Targeting mitochondrialdysfunction in neurodegenerativedisease: Part IIVictoria S Burchell, Sonia Gandhi, Emma Deas, Nicholas W Wood,Andrey Y Abramov & Helene Plun-Favreau††UCL Institute of Neurology, Department of Molecular Neuroscience, Queen Square,
London WC1N 3BG, UK
Importance of the field: With improvements in life expectancy over the past
decades, the incidence of neurodegenerative disease has dramatically
increased and new therapeutic strategies are urgently needed. One possible
approach is to target mitochondrial dysfunction, which has been implicated
in the pathogenesis of numerous neurodegenerative disorders.
Areas covered in this review: This review examines the role of mitochondrial
dysfunction in neurodegeneration, drawing examples from common diseases
such as Alzheimer’s disease and rarer familial disorders such as Charcot-Marie-
Tooth. The review is provided in two parts. In part I we discussed the mito-
chondrial defects which have been most extensively researched (oxidative
stress, bioenergetic dysfunction, calcium mishandling). We focus now on
those defects which have more recently been implicated in neurodegenera-
tion; in mitochondrial fusion/fission, protein import, protein quality control,
kinase signalling and opening of the permeability transition pore.
What the reader will gain: An examination of mitochondrial defects observed
in neurodegeneration, and existing and possible future therapies to target
these defects.
Take home message: The mitochondrially-targeted therapeutics that have
reached clinical trials so far have produced encouraging but largely inconclu-
sive results. Increasing understanding of mitochondrial dysfunction has,
however, led to preclinical work focusing on novel approaches, which has
Mitochondria mediate numerous processes in neurons from biogenesis to cell death,and defects in mitochondrial function are a feature of almost all neurodegenerativediseases. Current treatments for these diseases primarily target the symptoms ratherthan the cause(s), and have varying therapeutic effects. As research into the role ofmitochondrial dysfunction in disease has progressed, interest in targeting thesedefects therapeutically has gained momentum. In part I of this review we discusseda number of new compounds currently in trials which directly target aspects ofmitochondrial dysfunction. These compounds perform well in vitro and in animalmodels but have frequently disappointed in clinical trials, although progress hasbeen made in the past decade in improving bioavailability and mitochondrial target-ing. Optimisation of these compounds is ongoing, and new compounds are beingdeveloped all the time. As research into neurodegeneration continues, newmitochondrial defects are implicated in disease and new mechanisms elucidated
which may be the target of future therapeutic compounds. Inthis part of the review we focus on these defects and potentialstrategies to target them.
2. Mitochondrial fission/fusion andtrafficking in neurodegeneration
Mitochondria are highly dynamic and mobile, constantlychanging their morphology from long thread-like structuresto small round bodies that can be trafficked throughout thecell. This enables the cell to rearrange its mitochondriaaccording to the local metabolic demands of each cellularregion, vital for proper neuronal function.This morphology is determined by the balance of fusion
and fission: a shift towards fission produces punctatemitochondria, while a shift towards fusion produces networksof connected mitochondria. The mechanisms of mitochon-drial fission and fusion have been extensively reviewed else-where [1,2] and will not be covered in detail here. In brief,mitochondrial dynamics are controlled by a number of largedynamin-related GTPases. Fission is primarily mediated bythe enzyme dynamin-related protein 1 (Drp1), which isrecruited to the mitochondria and interacts with Fis1, ananchor protein in the outer membrane. Fusion is a concertedprocess requiring the action of both inner and outer
membrane enzymes. Outer membrane fusion is mediatedmainly by two large dynamin-related GTPases, mitofusins1 and 2 (Mfn1 and Mfn2), which oligomerise to tether neigh-bouring mitochondria together. Inner membrane fusion ismediated by the GTPase Opa1.
In a healthy neuron, tight regulation of fusion and fissionenables the formation of continuously rearranging mito-chondrial networks. This fusion/fission process is importantfor several reasons. It helps maintain mitochondrial function,since fusion allows damaged proteins to be compensatedfor by functional complementation (for review see [3]) andfission enables damaged mitochondria to be targeted to theautophagosome (mitophagy). Within a fused network themitochondria are electrically connected, enabling rapidcommunication over short distances [4]. Over longer distances,communication is facilitated by the trafficking of mitochon-dria which bud off one network and fuse to another. Thesemitochondria move bidirectionally along microtubules, coor-dinated by interaction with the motor proteins dynein (retro-grade transport) and kinesin (anterograde transport), whileshort-range movements along F-actin tracks are mediated bymyosins. Mitochondria are distributed throughout the neuronaccording to the energy demands of different regions. Thesynapse has particularly high ATP requirements in orderto maintain local ionic concentrations, so mitochondrialtrafficking to and proliferation at the synapse are essentialfor synaptic maintenance (see [5] for review).
2.1 Role in disease pathogenesisDefects in either fusion or fission can have severe conse-quences for the cell (Figure 1) [6]. Mutations in Opa1 causeautosomal dominant optic atrophy (ADOA), a progressivedegeneration of the optic nerve and retinal ganglion cells,while mutations in Mfn2 cause Charcot-Marie-Tooth sub-type 2A (CMT2A), a peripheral neuropathy causing axonaldegeneration of sensory and motor neurons (see table inpart I). In these cells, impaired fusion produces highly frag-mented mitochondria and trafficking defects. In addition,alterations to the mitochondrial ultrastructure [7,8] affectADP uptake into the matrix and subsequent ATP produc-tion [9]. Mutations in the fission machinery are extremelyrare because of the severity of the phenotype: one mutationin Drp1 has been observed in a paediatric patient with symp-toms similar to those observed in CMT2A and ADOA, butdisease progression was much more rapid and the patientdied aged 37 days. Inhibiting fission in cultured cells producesa highly fused and immobile reticulum of mitochondria: thisprevents mitophagy, resulting in an accumulation of damagedproteins in the mitochondria. Loss of either fusion or fissionalso results in damaged or absent mitochondrial DNA(mtDNA) and bioenergetic defects, characterised by a reduc-tion in oxygen consumption, ATP production and mito-chondrial membrane potential (Dym). This increasesreactive oxygen species (ROS) production, further damagingmtDNA. Mitochondrial fission is required for mitochondrial
Article highlights.
. Mitochondria mediate numerous processes in neuronsfrom biogenesis to cell death, and defects inmitochondrial function are a feature of almost allneurodegenerative diseases.
. Defects in mitochondrial fission or fusion andtranslocation lead to impaired mitochondrial functionand are observed in numerous neurodegenerativediseases.
. Mutations that impair import of certain proteins intomitochondria underlie a number of neurodegenerativediseases, and a better understanding of the mechanismsinvolved may be useful in targeting novel therapeuticcompounds to the mitochondria.
. Protein quality control in the mitochondria, includingrefolding of misfolded proteins and degradation ofdamaged proteins, is essential for proper mitochondrialfunction.
. The importance of kinase activity at the mitochondria isincreasingly recognised, and these kinases and theirsubstrates may be interesting targets for futuremitochondrial therapeutics.
. Opening of the mitochondrial permeability transitionpore is triggered by mitochondrial dysfunction andusually leads to cell death, making it an attractivetherapeutic target.
. While these mitochondrial defects are less wellcharacterised than those discussed in part I of thisreview, they represent interesting avenues for futureresearch and potentially novel therapeutic strategies.
This box summarises key points contained in the article.
Targeting mitochondrial dysfunction in neurodegenerative disease: Part II
498 Expert Opin. Ther. Targets (2010) 14(5)
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Not
re D
ame
Aus
tral
ia o
n 04
/27/
13Fo
r pe
rson
al u
se o
nly.
proliferation and therefore cell proliferation, and is implicatedin apoptosis [10]. It is important to note, however, thatGTPases involved in fusion and fission may have effects inde-pendent of their roles in mitochondrial dynamics: Opa1-mediated cristae remodelling in apoptosis, for example, hasbeen shown to be independent from its role in fusion [11].
Dysfunction of the fusion/fission machinery is not limited tothese diseases, however. Triggers for mitochondrial fissioninclude oxidative stress and mtDNA damage [12], and recentevidence indicates that cytosolic calcium also plays arole [13,14]. All of these are common features of neuro-degenerative diseases (see part I of this review [15]). Consistentwith this, mitochondrial fragmentation is observed in neuronstreated with amyloid--beta (Ab), a peptide implicated inAlzheimer’s disease (AD) pathogenesis, or with rotenone and3-nitropropionic acid (3-NP), toxins used to model Parkinson’sdisease (PD) and Huntington’s disease (HD) respectively [16].Fragmentation and clustering of mitochondria in the soma isalso observed in the brains of patients with sporadic AD, withaltered expression levels of several fusion/fission proteins [17].
A number of disease-causing mutations have also beenlinked to fission/fusion (see part I of this review [15]). Mutanthuntingtin (htt) forms foci on the outer membrane in a similar
manner to Drp1, and induces ultrastructural changes andabnormal calcium buffering consistent with a defect in mito-chondrial dynamics (for review see [12]). It has recently beenshown that overexpressing htt with 74 or 138 (but not 17 or28) poly-glutamine repeats reduces mitochondrial movementand sensitises cells to oxidative stress-induced fission [18]. Twogenes known to underlie autosomal recessive PD, PINK1 andparkin, have also been linked to mitochondrial fusion and fis-sion in flies and mammals (reviewed in [19]). A recent studyshowed that PINK1-deficient neuroblastoma cells exhibit mito-chondrial fragmentation due to calcineurin-dependent dephos-phorylation of Drp1 [14], consistent with a role for PINK1 incalcium homeostasis (see part I of this review [15]). Poly-glutamine expansion of the gene PPP2R2B, which encodes asubunit of protein phosphatase 2A, causes spinocerebellar ataxiatype 12 (SCA12) [20]. Recent evidence has shown that this pro-tein is a pro-apoptotic phosphatase which is translocated to themitochondrial outer membrane and mediates its effect byinducing mitochondrial fragmentation [21].
Impaired mitochondrial trafficking has also been impli-cated in neurodegenerative disease. Tau, the principal compo-nent of neurofibrillary tangles (NFTs) in AD pathology, is amicrotubule-binding protein and tau overexpression has
Fragmentedmitochondria
Functional mtDNAand proteins
Normal
Dynamic networks ofmitochondria
Excess fusion
Fused reticulum Trafficking defects → ↓ ATP at synapse
Trafficking defects → ↓ ATP at synapse
Ultrastructuralabnormalities
No mtDNA
Damaged mitochondrialcomponents
Excess fission
Mitochondria at synapse → synaptic maintenance
Normal cristaefolding
Figure 1. Effects of dysregulation of the fusion/fission machinery. A comparison of mitochondria in a healthy neuron with
one in which either fusion or fission is impaired.
been shown to alter the kinesin-dependent trafficking of vari-ous organelles, including mitochondria [22]. Trinczek et al.further suggest that tau could control intracellular traffickingby reducing the attachment of kinesin to microtubules [23].Studies in Drosophila and mouse HD models have shownthat htt is important for axonal trafficking of mitochondria(for review see [24]), and mitochondrial transport has alsobeen shown to be disrupted in many models of spastic para-plegia. For example, several mutations have been identifiedin the kinesin motor protein KIF5A in patients with spasticparaglegia type 10 (SPG10; for review see [25]). Finally,PINK1 has recently been shown to form a multiprotein com-plex with Miro and Milton [26], two proteins which, togetherwith kinesin-1, play a crucial function in anterogrademitochondrial transport along microtubules.
2.2 Therapeutic potentialMitochondrial fission and fusion are finely balanced processeswhich vary greatly between cells, making them difficulttherapeutic targets. A chemical inhibitor of Drp1 GTPaseactivity has however been identified and shown to protectmammalian cells against apoptosis [27]. This protection arisesfrom inhibition of mitochondrial outer membrane permeabi-lisation, which appears to be a distinct role of Drp1 unrelatedto its role in fission. Mitochondrial fission is also inhibited bythe compound, named mdivi1, and the authors suggest thismay be of benefit in diseases of the mitochondrial fusionmachinery such as CMT2A or ADOA. It will be interestingto see the effect of mdivi1 in cell and animal models of thesediseases. It would be expected that mitochondrial morphologycould be rescued in this manner, but inhibiting fission in asystem in which fusion is already impaired might have seriousconsequences for mitochondrial maintenance, particularlywith age.The increasing focus on fission and fusion as a mechanism
of mitochondrial maintenance has led some to suggest thatthis could be exploited to repair mtDNA in disease [3,28].In most neurodegenerative diseases mtDNA mutations areheteroplasmic, so wild type mtDNAs capable of comple-menting the mutants are present in the cell. It has thereforebeen suggested that increasing fusion might allow transcom-plementation to restore bioenergetic function [3]. In inheri-ted mitochondrial diseases, wild type mitochondria can beintroduced to the cell by ooplasmic transplantation, wherebyooplasm containing mitochondria from a donor cell isinjected into the fertilised egg before implantation. Inthe USA, this has been successfully used in a clinical settingto produce healthy offspring from mothers with heritablemitochondrial diseases. Although donor mitochondria arepresent at lower levels than the host mitochondria, thesemitochondria are able to replicate and are still detectableby mtDNA fingerprinting in offspring after one year [29].Very recently, nuclear transplantation, in which the pronu-cleus is transferred from the fertilised egg into anotheroocyte, has also been demonstrated to prevent inheritance
of mitochondrial disease in apes [30] and permissionhas been granted in the UK to test the approach inhuman embryos.
The majority of mitochondrial proteins are encoded bynuclear genes, synthesised in the cytosol and imported intomitochondria through protein translocation machinerieslocated in the four intra-mitochondrial compartments:the outer and inner mitochondrial membranes (OMMand IMM, respectively), the intermembrane space (IMS)and the matrix. Import of mitochondrial proteins is adynamic and tightly regulated process that can be altered inresponse to various conditions, such as aging and oxidativestress (for review see [31]). Defective import of mitochondrialproteins has been shown to lead to various disorders includingneurodegenerative diseases. This can result from mutations inthe mitochondrial targeting signal (MTS) or deficiencies inthe translocation machinery or in the mitochondrial chaper-ones required for the correct folding and assembly of newlysynthesised mitochondrial proteins.
3.1 Mutations in the MTSAn MTS is a sequence of amino acids, rich in hydroxyl aminoacids and normally lacking acidic residues, that is recognisedby a mitochondrial receptor that permits mitochondrialproteins synthesised in the cytoplasm to be imported intomitochondria. The precise sequence is not fully defined, butmutations in this region can prevent the protein reaching itsfinal destination, ultimately producing a disease state [31].
The first report of an MTS polymorphism in humansidentified a structural mutation in the MTS of the mito-chondrial superoxide dismutase MnSOD (or SOD2), in aJapanese population. Using a predicted secondary structure,the authors obtained evidence of the capability of thepolymorphism to influence mitochondrial transport. Thispolymorphism has been associated with PD [32], although thisassociation has since been questioned [33,34].
Pyruvate dehydrogenase (PDH) complex deficiency isanother illustration of how a defect in the mitochondrialimport machinery can lead to neurodegeneration. PDHcomplex deficiency is one of the most common neurodegener-ative disorders associated with abnormal mitochondrialmetabolism. The clinical symptoms of patients with a PDHcomplex deficiency can vary considerably and range fromintermittent ataxia to a progressive disease with mental retar-dation and neurological complications. The vast majority ofcases are due to mutations in the X-linked PDH E1a gene(PDHA1). A base substitution was identified in the MTS ofthe PDHA1 gene, resulting in an arginine-to-proline substitu-tion at codon 10 (R10P) [35]. The activity of PDH and PDHE1 has been shown to be greatly reduced in the fibroblastsof these patients [36] and mitochondrial import assays showed
Targeting mitochondrial dysfunction in neurodegenerative disease: Part II
500 Expert Opin. Ther. Targets (2010) 14(5)
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Not
re D
ame
Aus
tral
ia o
n 04
/27/
13Fo
r pe
rson
al u
se o
nly.
that the mutation caused a decrease in the enzymeimport efficiency.
Interestingly, several mutations have been identified inthe MTS of the PD-associated gene PINK1 (for reviewsee [19]). Further studies will be necessary to determinewhether these mutations can affect the mitochondrial importof the PINK1 kinase.
3.2 Deficiencies in translocation machineryAll nuclear-encoded mitochondrial proteins initially entermitochondria via the translocase of the OMM, the TOMcomplex, which mediates insertion into the OMM. Proteinsdestined for the IMS, IMM or matrix possess a cleavableN-terminal MTS, and are addressed to their final destinationby the combined actions of the translocation machinery andthe information encoded by the protein itself (for reviewsee [37]). The TOM complex cooperates with translocases ofthe IMM, TIM22 and TIM23, to mediate insertion intothe IMM or translocation to the matrix respectively.
The human Deafness Dystonia Peptide 1 (DDP1, which ishomologous to the fungal protein TIM8), in complex withTIM13, is directly involved in import by human TIM23.Mutations in the DDP1 gene cause the neurodegenerativedisorder Deafness Dystonia Syndome (DDS), a rare X-linkedcondition characterised by the association of dystonia andprogressive postlingual sensorineural hearing impairment [38].Although various hypotheses have been proposed to explainhow dysfunctional DDP/TIM8 could lead to DDS, it isbelieved that defective TIM8 might impair the biogenesis ofthe TIM23 complex, and therefore the import of matrixproteins ([39]; see table in part I).
3.3 Deficiencies in the molecular chaperonesMitochondria also contain proteases and chaperonesessential to correct protein folding. Mutations in paraplegin(a mitochondrial AAA-protease family member involvedin respiratory chain complex assembly) and in hsp60(a mitochondrial chaperone involved in protein folding)have been implicated in spastic paraplegia types 7 and 13(SPG7 and SPG13), respectively ([40,41], see table in part I).SPG7 and SPG13 are both forms of hereditary spasticparaplegias (HSPs).
3.4 Therapeutic potentialThis field is still relatively unexplored, and the mechanismsand significance of mitochondrial protein import will needto be further elucidated before they can be targeted therapeu-tically. However, pyruvate dehydrogenase (PDH) deficiency,which as we have seen arises from a mutation in the MTS ofthe protein, has been suggested as a possible target for genetherapy. An adenovirus containing full-length PDH wasused successfully to deliver wild-type PDH into mitochon-dria in cultured cells and in vivo in rats [42] and this approachmay be useful in other cases of import deficiency. A betterunderstanding of the mechanisms involved in mitochondrial
import may furthermore be useful in effectively targetingnovel therapeutic compounds to the mitochondria.
4. Protein quality control and mitochondria
The accumulation of non-functional and potentially toxicproteins can be detrimental to the cell, which has thereforedeveloped systems to control the quality of proteins. Thesesystems rely on an army of chaperones to assist the folding/refolding of misfolded proteins, or to target irreparablydamaged proteins for degradation. In most cases, the selectivedegradation of misfolded and potentially toxic proteinsinvolves their ubiquitination and subsequent degradation bythe 26S proteasome (see [43]).
4.1 Ubiquitin-dependent protein quality control
in mitochondriaProteins are typically targeted for proteasomal degradationafter covalent modification with ubiquitin by a systemtermed the ubiquitin--proteasome system (UPS). Linkage ofubiquitin involves three types of enzymes: E1 (ubiquitin-activating enzyme); E2 (ubiquitin-conjugating enzyme); andE3 (ubiquitin ligase). Dysfunction of the UPS has longbeen implicated in the pathology of neurodegenerativediseases by the appearance of ubiquitin-positive neuronalinclusions, including tau-containing NFTs in AD and othertauopathies [44], a-syn-containing Lewy bodies in Parkin-sonian disorders [45] and, most recently, TDP-43-containingaggregrates in amyotrophic lateral sclerosis (ALS) andother diseases [46]. Ubiquitin-dependent protein qualitycontrol (PQC) occurs in the cytosol, the endoplasmic reti-culum (ER) and the nucleus, and until recently UPSdysfunction was considered separate to the mitochondrialdysfunction observed in neurodegenerative disease. How-ever, a growing number of studies now indicate a role ofthe ubiquitin--proteasome pathway in the regulation ofmitochondrial processes (for review see [47]).
Proteasome dysfunction appears to be of particular impor-tance in the pathogenesis of PD. Depletion of 26S protea-somes in mouse neurons causes neurodegeneration andLewy-like inclusions containing mitochondria, suggestingthat mitochondria play a direct role in Lewy body formationand/or function [48]. Interestingly the three PD-associatedproteins parkin, PINK1, and DJ-1 have been shown to forma ubiquitin-E3-ligase complex detected in both cytosolic andmitochondrial fractions, promoting ubiquitination and degra-dation of parkin substrates in neuroblastoma cells andhuman brain lysates [49]. This suggests a direct link betweenthe ubiquitin pathway, mitochondria, and disease patho-genesis. Furthermore, mitochondrial PQC by the proteasomehas been shown to be ubiquitin-dependent and to involve themitochondrial high temperature requirement proteinA2 (HtrA2) [50]. Interestingly HtrA2 has also been recentlysuggested to cleave parkin, thereby inactivating its ubiquitinligase activity [51].
Mitochondria have also been linked to impaired protea-somal degradation as a mechanism of cell death. Aggregationof poly-glutamine-containing N-terminal fragments of httinduces mitochondria-dependent apoptotic cell death as aresult of proteasome inhibition [52], implicating mitochondriaand UPS dysfunction in HD. The same has also been shownfor aggregates of intracellular Ab and amyloid precursorprotein (APP) fragment C99 in AD [53].
4.2 Ubiquitin-independent protein quality controlIn addition to the mitochondrial ubiquitin-dependent PQC,the mitochondrial chaperones and ATP-dependent proteases(including the AAA-protease complexes) monitor proteinquality in the matrix of the organelle. The crucial importanceof this chaperone network has recently been reviewedelsewhere [54].The importance of the mitochondrial ubiquitin-
independent PQC in neurodegenerative diseases is high-lighted by findings that mutations in the chaperone proteinhsp60 and the AAA-protease subunit paraplegin underlieforms of hereditary spastic paraplegia [40,41]. Expression ofthe mitochondrial heat shock protein hsp70 is also decreasedin PD brains and in a cellular model of PD [55], and phos-phorylation of the mitochondrial chaperone TNF-associatedprotein 1 (TRAP1, also called hsp75) by PINK1 has beenshown to protect against oxidative stress [56].Accumulation of defective proteins in the mitochondrial
matrix is known to activate retrograde signalling pathwaysfrom the mitochondria to the nucleus. These responses arereferred to as mitochondrial unfolded protein responses(UPRmt), by analogy to the UPR in the ER [57]. Moisoi et al.showed that the loss of the protease HtrA2 contributed todefects in the electron transport chain (ETC) and enhancedROS production and accumulation of unfolded proteins inmitochondria. Subsequent mitochondrial dysfunction resultedin activation of a brain-specific nuclear transcriptional responseto stress, suggesting an important role for the UPRmt in thepathogenesis of neurodegeneration [58].
4.3 MitophagyThe other protein degradation route is the autophagy/lysosome pathway. Autophagy involves the sequestration ofcytoplasm by double-layered membranes to form vesiclescalled autophagosomes, which ultimately fuse with lysosomes,in which their contents are degraded. However, it is becomingincreasingly apparent that autophagy can also be a selectiveprocess that degrades various organelles, such as mitochon-dria, ribosomes or peroxysomes. Autophagic degradation ofmitochondria is called mitophagy. Mitophagy is a selectiveprocess [59] suggested to play an important role in PQC.Although the molecular mechanism is still poorly under-
stood, a growing body of evidence suggests a role for mitophagyin the pathogenesis of neurodegenerative diseases, includingHD [60] and PD. In the latter, the level of mitophagy hasbeen shown to correlate with ERK activity levels [61],
supporting a role for the ERK/MAPK pathway in regulatedautophagy in PD. Loss of PINK1 function has also been shownto promote mitophagy through effects on oxidative stress andmitochondrial fission [62], and Parkin has been shown topromote mitophagy of dysfunctional mitochondria [63].
4.4 Therapeutic potentialMitochondrial protein degradation is a relatively new field, andthere remain unanswered questions about the mechanismsinvolved and its significance in neurodegenerative disease. Asunderstanding advances, however, these systems may be animportant target for future therapeutic compounds.
5. Mitochondrial kinases: new targets forneurodegeneration?
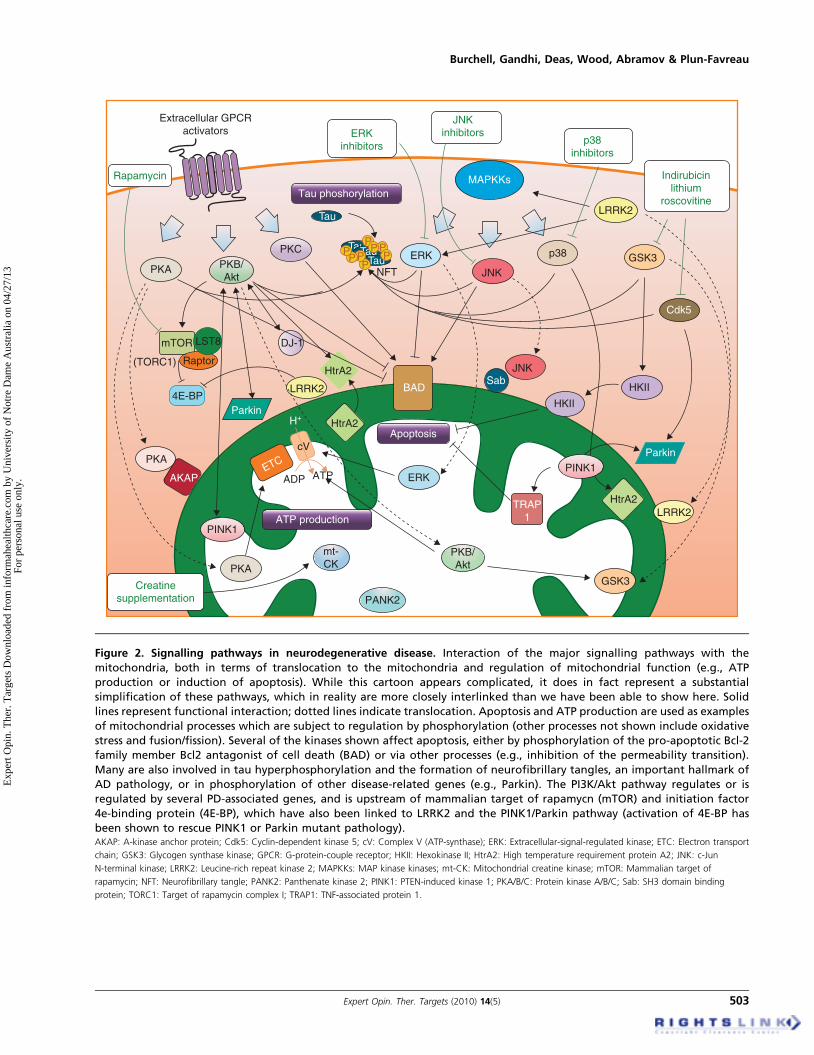
Mitochondria have long been viewed as an unlikely site forsignalling by reversible protein phosphorylation. This ispartly due to the compartmentalisation of the organellefrom the rest of the cell and to the absence of an MTS inmost signalling proteins. However, in recent years numerousstudies have demonstrated that phosphorylation is crucial forregulation of mitochondrial processes (for review see [64]),raising several interesting questions. Are most of the neces-sary kinases normally present within mitochondria? Or dothey only gain entry following injury? Recent studies haveconsistently shown that certain components of well-knownkinase signalling cascades can be specifically targeted tomitochondria (Figure 2), where they may be imported viaan MTS or associate with scaffold proteins such as A-kinase anchor protein (AKAP) or SH3 domain binding pro-tein (Sab) on the surface (see [65]). These kinases have beenshown to play key roles in the regulation of mitochondrialprocesses including ATP production and apoptosis. Effectson apoptosis may be mediated via phospho-activation or -inhibition of Bcl2 family members such as Bcl2 antagonistof cell death (BAD) (e.g., MAPKs, reviewed in [65]), by inter-fering with permeability transition pore (PTP) opening (e.g.,hexokinase II; [66]), or by suppressing cytochrome c release(e.g., phosphorylated TRAP1; [56]).
Mitochondrial kinases have been directly implicated inneurodegenerative disease pathogenesis, since mutations inthe genes encoding pantothenate kinase 2 (PANK2) andPINK1, both of which harbour a predicted MTS, are associ-ated with recessive progressive extrapyramidal and dementiasyndromes and PD respectively [67,68]. This section willspecifically focus on kinases which can translocate to or actat the mitochondria.
5.1 Mitogen activated protein kinasesMAPKs are a family of conserved serine/threonine proteinkinases that play a critical role in transducing multiple signalsfrom the cell surface to the nucleus in all eukaryotic species.More recently, MAPKs have emerged as important regulatorsof neuronal responses to both functional and pathological
Targeting mitochondrial dysfunction in neurodegenerative disease: Part II
502 Expert Opin. Ther. Targets (2010) 14(5)
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Not
re D
ame
Aus
tral
ia o
n 04
/27/
13Fo
r pe
rson
al u
se o
nly.
Tau
Tau
TauTauP
PPP
PPP
P
PKA
PANK2
TRAP1
HtrA2
Parkin
PKB/Akt
ERK
ATP production
ADP ATP
H+HtrA2
Parkin4E-BP
AKAP
mTOR
PKA
PKCERK
JNKNFT
Tau phoshorylation
JNKinhibitors
LRRK2
p38 GSK3
Cdk5
HKII
p38inhibitors
Extracellular GPCRactivators
PKB/Akt
HtrA2
ETC
Apoptosis
BADSab
JNK
PINK1
mt-CK
Indirubicinlithium
roscovitine
MAPKKsRapamycin
Creatinesupplementation
LRRK2
GSK3
PINK1
cV
LRRK2
DJ-1
HKII
PKA
ERKinhibitors
(TORC1) Raptor
LST8
Figure 2. Signalling pathways in neurodegenerative disease. Interaction of the major signalling pathways with the
mitochondria, both in terms of translocation to the mitochondria and regulation of mitochondrial function (e.g., ATP
production or induction of apoptosis). While this cartoon appears complicated, it does in fact represent a substantial
simplification of these pathways, which in reality are more closely interlinked than we have been able to show here. Solid
lines represent functional interaction; dotted lines indicate translocation. Apoptosis and ATP production are used as examples
of mitochondrial processes which are subject to regulation by phosphorylation (other processes not shown include oxidative
stress and fusion/fission). Several of the kinases shown affect apoptosis, either by phosphorylation of the pro-apoptotic Bcl-2
family member Bcl2 antagonist of cell death (BAD) or via other processes (e.g., inhibition of the permeability transition).
Many are also involved in tau hyperphosphorylation and the formation of neurofibrillary tangles, an important hallmark of
AD pathology, or in phosphorylation of other disease-related genes (e.g., Parkin). The PI3K/Akt pathway regulates or is
regulated by several PD-associated genes, and is upstream of mammalian target of rapamycn (mTOR) and initiation factor
4e-binding protein (4E-BP), which have also been linked to LRRK2 and the PINK1/Parkin pathway (activation of 4E-BP has
been shown to rescue PINK1 or Parkin mutant pathology).AKAP: A-kinase anchor protein; Cdk5: Cyclin-dependent kinase 5; cV: Complex V (ATP-synthase); ERK: Extracellular-signal-regulated kinase; ETC: Electron transport
chain; GSK3: Glycogen synthase kinase; GPCR: G-protein-couple receptor; HKII: Hexokinase II; HtrA2: High temperature requirement protein A2; JNK: c-Jun
stimuli (for review see [69,70]). The MAPK family encompassesthe extracellular signal-regulated protein kinases (ERKs) andthe stress-activated protein kinases (SAPKs), which includeboth JNKs and p38 kinases. Whilst the role of MAPKs in neu-rodegenerative disease remains elusive, a specific role forthe MAPK pathway has been suggested in oxidative stress,one of the most common mechanisms implicated in neuronalcell death in neurodegenerative disease (for review see [70,71]).Historically, the ERK cascade is considered a critical path-
way for mitogenesis and differentiation, whereas activation ofthe JNK and p38 pathways is involved in the stress response.All three pathways are however thought to play a role in thephysiopathology of PD and dementia with Lewy bodies(DLB). Phospho-ERK, phospho-JNK and phospho-p38were all detected in neurons in DLB [72], and phospho-ERKwas specifically detected in mitochondria of dopaminergicneurons in PD and DLB disease brains (for review see [69]).The mitochondrial protease HtrA2 has been shown to bephosphorylated after activation of the p38 pathway in aPINK1-dependent manner [73], suggesting an important rolefor PINK1 and p38 kinases in the pathogenesis of PD. Inter-estingly, the PD-associated kinase leucine-rich repeat kinase2 (LRRK2), which shows a degree of localisation to the mito-chondria [74], appears to activate ERK signalling in SH-SY5Ycells [75]. Furthermore LRRK2 has been shown to phospho-rylate MKK3/6 and MKK4/7 (the upstream kinases of p38and JNK respectively) in vitro, emphasizing the role of thesepathways in the pathogenesis of PD [76]. The MAPKs arealso thought to play a role in the physiopathology of ADand may be of particular importance in the phosphorylationof tau (for review see [70]). Hyperphosphorylation of tau isrequired for its aggregation to form the NFTs observed inthe brains of patients with tauopathies [77], and is thus impor-tant in AD pathology. Finally the activation of the JNKcascade has been observed in cellular and animal modelsof HD, where JNK activation precedes the neuronal cell deathcaused by expanded htt [78,79]. Taken together these studiesindicate that ERK, p38 and JNK play important roles in thepathogenesis of neurodegenerative disorders. Nonetheless,more experiments will be required to further investigate theprecise role of MAPKs in these diseases.
5.2 Protein kinases A, B and CMany other kinases have been shown to localise or translocateto the mitochondria, in particular protein kinases A, B (alsocalled Akt) and C (for review see [65]). The PI3K/Akt pathwayhas been associated with all three of the PD-associatedproteins PINK1, parkin and DJ-1, suggesting an importantrole of this signalling pathway in the pathogenesis of PD (forreview see [80]). PKCd is associated with 6-hydroxydopamine-induced neurotoxicity and Parkinsonian neurodegenera-tion [81]. These kinases are also thought to play a role inthe physiopathology of AD, since the Akt and PKA pathwayshave both been shown to phosphorylate tau. Furthermore,deregulation of Akt and alteration of mTOR (a downstream
signalling component of the Akt pathway) activity has beenfound to be associated with AD pathology (for reviewsee [82]). Although mTOR is primarily located in the cytosol,subcellular fractionation revealed that it can be purified in themitochondrial fraction, where it has been shown to be neces-sary for the maintenance of mitochondrial oxidative func-tion [83]. Very recent studies in flies have also suggested arole for 4E-BP, a downstream target of Akt and mTOR,in PD [84,85]. Interestingly rapamycin, an inhibitor of themTOR signalling pathway, extends healthy lifespan not onlyin invertebrates but also in mice [86]. These findings mayhave implications for further development of interventions tar-geting the mTOR pathway for the treatment and preventionof neurodegenerative diseases, for most of which age is theprimary risk factor.
5.3 Proline-directed kinasesOver-activation of proline-directed kinases such as cyclin-dependent kinase 5 (Cdk5) and glycogen synthase kinase3 (GSK3) has also been implicated in the aberrant phospho-rylation of tau, and therefore the pathogenesis of AD [87].Cdk5 has also been shown to promote oxidative stress andmitochondrial dysfunction, two events that play a pivotalrole in neurodegeneration [88]. Moreover Cdk5 has beenshown to phosphorylate the PD-associated protein parkin,thereby regulating its E3 ubiquitin ligase activity [89].
5.4 Other kinasesWhile several kinase pathways are subject to regulation byreactive oxygen and nitrogen species (ROS and RNS; forreview see [65]), others have been implicated in ROS produc-tion and regulation. Mitochondrial hexokinase (mt-HK)(a glycolytic enzyme) and creatine kinase (mt-CK) (anenergy-buffering enzyme) activities have both been shownto regulate mitochondrial ROS generation in rat [90].Gimenez-Cassina et al. have recently shown that mt-HK IIis a promoter of neuronal survival under the regulation ofGSK3. This kinase may therefore be relevant to neurode-generative diseases in which mitochondrial dysfunction is aprominent feature [66]. Finally it has been suggested thatany disturbance of mt-CK function may exacerbate the ADand ALS disease process.
5.5 Therapeutic potentialKinase inhibitors may have therapeutic potential for severalneurodegenerative diseases and for AD in particular, sincetau hyperphosphorylation is directly implicated in diseasepathogenesis (see [91] for review). Inhibitors of the MAPKpathways have been developed for other diseases (for example,the JNK inhibitor ACTIMID has just completed a Phase Itrial and is entering a Phase II trial for prostate cancer) butmay also be useful in treating neurodegenerative disease.The JNK inhibitor SP600125 and the p38 inhibitorSB239063 might have therapeutic potential since theyhave recently been shown to prevent amyloid-induced
Targeting mitochondrial dysfunction in neurodegenerative disease: Part II
504 Expert Opin. Ther. Targets (2010) 14(5)
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Not
re D
ame
Aus
tral
ia o
n 04
/27/
13Fo
r pe
rson
al u
se o
nly.
neuroinflammation [92] and to protect against MPP+-induced neuronal death in cultured neurons [93], respec-tively. ERK2 inhibitors may have potential in AD, sincean ERK2 inhibitor K252a was shown to inhibit tau hyper-phoshorylation and prevent severe motor impairments intau transgenic mice [94].
Inhibitors of the proline kinases GSK3 and Cdk5 haveadditionally generated interest in AD because of their role intau phosphorylation. Indirubin-3¢-monoxime, an inhibitorof cyclin-dependent kinases, was shown to reduce tau phos-phorylation and b-amyloid toxicity in cultured cells [95].Similarly lithium was shown to block production of Ab byinhibiting GSK3a [96], although a subsequent study foundthat it reduced tau phosphorylation but had no effect on Abproduction [97]. Derivatives of roscovitine, a broad-specificityblocker of cyclin-dependent kinases and casein kinase 1, havealso been suggested as therapeutics for AD [98].
These findings are very encouraging, but a number of issuesstill need to be addressed. The signalling pathways discussedhere form a highly interconnected network, as illustrated bythe convergence of the ERK and p38 pathways in AD [99].One of the challenges for scientists will therefore be to eluci-date the interactions between these pathways in order to fullyunderstand the downstream effects of pharmacological inhibi-tion. Specificity can also be a problem for kinase inhibitors,since they are often targeted to the highly conserved ATPbinding site and can therefore have off-target effects. A novelseries of substrate-competitive JNK inhibitors have howeverbeen found to be more selective [100], and may open avenuesfor future research. Another interesting area for future investi-gation will be those mitochondrial kinases (such as PINK1)whose physiological function and substrates remain unknownor poorly understand, since these substrates are potentiallyimportant therapeutic targets. If these challenges can beaddressed, the therapeutic potential of these pathways couldbe significant.
6. The mitochondrial permeability transition
Neuronal death in neurodegenerative disease can occur viavarious routes, including apoptosis (caspase-dependent pro-grammed cell death), and necrosis (caspase-independent celllysis). Apoptosis may be induced by an intrinsic pathwayinvolving cristae remodelling and outer membrane permeabi-lisation, or an extrinsic pathway triggered by direct activationof caspases independent of mitochondria. These pathwayshave been extensively reviewed elsewhere (e.g., [101]) and arebeyond the scope of this review. Apoptosis may be inducedin response to mitochondrial dysfunction by several indirectmechanisms: for example, redox signalling pathways canlead to transcriptional upregulation or activation of apoptoticmolecules in response to cytosolic ROS [102]. Alternativelymitochondrial dysfunction can directly trigger apoptotic ornecrotic cell death by causing opening of the PTP (for reviewsee [103]).
6.1 The permeability transition poreThe PTP is a voltage dependent pore in the IMM, permeableto water and solutes smaller than 1.5 kDa. The molecularcomposition of the pore remains controversial (for reviewsee [104]) and it can be activated or inhibited by numerouscompounds. Prolonged opening of the PTP causes collapseof the Dym as the proton gradient equilibrates acrossthe IMM, and subsequent failure of ATP production atthe ETC. This may lead to necrotic cell death if sufficientnumbers of mitochondria undergo the permeability transitionsimultaneously; alternatively, mitochondrial swelling dueto the colloidal osmotic pressure can cause unfolding of thecristae and rupture of the OMM, releasing pro-apoptoticmolecules such as cytochrome c, HtrA2 and apoptosis-inducing factor (AIF) into the cytosol and resulting inapoptotic cell death.
PTP opening is regulated by factors that include mitochon-drial calcium, redox potential, pH and kinase activity [105],and can thus be triggered by several of the mitochondrialdysfunctions discussed in this review. The PTP appears tohave at least three redox-active sites and is activated bymitochondrial ROS [106], another mechanism by which oxida-tive stress induces cell death. Respiratory chain dysfunctionaffects pH and Dym, both of which are regulators of PTPopening (see [105]), and complex I deficiency has recentlybeen shown to shift the voltage threshold for PTP openingto near resting potentials [107]. Calcium overload is an impor-tant trigger of PTP opening (see part I of this review [15]), andin cells with impaired calcium homeostasis (e.g., impairedmitochondrial Na+/Ca2+ exchanger) the PTP opens morereadily [108]. Various kinases have also been shown tomodulate PTP opening (see [105]), including GSK-3b andthe stress activated kinase p38.
6.2 Therapeutic potentialPTP opening is a common pathway to cell death for many ofthe mitochondrial defects observed in neurodegenerativedisease, and therefore an interesting therapeutic target. Thepharmacology of the pore is complex, and several inhibitorshave numerous additional effects. One inhibitor, the antibi-otic cyclosporin A, has however been proposed for thetreatment of traumatic brain injury [109] and for Ullrichcongenital muscular dystrophy [110], while another, sangli-fehrin, is under investigation in cardiac dysfunction [111].As understanding of the pore composition improves, PTPinhibition may be an exciting new therapeutic target innumerous disease areas.
7. Expert opinion and conclusions
In the first part of this review we discussed the roles of oxida-tive stress, bioenergetic dysfunction and calcium mishandlingin neurodegeneration, and the therapies which currentlytarget these mitochondrial defects. This has been an area ofactive research for some time, with therapies such as coenzyme
Q10 (CoQ10) going into clinical trials nearly a decade ago, butas yet the efficacy of these compounds has not been conclu-sively demonstrated. In this part of the review we have focusedinstead on mitochondrial defects which have more recentlybeen linked to neurodegeneration. In some cases, potentialtherapeutic compounds are already approved for use in otherdiseases, for example MAPK inhibitors for prostate cancer orcyclosporin A as an antibiotic. Other defects such as deficientprotein import and degradation are less well understood andmore difficult to target, but as research continues in theseareas and molecular mechanisms become better defined theymay become targets for new therapeutic strategies. Otherforms of mitochondrial therapeutics may also be increasinglyimportant. In this review we have focused primarily on phar-macological intervention but gene therapy is an alternativeapproach, enabling correction of enzyme deficiencies such aspyruvate dehydrogenase deficiency [42], downregulation ofgenes such as a-syn [112] or upregulation of genes such asapoptosis inhibitors [113]. Work in PD cybrid cells suggeststhat mitochondrial gene therapy, whereby wild type mtDNAis targeted to the mitochondria using a novel technology, mayalso have potential [114]. These approaches are still relativelynovel, but early reports in cultured cells and animal modelsare promising.Another topic discussed in detail in part I of this review
[15] is the degree of interrelation between the mitochondrial
defects observed in neurodegeneration. In this part we haveseen a continuation of this theme (Figure 3), with mitochon-drial fragmentation induced in response to oxidative stressand cytosolic calcium and deficiencies in the fusion/fissionmachinery inducing bioenergetic defects and ROS produc-tion. This interrelation may have implications in consideringtherapeutic strategies, since targeting one defect mayaffect another.
In this review we have discussed the potential of mitochon-drial therapies at length, but it is important to understandtheir limitations. Patients with neurodegenerative diseasesonly show symptoms after a substantial amount of neuronaldeath, and no mitochondrial therapy can replace those lostneurons. By targeting sick mitochondria in the survivingneurons these therapies might partially improve functionand slow disease progression, but it is likely that they willbe most effective if given to patients in the earliest diseasestages. Early diagnosis is critical for effective treatment ofneurodegenerative disease, and research is ongoing to findbiomarkers (e.g., oxidative damage in platelets) for presy-mptomatic disease. Mitochondrial therapies might alsobe useful in combination with existing therapies, to slowdisease progression while the symptoms are alleviated bytraditional means.
Other approaches are also under investigation. In somePD patients foetal stem cell grafts have been effective [115],
Mitochondrialremodelling→ apoptosis
PTP opening
Calciumdyshomeostasis
Fusion/fissionimbalance
UltrastructuralchangesMitochondrialdamage
Bioenergeticdysfunction/
energy decline
ROS →fission
↑O2• fromETC
Ox damagePARP activation
Oxidativestress
MAPKs
Redoxsignalling
Altered kinaseactivity
NOX/NOS/MAOactivation
PTP openingPARP1 signalling
Necrosis
Ca2+ → fission
Cell death
Apoptosis
Ca2+-activatedkinase
Figure 3. Routes to cell death. Different types of mitochondrial dysfunction interact in numerous and complex ways: this
figure highlights a few examples. For more information on oxidative stress, bioenergetic dysfunction and calcium
dyshomeostasis, see part I of this review.ETC: Electron transport chain; MAO: Monoamine oxidase; MAPK: Mitogen-activated protein kinase; NOS: Nitric oxide synthase; NOX: NADPH oxidase; O2.: Super-
Targeting mitochondrial dysfunction in neurodegenerative disease: Part II
506 Expert Opin. Ther. Targets (2010) 14(5)
Exp
ert O
pin.
The
r. T
arge
ts D
ownl
oade
d fr
om in
form
ahea
lthca
re.c
om b
y U
nive
rsity
of
Not
re D
ame
Aus
tral
ia o
n 04
/27/
13Fo
r pe
rson
al u
se o
nly.
and mesenchymal stem cells have been safely used in ALSpatients with mixed clinical results (for review see [116]). Intwo PD patients successfully receiving grafts, Lewy bodieshave recently been found in the grafted neurons upon post-mortem [117], raising interesting questions about themechanism of host to graft transfer of pathology, and possi-ble shortcomings of stem cell therapy as a cure for neurode-generative disease. The patients in question had not sufferedany impaired function in the grafted neurons, however, andthere is every evidence that grafted neurons can remain func-tionally integrated and continue to release dopamine over adecade after the operation. In the future induced pluripotentstem cells (iPS cells) may be used to avoid immune reactionsto the graft and ethical considerations. iPS cell therapies aimto replace the missing neurons using patient fibroblastswhich have been reprogrammed to iPS cells. iPS cells havesuccessfully been generated from a patient with ALS and dif-ferentiated into motor neurons in vitro [118]. If this approachworks in patients, the therapeutic potential could beenormous. This technique is still its infancy however, and it
will be several years before it is ready for use in patients. Mean-while, research into mitochondrial dysfunction is producingever more potential targets, and existing compounds are show-ing promise. As emerging biomarkers enable earlier diagnosis,these therapies could play a key role in the future treatmentof neurodegenerative disease.
Declaration of interest
VS Burchell is funded by Medical Research Council (MRC)and Brain Research Trust. H Plun-Favreau is funded bythe MRC and the Clinical Research & DevelopmentCommittee (CRDC) from University College London Hos-pital (UCLH) charities. S Gandhi is funded by the WellcomeTrust. E Deas is funded by the Parkinson’s Disease Societyand Brain Research Trust. NW Wood is funded by theMedical Research Council, Parkinson’s Disease Society andWellcome Trust. AY Abramov is funded by the Parkinson’sDisease Society and the Clinical Research & DevelopmentCommittee. The authors state no conflict of interest.