & Carol L. Krumhansl
RESULTS
A mixed-design analysis of variance (ANOVA) for each of the depen- dent measures determined the effects of pitch information, musical experi- ence, and excerpt. For effects including excerpt, we adjusted the degrees of freedom using the Greenhouse-Geisser epsilon to correct for violations of sphericity. The results of the statistical tests appear in Table 1.
The top of Figure 4 shows the percentage of taps that were on the down- beat, the upbeat, the percentage of off-beat taps, and the percentage of aperiodic taps. About 80% of the time, subjects tapped on the downbeat, and most of the remaining time they tapped on the upbeat. For percentage of taps on the downbeat, there were no effects of pitch information, F(l, 14) = 1.34, p > .05, or musical experience, F(l, 14) = .13, p > .05, but there was a significant effect of excerpt, F(7, 98) = 4.95, e = .579, p < .005. Additionally, pitch information and excerpt significantly interacted, P(7, 98) = 4.87, e = .658, p < .005. Significant main effects and interactions with excerpt are described in more detail later.
About 2% of the taps were aperiodic. Most of these came in small groups; others were single deviant taps. Neither pitch information, F(l, 14) = .13, p > .05, nor musical experience, JF(1, 14) = 3.18, p > .05, had a significant effect on this variable. Excerpts, however, differed from each other, P(7, 98) = 3.55, e = .420, p < .025. In this experiment, no subject tapped off- beat.
The bottom of Figure 4 displays the average number of switches in tap- ping mode, which occurred very infrequently. Neither the presence of pitch information, F(l, 14) = .53, p > .05, nor musical experience, F(l, 14) = .23, p > .05, affected the number of switches. There was also no significant difference among excerpts, P(7, 98) = 2.65, e = .345, p > .05. To examine possible effects of transitions from one section of the music to the next, we compared the number of switches and aperiodic taps near and far from section transitions. In the music used, 23.3% of the time was near a section transition, defined as within 4 s of a transition from one section of the music to another. Accordingly, 23.3% of the switches and aperiodic taps should occur near the transitions if they are randomly distributed. Signifi- cantly more switches, x2(l> N = 102) = 12.70, p < .001, and aperiodic taps, X2(l, N = 753) = 65.15, p < .001, were within 4 s of the section transitions than would be expected by chance.
The top of Figure 5 shows the proportion of musically experienced and inexperienced subjects tapping at different numbers of beats after the start of the music for the pitch-varied and monotonic versions. The bottom of Figure 5 shows this information in terms of mean BST. It appears as though musically experienced subjects began tapping earlier, but this effect was not statistically reliable, P(l, 14) = 4.45, p = .053. Neither was pitch infor-
Tapping to Ragtime 465
Fig. 4. Percentage of taps on the downbeat, upbeat, percentage of off-beat taps, and per- centage of aperiodic taps for musically experienced and inexperienced subjects in Experi- ment 1 (top). Mean switches in tapping mode (+SE) for musically experienced and inexpe- rienced subjects for pitch-varied and monotonic stimuli in Experiment 1 (bottom).
mation significant, F(l, 14) = 3.04, p > .05, although there was a signifi- cant effect of excerpt, F(7, 98) = 3.46, 8 = .360, p < .05.
Subjects tapped with an ITI of -600 ms (corresponding to the quarter- note level) on 87% of the trials, -300 ms on 9% of the trials, and -1200 ms on 3% of the trials. The CV, shown at the top of Figure 6, was about 4% of the tapping period or about 24 ms. There was no effect of pitch
466 Joel Snyder 8c Carol L. Krumhansl
Fig. 5. Experiment 1 beats to start tapping (BST). Proportion of trials in which subjects started tapping at different elapsed quarter-note beats from the first note of the trial for musically experienced and inexperienced subjects for pitch-varied and monotonic stimuli (top). Mean BST (+SE) for musically experienced and inexperienced subjects for pitch- varied and monotonic stimuli (bottom).
information, F(l, 14) = .98, p > .05, or musical training, F(l, 14) = 1.76, p > .05. Individual excerpts differed from each other on CV, F(7, 98) = 2.79, e = .523, p < .05. However, no interactions appeared with any combination of pitch information, musical experience, and excerpt.
Tapping to Ragtime 467
Fig. 6. Mean timing coefficients (+SE) in Experiment 1. Coefficient of variability (CV, top), coefficient of delay (CDel, middle), and coefficient of deviation (CDev, bottom) shown for
musically experienced and inexperienced subjects for pitch-varied and monotonic stimuli.
The middle of Figure 6 shows CDel to be near 1 % of the tapping period, corresponding to a tap delay of approximately 6 ms. There were no effects of pitch information, F(l, 14) = .00, p > .05, musical experience, F(l, 14) =
.13, p > .05, or excerpt, F(7, 98) = 3.12, e = .274, p > .05. As seen in the bottom of Figure 6, CDev was generally around 4% of the tapping period
468 Joel Snyder &; Carol L. Krumhansl
or 24 ms. No significant effects of pitch information, F(l, 14) = 1.83, p > .05, or musical experience, F(l, 14) = 1.02, p > .05, surfaced on this mea- sure, although excerpt was significant, F(7, 98) = 3.47, e = .395, p < .05.
DISCUSSION
Subjects in Experiment 1 demonstrated the following general pulse-find- ing behavior. Approximately 80% of all taps were on the downbeat, most of the remaining taps were on the upbeat, and only about 2% were aperi- odic taps. Switches between modes of tapping were rare (much less than once per trial on average), and when they occurred they were often near transitions from one section of the music to the next. Subjects tapped about 90% of the time with an ITI of approximately 600 ms, sometimes at twice that rate, and very rarely at half that rate. The BST was about 5 on average for musically experienced subjects and about 9 for musically inexperienced subjects. The variability of the tapping period, CV, was near 4% of the tapping period (-24 ms). On average, tapping occurred just slightly behind the beat, by about 1 % of the tapping period (~6 ms), as measured by CDel. Finally, subjects' average tap deviation from the beat in absolute values, CDev averaged around 4% of the tapping period (-24 ms).
As shown in Table 1, we found no differences between pitch-varied and monotonic versions on any of the performance measures. The lack of dif- ferences between pitch-varied and monotonic versions was unanticipated given previous experimental results (Dawe et al., 1993, 1994, 1995) and
Table 1 Summary of Experimental Results Showing the Statistical Significance
and Direction of Main Effects and the Significant Interactions in Experiment 1
Performance Pitch Musical Excerpt Significant Measure Information Experience Number Interactions
Note - n.s. = not significant; BST = beats to start tapping; CV = coefficient of variability; CDel = coefficient of delay; CDev = coefficient of deviation.
*p < .05.
Tapping to Ragtime 469
musical intuitions suggesting that pitch information, especially harmony, is important for the perception of musical rhythm and meter. We will return to this issue when we consider differences between the methods of these experiments and when we consider rhythmic characteristics of the excerpts.
In this experiment, we also found no effect of musical experience on any of the performance measures. Musically experienced subjects did not differ statistically from musically inexperienced subjects on any measures, although some trends favored musically experienced subjects. Whether these results will generalize to other styles of music is questionable given recently re- ported effects for musical experience using classical piano works (Drake et al., 2000). This divergence of findings suggests that pulse-finding differ- ences as a function of musical experience are not due to general motor control differences. Instead, style-dependent pulse-finding differences as a function of musical experience may indicate the presence of more percep- tual difficulties (e.g., temporal periodicity extraction) with less musical ex- perience.
Experiment 2
Experiment 2 further investigated what information subjects use to find and follow the pulse in piano ragtime music. In this music, the left hand usually plays a pattern of alternating low and high notes, often with chords as the second part of the alternation. This steady left-hand part seems a likely cue for finding the pulse. In this experiment, we tested the impor- tance of the left-hand part for pulse finding by measuring the effects of eliminating it (i.e., with stimuli consisting of only the right-hand notes). We also attempted a replication of the findings from Experiment 1 regarding the performance benchmarks and the lack of importance of pitch varia- tions. In addition, these stimuli allowed a test of the hypothesis that pitch variations may be more important in music with relatively weak temporal cues to pulse by looking for effects of pitch within the right-hand versions. As shown in Appendix B, we used shorter excerpts to lessen the perfor- mance disruptions due to section transitions observed in the first experi- ment.
METHOD
Subjects
Twelve musically experienced Cornell University students (male = 5, female = 7; mean years playing music =12) participated for extra credit in undergraduate psychology courses or received $5. One of these subjects reported having congenital high-frequency hearing loss, and one reported having absolute pitch. Two additional subjects' data were not in-
470 Joel Snyder & Carol L. Krumhansl
eluded because of computer malfunction and another because the subject failed to follow instructions. On a 7-point scale, the average familiarity with ragtime music was 3.9. Al- though this average familiarity is higher than for the musically experienced subjects in Ex- periment 1, the difference was not statistically reliable, t(lS) = 1.74, p = .10.
Materials and Stimuli
The same eight modified pieces from Experiment 1 were used. The length of each ex- cerpt was -40 s in this experiment, again terminating at the end of a section to avoid awk- ward endings. Four versions of the ragtime excerpts were used, as illustrated in Figure 7 for the beginning of one of the excerpts: (1) full pitch-varied, (2) full monotonic, (3) right-hand pitch-varied (a pitch-varied version of only the right-hand notes), and (4) right-hand mono- tonic (a monotonic version of only the right-hand notes). The stimuli were presented using the same equipment as in Experiment 1.
Procedure
The procedure was also similar to Experiment 1. The full and right-hand stimuli were presented in separate blocks, with pitch-varied and monotonic versions interleaved within blocks, as before. We counterbalanced whether subjects heard the full or the right-hand block first.
RESULTS
Repeated-measure analysis of variance (ANOVA) tested three within- subjects factors: pitch information, full/right-hand, and excerpt. We used
Fig. 7. The introduction from Glad Cat, showing full pitch-varied, full monotonic, right- hand (RH) pitch-varied, and right-hand monotonic versions. The full pitch-varied and full monotonic versions are identical to the pitch-varied and monotonic versions in Experiment 1, respectively. All of the notes normally played by the left hand were absent in the right- hand versions.
Tapping to Ragtime 471
the same degrees of freedom adjustments for sphericity violations as in Experiment 1. The right-hand-only versions of one excerpt, Pine Apple, differed erroneously from the full versions in their construction. Experi- ment 2 ANOVAs were performed with and without data from Pine Apple. Main effects of pitch information and full/right-hand did not differ accord- ing to whether the data from Pine Apple were included. The main effects of excerpt for aperiodic and CDel were significant with all excerpts included but not when Pine Apple was excluded. In addition, the three-way interac- tion for off-beat tapping was not significant with all excerpts but became significant when we removed Pine Apple. Data from Pine Apple were not used in the results we present, which are summarized in Table 2.
The top of Figure 8 shows the percentage of taps on the downbeat, upbeat, the percentage of off-beat taps, and the percentage of aperiodic taps. Subjects tapped significantly more on the downbeat for the pitch-varied than mono- tonic versions, JF(1, 11) = 25.90, p < .001, and more for the full than the right- hand versions, F(l, 11) = 50.67, p < .001. We also observed a significant difference between excerpts, F(6, 66) = 27.08, 8 = .478, p < .001. For percent- age of taps on the downbeat, we detected an interaction between pitch infor- mation and full/right-hand, F(l, 11) = 8.82, p < .025. We also found interac- tions between pitch information and excerpt, F(6, 66) = 3.37, e = .547, p < .05, and between pitch information, full/right-hand, and excerpt, P(6, 66) = 4.37, 8 = .532,p<.01.
Table 2 Summary of Experimental Results Showing the Statistical Significance
and Direction of Main Effects, and the Significant Interactions in Experiment 2
Performance Pitch Excerpt Significant Measure Information Full/Right-Hand Number Interactions
Downbeat Pitch>Mono* Full>right-hand* * Pitch x full/right-hand
Pitch x excerpt Full/right-hand x excerpt Pitch x full/right-hand x
excerpt Off-beat n.s. Right-hand>full*
* Full/right-hand x excerpt Pitch x full/right-hand x
Note - n.s. = not significant; BST = beats to start tapping; CV = coefficient of variability; CDel = coefficient of delay; CDev = coefficient of deviation.
*p < .05.
472 Joel Snyder & Carol L. Krumhansl
Fig. 8. Percentage of taps on the downbeat, upbeat, percentage of off-beat taps, and per- centage of aperiodic taps for full pitch-varied, full monotonic, right-hand (RH) pitch-var- ied, and right-hand monotonic stimuli in Experiment 2 (top). Mean switches (+SE) in tap- ping mode for full pitch-varied, full monotonic, right-hand pitch-varied, and right-hand monotonic stimuli in Experiment 2 (bottom).
Results for tapping off-beat were similar (but opposite in direction) to those for tapping on the downbeat. Somewhat more off-beat tapping oc- curred for monotonic than for pitch-varied stimuli, although this effect was not significant, F(l, 11) = 3.84, p > .05. Significantly more off-beat tapping occurred for right-hand than full versions, F(l, 11) = 45.30, p < .001. Excerpts also differed from each other on this measure, F(6, 66) = 16.11, e = .481, p < .001. Last, we detected interactions between full/right-
Tapping to Ragtime 473
hand and excerpt, F(6, 66) = 18.33, e = .559, p < .001, and between pitch information, full/right-hand, and excerpt, F(6, 66) = 2.81, e = .544, p < .05.
We found slightly more aperiodic tapping on monotonic than pitch- varied versions, but the difference was not significant, F(l, 11) = 4.02, p > .05. Aperiodic tapping also occurred less on the full than on the right-hand versions, JF(1, 11) = 48.69, p < .001, but excerpts did not differ from each other, JF(6, 66) = 2.97, e = .462, p > .05. A significant interaction appeared between full/right-hand and excerpt, F(6, 66) = 3.66, e = .497, p < .025.
The bottom of Figure 8 shows switches in tapping mode. There was no statistical difference between pitch- varied and monotonic versions, P(l, 11) = 2.56, p > .05. Subjects made fewer switches in the full versions than in the right-hand versions, F(l, 11) = 12.06, p < .01, and there were signifi- cant differences between excerpts, F(6, 66) = 9.59, e = .402, p < .001. We also observed a significant interaction between full/right-hand and excerpt, F(6, 66) = 3.91, e = .367, p < .05.
The top of Figure 9 shows the proportions of subjects tapping at differ- ent numbers of beats after the start of the music, and the bottom of Figure 9 shows this as mean BST. We observed no effects on BST for pitch infor- mation, F(l, 11) = 4.29, p > .05, for full/right-hand, F(l, 11) = .50, p > .05, or for excerpt, F(6, 66) = 2.57, e = .384, p > .05. No significant interactions occurred among pitch information, full/right-hand, and excerpt for BST.
On 96% of the trials, subjects tapped with an ITI of -600 ms, corre- sponding to the quarter-note level. The rest of the trials consisted of sub- jects tapping twice as fast, with an ITI of -300 ms. No subjects tapped at -1200 ms ITI. As displayed in the top of Figure 10, pitch information did not affect the CV, F(l, 11) = .66 , p > .05, though there was a strong effect for higher values on right-hand than full versions, F(l, 11) = 52.63, p < .001. No significant effect of excerpt surfaced for CV, P(6, 66) - 1.93, e = .645, p > .05. Full/right-hand and excerpt interacted significantly, F(6, 66) = 2.92, e = .568, p < .05.
CDel, as displayed in the middle of Figure 10, showed a different pattern than in Experiment 1. Subjects tapped slightly more ahead of the beat with pitch-varied versions than with monotonic versions, JP(1, 11) = 8.28, p < .025. The anticipation was approximately 4 ms on average. No difference occurred in CDel between full and right-hand versions, F(l, 11) = .63, p > .05, or between excerpts, F (6, 66) = 3.09, e = .391, p > .05, but we found a significant interaction between full/right-hand and excerpt, F( 6, 66) = 3.01, e = .464, p < .05. As displayed at the bottom of Figure 10, there was no effect of pitch information for CDev, JF(l,ll) = .01,p>.05, whereas higher values appeared for right-hand than full versions, F(l, 11) = 48.01, p < .001. Excerpts differed from each other on CDev, F(6, 66) = 3.17, e = .669, p < .025, but pitch information, full/right-hand, and excerpt did not inter- act in any combination for this measure.
474 Joel Snyder & Carol L. Krumhansl
Fig. 9. Experiment 2 beats to start tapping (BST). Proportion of trials in which subjects started tapping at different elapsed quarter-note beats from the first note of the trial for full pitch-varied, full monotonic, right-hand (RH) pitch-varied, and right-hand monotonic stimuli (top). Mean BST (+S£) for full pitch-varied, full monotonic, right-hand pitch-varied, and right-hand monotonic stimuli (bottom).
DISCUSSION
The general pulse-finding abilities found in this experiment for the full versions were similar to those of Experiment 1. The main additional find- ing of this experiment was that a number of measures showed poorer per-
Tapping to Ragtime 475
Fig. 10. Mean timing coefficients in Experiment 2. Coefficient of variability (CV, top), coef- ficient of delay (CDel, middle), and coefficient of deviation (CDev, bottom) shown for full pitch-varied, full monotonic, right-hand pitch-varied, and right-hand monotonic stimuli. (Coefficients are +SE for CV and CDev, ±SE for CDel.)
formance for the right-hand-only versions. Without the left-hand accom- paniment, the percentage of tapping on the downbeat significantly decreased, and the percentages of off-beat and aperiodic taps increased. Also, more switches between tapping modes occurred, indicating greater instability of synchronization for right-hand-only versions. Finally, the right-hand-only
476 Joel Snyder 8c Carol L. Krumhansl
versions produced more variable tapping and taps farther from the beat in absolute value. Thus, removing the left-hand notes generally produced dec- rements in tapping performance as assessed on a number of measures. With CDel, subjects tapped very slightly ahead of the beat in the pitch-varied versions (unlike in the earlier experiment) and very slightly behind the beat in the monotonic versions (as in the earlier experiment).
As in Experiment 1, pitch information generally had little effect. Two measures showed pitch effects in Experiment 2, however. More down-beat taps occurred for the pitch-varied than the monotonic versions (especially for the full versions), and taps occurred somewhat earlier relative to the beat for the pitch-varied versions than for the monotonic versions. These discrepancies are difficult to explain but could potentially arise from ex- posing subjects to two pitch-varied versions of each excerpt in Experiment 2, but just one pitch-varied version of each excerpt in Experiment 1. With the exceptions of percentage of tapping on the downbeat and tap delay, this experiment replicated the previous finding that removing pitch infor- mation had little effect on tapping performance. Moreover, the lack of in- teractions between full/right-hand and pitch information, except on per- centage of down-beat taps, suggested that pitch cues were not used more when the left hand was absent. This one interaction demonstrated that while removing pitch information resulted in less tapping on the down- beat, the effect was larger for the full versions. This was against the predic- tion that removing the left-hand part would result in greater use of pitch information.
Excerpt Main Effects and Interactions
Since most performance measures in the two experiments showed sig- nificant excerpt effects, we determined whether the same excerpts exhib- ited poor performance across measures. For Experiment 1, American Beauty had the least down-beat tapping, the most aperiodic tapping, the highest BST, the highest CDev, the second most switches, and the second highest CV. Sensation had the most switches, the highest CV, and the second most aperiodic tapping. For Experiment 2, American Beauty showed the least down-beat tapping, the most aperiodic tapping, the most switches, the high- est BST, the highest CV, the second most off-beat tapping, and the second highest CDev. Sensation showed the most off-beat tapping, the second least down-beat tapping, the second most aperiodic tapping, the most switches, and the second highest CV. These were the only two excerpts that showed consistently poor performance. American Beauty may show poor perfor- mance because the right hand contained frequent long notes and changing rhythmic patterns, and because it was the only excerpt to begin with a
Tapping to Ragtime 477
pickup in both left- and right-hand parts. The reason for the poor perfor- mance on Sensation is less clear, although it is possible that the relatively large number of different sections in this excerpt, or the fact that the right hand began on the second sixteenth-note metrical position, caused diffi- culty.
In both experiments, we detected interactions between pitch informa- tion and excerpt for down-beat tapping. For Experiment 1, this was mostly due to less tapping on the downbeat for American Beauty when pitch in- formation was absent. In Experiment 2, American Beauty and Blue Goose showed less down-beat tapping for the monotonic than the pitch-varied versions. American Beauty was the only excerpt beginning with a left-hand pickup on the next to final sixteenth-note position (upbeat), and with a right-hand pickup on the final sixteenth-note position of the measure, thus eliminating the opening note as a cue to the downbeat in all stimulus ver- sions. Accounting for the three-way interaction of pitch information, full/ right-hand, and excerpt in Experiment 2, American Beauty had a relatively high percentage of down-beat tapping for the full pitch- varied version only. This shows that subjects only overcame the pickups in American Beauty when pitch information was present in the left hand. In contrast, Blue Goose showed a decrease in down-beat tapping when pitch information was ab- sent regardless of whether the left hand was present. This suggests that right-hand pitch cues carry information about the downbeat in this ex- cerpt. The most obvious right-hand pitch cue in Blue Goose is the presence of a melodic pattern that opened many of the measures with three consecu- tive intervals of -1, +1, and -5 semitones (i.e., descending minor second, ascending minor second, and descending perfect fourth).
In Experiment 2, the interaction of full/right-hand and excerpt for down- beat tapping resulted from American Beauty and Sensation showing larger deleterious effects of removing the left hand than the other excerpts. For American Beauty, this is attributable to the right-hand characteristics de- scribed earlier. For Sensation, this is most likely because the right hand began with a note on the second sixteenth-note, a very weak metrical posi- tion. However, the presence of the left hand in Sensation, which began on the downbeat, seems to have enabled subjects to find the downbeat in the full versions. The interaction between full/right-hand and excerpt with ape- riodic tapping in Experiment 2 occurred largely because each excerpt showed a different increase in aperiodic tapping when the left hand was absent. The largest increase was in American Beauty, consistent with the right- hand characteristics described above. With switches, we observed a signifi- cant interaction between full/right-hand and excerpt in Experiment 2, mostly due to American Beauty exhibiting a more deleterious effect of removing the left hand than the other excerpts. For CV, the interaction between full/ right-hand and excerpt in Experiment 2 was mostly due to different sized
478 Joel Snyder & Carol L. Krumhansl
increases in CV when the left hand was absent, depending on the excerpt. Finally, the significant interaction between full/right-hand and excerpt for CDel in Experiment 2 resulted from a slight increase when the left hand was absent for Lily Queen, while the other excerpts showed a slight de- crease. This is difficult to explain because Lily Queen does not stand out from the other excerpts, except in its relatively fast tempo.
Music Analysis
In order to isolate musical dimensions that may be cues to pulse in the ragtime pieces, we analyzed the full pitch-varied versions from Experiment 2 on a number of features. The first two features were general aspects of rhythmic structure relevant to pulse finding. These were right-hand synco- pation (defined as a half-measure pattern in which emphasized notes occur on weak sixteenth-note metrical positions and one or more strong sixteenth- note metrical positions contain rests or tied notes), and presence of the standard ragtime left-hand pattern (defined as a pattern of alternating low and high eighth notes with no faster notes in between). Across the excerpts, 40% of half-measures in the right hand contained syncopation, and 67% of the left-hand half-measures contained the standard ragtime left-hand pattern. This coarse analysis fits well with the general notion that ragtime usually has a regular left-hand part and frequently contains syncopation in the right hand.
The next analysis considered the distributions of musical events at each sixteenth-note metrical position. These may provide cues to the phase of the metrical structure, that is, the location of relatively strong beats within the metrical period. We calculated average event distributions at the six- teenth-note level for the following musical dimensions: number of right- hand note onsets, right-hand chords (0 = absent, 1 = present), melody note duration in sixteenth notes, melody note pitch value (60 = middle C, 1 scale unit = 1 semitone), melodic interval size in semitones, melodic con- tour change (0 = absent, 1 = present), number of left-hand note onsets, and left-hand chords (0 = absent, 1 = present). For melodic interval size, the interval is attributed to the metrical position of the second tone constitut- ing the interval (e.g., with notes C5 D5, the interval occurs at D5). A melodic contour change is attributed to the metrical position where the middle of three tones defining the change occurs (e.g., with notes C5 D5 C5, the con- tour change occurs at D5).
The first three rows of graphs in Figure 1 1 show the average event distri- butions across excerpts for the right-hand musical dimensions. Since sub- jects tapped primarily on the first and fifth sixteenth-note metrical posi- tions (i.e., the downbeat), we looked for peaks at these positions. In the
Tapping to Ragtime 479
Fig. 11. Mean distribution of musical events by sixteenth-note metrical position across the Experiment 2 full pitch-varied stimuli. Metrical positions 1 and 5 correspond to the down- beat, and positions 3 and 7 correspond to the upbeat. From left to right, starting with the top row: number of right-hand (RH) note onsets, right-hand chords, melody note duration in sixteenth-notes, melody note pitch value, melodic interval size in semitones, melodic contour change, number of left-hand (LH) note onsets, and left-hand chords.
right hand, modest peaks at the first metrical position appeared for right- hand note onsets, right-hand chords, and melody note duration. In addi- tion, right-hand note onsets, right-hand chords, and melodic interval size showed peaks at the fifth metrical position. The large intervals on the fifth metrical position could form part of a larger melodic pattern that has a consistent phase structure in ragtime. Among the right-hand musical di- mensions, only the note onsets and chords contained peaks at both the first and fifth metrical positions, and no pitch features showed a similarly clear pattern. For the left hand, the bottom row of Figure 1 1 displays results of the event distributions for note onsets and chords. In contrast to the right- hand musical analyses, the left-hand note onsets and chords showed a very clear pattern of peaks. Strong peaks appeared at the odd metrical positions
480 Joel Snyder & Carol L. Krumhansl
for both musical dimensions. There were very few left-hand onsets and no chords on even metrical positions.
Next we employed auto-correlation analysis as a way of discovering repeating patterns that may serve as cues to the periodic metrical struc- ture (Brown, 1993; Vos et al., 1994) for each of the excerpts (for a theoretical account of autocorrelations, see Cryer, 1986). Autocorrelations detect periodicities in a series of numbers by correlat- ing the series with itself using relative lags (or shifts) of the time series. The results are displayed in autocorrelograms (here averaged across excerpts), which plot correlation coefficients as a function of lag. A peak in an autocorrelogram at a lag of 4 indicates a periodicity in the music at that duration (i.e., a period of 4 sixteenth notes). In the present analysis, correlations at lags of 2, 4, and 8 were of interest because these corresponded to observed metrical distances between taps. We analyzed the same musical dimensions as in the metrical position analysis for each excerpt.
The first three rows of graphs in Figure 12 show the average right- hand autocorrelation coefficients across excerpts for lags of 1-8 six- teenth notes. After Bonferroni corrections for multiple t tests, no peaks significantly different from 0 appeared at two, four, or eight sixteenth notes for note onsets, chords, note duration, pitch value, or contour changes. The only significant peak in the right-hand dimensions was at four sixteenth-notes for melodic interval size, t(7) = 6.12, p < .001. In contrast, the bottom row of graphs in Figure 12 shows peaks signifi- cantly different from 0 at periods two, four, and eight for left-hand note onsets and chords (ps< .05) except in the case of left-hand chords at two sixteenth notes, t(7) = 3.48, p > .05. It is noteworthy that four sixteenth notes is the lag with the highest correlations in the two left- hand autocorrelograms, corresponding to the predominant tapping pe- riod of a half-measure.
To summarize, the musical analyses across excerpts demonstrated a large amount of syncopation in the right hand and a high prevalence of the left-hand pattern of eighth notes. This is consistent with conven- tional descriptions of ragtime music (Berlin, 1980, 1985; Jasen & Tichenor, 1978). It is also consistent with the behavioral finding that removing the left-hand notes caused significant disruptions in tapping performance. Event distributions and autocorrelational results for the right-hand note onsets, right-hand chords, melody note duration, me- lodic interval size, and melodic contour change indicated that these are sometimes modest cues for where to tap in the measure (phase) and how often to tap (period). In contrast, the event distributions and autocorrelations for left-hand note onsets and chords indicated these to be very strong cues to phase and period.
Tapping to Ragtime 481
Fig. 12. Mean autocorrelation coefficients for lags of 1-8 sixteenth-notes across the Experi- ment 2 full pitch-varied stimuli. Periods of 4 and 8 correspond to durations of a quarter- note and a half-note, respectively. From left to right, starting with the top row: number of
right-hand (RH) note onsets, right-hand chords, melody note duration in sixteenth notes, melody note pitch value, melodic interval size in semitones, melodic contour change, num- ber of left-hand (LH) note onsets, and left-hand chords.
General Discussion
We took two approaches to understanding the musical cues used for pulse finding. The first approach was to experimentally manipulate the musical materials. Both experiments contained pitch-varied and monotonic versions of the same excerpts. The results showed remarkably little differ- ence in performance between the two versions. The second experiment pre- sented versions with both left-hand and right-hand parts of the music or only the right-hand part. Removing the left-hand part produced noticeable decrements in performance. The second approach analyzed the music for the presence of cues to pulse in the right-hand and left-hand parts, espe- cially those that corresponded to the predominant period and phase of
482 Joel Snyder d>c Carol L. Krumhansl
subjects' tapping. Together, these approaches uncovered some possible fea- tures important to pulse finding, especially in the left-hand part.
The relatively small effect of removing pitch variations for most excerpts directed the focus to temporal cues that would be available in both pitch- varied and monotonic versions. Some candidate cues in the right-hand part were the relatively frequent note and chord onsets on the downbeat (com- pared with the upbeat), the relatively long duration of these notes, and the relative size of melodic intervals at these positions. The left-hand part was characterized by a succession of events at the eighth-note level, producing periodicities corresponding to the eighth-note, quarter-note, and half-note levels. In combination, these cues appeared sufficient for rapidly finding and maintaining the pulse. However, when the left-hand part was absent, performance deteriorated. This is understandable because the right-hand part was frequently syncopated, and because the right-hand cues found in the musical analyses were extremely weak or nonexistent, depending on the specific right-hand musical dimension, compared with the left-hand cues.
The finding that musically inexperienced subjects performed similarly to musically experienced subjects contrasts with results of one recent study that used classical piano music (Drake et al., 2000). Whether this discrep- ancy is due to differences in stimulus materials or the performance mea- sures used is currently unclear and therefore deserves systematic investiga- tion. Such an investigation could serve to establish what aspects (e.g., metrically consistent tapping) of pulse finding vary with musical experi- ence and how they might depend on musical style.
The two experiments showed that subjects performed well in absolute terms when tapping to music. Based on the performance of musically expe- rienced subjects on the full pitch- varied versions, we propose the following benchmarks for pulse finding in this style of music: (1) 90% of taps on the downbeat, (2) no off-beat tapping, (3) less than 1% aperiodic taps, (4) an average of 0.1 switch between tapping modes per excerpt, (5) 90% tapping at -600 ms ITI, (6) 4 beats to start tapping (BST, about 2400 ms), (7) 4% CV (about 24 ms), (8) ±1% CDel (about 6 ms behind or ahead of the beat), and (9) 4% CDev (about 24 ms).
These benchmark values may differ with other styles of music, but the findings are generally in line with previous results. That subjects prefer to tap with a 600 ms ITI is consistent with studies of spontaneous tapping and other rhythmic behaviors (Clarke, 1999; Fraisse, 1982; van Noorden & Moelants, 1999). Subjects in the current study showed BSTs similar to values reported by Fraisse (1982) for human synchronization with isochro- nous patterns and similar to some rule-based pulse-finding models (Desain &C Honing, 1999). The range of typical values across these reports is 2-4 beats. Once the tapping began, it was remarkably stable. Switches in tap-
Tapping to Ragtime 483
ping mode occurred infrequently and often appeared at transitions between sections of the music. Other behavioral data on switches in tapping mode are not available for comparison, to our knowledge. However, the relative instability of some pulse-finding models suggests there may be a role for top-down processes in maintaining a tapping mode once it is established (Scheirer, 1998).
The typical variability of ITI observed in this study (CV = 4%, or about 24 ms) is similar in magnitude to other studies of synchronization (Scheirer, 1998). Subjects' tapping to isochronous auditory patterns typically antici- pates the beat by around 40 ms (Mates et al., 1992; Vos et al., 1995), although when the pacing intervals are filled with other tones, anticipation is greatly diminished (Wohlschlager & Koch, 2000). This latter fact is con- sistent with the current study, which found no clear anticipation in music with multiple notes per tapped pulse. The average tap delay was small, about 6 ms (behind the beat) in Experiment 1 and about -4 ms (ahead of the beat) in Experiment 2.
Although the precise relationship between the perception of musical pulse and other rhythmic behaviors is unclear, pulse perception may share simi- lar or identical mechanisms with limb coordination and time perception (Ivry & Hazeltine, 1995; Large & Jones, 1999; McAuley, 1995; McAuley 8c Kidd, 1998). It should be noted that others have argued, based on em- pirical studies of different rhythmic behaviors, that a common timing mecha- nism does not underlie motor control across tasks (e.g., Robertson et al., 1999). Furthermore, researchers disagree on the precise mechanism sup- porting temporal cognition in the 300-1500 ms range, especially whether the phase is represented in addition to the period of time intervals (Gibbon, Malapani, Dale, & Gallistel, 1997; Ivry, 1996; Large & Jones, 1999; McAuley, 1995).
We now provide a brief description of pulse-finding models. It is impor- tant to note that the current study does not provide evidence for one type of pulse-finding mechanism over another. However, data such as ours might be used to test the various types of models. One early class of models for meter and pulse applied rule-based algorithms that use information such as melodic patterning and the location of long notes to create abstract hypoth- eses about the period and phase of the pulse (e.g., Desain & Honing, 1999; Longuet-Higgins &c Lee, 1982; Steedman, 1977). These models can be ex- tremely useful for testing the contributions of different types of information to pulse and meter. The main limitation with rule-based models is that they do not address how we accommodate for the variability found in human musical performance (Palmer, 1997). This limitation also applies to the approach of Parncutt (1994). His psychoacoustic model quantifies the relationship between stimulus characteristics and perceived accent and pulse salience, relationships that can be used by other types of pulse-finding models (e.g., Toiviainen, 1998).
484 Joel Snyder & Carol L. Krumhansl
An influential theory of rhythmic cognition is that of Jones and her col- leagues, known as Dynamic Attending Theory (Jones, 1976; Jones & Boltz, 1989; Large & Jones, 1999). This theory postulates that people synchro- nize with an external pattern by generating internal rhythmic expectancies for when events will occur. The period and phase of these expectancies, or attentional rhythms, can adapt to correct for temporal variability in the attended pattern. Influenced by this notion, a number of computational models use adaptive oscillators as basic computational units (Eck, Gasser, & Port, 2000; Gasser et al., 1999; Large & Jones, 1999; Large & Kolen, 1994; McAuley, 1995; McAuley & Kidd, 1998; Toiviainen, 1998). Note that these adaptive models are different from the theory of pulse sensation proposed by van Noorden and Moelants (1999), who used a damped lin- ear oscillator (e.g., a mass and spring system). For simulating musical pulse, an adaptive oscillator is particularly attractive because it can adjust its pe- riod and phase in response to temporal deviations from isochrony. This property, in principle, would enable listeners, performers, and dancers to synchronize with performed music in real time. However, obtaining direct behavioral evidence for oscillators has been difficult (McAuley, 1995; McAuley & Kidd, 1998; Pashler, 2001; but see Large & Jones, 1999). In addition, some investigators argue that timing may instead be an emergent property of coded trajectories within a distributed neural system or that it depends on spatially coded interval representations (Buonomano, Hickmott, & Merzenich, 1997; Conditt & Mussa-Ivaldi, 1999; He, Hashikawa, Ojima, & Kinouchi, 1997; Ivry & Hazeltine, 1995; Keating & Thach, 1997; Penhune, Zatorre, & Evans, 1998; Scheirer, 1998; Todd, 1994; Todd et al., 1999; Wright, Buonomano, Mahncke, & Merzenich, 1997).
Another approach to pulse finding relies on linear filtering of the acous- tic signal in order to extract periodicities (Scheirer, 1998; Todd, 1994; Todd et al., 1999). Such an approach has the advantage of being motivated by physiology, in particular the filtering characteristics of sensory systems. Applications of this approach have shown how the complex temporal struc- ture of music (i.e., grouping and meter) can be recovered by passive filter- ing mechanisms (Todd, 1994; Todd et al., 1999), how human musical per- formances can be tracked without assuming nonlinear oscillators (Scheirer, 1998; Todd et al., 1999), and how uninterrupted summation of tone pro- cessing can explain durational accent (Todd, 1994; Todd et al., 1999). Since filtering models and oscillator models can both track temporal variability in music and represent pulse and meter, determining which approach best describes human pulse-finding mechanisms will likely require detailed com- putational analysis and experimentation.
Most models of pulse assume that pitch information is not vital, an ap- proach largely consistent with the present behavioral results. Several ef- forts (Desain & Honing, 1999; Eck et al., 2000; Gasser et al., 1999; Large & Jones, 1999; Large & Kolen, 1994; McAuley, 1995; McAuley & Kidd,
Tapping to Ragtime 485
1998; Scheirer, 1998; Todd, 1994; Todd et al., 1999; Toiviainen, 1998) have shown that computational models that rely on temporal information can simulate human time perception, determine the pulse and meter of music, and follow temporally modulating patterns. Despite the successes of many of these time-based models and the current study's findings, it remains a possibility that pitch information is used when it is a reliable cue to pulse, as suggested by the behavioral results found for some excerpts in this study.
Indeed, the studies of Dawe et al. (1993, 1994, 1995) showed the impor- tance of harmonic rhythm (i.e., the temporal position of a chord change) in relatively simple stimuli. Their studies showed that this cue predominantly determined the rhythmic organization when it was compared with tempo- ral duration of tones and melodic grouping. In their studies, however, the various cues were put into conflict. With music, in contrast, we would expect that the cues would generally covary, although further music analy- sis directed toward this point is needed. Another important issue that mu- sic analysis could address is whether harmonic accents are generally as regular as temporal accents. If not, we might expect them to receive rela- tively little attention in pulse finding. On the other hand, if harmonic ac- cents are as useful as temporal cues, one or both of these cues may be used.
In summary, this study suggested some general benchmarks for pulse finding in humans with piano ragtime music. These abilities are impressive and do not show large improvements with musical training. We hope that data such as these play an increased role by informing computational mod- eling of sensory-motor synchronization (e.g., the current data have been modeled by Large, 2000; Toiviainen &c Snyder, 2000) and encouraging music analysis that more fully characterizes the musical structures on which this ability is based. In addition, studies of various musical styles, more specific and theoretically motivated manipulations of the music, and the use of performed musical stimuli are important aims. The latter issue deserves particular attention (as in Drake et al., 2000) because one of the funda- mental accomplishments of synchronizing systems is their ability to ac- commodate temporal noise, tempo change, and expressive timing.2
2. The preparation of this article occurred while Joel Snyder was a predoctoral trainee on National Institute of Mental Health (NIMH) training grant T32 MH 19389, "Multidisciplinary Training in Developmental Psychology." We thank Peter Desain, Ed Large, Steve McAdams, and Neil Todd for their valuable comments on previous drafts of this paper. We also gratefully acknowledge the assistance of Angelica Goss-Bley in running sub- jects and analyzing data, and Anne Briard in analyzing the music.
References
Berlin, E. A. (1980). Ragtime: A musical and cultural history. London: University of Cali- fornia Press.
Berlin, E. A. (1985). Ragtime songs. In J. E. Hasse (Ed.), Ragtime: Its history, composers, and music. New York: Schirmer Books
486 Joel Snyder &c Carol L. Krumhansl
Bertenthal, B. I., Rose, J. L., & Bai, D. L. (1997). Perception-action coupling in the develop- ment of visual control of posture. Journal of Experimental Psychology: Human Percep- tion and Performance, 23, 1631-1643.
Brown, J. C. (1993). Determination of the meter of musical scores by autocorrelation. Jour- nal of the Acoustical Society of America, 94, 1953-1957.
Buonomano, D. V., Hickmott, P. W., & Merzenich, M. M. (1997). Context-sensitive synap- tic plasticity and temporal-to-spatial transformations in hippocampal slices. Proceed- ings of the National Academy of Sciences USA, 94, 10403-10408.
Clarke, E. F. (1999). Rhythm and timing in music. In D. Deutsch (Ed.), Psychology of music (2nd ed., pp. 473-500). New York: Academic Press.
Conditt, M. A., & Mussa-Ivaldi, F. A. (1999). Central representation of time during motor learning. Proceedings of the National Academy of Sciences USA, 96, 11625-11630.
Cryer, J. D. (1986). Time series analysis, Boston: Duxbury Press. Cummins, E, & Port, R. (1998). Rhythmic constraints on stress timing in English. Journal
of Phonetics, 26, 145-171. Dawe, L. A., Platt, J. R., & Racine, R. J. (1993). Harmonic accents in inference of metrical
structure and perception of rhythm patterns. Perception and Psychophysics, 54, 794- 807.
Dawe, L. A., Platt, J. R., & Racine, R. J. (1994). Inference of metrical structure from per- ception of iterative pulses within time spans defined by chord changes. Music Percep- tion, 12, 57-76.
Dawe, L. A., Platt, J. R., & Racine, R. J. (1995). Rhythm perception and differences in accent weights for musicians and nonmusicians. Perception and Psychophysics, 57, 905- 914.
Desain, P., & Honing, H. (1999). Computational models of beat induction: The rule-based approach. Journal of New Music Research, 28, 29-42.
Deutsch, D. (1983). The generation of two isochronous sequences in parallel. Perception and Psychophysics, 34, 331-337.
Diedrich, F. J., & Warren, W. H., Jr. (1995). Why change gaits? Dynamics of the walk-run transition. Journal of Experimental Psychology: Human Perception and Performance, 21, 183-202.
Drake, C, Penel, A., & Bigand, E. (2000). Tapping in time with mechanically and expres- sively performed music. Music Perception, 18, 1-23.
Eck, D., Gasser, M., & Port, R. (2000). Dynamics and embodiment in beat induction. In P. Desain & L. Windsor, Rhythm perception and performance (pp. 157-170). Lisse, The Netherlands: Swets and Zeitlinger.
Essens, P. J., & Povel, D. J. (1985). Metrical and nonmetrical representations of temporal patterns. Perception and Psychophysics, 37, 1-7.
Fraisse, P. (1982). Rhythm and tempo. In D. Deutsch (Ed.), Psychology of music (pp. 149- 180). New York: Academic Press.
Gasser, M., Eck, D., & Port, R. (1999). Meter as mechanism: A neural network that learns metrical patterns. Connection Science, 11, 187-216.
Gibbon, J., Malapani, C, Dale, C. L., & Gallistel, C. R. (1997). Toward a neurobiology of temporal cognition: Advances and challenges. Current Opinion in Neurobiology, 7, 170- 184.
He, J., Hashikawa, T, Ojima, H., & Kinouchi, Y. (1997). Temporal integration and duration tuning in the dorsal zone of cat auditory cortex. Journal of Neuroscience, 1 7, 2615-2625.
Helmuth, L. L., & Ivry, R. B. (1996). When two hands are better than one: Reducing timing variability during bimanual movements. Journal of Experimental Psychology: Human Perception and Performance. 22. 278-293.
Ivry, R. B. (1996). The representation of temporal information in perception and motor control. Current Opinion in Neurobiology, 6, 851-857.
Ivry, R. B., &c Hazeltine, R. E. (1995). Perception and production of temporal intervals across a range of durations: Evidence for a common timing mechanism. Journal of Ex- perimental Psychology: Human Perception and Performance, 21, 3-18.
Tapping to Ragtime 487
Jasen, D. A., & Tichenor, T. J. (1978). Rags and ragtime: A musical history. New York: Seabury Press.
Jones, M. R. (1976). Time, our lost dimension: Toward a new theory of perception, atten- tion, and memory. Psychological Review, 83, 323-355.
Jones, M. R., & Boltz, M. (1989). Dynamic attending and responses to time. Psychological Review, 96, 459-491.
Jones, M. R., & Pfordresher, P. Q. (1997). Tracking musical patterns using joint accent structure. Canadian Journal of Experimental Psychology, 51, 271-290.
Keating, J. G., & Thach, W. T. (1997). No clock signal in the discharge of neurons in the deep cerebellar nuclei. Journal of Neurophysiology, 77, 2232-2234.
Klapp, S. T, Hill, M. D., Tyler, J. G., Martin, Z. E., Jagacinski, R. J., & Jones, M. R. (1985). On marching to two different drummers: Perceptual aspects of the difficulties. Journal of Experimental Psychology: Human Perception and Performance, 11, 814-827.
Large, E. W. (1994). Dynamic representation of musical structure. Unpublished doctoral dissertation, The Ohio State University, Columbus.
Large, E. W. (2000). On synchronizing movements to music. Human Movement Science, 19,527-566.
Large, E. W., & Jones, M. R. (1999). The dynamics of attending: How people track time- varyine events. Psychological Review, 106, 119-159.
Large, E. W., & Kolen, J. F. (1994). Resonance and the perception of musical meter. Con- nection Science, 6, 177-208.
Longuet-Higgins, H. C, & Lee, C. S. (1982). The perception of musical rhythms. Percep- tion, 11, 115-128.
Mates, J., Radii, T., & Poppel, E. (1992). Cooperative tapping: Time control under differ- ent feedback conditions. Perception and Psychophysics, 52, 691-704.
McAuley, J. D. (1995). Perception of time as phase: Toward an adaptive-oscillator model of rhythmic pattern processing. Unpublished doctoral dissertation, Indiana University, Bloomineton.
McAuley, J. D., & Kidd, G. R. (1998). Effect of deviations from temporal expectations on tempo discrimination of isochronous tone sequences. Journal of Experimental Psychol- ogy: Human Perception and Performance, 24, 1786-1800.
Palmer, C. (1997). Music performance. Annual Review of Psychology, 48, 115-138. Parncutt, R. (1987). The perception of pulse in musical rhythm. In A. Gabrielsson (Ed.),
Action and perception in rhythm and music. Stockholm: Royal Swedish Academy of Music.
Parncutt, R. (1994). A perceptual model of pulse salience and metrical accent in musical rhythms. Music Perception, 11, 409-464.
Pashler, H. (2001). Perception and production of brief durations: Beat-based versus inter- val-based timing. Journal of Experimental Psychology: Human Perception and Perfor- mance, 27, 485-493.
Penhune, V. B., Zatorre, R. J., & Evans, A. C. (1998). Cerebellar contributions to motor timing: A PET study of auditory and visual rhythm reproduction. Journal of Cognitive Neuroscience, 10, 752-765.
Peper, C. L. E., Beek, P. J., & van Wieringen, P. C. W. (1995). Multifrequency coordination in bimanual tapping: Asymmetrical coupling and signs of supercriticality. Journal of Experimental Psychology: Human Perception and Performance, 21, 1117-1138.
Povel, D. J. (1981). Internal representation of simple temporal patterns. Journal of Experi- mental Psychology: Human Perception and Performance, 7, 3-18.
Povel, D. J. (1984). A theoretical framework for rhythm perception. Psychological Re- search, 45, 315-337.
Povel, D. J. (1985). Time, rhythms, and tension: In search of the determinants of rhyth- micity. In J. A. Michon, & J. L. Jackson (Eds.), Time, Mind, and Behavior (pp. 215- 225). Berlin: Springer.
Povel, D. J., & Collard, R. (1982). Structural factors in patterned finger tapping. Acta Psychologica, 52, 107-123.
488 Joel Snyder & Carol L. Krumhansl
Povel, D. J., & Essens, P. (1985). Perception of temporal patterns. Music Perception, 2, 411-440.
Robertson, S. D., Zelaznik, H. N., Lantero, D. A., Bojczyk, K. G., Spencer, R. M., Doffin, J. G., & Schneidt, T. (1999). Correlations for timing consistency among tapping and draw- ing tasks: Evidence against a single timing process for motor control. Journal of Experi- mental Psychology: Human Perception and Performance, 25, 1316-1330.
Scheirer, E. D. (1998). Tempo and beat analysis of acoustic musical signals. Journal of the Acoustical Society of America, 103, 588-601.
Schmidt, R. C, Beek, P. J., Treffner, P. J., & Turvey, M. T. (1991). Dynamical substructure of coordinated rhythmic movements. Journal of Experimental Psychology: Human Per- ception and Performance, 17, 635-651.
Schmidt, R. C, Bienvenu, M., Fitzpatrick, P. A., & Amazeen, P. G. (1998). A comparison of intra- and interpersonal interlimb coordination: Coordination breakdowns and coupling strength. Journal of Experimental Psychology: Human Perception and Performance, 24, 884-900.
Schmuckler, M. A. (1997). Children's postural sway in response to low- and high-frequency visual information for oscillation. Journal of Experimental Psychology: Human Percep- tion and Performance, 23, 528-545.
Steedman, M. J. (1977). The perception of musical rhythm and metre. Perception, 6, 555- 569.
Todd, N. P. M. (1994). The auditory "primal sketch": A multiscale model of rhythmic grouping. Journal of New Music Research, 23, 25-70.
Todd, N. P. M. (1999). Motion in music: A neurobiological perspective. Music Perception, 17, 115-126.
Todd, N. P. M., O'Boyle, D. J., & Lee, C. S. (1999). A sensory-motor theory of rhythm, time perception, and beat induction. Journal of New Music Research, 28, 5-28.
Toiviamen, P. (1998). An interactive MIDI accompanist. Computer Music Journal, 22, 63- 75.
Toiviainen, P., & Snyder, J. (2000). The time-course of pulse sensation: Dynamics of beat induction. In C. Woods,G. B. Luck, R. Brochard, S. A. O'Neill, & J. A. Sloboda (Eds.), Proceedings of the Sixth International Conference on Music Perception and Cognition [CD-ROM1. Keele, Staffordshire, UK: Department of Psychology.
van Noorden, L., & Moelants, D. (1999). Resonance in the perception of musical pulse. Journal of New Music Research, 28, 43-66.
Vos, P. G., Mates, J., & van Kruysbergen, N. W. (1995). The perceptual centre of a stimulus as the cue for synchronization to a metronome: Evidence from asynchronies. The Quar- terly Journal of Experimental Psychology, 48 A, 1024-1040.
Vos, P. G., van Dijk, A., & Schomaker, L. (1994). Melodic cues for metre. Perception, 23, 965-976.
Wohlschlager, A., & Koch, R. (2000). Synchronization error: An error in time perception. In P. Desain & L. Windsor, Rhythm perception and performance (pp. 115-127). Lisse, The Netherlands: Swets and Zeitlineer.
Wright, B. A., Buonomano, D. V., Mahncke, H. W, & Merzenich, M. M. (1997). Learning and generalization of auditory temporal-interval discrimination in humans. Journal of Neuroscience, 17, 3956-3963.
Note - Tempo is in quarter-note beats per minute (bpm) and interbeat interval (IBI) in milliseconds. Under Form, Intro stands for a four-measure introduction, Pickup stands for an introduction shorter than one measure, and letters stand for unique sixteen-measure sections.
Appendix B
Experiment 2 Excerpt Information
Title Duration (s) Tempo (bpm) Tempo (IBI) Form
Pine Apple* 41 92 651 AA Glad Cat 30 86 697 IntroA Lily Queen 41 102 587 IntroAA Sensation 40 96 624 AB Blue Goose 29 94 639 IntroA American Beauty 41 97 617 PickupAA Nonpareil 45 95 631 IntroAA Chrysanthemum 46 92 651 IntroAA
Note - Tempo is in quarter-note beats per minute (bpm) and interbeat interval (IBI) in milliseconds. Under Form, Intro stands for a four-measure introduction, Pickup stands for an introduction shorter than one measure, and letters stand for unique sixteen-measure sections.