99
TD ! DFT applied to biological problems Marcus Elstner TU Braunschweig

TD!DFT applied to biological problems
Marcus ElstnerTU Braunschweig

0) biological structures

N
Biological structures: proteins, DNA, lipids
Amino acids: 20DNA bases: 4 (A,C,G,T)
lipids

- contain (mainly) H, C, N, O and S
- units: amino acids
N
Proteins

- contain (mainly) H, C, N, O and S
- units: amino acids
N
Proteins

Proteins

Proteins

Proteins

Photosynthetic Reaction CenterAquaporin
Photochemistry
bR
Different representations of backbone

DNA

1) Intro

Optical properties in biological systems: examples
• bioenergetics: photosynthesis
• vision
• avoiding radiation damage: DNA protection and repair
• biological fluorescence: from jellyfish to biological markers

Three pigments, same chromopor: what determines the color?
Vision

• two different types of cells involved in vision
• rod cells for dim-light vision (500 nm, rhodopsin)
• cone cells (425nm, 533 nm, 560 nm) for color discrimination
Vision

• 7-helix proteins
• G-protein coupled receptors (GPCR’s)
• cone cells (425nm, 533 nm, 560 nm) for
color discrimination
• differ in amino acid composition
Vision

Reaction coordinate efficiencyalcohol not specific 10 ps 0.1
Rh cis-trans <0.5 ps 0.65
- intiates structural response => signalling state
cis trans
After photon absorption:

Questions:
cis trans
can we calculate the
excitation energy of
Rhodopsins?
can we describe the
isomerization
process?

Answers:
cis trans
Rh exp: 500 nm
TD-B3LYP: 505 nm
(JPCB 112 2007 6814)
(also Biophys J. 87 2004 2931)
DFT-QM/MM simulation
of isomerization
process:
JACS 126 2004 15328
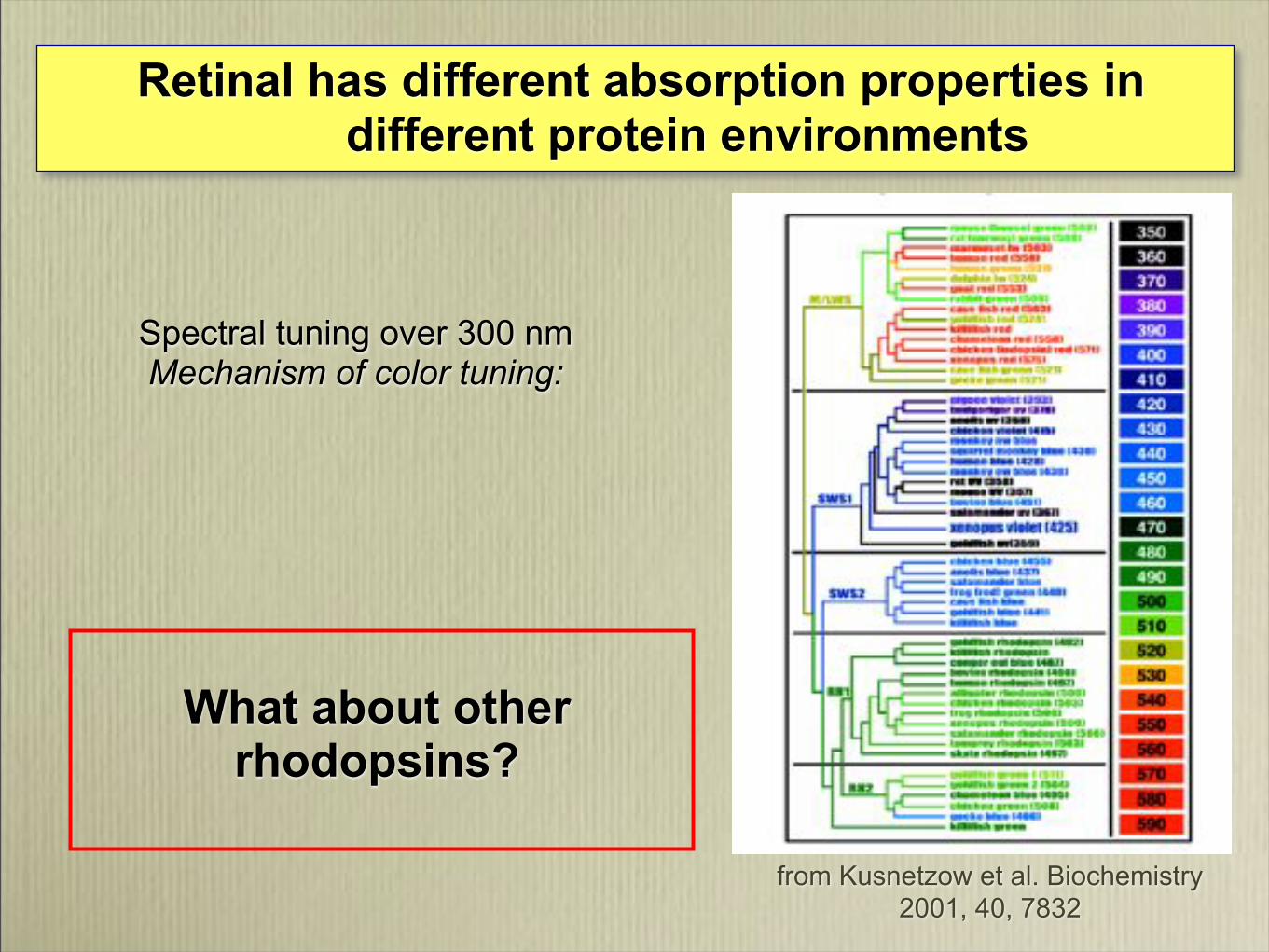
Spectral tuning over 300 nmMechanism of color tuning:
Retinal has different absorption properties in different protein environments
What about other rhodopsins?
from Kusnetzow et al. Biochemistry
2001, 40, 7832

e.g. Bacteriorhodopsin
• transmembrane protein
• 7 a-helices
• retinal chromophor

e.g. Bacteriorhodopsin
• transmembrane protein
• 7 a-helices
• retinal chromophor

Bioenergetics
bR is simplest light driven proton pump

bR photocycle
absorbs at 570 nm, e.g. 70 red-shifted with respect to Rh

and one more: SRII
SRII absorbs as Rh at 500 nm, i.e. 70 red-shifted with respect to bR

TD-DFT (B3LYP) exp.
bR 2.57 eV 2.18 eV (570nm)
SRII 2.58 eV 2.48 eV (500nm)
Rh 2.52 eV 2.49 eV (498nm)
JCTC 3 (2007) 605
JPCB 112 2007 6814
Theor Chem Acc (2003) 109:125
Is TD-DFT color blind?

cis trans
Further
DFT-QM/MM simulation
of isomerization
process:
JACS 126 2004 15328
why did they heat the
chromophore to
690K?

Some other interesting systems
• bioenergetics: photosynthesis
• vision
• Avoiding radiation damage: DNA protection and repair
• Biological fluorescence: from jellyfish to biological markers

1! light absorption
2! proton transfer
3! ATP synthesis
Bacterial Photosynthesis
" bacterial reaction center" bacteriorhodopsin

bacteriorhodopsin bacterial reaction center
Bacterial Photosynthesis

- photon absorption- energy transfer- electron transfer- proton transfer
- QB movement:
large structural transitions
Bacterial Reaction Center

Special pair
special pair: chlorophyll
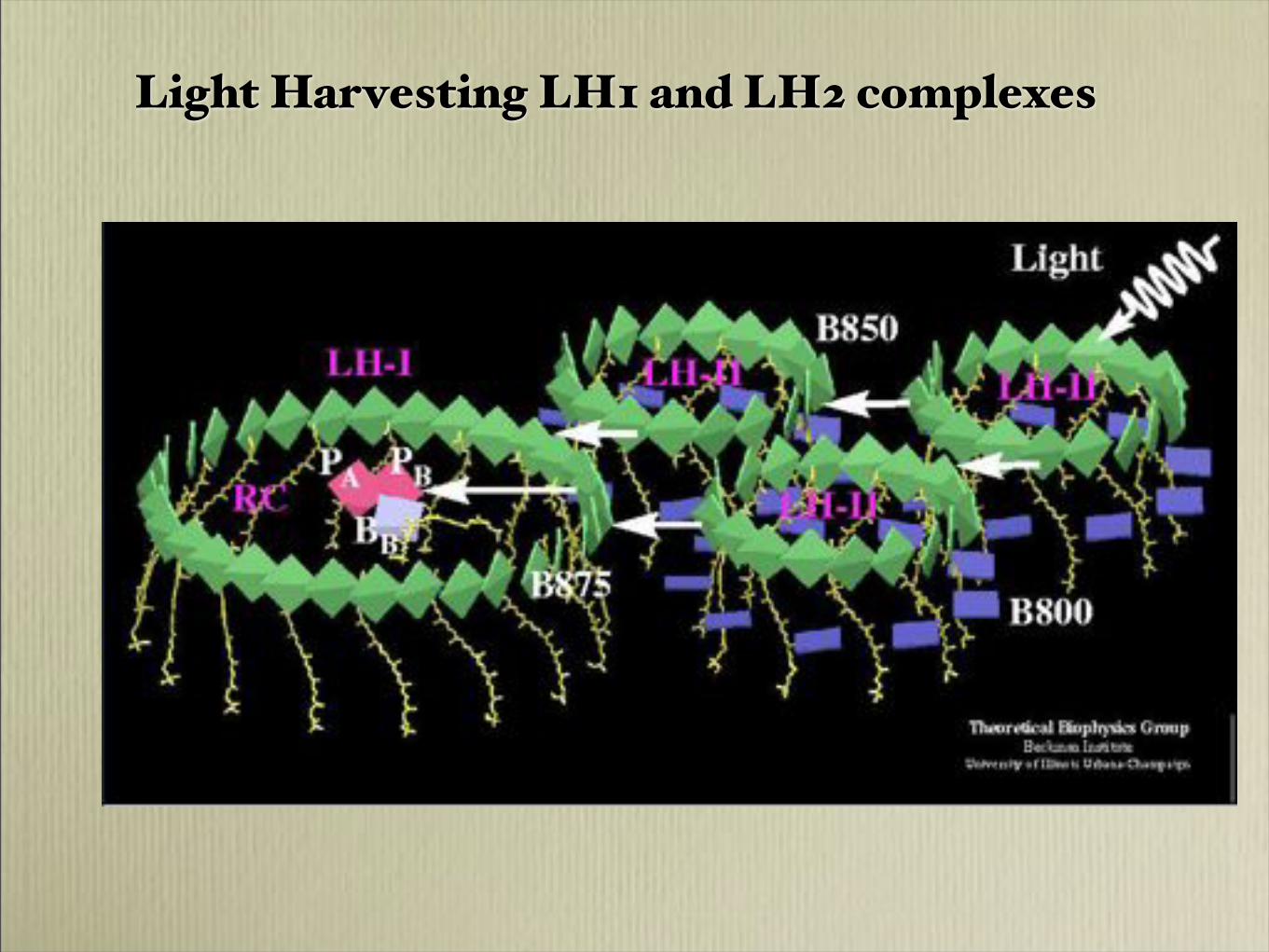
Light Harvesting LH1 and LH2 complexes

- Antenna complex
- BChl800 (green)- BChl850 (red) - carotenoids (yellow)
LH2
from Sundström ARPC 2007

Chromophors
Mg ! porphyrenepolyenes

- Shomomura discovered GFP from Aequorea victoria
- Similar proteins exist in a big number of jellyfish with different wavelenghts of emission
- GFP can be expressed by bacteria
- chromophore build from amino acids: good marker for molecular biology
Green fluorescent protein (GFP)

- green fluorescence 508 nm
- dual absorption: 395 nm (neutral form) A 475 nm (anionic form) B 4:1 fraction (A:B)
- Conversion via PT and structural change of Thr 203

DNA
how does it avoid radiation damage?
- rapid radiationless de-excitation via conical intersections - N-H stretch coordinate - out-of-plane motion
- transport of radical cations over large distcances

2) simulating biological structures

simulating biological structures
complexity has 2 dimensions:
1) size: number of atoms2) time scale of processes: need MD to sample conformational space
environment of acitve site looks caotic,but is highly structured from a functional perspective
=> do not neglect environment, otherwise you loose the most important point!

active site: chromophore + X
• Size: 4000-15000 atoms (water)
• actives site:50-100 atoms
• excited states, chemical reactions => QM
• size, ns MD simulations => MM
The computational problem

models in theoretical chemistry/biophysics
CI, MPCASPT2
Length scale
Continuum Electrostatics “Coarse graining”
Force Fields:MM
f
s
ps
ns
tim
e
ApproximativeMethods
HF, DFT
nm10"" 100"" 1000"" 10.000 " atoms

© Grubmüller
Molecular Mechanics: MM

Quelle: GrubmüllerMPI Göttingen

For Protein- and DNA ok!
Problems.:
- Polarization
- Charge transfer
- no reactions!
kb
k!
k"
qjqi
fixed point charges
Molecular Mechanics: MM

models in theoretical chemistry/biophysics
CI, MPCASPT2
Length scale
Continuum Electrostatics “Coarse graining”
Force Fields:MM
f
s
ps
ns
tim
e
ApproximativeMethods
HF, DFT
nm10"" 100"" 1000"" 10.000 " atoms

Continuum electrostatics
!"#$%&'()*&+$#+#,-.$/&+0)1+%$0",##$
0#,2%3
!"1%$/&+0,145*&+$/")+-#%$)''$6,&6#,*#%7$'18#
#+#,-1#%7$9&,/#%$:1;#;$-#&2#0,.<7$(14;$9,#=5#+/1#%$>$
!#?0

Continuum electrostatics
9&,2)*&+$&9$/)(10.$)+@$A@B$1+0#,)/*&+%$&9$%&'50#$C10"$%&'(#+0
2&@#'#@$)%$%52
&(#,$%5,9)/#$),#)%$D1$
&9$)0&2%

Continuum electrostatics
E+0#,)/*&+$&9$2&'#/5'),$/"),-#%$C10"$#'#/0,&%0)*/$6&0#+*)'$F:,<

Continuum electrostatics
G&(#$0&$9,##$#+#,-.$%5,9)/#$4.$H%)26'1+-I$&(#,$6&%%14'#$%&'(#+0$
/&+0,145*&+%$:#+0,&6.<$)+@$4.$1+/'5@1+-$0"#$6A$0#,2;
!"1%$/)+$4#$#J#/*(#'.$@&+#$@5,1+-$6),)2#0,1K)*&+$&9$0"#$2&@#'
L,##$#+0")'6.3
M)'/5')0#$,#)/*&+$N#'@

Continuum electrostatics
G&(#$0&$9,##$#+#,-.$%5,9)/#$4.$H%)26'1+-I$&(#,$6&%%14'#$%&'(#+0$
/&+0,145*&+%$:#+0,&6.<$)+@$4.$1+/'5@1+-$0"#$6A$0#,2;
!"1%$/)+$4#$#J#/*(#'.$@&+#$@5,1+-$6),)2#0,1K)*&+$&9$0"#$2&@#'
L,##$#+0")'6.3
M)'/5')0#$,#)/*&+$N#'@
O/"1#(#2#+03$
C#$@&$+&0$")(#$0&$%)26'#$&(#,$)''$%&'(#+0$@#-,##%$&9$9,##@&2P

Continuum electrostatics
Q&,+$2&@#'$:RSTU<
V#+#,)'1K#@$Q&,+3
N+@$#261,1/)'$9&,25'),7$C"1/"$
@#0#,21+#%$0"#$Q&,+$,)@15%$&9$)0&2$1$
1+%1@#$0"#$6,&0#1+

Continuum electrostatics
W&1%%&+$Q&'0K2)++3$%&'(#$W&1%%&+$#=;$&+$)$-,1@$>$
>$C10"$)+@$C10"&50$/&+*+552$),&5+@$0"#$2&'#/5'#3$-#0$,#)/*&+$N#'@

Continuum electrostatics
W&1%%&+$Q&'0K2)++3$%&'(#$W&1%%&+$#=;$&+$)$-,1@$>$
>$C10"$)+@$C10"&50$/&+*+552$),&5+@$0"#$2&'#/5'#3$-#0$,#)/*&+$N#'@
!"#+$650$1&+%$C10"$/&+/#+0,)*&+$+1$)+@$/"),-#$=1

Poisson Boltzmann (PB) vs. Generalized Born (GB)
WQ$(#,.$#?6#+%1(#7$#(#+$9&,$GG3$/)+$+&0$4#$@&+#$
#?0#+%1(#'.$)'&+-$GX$%125')*&+%
V#+#,)'1K#@$%&'(#+0$4&5+@),.$6&0#+*)'$:VDQW$4.$Q;$
Y&5?<
!$N?$)+$&50#,$,#-1&+
!$/)'/5')0#$,#)/*&+$N#'@$)+@$%&'()*&+$9,##$#+#,-.$5%1+-$
)$4)%1%$%#0$9&,$0"#$1++#,$,#-1&+
"$A#,.$-&&@$9&,$6,&4'#2%7$C"#,#$&+#$/)+$+#-'#/0$0"#$
2&*&+$&9$0"#$&50#,$6),0
&50#,$,#-1&+3$N?#@
1++#,$
,#-1&+
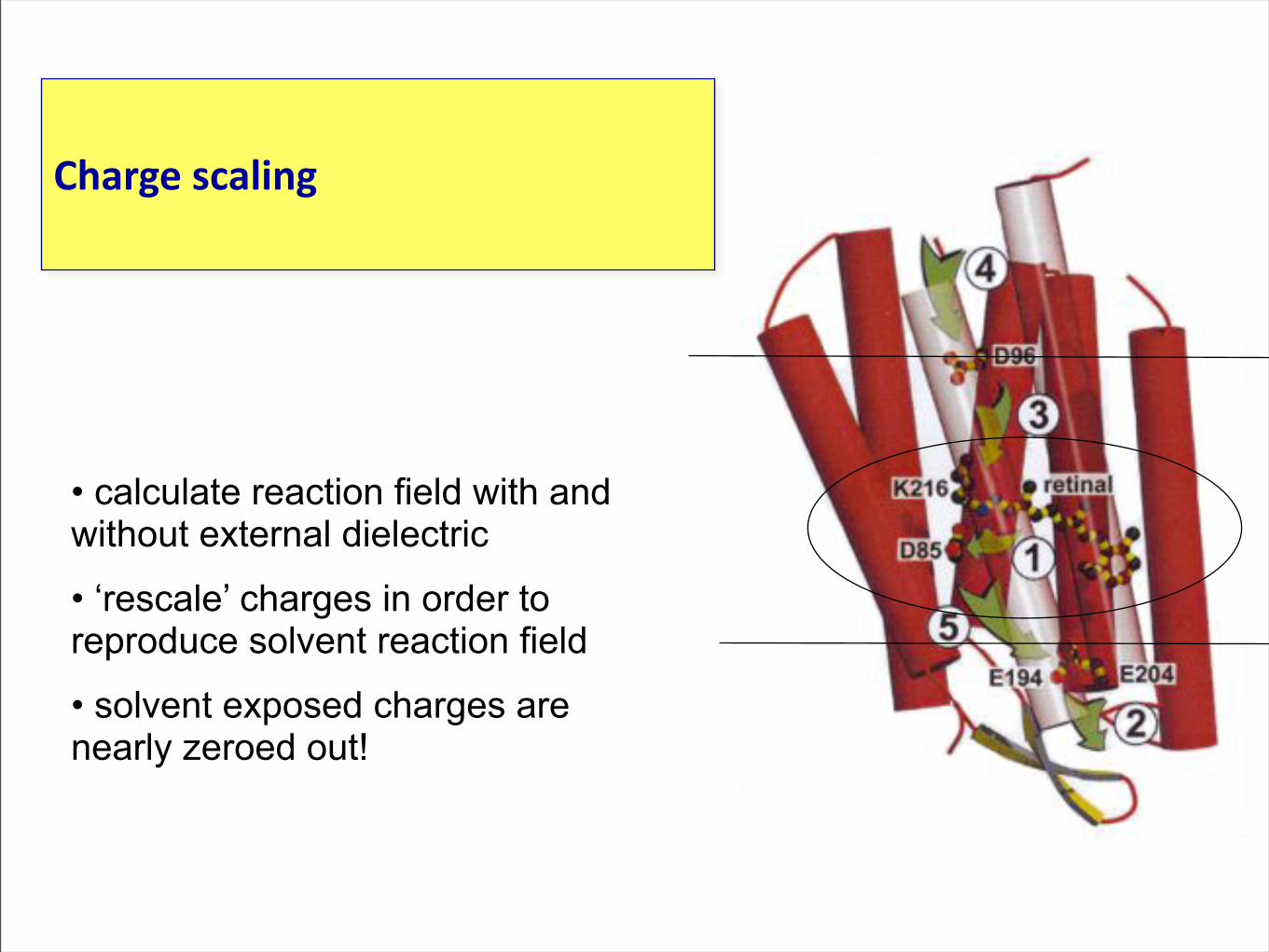
• calculate reaction field with and without external dielectric
• ‘rescale’ charges in order to reproduce solvent reaction field
• solvent exposed charges are nearly zeroed out!
!"#$%&'()#*+,%

3) QM/MM

simulating biological structures
complexity has 2 dimensions:
1) size: number of atoms2) time scale of processes: need MD to sample conformational space
environment of acitve site looks caotic,but is highly structured from a functional perspective
=> do not neglect environment, otherwise you loose the most important point!

! chemical reaction which needs QM treatment
! immediate environment: electrostatic and steric interactions
! solution, membrane: polarization and structural effects on protein and reaction!
" 10.000... - several 100.000 atoms
Computational problem I: number of atoms

combining models in multi-scale approaches
CI, MPCASPT2
Length scale
Continuum Electrostatics “Coarse graining”
Force Fields:MM
f
s
ps
ns
tim
e
ApproximativeMethods
HF, DFT
nm10"" 100"" 1000"" 10.000 " atoms

~ 1.000-100.000 atoms
~ ns MD simulations
(MD, umbrella sampling)
- chemical reactions
- excited states, spectroscopy
QM
Combined QM/MM methods
In many cases, the site of interest is localized" apply QM locally
Recent review: Senn & Thiel, Top Curr Chem #2007! 268: 173

1976 Warshel und Levitt
1986 Singh und Kollman
1990 Field, Bash und Karplus
QM • semi-empirical methods• quantum chemistry : DFT, HF, MP2, LMP2• DFT ‘plane wave‘ codes: CPMD
MM• CHARMM, AMBER, GROMOS, SIGMA,TINKER, ...
Combined QM/MM methods
Recent review: Senn & Thiel, Top Curr Chem #2007! 268: 173

!=80
QM region
Molecular Mechanics (MM) region
Effects:
- polarization of QM region through MM
- steric interactions
Main effect e.g. for catlytic efficiency of proteins
Recent review: Senn & Thiel, Top Curr Chem #2007! 268: 173
Combined QM-MM methods

MM
QM
! Mechanical embedding: only steric effects
! Electrostatic embedding: polarization of QM due to MM
! Electrostatic embedding + polarizable MM
QM/MM Methods

! Mechanical embedding: only steric effects
! Electrostatic embedding: polarization of QM due to MM
! Electrostatic embedding + polarizable MM
! Larger environment: - box + Ewald summ.
- continuum electrostatics
- coarse graining
MM
QM
? ?
QM/MM Methods

- subtractive: several layers: QM-MM
doublecounting on the regions is subtracted
- additive: different methods in different regions +
interaction between the regions
MM
QM
Subtractive vs. additive models

total energy
QM
MM=
-+ MM
Subtractive QM/MM: ONIOM Morokuma and co.: GAUSSIAN

total energy QM= +
QM+
MM
MM
interaction
Additive QM/MM

Additive QM/MM: linking

Additive QM/MM:

Additive QM/MM:
Elecrostatic mechanical embedding
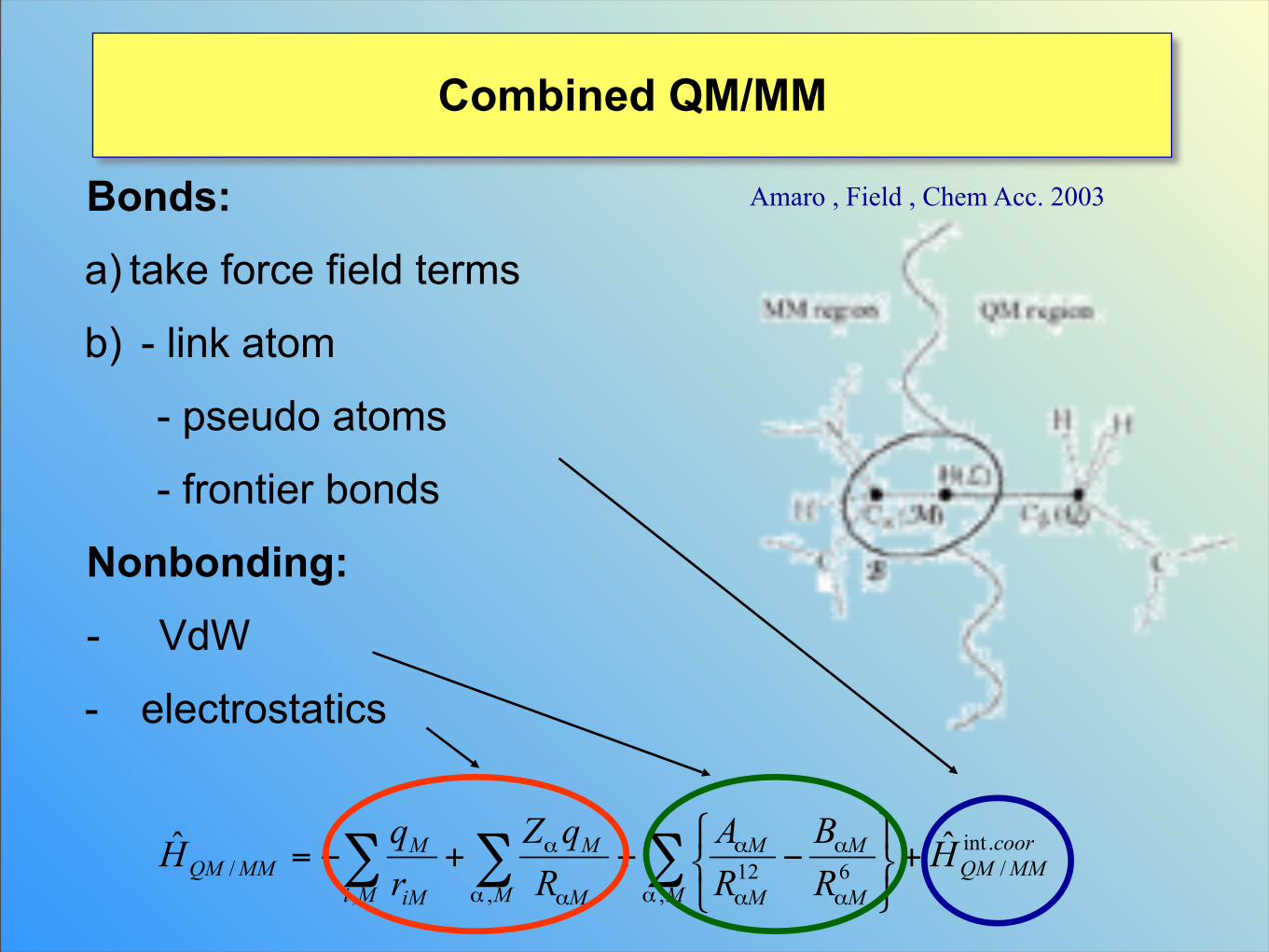
Combined QM/MM
Amaro , Field , Chem Acc. 2003Bonds:
a) take force field terms
b) - link atom
- pseudo atoms
- frontier bonds
Nonbonding:
- VdW
- electrostatics

Combined QM/MM
Reuter et al, JPCA 2000
Bonds:
a)
from force field

Combined QM/MM: link atom
a) constrain or not?
(artificial forces)
relevant for MD
b) Electrostatics
- QM-MM:
exclude MM-host
exclude MM-hostgroup
- DFT, HF: gaussian broadening of MM point charges, pseudopotentails (e spill out)
- J. Chem. Phys. 2002, 117, 10534 J. Phys. Chem. B 2005, 109, 9082

Combined QM/MM: frozen orbitals
Warshel, Levitt 1976
Rivail + co. 1996-2002
Gao et al 1998
Reuter et al, JPCA 2000

Combined QM/MM: Pseudoatoms
Amaro & Field ,T Chem Acc. 2003
Pseudobond- connection atom
Zhang, Lee, Yang, JCP 110, 46
Antes&Thiel, JPCA 103 9290
No link atom: parametrize C! H2 as pseudoatom
X

Nonbonding terms:
VdW
- take from force field
- reoptimize for QM level
Coulomb:
which charges?
Combined QM/MM
Amaro & Field ,T Chem Acc. 2003

Tests:
- C-C bond lengths, vib. frequencies
- C-C torsional barrier
- H-bonding complexes
- proton affinities, deprotonation
energies
Combined QM/MM

- parametrization of methods for all regions required
e.g. MM for Ligands
SE for metals
+ QM/QM/MM conceptionally simple and applicable
Subtractive vs. additive QM/MM

PW implementations
(most implementations in LCAO)
- periodic boundary conditions and large box!
lots of empty space in unit cell
- hybride functionals have better accuracy: B3LYP, PBE0 etc.
+ no BSSE
+ parallelization (e.g. DNA with ~1000 Atoms)
Local Orbital vs. plane wave approaches:

• QM and MM accuracy
• QM/MM coupling
• model setup: solvent, restraints
• PES vs. FES: importance of sampling
All these factors CAN introduce errors in similar magnitude
Problems

Z$1+$2&%0$6,&0#1+%3$/)0)'.*/$#J#/0$@5#$0&$#'#/0,&%0)*/$1+0#,)/*&+%P
'#%%$126&,0)+03
[$@#%&'()*&+
[$%0#,1/$#J#/0%$$[$$$+#),$)\)/$/&+9&,2)*&+$:]OM<
[$/&"#,#+0$@.+)21/%^
-,.&$(/#,.+,%'/"&'#)01,'12'&,345&(678#$("&*9'O++5;$Y#(;$Q1&6".%;$Q1&2&';$D0,5/0;$TUU_;$_T3`Tab`_:

Example bR: 1st step, excited states dynamics

2nd step: proton transfer

Retinal
Asp85
Thr89
Asp212
w402
direct proton transfer
How the protein shapes the barrier
•MM: CHARMM
•QM: DFT-B3LYP and semi-empirical SCC-DFTB
• Minimum Energy Path (MEP)
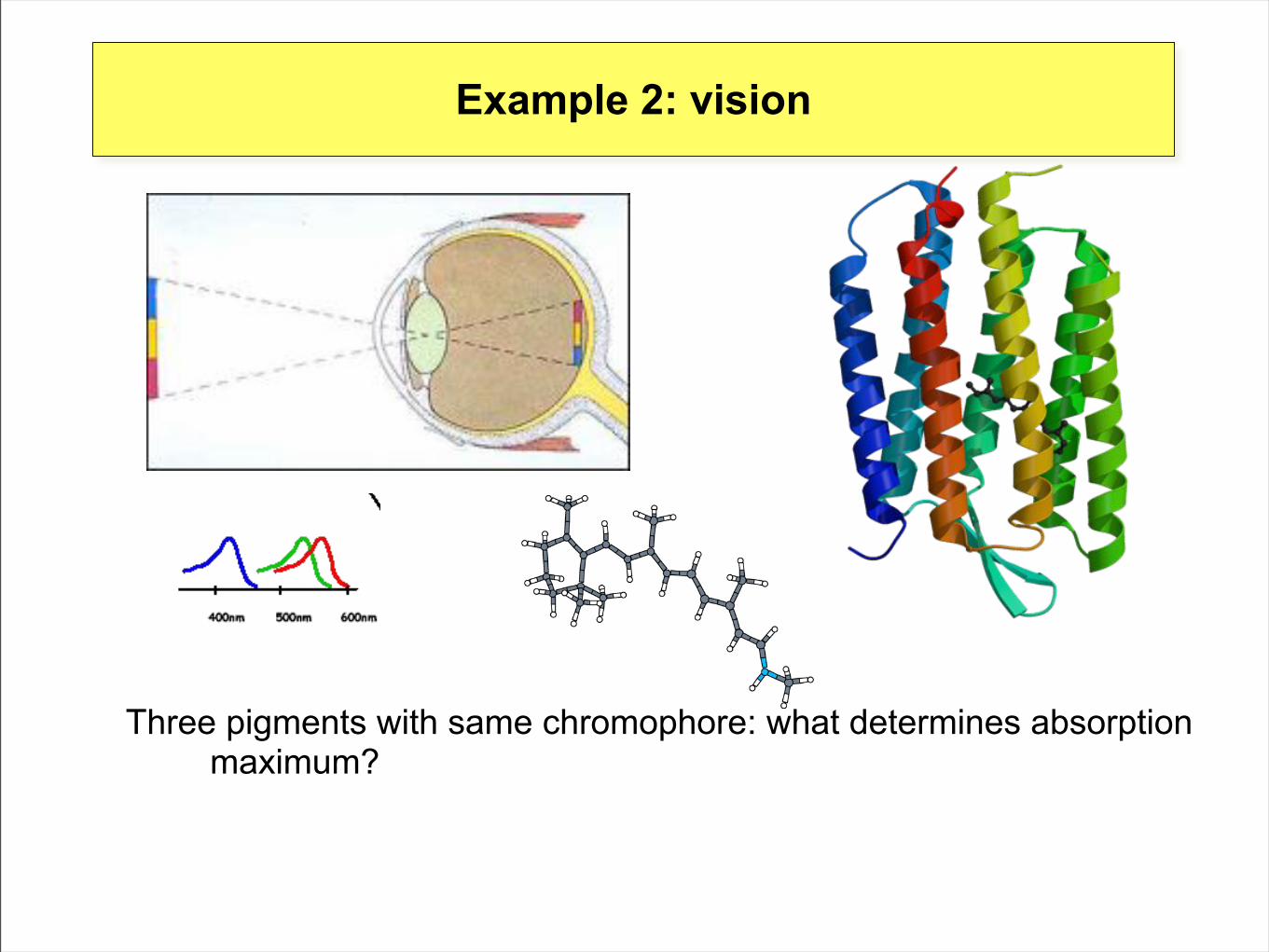
Example 2: vision
Three pigments with same chromophore: what determines absorption maximum?

Absorption over 300 nm“Tuning” by protein environment(opsin-shift)
‘Spectral tuning’
[$steric interactions: twist
- interaction with polar groups in environment
- H-bonding with counterions

Absorption over 300 nm“Tuning” by protein environment(opsin-shift)
‘Spectral tuning’
[$steric interactions: twist
- interaction with polar groups in environment
- H-bonding with counterions
-nearby amino acids have functional role: a single mutation can have drastic effects

;<
Long range forces in Biology
<<
Solvation of whole protein can be important:
=> a) periodic boundary: box filled with waterb) continuum electrostatic c) charge scaling

Multi-scale methods:QM1/QM2/MM/continuum
!linearized PB eq.
!=80
!=2

Computational problem II: sampling with MD
!$flexibility: not one global minimum
" conformational entropy
! solvent relaxation
" ps – ns timescale (timestep ~ 1fs)

Different energy profiles for different
protein conformations
‘Problem’ of potential energy
c")+-$#0$)'$dWMQ$RUe$:TUU_<$```aS

Different energy profiles for different
protein conformations
‘Problem’ of potential energy
c")+-$#0$)'$dWMQ$RUe$:TUU_<$```aS
A) One always has to average over the different conformations of the environment :
Total energy" inner energyE" U
B) Entropy is often as important as accurate total energy : U" F

$
G&'#/5'),$X.+)21/%$:GX<3$
]52#,1/)'$1+0#-,)*&+$&9$]#C0&+%$$#=5)*&+$&9$2&*&+$Lf2g)$$C10"$
*2#%0#63$hR9%
"$0,)i#/0&,1#%
"$R6%$@.+)21/%3$RUUU$9&,/#$#()'5)*&+%
Molecular Dynamics: MD

First Molecular Dynamics Simulation (MD) of a protein: 9.2 ps
G/$M)22&+7$V#'1+j$k),6'5%7$])05,#$Tle7$=>??
QW!E
:4&(1+#$6)+/,#)*/$0,.6%1+$
1+"1410&,<
am$)21+&$)/1@%

First Molecular Dynamics Simulation (MD) of a protein: 9.2 ps
G/$M)22&+7$V#'1+j$k),6'5%7$])05,#$Tle7$=>??
QW!E
:4&(1+#$6)+/,#)*/$0,.6%1+$
1+"1410&,<
am$)21+&$)/1@%
,&&.&.6
)$4(/#*'(/$@)/@$&

BPTI: 210ps and water
n#(1\$j$D"),&+7$W]OD$ma7$=>AAB

BPTI: 210ps and water
n#(1\$j$D"),&+7$W]OD$ma7$=>AAB
/1.#46''C=DDBDDD'#/15(
''''C=DE=DD',(
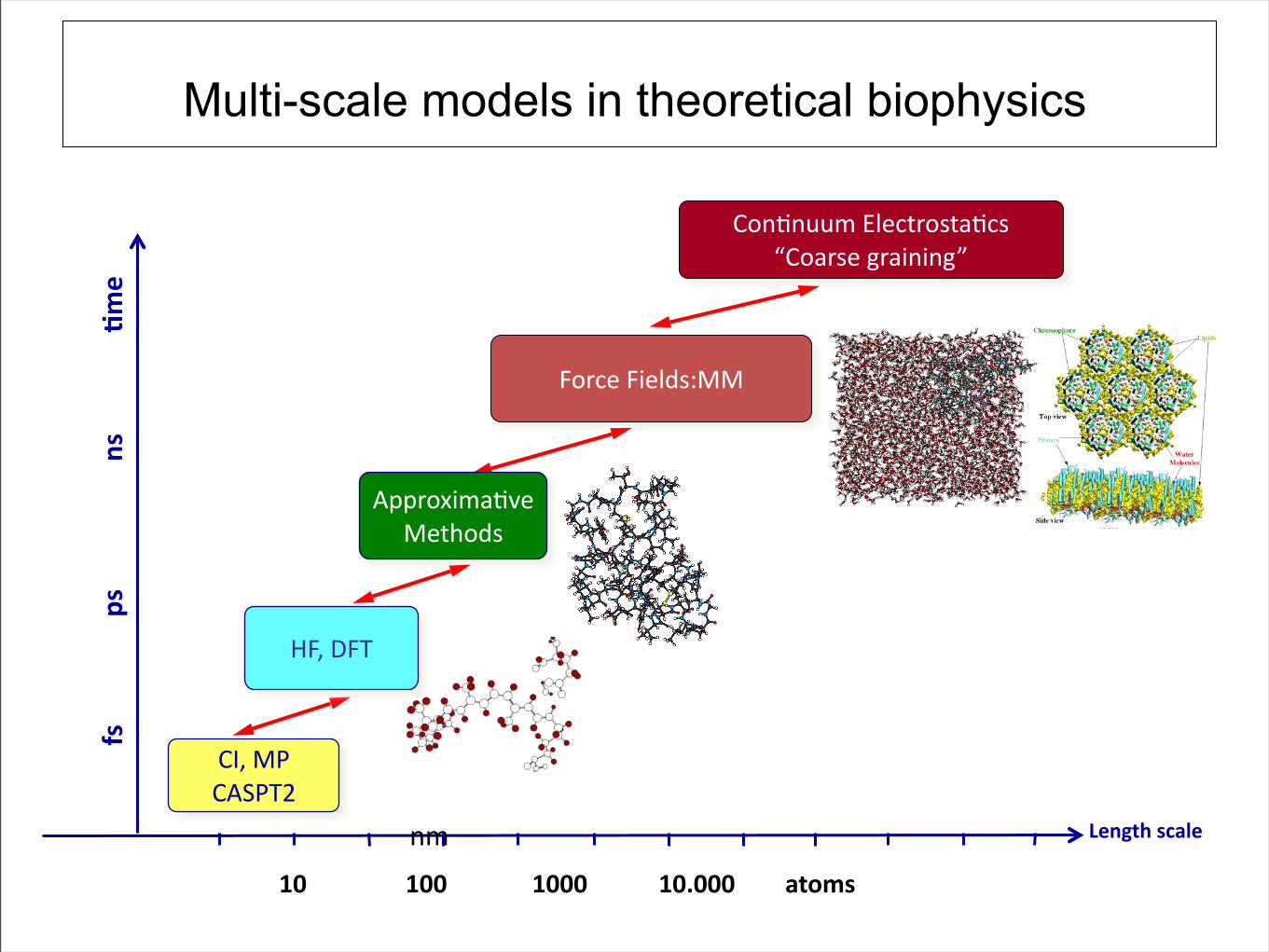
Multi-scale models in theoretical biophysics
ME7$GW
MODW!T
F&,%/"'()#*&
M&+*+552$o'#/0,&%0)*/%$
pM&),%#$-,)1+1+-q
L&,/#$L1#'@%3GG
''''''''''2(''''''''''''''''G(''''''''''''''''''',('''''''''''''''05&
O66,&?12)*(#
G#0"&@%
rL7$XL!
+2
=D' ' =DD' ' =DDD' ' =DBDDD' #/15(

Multi-scale models in theoretical biophysics
ME7$GW
MODW!T
F&,%/"'()#*&
M&+*+552$o'#/0,&%0)*/%$
pM&),%#$-,)1+1+-q
L&,/#$L1#'@%3GG
''''''''''2(''''''''''''''''G(''''''''''''''''''',('''''''''''''''05&
O66,&?12)*(#
G#0"&@%
rL7$XL!
+2
=D' ' =DD' ' =DDD' ' =DBDDD' #/15(
problem: time step~ 1fs
s: energy + forces
min – h: energy + forces
h – days: energy + forces

















